
Фотосинтез и его фазы (световая и темновая). Схема фотосинтеза у растений
упрощенная и полная схема фотосинтеза, определение
ФОТОСИНТЕЗ (от фото... и греч. synthesis — соединение) — образование клетками автотрофных организмов органических веществ из неорганических при участии и за счет энергии солнечного света. У растений и цианобактерий донором электронов является вода и в процессе фотосинтеза выделяется кислород (оксигенный тип фотосинтеза). Фотосинтезирующие бактерии, использующие иные, чем вода, доноры электронов, кислород не выделяют (аноксигенный). Обязательное условие оксигенного фотосинтеза — наличие фотосистем I и II, где главным компонентом является пигмент хлорофилл в двух ею модификациях: a и b. Фотосинтез и продукты превращения углеводов служат основой биосинтеза биологически активных веществ в растениях. Суммарная реакция фотосинтеза имеет следующий вид:

Фотосинтез включает две фазы — световую, связанную со светом и пигментами и проходящую в гранах, и темновую фазу, не требующую ни света, ни пигментов и осуществляющуюся в строме. Световая фаза заключается в образовании восстановительного потенциала в виде восстановленной формы NADPH и запаса энергии в виде АТР. Темновая фаза заключается в фиксации СО2 и ее восстановлении до углевода. При этом расходуются поступившие в строму NADPH и АТР и все реакции катализируются специфическими ферментами. Известны три разных механизма темновых реакций фотосинтеза у высших растений. Основной механизм — фиксация СО2 в цикле фотосинтетического восстановления углерода, называемого циклом Кальвина. В последнее время его стали называть С3-путем или С3-типом фотосинтеза.
Ключевая реакция цикла Кальвина — взаимодействие СО2 с D-рибулозо-1,5-дифосфатом (РДФ), в результате чего получается 3-фосфоглицериновая кислота (3-ФГК), которая фосфорилируется с образованием 1,3-дифосфоглицериновой кислоты (1,3-ДФГК) с участием АТР, синтезированной в световой фазе фотосинтеза. Далее 1,3-ДФГК восстанавливается при посредстве NADPH до 3-фосфоглицеринового альдегида (3-ФГА). На этой стадии СО2 уже восстановлена до триозофосфата, т. е. до уровня углевода. Затем 3-ФГА частично изомеризуется в фосфат дигидроксиацетона (ДГАФ).
При взаимодействии ДГАФ и 3-ФГА получается фруктозо-1,6-дифосфат, затем преобразующийся в фруктозо-6-фосфат.
Таким образом, синтезируется фосфат гексозы, который либо идет на образование крахмала или др. полисахаридов, либо превращается в фосфаты др. сахаров или потребляется на другие нужды растительного организма.
Однако для того чтобы снова могла осуществиться фиксация СО2, необходима регенерация ее акцептора, т. е. дифосфата рибулозы. В результате ферментативных превращений фосфата глицеринового альдегида, фосфата дигидроксиацетона и фруктозодифосфата в качестве промежуточных соединений возникают эритрозофосфат, седогептулозофосфат, а также фосфаты ксилулозы и рибозы, которые затем превращаются в рибулозо-1,5-дифосфат — акцептор СО2.
На схеме 1 представлен цикл восстановления углерода при фотосинтезе, или цикл Кальвина.
Схема 1. Цикл восстановления углерода при фотосинтезе, или цикл Кальвина (по Гудвину и Мерсер, 1986)

Рассмотренный путь восстановления СО2 — главный, но не единственный. Образовавшиеся в процессе фотосинтеза запасные полисахариды потребляются растением на путях дыхания и брожения. Эти процессы — источник энергии, источник многочисленных лабильных соединений, которые синтезируются в качестве промежуточных продуктов и вместе с тем служат исходным материалом для разнообразных синтетических реакций.
Полисахариды подвергаются гликолизу, который происходит по реакциям, свойственным фотосинтезу, но идущим в обратном направлении с участием других ферментов до образования глицериновой кислоты. Глицериновая кислота далее может подвергаться различным превращениям, в т. ч. окисляться до ацетила. Ацетил, активированный СоА, включается в многочисленные процессы биосинтеза. Ацетил-СоА получается не только на путях окисления углеводов, но и на путях окисления жирных кислот и аминокислот.
На схеме 2 приведены упрощенные пути биосинтеза разных групп действующих веществ, неразрывно связанные с фотосинтезом.
В этой схеме ацетил-СоА занимает центр, место и идет на построение разнообразных биологически активных веществ.
Схема 2. Упрощенная схема путей биосинтеза групп действующих веществ в С3-растениях

www.9lc.com
Фотосинтез у растений общая схема
Фотосинтез у растений общая схема [c.333]Источником углеводов в природе служит процесс фотосинтеза — превращение в зеленых листьях растений углекислого газа воздуха в углеводы. Энергию для этого процесса дает солнечный свет. Фотосинтез служит единственным источником органических веществ в живой природе, поскольку животные неспособны синтезировать органические вещества нз неорганических они лишь перерабатывают органические веи ества, накопленные растениями. Велика роль продуктов фотосинтеза и в качестве источников энергии и каменный уголь, и нефть, и газ, и тем более древесина — все это консервированная солнечная энергия , накопленная за счет фотосинтеза. Общий результат фотосинтеза можно выразить схемой [c.317]
В этой работе, проведенной в нашей лаборатории Л. А. Филипповой, у нескольких видов растений высокогорных пустынь Восточного Памира были определены дневные изменения интенсивности фотосинтеза. Результаты определений (табл. 5), проведенных на срезанных листьях, выражены в миллиграммах СО2, поглощенной на 1 г сухого веса листьев за 10 мин. фотосинтеза в токе радиоактивной углекислоты. Материал, фиксированный спиртом, был взят для количественного радиохимического анализа, проведенного по предложенной выше схеме. Получив данные о весе и удельной радиоактивности всех фракций органических веществ, содержащих , вычислялось общее количество импульсов в минуту для каждой фракции. Зная последнюю величину и удельную радиоактивность углекислоты, взятой для опыта, по методике, описанной на стр. 16, определялось количество миллиграммов СО2, вошедшее в каждую фракцию органических веществ листа. [c.48]СОа + НаО = [НСНО] 4- Оа где [НСНО] обозначает образующиеся в клетках растений первичные продукты ассимиляции, которые затем уже без участия света конденсируются в углеводы, превращаются в карбоновые кислоты, белки и т. д. Эта простая схема в общих чертах передает лишь внешний результат первых стадий фотосинтеза и не поясняет ни его путей, ни внутреннего механизма. [c.473]
Фотосинтез происходит не во всех частях растения. Тем из них, которые удалены от фотосинтезирующих структур, например корням, нужна специальная транспортная система снабжения ассимилятами. У сосудистых растений органические продукты переносятся из главных органов фотосинтеза — листьев — ко всем остальным частям растения по флоэме. На рис. 13.21 приведена общая схема связей между автотрофными клетками, образующими органические питательные вещества, и клетками, получающими эти вещества. Как видно из этого рисунка, органические вещества у растений могут перемещаться по побегам как вверх, так и вниз. Это отличает флоэму от ксилемы, по которой транспорт осуществляется только вверх. Следует также отметить, что запасающие органы в разное время могут функционировать то как источники ассимилятов, то как их потребители. [c.129]
Последовательность реакций, в которых диоксид углерода связывается в процессе фотосинтеза, была впервые предложена в 50-х годах Кальвином ее часто называют циклом Кальвина или фотосинтетическим циклом восстановления углерода (см. схему 4). В отличие от световой реакции, свойственной только фотосинтезирующим тканям, синтез углеводов из диоксида углерода имеет много общего с реакциями, используемыми для синтеза углеводов в нефотосинтезирующих организмах. Тем не менее поражают масштабы этого процесса в зеленых растениях по самым минимальным оценкам растения ежегодно связывают около 35-10 кг углерода, причем для получения каждого грамма связанного углерода растение должно переработать более 6250 л воздуха. Хотя 99 % диоксида углерода, усваиваемого растениями из воздуха, связывается в процессе фотосинтетических реакций на свету, существуют и процессы темнового карбоксилирования [2], отличающиеся высокой скоростью и вносящие значительный вклад в общее количество связываемого углерода некоторых растений, в особенности суккулентов (сем. rassula eae). [c.398]
Можно сделать одно возражение по поводу механизма (19.13). Реакция (17.13б), повидимому, одинакова с реакцией, при которой восстановители-заменители заменяют воду в фотоеинтезе, бактерий и анаэробно адаптированных водорослей. Если эта реакция может происходить у всех зеленых растений (т. е. не нуждается в посредничестве гидрогеназы), возникает вопрос, почему все они не могут восстанавливать двуокись углерода за счет клеточных или введенных извне органических водородных доноров, т. е. осуществлять фоторедукцию с органическими восстановителями вместо фотосинтеза (Этот вопрос уже ставился в главе VI.) На это следует ответить, что фоторедукция возможна, но у фотосинтетических активных растений вероятность, что o hl прореагирует с аодой, настолько выше вероятности его реакции с другим водородным донором (А — в реакции (19.13в), HgR—в схеме на фиг. 11), что эта последняя реакция остается незаметной, С другой стороны, лри фотоокислении лишь малая часть молекул оСЫ, реагирующих с А, вызовет общее химическое изменение, так как реакция оСЫ с HgO [c.561]
Данные, подтверждающие концепцию о существовании двух различных фотосистем, еще не позволяют оценить относительный вклад каждой фотореакции в общий процесс. Одно из слабых мест в схеме переноса электронов, представленной на фиг. 219,— допущение равного участия обеих фотосистем по одному фотону на эквивалент. Тогда возникает вопрос, как поглощенные кванты распределяются между двумя фотореакциями, так чтобы обеспечить образование первичных фотопродуктов в нужном соотношении Были выдвинуты две гипотезы. Согласно первой из них — так называемой гипотезе раздельной упаковки ( separate pa kage ), существуют две полностью раздельные фотосистемы, причем каждая из них имеет свой собственный набор пигментов. Передача энергии может происходить между пигментами данной системы, но не от одной системы к другой. По этой гипотезе выход может быть максимальным (т. е. усиление отсутствует) именно при тех длинах волн, при которых поглощение каждой пигментной системы и выходы отдельных фотореакций равны. Тогда анализ спектров действия, приведенных на фиг. 226 и 227, наводит на мысль, что вспомогательные пигменты, которые сенсибилизируют фотосинтез очень эффективно, почти поровну разделены между двумя системами (с фотосистемой II связано несколько больше пигмента). Обе системы содержат также одну или более форм хлорофилла а. У зеленых растений эти системы содержат примерно равное количество хлорофилла а, за исключением длинноволнового компонента, который сенсибилизирует только систему I. У сине-зеленых и красных водорослей система I содержит значительно больше хлорофилла а, чем система II. Такое несоответствие приводит к тому, что не все фотопродукты системы I находят партнеров по реакции из системы II, и, следовательно, общая эффективность в той области, в которой в основном поглощает хлорофилл, будет низкой. [c.570]
В 1894 г. Эмиль Фишер впервые четко сформулировал определение асимметрического синтеза на основании своих экспериментов по превращению моносахаридов в высшие гомологи по схеме циангидрипового синтеза, сопоставив этот процесс непосредственно с биохимическим процессом образования оптически активных сахаров в растениях [1]. Он высказал предположение, что двуокись углерода и вода, конденсируясь под влиянием солнечного света и хлорофилла, образуют формальдегид, который затем вступает в реакцию конденсации с другой молекулой формальдегида или с простейшими углеводами, причем под направленным воздействием оптически активных соединений в хлорофиллсодержащих зернах клетки реакция протекает таким образом, что вхождение каждого следующего асимметрического атома в цепь приводит к образованию только одной из двух возможных стерео-изомерных форм. В результате образуется молекула моносахарида. прочно связанная с хлорофиллом. Это образование затем распадается на оптически активный моносахарид и регенерированный хлорофилл — катализатор, способный вновь участвовать в этом цикле [1, 2]. Химический путь превращения углерода при фотосинтезе, установленный в настоящее время, в деталях имеет мало общего с этой упрощенной схемой, предложенной в конце прошлого столетия. Тем пе менее представления об асимметри-ческол синтезе, выдвинутые Фишером, в общих чертах остаются в силе и до настоящего времени. [c.11]
Одно из центральных мест принадлежит фосфорной кислоте в процессах аэробного дыхания и фотосинтеза. Речь идет о процессах окислительного и фотосинтетического фосфорилирования, которые подробно разбирались в главах Дыхание и Фотосинтез . Именно этим и определяется влияние фосфора на общий уровень метаболитической активности тканей растения, на активность синтетических процессов и, в частности, на синтез белков. На рис. 125 дана схема, иллюстрирующая современные представления о роли фосфора в энергетическом обмене. [c.414]
Рассмотрим общие особенности функционирования и организации ННФ. В фотосинтезе высших растений перенос электрона от воды (окислительновосстановительный потенциал полувосстановления, 1/2, при pH 7 равен -1-0,82 В) к НАДФ Eij2 —0,32 В) осуществляется при затрате энергии двух квантов света с участием соответственно двух функционально сопряженных и работающих последовательно фотосистем (ФС П и ФС I). Схема взаимодействия фотосистем и промежуточных переносчиков в ЭТЦ фотосинтеза высших растений показана на рис. XXVH.2 там же даны соответствующие значения 1/2 компонентов ЭТЦ. [c.280]
Дискретность и целостность. Жизнь характеризуется диалектическим единством противоположностей, она одновременно и целостностна и дискретна (от лат. упрощенной схеме это можно представить так животные-хищники для своего питания нуждаются в существовании растительноядных, живот-ные-растительноядные—в существовании растений, растения в процессе фотосинтеза поглощают из атмосферы СОг, выделение которого в атмосферу связано с жизнедеятельностью живых организмов кроме того, растения из почвы получают ряд минеральных веществ, количество которых не истощается благодаря разложению органических веществ, осуществляемому бактериями, и т. д. Органический мир целостен, так как составляет систему взаимосвязанных частей, и в, то же время он дискретен. Ои состоит из единиц—организмов, или особей. Каждый живой организм дискретен, так как состоит из органов, тканей, клеток, но вместе с тем каждый из органов, обладая определенной автономностью, действует как часть целого. Каждая клетка состоит из органоидов, но функционирует как единое целое. Наследственная информация осуществляется генами, но ни один из генов вне всей совокупности не определяет развитие признаков и т. д. Идя далее, следует сказать, что жизнь связана с молекулами белков и нуклеиновых кислот, но только их единство, целостная система обусловливает существование живого. [c.13]
chem21.info
Схема процесса фотосинтеза: магия света
Фотосинтез – процесс преобразования органики из неорганических веществ – углекислоты и воды. В этом процессе непосредственное участие принимает энергия солнечного света, без нее он не возможен вообще. Формула, объясняющая этот процесс:
6СО2+6Н2О+свет→С6Н12О6+6О2.
У высших растений, таких как цветы или деревья, органом, ответственным за процессы фотосинтеза является лист.
Главными органоидами выступают входящие в состав листа хлоропласты. В них встроены фотосинтетические вещества – пигменты под названиями хлорофиллы и каротиноиды. Различают следующие типы хлорофилла — a, b, c, d, главным среди них является хлорофилл типа a. Хлорофиллы поглощают красный и сине-фиолетовый лучи из спектра, в свою очередь отражают зеленый, что и позволяет наблюдать у растений характерную окраску. Схема процесса фотосинтеза наглядно показывает, что это сложный многоступенчатый процесс, притекающий в несколько этапов.
Световая фаза: первый этап фотосинтеза
Эта фаза, как можно понять из названия, связана с солнечной энергией, то есть все процессы происходят исключительно в присутствии света в мембранах тилакоидов. Под действием света электроны хлорофилла становятся активными, покидают молекулу, откуда попадают на внешнюю сторону мембраны тилакоида. Окисленные молекулы хлорофилла подвергаются процессу восстановления, отбирая электроны у воды. Выглядит это следующим образом: Н2О + квант света = Н++ ОН-.
Кислород удаляется из процесса, попадая в окружающее пространство протоны копятся внутри тилакоида. Затем они проникают через специальные каналы АТФ-синтетазы, так происходит еще одно превращение веществ, называемое процессом фосфорилирования АДФ, результатом которого становится появление родственного ей вещества АТФ. Таким образом, в этой фазе происходит процесс фотолиза воды, а АТФ и НАДФ транспортируются в строму хлоропласта, где продолжают, участив в темновой фазе.
Темновая фаза: бессветовой этап
Схема процесса фотосинтеза указывает на то, что этот этап происходит в структурной единице хлоропласта — строме. Для запуска реакций здесь вовсе не нужна энергия солнечного света. Реакции второго этапа — темновой фазы представляют собой химическую цепочку поэтапных превращений СО2, в результате которых образуется глюкозы и другие вещества органической природы.
Открывает химические превращения реакция фиксации СО2. В этих превращениях активно используются энергии веществ АТФ и НАДФН2, возникших на предыдущем этапе. Цикл химических превращений в темновой фазе называется циклом Кальвина
Кроме глюкозы (С6Н12О6), в процессе взаимодействия и превращения веществ и молекул образуются разнообразные аминокислоты, спирты (глицерин) и жирные кислоты, а также нуклеотиды.
Значение фотосинтеза для живой природы
Благодаря явлению фотосинтеза, происходит восстановление окружающей среды, ее очищение от загрязняющих веществ, так как ежегодно поглощаются миллиарды тонн, СО2 , в замен образуется столько же кислорода. Этот процесс является основным источником образования органических веществ в природе. Из кислорода образуется уникальный защитный слой – озоновый, который оберегает все живое от ультрафиолетовой космической радиации.
Механизм фотосинтеза — видео
life-students.ru
Фотосинтез и его фазы (световая и темновая)
Фотосинтез — уникальная система процессов создания с помощью хлорофилла и энергии света органических веществ из неорганических и выделения кислорода в атмосферу, реализуемая в огромных масштабах на суше и в воде.
Фотосинтез происходит в клетках зелёных растений с помощью пигментов, главным образом хлорофилла, находящегося в хлоропластах клетки. Его продуктами являются мономеры углеводов (моносахариды: глюкоза, фруктоза и др.).
В основе фотосинтеза лежит окислительно-восстановительный процесс, в котором электроны переносятся от донора-восстановителя (вода, водород и др.) к акцептору (CO2 , ацетат и др.) с образованием восстановленных соединений (углевода) и выделением кислорода, если окисляется вода. Фотосинтезирующие бактерии часто используют другие доноры, а не воду, кислород при этом они не выделяют.
В системе процессов фотосинтеза различают два цикла реакций, как две фазы, последовательно и непрерывно идущие друг за другом — световую и темновую (рис. 62).
Световая фаза фотосинтеза характеризуется тем, что здесь все процессы происходят только при участии энергии света, поэтому её и называют световой. Связывание солнечной (электромагнитной) энергии происходит преимущественно на мембранах тилакоидов хлоропласта. Размещающийся здесь хлорофилл и другие пигменты собраны в функциональные единицы-комплексы — пигментные системы, получившие название фотосистемы.
 |
| Рис. 62. Схема фотосинтеза |
Таким образом, светособирающие и пигментно-белковые комплексы фотосистемы I и фотосистемы II обеспечивают процесс фотосинтеза необходимой энергией в виде макроэнергетических соединений НАДФ•Н и АТФ. В этом заключается основная функция световой фазы фотосинтеза. Она реализуется только при участии света и с помощью пигментов, размещённых в тилакоидной мембране хлоропластов.
Темновая фаза фотосинтеза проходит в строме хлоропласта без непосредственного поглощения света, в любое время суток. В процессе световой фазы фотосинтеза накапливается достаточно высокий уровень АТФ и НАДФ•Н. Однако сами по себе эти высокоэнергетические соединения не способны синтезировать углеводы из CO2. Поэтому становится очевидным, что и темновая фаза фотосинтеза — сложный процесс, включающий большое количество реакций. Материал с сайта http://doklad-referat.ru
Все процессы темновой фазы фотосинтеза идут без непосредственного потребления света, но в них большую роль играют высокоэнергетические вещества (АТФ и НАДФ•Н), образующиеся с участием энергии света, во время световой фазы фотосинтеза. В процессе темновой фазы энергия макроэнергетических связей АТФ преобразуется в химическую энергию органических соединений молекул углеводов. Это значит, что энергия солнечного света как бы консервируется в химических связях между атомами органических веществ, что имеет огромное значение в энергетике биосферы и конкретно для жизнедеятельности всего живого населения нашей планеты.
Фотосинтез происходит в хлоропластах клетки и представляет собой синтез углеводов в хлорофиллоносных клетках, идущий с потреблением энергии солнечного света. Различают световую и темповую фазы фотосинтеза. Световая фаза при непосредственном потреблении квантов света обеспечивает процесс синтеза необходимой энергией в виде НАДН и АТФ. Темновая фаза — без участия света, но путем многочисленного ряда химических реакций (цикл Кальвина) обеспечивает образование углеводов, главным образом глюкозы. Значение фотосинтеза в биосфере огромно.
Вопросы по этому материалу:Где происходит фотосинтез в растительной клетке?
Какие превращения происходят во время световой фазы фотосинтеза?
Что является результатом световой фазы фотосинтеза?
Где протекают темновые реакции фотосинтеза?
doklad-referat.ru
Упрощенная схема фотосинтеза: как появляется кислород? | We are students
Фотосинтез представляет собой процесс образования автотрофными организмами органических веществ (крахмал, сахар) из неорганических. Процесс осуществляется с помощью солнечного света, который поглощается пигментами растений. Упрощенная схема фотосинтеза наглядно это демонстрирует.
В ходе фотосинтеза происходит выработка кислорода, необходимого для поддержания жизнедеятельности животных, растений и человека.
Как происходит процесс фотосинтеза
Реакция фотосинтеза имеет следующий вид:

Органическими веществами выступают практически все соединения углерода (кроме нитридов и оксидов). Фотосинтез образовывает такие органические вещества, как аминокислоты, углеводы и жирные кислоты. Неорганическими веществами выступает диоксид углерода и вода.
Диоксид углерода растения получают из воздуха, поступает он в листья, а уже синтезированный кислород выделается через устьица. Устьица – это своеобразные поры, отверстия на эпидермисе. За работу этих пор отвечают зеленые клетки, которые тоже осуществляют фотосинтез, называются они замыкающими. При попадании солнечного света на эти клетки, они растягиваются, тем самым открывая устьица все шире. Таким образом, диоксид углерода (CO2) попадает в нижние слои листа. Свет в процессе фотосинтеза – это не только катализатор, но и реагент.
Упрощенная схема фотосинтеза внесет ясность, так как весь процесс можно будет отследить наглядно.
Стадии фотосинтеза
Процесс фотосинтеза включает в себя две фазы:
- Световой этап;
- Темновой этап.
Световая фаза может проходить лишь только в случае попадания солнечных лучей. Осуществляется фаза в хлоропластах. Хлоропласты представляют собой пластиды, которые окрашены пигментом хлорофиллом в зеленый цвет. Именно этот пигмент и поглощает солнечный свет, образуя при этом кислород. Итак, солнечный свет трансформируется в молекулы АТФ. АТФ – это нуклеотид, предназначенные для транспортировки энергии реакций. С помощью света также происходит образование молекулы НАДФ – она образовывается посредством отщепления водорода от воды.
Темновой этап происходит в стромах листьев. С помощью энергетических молекул, произведенных в световой фазе, осуществляется синтез, результатом которого являются органические вещества.
Частота фотосинтеза зависит от нескольких факторов: температурного режима, концентрации углерода, интенсивности солнечных лучей, минерального питания растения.
Упрощенная схема фотосинтеза:
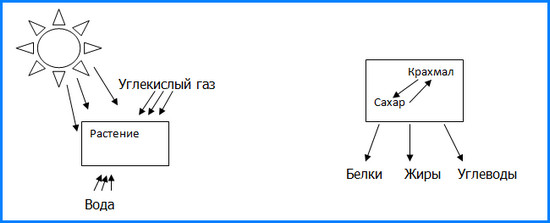
Схема показывает три составляющих фотосинтеза — вода внутри зеленого листа, солнечный свет и углекислый газ. Таким образом, происходит запуск синтезирования.
Фотосинтез – это самый главный процесс, благодаря которому существуют все живые организмы на планете. Массовая вырубка лесов чревата печальными последствиями. В худшем случае, будет чувствоваться нехватка кислорода на Земле.
Процесс фотосинтеза — видео
westud.ru
Фотосинтез
Фотосинтез (от греч. φωτο- - свет и греч. σύνθεσις - синтез, совокупность) - процесс синтеза органических соединений с углекислого газа и воды с использованием энергии света и при участии фотосинтетических пигментов : ( хлорофилл в растений, хлорофилл, бактериохлорофилл и бактериородопсина в бактерий), часто с выделением кислорода в качестве побочного продукта. Это чрезвычайно сложный процесс, включающий длинную последовательность координированных биохимических реакций. Он происходит в высших растениях, водорослях, многих бактериях, некоторые архея и простейших - организмах, известных вместе как фототрофы. Сам процесс играет важную роль в круговорота углерода в природе.
Фотосинтез - единственный процесс в биосфере, который приводит к усвоению энергии Солнца и обеспечивает существование как растений, так и всех гетеротрофных организмов. [1]
Обобщенное уравнение фотосинтеза ( брутто-формула) имеет вид:
6СО 2 + 6Н 2 О = С 6 Н 12 О 6 + 6О 21. Типы фотосинтеза
Различают оксигенной и аноксигенным типа фотосинтеза. Оксигенной наиболее распространенный, его осуществляют растения, цианобактерии и прохлорофиты. Аноксигенным фотосинтез проходит в пурпурных, некоторых зеленых бактериях и гелиобактериях.
Выделяют три этапа фотосинтеза: фотофизических, фотохимический и химический. На первом этапе происходит поглощение фотонов света пигментами, их переход в возбужденное состояние и передача энергии к другим молекулам фотосистемы. На втором этапе происходит разделение зарядов в реакционном центре, перенос электронов по фотосинтетической електронотранспортному цепи, заканчивающийся синтезом АТФ и НАДФН. Первые два этапа вместе называют светозависимый стадией фотосинтеза. Третий этап происходит уже без обязательного участия света и включает биохимические реакции синтеза органических веществ с использованием энергии, накопленной на светозависимый стадии. Чаще всего в качестве таких реакций рассматривается цикл Кальвина и глюконеогенез, образование сахаров и крахмала из углекислого газа воздуха.
2. Пространственная локализация
Фотосинтез растений осуществляется в хлоропластах, обособленных двомембранних органеллах клетки. Хлоропласты могут быть в клетках плодов, стеблей, однако основным органом фотосинтеза, анатомически приспособленным к его осуществлению, является листья. В листке богатая хлоропластами ткань - палисадная или фотосинтезирующей / столбчатая / хлорофилоносным, паренхима. В некоторых суккулентов с вырожденным листьями (например, кактусов) основная фотосинтетическая активность связана со стеблем.
Свет для фотосинтеза захватывается более полно благодаря плоской форме листа, обеспечивающей большое отношение поверхности к объему. Вода доставляется из корня развитой сетью сосудов (прожилок листа). Углекислый газ поступает отчасти с помощью диффузии через кутикулу и эпидермис, однако большая его часть диффундирует в листья через устьица и по листу через межклеточное пространство. Растения, которые осуществляют С 4 и CAM фотосинтез сформировали особые механизмы для активной ассимиляции углекислого газа.
Внутреннее пространство хлоропласта заполнено бесцветной веществом (стромой) и пронизано мембранами ( ламелями), которые, соединяясь друг с другом, образуют тилакоиды, которые в свою очередь группируются в стопки, так называемые грани. Внутришньотилакоидний пространство отделен и не сочетается с остальными стромы, предполагается также, что внутреннее пространство всех тилакоидов соединен между собой. Световые стадии фотосинтеза связаны с мембранами, автотрофные фиксация углекислого газа происходит в строме.
В хлоропластах есть свои ДНК, РНК, рибосомы (70 S типа), идет синтез белка (хотя этот процесс и контролируется из ядра). Они не синтезируются вновь каждый, а образующиеся путем деления предыдущих. Все это позволило считать их предшественниками свободных цианобактерий, вошедших в состав эукариотической клетки в процессе симбиогенеза.
Цианобактерии и другие фотосинтетизуючи бактерии и археи, таким образом, сами выполняют функции хлоропластов растений и фотосинтетический аппарат их клеток не вынесен в особую органеллы. Их тилакоиды, однако, не образуют стопок, а формируют различные складчатые структуры (в одной цианобактерии Gloeobacter violaceus тилакоиды отсутствуют вовсе, а весь фотосинтетический аппарат находится на цитоплазматической мембране, не создает углублений). У них и у растений также имеются различия в свитлозбираючому комплексе (см. ниже) и в составе пигментов.
3. Световая (светозависимый) стадия
В ходе световой стадии фотосинтеза образуются высокоэнергетические продукты: аденозинтрифосфат, который служит в клетке источником энергии, и НАДФН, который используется как восстановитель. Как побочный продукт выделяется кислород.
3.1. Фотохимическая суть процесса

Двух стадийный фотосинтез ("Z-схема")
Хлорофилл имеет два уровня возбуждения (с этим связано наличие двух максимумов на его спектре поглощения): первый связан с переходом на более высокий энергетический уровень электрона системы связанных двойных связей, второй - с возбуждением неспаренных электронов азота и кислорода порфиринового ядра. При неизменном спине электрона формируются синглетный первый и второй возбужденное состояние, при измененном - триплетные первый и второй.
Второй возбужденное состояние наиболее высокоэнергетическое, нестабильный, и хлорофилл за 10 -12 с переходит с него на первый, с потерей 100 кДж / моль энергии только в виде тепла. С первого синглетного и триплетного состояний молекула может переходить в основной с выделением энергии в виде света ( флуоресценция) или тепла, с переносом энергии на другую молекулу, или, поскольку электрон на высоком энергетическом уровне слабо связан с ядром, с переносом электрона на другое соединение.
Вторая возможность реализуется в светособирающих комплексах, первая - в реакционных центрах, где переведен в возбужденное состояние при поглощении кванта света хлорофилл становится донором электрона (восстановителем) и передает его на первичный акцептор. Чтобы предотвратить возвращение электрона на положительно заряженный хлорофилл, первичный акцептор передает его вторичному. Кроме того, время жизни полученных соединений выше, чем в возбужденной молекулы хлорофилла. Происходит стабилизация энергии и разделения зарядов. Для дальнейшей стабилизации вторичный донор электронов окисляет положительно заряженный хлорофилл, первичным донором же в случае оксигенной фотосинтеза является вода.
Проблемой, с которой сталкиваются при этом организмы с оксигенной типом фотосинтеза, является различие окислительно-восстановительных потенциалов воды (E 0 = +0,82 В) и НАДФ + (E 0 = -0,32 В). Хлорофилл при этом должен иметь в основном состоянии потенциал, больше +0,82, в чтобы окиснюваты воду, но при этом иметь в возбужденном состоянии потенциал меньший, чем -0,32, В чтобы восстанавливать НАДФ +. Одна молекула хлорофилла не может отвечать обоим требованиям. Поэтому сформировались две фотосистемы и для полного проведения процесса необходимо два кванта света и два хлорофиллы различных типов.
3.2. Светособирающего комплексы

Хлорофилл выполняет две функции: поглощение и передачу энергии. Более 90% всего хлорофилла хлоропластов входит в состав светособирающих комплексов (СЗК), которые выполняют роль антенны, которая передает энергию к реакционному центру фотосистем I или II. Кроме хлорофилла, в СЗК является каротиноиды, а в некоторых водорослей и цианобактерий - фикобилины, роль которых заключается в поглощении света тех длин волн, которые хлорофилл поглощает сравнительно слабо.
Передача энергии идет резонансным путем (механизм Ферстера) и занимает для одной пары молекул 10 -10 -10 -12 сек., Расстояние, на которое осуществляется перенос, составляет около 1 нм. Передача сопровождается некоторыми потерями энергии (10% от хлорофилла а к хлорофиллу b, 60% каротиноидов в хлорофилла), из-за чего возможна только от пигмента с максимумом поглощения при меньшей длине волны к пигмента с большей длиной волны в максимуме поглощения. Именно в таком порядке взаимно локализуются пигменты ССК, причем наиболее длинноволновые хлорофиллы находятся в реакционных центрах. Обратный переход энергии невозможен.
СЗК растений расположен в мембранах тилакоидов, у цианобактерий основная его часть вынесена за пределы мембран в прикрепленные к ним фикобилисомы - палочковидные полипептидные -пигментные комплексы, в которых находятся различные фикобилины: на периферии фикоеритрины (с максимумом поглощения при 495-565 нм), за ними фикоцианины (550-615 нм) и аллофикоцианины (610-670 нм), последовательно передают энергию на хлорофилл а (680-700 нм) реакционного центра.
3.3. Основные компоненты электронно-транспортной цепочки
3.3.1. Фотосистема II
Фотосистема - совокупность СЗК, фотохимического реакционного центра и переносчиков электрона. Светособирающего комплекс II содержит 200 молекул хлорофилла а, 100 молекул хлорофилла b, 50 молекул каротиноидов и 2 молекулы феофитину. Реакционный центр фотосистемы II является пигмент-белковым комплексом, расположенным в тилакоидних мембранах и окруженным СЗК. В нем находится димер хлорофилла а с максимумом поглощения при 680 нм (П680). На него в конце концов передается энергия кванта света с СЗК, в результате чего один из электронов переходит в высший энергетическое состояние, связь его с ядром ослабляется и возбужденная молекула П680 становится сильным восстановителем (E 0 = -0,7 В).
П680 восстанавливает феофитин, в дальнейшем электрон переносится на хиноны, входящие в состав ФС II, и далее на пластохинона, транспортируется в восстановленной форме в b 6 f комплекса. Одна молекула пластохинона переносит 2 электрона и 2 протоны, которые берутся из стромы. Заполнение электронной вакансии в молекуле П680 происходит за счет воды. В состав ФС II входит водоокислюючий комплекс, содержащий в активном центре ионы марганца в количестве 4 штук. Для образования одной молекулы кислорода требуется две молекулы воды, которые дают 4 электрона. Поэтому процесс проводится в 4 такта, и для его полного осуществления требуется 4 кванты света. Комплекс находится со стороны внутришньотилакоидного пространства и полученные 4 протона выбрасываются в него.
Таким образом, суммарный результат работы ФС II - это окисление 2 молекул воды с помощью 4 квантов света с образованием 4 протонов в внутришньотилакоидному пространстве и 2 восстановленных пластохинонив в мембране.
3.3.2. b 6 f или b / f-комплекс
b 6 f комплекс является насосом перекачивает протоны с стромы в внутришньотилакоидний пространство и создает градиент их концентрации за счет энергии, выделяющейся в окислительно-восстановительных реакциях электронно-транспортной цепочки. 2 пластохиноны обеспечивают перекачку 4 протонов. В дальнейшем трансмембранный протонный градиент ( pH стромы, близкий к 8, внутришньотилакоидного пространства - 5) используется для синтеза АТФ трансмембранным ферментом АТФ-синтазы.
3.3.3. Фотосистемы I
Светособирающего комплекс I содержит примерно 200 молекул хлорофилла.
В реакционном центре первой фотосистемы находится димер хлорофилла а с максимумом поглощения при 700 нм (П700). После возбуждения квантом света он восстанавливает первичный акцептор - хлорофилл а тот - вторичный ( витамин K 1), после чего электрон передается на филлохинон, от него на ферридоксинов, который и восстанавливает НАДФ с помощью фермента ферридоксинов-НАДФ-редуктазы. Белок пластоцианин, окисленный в b 6 f комплексе, транспортируется к реакционному центра первой фотосистемы со стороны внутришньотилакоидного пространства и восстанавливает окисленный П700.
3.4. Циклический и псевдоцикличний транспорт электрона
Помимо полного нециклического пути электрона, описанного выше, обнаружено циклический и псевдоцикличний. Суть циклического пути заключается в том, что ферридоксинов вместо НАДФ восстанавливает пластохинон, который переносит его назад на b 6 f комплекс. В результате образуется больше протонный градиент и больше АТФ, но не возникает НАДФН. При псевдоцикличному пути ферридоксинов восстанавливает кислород, который в дальнейшем превращается в воду и может быть использован в фотосистеме II. При этом также не образуется НАДФН.
4. Темновая стадия

В темновой стадии с участием АТФ и НАДФН происходит восстановление CO 2 к глюкозе. Хоча світло не потрібне для здійснення даного процесу, воно бере участь у його регуляції.
4.1. С 3 - фотосинтез, цикл Кальвіна
Цикл Кальвіна, або відновлювальний пентозо-фосфатний цикл, складається з трьох стадій:
На першій стадії до рибулезо-1,5-біфосфату приєднується CO 2 під дією ферменту рибулезобифосфат-карбоксилаза/оксигенази (Rubisco). Цей білок складає основну фракцію білків хлоропласту і, ймовірно, найбільш поширений фермент в природі. В результаті утворюється проміжне нестійке з'єднання, що розпадається на дві молекули 3-фосфогліцеринової кислоти (ФГК). У другій стадії ФГК в два етапи відновлюється. Спочатку вона фосфорилюєтся АТФ під дією фосфорогліцерокінази, потім НАДФН при дії тріозофосфатдегідрогенази, її карбоксильна група окислюється до альдегідної і вона стає вуглеводом (ФГА).
У третій стадії беруть участь 5 молекул ФГА, які через утворення 4-, 5-, 6- і 7-вуглецевих зв'язків об'єднуються в 3 5-вуглецевих рибулезо-1,5-біфосфати, для чого необхідні 3 АТФ. Нарешті, дві ФГА необхідні для синтезу глюкози. Для утворення однієї молекули глюкози потрібно 6 обертів циклу, 6 CO 2, 12 НАДФН і 18 АТФ.
4.2. С 4 - фотосинтез, цикл Хетча-Слека-Карпілова
При низькій концентрації розчиненого в стромі CO 2 Rubisco каталізує реакцію окислення рибулезо-1,5-біфосфату і його розпад на 3-фосфогліцеринову кислоту і фосфогліколеву кислоту, яка вимушено використовується в процесі фотодихання. Для збільшення концентрації CO 2 рослини С 4 типу змінили анатомію листка. Цикл Кальвіна у них локалізується в клітинах обкладання провідного пучка, в клітинах мезофілу ж під дією ФЕП-карбоксилази фосфоенолпіруват карбоксилуєтся з утворенням щавелеоцетової кислоти, яка перетворюється на малат або аспартат і транспортується в клітини обкладання, де декарбоксилуєтся з утворенням пірувату, що повертається в кліини мезофіла.
С 4 фотосинтез практично не супроводжується втратами рибулезо-1,5-біфосфату з циклу Кальвіна, тому ефективніший. Проте він вимагає не 18, а 30 АТФ на синтез 1 молекули глюкози. Це виправдовує себе в тропіках, де жаркий клімат вимагає тримати продихи закритими, що перешкоджає надходженню CO 2 в листок, а також при рудеральній життєвій стратегії.
4.3. САМ фотосинтез
При CAM (Crassulaceae acid metabolism) фотосинтезі відбувається розділення асиміляції CO 2 і циклу Кальвіна не в просторі, як у С 4, а в часі. Вночі у вакуолях клітин по аналогічному описаному вище механізму при відкритих продихах накопичується малат, вдень при закритих продихах йде цикл Кальвіна. Цей механізм дозволяє максимально економити воду, проте поступається в ефективності і С 4, і С 3. Він виправданий при стрестолерантній життєвій стратегії.
5. Аноксигенний фотосинтез
Аноксигенний фотосинтез властивий деяким бактеріям та археям (наприклад, пурпурним, деяким зеленим бактеріям та геліобактеріям тощо). Ці організми не використовують води у якості відновника, тому кисень (O 2) не є побічним продуктом синтезу. Замість води використовуються як сірководень (H 2 S) (дивіться малюнок 2) або йони двовалентного заліза (Fe ++), унаслідок чого на виході виникають елементарна сірка (S) і тривалентні іони заліза (Fe +++) відповідно, або молекулярний водень (H 2). Наприклад, фотосинтез з використанням H 2 S у якості відновника проходить такі стадії:
CO 2 + 2 H 2 S → (CH 2 O) + 2 S + H 2 OТут першим продуктом фотосинтезу виступає фіктивна хімічна сполука CH 2 O.
6. Значення фотосинтезу
Фотосинтез є основним джерелом біологічної енергії,$фотосинтезуючі автотрофи використовують її для утворення органічних речовин з неорганічних, гетеротрофи існують за рахунок енергії хімічних зв'язків, запасеної автотрофами, вивільняючи її в процесах аеробного та анаеробного дихання. Енергія, отримувана людством при спалюванні викопного палива (вугілля, нафта, природний газ, торф), також є запасеною в процесі фотосинтезу.
Фотосинтез является главным методом привлечения неорганического углерода в биологический цикл. Весь кислород атмосферы биогенного происхождения и является побочным продуктом фотосинтеза. Формирование окислительной атмосферы полностью изменило состояние земной поверхности, сделало возможным появление дыхания, а в дальнейшем, после образования озонового слоя, позволило жизни выйти на сушу.
7. Изучение фотосинтеза

Первые опыты по фотосинтезу были проведены Джозефом Пристли в 1770-1780-х годах, когда он обратил внимание на "порчи" воздуха в герметичном сосуде свечой (воздух переставал быть способен поддерживать горение, помещенные в него животные задыхались), горящего и "исправления" его растениями. Пристли сделал вывод, что растения выделяют кислород, необходимый для дыхания и горения, однако не отметил, что для этого растениям нужен свет. Это показал вскоре Ян Ингенхауз.
Позже было установлено, что, помимо выделения кислорода, растения поглощают углекислый газ и при участии воды синтезируют на свету органическое вещество. В 1842 Роберт Майер на основании закона сохранения энергии постулировал, что растения превращают энергию солнечного света в энергию химических связей. В 1877 Вильгельм Пфеффер назвал этот процесс фотосинтезом.
Хлорофилл был впервые выделен в 1818 году П. Ж. Пелетье и Жозефом Каванту. Разделить пигменты и изучить их отдельно удалось М. С. Цвету с помощью созданного им метода хроматографии. Спектры поглощения хлорофилла были изучены К. А. Тимирязева, он же, развивая положения Майера, показал, что именно поглощенные позволяют повысить энергию системы, создав вместо слабых связей С-О и О-Н высокоэнергетические С-С (до этого считалось, что в фотосинтезе используются желтые лучи не поглощаются пигментами листа). Сделано это было благодаря созданному им методу учета фотосинтеза по поглощенному CO 2, в ходе экспериментов по освещению растения светом разных длин волн (разного цвета) оказалось, что интенсивность фотосинтеза совпадает со спектром поглощения хлорофилла.
Окислительно-восстановительную сущность фотосинтеза (как оксигенной, так и аноксигенным) постулировал Корнелис ван Ниль. Это означало, что кислород в фотосинтезе образуется полностью из воды, экспериментально подтвердил в 1941 А. П. Виноградов в опытах с изотопной меткой. В 1937 Роберт Хилл установил, что процесс окисления воды (и выделение кислорода), а также ассимиляции CO 2, можно разъединить. В 1954 - 1958 Дэниэл И. Арнон установил механизм световых стадий фотосинтеза, а суть процесса ассимиляции CO 2 была раскрыта Мельвином Кальвином с использованием изотопов углерода в конце 1940-х, за эту работу в 1961 ему была присуждена Нобелевская премия.
В 1955 была выделена и очищена Rubisco. С 4 фотосинтез был описан Ю. С. Карпиловим в 1960 и М. Д. Хетч и К. Р. Слэк в 1966.
Примечания
- Никифорович А.А. Никифорович А.А. О Фотосинтез растений. - М.: Правда, 1948. - 31 с
Литература
nado.znate.ru
Фотосинтез
 Здравствуйте, уважаемые читатели блога репетитора биологии по Скайпу biorepet-ufa.ru.
Здравствуйте, уважаемые читатели блога репетитора биологии по Скайпу biorepet-ufa.ru.
Прошла уже неделя как я начал заполнять странички своего блога о самых сокровенный тайнах живого и вижу, что кто-то заходит их читать.
Это вселяет надежду на продолжение нашего диалога (пусть пока и монолога, но мысленно я, репетитор по биологии, уже давно общаюсь с вами).
Еще лежит снег, а Солнце сегодня припекает даже не по-весеннему — загорать можно!
Радуюсь, видя как радуются Солнцу цветы на подоконнике, оживающие после «зимней спячки». Так о чем же нам с вами сегодня говорить, как не о ФОТОСИНТЕЗЕ — этом проводнике между Солнцем и жизнью на нашей матушке Земле.
Фотосинтез как один из основных глобальных процессов в биосфере Земли
Все гетеротрофные организмы на Земле нуждаются в углероде, находящемся в органической форме. Только автотрофные организмы (растения и некоторые бактерии) способны ассимилировать углекислоту воздуха (неорганический углерод) и создавать органические вещества в процессе фотосинтеза.
Ошибки, допускаемые при сдаче ГИА и ЕГЭ
В школе, начиная с уроков по естествознанию, затем в курсе ботаники, а затем и в курсе общей биологии, изучению фотосинтеза уделяется значительное место.Но, как репетитор по биологии, вынужден обратить ваше внимание, что на экзамене именно по этой теме большинство из вас допускают много ошибок. И основная ошибка, как выяснилось, заключается часто в непонимании самой сути этого жизненно важного процесса.
СкажИте, как вы считаете, к какому типу клеточных реакций следует отнести фотосинтез? К энергетическому или пластическому обмену?
Полагаю, что и вы сейчас немного задумались. Кто-то скажет, конечно к энергетическому. Энергия Солнца переходит в энергию химических связей и превращается в хлоропластах клеток в универсальный источник клеточной энергии АТФ. И они будут правы…
Другие скажут, конечно же к пластическому, потому что при фотосинтезе создаются органические вещества, идет синтез веществ. И они тоже будут правы.
Что же выходит, процесс один, а правды две?
Дело в том, что сам термин фотосинтез уже включает в свое название оба эти процесса. Фото — перевод солнечной энергии растениями в энергию АТФ.
Это так называемое фотосинтетическое фосфорилирование (фотофосфорилирование), осуществляемое в хлоропластах. Хлоропласты — это такое расчудесное чудо! Только они на Земле способны работать на нескончаемом энергетическом топливе — энергии Солнца!!!
А гетеротрофы, помните, чтобы образовать АТФ должны обязательно, поглотив готовые органические вещества извне, произвести их окисление в митохондриях своих клеток, то есть они способны лишь к окислительному фосфорилированию.
Так откуда же берутся изначально для гетеротрофов готовые органические вещества? Они создаются растениями на втором этапе фотосинтеза — синтеза органических веществ.
Где создаются растениями органические вещества — спрашивает вас репетитор по биологии?
Да все в тех же хлоропластах! Хлоропласты не только преобразователи солнечной энергии, но и уловители неорганического углерода СО2 воздуха в цикле Кальвина.. В них же и происходит образование органических веществ.
Дорогие мои, если я вас сейчас своими восторгами по поводу наличия хлоропластов у растений только окончательно запутал, то вернитесь к началу странички, прежде чем решите освежить в памяти саму схему процесса, приведенную ниже.
Эта схема без какой-либо сложной «начинки». Она отражает лишь самое основное, саму суть процесса: что, где, когда.
 ФОТОСИНТЕЗ (хлоропласты)
ФОТОСИНТЕЗ (хлоропласты)
I.Световая стадия (на мембранах тилакоидов гран хлоропластов)
1) Создание отрицательно заряженного электрического поля снаружи тилакоидов за счет электронов хлорофилла, перешедших на более высокий энергетический уровень под действием квантов света.
2) Создание положительно заряженного электрического поля внутри тилакоидов за счет протонов водорода Н+, образующихся при фотолизе воды (под действием квантов света):Н2О = 4Н+ + 4е- + О2 (побочный продукт фотолиза воды)
3) Образуются высокоэнергетические молекулы АТФ и НАДФ*Н (НАДФ*Н выступает переносчиком водорода, который участвует в восстановлении СО2 до глюкозы в темновой фазе фотосинтеза).
II.Темновая стадия (в строме хлоропластов)
Это реакции восстановления СО2 в цикле КАЛЬВИНА с образованием углеводов (глюкозы)
___________________________________________________________________________________
Схема суммарного процесс фотосинтеза двух фаз:
 ___________________________________________________________________________________
___________________________________________________________________________________
Итак, световая стадия процесса так названа, потому что для нее солнечная энергия обязательна. Причем, убедительно прошу как репетитор по биологии, обратить ваше внимание на то, что энергия света в хлоропластах служит двум целям:
* Под действием света происходит возбуждение электронов хлорофилла — это пигментный белок хлоропластов и
* о чем чаще всего и забывают, в световую фазу происходит еще и фотолиз воды.
Таким образом, для первой световой стадии фотосинтеза необходимы солнечный свет, хлорофилл, вода. Основным итогом этой стадии является образование АТФ и НАДФ*Н.
Побочным продуктом реакции фотолиза воды является выделившийся молекулярный кислород О2. Хорошенький такой «побочный продуктик». Благодаря этому побочному продукту атмосфера нашей планеты состоит на 20% из кислорода. Именно кислород, образующийся при фотосинтезе растениями и обеспечивает процессы дыхания всех аэробных организмов на Земле.……………….
Обратите внимание, что вторая темновая стадия фотосинтеза не обязательно протекает только в темноте ночью. Она проходит и днем, но вот свет для второй стадии уже не нужен. Необходимы в наличии в строме хлоропластов для осуществления цикла Кальвина молекулы углекислоты СО2, АТФ и НАДФ*Н.
Конечно же основным итогом фотосинтеза является образование органических веществ (сначала моносахаридов — глюкозы, затем и полисахаридов — крахмала).………………
Вот и все. Думаю, что теперь никто из вас никогда не скажет, что для фотосинтеза нужен кислород и что при фотосинтезе выделяется СО2.
***************************************
У кого есть вопросы по статье к репетитору биологии по Скайпу, замечания, пожелания — прошу в комментарии.
www.biorepet-ufa.ru









