13. Репродуктивные органы цветковых растений. Репродуктивные органы растений
13. Репродуктивные органы цветковых растений
Покрытосеменные, или цветковые растения являются наиболее крупным отделом растительного царства, насчитывающим более 350 семейств, 13 тысяч родов и до 240 тысяч видов. В настоящее время покрытосеменные распространены по всему Земному шару – от тропиков до приполярных пустынь. Автотрофные цветковые растения – важнейший компонент биосферы, от которого зависят биосферные процессы обмена веществ и трансформации энергии, газовый состав атмосферы, климат, водный режим суши, характер процессов почвообразования. На суше цветковые часто доминируют в растительном покрове, только в немногих сообществах и типах растительности они уступают господство по общей массе другим группам растений – в хвойных лесах голосеменным, в некоторых типах тундр и на сфагновых болотах – мохообразным.
Покрытосеменные определяют самую возможность существования большинства наземных животных. Они формируют среду обитания для разнообразных консументов – потребителей первичного органического вещества. Различные группы гетеротрофов связаны с ними трофическими, консортивными связями. В процессе эволюции многие группы гетеротрофов смогли возникнуть только тогда, когда Землю завоевали покрытосеменные и образуемые ими сообщества. Для многих членистоногих и позвоночных характерна сопряженная с цветковыми эволюция. Человек как биологический вид тоже возник и эволюционировал после появления покрытосеменных, отличающихся высоким уровнем продуктивности и биохимическим разнообразием.
Покрытосеменные растения отличаются необычайным полиморфизмом, эволюционной пластичностью и могут существовать в разнообразных условиях обитания. Это единственная группа среди высших растений, представители которой сумели вторично освоить морскую среду, где стали доминировать на огромных акваториях.
Разнообразие размеров и жизненных форм цветковых растений, их приспособлений к конкретным условиям обитания велико. Представитель семейства рясковых – ряска малая (Lemna minor) не превышает 3-5 мм в длину и похожа на свободноплавающие слоевищные растения – мхи или водоросли (рис. 13 .1).

Рис. 13.1. Ряска малая (Lemna minor) – миниатюрное цветковое растение.
С другой стороны, гигантские эвкалипты (виды рода Eucalyptus из семейства миртовые Myrtaceae) вырастают высотой до 100 м, длина побегов тропических лиан – ротанговых пальм измеряется сотнями метров. Среди покрытосеменных преобладают автотрофы, но встречаются полупаразиты, паразиты и сапротрофы, полностью утратившие хлорофилл и способность к фотосинтезу. Среди покрытосеменных имеются также эпифиты и эпифиллы (эпифиты, живущие на листьях других растений), деревья, кустарники, кустарнички, полукустарники и травы (многолетники и однолетники), плотоядные растения с их разнообразными приспособлениями для улавливания и переваривания мелких животных (рис. 13.2).

Рис. 13.2. Насекомоядное цветковое растение росянка круглолистная (Drosera rotundifolia).
Отмечены разнообразные приспособления видов цветковых растений к опылению строго определенными агентами, к распространению плодов и семян, к перенесению неблагоприятных климатических периодов. Эти особенности дают цветковым растениям возможность полностью реализовать свой эволюционно-адаптационный потенциал. Ни одна другая группа растений не смогла выработать такого разнообразия приспособлений к определенным факторам среды.
Существенным признаком покрытосеменных считают цветок – отсюда одно из названий отдела – Anthophyta (antos – цветок, phyton – растение). Происхождение цветка как особого органа – это одна из ключевых проблем морфологии и филогении покрытосеменных. Дискуссии по этому вопросу, начавшиеся еще в XVIII в., продолжаются и в настоящее время.
Типичный цветок определяют как видоизмененный побег, обеспечивающий функции спороношения, развития гаметофитов, половое размножение, образование семян в плоде. В истории ботаники отмечено, что в поисках архетипа цветка за первичный примитивный цветок различные ученые принимали разные структуры.
Сторонники эвантовой теории считают первичным крупный обоеполый цветок, опыляемый насекомыми, с большим и часто неопределенным числом свободных частей, и тогда цветки однополые, олигомерные, голые рассматриваются как результат более поздних этапов редукционной эволюции.
Сторонники псевдантовой теории полагают, что эволюционные трансформации выражались в постепенном усложнении структуры цветка от однополых, анемофильных, с небольшим числом частей к обоеполым, полимерным, энтомофильным, с хорошо развитым околоцветником.
Следует учитывать, что в процессе эволюционного возникновения покрытосеменные сначала приобрели комплекс важнейших общих признаков (в первую очередь – уникальные особенности жизненного цикла), а уже потом выработали разные цветки. При этом в каждой эволюционной ветви покрытосеменных были осуществлены различные морфогенетические решения.
Существенными частями цветка являются андроцей, состоящий из одной, нескольких или многих тычинок (микроспорофиллов) и гинецей, который состоит из одного, нескольких или многих плодолистиков, или карпелл (мегаспорофиллов).
Плодолистики (карпеллы) могут быть свободными, и тогда каждый из них, срастаясь краями, образует пестик (один или несколько пестиков в цветке – по числу плодолистиков). При срастании нескольких плодолистиков формируется ценокарпный гинецей, пестик в нем всегда один.
Важнейшая часть пестика – завязь. Наличие завязи (замкнутой полости), в которой развиваются семязачатки (в отличие от голосеменных, у которых семязачатки расположены на поверхности открытых мегаспорофиллов), обычно считают важнейшим признаком покрытосеменных, откуда и другое название отдела – Angiospermae (от греческого angeion – сосуд, вместилище, sperma – семя). Полагают, что завязь обеспечивает лучшую защиту семяпочек (семязачатков). В полости завязи семязачатки оказываются в условиях влажной камеры, оптимальных для их развития. Благодаря образованию завязи расширяются и обогащаются возможности распространения растений: из цветка формируется плод, при диссеминации расселяются не только отдельные семена, но и целые плоды, у которых возникают разнообразные приспособления к распространению широким кругом агентов.
У цветковых растений и некоторых голосеменных спермин (мужские неподвижные половые клетки) доставляются к яйцеклетке с помощью особой клетки – пыльцевой трубки, которая обеспечивает высокую точность попадания спермия в зародышевый мешок (то есть к женскому гаметофиту с яйцеклеткой). Семязачатки у покрытосеменных растений скрыты в завязи пестика, поэтому необходимо приспособление для улавливания пыльцы. Таким аппаратом у покрытосеменных служит рыльце на верхушке столбика пестика (стилодия). Наличие рыльца – важная особенность покрытосеменных растений.
Архегониальные растения (папоротникообразные, голосеменные) характеризуются тенденцией к возникновению разноспоровости, которая сопровождается редукцией полового поколения в жизненном цикле. Покрытосеменные, у которых редуцированы гаметангии, представляют собой завершающий этап на этом пути. Предельное сокращение процессов микро- и мегаспорогенеза и ускорение развития гаметофитов, связанное с их редукцией, были, вероятно, важными эволюционными факторами. Женский гаметофит цветковых растений представлен зародышевым мешком, состоящим всего из 7 клеток, а мужской гаметофит представлен пыльцевым зерном (пылинкой), в которой только 2 клетки – сифоногенная и спермагенная (дающая 2 спермия).
Уникальной особенностью покрытосеменных является двойное оплодотворение, в котором участвуют 4 клетки: два спермия, яйцеклетка и центральная клетка зародышевого мешка. В ходе этого процесса один из cnepмиев оплодотворяет яйцеклетку, образуя диплоидную зиготу, которая впоследствии развивается в зародыш семени. Второй спермий сливается с диплоидным ядром центральной клетки зародышевого мешка, образуя триплоидную зиготу. Эта зигота впоследствии дает начало триплоидному эндосперму семени. Общий для всех покрытосеменных уникальный процесс двойного оплодотворения указывает на их эволюционное единство и монофилетическое становление.
Одно из следствий двойного оплодотворения – формирование питательной ткани – полиплоидного эндосперма (в отличие от гаплоидного эндосперма голосеменных, представляющего остаток женского гаметофита). Биологическое значение полиплоидного состояния запасающей ткани связано с тем, что оно обеспечивает более благоприятные условия для развития зародыша и поэтому могло иметь важную эволюционную роль.
На тканевом уровне подавляющее большинство покрытосеменных характеризуется наличием в ксилеме сосудов, тогда как почти у всех голосеменных проводящие элементы древесины представлены только трахеидами. Только у немногих цветковых проводящая система ксилемы гомоксильная, бессосудистая, унаследовавшая примитивность от предковых голосеменных форм.
Паренхиматизация цветковых позволила выработать специальные запасающие ткани и травянистые жизненные формы. В отличие от древесных растений, у трав возможно сокращение онтогенеза и ускорение темпов эволюции; травянистые растения способны существовать в таких крайних условиях среды, в которых деревья и кустарники жить не могут.
Остается нерешенной проблема геологического возраста цветковых растений. Считалось, что покрытосеменные растения возникли раньше мелового периода – в триасе или палеозое (255-210 млн. лет назад). Предполагалось, что первоначально небольшая, эта группа существовала в таких условиях, которые не благоприятствовали фоссилизации их остатков, например, в горных районах или в пустынях. Позже, в меловой период (145-63 млн. лет назад), покрытосеменные дали мощную вспышку развития и сформировали свое современное разнообразие.
Этому взгляду противоречат объективные данные палеоботаники. Многочисленные указания на нахождение ископаемых остатков покрытосеменных в домеловых осадочных породах в настоящее время достоверно не доказаны. Имеются косвенные указания на возможность существования покрытосеменных в юре (210-145 млн. лет назад). Например, с этого времени известны некоторые формы насекомых, современные родственники которых настолько тесно связаны с цветковыми растениями, что допустить их независимое от последних существование трудно.
Древнейшие, достоверно определяемые ископаемые остатки цветковых растений в виде фоссилизованных листьев, цветков, плодов, древесины и пыльцевых зерен встречаются лишь с начала мелового периода (135 млн. лет назад). В этих отложениях они очень немногочисленны и фрагментарны. Еще недавно считалось, что к середине мелового периода (120 млн. лет назад) покрытосеменные появились в массе и выступали как «победители в борьбе за существование» с голосеменными и папоротникообразными. Сейчас палеоботаники отодвигают начало расцвета покрытосеменных на более позднее время – конец мелового периода (80-63 млн. лет назад).
Предполагают, что родиной цветковых могли быть тропические или субтропические регионы Земли. Исследователи считают, что это были районы тропической Юго-Восточной и Восточной Азии и Меланезия. В этом районе сосредоточены древние и считающиеся примитивными таксоны цветковых. Вероятно, что отсюда шла миграция древнейших цветковых в другие области Земли. Еще одна точка зрения заключается в признании центром происхождения покрытосеменны[ древнего континента Южного полушария – Гондваны. Считается, что цветковые могли возникнуть и расселиться по этому обширному континенту до начала раскола Гондваны на современные континенты Южного полушария.
К началу XX в. сложились две противостоящие друг другу системы взглядов на происхождение покрытосеменных от более примитивных предковых форм растений. Одна из них, связанная с именами австрийского ботаника Р. Веттштейна и классика немецкой ботаники А. Энглера, была основана на представлении о происхождении цветковых от эфедроподобных или гнетоподобных голосеменных предков. Р.Веттштейном была разработана оригинальная концепция происхождения цветка – так называемая псевдантовая гипотеза (от греческого «псевдантиум» – ложный цветок). Согласно этой гипотезе цветок представляет собой собрание мелких разнополых стробилов, претерпевших в процессе эволюции ряд преобразований: редукцию, сближение и срастание. На основе этой гипотезы наиболее архаичными следует считать семейства с тычиночными и пестичными невзрачными ветроопыляемыми цветками (ивовые, казуариновые).
Другая концепция восходит к построениям И.В.Гёте и А.П.Декандоля, то есть к концу XVIII – началу XIX веков. В эпоху филогенетической систематики это направление ознаменовалось появлением стробилярной (от греческого «стробилюс» – шишка хвойного)гипотезы, которую сформулировали Е.Арбер и Дж. Паркин . Другое название этой гипотезы – эвантиевая (от греческого «эвантиум» – настоящий цветок). Согласно стробилярной гипотезе наиболее вероятными предками покрытосеменных были мезозойские голосеменные растения – беннеттиты. С этих позиций цветок – это метаморфизированный (видоизмененный) укороченный спороносный побег, первоначально напоминавший шишку голосеменных. Мегаспорофиллы, несущие семязачатки, в процессе метаморфоза превратились в плодолистики (карпеллы), а несущие пыльники микроспорофиллы – в тычинки. Эти видоизменения многие исследователи связывают с приспособлением покрытосеменных к насекомоопылению (энтомофилии). Примитивные цветки имели значительные размеры и выпуклое цветоложе, на котором располагались многочисленные микро- и мегаспорофиллы. Микроспорофиллы эволюционировали в тычинки, а мегаспорофиллы – в карпеллы (плодолистики), которые образовали пестики. Цветки приобрели строение, сходное с тем, которое наблюдается у современных магнолиевых, лютиковых, кувшинковых. Поэтому перечисленные семейства обычно располагаются в основании системы покрытосеменных как наиболее древние и архаичные. Дальнейшая эволюция цветка в пределах покрытосеменных имела редукционный характер.
Обе эти гипотезы являются фолиарными (от латинского «фолиум» – лист), так как исходят из представлений, что цветки образовались из листостебельных спороносных побегов. Фолиарным гипотезам противопоставляются различные теломные (от греческого «телома» – конечные побеги) гипотезы (например, теория гонофилла Р. Мелвилла), широко распространенные в 60-70-е годы ХХ в. Согласно этим гипотезам, все части цветка могут быть выведены из теломов – осевых структур, свойственных ископаемым растения – риниевым.
Палеоботаника в последние десятилетия дала много нового материала о древнейших, ранне- и среднемеловых покрытосеменных. Эти данные позволяют сделать несколько заключений. Во-первых, уже самые ранние из известных нам цветковых отличаются большим полиморфизмом. Очень разнообразны пыльцевые зерна по форме, размерам, скульптуре поверхности, характеру поровости, числу и строению апертур. Они обнаруживают сходство с пыльцевыми зернами порядков и семейств современных растений. Листья меловых цветковых также весьма разнообразны и напоминают современные фикусы, протейные, дубы, акации, аралию, подорожник, известны и специализированные щитовидные листья.
Большим полиморфизмом отличаются и органы размножения – цветки и плоды. У меловых цветковых известны как одиночные и относительно крупные цветки, так и мелкие, собранные в разнообразно устроенные соцветия с простым или двойным околоцветником, с апокарпным или ценокарпным гинецеем, с верхней или нижней завязью, с разным числом тычинок и плодолистиков. Наряду со вскрывающимися плодами типа листовок обнаружены и не-вскрывающиеся плоды. Вместе с древесными формами в меловую эпоху существовали многочисленные травянистые покрытосеменные.
Во-вторых, если раньше часто утверждали, будто в меловую эпоху покрытосеменные были представлены современными таксонами, то сейчас большинство исследователей считают, что раннемеловые Angiospermae принадлежали к вымершим порядкам и семействам. Этот факт важен как доказательство того, покрытосеменные не оставались неизменными с момента зарождения.
В целом сейчас большинство специалистов считают, что предков Angiospermae нужно искать среди каких-то древних и малоспециализированных голосеменных растений. Не оставлена идея их генетической связи с беннеттитовыми. Допускают, что магнолиевые (как наиболее примитивные цветковые) лишь сохранили наибольшее число архаичных признаков, но сами, как и все прочие таксоны цветковых, произошли от какой-то вымершей, еще более древней гипотетической группы покрытосеменных.
В настоящем учебном пособии принята система цветковых растений, разработанная А.Л.Тахтаджяном в 1987 г. В данной системе древнейшей группой покрытосеменных считается порядок магнолиевых (Magnoliales), от предков которых, вероятно, произошли современные покрытосеменные. В этом смысле приведенная система монофилетична ( рис. 13.3).

Рис. 13.3. Система покрытосеменных растений (по А.Л.Тахтаджяну).
Цветок – особый побег покрытосеменных растений, ответственный за формирование спор двух типов (микро- и мегаспор), развитие мужского и женского гаметофитов, образование гамет (яйцеклеток и спермиев), опыление, оплодотворение и формирование семян. Цветок в процессе своего развития превращается в плод с семенами. Во многих отношениях он уникален и столь характерен для этой группы, что покрытосеменные нередко называют цветковыми растениями. Исключительная роль цветка как особой морфологической структуры связана с тем, что в нем полностью совмещены все процессы бесполого и полового размножения. В цветках осуществляются микро- и мегаспорогенез, развитие гаметофитов, микро- и мегагаметогенез, опыление, оплодотворение и формирование зародышей нового поколения спорофитов. Завершается онтогенез цветка образованием плода с семенами. Особенности строения цветка обеспечивают осуществление всех этих функций с наименьшими затратами пластических веществ и энергии.
studfiles.net
Морфология репродуктивных органов растенийПри изучении семян и плодов сорных растений необходимо отчетливо представить структуру всех органов (цветка, пестика, семяпочки), в процессе развития которых они образуются. Характеристика морфологических признаков репродуктивных органов начинается с описания репродуктивных органов покрытосеменных (Angiospermae), к которым принадлежат все виды сорных растений, включенные в настоящий определитель. При изложении общих основ морфологии приводятся лишь ведущие сведения, без которых нельзя обойтись при изучении и определении зачатков растений.
Растение имеет три основных репродуктивных органа: - Цветок ( о нём мы расскажем в этой статье)
Рассмотрим первый из них - цветок.
ЦВЕТОК
Цветок является укороченным измененным репродуктивным побегом, назначение которого — генеративное размножение растений. Оно выполняется пестиком и тычинками, косвенно той же цели служит и околоцветник.
Цветок не всегда имеет все указанные части. Так, околоцветник нередко отсутствует. Цветки с тычинками и пестиками называют обоеполыми. Цветки, имеющие только тычинки или пестики — однополыми. Однополые цветки тычиночные — это мужские, пестичные — это женские.
Цветоложе — укороченная стеблевая часть цветка, на которой расположены с определенной правильностью по спирали или (чаще) сближенными кольцами (кругами) остальные части цветка.
Цветоложе может быть выпуклым, плоским и вогнутым. В зависимости от его строения, положения завязи и места формирования других частей цветка в отношении завязи цветок бывает подпестичным, около-пестичным и надпестичным.
Околоцветник состоит из листочков околоцветника и называется простым, если листочки его по форме и.окраске одинаковые, и двойным, если наружные листочки отличаются от внутренних.
Наружные листочки двойного околоцветника, обычно травянистые, называют чашелистиками, а вместе — чашечкой. Внутренние листочки двойного околоцветника, обычно яркоокрашенные, — это лепестки, а вместе они составляют венчик.
Листочки чашечки и венчика могут быть свободными или сросшимися между собой в разной степени. В первом случае чашечка свободнолистная, а венчик свободнолепестный, во втором случае чашечка спайнолистная, а венчик спайнолепестный. На спайнолистной чашечке и спайнолепестном венчике можно различить нижнюю часть — трубочку и верхнюю — зубцы, которые у спайнолепестного венчика образуют его отгиб, пограничную часть между трубкой венчика и отгибом называют зевом,
В строении цветка обычно наблюдается лучевая или двухсторонняя симметрия, реже встречается цветок асимметричный. В первом случае цветок правильный, или актиноморфный; при этом лепестки венчика, как и чашелистики, сходны между собой, в результате чего через ось такого цветка можно провести несколько плоскостей симметрии. У неправильного, или зигоморфного, цветка листочки околоцветника различаются по форме и величине, поэтому цветок может быть разделен только по одному направлению на две симметричные половины (Lamiaceae, Delphinium и др.). У асимметричного цветка невозможно провести ни одной плоскости симметрии.
Внутри цветка вслед за листочками околоцветника располагаются тычинки. Число их различно у разных видов растений —от одной до многих десятков. Совокупность всех тычинок цветка называется андроцеем.
Тычинка состоит из пыльника, связника и тычиночной нити, которая отсутствует только у сидячих пыльников. Тычинки у некоторых растений могут срастаться нитями в общую тычиночную трубку (Malvaceae) или пыльниками (Cucurbitaceae).
Пыльник обычно состоит из двух пар параллельно сложенных пыльцевых мешков, соединенных между собой связником, к которому и прикрепляется тычиночная нить. Внутри каждого пыльника находится масса пыльцы, состоящая из пыльцевых зерен.
В центральной части цветка располагается пестик, образованный одним, двумя или многими листочками — плодолистиками. Вся совокупность плодолистиков цветка называется гинецеем. Он бывает сростнолистным, или синкарпным, когда все его плодолистики, срастаясь краями, образуют один пестик, и раздельнолистным, или апокарпным, когда каждый плодолистик образует отдельный пестик. В зависимости от числа плодолистиков апокарпный гинецей может состоять из одного пестика и быть мономерным или из многих пестиков и быть полимерным.
С точки зрения эволюционной морфологии наиболее примитивным является апокарпный гинецей. Из него путем срастания плодолистиков возник синкарпный гинецей, который в процессе эволюции покрытосеменных постепенно видоизменялся.
Пестик является своеобразным замкнутым органом, присущим только покрытосеменным растениям. Плодолистики, образующие его, срастаются между собой различными способами: полностью в один пестик (от завязи до рыльца), только завязями или завязями и столбиками.
Определить число плодолистиков, участвующих в формировании пестика, можно по числу средних жилок на стенках завязи, по числу свободных столбиков и долей рыльца. Поскольку плодолистик является органом листового происхождения, пестик, образованный одним плодолистиком, представляет собой свернутый в продольную трубочку листок с соединенными в шов краями.
Обычно пестик состоит из трех частей: нижней расширенной и полой — завязи, средней суженной — столбика и верхней железистой и более или менее ворсинчатой —рыльца. Типичная форма пестика — бутылочная. Полость завязи содержит одну, две и больше семяпочек (семязачатков).
Возвышения ткани внутренней полости завязи, служащие местом прикрепления семяпочек, называются семяносцами. Положение семяпочек в завязи зависит от ее строения. В завязи, образованной только одним плодолистиком, семяпочка может прикрепляться к сросшимся краям плодолистика — брюшному шву или к средней линии плодолистика — спинному шву. И в том и в другом случае семяносец называется постенным или стенным. Однако семяпочка иногда прикрепляется к колонке, возвышающейся в центре завязи, или непосредственно ко дну завязи; тогда семяносец называется центральным или основным.
Одногнездную завязь могут иметь и пестики, состоящие из нескольких сросшихся между собой плодолистиков, причем положение семяносцев здесь также варьирует. При срастании плодолистиков только своими краями семяпочки могут располагаться по линиям сращения краев плодолистиков или по спинным швам; далее края плодолистиков могут заворачиваться и выдвигаться несколько внутрь завязи, увлекая за собой и семяпочки. Семяносцы таких завязей называются постенными. Кроме того, подобно завязи пестика, образованного одним плодолистиком, одногнездная завязь пестика, возникшая из нескольких плодолистиков, может иметь также центральный семяносец.
Иногда плодолистики настолько сильно вдаются внутрь завязи, что срастаются посредине своими краями, причем соприкасающиеся друг с другом стенки их прирастают одна к другой, образуя полные перегородки. Здесь мы имеем завязь, разделенную на несколько не сообщающихся между собой гнезд—многогнездную. Такие завязи по числу гнезд называют двух-, трехгнездными. Если же перегородка неполная, то есть хотя бы где-нибудь между гнездами имеется небольшое сообщение, завязь называется одногнездной.
Только те перегородки называются истинными, которые представляют собой завернувшиеся внутрь края плодолистиков. Перегородки, образованные позднейшими выростами ткани, относят к ложным. Наличием этих выростов объясняется, почему завязь одногнездного пестика становится иногда двугнездной, у двугнездного — четырехгнездной и т. п.
Завязь по отношению к другим частям цветка может располагаться по-разному в зависимости от формы цветоложа. Если завязь находится на плоском или выпуклом цветоложе, а тычинки и околоцветник прикреплены вокруг ее основания или ниже, то завязь называется свободной или верхней. Когда завязь заключена в вогнутом цветоложе, сросшемся с ней, а тычинки и околоцветник прикреплены над ней, завязь называют приросшей или нижней. Если только нижняя часть завязи заключена в вогнутом цветоложе, а тычинки и околоцветник прикреплены вокруг ее середины, то завязь полу нижняя.
Столбиков в пестике может быть один или несколько, обычно их число соответствует числу плодолистиков. Если столбик один, то он бывает простым или делящимся на лопасти.
Столбик способствует выдвижению рыльца и проведению в завязь пыльцевых трубок. Внутри столбика имеется канал, проводящий пыльцевую трубку в глубь завязи, или особая рыхлая ткань, питающая пыльцевую трубку. В зависимости от структуры столбика выделены следующие их типы:
открытый — с каналом, выстланным обычной эпидермой; полузакрытый — с каналом, выстланным железистыми клетками; закрытый — с каналом, выполненным проводниковой тканью, по межклетникам которой продвигаются пыльцевые трубки.
Первый тип столбиков преобладает у представителей однодольных, два последних типа свойственны двудольным. Столбик обычно выходит из верхушки завязи, реже он прикрепляется сбоку (называется боковым) или у самого основания завязи (называется тогда гинобазическим).
После оплодотворения столбик обычно отпадает, реже сохраняется на плодах.
Рыльцем называется часть пестика, улавливающая пыльцу. Пыльца удерживается рыльцем благодаря железистым сосочкам, волоскам и бугоркам, заметным обычно только под лупой. Попав на рыльце, пыльца начинает прорастать и, образуя пыльцевую трубку, проникает внутрь завязи.
Если столбика нет или столбик очень короткий, то рыльце называется сидячим. Рыльца по величине и форме могут быть самыми различными. Редко рыльце настолько слабо обозначено, что представляет липкую площадку. Если у растений пыльца переносится ветром, то поверхность рыльца сильно увеличена, что облегчает улавливание пыльцы. Обычно число углов, или лопастей, рыльца равно числу плодолистиков, образующих пестик, или вдвое больше него.
Семяпочкой (семязачатком) называется зачаток будущего семени, формирующийся в завязи пестика. Место его заложения носит название семяносца или плаценты.
Вначале семяпочка появляется в виде микроскопически малого бугорка, у которого тут же начинается наружная и внутренняя дифференциация клеток. Почти одновременно с развитием центральной части семяпочки, называемой ядром или нуцеллусом (nucellus), развиваются покровы, или интегументы. Семяпочка чаще имеет два покрова — наружный и внутренний, как это наблюдается у большинства однодольных, а из двудольных — у многих раздельнолепестных. Обычно один покров свойствен семяпочке спайнолепестных двудольных. Реже в пределах одного и того же рода встречается то один, то два покрова (Delphinium), еще реже они вовсе отсутствуют (голая семяпочка). Как крайне редкое отклонение можно обнаружить семяпочки с тремя покровами, когда третий покров формируется в результате расщепления наружного покрова или представляет собой новое образование, возникшее из основания семяпочки.
На верхушке семяпочки покровы ее не смыкаются, образуя канал для проникновения пыльцевых трубок в зародышевый мешок. Этот канал носит название пыльцевхода, семявхода или микропиле (micropyle).
Существуют различные структуры микропиле: у одних — это узкие полые каналы, у других они заполнены вытянутыми клетками, принимающими форму волосков (Asteraceae), у третьих — железистыми волосками, очевидно, служащими для питания пыльцевых трубок. В тех случаях, когда покровы семяпочки полностью смыкаются или нуцеллус выдается из покрова, микропиле отсутствует.
Семяпочка прикрепляется к плаценте при помощи ножки, называемой семяножкой или фуникулусом (funiculus). Последняя бывает как сильно удлиненной, так и укороченной настолько, что кажется сидячей, например у злаковых (Роасеае), некоторых сложноцветных (Asteraceae) и др.
Семяножка служит не только для укрепления семяпочки, но и для проведения питательных веществ из завязи. Нижняя часть семяпочки в месте перехода ее в семяножку называется халазой. Место прикрепления семяпочки к семяножке, заметное и на поверхности семени, называется рубчиком.
Форма семяпочки обычно овальная или более или менее округлая. По положению семяпочки относительно семяножки различают следующие основные типы.
Прямая, или атропная, семяпочка формируется при равномерном разрастании покровов по всем направлениям, в результате чего нуцеллус составляет продолжение семяносца. Подобное строение семяпочки наблюдается нечасто, например у гречишных (Polygonaceae), у крапивы (Urtica) и др.
Обращенная, или анатропная, семяпочка, оставаясь выпрямленной, загнута на 180 ° и вследствие этого приращена боковой стороной к удлиненной семяножке. У места, где кончается это сращение, образуется вторичный рубчик, который лежит около семявхода. Первичный рубчик, оставаясь на своем месте под халазой, противолежит семявходу. Между первичным и вторичным рубчиками возникает возвышение, называемое семенным швом. Обращенная семяпочка наиболее распространена среди покрытосеменных, она встречается у однодольных и двудольных, у раздельнолепестных и спайнолепестных.
Изогнутая, или кампилотропная, семяпочка образуется в том случае, когда одна сторона ее развивается сильнее, чем другая. При этом искривляются ядро семяпочки и ее покровы, а семявход приближается к хал азе и рубчику, находясь в стороне от них. Наблюдается у крестоцветных (Brassicaceae), гвоздичных (Caryophyllaceae), пасленовых (Solanaceae) и др.
При исследовании строения семяпочки не всегда удается отнести их к одной из этих трех категорий: попадаются промежуточные формы, как полуобратная, полуизогнутая и т. п.
Число семяпочек в завязи, их форма, наличие или отсутствие покровов у семяпочек, а также структура нуцеллуса имеют важное систематическое и филогенетическое значение.
Зародышевый мешок, находящийся внутри ядра семяпочки, является наиболее сложной и вакной ее частью, поскольку в дальнейшем, после оплодотворения, в нем развивается зачаток будущего растения — зародыш. Формирование зародышевого мешка начинается с обособления одной из клеток нуцеллуса (против микропиле) в первичную археспориальную клетку.
Зрелый зародышевый мешок обычно состоит из шести клеток и двух полярных ядер. У многих видов полярные ядра сливаются еще до оплодотворения и возникает вторичное, или центральное ядро, располагающееся в центральной клетке зародышевого мешка.
Яйцевой аппарат в зрелом зародышевом мешке состоит из яйцеклетки и двух синергид. Этот комплекс клеток имеет очень своеобразные морфологические и физиологические особенности. Яйцеклетка обычно занимает среднее положение в яйцевом аппарате, она крупнее синергид, имеет удлиненную или грушевидную форму, содержит крупное ядро, расположенное в нижней части клетки, и густую цитоплазму. Синергиды — сестринские клетки, по строению и величине довольно однотипны. Они играют существенную роль в движении пыльцевых трубок к зародышевому мешку и яйцевому аппарату, а также способствуют растворению пыльцевых трубок.
Вблизи яйцевого аппарата располагается центральная клетка зародышевого мешка, включающая два полярных ядра.
В халазальном конце зародышевого мешка развивается комплекс антиподов из трех клеток, обладающий большой изменчивостью.
Оплодотворение у растений осуществляется при помощи опыления, прорастания пыльцевых трубок и слияния генеративных клеток.
Большинство покрытосеменных растений имеет обоеполые цветки, в которых обычно тычинки и пестики закладываются в разное время, благодаря чему обеспечивается перекрестное опыление. Последнее у цветковых растений происходит преимущественно при переносе пыльцы ветром или насекомыми.
Пыльцевые зерна, попадающие на рыльце пестика, начинают прорастать. Рыльце в это время выделяет раствор сахара (с примесью азотистых веществ), которым и питаются прорастающие пыльцевые трубки.
Направление роста пыльцевых трубок обусловливается явлением хемотропизма, то есть стремлением их к химическим веществам, имеющимся в завязи и на семяпочке.
В это время в пыльцевую трубку переходит содержимое пыльцевого зерна, состоящее из двух спермиев и вегетативного ядра либо из генеративной клетки и вегетативного ядра. В последнем случае спермии формируются уже в пыльцевой трубке. Подойдя к семяпочке, пыльцевая трубка в большинстве случаев проникает через микропиле в ее зародышевый мешок.
Оплодотворяются покрытосеменные растения следующим образом. Первая из проросших пыльцевых трубок, попадая в зародышевый мешок, изливается в одну из синергид, содержимое которой проникает в зону яйцевого аппарата, обычно в щель между яйцеклеткой и центральной клеткой. После излияния пыльцевой трубки в синергиду спермии попадают в плазму зародышевого мешка. Затем один из спермиев проникает в яйцеклетку и сливается с ее ядром, а другой спермий сливается с ядром центральной клетки. Как правило, этому предшествует слияние полярных ядер и образование ядра центральной клетки зародышевого мешка.
Впоследствии из оплодотворенной яйцеклетки (зиготы) образуется зародыш, а из оплодотворенного ядра центральной клехки — эндосперм.
Этот сложный половой процесс, имеющий универсальное значение для покрытосеменных растений, был назван двойным оплодотворением. Его открыл и изучил в 1898 г. академик С. Г. Навашин. Биологический смысл двойного оплодотворения у покрытосеменных заключается в том, что эндосперм как питательный материал для зародыша развивается раньше, чем зародыш. Лишь после того, как эндосперм в значительной степени сформирован, начинается развитие зародыша. В тех случаях, когда оплодотворение не произойдет, эндосперм не развивается, поскольку на его образование материнский организм должен истратить много энергии и питательного материала. При недоразвитии и аномалиях в развитии эндосперма семяпочка обычно превращается в неполноценное семя.
 Естествознание | Естествознание | 51 Морфология репродуктивных органов растений | растение, цветок, пестик, клетка, ядро, морфология |
www.abakbot.ru
13. Репродуктивные органы цветковых растений
Покрытосеменные, или цветковые растения являются наиболее крупным отделом растительного царства, насчитывающим более 350 семейств, 13 тысяч родов и до 240 тысяч видов. В настоящее время покрытосеменные распространены по всему Земному шару – от тропиков до приполярных пустынь. Автотрофные цветковые растения – важнейший компонент биосферы, от которого зависят биосферные процессы обмена веществ и трансформации энергии, газовый состав атмосферы, климат, водный режим суши, характер процессов почвообразования. На суше цветковые часто доминируют в растительном покрове, только в немногих сообществах и типах растительности они уступают господство по общей массе другим группам растений – в хвойных лесах голосеменным, в некоторых типах тундр и на сфагновых болотах – мохообразным.
Покрытосеменные определяют самую возможность существования большинства наземных животных. Они формируют среду обитания для разнообразных консументов – потребителей первичного органического вещества. Различные группы гетеротрофов связаны с ними трофическими, консортивными связями. В процессе эволюции многие группы гетеротрофов смогли возникнуть только тогда, когда Землю завоевали покрытосеменные и образуемые ими сообщества. Для многих членистоногих и позвоночных характерна сопряженная с цветковыми эволюция. Человек как биологический вид тоже возник и эволюционировал после появления покрытосеменных, отличающихся высоким уровнем продуктивности и биохимическим разнообразием.
Покрытосеменные растения отличаются необычайным полиморфизмом, эволюционной пластичностью и могут существовать в разнообразных условиях обитания. Это единственная группа среди высших растений, представители которой сумели вторично освоить морскую среду, где стали доминировать на огромных акваториях.
Разнообразие размеров и жизненных форм цветковых растений, их приспособлений к конкретным условиям обитания велико. Представитель семейства рясковых – ряска малая (Lemna minor) не превышает 3-5 мм в длину и похожа на свободноплавающие слоевищные растения – мхи или водоросли (рис. 13 .1).

Рис. 13.1. Ряска малая (Lemna minor) – миниатюрное цветковое растение.
С другой стороны, гигантские эвкалипты (виды рода Eucalyptus из семейства миртовые Myrtaceae) вырастают высотой до 100 м, длина побегов тропических лиан – ротанговых пальм измеряется сотнями метров. Среди покрытосеменных преобладают автотрофы, но встречаются полупаразиты, паразиты и сапротрофы, полностью утратившие хлорофилл и способность к фотосинтезу. Среди покрытосеменных имеются также эпифиты и эпифиллы (эпифиты, живущие на листьях других растений), деревья, кустарники, кустарнички, полукустарники и травы (многолетники и однолетники), плотоядные растения с их разнообразными приспособлениями для улавливания и переваривания мелких животных (рис. 13.2).

Рис. 13.2. Насекомоядное цветковое растение росянка круглолистная (Drosera rotundifolia).
Отмечены разнообразные приспособления видов цветковых растений к опылению строго определенными агентами, к распространению плодов и семян, к перенесению неблагоприятных климатических периодов. Эти особенности дают цветковым растениям возможность полностью реализовать свой эволюционно-адаптационный потенциал. Ни одна другая группа растений не смогла выработать такого разнообразия приспособлений к определенным факторам среды.
Существенным признаком покрытосеменных считают цветок – отсюда одно из названий отдела – Anthophyta (antos – цветок, phyton – растение). Происхождение цветка как особого органа – это одна из ключевых проблем морфологии и филогении покрытосеменных. Дискуссии по этому вопросу, начавшиеся еще в XVIII в., продолжаются и в настоящее время.
Типичный цветок определяют как видоизмененный побег, обеспечивающий функции спороношения, развития гаметофитов, половое размножение, образование семян в плоде. В истории ботаники отмечено, что в поисках архетипа цветка за первичный примитивный цветок различные ученые принимали разные структуры.
Сторонники эвантовой теории считают первичным крупный обоеполый цветок, опыляемый насекомыми, с большим и часто неопределенным числом свободных частей, и тогда цветки однополые, олигомерные, голые рассматриваются как результат более поздних этапов редукционной эволюции.
Сторонники псевдантовой теории полагают, что эволюционные трансформации выражались в постепенном усложнении структуры цветка от однополых, анемофильных, с небольшим числом частей к обоеполым, полимерным, энтомофильным, с хорошо развитым околоцветником.
Следует учитывать, что в процессе эволюционного возникновения покрытосеменные сначала приобрели комплекс важнейших общих признаков (в первую очередь – уникальные особенности жизненного цикла), а уже потом выработали разные цветки. При этом в каждой эволюционной ветви покрытосеменных были осуществлены различные морфогенетические решения.
Существенными частями цветка являются андроцей, состоящий из одной, нескольких или многих тычинок (микроспорофиллов) и гинецей, который состоит из одного, нескольких или многих плодолистиков, или карпелл (мегаспорофиллов).
Плодолистики (карпеллы) могут быть свободными, и тогда каждый из них, срастаясь краями, образует пестик (один или несколько пестиков в цветке – по числу плодолистиков). При срастании нескольких плодолистиков формируется ценокарпный гинецей, пестик в нем всегда один.
Важнейшая часть пестика – завязь. Наличие завязи (замкнутой полости), в которой развиваются семязачатки (в отличие от голосеменных, у которых семязачатки расположены на поверхности открытых мегаспорофиллов), обычно считают важнейшим признаком покрытосеменных, откуда и другое название отдела – Angiospermae (от греческого angeion – сосуд, вместилище, sperma – семя). Полагают, что завязь обеспечивает лучшую защиту семяпочек (семязачатков). В полости завязи семязачатки оказываются в условиях влажной камеры, оптимальных для их развития. Благодаря образованию завязи расширяются и обогащаются возможности распространения растений: из цветка формируется плод, при диссеминации расселяются не только отдельные семена, но и целые плоды, у которых возникают разнообразные приспособления к распространению широким кругом агентов.
У цветковых растений и некоторых голосеменных спермин (мужские неподвижные половые клетки) доставляются к яйцеклетке с помощью особой клетки – пыльцевой трубки, которая обеспечивает высокую точность попадания спермия в зародышевый мешок (то есть к женскому гаметофиту с яйцеклеткой). Семязачатки у покрытосеменных растений скрыты в завязи пестика, поэтому необходимо приспособление для улавливания пыльцы. Таким аппаратом у покрытосеменных служит рыльце на верхушке столбика пестика (стилодия). Наличие рыльца – важная особенность покрытосеменных растений.
Архегониальные растения (папоротникообразные, голосеменные) характеризуются тенденцией к возникновению разноспоровости, которая сопровождается редукцией полового поколения в жизненном цикле. Покрытосеменные, у которых редуцированы гаметангии, представляют собой завершающий этап на этом пути. Предельное сокращение процессов микро- и мегаспорогенеза и ускорение развития гаметофитов, связанное с их редукцией, были, вероятно, важными эволюционными факторами. Женский гаметофит цветковых растений представлен зародышевым мешком, состоящим всего из 7 клеток, а мужской гаметофит представлен пыльцевым зерном (пылинкой), в которой только 2 клетки – сифоногенная и спермагенная (дающая 2 спермия).
Уникальной особенностью покрытосеменных является двойное оплодотворение, в котором участвуют 4 клетки: два спермия, яйцеклетка и центральная клетка зародышевого мешка. В ходе этого процесса один из cnepмиев оплодотворяет яйцеклетку, образуя диплоидную зиготу, которая впоследствии развивается в зародыш семени. Второй спермий сливается с диплоидным ядром центральной клетки зародышевого мешка, образуя триплоидную зиготу. Эта зигота впоследствии дает начало триплоидному эндосперму семени. Общий для всех покрытосеменных уникальный процесс двойного оплодотворения указывает на их эволюционное единство и монофилетическое становление.
Одно из следствий двойного оплодотворения – формирование питательной ткани – полиплоидного эндосперма (в отличие от гаплоидного эндосперма голосеменных, представляющего остаток женского гаметофита). Биологическое значение полиплоидного состояния запасающей ткани связано с тем, что оно обеспечивает более благоприятные условия для развития зародыша и поэтому могло иметь важную эволюционную роль.
На тканевом уровне подавляющее большинство покрытосеменных характеризуется наличием в ксилеме сосудов, тогда как почти у всех голосеменных проводящие элементы древесины представлены только трахеидами. Только у немногих цветковых проводящая система ксилемы гомоксильная, бессосудистая, унаследовавшая примитивность от предковых голосеменных форм.
Паренхиматизация цветковых позволила выработать специальные запасающие ткани и травянистые жизненные формы. В отличие от древесных растений, у трав возможно сокращение онтогенеза и ускорение темпов эволюции; травянистые растения способны существовать в таких крайних условиях среды, в которых деревья и кустарники жить не могут.
Остается нерешенной проблема геологического возраста цветковых растений. Считалось, что покрытосеменные растения возникли раньше мелового периода – в триасе или палеозое (255-210 млн. лет назад). Предполагалось, что первоначально небольшая, эта группа существовала в таких условиях, которые не благоприятствовали фоссилизации их остатков, например, в горных районах или в пустынях. Позже, в меловой период (145-63 млн. лет назад), покрытосеменные дали мощную вспышку развития и сформировали свое современное разнообразие.
Этому взгляду противоречат объективные данные палеоботаники. Многочисленные указания на нахождение ископаемых остатков покрытосеменных в домеловых осадочных породах в настоящее время достоверно не доказаны. Имеются косвенные указания на возможность существования покрытосеменных в юре (210-145 млн. лет назад). Например, с этого времени известны некоторые формы насекомых, современные родственники которых настолько тесно связаны с цветковыми растениями, что допустить их независимое от последних существование трудно.
Древнейшие, достоверно определяемые ископаемые остатки цветковых растений в виде фоссилизованных листьев, цветков, плодов, древесины и пыльцевых зерен встречаются лишь с начала мелового периода (135 млн. лет назад). В этих отложениях они очень немногочисленны и фрагментарны. Еще недавно считалось, что к середине мелового периода (120 млн. лет назад) покрытосеменные появились в массе и выступали как «победители в борьбе за существование» с голосеменными и папоротникообразными. Сейчас палеоботаники отодвигают начало расцвета покрытосеменных на более позднее время – конец мелового периода (80-63 млн. лет назад).
Предполагают, что родиной цветковых могли быть тропические или субтропические регионы Земли. Исследователи считают, что это были районы тропической Юго-Восточной и Восточной Азии и Меланезия. В этом районе сосредоточены древние и считающиеся примитивными таксоны цветковых. Вероятно, что отсюда шла миграция древнейших цветковых в другие области Земли. Еще одна точка зрения заключается в признании центром происхождения покрытосеменны[ древнего континента Южного полушария – Гондваны. Считается, что цветковые могли возникнуть и расселиться по этому обширному континенту до начала раскола Гондваны на современные континенты Южного полушария.
К началу XX в. сложились две противостоящие друг другу системы взглядов на происхождение покрытосеменных от более примитивных предковых форм растений. Одна из них, связанная с именами австрийского ботаника Р. Веттштейна и классика немецкой ботаники А. Энглера, была основана на представлении о происхождении цветковых от эфедроподобных или гнетоподобных голосеменных предков. Р.Веттштейном была разработана оригинальная концепция происхождения цветка – так называемая псевдантовая гипотеза (от греческого «псевдантиум» – ложный цветок). Согласно этой гипотезе цветок представляет собой собрание мелких разнополых стробилов, претерпевших в процессе эволюции ряд преобразований: редукцию, сближение и срастание. На основе этой гипотезы наиболее архаичными следует считать семейства с тычиночными и пестичными невзрачными ветроопыляемыми цветками (ивовые, казуариновые).
Другая концепция восходит к построениям И.В.Гёте и А.П.Декандоля, то есть к концу XVIII – началу XIX веков. В эпоху филогенетической систематики это направление ознаменовалось появлением стробилярной (от греческого «стробилюс» – шишка хвойного)гипотезы, которую сформулировали Е.Арбер и Дж. Паркин . Другое название этой гипотезы – эвантиевая (от греческого «эвантиум» – настоящий цветок). Согласно стробилярной гипотезе наиболее вероятными предками покрытосеменных были мезозойские голосеменные растения – беннеттиты. С этих позиций цветок – это метаморфизированный (видоизмененный) укороченный спороносный побег, первоначально напоминавший шишку голосеменных. Мегаспорофиллы, несущие семязачатки, в процессе метаморфоза превратились в плодолистики (карпеллы), а несущие пыльники микроспорофиллы – в тычинки. Эти видоизменения многие исследователи связывают с приспособлением покрытосеменных к насекомоопылению (энтомофилии). Примитивные цветки имели значительные размеры и выпуклое цветоложе, на котором располагались многочисленные микро- и мегаспорофиллы. Микроспорофиллы эволюционировали в тычинки, а мегаспорофиллы – в карпеллы (плодолистики), которые образовали пестики. Цветки приобрели строение, сходное с тем, которое наблюдается у современных магнолиевых, лютиковых, кувшинковых. Поэтому перечисленные семейства обычно располагаются в основании системы покрытосеменных как наиболее древние и архаичные. Дальнейшая эволюция цветка в пределах покрытосеменных имела редукционный характер.
Обе эти гипотезы являются фолиарными (от латинского «фолиум» – лист), так как исходят из представлений, что цветки образовались из листостебельных спороносных побегов. Фолиарным гипотезам противопоставляются различные теломные (от греческого «телома» – конечные побеги) гипотезы (например, теория гонофилла Р. Мелвилла), широко распространенные в 60-70-е годы ХХ в. Согласно этим гипотезам, все части цветка могут быть выведены из теломов – осевых структур, свойственных ископаемым растения – риниевым.
Палеоботаника в последние десятилетия дала много нового материала о древнейших, ранне- и среднемеловых покрытосеменных. Эти данные позволяют сделать несколько заключений. Во-первых, уже самые ранние из известных нам цветковых отличаются большим полиморфизмом. Очень разнообразны пыльцевые зерна по форме, размерам, скульптуре поверхности, характеру поровости, числу и строению апертур. Они обнаруживают сходство с пыльцевыми зернами порядков и семейств современных растений. Листья меловых цветковых также весьма разнообразны и напоминают современные фикусы, протейные, дубы, акации, аралию, подорожник, известны и специализированные щитовидные листья.
Большим полиморфизмом отличаются и органы размножения – цветки и плоды. У меловых цветковых известны как одиночные и относительно крупные цветки, так и мелкие, собранные в разнообразно устроенные соцветия с простым или двойным околоцветником, с апокарпным или ценокарпным гинецеем, с верхней или нижней завязью, с разным числом тычинок и плодолистиков. Наряду со вскрывающимися плодами типа листовок обнаружены и не-вскрывающиеся плоды. Вместе с древесными формами в меловую эпоху существовали многочисленные травянистые покрытосеменные.
Во-вторых, если раньше часто утверждали, будто в меловую эпоху покрытосеменные были представлены современными таксонами, то сейчас большинство исследователей считают, что раннемеловые Angiospermae принадлежали к вымершим порядкам и семействам. Этот факт важен как доказательство того, покрытосеменные не оставались неизменными с момента зарождения.
В целом сейчас большинство специалистов считают, что предков Angiospermae нужно искать среди каких-то древних и малоспециализированных голосеменных растений. Не оставлена идея их генетической связи с беннеттитовыми. Допускают, что магнолиевые (как наиболее примитивные цветковые) лишь сохранили наибольшее число архаичных признаков, но сами, как и все прочие таксоны цветковых, произошли от какой-то вымершей, еще более древней гипотетической группы покрытосеменных.
В настоящем учебном пособии принята система цветковых растений, разработанная А.Л.Тахтаджяном в 1987 г. В данной системе древнейшей группой покрытосеменных считается порядок магнолиевых (Magnoliales), от предков которых, вероятно, произошли современные покрытосеменные. В этом смысле приведенная система монофилетична ( рис. 13.3).

Рис. 13.3. Система покрытосеменных растений (по А.Л.Тахтаджяну).
Цветок – особый побег покрытосеменных растений, ответственный за формирование спор двух типов (микро- и мегаспор), развитие мужского и женского гаметофитов, образование гамет (яйцеклеток и спермиев), опыление, оплодотворение и формирование семян. Цветок в процессе своего развития превращается в плод с семенами. Во многих отношениях он уникален и столь характерен для этой группы, что покрытосеменные нередко называют цветковыми растениями. Исключительная роль цветка как особой морфологической структуры связана с тем, что в нем полностью совмещены все процессы бесполого и полового размножения. В цветках осуществляются микро- и мегаспорогенез, развитие гаметофитов, микро- и мегагаметогенез, опыление, оплодотворение и формирование зародышей нового поколения спорофитов. Завершается онтогенез цветка образованием плода с семенами. Особенности строения цветка обеспечивают осуществление всех этих функций с наименьшими затратами пластических веществ и энергии.
studfiles.net
Введение
Развитие высших растений (мохообразные, плаунообразные, хвощеобразные, папоротникообразные, голосеменные и покрытосеменные) представляют собой сложный многофункциональный процесс, однако, условно его можно разделить на два периода: период формирования вегетативной сферы и период формирования репродуктивной сферы.
Особенности развития вегетативной сферы – развитие и строение вегетативных органов и образующих их тканей – изложены в 1-ой части курса лекций по морфологии растений (Т.А.Сауткина, В.Д.Поликсенова, 2004г.).
Благодаря вегетативным органам, которые выполняют функции почвенного и воздушного питания, транспирации, газообмена, выделения секреторных и экскреторных веществ, поддерживается индивидуальная жизнь каждой особи. На определенном этапе развития, когда вполне разовьются вегетативные органы, на растении начинают формироваться репродуктивные органы (спорангии, гаметангии) (иначе говоря, начинает развиваться репродуктивная сфера), в которых происходят процессы, связанные с размножением растений.
Размножение растений, т.е. способность оставлять после себя потомство, одно из основных свойств всех живых организмов. Оно поддерживает непрерывность и преемственность жизни видов и в целом жизни на Земле. У растений процесс размножения протекает очень сложно и сопряжен с чередованием в жизненном цикле гаметофитного (полового) и спорофитного (бесполого) поколений, т.е. в жизненном цикле имеет место смена фаз развития. Смена фаз развития происходит благодаря строгому чередованию в жизненном цикле высших растений двух репродуктивных процессов: спорообразования и полового процесса.
Все вопросы, касающиеся размножения представителей различных таксонов и чередования поколений у них, изложены в пособии авторов (Т.А.Сауткина, В.Д.Поликсенова. Размножение растений. Минск, 2001. 47 с.). Однако наиболее сложно и наиболее совершенно осуществляется развитие покрытосеменных растений, которые известный русский ботаник профессор Московского университета М.И. Голенкин (1864 - 1941) назвал «победителями в борьбе за существование». Уникальной особенностью покрытосеменных растений является то, что у них возникает особый репродуктивный орган – цветок, в котором протекают все процессы, связанные с размножением покрытосеменных растений.
Во 2-ой части курса лекций по морфологии растений речь пойдет о репродуктивной сфере покрытосеменных растений: о цветке, соцветиях, плодах и семенах. Кроме того, будет дано представление о строении зародыша, об особенностях прорастания семян двудольных и однодольных растений, о специфических условиях, необходимых для прорастания, а также о ранних фазах постэмбрионального развития растений.
Цветок как особый репродуктивный орган. Развитие представлений о цветке
Цветок (лат. – flos, греч. - anthos) – орган размножения цветковых, или покрытосеменных, растений. Развитие представлений о цветке как репродуктивном органе покрытосеменных растений обычно связывается с именем Иогана Вольфганга Гёте (1749 -- 1832) «Опыт объяснения метаморфоза у растений» (1790). Именно в этой работе Гёте впервые высказал предположение о том, что все элементы цветка представляют не что иное, как метаморфизированные листья вегетативного побега. Это предположение следует рассматривать как первую попытку выяснить морфологическую природу частей цветка. Идею Гёте о фолиарном (от лат. folium - лист) происхождении цветка поддержал О. П. Декандоль (1778 -- 1841), и на протяжении многих лет ботаники рассматривали цветок как вегетативный побег с сильно укороченной осью.
Во второй половине Х1Х – начале ХХ вв. под влиянием идей Ч. Дарвина (1809 - 1882) многие ботаники (Van Tieghem, 1891; Бекетов, 1897; Кернер, 1902; Буш, 1924; Кузнецов, 1936; Комаров, 1941) несколько переосмыслили первоначальное понимание природы цветка. Развивая фолиарную теорию Гёте, они стали рассматривать цветок как «видоизмененный (метаморфизированный) вегетативный побег». Представление о цветке как о метаморфизированном вегетативном побеге подтверждалось тем, что ботаники находили сходство в онтогенетическом развитии цветка и вегетативного побега, спорофиллов и листьев.
Такое представление о цветке господствовало вплоть до сороковых годов ХХ столетия. Приходится констатировать, что не только представление о морфологической природе элементов цветка складывалось постепенно, но и объем понятия «цветок» также сложился не сразу. Термин «Anthophyta» (от греч. anthos – цветок, phyton - растение) -- «цветковые растения» -- впервые был введен директором Берлинского ботанического сада профессором А. Брауном (1805 - 1877) для обозначения голосеменных и покрытосеменных растений. А. Браун называл цветком репродуктивные органы и тех, и других. Приблизительно в таком же объеме рассматривал понятие «цветок» и видный австрийский ботаник Р. Веттштейн (1863 – 1931). Однако были ботаники, которые еще более широко трактовали это понятие, Так немецкие ботаники Г. Галлир (1868 - 1932) и А. Энглер (1844 - 1930) считали, что можно говорить о цветке и у папоротников, если фертильные вайи у них сгруппированы как у страусника обыкновенного (Matteuccia struthiopteris). И только с 1907 г. по предложению английских палеоботаников Э. Арбера и Дж. Паркина понятие «цветковые растения» стали применять только в отношении покрытосеменных, а цветок стал рассматриваться как особый репродуктивный орган покрытосеменных растений. В цветке происходят все процессы, связанные с размножением покрытосеменных растений: образование микро- и макроспор, «прорастание» спор и образование мужского (пыльцы) и женского (зародышевого мешка) гаметофитов, формирование мужских (спермиев) и женских (яйцеклеток) гамет, процесс двойного оплодотворения, образование зиготы и эндосперма, развитие семени, включающего зародыш – зачаток нового спорофита, запас питательных веществ, образование плода.
Благодаря тщательным исследованиям всех выше перечисленных процессов, которые провели ботаники разных стран, с сороковых годов ХХ столетия в морфологии растений твердо установилось представление о том, что часть элементов цветка – это результат метаморфоза вегетативных листьев, а часть образовалась в результате метаморфоза спорофиллов (спороносных листьев).
Таким образом, учитывая морфологическую природу элементов цветка и особенности его функционирования, понятие «цветок» можно определить следующим образом: «Цветок – это укороченный метаморфизированный спороносный побег с ограниченным ростом, приспособленный для размножения».
ГИПОТЕЗЫ ПРОИСХОЖДЕНИЯ ЦВЕТКА
В связи с тем, что цветковые растения играют большую роль в жизни человека, вопрос об их происхождении, а также о происхождении их специализированного репродуктивного органа – цветка – занимали и занимают в научных исследованиях ботаников разных профилей первостепенное место. Несмотря на то, что морфологи, систематики, эмбриологи, палеоботаники накопили большое количество фактического материала, до сих пор не потеряли своего значения слова великого натуралиста Ч. Дарвина «об ужасной тайне, окружающей цветковые растения».
Действительно, вопрос о морфологической природе частей цветка не разрешил проблемы происхождения цветка как особого репродуктивного органа.
Попытка найти возможных предков покрытосеменных растений и на основании изучения их репродуктивных органов попытаться найти репродуктивный орган, который мог бы быть предшественником цветка, «заставили» ботаников обратить внимание на ископаемые голосеменные. Так возникли псевдантовая (от греч. pseudo – ложь, anthos – цветок ) и эуантовая, или стробилярная (от греч. eu – истина, anthos – цветок) теории происхождения цветка.
Псевдантовая теория происхождения цветка была сформулирована Р. Веттштейном в1901 г. В качестве предковой группы покрытосеменных растений он рассматривал ископаемые голосеменные эфедровые, у которых типичных шишек нет, но существуют своеобразные структуры, которые называют «микростробилами» и «мегастробилами». Отдельный микростробил внешне похож на тычинку покрытосеменных, а группа микростробилов, окруженных кроющими листьями, -- на их соцветие. Мегастробил состоит из одного семязачатка (семяпочки) и окружен покровом. Р. Веттштейн полагал, что из микростробила в результате редукции кроющих листьев мог возникнуть тычиночный цветок с простым околоцветником, а из мегастробила – голый женский цветок. Согласно представлению Веттштейна, обоеполый цветок покрытосеменных гомологичен «соцветию» (собранию микро- и мегастробилов) эфедровых, но показать, каким образом из однополых цветков могли образоваться обоеполые, убедительно доказать не удалось. Постепенно ботаники от этой гипотезы отказались. Однако в свое время псевдантовая гипотеза сыграла определенную роль и была положена в основу ряда систем покрытосеменных растений. Самой известной из таких систем является система А. Энглера (1844 -- 1930), единственная система, филогенетические связи в которой прослеживаются до рода. До настоящего времени ею пользуются в ряде крупных Гербариев, в том числе и в Гербарии Ботанического института имени В. Л. Комарова Российской Академии наук (Санкт- Петербург).
К концу ХIХ началу ХХ веков создались предпосылки для осмысливания путей происхождения цветка в процессе эволюции. Опираясь на данные палеоботаники и морфологии, американский ботаник и филогенетик Ч. Бэсси (1845 - 1915) отмечает, что репродуктивный орган покрытосеменных состоит из стебля, на котором развиваются спороносные и стерильные листья, а позднее высказывает предположение, что покрытосеменные произошли от стробилоносных саговниковых. Независимо от Ч. Бэсси немецкий систематик Г. Галлир в 1902 г. на основании изучения позеленевшего пролиферирующего цветка водосбора (Aquilegia vulgaris) рассматривает цветок как видоизмененный репродуктивный побег. Таким образом, Ч. Бэсси и Г. Галлир фактически явились основоположниками эуантовой, или стробилярной, гипотезы происхождения цветка. Однако окончательно она была сформулирована несколько позже.
В 1906 г. Виланд описал обоеполый стробил ископаемых голосеменных – беннетитовых. Это открытие позволило Э. Арберу и Дж. Паркину окончательно сформулировать стробилярную гипотезу происхождения цветка. Согласно этой гтпотезе, цветок представляет собой обоеполый стробил, приспособленный для размножения. Цветок в представлении Э. Арбера и Дж. Паркина – это новейшая модификация стробила – антостробил. Э. Арбер Дж. Паркин считают, что беннетитовые, очевидно, не прямые предки покрытосеменных растений, но, возможно, они являются предками каких-то гипотетических полупокрытосеменных, которые до сих пор не выявлены. В своем труде «О происхождении покрытосеменных» (1907) авторы устанавливают признаки примитивности и специализации цветка и высказывают свою точку зрения на филогенетическую систему покрытосеменных. Она диаметрально противоположна точке зрения Р. Веттштейна. Если Р. Веттштейн считал наиболее примитивными семействами те, представители которых имели мелкие раздельнополые ветроопыляемые цветки без околоцветника или с редуцированным околоцветником, то, по Арберу и Паркину, самые примитивные семейства включают представителей с крупными обоеполыми насекомоопыляемыми цветками, имеющими простой или двойной околоцветник.
Открытие псилофитовых (риниообразных), признанных наиболее примитивными высшими растениями, дало основание вновь вернуться к проблеме происхождения цветка и выдвинуть теломную теорию. Основоположником ее является В. Циммерман (1930). Он считает, что все органы высших растений развились из простых и недифференцированных элементов, названных им теломами. Согласно представлениям В. Циммермана, стерильные телломы – филлоиды и фертильные теломы, несущие спорангии, могли срастаться и формировать синтеломы. Цветок, по В. Циммерману, представляет собой синтелом, процесс возникновения которого происходил параллельно процессу формирования вегетативного побега. И хотя детально эта теория не разработана, она имела большое значение для решения некоторых вопросов о природе элементов цветка. С 40-х – 50-х годов ХХ столетия – периода особенно острых дискуссий сторонников теломной теории и приверженцев классической морфологии, цветок перестали определять как «метаморфизированный вегетативный побег».
В связи с появлением новых научных данных появляются и новые гипотезы происхождения цветка, однако, фактически ни одна из них не является достаточно убедительной. Суммируя все знания в этой области известный советский ботаник Б. М. Козо-Полянский (1890 -1957) вынужден был констатировать: «Не известны ни предок цветковых растений, ни конкретный орган, исторически исходный для цветка».
studfiles.net
репродуктивные органы - это... Что такое репродуктивные органы?
репродуктивные органыАнатомия и морфология высших растений. Словарь терминов. — М.: Дрофа. Коровкин О.А.. 2007.
- репродуктивная часть монокарпического побега
- репродукция
Смотреть что такое "репродуктивные органы" в других словарях:
Репродуктивные органы — * рэпрадуктыўныя органы * reproductive organs органы, связанные с функцией размножения полового (генеративные органы) или вегетативного … Генетика. Энциклопедический словарь
РЕПРОДУКТИВНЫЕ ОРГАНЫ — (от лат. rе приставка, здесь означающая возобновление, повторение и produco создаю), органы растений и животных, выполняющие функции размножения. У растений Р. о. представляют собой разл. структуры, обеспечивающие вегетативный, бесполый и половой … Биологический энциклопедический словарь
репродуктивные органы — ЭМБРИОЛОГИЯ ЖИВОТНЫХ РЕПРОДУКТИВНЫЕ ОРГАНЫ – органы, обеспечивающие процесс размножения … Общая эмбриология: Терминологический словарь
репродуктивные органы — reprodukcijos organai statusas T sritis augalininkystė apibrėžtis Organai, atliekantys dauginimosi funkciją. atitikmenys: angl. reproductive organs rus. репродуктивные органы … Žemės ūkio augalų selekcijos ir sėklininkystės terminų žodynas
Репродуктивные органы — органы воспроизведения, органы растений и животных, выполняющие функции размножения. У многих низших и всех высших растений в циклах развития осуществляется смена ядерных фаз, приводящая к чередованию бесполого (Спорофит) и полового… … Большая советская энциклопедия
РЕПРОДУКТИВНЫЕ ОРГАНЫ — органы, связанные с функцией полового размножения. См. Половые органы … Энциклопедический словарь по психологии и педагогике
РЕПРОДУКТИВНЫЕ ОРГАНЫ — органы полового размножения растений … Словарь ботанических терминов
репродуктивные органы — органы растения, создающие половое потомство: цветок, соцветие, семена, ягоды … Сельскохозяйственный словарь
Репродуктивные органы — у растений суть органы размножения; их противополагают вегетативным, рабочим органам, назначение которых состоит в удовлетворении всех остальных жизненных потребностей организма. Различают два основных типа Р. органов: органы бесполого… … Энциклопедический словарь Ф.А. Брокгауза и И.А. Ефрона
РЕПРОДУКТИВНЫЕ ОРГАНЫ — растений, органы, обеспечивающие вегетативное, бесполое и половое размножение. Р. о. мн. водорослей, а также мхов, папоротников, хвощей и плаунов с выраженным чередованием поколений (спорофит и гаметофит) представлены спорангиями (обеспечивают… … Сельско-хозяйственный энциклопедический словарь
plant_anatomy.academic.ru
Вегетативные и репродуктивные органы растений
Растения состоят из таких органов, как вегетативные и репродуктивные. Каждый из них отвечает за определенные функции. Вегетативные – за развитие и питание, а репродуктивные органы растений участвуют в размножении. К ним относят цветок, семя и плод. Они отвечают за "рождение" потомства.

Вегетативные органы
Появление вегетативных органов было связано с необходимостью получать питательные вещества из почвы. К ним относятся:
- Корень - основной орган каждого растения, произрастающего в земле.
- Побег.
- Стебель.
- Листья, отвечающие за фотосинтез.
- Почки.
Корень характерен для всех растений, так как он удерживает их и питает, добывая из воды полезные вещества. Именно от него идут побеги, на которых вырастают листья.
При посеве семян первым прорастает корешок. Он является главным органом растения. После того, как корень наберется сил, появляется система побегов. Затем формируется стебель. На нем располагаются боковые побеги в виде листьев и почек.
Стебель поддерживает листья и проводит к ним от корней питательные вещества. Также он может запасать в себе воду на время засухи.
Листья отвечают за фотосинтез и газообмен. У некоторых растений они выполняют и другие функции, такие как запас веществ или размножение.
В процессе эволюции органы видоизменяются. Это дает возможность растениям приспосабливаться и выживать в природе. Появляются новые виды, которые все более уникальны и неприхотливы.
Корень
Вегетативный орган, который удерживает стебель, участвует в процессе всасывания воды и питательных веществ из почвы на протяжении всей жизни растения.
Он возник после появление суши. Корень помог растениям приспособиться к изменениям на земле. В современном мире остались еще бескорневые – мох и псилотообразные.
У покрытосеменных растений развитие корня начинается с попадания зародыша в землю. По мере развития появляется устойчивый орган, из которого прорастает побег.
Корень защищен чехликом, который помогает получать полезные вещества. Это происходит благодаря его строению и содержанию большого количества крахмала.
Стебель
Осевой вегетативный орган. Стебель несет на себе листья, почки и цветы. Он является проводником питательных веществ от корневой системы к другим органам растения. Стебель у травянистых видов также способен к фотосинтезу, как и листья.
Он способен выполнять следующие функции: запасающая и размножения. Структура стебля представляет собой конус. Эпидермис, или ткани, являются у некоторых видов растений первичной корой. У цветоносов она более рыхлая, а у побегов, например у подсолнечника, пластинчатая.
Функция фотосинтеза выполняется благодаря тому, что в составе стебля есть хлоропласт. Это вещество преобразует углекислый газ и воду в органические продукты. Запас веществ происходит благодаря крахмалу, который не расходуется в период роста.
Интересно, что у однодольных растений стебель сохраняет свою структуру на протяжении всего цикла жизни. У двудольных он изменяется. Это можно увидеть по срезу деревьев, где образуются годичные кольца.
Лист
Это боковой вегетативный орган. Листья отличаются по внешнему виду, строению и функциям. Орган участвует в фотосинтезе, газообмене и транспирации.
Эволюция растений привела к появлению ловчих видов. У них листья ловят насекомых и питаются ими. Этот орган у некоторых видов растений превращается в колючки или усики, тем самым выполняя защитную функцию от животных.
У листа есть основание, которое соединяет его со стеблем. Через него в листья поступают питательные вещества. Основание может расти в длину или в ширину. Вслед за ним вырастают прилистники. У листа есть жилки, которые делятся на два вида: открытые и закрытые.
Продолжительность жизни у этого вегетативного органа невелика. Деревья сбрасывают листья, так как в них остаются продукты жизнедеятельности, оставшиеся после фотосинтеза.
Вегетативное размножение
У каждого растения есть свой цикл жизни. Выделяют два вида размножения с помощью вегетативных органов:
- Естественное.
- Искусственное.
Естественное размножение производится за счет листьев, плетей, корневых клубней, корневищ, луковиц.
Искусственное размножение:
- Делят куст. Корневищные растения разделяют на несколько частей и рассаживают.
- Второй способ - это укоренение черенков. Они могут быть не только корневыми, но и листовыми и стеблевыми.
- На материнском растении можно использовать отводки.
- Также популярен способ прививания. Это когда часть одного растения переносят на другое.
Вегетативные органы помогают так же, как и репродуктивные, в размножении. Растения играют большую роль в жизни человека и природы. На земле они занимают достаточно большое пространство.
Функция репродуктивных органов
Их значение в строении цветка обеспечивает размножение вида, защиту семян и их дальнейшее расселение. Репродуктивные органы покрытосеменных растений – это цветок, семя и плод. Они постепенно сменяют друг друга.
Цветок – это видоизмененный побег, который постепенно меняет облик. Семя, которое находится внутри, созревает и набирает питательные вещества. После оплодотворения оно превращается в плод. Он состоит из множества семян и околоплодника, который защищает их от внешней среды.
Вегетативные и репродуктивные органы растений всегда взаимодействуют. Друг без друга они не смогут выполнять свои функции.
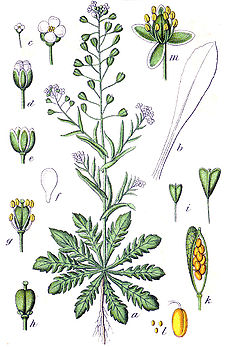
Цветок
В природе все устроено так, чтобы цветы проживали свой цикл заново. Как мы уже говорили, к репродуктивным органам растения относятся цветок, плод и семя. Они взаимосвязаны друг с другом, чтобы поддерживать жизнь и давать возможность рождаться новым поколениям.
Такой репродуктивный орган растения, как цветок, отвечает за опыление, оплодотворение и образование семян. Он является укороченным побегом, который видоизменяется в процессе роста.
Рассмотрим, из чего образуется цветок:
- Цветоножка – осевая часть.
- Чашечка. Состоит из чашелистиков и находится в нижней части соцветия.
- Венчик. Отвечает за окраску цветка и состоит из лепестков.
- Тычинка. В ней образуется пыльца, которая помогает в опылении.
- Пестик. Это место, где прорастает пыльца.
 Цветы делятся, в свою очередь, на обоеполые и однополые. Чем они отличаются? У обоеполых есть одновременно тычинка и пестик. Например, кукуруза и тыква. Однополые, или однодомные, имеют только один орган. К ним относятся крапива, конопля. Цветок является репродуктивным органом растения, который отвечает за семенное размножение.
Цветы делятся, в свою очередь, на обоеполые и однополые. Чем они отличаются? У обоеполых есть одновременно тычинка и пестик. Например, кукуруза и тыква. Однополые, или однодомные, имеют только один орган. К ним относятся крапива, конопля. Цветок является репродуктивным органом растения, который отвечает за семенное размножение.
Чаще всего образуются соцветия. Это группа из нескольких цветков. Они бывают простые и сложные, то есть с одной цветоножкой или с несколькими. Их число может достигать десятка тысяч на одном растении.
Соцветие – это группа цветков. Она расположена на концах побегов, а также веточках деревьев. Чаще всего соцветие образуется из мелких цветочков. Они, в свою очередь, делятся на простые и сложные. Первые имеют одну ось, на которой и располагаются цветы. Вторые имеют боковые ответвления.
Распространенные типы соцветий:
- Кисть – у черемухи, ландыша.
- Початок – у кукурузы.
- Корзинка – ромашка или одуванчик.
- Зонтики – у вишни.
- Щиток – у груши.
Сложные соцветия представляют собой несколько простых. Их происхождение связано с функцией оплодотворения. Чем больше количество цветов, тем быстрее пыльца переносится.
Плод
Репродуктивные органы растений в первую очередь выполняют функцию размножения. Плод защищает семена от их преждевременного распространения. Они бывают сухие или сочные. Семена образуются внутри плода, постепенно созревая. Некоторые из них оснащены приспособлениями, которые помогают распространиться, например, одуванчик разлетается по ветру.
Основные виды плодов:
- Односемянный с тремя слоями – вишня, абрикос, персик.
- Многосемянный с мякотью – виноград.
Сухой многосемянный плод бывает с перегородкой – капуста, и без нее – горох. У дуба – односемянный.
Репродуктивные органы цветковых растений устроены так, что семена распространяются несколькими способами:
- По воде.
- По воздуху.
- С помощью животных.
- Саморазбрасываются.
Органы устроены так, чтобы растения проходили процесс от зарождения корней до размножения. Плоды приспособились к тому, чтобы их переносили животные. Это обеспечивается такими приспособлениями, как зацепки, парашютики, цветовые акценты и приятный вкус.
Семя
Зная, какие органы растений относятся к репродуктивным, можно понять, как именно они размножаются. Семя воспроизводит потомство и расселяет его для последующего выращивания. Оно складывается из кожуры, зародыша и питательных веществ, поступающих от стебля.
В составе семени есть белки, жиры и углеводы. По сути, зародыш – это зачатки стебля, корня и листиков. Он является главной частью семени и бывает с одной или двумя семядолями.
Семена тоже подразделяются на несколько разных типов. У одних питательные вещества находятся в эндосперме, у других отсутствуют полностью ткани для запасов.
Семенная кожура защищает от воздействия внешней среды, ветра и животных. После созревания она помогает расселить растение. Некоторые виды запасают в кожуре питательные вещества.
Семена для людей и животных являются пищей. Их значение на земле достаточно высоко, как и у плода. Эти органы растений участвуют в цикле жизни насекомых и животных, тем самым обеспечивая их пищей.
Высшие растения
В растительном мире все устроено так, чтобы организмы имели возможность расти постоянно. Высшие растения имеют такие органы, как побег и корень. Они отличаются тем, что в процессе оплодотворения появляется зародыш.
Репродуктивные органы высших растений, взаимодействуя с вегетативными,сменяют свои жизненные фазы. Они включают в себя четыре отдела:
- Папоротникообразные произрастают в увлажненных местах. К ним относят хвощи и плауны. Их строение включает в себя корень, стебель и листья.
- Мохообразные являются промежуточной группой. Их тело состоит из ткани, однако сосудов они не имеют. Живут как во влажной, так и в сухой почве. Мох размножается не только спорами, но и половым и вегетативным способом.
- Голосеменные. Самые древние растения. Чаще всего к ним относят хвойные деревья и кустарники. Они не цветут, а их плоды образуют шишку с семенами внутри.
- Покрытосеменные. Самые распространенные растения. Они отличаются тем, что семена надежно укрыты под кожурой плода. Размножение происходит несколькими способами. Они отличаются тем, что имеют женские и мужские половые органы в строении.

Все эти растения растут и развиваются на земле уже достаточно продолжительное время. Они отличаются друг от друга способом размножения и наличием тех или иных органов. Однако нельзя не отметить, что растительность оказывает большое влияние на жизнь человека.
Цветковые растения
Этот вид является самым многочисленным в растительном мире. Цветковые, или покрытосеменные растения, росли на планете с древних времен. Папоротники в процессе эволюции разделились на множество видов.
Главные репродуктивные органы цветковых растений – это семена. Они защищены плодом, что помогает им лучше сохраняться до момента распространения. Интересно, что эта группа растений единственная, которая может образовывать многоярусные сообщества. В свою очередь, цветочные делятся на два подвида: однодольные и двудольные.
Главным отличием цветковых является то, что репродуктивные органы растений – цветок, плод и семя. Опыление происходит посредством ветра, воды, насекомых и животных. В строении растения есть женский и мужской заросток, а также происходит двойное оплодотворение.
Семя при прорастании насыщается водой и набухает, затем запасные вещества расщепляются и дают энергию для прорастания. Из зародыша появляется росток, который в дальнейшем становится цветком, деревом или травой.
Голосеменные растения
Этот вид появился миллионы лет назад. Голосеменные растения размножались спорами, а в процессе эволюции появились семена. По своему строению плод – это шишка. Семя располагается под чешуйками и ничем не защищено.
У голосеменных растений репродуктивные органы могут быть различного вида. У некоторых это шишки, у других они похожи на ягоды.
К ним относятся не только хвойные, но и лиственные деревья. В пустынях Кении растет удивительное растение, у которого всего два больших листа. Его родственником является эфедра. Это голосеменное растение, у которого есть небольшие круглые ягодки.
Процесс опыления
Как известно, к репродуктивным органам растения относятся цветок, плод и семя. Чтобы произошел процесс оплодотворения, необходимо опыление, которое помогает появлению потомства.
У покрытосеменных растений происходит слияние мужских и женских клеток. Это получается благодаря перекрестному опылению. Это процесс переноса пыльцы с одного цветка на другой. В некоторых случаях происходит самоопыление.
Для перекрестного опыления необходимы помощники. В первую очередь, это насекомые. Они лакомятся сладкой пыльцой и переносят ее с цветка на цветок на своих рыльцах и крылышках. После этого начинают свою работу репродуктивные органы растений. Цветы, которые опыляются с помощью насекомых, окрашены в яркие и сочные оттенки. После окраски их привлекает аромат. Насекомые чувствуют запах цветка, находясь от него на достаточно большом расстоянии.
Ветроопыляемые растения также оснащены специальными приспособлениями. Их пыльники достаточно свободно расположены, поэтому ветер переносит пыльцу. Например, тополь цветет во время ветров. Это дает возможность без препятствий разносить пыльцу с одного деревца на другое.
Есть растения, в опылении которым помогают маленькие птицы. Их цветы не имеют резкого аромата, но оснащены яркой красной окраской. Это привлекает птиц пить нектар, и одновременно происходит опыление.
Эволюция растений
После появления суши природа изменилась. Растения постепенно эволюционировали, и на смену папоротникам пришли цветы, кустарники и деревья. Это произошло из-за появления корневой системы, тканей и клеток.
Благодаря разнообразию репродуктивных органов покрытосеменных растений появлялось все больше видов и подвидов. Для размножения начали появляться споры и семена, в которых находились половые клетки.
Постепенно появились побеги, листья и плоды. После выхода на сушу растения развивались в двух направлениях. Одни (гаметофитные) имели две фазы развития, другие (спорофитные) переходили из одного цикла в другой.
Растения приспосабливались и развивались. Споровые виды стали достигать 40-метровой высоты. Стали появляться все новые репродуктивные органы растений. Их эволюция зависела от воздействия внешней среды.
Внутри семени формировался зародыш, который после оплодотворения и распыления прорастал. Попадая в землю, он питался полезными веществами и превращался в росток.
Эволюция процесса оплодотворения привела к появлению покрытосеменных растений, у которых семена были защищены плодом.
Значение растений для человека
Польза природного мира для людей бесценна. Растения не только выделяют газы, соли и воду, но и превращают неорганические вещества в необходимые для жизни. С помощью корневой системы, побегов и листьев происходит газообмен.
Зеленые растения накапливают в себе ценные органические вещества, очищают воздух от углекислого газа, при этом насыщая его кислородом.
Благодаря природным ресурсам люди получают более ценные продукты, необходимые для жизни. Растения становятся пищей для животных и человека. Их используют для лечения различных болезней, в производстве косметики.
Так как репродуктивным органом растения является плод и семя, то они стали незаменимыми в питании людей. Ягоды, которые растут на кустарниках, любят практически все. Что интересно, уголь и нефть также произошли от растительности. Торфяники - это зарождение водорослей и папоротников.
Вегетативные и репродуктивные органы цветковых растений играют важную роль в их жизни. Они отвечают за питание, развитие и размножение. Когда жизненный цикл заканчивается, семена распространяются вокруг и прорастают новые растения.
загрузка...
buk-journal.ru
Репродуктивные органы высших растений
Репродуктивные органы высших растений
Быстрое освоение суши наземными растениями объясняется не только совершенством их вегетативных органов, но теми принципиальными изменениями, которые претерпели репродуктивные органы - гаметангии и спорангии . Основное назначение репродуктивных органов заключается в образовании и расселении потомства, обеспечивающего сохранение и процветание вида. Поэтому в условиях наземной среды обитания они должны быть более надежно защищены, чем одноклеточные гаметангии и спорангии водорослей. В ходе эволюции у всех высших растений сформировались многоклеточные спорангии и гаметангии, имеющие стенку из живых клеток. Цитоплазма клеток стенки, прочно удерживая влагу, предохраняет развивающиеся гаметы и споры от высыхания. Мужской половой орган - антеридий - имеет овальную или шаровидную форму ( рис. 13 ). Под однослойной стенкой располагается сперматогенная ткань, из которой формируются жгутиковые сперматозоиды. Ко времени их созревания при наличии воды стенка вскрывается, и сперматозоиды по воде двигаются по направлению к архегонию . Женский половой орган - архегоний - имеет колбовидную форму; верхняя узкая часть называется шейкой, нижняя расширенная - брюшком. Внутри шейки под защитой стенки расположены шейковые канальцевые клетки, в брюшке - одна-две брюшные канальцевые клетки. На дне брюшка помещается одна крупная яйцеклетка. К моменту ее созревания шейковые и брюшные канальцевые клетки, а также верхние клетки стенки ослизняются, и часть слизи выходит за пределы архегония. Она содержит вещества, действующие хемотаксически на сперматозоиды, которые подплывают к архегонию, по слизи шейки двигаются по направлению к яйцеклетке и оплодотворяют ее ( рис. 13 ).
Возможный путь возникновения многоклеточных архегониев и антеридиев высших растений рисует теория Дэвиса, созданная в 1903 г. Согласно этой теории, гаметангии высших растений произошли из многокамерных гаметангиев водорослей, подобных тем, которые имеются у эктокарпуса (Ectocarpus) из отдела бурых и у хетонемы (Chaetonema) из отдела зеленых . У них каждая клетка многокамерного гаметангия оказывается фертильной, т.е. способной образовывать изогаметы. Первые наземные растения, по представлению Дэвиса, обладали изогамным половым процессом. По мере освоения суши периферические клетки гаметангиев подвергались стерилизации и превращались в стенку, которая защищает внутренние фертильные клетки от высыхания. В дальнейшем происходило разделение полов гаметангиев.
Формирование антеридиев сопровождалось увеличением количества сперматогенных клеток и уменьшением их размеров. Увеличение количества сперматозоидов повышало вероятность полового процесса, а уменьшение их размеров способствовало передвижению сперматозоидов в самых тонких пленках воды. При формировании архегониев происходило увеличение размеров яйцеклеток и постепенное сокращение их числа до одного. Этот процесс биологически оправдан, так как одна крупная яйцеклетка, содержащая большой запас питательных веществ, способна обеспечить развитие наиболее полноценного потомства. Стерилизация внутренних клеток архегония и превращение их в брюшные и шейковые канальцевые клетки способствовало лучшему проведению сперматозоидов к яйцеклетке. Эта теория находит подтверждение в наличии так называемых смешанных гаметангиев , которые встречаются среди мохообразных . В одних случаях в гаметангиях могут развиваться одновременно яйцеклетки и сперматозоиды, в других случаях в архегонии развивается несколько яйцеклеток.
В результате полового процесса образуется диплоидная зигота. Она формирует диплоидный спорофит , который заканчивает свое развитие образованием многоклеточного спорангия со спорами . У всех высших растений, за исключением мохообразных , спорангии возникают на специальных органах, которые широко трактуются как спорангиофоры, т.е. носители спорангиев . Форма спорангиофоров в разных группах может быть разнообразной; в тех случаях, когда они имеют плоскую листовидную форму, их называют спорофиллами . Стенка спорангия может быть однослойной либо многослойной. Ее формирование в ходе эволюции могло происходить аналогично формированию стенки гаметангиев, т.е. путем стерилизации периферических спорогенных клеток.
В спорангиях всех высших растений в результате мейотического (редукционного) деления возникают тетрады гаплоидных спор. Характерной чертой всех высших растений является наличие в оболочке спор спорополлинина - вещества, близкого по физическим и химическим свойствам к кутину , поэтому обычно оболочки спор называют кутинизированными. Благодаря большой стойкости к химическим воздействиям и водонепроницаемости оболочек споры могут длительное время (иногда десятилетиями) сохранять свою жизнеспособность. Из гаплоидных спор формируется гаплоидное половое поколение - гаметофит . Мохообразные унаследовали гаметофитный тип цикла воспроизведения, при котором ведущее положение занимает гаметофит. Спорофит представлен лишь коробочкой и гаусторией (иногда развивается ножка), т.е. он не является самостоятельно живущим поколением. Поскольку половой процесс у мохообразных осуществляется сперматозоидами, т.е. при участии воды, гаметофит должен быть связан с влажными местообитаниями и не может по той же причине достигать больших размеров. Кроме того, гаплоидный гаметофит обладает меньшим генетическим потенциалом, чем диплоидный спорофит . Поэтому эта линия эволюции оказалась боковой, тупиковой. У всех остальных высших растений спорофит занимает ведущее место в цикле воспроизведения. Поскольку он обеспечивает размножение кутинизированными спорами, то является типично наземным растением.
Диплоидный набор хромосом расширил возможности формообразовательных процессов, поэтому огромное разнообразие жизненных форм высших растений принадлежит бесполому диплоидному поколению. Обильное ветвление и создание крупных размеров спорофита приводило к колоссальной продуктивности спор и к их эффективному рассеиванию.
Гаметофит , назначение которого состоит в осуществлении полового процесса, подвергся у большинства высших растений значительной редукции и существует в виде недифференцированного таллома , получившего название заростка, или проталлиума (prothallium) . Широко распространено мнение, что предковые формы высших растений обладали изоморфным чередованием поколений, и от этого типа в дальнейшем эволюция пошла по двум направлениям - гаметофитному и спорофитному. Однако высказывается мысль и о том, что уже исходная группа водорослей обладала разными типами цикла воспроизведения и дала две независимые линии эволюции.
Ссылки:
medbiol.ru