Растительная клетка как живая система. Раздражение клетки растений
Реакция клеток на раздражение
Раздражимость. На любой организм постоянно действуют разнообразные факторы внешней среды, например: свет, температура, давление, звук, электрический ток, сила тяжести и др. Действие всех внешних факторов-раздражителей вызывает у организма ответные реакции, в основе которых лежит свойство раздражимости клеток. Раздражимостью называют способность организмов, а также клеток отвечать на воздействия внешней среды определенными реакциями.
Раздражимость можно наблюдать у любых клеток и организмов. У простейших, например у амеб, эвглен, инфузорий, реакция на изменение условий среды проявляется в передвижении их по отношению к раздражителю. Такие движения называются таксисами.
Если простейшие движутся по направлению к раздражителю, то такие движения их именуются положительным таксисом; движения же простейших от раздражителя носят название отрицательного таксиса. Те движения, которые возникают в ответ на действие света, получили название фототаксиса. Пример фототаксиса — движение зеленых жгутиконосцев по направлению к источнику освещения: если аквариум, в котором находятся эвглены, одинаково освещен со всех сторон, то эвглены равномерно распределяются по всей толще воды. Если же наиболее сильно осветить лишь какую-либо одну часть аквариума, то эвглены скапливаются именно в этой освещенной части, проявляя положительный фототаксис по отношению к свету.
Движения простейших, вызванные действием химических веществ, именуются хемотаксисами. Хемотаксис можно наблюдать у инфузории туфельки: если в пробирку налить воду с находящимися в ней инфузориями, то через небольшой промежуток времени они все соберутся в верхнем слое воды, богатом кислородом. Инфузориям необходим кислород для дыхания, и они по отношению к нему проявляют положительный хемотаксис. Те движения простейших, которые возникают под влиянием изменения температуры, называются термотаксисом. Термотаксис можно также легко наблюдать у инфузории туфельки. Для этого туфелек вместе с небольшим количеством среды, в которой они находятся, помещают в тонкий стеклянный капилляр, который с одной стороны охлаждается льдом, а с другой подогревается горячей водой до температуры 38—40° С. Туфельки, сначала равномерно распределявшиеся по всей длине капилляра, начинают двигаться от слишком холодных и слишком горячих его участков, проявляя к ним отрицательный термотаксис и собираясь в средней зоне с температурой 24—26° С, которая для них служит оптимальной, т. е. наилучшей для жизни. Именно к этой температуре они обнаруживают отчетливо выраженный положительный термотаксис.
Явление раздражимости хорошо выражено и у клеток растений. Чаще всего у растений встречаются проявления раздражимости в форме медленных двигательных реакций. Такие медленные движения, направленные к раздражителю или от него, называются тропизмами. У растений широко распространены фототропизмы — движения, возникающие в ответ на действие света. Растения тянутся к свету, изгибаются по направлению к нему, и в основе этой реакции лежит свойство раздражимости их клеток.
Иногда же клетки растений быстро реагируют на действие раздражителей. Примером может служить быстрая реакция у растения, известного под названием «стыдливая мимоза». При любом прикосновении к мимозе, при помещении в темноту или в условия повышенной температуры листья ее складываются и как бы увядают. Как только действие раздражителя прекращается, листья мимозы принимают прежнее положение. В основе этой быстрой реакции мимозы лежит также свойство раздражимости ее клеток. Еще пример быстрой реакции растения на действие раздражителя. На болотах, а иногда и по берегам ручьев растет росянка — растение, питающееся насекомыми. Росянка — небольшое растение с розеткой стелющихся листьев, похожих на лопаточки. Поверхность каждого листа покрыта чувствительными волосками красного цвета. Кончик каждого волоска утолщен и покрыт капельками блестящего, как роса, и липкого, как клей, сока. Если на такой лист сядет насекомое, например комар или небольшой жук, то клейкий сок волосков сразу же затрудняет его движения и насекомое оказывается в западне. Волоски листа, задетые насекомым, быстро складываются над пойманной добычей и обильно поливают ее соком. Сок, выделенный секреторными клетками листа, содержит ферменты, под действием которых расщепляются белки. Насекомое переваривается и через несколько часов всасывается. После этого волоски листа поднимаются, и лист снова готов к «охоте».
Многоклеточным растениям свойственны различные тропизмы как результат раздражимости их клеток под действием факторов внешней среды. Очень важен в жизни растения геотропизм: положительный геотропизм корней определяет их рост по направлению к центру Земли, а отрицательный геотропизм стеблей поднимает фотосинтезирующие органы растений вверх. Во втором случае играет роль и другой раздражитель — свет. Благодаря фототропизму растение растет вверх. Некоторые растения (например, подсолнечник) способны в течение дня поворачивать свое соцветие так, что оно все время обращено к солнцу (гелиотропизм).
Раздражимость высших многоклеточных животных часто принимает очень сложные формы и связана со специальной системой органов — нервной системой. Нервная система обеспечивает очень тонкое и разнообразное реагирование животного на всевозможные внешние раздражители. Одна из форм такого реагирования — активное движение животного.
По сравнению с многоклеточными животными реакции одноклеточных организмов и растений, возникающие в ответ на действие раздражителя, относительно просты: клетки их непосредственно взаимодействуют с внешней средой. У сложноорганизованных многоклеточных животных и у человека нервная система в процессе эволюции стала основным посредником между организмом и окружающей средой. Человек и животные получают информацию об изменениях внешней и внутренней среды посредством рецепторов — особых клеток, обладающих высокой чувствительностью к воздействию разнообразных раздражителей.
У человека 5 видов внешних рецепторов, которые известны вам из курса физиологии (вспомните и назовите их). Имеется и множество внутренних рецепторных клеток. Например, по всему телу рассеяны болевые рецепторные клетки, в стенках крупных кровеносных сосудов находятся чувствительные клетки, реагирующие на изменение концентрации CO2 в крови.
Раздражимость — один из основных признаков жизни. Пока организм жив, он раздражим. С прекращением жизни раздражимость исчезает. Огромное значение раздражимости клеток и организмов заключается в том, что она позволяет всем живым существам находиться в постоянной связи с окружающим миром, дает возможность приспосабливаться к нему. Раздражимость клеток связана в первую очередь с теми большими изменениями, которые происходят в белках, входящих в состав мембран цитоплазмы и ядра каждой клетки. При действии раздражителей, как это стало известно сейчас, происходят изменения в структуре белковых молекул. Способность к изменению структуры в ответ на действие раздражителей — это, по-видимому, одно из первичных элементарных свойств белков, которое возникло в процессе эволюции организмов.
Движение. В теснейшей связи с раздражимостью находится способность клеток и организмов совершать движения. Основу движения составляет сократимость цитоплазмы клеток. Сократимость — одно из основных свойств цитоплазмы живых клеток.
Как правило, растения неподвижно растут на одном месте, и исключение составляют только некоторые одноклеточные водоросли (например, диатомовые), способные к самостоятельному передвижению. Мы уже видели, что на действие таких внешних раздражителей, как свет, растения отвечают движениями листьев и побегов. Кроме того, у растений движения проявляются в росте.
В клетках всех растений постоянно происходит движение цитоплазмы. Эти движения называются токами цитоплазмы. Их можно видеть с помощью микроскопа у водорослей, в клетках листьев традесканции и в других растительных клетках. Токи цитоплазмы имеются также в клетках животных, и их легко наблюдать, например, у таких простейших, как инфузории.
Способность к передвижениям во внешней среде характерна для многих видов бактерий, простейших, для огромного большинства многоклеточных животных. У организмов, способных к передвижениям во внешней среде, различается 4 типа движения клеток: амебоидное, ресничное, жгутиковое и мышечное.
biofile.ru
ФИЗИОЛОГИЯ РАСТИТЕЛЬНОЙ КЛЕТКИ
Все клетки обладают такими свойствами живого как способность к самовоспроизведению, метаболизм (обмен веществ), раздражимость, рост, изменчивость и адаптация к внешней среде. Однако надо учитывать, что каждая клетка многоклеточного организма находится в тесном взаимодействии с другими клетками и что организм - это единое целое, а не сумма клеток.
Клетка обладает сложной структурной организацией и представляет собой систему, дифференцированную на отдельные органеллы. Растительная клетка имеет клеточную стенку и протопласт. Протопласт состоит из ядра с ядрышком, цитоплазмы и включенных в нее мембранных (вакуоль, пластиды, митохондрии, аппарат Гольджи, лизосомы, эндоплазматический ретикулум) и немембранных (микротрубочки, рибосомы) органелл. Все органеллы погружены в матрикс цитоплазмы - гиалоплазму или основную плазму (рис. 2.1).
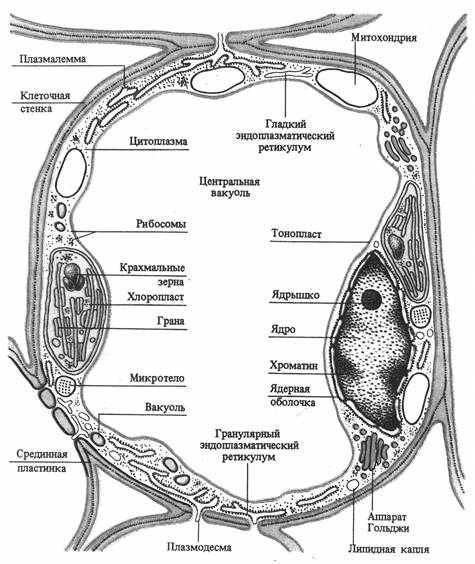
Рис. 2.1. Схема строения клетки мезофилла листа (по В. В. Полевому).
Все окруженные полупроницаемой мембраной компоненты клетки представляют собой замкнутые образования, в которых происходят разнообразные биохимические реакции. Цитоплазматический матрикс также разделен на отсеки эндоплазматической сетью. Тем самым достигается дополнительное пространственное разделение (компартментализация).
Основные физиологические функции структурных компонентов клетки: клеточная стенка - обеспечение прочности, защита, опорная функция; плазмалемма - транспорт веществ из и внутрь клетки, защита, рецепция; ядро - хранение и передача генетической информации; ядрышко - синтез РНК; вакуоль - осморегуляция, запасание веществ, переваривание; основная плазма - гликолиз; митохондрии - дыхание; хлоропласты – фотосинтез; аппарат Гольджи - секреция, образование компонентов клеточной стенки; эндоплазматическая ретикулум - транспорт веществ в клетке; лизосомы - внутриклеточное пищеварение; сферосомы - накопление и хранение жира; микротела (пероксисомы, глиоксисомы) - фотодыхание, глиоксалатный цикл; микротрубочки - ориентация микрофибрилл целлюлозы; рибосомы - синтез белка.
2.1. Раздражимость
Это способность клетки реагировать на действие внешних и внутренних факторов – раздражителей и передавать возбуждение в другие клетки. Восприятие раздражения обозначают термином перцепция или рецепция. У растений нет органов чувств, но есть рецепторные белки и клетки, воспринимающие разные воздействия. Различают фото-, хемо- и механорецепторы.
Возбуждение представляет собой измененное состояние клетки. В отсутствие раздражения растительная клетка имеет отрицательный потенциал покоя от -50 до -200 мВ, то есть протоплазма заряжена отрицательно по отношению к наружной поверхности. Это связано с тем. что внутри клетки находится больше ионов хлора и калия, но меньше ионов кальция, чем снаружи.
В ответ на раздражение возникает потенциал противоположного знака - потенциал действия, который может распространяться и на время сравняться или превысить потенциал покоя. Потенциал действия образуется в результате выхода ионов хлора из клетки и поступления ионов кальция в клетку.
Раздражение воспринимается клеткой, если его сила превышает пороговую величину. Следующие друг за другом подпороговые раздражения могут суммироваться, если интервалы между ними незначительны. Минимальное время, необходимое для рецепции, называют временем презентации.
Установлены следующие законы раздражимости:
1. Закон силы раздражения: чем больше раздражение, тем сильнее ответная реакция.
2. Закон длительности раздражения: чем длительнее раздражение, тем сильнее ответная реакция.
3. Закон количества раздражения: чем больше сила раздражения, тем меньше время презентации.
4. Закон градиента раздражения: чем выше скорость нарастания силы раздражителя, тем больше ответная реакция.
При действии чрезмерно сильных и (или) длительных воздействий чувствительность клеток к раздражителю снижается и клетки могут погибнуть.
Реституция - это восстановление исходного состояния после раздражения. Она идет с затратой энергии и тормозится под действием наркотиков, ингибиторов дыхания, при недостатке кислорода и снижении температуры. Во время реституции клетки находятся в рефрактерном периоде, когда новое раздражение не вызывает возбуждения.
2.2. Репликация, транскрипция и трансляция
Хранение наследственной информации осуществляется дезоксирибонуклеиновой кислотой (ДНК). ДНК - это полимер, мономерами которого являются дезоксирибонуклеотиды. В их состав входят углевод 2-дезокси-b-D-рибоза, остаток фосфорной кислоты и азотистые основания четырех типов: два пуриновых - аденин и гуанин, и два пиримидиновых - тимин и цитозин. Молекула ДНК состоит из двух полинуклеотидных цепочек, скрепленных между собой водородными связями между азотистыми основаниями. Полинуклеотидная цепочка образована чередующимися остатками 2-дезокси-b-D-рибозы и фосфорной кислоты, которая присоединена к 5 атому углерода кольца углевода с одной стороны и к 3 атому с другой стороны. Азотистые основания ответвляются от цепочки, присоединяясь к 1 атому углерода кольца углевода. Азотистые основания, противостоящие друг другу в полинуклеотидных цепочках, комплементарны друг другу: пуриновому основанию соответствует пиримидиновое (аденину – тимин, гуанину – цитозин). У каждой цепочки молекулы ДНК один конец заканчивается пятым (фосфатным), а другой - третьим (гидроксильным) углеродным атомом углевода. Они обозначаются как 5’ и 3’- концы. Цепочки в молекуле ДНК антипараллельны. В одной цепочке нуклеотиды связаны в направлении 5’ ® 3’, а в другой - 3’ ® 5’. Полинуклеотидные цепочки образуют двойную спираль. Каждый виток спирали содержит 10 пар азотистых оснований (рис. 2.2).

Рис. 2.2. Схема строения молекулы ДНК в виде двойной спирали (а) и прямой лесенки (б) (по В. Олфри и А. Мирскому).
1 – остаток 2-дезокси-β-D-рибозы, 2, 3, 4, 5 – азотистые основания (тимин, аденин, гуанин, цитозин, соответственно), 6 – остатки фосфорной кислоты.
Во время репликации (синтеза) ДНК каждая из цепей родительской ДНК служит матрицей для образования комплементарной дочерней цепи из предшественников – дезоксирибонуклеозидтрифосфатов: дезоксиаденозин-, дезоксицитидин-, дезоксигуанозин- и дезокситимидинтрифосфата. При их полимеризации происходит освобождение молекул пирофосфата, которые расщепляются пирофосфатазой. Репликация ДНК осуществляется по полуконсервативному механизму: одна из цепей дочерней молекулы ДНК является частью родительской молекулы, а другая – вновь синтезированной.
ДНК-полимераза способна синтезировать ДНК только в направлении от 5¢-конца к 3¢-концу. Поэтому на одной цепи ДНК образование дочерней цепи, называемой ведущей, происходит непрерывно, синтез другой цепи происходит прерывисто в виде коротких фрагментов Оказаки, получивших свое название в честь ученого, впервые их обнаружившего. Эта вторая дочерняя цепь получила название отстающей. Фрагменты Оказаки также синтезируются в направлении 5¢ ® 3¢, но перемещение ДНК-полимеразы вдоль матричной цепи ДНК при образовании каждого фрагмента противоположно направлению движения при синтезе ведущей цепи. Ведущая и отстающая цепи ДНК синтезируются координировано, что обеспечивается димеризацией ДНК-полимеразных комплексов у бактерий и наличием разных ДНК-полимераз у эукариотов. Для соединения двух фрагментов Оказаки ДНК-лигазой сначала удаляется РНК-затравка с помощью экзонуклеазы и РНКазы Н – нуклеазы, специфически расщепляющей РНК в ДНК-РНК-гибридах.
Процесс репликации ДНК разделяют на три этапа: инициацию, элонгацию и терминацию. Для начала репликации необходим праймер (затравка) – короткий олигодезокси- или олигорибонуклеотид, комплементарный соответствующему участку ДНК-матрицы.
В ходе репликации ДНК ее цепи расходятся из точки репликации, образуя Y-подобную структуру, которая называется репликативной вилкой. Именно в этом месте локализован репликативный комплекс, состоящий из нескольких белков. ДНК-хеликаза перемещается в репликативной вилке впереди ДНК-синтезирующего комплекса, расплетает цепи родительской ДНК и стимулирует образование затравок праймазой. ДНК-синтезирующий комплекс содержит ДНК-полимеразу, подвижный связывающий белок RPA и ДНК-зависимую АТФазу, которая узнает и связывается с единственным праймером ведущей цепи ДНК и с каждым праймером фрагментов Оказаки отстающей цепи, что делает возможным присоединение к праймерам ДНК-полимеразы и белка RPA. RPA связывается с образующимися одноцепочечными участками, облегчая процесс расплетения. RPA охватывает молекулу ДНК позади ДНК-зависимой АТФазы, стимулирует АТФазную активность и оставляет 3¢-конец праймера доступным для ДНК-полимеразы. ДНК-полимераза начинает элонгацию цепей ДНК, присоединяя первый дезоксирибонуклеозидмонофосфат к 3¢-концевому нуклеотиду РНК-затравки.
Передача информации от ДНК осуществляется посредством информационной или матричной рибонуклеиновой кислоты (мРНК). Синтез мРНК называется транскрипцией. Молекула мРНК, комплементарная одной из цепей матричной ДНК, образуется в ходе сополимеризации четырех рибонуклеозидтрифосфатов (аденин-, гуанозин-, цитозин- и урацилтрифосфата) с образованием 3¢-5¢-фосфодиэфирных связей и освобождением неорганического пирофосфата. Транскрипцию осуществляет фермент ДНК-зависимая РНК-полимераза. Синтез мРНК молекулами РНК-полимеразы начинается в определенных местах ДНК, называемых промоторами, и завершается на особых нуклеотидных последовательностях – терминаторах. Совокупность нуклеотидов ДНК, заключенных между промотором и терминатором, называют транскрипционной единицей или транскриптоном.
Процесс транскрипции подразделяют на 4 стадии: связывание РНК-полимеразы с ДНК и распознавание промотора, инициация, элонгация и терминация. Предполагается, что после первоначального непрочного связывания с ДНК в случайном месте молекула РНК-полимеразы перемещается вдоль двойной спирали ДНК до тех пор, пока не обнаружит последовательность нуклеотидов промотора. В этом месте связывание молекулы фермента с ДНК становится более прочным. Инициация транскрипции начинается с образования на промоторе предъиниционного комплекса, состоящего из РНК-полимеразы и матричной ДНК. После сборки предъинициационный комплекс претерпевает температурно-зависимые конформационные изменения, которые сопровождаются локальным плавлением, то есть расплетением двойной спирали ДНК, и комплекс становится способным к транскрипции. При наличии рибонуклеозидтрифосфатов происходит образование первых фосфодиэфирных связей в молекуле синтезируемой мРНК, после чего начинается стадия элонгации, то есть последовательное удлинение синтезируемой молекулы мРНК.
В 1992 г. М. Чэмберлен с сотрудниками разработали общую модель элонгации мРНК, согласно которой перемещение РНК-полимеразы вдоль ДНК и присоединение нуклеотидов к растущей цепи мРНК в активном центре фермента разделены во времени. Это разделение возможно потому, что у РНК-полимеразы имеется два сайта (участка), удерживающих растущую цепь мРНК, и два участка связывания ДНК-матрицы. Молекула РНК-полимеразы перемещается вдоль ДНК подобно гусенице: когда один сайт связывания ДНК фиксирован, другой перемещается вперед. ДНК-зависимые РНК-полимеразы фагов, состоящие из одной субъединицы, синтезируют РНК в условиях in vitro со скоростью 200-400 нуклеотидов в секунду. При перемещении фермента вдоль матрицы цепи ДНК подвергаются плавлению и повторному отжигу, в результате которого восстанавливается исходная структура ДНК. Стадия элонгации заканчивается после достижения РНК-полимеразой терминатора транскрипции. Затем синтезированная РНК и РНК-полимераза освобождаются из транскрипционного комплекса. Только минус-цепь ДНК служит матрицей для синтеза мРНК.
Участки ДНК, несущие информацию о строении белка - экзоны, разделены неинформативными интронами. В процессе транскрипции считывается информация как с экзонов, так и с интронов. Образуется предшественник мРНК - про-мРНК. Молекулы про-мРНК претерпевают созревание - процессинг. В ядре из про-мРНК происходит вырезание интронов и объединение экзонов - сплайсинг. К образовавшейся мРНК прикрепляется особая группировка поли-А и к одному из концов мРНК присоединяется защитная химическая группировка КЭП. После этого мРНК соединяется с белком, образуя инфорсому. Она выходит через поры в ядерной оболочке в цитоплазму. мРНК высвобождается из инфорсомы и одноцепочечная неспирализованная молекула мРНК присоединяется к участку малой субъединицы рибосомы, который примыкает к большой субъединице. К рибосоме прикрепляется небольшой участок цепи мРНК, содержащий один кодон, состоящий из трех азотистых оснований. Один кодон соответствует одной аминокислоте. Однако некоторые аминокислоты кодируются несколькими разными кодонами. Таким образом, первый этап синтеза белка - трансляции заключается в образовании комплекса между мРНК и рибосомой.
Перенос аминокислот в цитоплазме к рибосомам осуществляется транспортными РНК ( тРНК или 4S-РНК) из 70-80 нуклеотидов с молекулярной массой 25-30 кД. Они составляют почти 10 % от всей клеточной РНК и растворены в гиалоплазме. Поэтому тРНК еще называют растворимой РНК. В состав белков входят 20 аминокислот и каждой аминокислоте соответствует своя тРНК. Благодаря определенному расположению комплементарных нуклеотидов полинуклеотидная цепочка тРНК образует вторичную структуру, получившей название “клеверного листа”. Рентгеноструктурный анализ позволил установить третичную структуру тРНК. Она оказалась составленной из двух стеблей наподобие латинской буквы L.
Каждая тРНК имеет триплет оснований - антикодон, ответственный за прикрепление к комплементарному кодону мРНК. Для того, чтобы аминокислота присоединилась к тРНК необходима ее активация или обогащение энергией. Активация аминокислоты происходит за счет ее реакции с аденозинтрифосфорной кислотой (АТФ) - вещества с макроэргическими связями (~). Реакция идет при участии фермента аминоацилсинтетазы (кодазы). Образовавшийся аминоациладенилат остается связанным с ферментом и вступает в реакцию с тРНК с образованием аминоацил-тРНК.
Аминоацил-тРНК антикодоном присоединяется к кодону мРНК на малой субъединице рибосомы. После того как первая аминокислота со своей тРНК вошла в малую субъединицу рибосомы, происходит смыкание малой и большой субъединиц рибосомы. После смыкания субъединиц тРНК вместе с аминокислотой переносится на большую субъединицу. Одновременно мРНК перемещается на один кодон. В результате в малую субъединицу входит следующий кодон, кодирующий другую аминокислоту. К этому кодону с помощью антикодона присоединяется вторая тРНК со своей аминокислотой. В рибосоме оказываются две аминокислоты, ориентированные друг около друга таким образом, что карбоксильная группа первой аминокислоты оказывается рядом с аминогруппой второй аминокислоты. В результате сближения этих групп и с участием фермента большой субъединицы рибосомы пептидилтрансферазы аминокислоты соединяются пептидной связью. Образовавшийся дипептид присоединен ко второй тРНК, а первая тРНК освобождается и уходит в цитоплазму. В результате дипептид со второй тРНК оказывается связанным с большой субъединицей рибосомы, а мРНК перемещается еще на один кодон (рис. 2.3). Присоединение аминокислотных остатков - элонгация повторяется многократно, пока не образуется полипептидная цепочка белка. Окончание образования полипептидной цепочки - терминация связано с тем, что в малую субъединицу вступает терминальный кодон мРНК. Образовавшаяся полипептидная цепочка покидает рибосому. Большое значение имеет объединение рибосом в цепочки - полисомы. В этом случае одна молекула мРНК может последовательно присоединяться к ним и служить матрицей для синтеза нескольких одинаковых молекул белка. Когда синтез белка закончен, мРНК распадается. Синтез белковой молекулы идет с большой скоростью и поэтому время жизни мРНК невелико (от нескольких секунд до 1-2 минут). Однако, на определенных фазах развития растений (например, в набухающих и прорастающих семенах) синтезируются так называемые долгоживущие молекулы мРНК.

Рис. 2.3. Схема синтеза полипептидной цепи в рибосоме (по В. В. Полевому).
В заключение можно сказать, что из поколения в поколение передаются молекулы ДНК, которые несут в себе информацию о составе белковых молекул. План построения белка записан в ДНК с помощью триплетного кода, представленного чередованием азотистых оснований. Под влиянием внешних условий или спонтанно ДНК может изменяться. Эти изменения могут быть полезными, бесполезными и вредными. Полезные изменения, дающие организмам преимущество в борьбе за существование, могут закрепляться по наследству в ходе естественного или искусственного отбора.
Возникающие в процессе трансляции белки являются полимерами, мономерами которых служат аминокислоты. Белки представляют собой цепочки остатков аминокислот, соединенных между собой пептидными связями. Молекулярная масса белков зависит от количества мономеров и колеблется от нескольких тысяч до миллионов. Разнообразие белков определяется различной последовательностью аминокислотных остатков. Белки, также как и аминокислоты - амфотерные соединения, то есть имеют положительный и отрицательные заряды. У каждого белка своя изоэлектрическая точка - значение рН, при которой молекула белка не имеет заряда.
Молекула белка имеет первичную, вторичную, третичную и четвертичную структуру. Первичная структура определяется последовательностью аминокислотных остатков в белковой молекуле. Между аминокислотами, входящими в полипептидную цепочку, возможны водородные и другие связи. В результате этого полипептидная цепочка приобретает особенное расположение в пространстве, чаще всего в виде спирали. Это вторичная структура. По конформации полипептидной цепи различают фибриллярные и глобулярные белки. Фибриллярные белки сохраняют вытянутую форму спирали. У глобулярных белков спираль сворачивается в шарообразную глобулу, которая представляет собой третичную структуру белка. Она поддерживается кроме водородных связей гидрофобными взаимодействиями и дисульфидными связями, возникающими между двумя сульфгидрильными (SH) группами. Белки могут состоять из нескольких полипептидных цепочек и их взаимное расположение в пространстве и вокруг друг друга представляет собой четвертичную структуру.
2.3. Регуляции ферментативной активности в клетке
Химические процессы в клетке протекают с большой скоростью благодаря действию биологических катализаторов - ферментов или энзимов. Белковую часть фермента называют апоферментом. Небелковый компонент, прочно связанный, называют простетической группой, слабо связанный, обслуживающий несколько ферментов - коферментом. В состав простетических групп и коферментов входят металлы (железо, медь, цинк), витамины и их производные. Эндоферменты функционируют в клетке, а экзоферменты выделяются из клетки или локализуются в плазмалемме и действуют снаружи от нее.
Ферменты обладают специфичностью как к субстратам (субстратная специфичность), так и к определенным химическим реакциям (специфичность действия). Названия ферментов обычно заканчиваются суффиксом “аза”, за исключением некоторых общепринятых названий, например, пепсин, трипсин и другие. Название, как правило, отражает природу субстрата (хитиназа расщепляет хитин), или функцию фермента (аминотрансфераза переносит аминогруппы), или же то и другое вместе (алкогольдегидрогеназа). Ферменты разделяют на 6 классов: 1) оксиредуктазы - катализируют окислительно-восстановительные реакции, 2) трансферазы - перенос целых атомных группировок от одного соединения к другому, 3) гидролазы - распад органических соединений с участием воды, 4) лиазы - присоединение какой-либо атомной группировки к органическим соединениям или отщепление от субстратов определенной группы без участия воды, 5) изомеразы - превращение одних изомеров в другие, 6) лигазы или синтетазы - синтез органических соединений, происходящий при участии АТФ с использованием энергии этой кислоты.
Ферменты, катализирующие одну и ту же реакцию и встречающиеся у одного организма, но различающиеся по своим физико-химическим свойствам (например, по электрофоретической подвижности, следовательно, по молекулярной массе и заряду) называют изоферментами или изозимами. Наличие изоферментов позволяет организмам лучше приспосабливаться к меняющимся условиям внешней среды.
Катализ осуществляется в результате образования фермент-субстратного комплекса, что приводит к сближению реагирующих молекул или созданию напряженных химических связей путем их растягивания. Субстрат должен соответствовать активному центру не только пространственно, но и по распределению зарядов, расположению групп атомов и так далее. Окончательная подгонка происходит лишь в процессе взаимодействия субстрата с ферментом, претерпевающим при этой реакции конформационные изменения. Продукты реакции отделяются от фермента и молекулы фермента регенерируются. Благодаря своей способности регенерироваться, то есть возвращаться к первоначальному состоянию, одна и та же молекула фермента может катализировать большой объем превращений.
Скорость и направленность ферментативных реакций в клетке зависит от количества фермента, температуры и рН. Как всякая химическая реакция, ферментативные реакции зависят от температуры, что принято оценивать величиной температурного коэффициента (Q10), который показывает во сколько раз данный процесс ускоряется при повышении температуры на 10 Со. Поскольку ферменты являются белками, то повышение температуры свыше 35-40 Со вызывает их частичную инактивацию, а дальнейшее повышение температуры приводит уже к необратимой денатурации. У каждого фермента имеется свой оптимум рН, при котором лучше всего проявляется его активность. Это связано с тем, что рН влияет на заряд функциональных групп фермента, составляющих его активный центр, а от заряда зависит возможность образования фермент-субстратного комплекса.
Известны следующие механизмы внутриклеточной регуляции функционирования ферментов:
1. Метаболитная регуляция. Она происходит в результате изменения концентрации метаболитов и не затрагивает активность или число ферментных молекул. Различают регуляцию в местах разветвления путей обмена веществ и регуляцию по принципу обратной связи. В первом случае ферменты конкурируют за один и тот же субстрат и выбор пути определяется концентрацией общего для конкурирующих ферментов субстрата и степенью сродства фермента к субстрату. Под обратной связью понимается влияние более позднего члена цепи взаимосвязанных реакций на более ранний.
2. Ферментная регуляция. При этом типе регуляции изменяется активность ферментов. Изменение ферментативной активности может осуществляться несколькими путями: а) Обратимое или необратимое превращение неактивных предшественников ферментов - зимогенов в активные ферменты. Например, b-амилаза инактивирована в запасающих клетках эндосперма семян злаков из-за соединения с запасными белками посредством дисульфидных связей ( -S-S-). К началу прорастания семян из живых клеток алейронового слоя в эндосперм поступают вещества, разрушающие дисульфидные связи. Активированная b-амилаза принимает участие в гидролизе запасного крахмала; б) Изменение активности фермента под влиянием эффекторов. Связываясь с ферментом, эффекторы могут повышать его активность - это положительные эффекторы - активаторы или уменьшать ее - это отрицательные эффекторы - ингибиторы. Эффектор может влиять на активность фермента, взаимодействуя с активным центром (изостерический эффект) или изменяя конформацию ферментной молекулы в результате связывания с ее аллостерическим центром (аллостерический эффект). Изостерический эффект происходит в том случае, когда эффектор и субстрат похожи по своему строению и конкурируют друг с другом за активный центр фермента. Такой тип ингибирования называют конкурентным ингибированием.
3. Генная регуляция. В этом случае изменяется количество ферментных молекул в клетке из-за включения или выключения синтеза ферментов. Регулирующие факторы действуют на ДНК, РНК или рибосомы.
4. Мембранная регуляция. Различают контактную и дистанционную мембранную регуляцию активности ферментов. Контактная регуляция – связывание ферментов с мембранами или их освобождение меняет их активность. Дистанционная мембранная регуляция активности ферментов осуществляется косвенным путем в результате транспорта через мембраны субстратов и коферментов, удаления продуктов реакции, ионных и рН сдвигов в компартментах клетки.
2.4. Поступление воды в клетку
При температуре выше абсолютного нуля все молекулы находятся в постоянном движении. Диффузия - это процесс, ведущий к равномерному распределению молекул газов или растворенного вещества и растворителя благодаря их постоянному движению. Диффузия всегда направлена от большей концентрации вещества к меньшей. Количество вещества J, диффундирующего в единицу времени через воображаемое поперечное сечение, зависит от величины градиента dc/dx(градиент – мера изменения какого-либо параметра с расстоянием или временем) и от природы диффундирующего вещества, влияющего на коэффициент диффузии D. Это 1-й закон диффузии Фика, который описывается следующим уравнением:
J = - D · dc/dx,
так как суммарный поток направлен в сторону области меньшей концентрации, в уравнении стоит знак минус.
Скорость диффузии уменьшается с увеличением ее продолжительности. Проходимое путем диффузии расстояние пропорционально не времени, как при равномерном движении, а ее квадратному корню. Это 2-й закон диффузии Фика:
dc/dt = D · dc2/dx2,
где dc/dt – изменение концентрации вещества во времени.
Диффузия воды через полупроницаемую мембрану называется осмосом. Полупроницаемая мембрана - это мембрана хорошо проницаемая для воды и непроницаемая или плохо проницаемая для растворенных в воде веществ. Осмотическая ячейка - это пространство, окруженное полупроницаемой мембраной и заполненное каким-либо водным раствором. Все клеточные мембраны, в том числе плазмалемма и тонопласт, являются полупроницаемыми мембранами. Вода проходит в клетку через водные поры в плазмалемме, образованные специальными белками аквапоринами.
Внутри осмотической ячейки раствор развивает осмотическое давление p:
p = i · c · R · T,
где с – концентрация раствора в молях, Т - абсолютная температура, R - газовая постоянная 0,082 л · атм/град · моль, i – изотонический коэффициент, равный 1 + a(n-1), где a - степень электролитической диссоциации, n – число ионов, на которые распадается молекула электролита.
Благодаря осмотическому притоку воды в клетку там возникает гидростатическое давление, называемое тургорным. Это давление прижимает цитоплазму к клеточной стенке и растягивает ее. Клеточная стенка имеет ограниченную эластичность и оказывает равное противодавление. Эластическое растяжение ткани благодаря тургорному давлению ее клеток придает твердость не одревесневшим частям растений. Завядающие побеги становятся дряблыми, так как при потере воды тургорное давление падает. Тургорное давление противодействует притоку воды в клетку. Давление, с которым вода осмотически притекает в клетку, равно таким образом, разности между осмотическим давлением p и тургорным давлением P. Эту величину называют сосущей силой S: S = p - P. Вода поступает в клетку из внешнего раствора, если его потенциальное осмотическое давление меньше сосущей силы клетки и, наоборот, вода выходит из клетки в раствор с более высоким потенциальным осмотическим давлением.
При термодинамической трактовке сосущая сила заменяется водным потенциалом yw. Водный потенциал можно определить как работу, необходимую для того, чтобы поднять потенциал связанной воды до потенциала чистой, то есть свободной воды. Термин водный потенциал не совсем точен. Правильнее, но менее употребителен термин разность потенциалов воды, поскольку он определяется разностью между химическими потенциалами воды в системе mw (например, вакуоле) и чистой воды mow при атмосферном давлении. Абсолютные значения mw и mow неизвестны, но их разность можно определить. Она всегда отрицательна. Потенциал воды в растворе, растении, почве и атмосфере меньше 0. Потенциал чистой воды равен 0.
Можно также заменить p и P на потенциалы, а именно на осмотический потенциал yp (отрицательный) и потенциал давления yр (как правило, положительный). В таком случае осмотическое уравнение превращается в уравнение потенциала воды:
- yw = - yp - yр ( размерность бар = эрг · см-3 ·106)
Величину осмотического потенциала можно определить плазмолитическим методом. Плазмолиз - это процесс, обусловленный потерей воды клеткой. Он проявляется в отходе протопласта от клеточной стенки. В отдельных местах цитоплазма может в течение более или менее продолжительного времени сохранять связь с клеточной стенкой, образуя так называемые нити Гехта. Наблюдаются различные формы плазмолиза: выпуклый плазмолиз при небольшой вязкости цитоплазмы и вогнутый плазмолиз при высокой вязкости цитоплазмы (рис. 2.4). При переносе плазмолизированных тканей в гипотонический раствор или чистую воду вода поступает в клетку и происходит деплазмолиз. Количество воды в клетке увеличивается, объем вакуоли возрастает и она прижимает цитоплазму к клеточной стенке. Плазмолитический метод основан на подборе изоосмотического (изотонического) раствора, то есть имеющего осмотический потенциал равный осмотическому потенциалу клетки. Раствор, при котором начался плазмолиз, имеет осмотический потенциал примерно равный осмотическому потенциалу клетки. Зная концентрацию наружного раствора в молях, можно вычислить осмотический потенциал клетки.

Рис. 2.4. Формы плазмолиза.
1 – последовательные этапы плазмолиза в клетках листа мха, 2 – выпуклая форма плазмолиза (колпачковый плазмолиз) в клетке эпидермиса чешуи лука с окрашенной антоцианом вакуолью: а – ядро, б – цитоплазма, в – вакуоль (по Д. А. Сабинину – цит. по С. И. Лебедеву).
Иногда при сильном завядании протопласт не отстает от клеточной стенки как при плазмолизе, а сжимается и тянет ее за собой. При этом клеточная стенка прогибается. Это явление называют циторризом. Развивается натяжение или отрицательное давление стенки и потенциал тургорного давления приобретает отрицательное значение. В этом случае величина водного потенциала определяется уже не разностью, а суммой осмотического потенциала и потенциала давления: -yw = -yp + yp.
Величина осмотического потенциала позволяет судить о способности растения поглощать воду из почвы и удерживать ее, несмотря на иссушающее действие атмосферы. Осмотический потенциал колеблется у разных растений в пределах от -5 до -200 бар. У водных растений осмотический потенциал около -1 бара. У большинства растений средней полосы осмотический потенциал колеблется от -5 до -30 бар, растения степей и пустынь имеют более отрицательный осмотический потенциал. Осмотический потенциал различен и у разных жизненных форм. У деревьев он более отрицателен, чем у кустарников и травянистых растений, соответственно. У светолюбивых растений осмотический потенциал более отрицателен, чем у теневыносливых растений.
Поступление воды в клетку обусловлено не только осмотическим давлением, но и силой набухания. Набуханием называют поглощение жидкости или пара высокомолекулярным веществом (набухающим телом), сопровождаемое увеличением объема. Явление набухания обусловлено коллоидальными и капиллярными эффектами. В протоплазме преобладает набухание на коллоидальной основе (гидратация коллоидов), а в клеточной стенке наблюдаются оба эффекта: капиллярный - накопление воды между микрофибриллами и в межмицеллярных пространствах и коллоидальный - гидратация полисахаридов, особенно гемицеллюлоз.
У некоторых частей растений поглощение воды происходит исключительно путем набухания, например, у семян. Вода диффундирует в набухающее тело. Благодаря большому сродству набухающего тела к воде при набухании может возникать давление набухания в несколько сотен атмосфер. Силу набухания обозначают термином матричный потенциал yt.
Таким образом, для клетки характерны следующие уравнения водного потенциала:
вакуоль: -yw = -yp - yp
протоплазма: -yw = -yp - yp - yt
клеточная стенка: -yw = - yt
Вода в клетку может поступать также в процессе пиноцитоза, когда часть плазмалеммы под влиянием различных причин, чаще всего в результате адсорбции на плазмалемме крупных молекул и вирусных частиц, прогибается внутрь клетки, внешние края такой инвагинации смыкаются и виде пузырька - везикулы с адсорбированной частицей и внешним раствором проходит внутрь цитоплазмы.
2.5. Поступление ионов в клетку
Все неорганические питательные вещества поглощаются в форме ионов, содержащихся в водных растворах. Поглощение ионов клеткой начинается с их взаимодействия с клеточной стенкой. Ионы могут частично локализоваться в межмицеллярных и межфибриллярных промежутках клеточной стенки, частично связываться и фиксироваться в клеточной стенке электрическими зарядами.
Поступившие ионы легко вымываются. Объем клетки, доступный для свободной диффузии ионов, получил название свободного пространства. Свободное пространство включает межклетники, клеточные стенки и промежутки, которые могут возникать между клеточной стенкой и плазмалеммой. Иногда его называют кажущееся свободное пространство (КСП). Термин “кажущееся” означает, что его объем зависит от объекта и природы растворенного вещества. КСП занимает в растительных тканях 5-10 % объема. Свободное пространство всего растения получило название апопласт, в отличие от симпласта - совокупности протопластов всех клеток.
Поглощение и выделение веществ в КСП - физико-химический пассивный процесс, не зависимый от температуры и ингибиторов энергетического и белкового обменов. Клеточная стенка обладает свойствами ионообменника, так как в ней адсорбированы ионы Н+ и НCO-3, обменивающиеся в эквивалентных количествах на ионы внешнего раствора. В клеточную стенку входят амфотерные белковые соединения, заряд которых меняется при изменение рН. Поэтому адсорбция ионов зависит от величины рН. Из-за преобладания отрицательных фиксированных зарядов в клеточной стенке происходит первичное концентрирование катионов (особенно двух- и трехвалентных).
Для того, чтобы проникнуть в цитоплазму и включиться в обмен веществ, ионы должны пройти через плазмалемму. Транспорт ионов через мембрану может быть пассивным и активным. Пассивное поглощение не требует затрат энергии и осуществляется путем диффузии по градиенту концентрации вещества, для которого плазмалемма проницаема. Пассивное передвижение ионов определяется не только химическим потенциалом m, как это имеет место при диффузии незаряженных частиц, но и электрическим потенциалом e. Оба потенциала объединяют в виде электрохимического потенциала `m: `m = m + nFe, где m - химический, e - электрический, `m - электрохимический потенциалы, n - валентность иона, F - константа Фарадея. Любая разность электрических потенциалов, которая возникает на мембранах, вызывает соответствующее перемещение ионов.
Электрический потенциал на мембране - трансмембранный потенциал может возникнуть по следующим причинам: 1) если поступление ионов идет по градиенту концентрации, но благодаря разной проницаемости мембр
Похожие статьи:
poznayka.org
2. Раздражимость и реакция клетки на повреждающее воздействие
Жизнь клетки протекает в постоянном взаимодействии с окружающей средой, изменения в которой не могут не отражаться на функциональном состоянии клетки. В определенном диапазоне действия внешнего фактора уровень клеточных функций меняется градуально. Если же изменения превышают обычную норму, то внешний агент становится раздражителем. Раздражитель - это внешнее воздействие, достигшее пороговой силы. Способность живых структур отвечать на действие раздражителя называется раздражимостью.
В качестве раздражителей может выступать любой вид энергии - механическая, химическая, электрическая, световая, тепловая. Сила ответной реакции, эффект раздражения, определяется количеством раздражения - произведением силы раздражителя на время его действия. Т.о., слабый агент, если он действует достаточно долго, может оказывать такое же действие, как и сильный. Другим важным свойством раздражимости является суммация раздражения - отдельные внешние воздействия различной напряженности могут суммироваться, достигая порогового значения. Так, усик вьющегося растения совершает в воздухе колебательные движения. Если подвести к нему палочку, то вначале он, столкнувшись с ней, отталкивается несколько раз. Когда в результате столкновений происходит суммация раздражения и достигаемся пороговая сила, возникает ответная реакция и усик обвивается вокруг опоры.
Явления суммации раздражения имеет большое приспособительное значение. Если бы каждое слабое воздействие вызывало ответную реакцию, то этих реакций было бы бесчисленное множество и организм тратил бы слишком много энергии. Суммация раздражения позволяет экономно расходовать энергию, затрачивая ее лишь в ответ на раздражитель достаточной силы.
Русский физиолог Н. В. Введенский впервые отметил единство признаков возбуждения и повреждения (1901). Д. Н. Насоновым и В. Я. Александровым было установлено, что в ответ на раздражители и повреждающие агенты в клетке возникают неспецифические (не зависящие от природы действующего агента изменения, которые проявляются рядом ответных реакций) и специфические (будут рассматриваться в разделе «Устойчивость»).
НЕСПЕЦИФИЧЕСКИЕ РЕАКЦИИ:
Уменьшение степени дисперсности цитоплазмы («оструктуривание»). Так, при малом и среднем увеличениях микроскопа цитоплазма не видна из-за высокой степени дисперсности Однако, если клетку уколоть, добавить гипертонический раствора подействовать электрическим током, цитоплазма сразу станет заметной, так как размеры составляющих ее частиц увеличатся.
Увеличение общей проницаемости. При повреждающем воздействии из клетки в окружающую среду выходят самые разнообразные вещества, которые в неповрежденной клетке прочно удерживаются: чем сильнее увеличивается проницаемость, тем больше степень повреждения. На этом основаны методы диагностики устойчивости к морозу, засухе и другим неблагоприятным факторам среды. Растительные ткани погружают в 6идисцтиллированную воду, после чего анализируют состав образовавшуюся раствора(табл. ).
Экзоосмос электролитов из тканей узла кущения растений озимой пшеницы, различающихся по морозоустойчивости (по Э.А.Барашковой и др., 1983)
| Сорт | Морозоустойчивость по методу прямого промораживания | Экзоосмос электролитов после промораживания (% от экзоосмоса из непромороженных растений) |
| Тормовчанка | Выше средней | 264 |
| Лютестенс 1060/10 | То же | 368 |
| Московская озимая | То же | 378 |
| РПГ 51/49 | Средняя | 579 |
| Безостая 1 | То же | 500 |
| Лада | Слабая | 501 |
| Гектор | То же | 603 |
Повышение у цитоплазмы и ядра сродства к витальным (прижизненным) красителям. На этом показателе основан способ определения жизнеспособности семян (тетрозол).
Неспецифичность ответных реакций на раздражители и повреждающие агенты разного рода связана с общими изменениями в структуре клеточных белков. Любой из внешние факторов, достигших пороговой силы, нарушает слабые связи, поддерживающие специфическую конформацию молекул. Упрощение структуры, «развертывание» глобулы представляет собой денатурацию, которая может быть обратимой или необратимой (коагуляция). Раскручивание белковых глобул приводит к уменьшению дисперсности цитоплазмы, повышению вязкости, сродства к красителям, дестабилизации мембран. Последнее обстоятельство ярко проявляется еще в одной неспецифической реакции клетки: при действии раздражителя и повреждающего фактора МП падает.
При достижении внешним агентом пороговой силы МП начинает снижаться, иногда происходит перезарядка мембраны - поверхность клетки негативируется. Так, при действии ядом цианидом на колеоптили овса МП упал с -119 до -42 мВ: в отсутствие кислорода МП гипокотиля вигны составил лишь -40 мВ, тогда как в контрольных условиях он был равен -111 мВ.
В ответ на сильное раздражение (механическое, химическое, температурное, электрическое, фото- и радиостимулы) может возникнуть другая биоэлектрическая реакция - потенциалы действия (ПД). Суть этого явления. При резком изменении какого-либо фактора увеличивается ионная проводимость мембраны; за счет входящего в клетку Са2+ происходит деполяризация мембраны, при этом внешняя поверхность может стать отрицательной по отношению к внутренней. Затем за счет выходящего ионного тока К+ потенциал возвращается к исходному уровню - возбужденный участок мембраны сам становится источником возбуждения для соседнего участка, где повторяются уже описанные двухфазные изменения. Потенциалы как бы бегут вдоль мембраны (рис.). Так возникает и распространяется ПД, с помощью которого по растению передаются сигналы о происходящих во внешней среде изменениях. После достижения ПД какого-либо органа наступает ответная физиологическая реакция. Особенно хорошо она проявляется у «чувствительных» растений. Например, у мимозы вслед за механическим раздражением листа регистрируется ИД, который достигая листовой подушечки, вызывает в ней изменения тургора, что, в свою очередь, приводит к опусканию листочков.
И у обычных растений ПД вызывает физиологический эффект. В МСХА профессором П.С.Беликовым (1964) было показано, что раздражение корней растений пшеницы концентрированными солями или высокой температурой генерируют ПД, который, распространяясь достигает листа. Этот сигнал вызывает изменения фотосинтетической активности.
Не только биопотенциалы, но и другие показатели при действии раздражителей изменяются волнообразно, двухфазно. Так, сразу же после начала теплового воздействия вязкость цитоплазмы сначала уменьшается, а уже затем увеличивается: скорость движения цитоплазмы сначала возрастает, а потом снижается; выход веществ из клетки изменяется волнообразно (рис. ).
Колебательный характер ответных реакций объясняется тем, что при действии на клетку раздражителя в ней одновременно наблюдается несколько функциональных состояний (альтераций) - повреждение, возбуждение, закалка, репарация. Повреждение выражается в нарушении структуры и функций клетки. Возбуждение приводит к активации клеточной деятельности, в результате чего возникает закалка - действие раздражителя воспринимается в меньшей степени. На фоне закалки происходят восстановление исходных структур и функций - репарация. Клетка адаптируется к данному уровню внешнего фактора. Перечисленные процессы имеют разную направленность и противоречивый характер. В результате их взаимодействия переход клетки из одного состояния в другое не бывает плавным, а представляет собой колебательный процесс.
Характерными особенностями временного хода колебательного процесса являются время наступления ответной реакции, амплитуда и длина волны. Уровень каждого из этих параметров может служить критерием силы повреждения. Эти параметры могут быть использованы для диагностики устойчивости к различным воздействиям (рис. ).
Изменение структуры растительной клетки при действии патогенных микроорганизмов.
Агроном должен четко представлять, что происходит в растительном организме на всех уровнях его организации при заболевании. Знание ответных реакций на действие патогена поможет правильно оценить степень заболевания, своевременно применить нужные агротехнические меры, наконец, выявить иммунные формы, сорта и индивидуальные растения, необходимые при селекции на устойчивость к болезням.
Влияние инфекции на клеточные структуры значительно и разнообразно. Однако клетка устойчивого организма способна преодолевать пагубное действие инфекции, выздоравливать, и это выражается в сохранении и новообразовании клеточных структур и восстановлении их функций.
Типичные изменения в клетках иммунных и неиммунных форм растений при действии инфекции.
| Структура, св-ва клетки | Неиммунные формы | Иммунные формы |
| Проницаемость протоплазмы | Сильно возрастает | Изменяется мало |
| Гидрофильность цитоплазмы | уменьшается | Слабо меняется |
| Ядро | Уменьшается в размерах | Сначала уменьшается, затем увеличивается |
| Пластиды | Набухают ламелы и граны разрушаются | Структура сохраняется |
| Митохондрии | Набухают, кристы исчезают | - // - |
| Рибосомы | Новых нет | новообразование |
Одним из важнейших условий, обеспечивающих жизнь, является способность к внутренней регуляции клеточных процессов. Существует несколько систем регулирования: внутриклеточные, межклеточные и организменные. Их совместная деятельность приводит к быстрым, направленным и экономичным перестройкам, обеспечивающие поддержание гомеостаза и оптимальный уровень функций в условиях непостоянной внешней среды при выполнении наследственной программы.
Внутриклеточные системы регуляции: регуляция активности ферментов, генетическая, мембранная. В основе их лежит рецепторно-конформационный принцип.
Генетическая. Обеспечивает включение и выключение отдельных генов, ответственных за синтез специфических белков, и осуществляется за счет уникальной структуры ДНК и саморегуляции её деятельности.
Мембранная регуляция осуществляется благодаря сдвигам в мембранном транспорте, связыванию или освобождению ферментов и регуляторных белков и путем изменения активности мембранных ферментов. Особое значение имеет система мембранных фото-, хемо- и механорецепторов, позволяющих оценить изменения внешней среды и в соответствии с ними изменять свойства мембран.
Энергетическая. Выполняется системами, ответственными за новообразование энергоемких молекул, прежде всего АТФ, способные снабжать энергией все виды работ в клетке (в т. ч. - синтетические).
Межклеточные системы регуляции развиваются с появлением многоклеточных организмов и включают: трофическую, гормональную и электрофизиологическую.
Трофическая - осуществляет наиболее простой способ связи между клетками, тканями и органами. Она носит количественный характер. Реализуется с помощью химических веществ, продуктов метаболизма (наземные органы зависят от МВ, поступающих от корней, подземные - от ассимилятов, образующихся в листьях). Они могут использоваться как источники синтетического и энергетического обмена, а также выполнять в клетке «тонкую настройку», выступая как регуляторы ферментативной активности.
Гормональная регуляция осуществляется за счет фитогормонов, в малых количествах необходимых для запускания физиологических и морфо-физиологических процессов - один из самых тонких механизмов структурных и функциональных изменений. О специфических веществах - гормонах - и принципах гормонального регулирования процессов жизнедеятельности будет рассказано в разделе «Рост и развитие растений».
Электрофизиологическая система (является аналогичной нервной системе животных). Между различными частями растений существуют стационарные (точнее медленно изменяющиеся разности потенциалов (электротонические поля и токи), Наблюдаются местные и распространяющиеся ПД.
Электротонические поля и токи: верхушечная почка заряжена полдожительно по отношению к базальным частям; центр стебля электроположителен по отношению к наружной поверхности; верхушка колеоптиля отрицательна по отношению к основанию; у корней и корневых волосков заряд положительный; наземная часть +, корни - (ток порядка 0,1...0,4 мкА). В опытах под влиянием эл. Тока активируется Н+-помпа, повышается метаболическая активность, происходят изгибы, удлинение.
Потенциалы действия
studfiles.net
Ответ растений на раздражения - СТРОЕНИЕ И ЖИЗНЕДЕЯТЕЛЬНОСТЬ - РАСТЕНИЯ - Биология 7 класс
Раздел 1 РАСТЕНИЯ
Тема 1. СТРОЕНИЕ И ЖИЗНЕДЕЯТЕЛЬНОСТЬ
15. Ответ растений на раздражения
Основное понятия: РАЗДРАЖИМОСТЬ
Мы должны сделать вывод, что если проростки освещаются сбоку, то от их верхней части к нижней передается некий стимул, что заставляет последнюю выгибаться.
Ч. Дарвин
Знакомьтесь
Английский биолог Чарлз Дарвин (1809-1882) первым доказал наличие раздражения у растений (ил. 15.1). По его мнению, у растений есть две зоны — чувствительная и двигательная. Первая зона воспринимает раздражение, а другая — совершает движения. Например, при действии света на проростки овса их верхушка осуществляет восприятие этого влияния, а участок, которая расположена под верхушкой, отвечает на это раздражение, изгибаясь в сторону света.

Ил. 15.1. Ч. Дарвин
ОСНОВНОЕ СОДЕРЖАНИЕ
Раздражимость — это способность живых клеток, тканей или целого организма реагировать на внешние или внутренние воздействия. Этот признак живых существ взято за основу их приспособления к условиям среды существования. Растения, как и все организмы, способны отвечать на различные изменения окружающей среды изменениями в своем организме. Например, листья комнатного растения, которое стоит на подоконнике, повернуты к свету. Если вы развернете вазон другой стороной к свету, то через некоторое время листья снова будут обращены в сторону лучшего освещения. Изменения положения тела растения или его частей в пространстве называют движением. Движения растений обусловлены различными факторами, в частности, влиянием света, температуры, влаги, химических веществ. Движения бывают положительными (в направлении к фактору) или негативными (в направлении от фактора), активными и пассивными. Пассивные движения возникают под воздействием ветра, течений воды (например перелеты плодов одуванчика) и тому подобное. Активные движения связаны с ростом и развитием растений. Ростовые процессы связаны с сезонными изменениями в природе характеризуются ритмичностью. Растениям свойственно состояние покоя, когда рост и развитие прекращаются (ил. 15.2).

Ил. 15.2. Движения растений
Как растения отвечают на раздражение?
Раздражение — это внешнее или внутреннее вгинів, который вызывает движение растения. Факторы, которые оказывают такое влияние, называют раздражителями (свет, прикосновение и тому подобное). Эти раздражения во многих случаях вызывают возбуждение в клетках, что проявляется в повышении их деятельности, следствием которой могут быть деление и рост клеток, изменение давления в середине клеток, которые и обусловливают движения растений. Например, в комахоїдної растения росянки круглолистной, которая растет на сфагновых болотах и торфяниках, на листьях есть липкие волоски, на кончиках которых сверкают прозрачные капли жидкости, которые привлекают мелких насекомых. Как только насекомое касается этих волосков, она прилипает к капель густой слизи и, пытаясь освободиться, раздражает другие волоски. Механическое раздражение приводит к возникновению возбуждения в листке, что предопределяет загибания его краев внутрь. Листок, совершая сократительные движения, медленно окутывает со всех сторон свою жертву и переваривает ее с помощью веществ, которые выделяют другие волоски.
Какое значение имеют ростовые движения растений?
Ростовые движения — это активные движения растений, связанные с процессами роста. У большинства растений они охватывают лишь отдельные органы — корень, побег, цветок. Ростовые движения являются результатом быстрого роста клеток на каком-то одном боку органа под влиянием факторов среды (ил. 15.3). Причиной ростовых движений у растений является изменение условий существования в течение суток, прежде всего освещенности и температуры. Ростовые движения разделяют на тропізми и настії. Тропізми — это ростовые движения в направлении, определяемом односторонним воздействием определенного фактора среды. Эти движения могут быть направлены в направлении к раздражителю (движение корзин подсолнечника в сторону к солнцу и т. п) или от него (рост корней плюща в направлении, противоположном направлению света). Результатом таких движений является то, что растение занимает в пространстве я «благоприятное положение и избегает всего опасного для ее жизни. Настії — это ростовые движения в направлении, что определяется внутренними факторами, а внешние воздействия лишь обусловливают их возникновение. Они бывают обусловлены неравномерным ростом нижней и верхней стороны листа, лепестков. их можно наблюдать в течение суток, когда свет периодически сменяется тьмой. У некоторых растений цветки раскрываются утром, а на ночь закрываются. Такие «сонные движения» цветков осуществляются в ответ на изменение света или температуры. Корзинки одуванчика, цветки кувшинки белой па ночь закрываются, а утром раскрываются. А у маттиолы и табака душистого цветки закрываются днем, а раскрываются ночью.
Какое значение имеют сократительные движения у растений?
Активность растений, которые ведут прикрепленный образ жизни, может быть связана не только с ростовыми, но и со скоротливими движениями. Сократительные движения - это активные движения растений, обусловленные изменением давления внутри определенных групп клеток, в результате чего изменяются их размеры. Примером таких движений является опускание листочков мимозы застенчивой при прикосновении к ним, раскрытие цветков тюльпана при переносе с холода в тепло, заглубление луковицы в грунт в течение осени при помощи сократительных корней и тому подобное. В бобовых (фасоль, клевер и др) в листьях есть слоеные подушечки. Это особые творения при основании черешка или листочка, где содержатся крупные клетки. Быстрая смена давления в верхних и нижних клетках вследствие перемещения воды приводит к тому, что листовая подушечка работает как шарнир, с помощью которого листья то опускаются, то поднимаются. Итак, ростовые и сократительные движения являются основными активными реакциями на раздражение факторов среды.

Ил. 15.3. Опыт, который демонстрирует ростовые движения растений
САМОПРОВЕРКА И ОБОБЩЕНИЯ ЗНАНИЙ
Содержание материала | Вы освоили эту тему, если можете: |
4. Общая характеристика растений | — назвать самые общие особенности строения и процессы жизнедеятельности растения; — указать условия, необходимые для жизнедеятельности растения; — привести примеры влияния условий среды на основные процессы жизнедеятельности растения |
5. Клетка растений | — назвать основные компоненты клетки растений; — объяснить, почему клетка является живой системой; — наблюдать и описывать строение клеток растения |
6. Ткани растений | — назвать растительные ткани; — указать значение образующих тканей для растения; — объяснить взаимосвязь строения и функций постоянных тканей растения |
7. Органы растений | — назвать органы растений; — указать значение вегетативных органов; — объяснить значение для репродуктивных органов растения |
8. Строение и функции корня | — назвать виды корней и типы корневых систем; — охарактеризовать строение корней; — назвать растения, имеющие корнеплоды, бульбокорени, корни - придирки, корни-присоски и т. п |
9. Минеральное питание растений | — объяснить значение минерального питания для растения; — охарактеризовать почва как источник питательных веществ; — привести примеры использования удобрений в сельском хозяйстве |
10. Побег и его строение | — охарактеризовать роль побега в обеспечении поступления, транспорта и выведения веществ; — назвать основные виды почек; — назвать растения, имеющие надземные и подземные видоизменения побега |
11. Стебель — ось побега | — назвать основные функции стебля; — объяснить, чем определяются особенности внутреннего строения стебля; — указать значение ветвления стебля |
12. Транспорт веществ в растении | — описать опыт, который дает возможность наблюдать транспорт веществ по растению; — обосновать необходимость отложения запасных веществ; — сделать вывод о связи органов в растительном организме |
13. Лист — боковая часть побега | — назвать растения с простыми и сложными листьями; — охарактеризовать строение листа и его роль в обеспечении процесса фотосинтеза, дыхания и испарения воды; — сравнить типы жилкования и листкорозміщення |
14. Функции листка | — сравнить фотосинтез и дыхание; — описать опыт, который дает возможность наблюдать испарение воды; — назвать растения, которые имеют видоизменения листа |
15. Ответ растений на раздражения | — объяснить значение раздражимости для растений; — описать опыты, которые дают возможность наблюдать ростовые движения растений; — привести примеры разных движений растений |
schooled.ru
Как клетки воспринимают раздражения
В сложных организмах некоторые клетки высоко специализированы для обнаружения изменений в окружающей среде. Свойства таких клеток были выявлены при изучении зрительных рецепторов мечехвоста.
Выживание каждого живого существа зависит в конечном счете от его способности реагировать на окружающий его мир и соответственно регулировать свою собственную внутреннюю среду. У большинства многоклеточных животных реакции на внешние воздействия и регуляция осуществляются специализированными рецепторными клетками, которые чувствительны к весьма разнообразным физическим, химическим и механическим раздражениям.
У многих животных, а также и у человека эти рецепторы обеспечивают информацию, которая значительно превосходит информацию, получаемую при помощи традиционных пяти органов чувств (органы зрения, слуха, обоняния, вкуса и осязания). Не чем иным, как органами чувств, следует считать также и рецепторы, управляющие изменениями внутренней среды (хотя мы обычно и не отдаем себе отчета в их существовании). Рецепторные органы в мышцах, называемые мышечными веретенами, дают непрерывную информацию относительно степени мышечного растяжения, а другие рецепторы ощущают движения суставов. Не будь этих рецепторов, нам было бы трудно двигаться или говорить. Рецепторные клетки гипоталамуса — одной из частей головного мозга — воспринимают изменения температуры крови, а особые чувствительные клетки каротидного синуса — изменения давления крови. Существуют также внутренние рецепторы, регулирующие уровень углекислоты в особых участках крупных артерий. Болевые рецепторы, широко распространенные по всему организму, реагируют на вредные воздействия почти любого типа, которые могут оказаться пагубными для организма.
Рецепторные клетки различаются не только по своим функциям и структуре, но также по характеру своих связей с нервными волокнами, соединяющими их с центральной нервной системой. Некоторые рецепторные клетки непосредственно образуют собственные нервные волокна; другие же образуют связи с нервными волокнами иного происхождения. Однако для всех рецепторов характерна одна общая функция: генерация нервных импульсов. Это не значит, что импульсы непременно возникают в самих рецепторных клетках. Например, никому еще не удалось обнаружить импульсы в фоторецепторах глаза позвоночных — в палочках и колбочках. Тем не менее при раздражении палочек и колбочек светом в них создаются физико-химические условия, вызывающие возникновение импульсов в лежащих позади нервных клетках. Типичные нервные импульсы легко обнаружить в самом зрительном нерве, который состоит из отростков ганглионарных клеток, отделенных от палочек и колбочек по крайней мере одной промежуточной группой нервных клеток.
Физиологи надеются когда-нибудь распутать всю сложную цепь событий, в результате которой рецепторная клетка в ответ на механическую деформацию, поглощение света или тепла или же раздражение определенной молекулой дает начало нервному разряду. Ни в одном случае не удалось проследить все эти события. Мы начнем наше обсуждение с последнего звена, общего для всех сенсорных рецепторов — с возникновения нервных импульсов. Затем мы подробно рассмотрим события, происходящие в одном определенном рецепторе — фоторецепторе мечехвоста (Limulus). Наконец, мы попробуем дать некоторые характеристики «производительности» рецепторов, действующих поодиночке и совместно с другими рецепторами.
Нервное волокно, или аксон, представляет собой нитевидное образование, отходящее от тела нервной клетки. Вся поверхностная мембрана клетки, включая и оболочку аксона, электрически поляризована; внутреннее содержимое клетки заряжено отрицательно по отношению к внешней среде, причем заряд этот составляет около 70 тысячных вольта. Эта разность потенциалов носит название мембранного потенциала. В ответ на соответствующее раздражение немедленно происходит местное изменение мембранного потенциала, дающее начало нервному импульсу, который затем распространяется по всему волокну.
В любом нервном волокне импульсы, по существу, одинаковы по величине и форме и распространяются с одинаковой скоростью. Это было впервые установлено лет 30 назад работами Э. Эдриана. Эдриан и его сотрудники обнаружили, что, изменяя интенсивность раздражения, приложенного к рецепторной клетке, можно вызвать изменение не величины импульсов, а только их частоты; чем выше интенсивность, тем больше частота нервных импульсов, порождаемых рецептором. Таким образом, все сенсорные восприятия, связанные со светом, звуком, положением мышц и т. п., передаются при помощи одного и того же кода, состоящего из единичных нервных импульсов. Животное способно декодировать различные сообщения, потому что рецептор каждого типа сообщается с высшими нервными центрами только при помощи своего особого набора нервных путей.
Эдриан и другие авторы изучали триггерную функцию, рецепторных клеток по отношению к сенсорным нервным импульсам. Эдриан предполагает, что рецептор каким-то образом уменьшает мембранный потенциал покоя своего нервного волокна; иными словами, он, по-видимому, вызывает местную деполяризацию оболочки аксона. Существование местных потенциалов в глазу было известно еще в 1865 г.; в других органах чувств подобные потенциалы были зарегистрированы гораздо позднее. Однако связь этих электрических изменений с возникновением нервных импульсов оставалась неясной. У некоторых животных с простыми глазами полярность местных изменений потенциала в рецепторах носит такой характер, что они, по-видимому, приводят к деполяризации сенсорных нервных волокон. Ввиду этого Р. Гранит предложил называть их «генераторными потенциалами». Согласно современной точке зрения, раздражение рецепторных клеток приводит к устойчивой местной деполяризации сенсорных нервных волокон, которые в результате генерируют серию импульсов.
Первые прямые указания относительно наличия генераторных потенциалов на клеточном уровне были получены одним из авторов этой статьи — Хартлайном. Он обнаружил нечто напоминающее генераторный потенциал, когда ему удалось зарегистрировать активность одиночного волокна зрительного нерва и его рецептора в сложном глазе мечехвоста. На этот потенциал налагалась серия нервных импульсов.
В 1950 г. Катц получил несомненные доказательства наличия генераторных потенциалов в более простом рецепторе — мышечном веретене позвоночных. При растяжении мышечного веретена удавалось зарегистрировать незначительную, но устойчивую деполяризацию в отходящем от него нервном волокне. При наблюдении с помощью осциллографа создавалось впечатление, что основная линия зарегистрированного сигнала слегка смещается вверх. На этот смещенный сигнал, или местный потенциал, налагалась серия «пиковых потенциалов», представляющих собой отдельные нервные импульсы. Чем сильнее было растяжение или чем быстрее оно производилось, тем сильнее был сдвиг потенциала и тем выше частота импульсов. Анализ многих подобных записей показал, что при постоянных условиях частота нервных импульсов прямо пропорциональна величине измененного потенциала. Если подвергнуть веретено местному наркозу, то импульсы исчезают, но сдвиг потенциала сохраняется. Катц пришел к выводу, что этот сдвиг потенциала представляет собой основное связующее звено между растяжением веретена и разрядом нервных импульсов; иными словами, он действительно представляет собой генераторный потенциал. Более того, этот потенциал можно обнаружить только в непосредственной близости от веретена; это означает, что он распространяется по нервному волокну пассивно, т. е. весьма плохо.
Важные данные, подтверждающие роль генераторного потенциала, получены С. Каффлером и К. Эйзагиром. Эти ученые использовали так называемые клетки Александровича — рецепторные клетки ракообразных, воспринимающие растяжение. Это большие одиночные рецепторные клетки, дендриты (короткие волокна) которых погружены в специализированные рецепторные мышцы. Каффлеру удалось ввести микроэлектрод в клетку и зарегистрировать ее мембранный потенциал, а также нервные импульсы в ее аксоне. Оказалось, что, нарушив форму дендритов путем растяжения рецепторной мышцы, можно вызвать деполяризацию тела клетки; эта деполяризация пассивно распространяется к месту возникновения импульса, которое, возможно, находится на аксоне, вблизи от места выхода последнего из тела клетки. Когда этот генераторный потенциал достигал критического уровня, клетка посылала серию нервных импульсов; чем больше деполяризация аксона превышала критический уровень, тем выше была частота импульсов.
Теперь мы располагаем многочисленными данными, указывающими на то, что рецепторная клетка служит пусковым механизмом, вызывающим возникновение нервных импульсов путем местной деполяризации близлежащего нервного волокна — либо своего собственного волокна, либо волокна какой-либо другой клетки. За немногими исключениями, отдельные волокна какого-либо нервного ствола неспособны многократно реагировать на непрерывно проходящий через него деполяризующий ток; они реагируют лишь в течение короткого времени, образованием одного или нескольких импульсов, а затем привыкают к раздражению и перестают реагировать на него. По-видимому, часть сенсорного нервного волокна, расположенная близ рецептора, обладает особой специализацией, благодаря которой она не столь быстро привыкает к генераторному потенциалу. Тем не менее почти во всех случаях, когда рецепторную клетку подвергают непрерывному раздражению, происходит некоторое привыкание или адаптация. Во всяком случае, инициация нервных импульсов в аксонах рецепторных клеток при помощи генераторного потенциала представляет собой, по-видимому, широко распространенное явление.
Остается решить, каким образом внешнее раздражение вызывает генераторный потенциал? Изучение большей части рецепторов не дает никаких сведений по этому вопросу. Только на примере фоторецептора можно получить точные сведения относительно первого шага в процессе возбуждения рецепторных клеток. Между тем исследование фоторецептора связано с большими трудностями. У большинства животных рецепторы глаза очень мелкие и плотно упакованы, а связанные с ними нервные структуры очень сложны и высокоорганизованы. Счастливое исключение составляет сложный глаз мечехвоста, на котором были получены первые данные о генераторном потенциале. В этом глазу рецепторные клетки очень крупные и их нейральная организация относительно проста.
Сложный глаз мечехвоста, имеющий резко выраженную фасеточность, состоит примерно из тысячи омматидиев («маленьких глазков»), каждый из которых образован всего каким-нибудь десятком клеток. В каждом омматидии клетки расположены в определенном порядке. Клетки сетчатки (рецепторы) располагаются радиально, подобно долькам мандарина, вокруг дендрита их нейрона — единственной эксцентрически расположенной клетки, имеющейся в каждом омматидии.
Хартлайн, X. Вагнер и Э. Мак-Николь (младший) установили при помощи микроэлектродов, что эта эксцентрическая клетка генерирует нервные импульсы, которые можно зарегистрировать в дистальных участках нервного волокна, выходящего из омматидия. Микроэлектрод позволяет также зарегистрировать генераторный потенциал омматидия. Ввиду сложного анатомического строения омматидия до сих пор не удалось точно установить место возникновения генераторного потенциала. Не была также обнаружена активность и в аксонах клеток сетчатки. Как и в случае рецепторов, воспринимающих растяжение у позвоночных и беспозвоночных, местная анестезия подавляет нервные импульсы, не нарушая генераторный потенциал. Более того, как и в случае упомянутых рецепторов, наблюдается пропорциональность между степенью деполяризации и частотой нервных импульсов.
Недавно М. Фуортес показал, что под действием света проводимость эксцентрической клетки возрастает. По его мнению, это возрастание связано с каким-то химическим посредником, который освобождается под влиянием света и воздействует на дендрит эксцентрической клетки. По-видимому, возросшая проводимость дендрита связана с деполяризацией, которая пассивно распространяется к месту возникновения импульсов, где она играет роль генераторного потенциала.
Светочувствительные клетки — единственные среди всех рецепторов — позволяют получить прямые экспериментальные сведения о первоначальных молекулярных явлениях, связанных с процессом восприятия раздражения. Прошло почти сто лет с тех пор, как стало известно, что зрительные рецепторные клетки как позвоночных, так и беспозвоночных имеют специальные органеллы, содержащие светочувствительный пигмент. У позвоночных этот красноватый пигмент, называемый родопсином, хорошо виден в наружных сегментах палочек. Спектр поглощения родопсина человека почти полностью соответствует кривой спектральной чувствительности человеческого глаза при сумеречном освещении, когда функционируют только палочки сетчатки. Это служит веским доводом в пользу того, что с родопсином связано первое активное явление в палочковом зрении: поглощение света фоторецепторной структурой. (Имеются сведения о наличии аналогичных пигментов и в наружных сегментах колбочек, но выделить и исследовать их оказалось труднее.)
Зрительные пигменты, как известно, относятся к сложным белкам; однако оказалось, что та их часть, которая поглощает свет и носит название хромофора, представляет собой более простое соединение: альдегид витамина А. В молекуле этого соединения имеется несколько двойных химических связей, и поэтому оно может существовать в различных молекулярных конфигурациях, известных под названием цис — и транс- изомеров. Работами Р. Хэббард, Дж. Уолда и их сотрудников установлено, что хромофор, представляющий собой 11-цис-альдегид витамина А, в результате поглощения света переходит в транс-форму. Эта фотохимическая реакция представляет собой первый шаг в цепи сложных и пока еще неизвестных нам химических и физических явлений, вызывающих возникновение генераторного потенциала рецепторной клетки и приводящих в конечном счете к возникновению импульсов в зрительном нерве. Это единственный случай, когда известен специфический молекулярный механизм, при помощи которого рецепторная клетка воспринимает изменения в окружающих условиях.
Дополнительные данные о том, что реакция на световое раздражение зависит от родопсина, можно получить, сравнивая спектр поглощения родопсина из глаза мечехвоста с чувствительностью глаза этого животного при различной длине волны. В 1935 г. К. Грехэм и Хартлайн измерили интенсивность вспышек света с разной длиной волны, необходимую для того, чтобы вызвать определенное число импульсов в зрительном нерве мечехвоста. При наложении кривой чувствительности, полученной в этом эксперименте, на кривую поглощения для родопсина мечехвоста, полученную Хэббард и Уолдом, оказалось, что эти две кривые почти полностью совпадают. При длине волны, равной примерно 5200 ангстремов, при которой наблюдается наиболее сильное поглощение света родопсином, глаз мечехвоста генерирует наибольшее число импульсов в единицу времени на данное количество полученной световой энергии. Оказалось, что глаз мечехвоста чувствителен к свету примерно тех же длин волн, что и глаз человека при слабом освещении, когда преобладает палочковое зрение.
Многие привычные ощущения, известные нам из повседневного опыта, — это проявления свойств отдельных сенсорных клеток. Самым простым из них представляется наша способность улавливать увеличение интенсивности какого-либо раздражения. Каждый раз, когда мы испытываем что-либо подобное, мы можем быть уверены, что в чувствительных волокнах, несущих информацию к мозгу, импульсы возникают с большей частотой, чем прежде. Нам хорошо знакомо также явление приспособления органов чувств: например, какой-нибудь резкий запах с течением времени ощущается слабее, хотя определения его интенсивности при помощи приборов показывают, что она остается на прежнем уровне.
Те, кто занимается фотографией, хорошо знают, что можно, различным образом комбинируя выдержку и диафрагму, получать всякий раз один и тот же результат; это равносильно утверждению, что интенсивность и продолжительность освещения (в известных пределах) взаимозаменяемы, когда речь идет о том, чтобы получить тот или иной фотохимический эффект. Этому же правилу подчиняется и глаз человека, подверженный действию коротких вспышек света, а также фоторецептор мечехвоста. Число нервных импульсов, возникающих при действии на омматидий слабого света в течение 0,1 секунды, примерно соответствует числу импульсов, возникающих при действии в 10 раз более яркого света в течение 0,01 секунды.
Нам известно также, что мы вовсе не замечаем колебаний интенсивности света, происходящих с очень высокой частотой, например, когда мы смотрим на экран кино или телевизора. Это явление, по-видимому, можно объяснить на основе наблюдений над генераторными потенциалами и нервными импульсами, возникающими при действии на омматидий мечехвоста света, мерцающего с разной частотой. Мерцание света можно обнаружить по колебаниям генераторного потенциала, что проявляется в виде серий импульсов. По мере возрастания частоты мерцаний частота импульсов все более приближается к той, которая наблюдается при непрерывном освещении и в конце концов становится неотличимой от нее. Как можно видеть из этих записей, «слияние мерцаний» непосредственно обусловлено характером генераторного потенциала, который (выравнивается при наивысшей частоте мерцаний.
Все описанные выше эксперименты производились на одиночных клетках или одиночных сенсорных единицах. Между тем в глазу, ухе и других органах рецепторные клетки сгруппированы в плотные образования и обычно функционируют совместно. Современные исследования показывают, что рецепторные клетки сложных органов чувств в сущности редко действуют независимо друг от друга. В таких органах между рецепторными клетками имеются нервные связи и в результате этих связей возникают новые функциональные свойства.
Хотя глаз мечехвоста имеет не столь сложное строение, как глаза позвоночных, тем не менее в нем четко проявляются результаты нервного взаимодействия. В глазу мечехвоста активность каждого фоторецептора до некоторой степени зависит от активности близлежащих омматидиев. Частота разрядов импульсов в волокне зрительного нерва, отходящего от данного омматидия, уменьшается при действии света на соседние омматидии. Поскольку каждый омматидий является соседом своих соседей, подавление активности оказывается взаимным. Это подавление осуществляется благодаря сплетению нервных волокон, синаптически связанных друг с другом и образующих войлокообразное переплетение, расположенное позади омматидиев. Возможно, что подавление происходит в результате уменьшения величины генераторного потенциала в месте возникновения нервных импульсов, — уменьшения, влекущего за собой снижение частоты этих импульсов.
При одновременном освещении двух соседних омматидиев каждый из них генерирует меньшее число импульсов, чем если бы то же самое количество света получил он один. Степень подавления, испытываемого каждым омматидием (при постоянных условиях), зависит только от частоты реакций другого омматидия. Чем больше расстояние между двумя омматидиями, тем менее выражено взаимное подавление. При одновременном освещении нескольких омматидиев подавляющее действие, испытываемое каждым из них, представляет собой сумму подавляющих влияний, исходящих от всех прочих омматидиев.
Подавляющее взаимодействие может вызвать весьма важные зрительные эффекты. Участки сетчатки, освещенные более интенсивно, действуют на менее интенсивно освещенные участки сильнее, нежели эти последние действуют на них. В результате этого различия в нервной деятельности участков сетчатки, освещенных по-разному, усугубляются. При этом возрастает контрастность и некоторые важные черты изображения на сетчатке чрезмерно подчеркиваются в ущерб верности изображения.
Это удалось показать, перемещая перед сложным глазом мечехвоста неравномерно освещенный объект, состоящий из двух прямоугольников — светлого и темного. Глаз был экранирован таким образом, что только один омматидий «следил» за объектом, расположение которого на сетчатке все время изменялось. При любом положении объекта регистрировалась постоянная частота разрядов. В результате удалось точно воспроизвести (в виде частоты импульсов) форму освещенного объекта. Затем экранировку удаляли и вновь перемещали перед глазом освещенный объект, так что на этот раз за ним могли «следить» все омматидии; однако регистрировались импульсы лишь с одного омматидия. На этот раз частота импульсов возрастала при прохождении светлого прямоугольника и уменьшалась при прохождении темного. Этого следовало ожидать, поскольку в районе светлого прямоугольника у соседних омматидиев, обращенных к более темной части объекта, частота разрядов была ниже, а, следовательно, их подавляющее действие было небольшим. Поэтому частота разрядов у рецепторов, обращенных к светлой части объекта, оказывалась выше, чем у рецепторов, освещенных в равной степени, но расположенных несколько дальше. Подобным же образом можно объяснить понижение частоты импульсов у омматидиев, обращенных к темной части объекта. Реальный результат реакции подобного характера состоит в подчеркивании контуров; этот эффект легко проверить на самом себе, если смотреть на узор, состоящий из ряда одинаковых серых полос, дающих всю гамму переходов от белого к черному. Живописцам хорошо известен эффект «пограничного контраста», и они нередко намеренно усиливают его в своих картинах. И как все мы хорошо знаем по карикатурам и другим контурным рисункам, одни очертания объекта содержат весьма значительную информацию о нем. Дж. Бекеши высказал предположение, что аналогичное подавляющее взаимодействие играет важную роль в органе слуха, повышая его способность различать высоту тона.
Имеются также сведения о том, что реакции многих органов чувств, возможно, изменяются под действием нервных влияний, идущих от центров высшей нервной деятельности. Так, чувствительность мышечного веретена позвоночных зависит от изменений длины волокон в этом веретене, а эта длина зависит как от «производительности» рецептора, так и от его взаимодействия с высшими нервными центрами. Чувствительность обонятельных рецепторов позвоночных, по всей вероятности, также может измениться под влиянием серии импульсов из высших центров. Аналогичные влияния, еще недостаточно хорошо понятые, по-видимому, испытывает сетчатка глаза.
Совершенно очевидно, что реакции сложных органов чувств определяются свойствами отдельных рецепторных клеток, влияниями, которые эти клетки оказывают друг на друга, и регулирующим действием со стороны других органов. Таким образом, активность рецепторных клеток вносит свой вклад в создание тех сложных типов нервной деятельности, которые дают возможность организмам выжить в мире, полном бесконечного разнообразия и постоянных изменений.
Автор: У. Миллер, Ф. Ратклифф и X. Хартлайн
Если вы нашли ошибку, пожалуйста, выделите фрагмент текста и нажмите Ctrl+Enter.
www.activestudy.info
Растительная клетка как живая система
Клетка - это мельчайшая единица жизни, которая характеризуется определенным типом обмена веществ, самостоятельным энергетическим циклом и способностью к саморегуляции.

Растительная клетка
Клетка - это открытая термодинамическая система, существующая при сопряженности потоков вещества, энергии и информации.
Поток информации - это отражение состояния либо отдельных клеток, либо их органоидов, выражающееся в их биологической функции.
Основой дифференциации метаболических процессов в клетке является компартментация, т.е. деление клетки на участки (компартменты), различающиеся по степени активности и составу содержащихся в них химических метаболитов и ферментов. Все компартменты в клетке разделяются биологическими мембранами.
Сами метаболиты и ферменты обладают достаточной подвижностью и переносятся в составе мелких вакуолей по ЭПС.
В растительной клетке различают три основных компартмента:
- свободное пространство (СП),
- цитоплазму,
- вакуоль.
В СП находятся углеводы и их приток и отток не контролируется клеточными мембранами.
В цитоплазме и сферосомах происходит биосинтез органических веществ и каждая из органелл, осуществляющая такой биосинтез, играет роль реакционных отсеков. Таким образом цитоплазма делится в свою очередь на множество мелких компартментов.
Вакуоль является компартментом, в котором сосредоточены запасы неорганических веществ, в том числе и воды, и простых органических веществ (органических кислот, биоз, триоз, тетроз).
Все процессы, происходящие в клетке, управляются тремя основными регулирующими системами:
- генетической, которая обеспечивает включение и выключение отдельных генов,
- гормональной, которая реализуется за счет синтеза в клетке специфических белков, либо запускается в клетке при поступлении гормона от других клеток организма,
- факторами внешней среды, которые обладают трофической регуляторной функцией и энергетической регуляторной функцией. Трофические факторы - это химические вещества, образующиеся в результате метаболизма. Они выступают и как участники обмена веществ, и как регуляторы ферментативной активности. Энергетический фактор - это синтезирующиеся в клетке макроэргические молекулы АТФ.
Клетке присущи свойства раздражимости и возбудимости. Способность живой структуры отвечать на действие раздражителя называется раздражимостью. Раздражимость является свойством любой живой клетки, в том числе и растительной. Раздражитель - это внешнее воздействие, достигшее пороговой силы. В качестве раздражителя может выступать любой вид энергии - механическая, химическая, электрическая, световая, тепловая. Раздражимость характеризуется такими свойствами, как количество раздражения и суммация раздражения.
Сила ответной реакции, эффект раздражения, определяется количеством раздражения - произведением силы раздражителя на время его действия. Слабый агент при длительном воздействии может оказать такое же действие, как и сильный агент при кратковременном влиянии на клетку.
Суммация раздражения - это накопление ряда отдельных одинаковых по природе воздействий в течение определенного времени, в результате которого достигается пороговая сила воздействия ( пример усика и опоры). Это явление обусловлено эволюционным приспособлением организмов, позволяющим избежать образование ответной реакции на слабые воздействия внешней среды.
В результате раздражения клетка способна превращать местное воздействие в возбуждение электрических сил в клетке в виде смены биоэлектрических потенциалов и передавать этот сигнал в другие клетки. Поскольку у растений, в отличие от животных, нет специальных клеток для восприятия и передачи раздражения, то свойство раздражения и возбуждения присуще всем растительным клеткам. Еще Дарвин показал наличие у растений сенсорных и моторных зон, существование которых обусловлено способностью растительных клеток передавать по тканям сигнал раздражителя.
При действии на клетку раздражителя в ней одновременно можно наблюдать несколько функциональных состояний:
- повреждение,
- возбуждение,
- закалка,
- репарация.
Повреждение выражается в нарушении структуры и функций клетки. Вначале, когда действие раздражителя незначительно, возможно обратимое повреждение протоплазмы, наблюдается торможение, которое характеризуется нечувствительностью к действию раздражителя. Если же действие раздражителя продолжается, то торможение переходит в повреждение и заканчивается гибелью клетки.
Возбуждение возникает у растений лишь при определенной длительности и частоте воздействия раздражителя, определяющей силу возникающего биоэлектрического потенциала. Взаимосвязь клеток происходит через плазмодесмы, по которым идет не только передвижение веществ, но и передача стимулирующих эффектов.
Закалка возникает в результате активации клеточной деятельности при возбуждении, когда действие раздражителя воспринимается в меньшей степени.
Репарация наблюдается на фоне закалки и заключается в восстановлении исходных структур и функций клетки. При этом клетка адаптируется к уровню внешнего фактора.
При усилении действия раздражителя процесс повторяется, но на другом уровне силы воздействия. Таким образом реакция живой растительной клетки, а, следовательно, и всего растения, на условия внешней среды носит колебательный характер.
Уровень биоэлектрического потенциала каждой стороны мембраны определяется индивидуальными особенностями организма или ткани, однако разность этих потенциалов по обеим сторонам мембраны составляет 0.5-1 вольт. При передаче возбуждения происходит смена заряженности биопотенциалов мембран и передача электрического импульса от мембраны к мембране.
Очень важным качеством живой клетки является ее проницаемость. Проницаемость цитоплазмы зависит от:
- характера веществ, которые содержатся в клетке,
- соотношения различных минеральных ионов (например, проницаемость клетки можно регулировать с помощью добавления одно - или двухвалентных катионов),
- температуры и других факторов внешней среды,
- характера веществ, поступающих в клетку (например, чем больше гидроксильных групп в поступающем веществе, тем хуже оно поступает в цитоплазму). Такое же свойство оказывают карбоксильные, аминные группы, высокий молекулярный вес, сложное пространственное строение поступающей в клетку молекулы. Диссоциированные молекулы хуже проникают в цитоплазму, чем нейтральные молекулы.
biofile.ru
ФИЗИОЛОГИЯ РАСТИТЕЛЬНОЙ КЛЕТКИ
Количество просмотров публикации ФИЗИОЛОГИЯ РАСТИТЕЛЬНОЙ КЛЕТКИ - 210
Терминологический словарь
Сборы и пошлины с участников сделок с недвижимостью
Государственная пошлина.
При нотариальном удостоверении договоров отчуждения недвижимости с их участников взимается государственная пошлина.
Под государственной пошлиной принято понимать установленный обязательный и действующий на всей территории Российской Федерации платеж, взимаемый за совершение юридически значимых действий либо выдачу документов уполномоченными на то органами или должностными лицами.
Государственная пошлина взимается в следующих размерах#S:#G1
· за удостоверение договоров, предметом которых является отчуждение недвижимого имущества (земельных участков, жилых домов, квартир, дач, сооружений и иного недвижимого имущества):
- детям, в т.ч. усыновленным, супругу, родителям, родным братьям и сестрам - 0,5 процента от суммы договора, но не менее 50 процентов от минимального размера оплаты труда;
- другим лицам - 1,5 процента от суммы договора, но не менее однократного размера минимального размера оплаты труда.
Регистрационные сборы.
При государственной регистрации сделок с недвижимым имуществом взимаются регистрационные сборы.
Контрольные вопросы:
Каким нормативным документом определяются категории земель выделяются по целевому назначению в Российской Федерации?
Что принято понимать под термином ʼʼустойчивое развитие территорийʼʼ?
Какие основные документы входят в систему регулирования градостроительной деятельности на уровне поселений?
В чем отличие Генерального плана от Правил застройки и землепользования?
Что такое "разрешенное использование территорий"?
Какая информация содержится в градостроительном плане земельного участка?
В чем преимущества налогообложения недвижимости как источника доходов бюджета?
Какие условия необходимы для создания эффективной системы налогообложения недвижимости?
Какие особенности существуют в налогообложении недвижимости в разных странах?
В чем главные особенности системы налогообложения недвижимости в России?
Арендная плата –плата за пользование (владение) имуществом, в том числе, недвижимым.
Государственный кадастр недвижимости (ГКН) –систематизированный свод сведений .. о недвижимом имуществе, а также о Государственной границе РФ, границах между субъектами Федерации, границах муниципальных образований и населенных пунктов.
Государственная регистрация прав на недвижимое имущество и сделок с ним -юридический акт признания и подтверждения государством возникновения, ограничения (обременения), перехода или прекращения прав на недвижимое имущество.
Девелопмент (развитие) недвижимости - качественное материальное преобразование объектов недвижимости, обеспечивающее возрастание их стоимости и ведущее одновременно, как правило, к увеличению совокупного фонда недвижимости.
Действительный (эффективный) валовой доход от объекта недвижимости -потенциальный валовой доход, скорректированный с учетом незанятости помещений, потерь от недобросовестных арендаторов, льгот по арендной плате, прочих доходов от объекта недвижимости.
Жилая недвижимость – здания и помещения в зданиях, предназначенные для постоянного проживания.
Земельный участок -часть поверхности земли, границы которой описаны и удостоверены в установленном порядке …а также все, что находится над и под поверхностью земельного участка.
Инвестиционная недвижимость - недвижимость, используемая в интересах получения дохода в виде арендной платы и прироста стоимости.
Индустриальная недвижимость – здания, сооружения, выполняющие роль материальных условий производства, но не являющиеся активной частью средств производства, непосредственно приносящими доход.
Коммерческая недвижимость – здания и помещения, создающие доход, являющиеся активной частью средств производства (в рамках рынка коммерческой недвижимости принято выделять торговую, офисную, складскую и гостиничную недвижимость).
Контрактная арендная плата (контрактная рента) –уровень арендной платы, зафиксированный в договоре аренды.
Наилучшее и наиболее эффективное использование земли - ϶ᴛᴏ такое использование, выбранное среди разумных, возможных и законных альтернативных вариантов, ĸᴏᴛᴏᴩᴏᴇ является физически возможным, достаточно обоснованным и финансово осуществимым, и ĸᴏᴛᴏᴩᴏᴇ приводит к наивысшей стоимости земли
Недвижимое имущество –родовое понятие, отражающее класс вещей, общим признаком которых является прочная связь с землей, так что в качестве недвижимого имущества могут выступать вещи, либо состоящие из земли, либо включающие в себя землю как базовый элемент.
Норма вакансий –доля незанятых помещений в имеющемся фонде недвижимости
Объект недвижимого имущества -количественно определенный участок и принадлежащие к нему здания, сооружения, многолетние насаждения и пр.
Операционная недвижимость- недвижимость, занимаемая собственником и используемая им для ведения определенной деятельности – производственной, административной, учебной и пр
Операционные расходы- расходы на обеспечение нормального функционирования объекта в соответствии с его предназначением и, соответственно, обеспечение воспроизводства действительного валового дохода.
Потенциальный валовой доход –потенциально возможный доход от объекта в виде рентных поступлений, который может принести объект недвижимости при стопроцентной занятости в течение всего периода владения.
Первичный рынок недвижимости -совокупность сделок, совершаемых с вновь созданными объектами недвижимости, вторичный – сделки, совершаемые с уже созданными объектами, находящимися и находившимися в эксплуатации.
Принадлежности -движимые по природе вещи, прикрепленные к недвижимому имуществу (земельному участку, зданию) и являющиеся его неотъемлемой (технологически или по закону, договору, описанию) частью
Принципы оценки недвижимости –методические правила, основывающиеся на положениях экономической теории и позволяющие учесть, как все многообразие факторов, присущих рынку недвижимости, влияет на стоимость недвижимости.
Проект девелопмента- сознательно реализуемая система последовательных взаимосвязанных мероприятий с использованием ограниченного объёма ресурсов в течение заданного периода времени, направленная на создание качественно новых объектов (комплексов объектов) недвижимости в заданные сроки в целях последующего возврата вложенных средств и получения дохода.
Рента- доход, который присваивается отдельными экономическими субъектами благодаря их исключительному экономическому положению.
Риск- возможность отклонения реальных результатов какой-либо операции или проекта в отрицательную сторону по сравнению с планируемыми; событие, способное нанести ущерб, имеющий денежное выражение.
Рынок недвижимости –совокупность рыночных механизмов, посредством которых передаются права на недвижимость и связанные с ней интересы, устанавливаются цены и недвижимость распределяется между различными конкурирующими вариантами ее использования в процессе создания, обращения и использования объектов недвижимости
Рыночная арендная плата (рыночная рента) –уровень арендной платы, складывающийся в каждый данный момент времени на рынке.
Рыночная стоимость –наиболее вероятная цена, которую должна достигать недвижимая собственность на конкурентном и открытом рынке с соблюдением всех условий справедливой сделки между продавцом и покупателем при условии, что каждый из них действует ответственно и осознанно и на цену сделки не влияют не относящиеся к ней факторы.
Структурирование рынка - выделение относительно обособленных частей рынка недвижимости, для которых характерны определенные особенности функционирования, отличающие эту часть рынка от иных, смежных его частей.
Тип недвижимости –функциональное назначение объекта недвижимости, предъявляющее определенные требования к местоположению и иным характеристикам объекта
Трансакционные издержки –издержки, которые несут участники сделок на рынке в связи с совершением этих сделок (трансакций).
Улучшения земли -любые произведенные человеком изменения земли, в качестве которых могут выступать как усовершенствования самой земли в результате произведенных на ней работ (мелиорация, дренаж, комплексная инженерная подготовка к застройке), так и построенные на земле здания и сооружения.
Чистый операционный доход –действительный валовой доход за минусом операционных расходов и отчислений на замещение элементов здания и оборудования, срок службы которых менее срока экономической жизни здания.
Все клетки обладают такими свойствами живого как способность к самовоспроизведению, метаболизм (обмен веществ), раздражимость, рост, изменчивость и адаптация к внешней среде. При этом нужно учитывать, что каждая клетка многоклеточного организма находится в тесном взаимодействии с другими клетками и что организм - это единое целое, а не сумма клеток.
Клетка обладает сложной структурной организацией и представляет собой систему, дифференцированную на отдельные органеллы. Растительная клетка имеет клеточную стенку и протопласт. Протопласт состоит из ядра с ядрышком, цитоплазмы и включенных в нее мембранных (вакуоль, пластиды, митохондрии, аппарат Гольджи, лизосомы, эндоплазматический ретикулум) и немембранных (микротрубочки, рибосомы) органелл. Все органеллы погружены в матрикс цитоплазмы - гиалоплазму или основную плазму (рис. 2.1).
Рис. 2.1. Схема строения клетки мезофилла листа (по В. В. Полевому).
Все окруженные полупроницаемой мембраной компоненты клетки представляют из себязамкнутые образования, в которых происходят разнообразные биохимические реакции. Цитоплазматический матрикс также разделен на отсеки эндоплазматической сетью. Тем самым достигается дополнительное пространственное разделение (компартментализация).
Основные физиологические функции структурных компонентов клетки: клеточная стенка - обеспечение прочности, защита͵ опорная функция; плазмалемма - транспорт веществ из и внутрь клетки, защита͵ рецепция; ядро - хранение и передача генетической информации; ядрышко - синтез РНК; вакуоль - осморегуляция, запасание веществ, переваривание; основная плазма - гликолиз; митохондрии - дыхание; хлоропласты – фотосинтез; аппарат Гольджи - секреция, образование компонентов клеточной стенки; эндоплазматическая ретикулум - транспорт веществ в клетке; лизосомы - внутриклеточное пищеварение; сферосомы - накопление и хранение жира; микротела (пероксисомы, глиоксисомы) - фотодыхание, глиоксалатный цикл; микротрубочки - ориентация микрофибрилл целлюлозы; рибосомы - синтез белка.
2.1. Раздражимость
Это способность клетки реагировать на действие внешних и внутренних факторов – раздражителей и передавать возбуждение в другие клетки. Восприятие раздражения обозначают термином перцепция или рецепция. У растений нет органов чувств, но есть рецепторные белки и клетки, воспринимающие разные воздействия. Различают фото-, хемо- и механорецепторы.
Возбуждение представляет собой измененное состояние клетки. В отсутствие раздражения растительная клетка имеет отрицательный потенциал покоя от -50 до -200 мВ, то есть протоплазма заряжена отрицательно по отношению к наружной поверхности. Это связано с тем. что внутри клетки находится больше ионов хлора и калия, но меньше ионов кальция, чем снаружи.
В ответ на раздражение возникает потенциал противоположного знака - потенциал действия, который может распространяться и на время сравняться или превысить потенциал покоя. Потенциал действия образуется в результате выхода ионов хлора из клетки и поступления ионов кальция в клетку.
Раздражение воспринимается клеткой, в случае если его сила превышает пороговую величину. Следующие друг за другом подпороговые раздражения могут суммироваться, в случае если интервалы между ними незначительны. Минимальное время, крайне важно е для рецепции, называют временем презентации.
Установлены следующие законы раздражимости:
1. Закон силы раздражения: чем больше раздражение, тем сильнее ответная реакция.
2. Закон длительности раздражения: чем длительнее раздражение, тем сильнее ответная реакция.
3. Закон количества раздражения: чем больше сила раздражения, тем меньше время презентации.
4. Закон градиента раздражения: чем выше скорость нарастания силы раздражителя, тем больше ответная реакция.
При действии чрезмерно сильных и (или) длительных воздействий чувствительность клеток к раздражителю снижается и клетки могут погибнуть.
Реституция - это восстановление исходного состояния после раздражения. Она идет с затратой энергии и тормозится под действием наркотиков, ингибиторов дыхания, при недостатке кислорода и снижении температуры. Во время реституции клетки находятся в рефрактерном периоде, когда новое раздражение не вызывает возбуждения.
2.2. Репликация, транскрипция и трансляция
Хранение наследственной информации осуществляется дезоксирибонуклеиновой кислотой (ДНК). ДНК - это полимер, мономерами которого являются дезоксирибонуклеотиды. В их состав входят углевод 2-дезокси-b-D-рибоза, остаток фосфорной кислоты и азотистые основания четырех типов: два пуриновых - аденин и гуанин, и два пиримидиновых - тимин и цитозин. Молекула ДНК состоит из двух полинуклеотидных цепочек, скрепленных между собой водородными связями между азотистыми основаниями. Полинуклеотидная цепочка образована чередующимися остатками 2-дезокси-b-D-рибозы и фосфорной кислоты, которая присоединена к 5 атому углерода кольца углевода с одной стороны и к 3 атому с другой стороны. Азотистые основания ответвляются от цепочки, присоединяясь к 1 атому углерода кольца углевода. Азотистые основания, противостоящие друг другу в полинуклеотидных цепочках, комплементарны друг другу: пуриновому основанию соответствует пиримидиновое (аденину – тимин, гуанину – цитозин). У каждой цепочки молекулы ДНК один конец заканчивается пятым (фосфатным), а другой - третьим (гидроксильным) углеродным атомом углевода. Οʜᴎ обозначаются как 5’ и 3’- концы. Цепочки в молекуле ДНК антипараллельны. В одной цепочке нуклеотиды связаны в направлении 5’ ® 3’, а в другой - 3’ ® 5’. Полинуклеотидные цепочки образуют двойную спираль. Каждый виток спирали содержит 10 пар азотистых оснований (рис. 2.2).
Рис. 2.2. Схема строения молекулы ДНК в виде двойной спирали (а) и прямой лесенки (б) (по В. Олфри и А. Мирскому).
1 – остаток 2-дезокси-β-D-рибозы, 2, 3, 4, 5 – азотистые основания (тимин, аденин, гуанин, цитозин, соответственно), 6 – остатки фосфорной кислоты.
Во время репликации (синтеза) ДНК каждая из цепей родительской ДНК служит матрицей для образования комплементарной дочерней цепи из предшественников – дезоксирибонуклеозидтрифосфатов: дезоксиаденозин-, дезоксицитидин-, дезоксигуанозин- и дезокситимидинтрифосфата. При их полимеризации происходит освобождение молекул пирофосфата͵ которые расщепляются пирофосфатазой. Репликация ДНК осуществляется по полуконсервативному механизму: одна из цепей дочерней молекулы ДНК является частью родительской молекулы, а другая – вновь синтезированной.
ДНК-полимераза способна синтезировать ДНК только в направлении от 5¢-конца к 3¢-концу. По этой причине на одной цепи ДНК образование дочерней цепи, называемой ведущей, происходит непрерывно, синтез другой цепи происходит прерывисто в виде коротких фрагментов Оказаки, получивших свое название в честь ученого, впервые их обнаружившего. Эта вторая дочерняя цепь получила название отстающей. Фрагменты Оказаки также синтезируются в направлении 5¢ ® 3¢, но перемещение ДНК-полимеразы вдоль матричной цепи ДНК при образовании каждого фрагмента противоположно направлению движения при синтезе ведущей цепи. Ведущая и отстающая цепи ДНК синтезируются координировано, что обеспечивается димеризацией ДНК-полимеразных комплексов у бактерий и наличием разных ДНК-полимераз у эукариотов. Важно заметить, что для соединения двух фрагментов Оказаки ДНК-лигазой сначала удаляется РНК-затравка с помощью экзонуклеазы и РНКазы Н – нуклеазы, специфически расщепляющей РНК в ДНК-РНК-гибридах.
Процесс репликации ДНК разделяют на три этапа: инициацию, элонгацию и терминацию. Стоит сказать, что для начала репликации необходим праймер (затравка) – короткий олигодезокси- или олигорибонуклеотид, комплементарный соответствующему участку ДНК-матрицы.
В ходе репликации ДНК ее цепи расходятся из точки репликации, образуя Y-подобную структуру, которая принято называть репликативной вилкой. Именно в данном месте локализован репликативный комплекс, состоящий из нескольких белков. ДНК-хеликаза перемещается в репликативной вилке впереди ДНК-синтезирующего комплекса, расплетает цепи родительской ДНК и стимулирует образование затравок праймазой. ДНК-синтезирующий комплекс содержит ДНК-полимеразу, подвижный связывающий белок RPA и ДНК-зависимую АТФазу, которая узнает и связывается с единственным праймером ведущей цепи ДНК и с каждым праймером фрагментов Оказаки отстающей цепи, что делает возможным присоединение к праймерам ДНК-полимеразы и белка RPA. RPA связывается с образующимися одноцепочечными участками, облегчая процесс расплетения. RPA охватывает молекулу ДНК позади ДНК-зависимой АТФазы, стимулирует АТФазную активность и оставляет 3¢-конец праймера доступным для ДНК-полимеразы. ДНК-полимераза начинает элонгацию цепей ДНК, присоединяя первый дезоксирибонуклеозидмонофосфат к 3¢-концевому нуклеотиду РНК-затравки.
Передача информации от ДНК осуществляется посредством информационной или матричной рибонуклеиновой кислоты (мРНК). Синтез мРНК принято называть транскрипцией. Молекула мРНК, комплементарная одной из цепей матричной ДНК, образуется в ходе сополимеризации четырех рибонуклеозидтрифосфатов (аденин-, гуанозин-, цитозин- и урацилтрифосфата) с образованием 3¢-5¢-фосфодиэфирных связей и освобождением неорганического пирофосфата. Транскрипцию осуществляет фермент ДНК-зависимая РНК-полимераза. Синтез мРНК молекулами РНК-полимеразы начинается в определенных местах ДНК, называемых промоторами, и завершается на особых нуклеотидных последовательностях – терминаторах. Совокупность нуклеотидов ДНК, заключенных между промотором и терминатором, называют транскрипционной единицей или транскриптоном.
Процесс транскрипции подразделяют на 4 стадии: связывание РНК-полимеразы с ДНК и распознавание промотора, инициация, элонгация и терминация. Предполагается, что после первоначального непрочного связывания с ДНК в случайном месте молекула РНК-полимеразы перемещается вдоль двойной спирали ДНК до тех пор, пока не обнаружит последовательность нуклеотидов промотора. В этом месте связывание молекулы фермента с ДНК становится более прочным. Инициация транскрипции начинается с образования на промоторе предъиниционного комплекса, состоящего из РНК-полимеразы и матричной ДНК. После сборки предъинициационный комплекс претерпевает температурно-зависимые конформационные изменения, которые сопровождаются локальным плавлением, то есть расплетением двойной спирали ДНК, и комплекс становится способным к транскрипции. При наличии рибонуклеозидтрифосфатов происходит образование первых фосфодиэфирных связей в молекуле синтезируемой мРНК, после чего начинается стадия элонгации, то есть последовательное удлинение синтезируемой молекулы мРНК.
В 1992 ᴦ. М. Чэмберлен с сотрудниками разработали общую модель элонгации мРНК, согласно которой перемещение РНК-полимеразы вдоль ДНК и присоединение нуклеотидов к растущей цепи мРНК в активном центре фермента разделены во времени. Это разделение возможно потому, что у РНК-полимеразы имеется два сайта (участка), удерживающих растущую цепь мРНК, и два участка связывания ДНК-матрицы. Молекула РНК-полимеразы перемещается вдоль ДНК подобно гусенице: когда один сайт связывания ДНК фиксирован, другой перемещается вперед. ДНК-зависимые РНК-полимеразы фагов, состоящие из одной субъединицы, синтезируют РНК в условиях in vitro со скоростью 200-400 нуклеотидов в секунду. При перемещении фермента вдоль матрицы цепи ДНК подвергаются плавлению и повторному отжигу, в результате которого восстанавливается исходная структура ДНК. Стадия элонгации заканчивается после достижения РНК-полимеразой терминатора транскрипции. Далее синтезированная РНК и РНК-полимераза освобождаются из транскрипционного комплекса. Только минус-цепь ДНК служит матрицей для синтеза мРНК.
Участки ДНК, несущие информацию о строении белка - экзоны, разделены неинформативными интронами. В процессе транскрипции считывается информация как с экзонов, так и с интронов. Образуется предшественник мРНК - про-мРНК. Молекулы про-мРНК претерпевают созревание - процессинᴦ. В ядре из про-мРНК происходит вырезание интронов и объединение экзонов - сплайсинᴦ. К образовавшейся мРНК прикрепляется особая группировка поли-А и к одному из концов мРНК присоединяется защитная химическая группировка КЭП. После этого мРНК соединяется с белком, образуя инфорсому. Она выходит через поры в ядерной оболочке в цитоплазму. мРНК высвобождается из инфорсомы и одноцепочечная неспирализованная молекула мРНК присоединяется к участку малой субъединицы рибосомы, который примыкает к большой субъединице. К рибосоме прикрепляется небольшой участок цепи мРНК, содержащий один кодон, состоящий из трех азотистых оснований. Один кодон соответствует одной аминокислоте. При этом некоторые аминокислоты кодируются несколькими разными кодонами. Τᴀᴋᴎᴍ ᴏϬᴩᴀᴈᴏᴍ, первый этап синтеза белка - трансляции состоит в образовании комплекса между мРНК и рибосомой.
Перенос аминокислот в цитоплазме к рибосомам осуществляется транспортными РНК ( тРНК или 4S-РНК) из 70-80 нуклеотидов с молекулярной массой 25-30 кД. Οʜᴎ составляют почти 10 % от всей клеточной РНК и растворены в гиалоплазме. По этой причине тРНК еще называют растворимой РНК. В состав белков входят 20 аминокислот и каждой аминокислоте соответствует своя тРНК. Благодаря определенному расположению комплементарных нуклеотидов полинуклеотидная цепочка тРНК образует вторичную структуру, получившей название “клеверного листа”. Рентгеноструктурный анализ позволил установить третичную структуру тРНК. Она оказалась составленной из двух стеблей наподобие латинской буквы L.
Каждая тРНК имеет триплет оснований - антикодон, ответственный за прикрепление к комплементарному кодону мРНК. Для того, чтобы аминокислота присоединилась к тРНК необходима ее активация или обогащение энергией. Активация аминокислоты происходит за счёт ее реакции с аденозинтрифосфорной кислотой (АТФ) - вещества с макроэргическими связями (~). Реакция идет при участии фермента аминоацилсинтетазы (кодазы). Образовавшийся аминоациладенилат остается связанным с ферментом и вступает в реакцию с тРНК с образованием аминоацил-тРНК.
Аминоацил-тРНК антикодоном присоединяется к кодону мРНК на малой субъединице рибосомы. После того как первая аминокислота со своей тРНК вошла в малую субъединицу рибосомы, происходит смыкание малой и большой субъединиц рибосомы. После смыкания субъединиц тРНК вместе с аминокислотой переносится на большую субъединицу. Одновременно мРНК перемещается на один кодон. В результате в малую субъединицу входит следующий кодон, кодирующий другую аминокислоту. К этому кодону с помощью антикодона присоединяется вторая тРНК со своей аминокислотой. В рибосоме оказываются две аминокислоты, ориентированные друг около друга таким образом, что карбоксильная группа первой аминокислоты оказывается рядом с аминогруппой второй аминокислоты. В результате сближения этих групп и с участием фермента большой субъединицы рибосомы пептидилтрансферазы аминокислоты соединяются пептидной связью. Образовавшийся дипептид присоединен ко второй тРНК, а первая тРНК освобождается и уходит в цитоплазму. В результате дипептид со второй тРНК оказывается связанным с большой субъединицей рибосомы, а мРНК перемещается еще на один кодон (рис. 2.3). Присоединение аминокислотных остатков - элонгация повторяется многократно, пока не образуется полипептидная цепочка белка. Окончание образования полипептидной цепочки - терминация связано с тем, что в малую субъединицу вступает терминальный кодон мРНК. Образовавшаяся полипептидная цепочка покидает рибосому. Большое значение имеет объединение рибосом в цепочки - полисомы. В этом случае одна молекула мРНК может последовательно присоединяться к ним и служить матрицей для синтеза нескольких одинаковых молекул белка. Когда синтез белка закончен, мРНК распадается. Синтез белковой молекулы идет с большой скоростью и в связи с этим время жизни мРНК невелико (от нескольких секунд до 1-2 минут). При этом, на определенных фазах развития растений (к примеру, в набухающих и прорастающих семенах) синтезируются так называемые долгоживущие молекулы мРНК.
Рис. 2.3. Схема синтеза полипептидной цепи в рибосоме (по В. В. Полевому).
В заключение можно сказать, что из поколения в поколение передаются молекулы ДНК, которые несут в себе информацию о составе белковых молекул. План построения белка записан в ДНК с помощью триплетного кода, представленного чередованием азотистых оснований. Под влиянием внешних условий или спонтанно ДНК может изменяться. Эти изменения бывают полезными, бесполезными и вредными. Полезные изменения, дающие организмам преимущество в борьбе за существование, могут закрепляться по наследству в ходе естественного или искусственного отбора.
Возникающие в процессе трансляции белки являются полимерами, мономерами которых служат аминокислоты. Белки представляют из себяцепочки остатков аминокислот, соединенных между собой пептидными связями. Молекулярная масса белков зависит от количества мономеров и колеблется от нескольких тысяч до миллионов. Разнообразие белков определяется различной последовательностью аминокислотных остатков. Белки, также как и аминокислоты - амфотерные соединения, то есть имеют положительный и отрицательные заряды. У каждого белка своя изоэлектрическая точка - значение рН, при которой молекула белка не имеет заряда.
Молекула белка имеет первичную, вторичную, третичную и четвертичную структуру. Первичная структура определяется последовательностью аминокислотных остатков в белковой молекуле. Между аминокислотами, входящими в полипептидную цепочку, возможны водородные и другие связи. В результате этого полипептидная цепочка приобретает особенное расположение в пространстве, чаще всего в виде спирали. Это вторичная структура. По конформации полипептидной цепи различают фибриллярные и глобулярные белки. Фибриллярные белки сохраняют вытянутую форму спирали. У глобулярных белков спираль сворачивается в шарообразную глобулу, которая представляет собой третичную структуру белка. Она поддерживается кроме водородных связей гидрофобными взаимодействиями и дисульфидными связями, возникающими между двумя сульфгидрильными (SH) группами. Белки могут состоять из нескольких полипептидных цепочек и их взаимное расположение в пространстве и вокруг друг друга представляет собой четвертичную структуру.
2.3. Регуляции ферментативной активности в клетке
Химические процессы в клетке протекают с большой скоростью благодаря действию биологических катализаторов - ферментов или энзимов. Белковую часть фермента называют апоферментом. Небелковый компонент, прочно связанный, называют простетической группой, слабо связанный, обслуживающий несколько ферментов - коферментом. В состав простетических групп и коферментов входят металлы (железо, медь, цинк), витамины и их производные. Эндоферменты функционируют в клетке, а экзоферменты выделяются из клетки или локализуются в плазмалемме и действуют снаружи от нее.
Ферменты обладают специфичностью как к субстратам (субстратная специфичность), так и к определенным химическим реакциям (специфичность действия). Названия ферментов обычно заканчиваются суффиксом “аза”, за исключением некоторых общепринятых названий, к примеру, пепсин, трипсин и другие. Название, как правило, отражает природу субстрата (хитиназа расщепляет хитин), или функцию фермента (аминотрансфераза переносит аминогруппы), или же то и другое вместе (алкогольдегидрогеназа). Ферменты разделяют на 6 классов: 1) оксиредуктазы - катализируют окислительно-восстановительные реакции, 2) трансферазы - перенос целых атомных группировок от одного соединения к другому, 3) гидролазы - распад органических соединений с участием воды, 4) лиазы - присоединение какой-либо атомной группировки к органическим соединениям или отщепление от субстратов определенной группы без участия воды, 5) изомеразы - превращение одних изомеров в другие, 6) лигазы или синтетазы - синтез органических соединений, происходящий при участии АТФ с использованием энергии этой кислоты.
Ферменты, катализирующие одну и ту же реакцию и встречающиеся у одного организма, но различающиеся по своим физико-химическим свойствам (к примеру, по электрофоретической подвижности, следовательно, по молекулярной массе и заряду) называют изоферментами или изозимами. Наличие изоферментов позволяет организмам лучше приспосабливаться к меняющимся условиям внешней среды.
Катализ осуществляется в результате образования фермент-субстратного комплекса, что приводит к сближению реагирующих молекул или созданию напряженных химических связей путем их растягивания. Субстрат должен соответствовать активному центру не только пространственно, но и по распределению зарядов, расположению групп атомов и так далее. Окончательная подгонка происходит лишь в процессе взаимодействия субстрата с ферментом, претерпевающим при этой реакции конформационные изменения. Продукты реакции отделяются от фермента и молекулы фермента регенерируются. Благодаря своей способности регенерироваться, то есть возвращаться к первоначальному состоянию, одна и та же молекула фермента может катализировать большой объём превращений.
Скорость и направленность ферментативных реакций в клетке зависит от количества фермента͵ температуры и рН. Как всякая химическая реакция, ферментативные реакции зависят от температуры, что принято оценивать величиной температурного коэффициента (Q10), который показывает во сколько раз данный процесс ускоряется при повышении температуры на 10 Со. Поскольку ферменты являются белками, то повышение температуры свыше 35-40 Со вызывает их частичную инактивацию, а дальнейшее повышение температуры приводит уже к необратимой денатурации. У каждого фермента имеется свой оптимум рН, при котором лучше всего проявляется его активность. Это связано с тем, что рН влияет на заряд функциональных групп фермента͵ составляющих его активный центр, а от заряда зависит возможность образования фермент-субстратного комплекса.
Известны следующие механизмы внутриклеточной регуляции функционирования ферментов:
1. Метаболитная регуляция. Она происходит в результате изменения концентрации метаболитов и не затрагивает активность или число ферментных молекул. Различают регуляцию в местах разветвления путей обмена веществ и регуляцию по принципу обратной связи. В первом случае ферменты конкурируют за один и тот же субстрат и выбор пути определяется концентрацией общего для конкурирующих ферментов субстрата и степенью сродства фермента к субстрату. Под обратной связью принято понимать влияние более позднего члена цепи взаимосвязанных реакций на более ранний.
2. Ферментная регуляция. При этом типе регуляции изменяется активность ферментов. Изменение ферментативной активности может осуществляться несколькими путями: а) Обратимое или необратимое превращение неактивных предшественников ферментов - зимогенов в активные ферменты. К примеру, b-амилаза инактивирована в запасающих клетках эндосперма семян злаков из-за соединения с запасными белками посредством дисульфидных связей ( -S-S-). К началу прорастания семян из живых клеток алейронового слоя в эндосперм поступают вещества, разрушающие дисульфидные связи. Активированная b-амилаза принимает участие в гидролизе запасного крахмала; б) Изменение активности фермента под влиянием эффекторов. Связываясь с ферментом, эффекторы могут повышать его активность - это положительные эффекторы - активаторы или уменьшать ее - это отрицательные эффекторы - ингибиторы. Эффектор может влиять на активность фермента͵ взаимодействуя с активным центром (изостерический эффект) или изменяя конформацию ферментной молекулы в результате связывания с ее аллостерическим центром (аллостерический эффект). Изостерический эффект происходит в том случае, когда эффектор и субстрат похожи по своему строению и конкурируют друг с другом за активный центр фермента. Такой тип ингибирования называют конкурентным ингибированием.
3. Генная регуляция. В этом случае изменяется количество ферментных молекул в клетке из-за включения или выключения синтеза ферментов. Регулирующие факторы действуют на ДНК, РНК или рибосомы.
4. Мембранная регуляция. Различают контактную и дистанционную мембранную регуляцию активности ферментов. Контактная регуляция – связывание ферментов с мембранами или их освобождение меняет их активность. Дистанционная мембранная регуляция активности ферментов осуществляется косвенным путем в результате транспорта через мембраны субстратов и коферментов, удаления продуктов реакции, ионных и рН сдвигов в компартментах клетки.
2.4. Поступление воды в клетку
При температуре выше абсолютного нуля все молекулы находятся в постоянном движении. Диффузия - это процесс, ведущий к равномерному распределению молекул газов или растворенного вещества и растворителя благодаря их постоянному движению. Диффузия всегда направлена от большей концентрации вещества к меньшей. Количество вещества J, диффундирующего в единицу времени через воображаемое поперечное сечение, зависит от величины градиента dc/dx(градиент – мера изменения какого-либо параметра с расстоянием или временем) и от природы диффундирующего вещества, влияющего на коэффициент диффузии D. Это 1-й закон диффузии Фика, который описывается следующим уравнением:
J = - D · dc/dx,
так как суммарный поток направлен в сторону области меньшей концентрации, в уравнении стоит знак минус.
Скорость диффузии уменьшается с увеличением ее продолжительности. Проходимое путем диффузии расстояние пропорционально не времени, как при равномерном движении, а ее квадратному корню. Это 2-й закон диффузии Фика:
dc/dt = D · dc2/dx2,
где dc/dt – изменение концентрации вещества во времени.
Диффузия воды через полупроницаемую мембрану принято называть осмосом. Полупроницаемая мембрана - это мембрана хорошо проницаемая для воды и непроницаемая или плохо проницаемая для растворенных в воде веществ. Осмотическая ячейка - это пространство, окруженное полупроницаемой мембраной и заполненное каким-либо водным раствором. Все клеточные мембраны, в т.ч. плазмалемма и тонопласт, являются полупроницаемыми мембранами. Вода проходит в клетку через водные поры в плазмалемме, образованные специальными белками аквапоринами.
Внутри осмотической ячейки раствор развивает осмотическое давление p:
p = i · c · R · T,
где с – концентрация раствора в молях, Т - абсолютная температура, R - газовая постоянная 0,082 л · атм/град · моль, i – изотонический коэффициент, равный 1 + a(n-1), где a - степень элект
referatwork.ru