
Морозостойкость и зимостойкость растений. Неморозостойкие растения примеры
Морозоустойчивость растений
Морозоустойчивость- это способность переносить температуру ниже О °С, низкие отрицательные температуры. Морозоустойчивые растения способны предотвращать или уменьшать действие низких отрицательных температур. Морозы в зимний период с температурой ниже -20 °С обычны для значительной части территории России. Воздействию морозов подвергаются однолетние, двулетние и многолетние растения. Растения переносят условия зимы в различные периоды онтогенеза. У однолетних культур зимуют семена (яровые растения), раскустившиеся растения (озимые), у двулетних и многолетних - клубни, корнеплоды, луковицы, корневища, взрослые растения. Способность озимых, многолетних травянистых и древесных плодовых культур перезимовывать обусловливается их достаточно высокой морозоустойчивостью. Ткани зимостойких растений могут замерзать, однако растения не погибают. Большой вклад в изучение физиологических основ морозоустойчивости внесли Я. А. Максимов (1952), Г. А. Самыгин (1974), И. И. Туманов (1979) и другие отечественные исследователи.
Замерзание растительных клеток и тканей и происходящие при этом процессы. Способность растении переносить отрицательные температуры определяется наследственной основой данного вида растений, однако морозоустойчивость одного я того же растения зависят от условии, предшествующих наступлению морозов, влияющих на характер льдообразования. Лед может образовываться как в протопласте клетки, так и в межклеточном пространстве. Не всякое образование льда приводит клетки растения к гибели.
Постепенное снижение температуры со скоростью 0,5-1 °С/ч приводит к образованию кристаллов льда прежде всего в межклеточниках и первоначально не вызывают гибели клеток. Однако последствия этого процесса могут быть губительными для клетки. Образование льда в протопласте клетки, как правило, происходит при быстром понижении температуры. Происходит коагуляция белков протоплазмы, кристаллами образовавшегося в цитозоле льда повреждаются клеточные структуры, клетки погибают. Убитые морозом растения после оттаивания теряют тургор, из их мясистых тканей вытекает вода.
Условия и причины вымерзания растений. Образующийся при медленном промерзании в межклеточниках и клеточных стенках лед оттягивает воду из клеток; клеточный сок становится концентрированным, изменяется рН среды. Выкристаллизовавшийся лед действует как сухой воздух, иссушая клетки и сильно изменяя их осмотические свойства. Кроме того, цитоплазма подвергается сжатию кристаллами льда. Образующиеся кристаллы льда вытесняют воздух из межклеточников, поэтому замерзшие листья становятся прозрачными.
Если льда образуется немного и клетки не были механически повреждены его кристаллами, то при последующем оттаивания такие растения могут сохранить жизнеспособность. Так, в листьях капусты при температуре -5...-6 °С образуется некоторое количество льда в межклеточниках. Однако при последующем медленном оттаивании межклеточники заполняются водой, которая поглощается клетками, и листья возвращаются в нормальное состояние.
Однако клетки, ткани и растения могут погибнуть от мороза. Основными причинами гибели меток растений при низких отрицательных температурах и льдообразовании являются чрезмерное обезвоживание клеток или механическое давление, сжатие клеток кристаллами льда, повреждающее тонкие клеточные структуры. Оба эти фактора могут действовать одновременно.
Летальность действия мороза определяется несколькими обстоятельствами. Последствия воздействия низких отрицательных температур зависят от оводненности тканей растения. Насыщенные водой ткани легко повреждаются, сухие же семена могут выносить глубокие низкие температуры (до -196 °С). Низкое содержание воды предохраняет от образования льда в растениях при промораживании. Разные растения, их клетки имеют свой критический предел обезвоживания и сжатия.
Гибель клеток, тканей и растений под действием морозов обусловливается необратимыми изменениями, происходящими в протопласте клеток: его коагуляцией, денатурацией коллоидов протопласта, механическим давлением льда, повреждающим поверхностные структуры цитоплазмы, кристаллами льда, нарушающими мембраны и проникающими внутрь клетки. Вредное влияние оказывает повышение концентрации и изменение рН клеточного сока, сопровождающие обезвоживание клеток.
Действие льда, особенно при длительном влиянии низких температур, сходно с обезвоживанием клеток при засухе. Признаками повреждения клеток морозом являются потеря ими тургора, инфильтрация межклеточников водой и выход ионов из клеток. Выход ионов К и сахаров из клеток, по-видимому, связан с повреждением мембранных систем их активного транспорта. Поврежденные растения при переносе в теплое место имеют вид ошпаренных, утрачивают тургор, листья быстро буреют и засыхают. При оттаивании мороженых клубней картофеля, корнеплодов кормовой и сахарной свеклы вода легко вытекает из тканей. Важно отметить, что состояние переохлаждения (без образования льда) растения переносят без вреда; при тех же температурах, но с образованием льда в тканях растения гибнут.
Растения по-разному реагируют на образование льда в тканях: клубни картофеля и георгина быстро погибают, капуста и лук переносят лишь умеренное промораживание, рожь и пшеница выдерживают на уровне узла кущения морозы до -15...-20 °С.
У устойчивых к морозу растений имеются защитные механизмы, в основе которых лежат определенные физико-химические изменения. Морозоустойчивые растения обладают приспособлениями, уменьшающими обезвоживание клеток.
При понижении температуры у таких растений отмечаются повышение содержания сахаров и других веществ, защищающих ткани (криопротекторы), это прежде всего гидрофильные белки, моно- и олигосахариды;
снижение оводненности клеток;
увеличение количества полярных липидов и снижение насыщенности их жирнокислотных остатков,
увеличение количества защитных белков.
На степень морозоустойчивости растений большое влияние (оказывают сахара, регуляторы роста и другие вещества, образующиеся в клетках. В зимующих растениях в цитоплазме накапливаются сахара, а содержание крахмала снижается. Влияние сахаров на повышение морозоустойчивости растений многосторонне (предохраняет от замерзания большой объем внутриклеточной воды, заметно уменьшает количество образующегося льда).
Сахара защищают белковые соединения от коагуляции при вымораживании; они образуют гидрофильные связи с белками цитоплазмы, предохраняя их от возможной денатурации, повышают осмотическое давление и снижают температуру замерзания цитозоля. В результате накопления сахаров содержание прочносвязанной воды увеличивается, а свободной уменьшается. Особое значение имеет защитное влияние сахаров на белки. сосредоточенные в поверхностных мембранах клетки. Сахара увеличивают водоудерживающую способность коллоидов протоплазмы клеток; связанная с коллоидами вода в виде гидратных оболочек биополимеров при низких температурах не замерзает и не транспортируется.
Криопротекторами являются также молекулы гемицеллюлоз (ксиланы, арабиноксиланы), выделяемые цитоплазмой в клеточную стенку, обволакивающие растущие кристаллы льда, что предотвращает образование крупных кристаллов, повреждающих клетку. Так клетки защищаются как от внутриклеточного льда, так и от чрезмерного обезвоживания.
Значительное количество защитных белков и модификации молекул липидов увеличивают структурированность клеток. У большинства растений возрастает синтез водорастворимых белков. Белковые вещества, частично гидролизуясь, увеличивают содержание свободных АК.
В тканях морозоустойчивых растений в конце лета и осенью накапливаются в достаточно количестве запасные вещества (прежде всего сахара), которые используются весной при возобновлении роста, обеспечивая потребности растении в строительном материале и энергии. Необходимо также учитывать устойчивость растении к болезням, вероятность развития которых увеличивается при повреждении тканей морозом.
Закаливание растений, его фазы. Морозоустойчивость - не постоянное свойство растений. Она зависит от физиологического состояния растении и условии внешней среды. Растения, выращенные при относительно низких положительных температурах, более устойчивы, чем выращенные при относительно высоких осенних температурах. Свойство морозоустойчивости формируется в процессе онтогенеза растения под влиянием определенных условии среды в соответствии с генотипом растения и связано с резким снижением темпов роста, переходом растения в состояние покоя.
Жизненный цикл развития озимых, двуручек, двулетних и многолетних растении контролируется сезонным ритмом светового и температурного периодов. В отличие от яровых однолетних растении они начинают готовиться к перенесению неблагоприятных зимних условии с момента остановки роста и затем в течение осени во время наступления пониженных температур.
Повышение морозоустойчивости растении тесно связано с закаливанием - постепенной подготовкой к воздействию низких зимних температур. Закаливание - это обратимая физиологическая устойчивость к неблагоприятным воздействия среды.
Способностью к закаливанию обладают не все растения. Растения южного происхождения не способны переносить морозы. Способность к закаливанию у древесных и зимующих травянистых растений северных широт, переживающих значительное понижение температуры в зимний период, в период летней вегетации отсутствует и проявляется только во время наступления осенних пониженных температур (если растение к этому времени прошло необходимый цикл развития). Процесс закалки приурочен лишь к определенным этапам развития растений. Для приобретения способности к закаливанию растения должны закончить процессы роста.
Разные органы растении имеют неодинаковую способность к закаливанию, например, листья листопадных деревьев (яблоня, груша, вишня) не обладают способностью к закаливанию, цветочные почки закаливаются хуже, чем листовые. У вегетирующих растений легко вымерзают растущие и не закончившие рост органы. Выносливость растении к низким температурам в этот период незначительная.
Эффект закаливания может не проявляться, если какая-либо причина (засуха, поздний посев, посадка и др.) произошла задержка развития растений. Так, если в течение дета у плодовых растении процессы роста из-за летней засухи не успели закончиться, то зимой это может привести к гибели растений. Дело в том, что засуха, приостанавливая рост летом, не позволяет растениям завершить его к осени. Одновременно при закалке должен произойти отток различные веществ из надземных органов в подземные зимующие (корневые системы, корневища, луковицы, клубни). По этой же причине закаливание травянистых и древесных растений ухудшает избыточное азотное питание, удлиняющее период роста до поздней осени, в результате растения не способны пройти процессы закаливания и погибнут даже при небольших морозах.
Яровые злаки при озимом посеве по сравнению с озимыми растут при более пониженных положительных температурах, в весенний период почти не снижают темпов роста и не способны к закаливанию. Большую роль в закаливании играют условия внешней среды.
На озимых культурах убедительно покатана необходимость света для процесса закаливания. Сокращение фотопериода служит для растений сигналом к прекращению роста и накопления ингибиторов в растениях. Вероятно, с этих процессов начинается формирование морозоустойчивости у растений.
Растения, выращенные при несоответствующем фотопериоде не успевают завершить летний рост и не способны к закаливанию. Установлено, что длинный день способствует образованию в листьях черной смородины фитогормонов - стимуляторов роста, а короткий - накоплению ингибиторов. В естественные условиях к закаливанию способен лишь организм в целом, при обязательном наличии корневой системы. По-видимому, в корнях вырабатываются вещества, повышающие устойчивость растения к морозу.
Фазы закаливания. По И. И. Туманову (1979) процесс закаливания растений требует определенного комплекса внешние условий и проходит в две фазы, которым предшествуют замедление роста и переход растении в состояние покоя. Прекращение роста и переход в состояние покоя - необходимые условия прохождения первой фазы закаливания. Однако само по себе оно лишь немного повышает морозоустойчивость растения. У травянистых растений переход в состояние покоя происходит в период первой фазы закаливания. У древесных покой наступает в начале осени, до прохождения первой фазы закаливания.
При переходе в состояние покоя изменяется баланс фитогормонов: уменьшается содержание ауксина и гиббереллинов и увеличивается содержание абсцизовой кислоты, которая, ослабляя и ингибируя ростовые процессы, обусловливает наступление периода покоя. Поэтому обработка растений озимой пшеницы, люцерны и других культур в этот период ингибиторами роста (например, хлорхолинхлоридом - ССС или трийодбензойной кислотой) повышает устойчивость растении к низким температурам.
Первая фаза закаливания проходит на свету и при низких положительных температурах в ночное время (днем около 10 °С, ночью около 2 °С), останавливающих рост, и умеренной влажности почвы. Озимые злаки проходят первую фазу на свету при среднесуточной температуре 0,5-2 °С за 6-9 дней, древесные за 30 дней. В эту фазу продолжается дальнейшее замедление и даже происходит полная остановка ростовых процессов.
Свет в этой фазе необходим не только для фотосинтеза, но и для поддержания ультраструктур клетки. В таких условиях за счет фотосинтеза образуются сахара, а понижение температуры в ночное время значительно снижает их расход на дыхание и рост. В результате в клетках растений накапливаются сахароза, другие олигосахариды, растворимые белки и т. д., в мембранах возрастает содержание ненасыщенных жирных кислот, снижается точка замерзания цитоплазмы, отмечается некоторое уменьшение внутриклеточной воды.
Благоприятные условия для прохождения первой фазы закаливания озимых растений складываются при солнечной и прохладной (дневная температура до 10 °С) погоде, способствующей накоплению в тканях растений углеводов и других защитных веществ. В естественных условиях оптимальный срок первой фазы закаливания озимых злаков до двух недель. За это время количество сахаров в растениях возрастает до 70 % на сухую массу или до 22 % на сырую массу, т. е. близко содержанию сахаров в корнеплодах лучших сортов сахарной свеклы.
Растения озимой пшеницы можно закалить и в темноте при 2 °С, если их корни или узлы кущения погрузить в раствор сахарозы. Такие растения выдерживают морозы до -20 °С (И. И. Туманов, 1979). Накапливающиеся в процессе закаливания сахара локализуются в клеточном соке, цитоплазме, клеточных органеллах, особенно в хлоропластах. При закаливании растений высокоморозоустойчивого сорта озимой пшеницы при температуре, близкой к О °С, количество сахаров в хлоропластах листьев увеличивалось в 2,5 раза, благодаря чему хлоропласты продолжали функционировать. Повышение содержания сахаров в хлоропластах коррелирует с морозоустойчивостью растений.
В хлоропластах содержатся те же формы сахаров что и в листьях: фруктоза, глюкоза, сахароза, олигосахара (Т. И. Трунова, 1970). Имеются данные, что при накоплении сахаров процесс фотофосфорилирования продолжается даже при отрицательных температурах. Более морозоустойчивые виды и сорта растений лучше накапливают сахар именно при сочетании пониженной температуры и умеренной влажности почвы. Дело в том, что в первой фазе закаливания происходит уменьшение содержания свободной воды, а излишняя влажность почвы при дождливой осени затрудняет этот процесс, повышается вероятность в последующем образования внутриклеточного льда и гибели растений.
Метаболические изменения, наблюдаемые во время первой фазы, могут быть вызваны изменением гормонального и энергетического балансов, что определяет синтез и активацию специфических ферментов, свойства веточных мембран закаленных тканей. Накапливающаяся в тканях абсцизовая кислота увеличивает проницаемость мембран для воды, водоотдачу теток. К. концу первой фазы закаливания все зимующие растения переходят в состояние покоя. Однако процессы закалки, перестройки процессов обмена веществ продолжаются.
Вторая фаза закаливания не требует света и начинается сразу же после первой фазы при температуре немного ниже О °С. Для травянистых растений она может протекать и под снегом. Длится она около двух недель при постепенном снижении температуры до -10...-20 °С и ниже со скоростью 2-3 °С в сутки, что приводит к частичной потере воды ветками, освобождению клеток тканей от избыточного содержания воды иди витрификации (переходу воды в стеклообразное состояние). Явление витрификации воды в растительных клетках наступает при резком охлаждении (ниже -20 °С). Стеклообразная растительная ткань долго сохраняет свою жизнеспособность.
При постепенном понижении температуры в межклеточниках образуется лед и начинают функционировать механизмы, предохраняющие подготовленные в первой фазе закаливания растения от чрезмерного обезвоживания. Накопившиеся в первой фазе закаливания сахара изменяют устойчивость биоколлоидов цитоплазмы к низким температурам, возрастает относительное количество коллоидно-связанной воды.
Вторая фаза обеспечивает отток из цитозоля клеток почти всей воды, которая может замерзнуть при отрицательной температуре. При критических температурах отток воды из клеток значительно ухудшается, появляется много переохлажденной воды, которая затем замерзает внутри протопласта и может привести к гибели клеток. Следовательно, чем менее морозоустойчиво растение, тем медленнее должна протекать вторая фаза закаливания.
Действующими факторами второй фазы закаливания являются обезвоживание, вызывающее сближение молекул в цитозоле, вязкость которого соответственно увеличивается; низкая температура, уменьшающая тепловое движение молекул в протопласте. В результате во второй фазе закаливания происходит перестройка белков цитоплазмы, накапливаются низкомолекулярные водорастворимые белки, более устойчивые к обезвоживанию, синтезируются специфические белки. Содержание незамерзающей (связанной) воды в тканях зимостойкой пшеницы почти в 3 раза выше по сравнению с незимостойкой.
Перестройка цитоплазмы увеличивает проницаемость ее для воды, способствует более быстрому оттоку воды в межклеточники, что снижает опасность внутриклеточного льдообразования. При обезвоживании, происходящем под влиянием льдообразования, наблюдаются сближение и деформация белковых молекул. связи между которыми могут рваться и не восстанавливаются, что пагубно для клетки. Очевидно, при таких условиях происходит быстрое смещение структурных частиц по отношению друг к другу, что приводит к разрушению субмикроскопической структуры протопласта (И. И. Туманов).
Цитоплазма закаленных растений более устойчива к механическому давлению. Поэтому важно наличие у молекул белков сульфгидрильных и других гидрофильных группировок, которые способствуют удержанию воды, препятствуют слишком сильному сближению молекул белка. Между содержанием сульфгидрильных групп и морозоустойчивостью клеток растений установлена положительная связь. Благодаря изменению свойств молекул белков и межмолекулярных связей в процессе закаливания постепенное обезвоживание приводит к переходу цитоплазмы из состояния золя в гель.
Первая фаза закаливания повышает морозоустойчивость растений с -5 до -12 °С, вторая увеличивает морозоустойчивость, например, у пшеницы до -18...-20 °С, у ржи -до -20...-25 °С. Растения, находящиеся в глубоком органическом покое, отличаются способностью к закаливанию и выдерживают промораживание до -195 °С. Так, черная смородина после наступления состояния глубокого покоя и завершения первой фазы закаливания переносила охлаждение до -253 °С (И. И. Туманов. 1979).
Не у всех растений процесс закаливания проходит в две фазы.
У древесных растений, имеющих в тканях достаточное количество сахаров, сразу же протекают процессы, свойственные второй фазе закаливания. Однако не все растения способны к закаливанию. Теплолюбивые растения (хлопчатник, рис, бахчевые культуры) при длительном пребывании при температурах немного выше О °С не только не становятся устойчивыми, но еще сильнее повреждаются или даже погибают, так как в них накапливаются ядовитые вещества, усиливающие губительное действие на растения низких температур.
Обратимость процессов закаливания. Морозоустойчивость представляет собой процесс, а не постоянное свойство растений. Процесс закаливания обратим, при этом морозоустойчивость растений снижается. Развитие процесса закаливания в значительной степени зависит от условий его протекания. Особенно заметное влияние на морозоустойчивость оказывают условия закаливания растений в осенний период, определяемые в первую очередь соотношением числа ясных дней с пониженными положительными температурами ночью и числа пасмурных, дождливых дней с относительно сближенными высокими температурами днем и ночью. Чем это отношение выше, тем лучше условия для закалки (табл.).
Критические температуры (°С) повреждения растений озимой пшеницы при разных условиях закалки
| Регион | Условия закалки | ||
| хорошие | средние | плохие | |
| Украина, Северный Кавказ Белоруссия | -20 | -16 | -14 |
| Центрально-Черноземная зона, сев. районы Нечерноземной зоны | -22 | -18 | -16 |
| Поволжье, Западная Сибирь, Северный Казахстан | -25 | -20 | -18 |
У хорошо закаленных растений благодаря высокой концентрации клеточного сока, пониженному содержанию воды образуется значительно меньше кристаллов льда, причем не в клетке а в межклеточниках. Такие растения погибают только при очень, сильных морозах. При закаливании происходят обратимые физиологические изменения. При неустойчивой осенней и зимней погоде приобретенная в процессе закалки морозоустойчивость снижается. Наблюдается тесная связь между морозоустойчивостью растений и ростовыми процессами. Переход к состоянию покоя всегда сопровождается повышением устойчивости, а от достояния покоя к росту - снижением. В связи с этим морозоустойчивость одного и того же вида растений довольно сильно меняется в течение года: летом она минимальная (растения могут погибнуть при температурах намного выше тех, которые они выдерживают зимой), осенью увеличивается, а в конце зимы и в начале весны опять снижается. Повышение температуры весной сопровождается противоположными закаливанию физиолого-биохимическими изменениями - происходит процесс раззакаливания растений. Весной растения часто гибнут даже от небольших заморозков.
Способы повышения морозоустойчивости. Основа решения этой задачи -селекция морозоустойчивых сортов растений, хорошо адаптирующихся к климатическим условиям данного региона. Следует еще раз отметить, что процесс закаливания представляет собой временную адаптацию цитоплазмы, определяющую степень устойчивости к последующим повреждениям низкими температурами. Морозоустойчивость же формируется в соответствии с генотипом в процессе онтогенеза под влиянием определенных условий внешней среды и связана с наступлением периода покоя, его глубиной и длительностью.
Агротехника конкретного вида растений (срок и способ посева и др.) должна максимально способствовать формированию в процессе закалки реализации возможной генетически детерминированной морозоустойчивости сорта. В северных и центральных районах России с неустойчивой весной и частым возвращением весенних заморозков более устойчивы и урожайны сорта озимых хлебов и плодовых многолетних культур с более глубоким зимним покоем, с поздним сроком возобновления весенней вегетации (***). Наоборот, в районах с устойчивым нарастанием положительных температур весной преимущество имеют рано вегетирующие виды и сорта растений.
Морозоустойчивость сортов озимой пшеницы определяется не только количеством сахаров, накопленных с осени, но и их экономным расходованием в течение зимы. У растений зимостойких сортов озимой пшеницы в зимний период с понижением температуры содержание моносахаридов (глюкоза, фруктоза) увеличивается за счет расщепления сахарозы на глюкозу и фруктозу, что снижает точку замерзания клеточного сока. Узел кущения злаков, корневая шейка бобовых - своеобразная кладовая энергетических ресурсов растения в зимний период и орган побегообразования весной.
Морозоустойчивость растений озимой пшеницы положительно коррелирует с содержанием сахаров в узлах кущения. В хороших посевах озимой пшеницы в листьях в декабре содержание растворимых углеводов достигает 18-24 % (на сухое вещество), а в узлах кущения - 39-42 %. В опытах более морозоустойчивый сорт озимой пшеницы Мироновская 808 расходовал за зиму всего 10 % углеводов, а менее устойчивый сорт Безостая 1-23 % углеводов. Растения, закладывающие узлы кущения глубоко (3-4 см), как правило, более морозоустойчивы, чем те, у которых узел кущения находится близко к поверхности (1-2 см). Глубина залегания узла кущения и мощность его развития зависят от качества семян, способа посева, обработки почвы.
На морозоустойчивость существенное влияние оказывают условия почвенного питания, особенно в осенний период. Устойчивость растений к морозу возрастает на постоянно известкуемых почвах при внесении под посев озимых калийно-фосфорных одобрений, тогда как избыточные азотные удобрения, способствуя процессам роста, делают растения озимых более чувствительными к морозам. На морозоустойчивость, как и на холодостойкость растений, положительное влияние оказывают микроэлементы (кобальт, цинк, молибден, медь, ванадий и др.)Например, цинк повышает содержание связанной воды, усиливает накопление сахаров, молибден способствует увеличению содержания общего и белкового азота.
Методы изучения морозоустойчивости растений. И. И. Тумановым с сотрудниками предложены лабораторные методы ускоренного определения морозоустойчивости различных культурных растений. Испытуемые растения после закаливания подвергают воздействию критических низких температур в холодильных камерах, что дает возможность выявить невымерзающие растения. Такая ускоренная оценка морозоустойчивости имеет большое преимущество перед обычным полевым способом оценки, так как последний требует много времени (иногда нескольких лет).
Другие надежные и удобные в исполнении лабораторные методы определения морозоустойчивости основаны на измерении вязкости цитоплазмы в клетках тканей исследуемых органов, определении электропроводности и др. Определение морозоустойчивости культурных растений мирового ассортимента показало, что страны СНГ обладают самыми устойчивыми их формами. Наиболее устойчивые сорта озимой пшеницы выведены опытными учреждениями юго-востока и северо-востока России, где природная обстановка благоприятствует выведению морозоустойчивых форм.
studfiles.net
Морозостойкость и зимостойкость растений. Закаливание растений. — Biosfera Wiki
Закаливание растений
Холодоустойчивость - способность растений длительное время переносить низкие положительные температуры (от 1 да 10°С). Она свойственна растениям умеренной зоны. К холодоустойчивым культурам относятся ячмень, овес, лен, вика и др.
Степень холодоустойчивости разных растений, а также разных органов одного и того же растения неодинакова. Основная причина гибели теплолюбивых растений от действия низких положительных температур связана прежде всего с дезорганизацией обмена нуклеиновых кислот и белков, нарушением проницаемости цитоплазмы (повышением ее вязкости), прекращением тока ассимилятов и накоплением токсических веществ в клетке.
Холодоустойчивость определяется способностью растений сохранять нормальную структуру цитоплазмы и изменять обмен веществ в период охлаждения и последующего повышения температуры.
Морозоустойчивость - способность растений переносить температуру ниже 0°С. Разные растения переносят зимние условия, находясь в различном состоянии. У однолетних растений зимуют семена, нечувствительные к морозам, у многих - защищенные слоем земли и снега клубни, луковицы и корневища. У озимых растений и древесных пород ткани под действием отрицательных температур могут замерзнуть и даже промерзнуть насквозь, однако растения не погибают. Способность этих растений перезимовывать обусловливается их достаточно высокой морозоустойчивостью.
Поврежденные морозом растения имеют вид как бы обваренных, они утрачивают тургор, листья их буреют и засыхают. При оттаивании клубней картофеля или корнеплодов сахарной свеклы вода легко вытекает из тканей. Такое явление длительное время объясняли разрывом клеточных стенок под влиянием льда, образующегося в тканях растений. Однако установлено, что лед образуется главным образом в межклетниках и клеточные стенки остаются неповрежденными. Гибель растений под влиянием морозов обусловливается изменениями, происходящими в протопласте, его коагуляцией. Физико-химические преобразования в протопласте происходят вследствие оттягивания воды образующимися в межклетниках кристаллами. Кроме того, протопласт подвергается сжатию со стороны растущих в межклетниках кристаллов. В результате наступает необратимая денатурация коллоидов протопласта клеток и отмирание тканей. Если льда образуется немного, то после оттаивания растение может остаться живым.
Однако образование льда в межклетниках неопасно, и после оттаивания листья возвращаются в нормальное состояние. Каждая клетка имеет свою границу обезвоживания и сжатия. Переход этих границ, а не только снижение температуры - причина гибели клеток. Такое явление следует рассматривать не как непосредственное влияние холода на протопласт, а как действие, вызывающее обезвоживание протопласта вследствие вымораживания воды. Убедительным доказательством этого служит состояние переохлаждения (без образования льда), которое растения переносят без вреда; при тех же температурах, но с образованием льда в тканях растения гибнут.
Нечувствительность к морозам достигается физико-химическими изменениями в клетках. В зимующих листьях и других частях растения накапливается много сахара, а крахмала в них почти нет. Сахар защищает белковые соединения от коагуляции при вымораживании, и поэтому его можно назвать защитным веществом. При достаточном количестве сахара в клетках повышаются водоудерживающие силы коллоидов протопласта, увеличивается количество прочносвязанной и уменьшается содержание свободной воды. Связанная с коллоидами вода при действии низких температур не превращается в лед. У ряда древесных пород в результате преобразования углеводов в коровой паренхиме накапливаются жиры и липоиды, которые не замерзают и проявляют защитное действие в зимний период. Кроме того, белковые вещества, частично гидролизуясь, переходят из менее устойчивой в более устойчивую форму азотсодержащих веществ-аминокислоты, которые при замерзании в меньшей мере подвергаются денатурации.
Степень морозоустойчивости клеток во многом зависит от состояния плазмалеммы, обмена веществ, образования сложных органических соединений и массы цитоплазмы. В клетках внутренние слои цитоплазмы повреждаются раньше, чем плазмалемма, которая способна к быстрому новообразованию за счет молекул остальной массы цитоплазмы. Недостаточно закаленные клетки не могут быстро восстанавливать плазмалемму. На степень морозоустойчивости растений большое влияние оказывают сахара, регуляторы роста и другие вещества, образующиеся в клетках.
Теория закаливания растений, повышающего их устойчивость к действию низких температур.
Сущность ее заключается в том, что у растений под влиянием низких положительных температур накапливаются сахара и другие соединения - первая фаза закаливания. Дальнейшее повышение морозоустойчивости происходит уже при отрицательных температурах, но не повреждающих клетки, - вторая фаза закаливания. Она идет сразу же после первой при температуре немного ниже 0°С. В этой фазе наблюдается частичная потеря воды клетками. Под действием сахаров, накопившихся в клетках, изменяются биоколлоиды и возрастает относительное почек узла кущения, что свидетельствует о приспособленности к прохождению озимыми растениями так называемого зимнего покоя. При этом глубина залегания узла кущения и мощность его развития зависят от качества семян, способа посева, обработки почвы и оказывают большое влияние на перезимовку озимых. Обычно, когда узел кущения находится близко от поверхности (1,5 см}, такие растения менее зимостойки, чем растения с более глубоким залеганием узла кущения (3-4 см).
Существует связь зимостойкости озимой пшеницы с содержанием сахаров в узлах кущения. Отмечено интенсивное использование растворимых углеводов во второй половине зимы. Узел кущения - своеобразная кладовая энергетических ресурсов растения в зимний период. Морозостойкость сортов озимой пшеницы определяется как количеством сахаров, накопленных в осенний период, так и экономным расходованием их в течение зимы.
biosfera.wiki
Зоны морозостойкости, карта, растения-индикаторы | WikiBotanika.ru
В последнее время прилавки магазинов и рынков пестрят экзотическими для нашей страны растениями. Да и знакомые культуры пополняются новыми разновидностями и сортами. Продавцы уверяют, что все эти культуры районированы и прекрасно приспособлены для выращивания в нашем непростом климате. Но так ли это на самом деле?
Климат нашей страны, отличающийся теплым или даже жарким летом, но холодной и продолжительной зимой, подходит, естественно, далеко не для всей флоры. Оценить способность растений выдержать холодную зиму помогает понятие, называемое зимостойкостью. Некоторые (мхи и лишайники) отлично переносят даже самые низкие зимние температуры, другие (береза, дуб) могут расти только в более щадящих условиях, ну а третьи (магнолия, олеандр) и вовсе не приспособлены для суровых зим.
Какие растения адаптированы для нашего климата знает, пожалуй, каждый опытный садовод. Но что делать с новинками рынка, как узнать переживет ли вновь приобретенный зеленый питомец суровую зиму? На самом деле, никакой проблемы здесь нет – в справочниках, документах и на этикетках экземпляров из питомников указывается зона морозостойкости, подходящая для растения. Давайте разберемся, что же это такое.
Что такое зоны морозостойкости
Зоны морозостойкости (или зимостойкости) – это климатические области земного шара, подходящие для выращивания тех или иных видов растений. Они основаны на температурной шкале, показывающей среднегодовую минимальную температуру в градусах Цельсия или Фаренгейта.

В настоящее время наиболее полной шкалой морозостойкости, используемой повсеместно, является так называемая шкала или зоны USDA.
Зоны USDA
Впервые шкала USDA была разработана Министерством сельского хозяйства США для нужд фермеров и растениеводческих хозяйств, а затем предложенное зонирование стало использоваться во всем мире. Шкала включает в себя 13 зон (от 0 до 12), каждая из которых разбита еще на 2 подзоны, границы которых определялись на основе минимальных среднегодовых температур. Вначале была описана только территория Соединенных Штатов, а затем и другие страны. Сейчас все регионы земного шара относятся к той или иной зоне по шкале USDA, а все виды растений разделены согласно приспособленности к различным климатическим условиям.
К примеру, растение, описанное как «выносливое к зоне 10», означает, что оно может выдерживать минимальную температуру -1 °C. Более устойчивое растение, «выносливое к зоне 9», может выдерживать минимальную температуру -7 °C и т.д.

Зонирование USDA на сегодняшний момент – универсальная шкала морозостойкости, используемая современными хозяйствами. Однако она имеет множество недостатков и не учитывает целый ряд важнейших факторов, таких как:
- Высота снежного покрова. Известно, что снег отлично защищает корневую систему растений от морозов, поэтому культуры, вымерзающие в бесснежных регионах, могут неплохо сохраняться там, где зимой выпадает много снега.
[!] Селекционер растений из Польши заметил одну интересную особенность: азалии, умирающие от холода в Восточной Европе, отлично зимовали в Московской области, там, где климат гораздо суровее. Оказалось, что рододендроны защищал снег, не дававший погибнуть корневой системе.
- Перепады температуры. Многие культуры боятся не столько морозов, сколько резких изменений температуры зимой. Внезапная оттепель, вызывающая таяние снега, и столь же внезапное похолодание приводит к гибели корней. Именно поэтому целый ряд растений, например некоторые виды бамбука, которые способны выдерживать сильные морозы, не могут выжить в нашем климате.
[!] В 60-х годах прошлого столетия из-за такого внезапного перепада зимней температуры (от -1°С до -29°С) сильно пострадала коллекция кленов ботанического сада МГУ.
- Весенние заморозки – еще один фактор, сдерживающий выращивание теплолюбивых видов. В нашей стране эта проблема особенно актуальна: в средней полосе морозы могут вернуться даже в конце мая.
[!] За примерами далеко ходить не надо. Майские холода 2017 года погубили значительную часть винограда в Крыму.
- Среднегодовое количество осадков. Влажность – важный климатический параметр, влияющий, в том числе и на адаптацию растений. Например, некоторые культуры неплохо переносят сухой холодный воздух, но не выживают во влажном климате.
- Другие факторы: разница между дневными и ночными температурами, рельеф местности (наличие возвышенностей и низин), виды почв, продолжительность светового дня, роза ветров и т.д.
Россия, поделенная на климатические зоны USDA, выглядит примерно так:

Некоторые города нашей страны относятся к следующим зонам морозостойкости: зона 1 – Тикси, Батагай, зона 2 – бо́льшая территория России, Красноярск, Иркутск, Новосибирск, зона 3 – Хабаровск, Магадан, Воркута, зона 4 – Москва и Московская область, Вологда, Архангельск, Челябинск, Уфа, зона 5 – Санкт-Петербург, Владивосток, Воронеж, Брянск, Саратов, зона 6 и 7 – Краснодар.
Что касается этикеток на растениях из питомников, то для Москвы и Санкт-Петербурга следует выбирать культуры, акклиматизированные для зон 1-4, а также 5a и, частично, 5b.
Как видите, зонирование USDA далеко от совершенства, поэтому работы в этом направлении постоянно продолжаются.
Альтернативы
Несовершенная система USDA побудила искать альтернативы. И сейчас используют не только вышеописанную концепцию, но и некоторые другие. Во-первых, USDA-зоны были дополнены, а их количество значительно увеличено. Кроме того, одним из средств определения зон морозостойкости стало использование так называемых «индикаторных растений». Известно, что ареал некоторых видов строго ограничен в зависимости от климата, а потому наличие какого-либо индикаторного растения говорит о принадлежности территории к той или иной зоне морозостойкости.
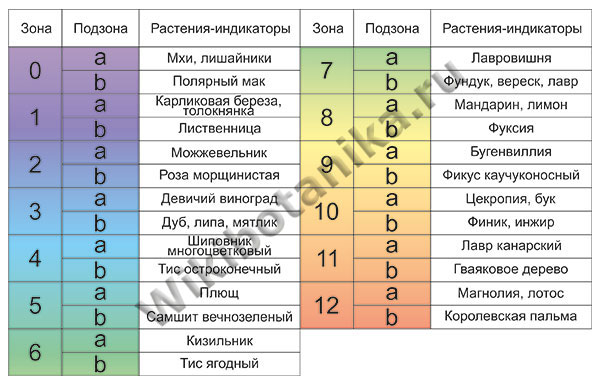
Кроме того, некоторые страны разработали свои климатические карты, основанные на многолетних метеорологических наблюдениях. Одной из таких стран стала Великобритания.
В 2012 году Королевское садоводческое общество Соединенного Королевства представило свой рейтинг морозостойкости растений. Этот рейтинг описывает все растения, встречающиеся в Великобритании, и широко используется английскими садоводами.
Английская система морозостойкости растений состоит из 9 частей:
- h2a (более 15°C) – выращивание в теплице;
- Н1в (от 10 до 15°С) и h2c (от 5 до 10°С) – выращивание на открытом воздухе только летом;
- Н2 (от 1 до 5°С) – культуры, способные переносить небольшое понижение температуры, но неустойчивые к заморозкам;
- Н3 (от -5 до 1°С) – культуры, хорошо переносящие климат прибрежных районов Великобритании;
- Н4 (от -10 до -5°С) – культуры, приспособленные к климату большей части Великобритании;
- Н5 (от -15 до -10°С) – культуры, приспособленные к климату большей части Великобритании и выдерживающие значительное понижение температуры зимой;
- Н6 (от -20 до – 15°С) – морозостойкие культуры, способные расти на всей территории Северной Европы;
- Н7 (менее -20°С) – наиболее морозостойкие культуры.
Зоны морозостойкости в СССР и России
В нашей стране работы по определению зон морозостойкости были начаты еще в начале XX века, а затем продолжились после октябрьской революции. И, хотя вначале карты были несовершенны, а климатические районы слишком обобщены, в 60-х годах работа была продолжена: количество зон увеличилось до 42 (до 60 вместе с подзонами). Работа получила название «Древокультурные районы СССР». Кроме зонирования была проведена работа и по разделению растений, встречающихся в СССР, по климатическим районам. Список состоял, главным образом, из древесных культур, однако количество видов, вошедших в него, весьма велико – около 700.
Полученные данные профессор Колесников А.И., руководивший работой, обобщил в издании «Декоративная дендрология». Эта книга до сих пор не утратила актуальности.
С подробным описанием древокультурных районов СССР вы можете ознакомиться в этой статье.
На этом работа не остановилась, а детализация зонирования продолжилась. Стали учитываться и другие факторы, влияющие на зимостойкость растений: не только минимальные, но и среднегодовые температуры зимних и летних месяцев, средняя и минимальная влажность, испаряемость и годовые осадки. Количество зон морозостойкости увеличилось до 76, а растения, рекомендуемые для каждой зоны, были разделены на три группы – «основная», «дополнительная» и «вспомогательная»:
- основная – растения, хорошо акклиматизированные для района,
- дополнительная – культуры, способные расти в рассматриваемой климатической зоне только при условии хорошего ухода (укрытия на зиму),
- вспомогательная – единично встречающиеся растения.
К сожалению, серьезных государственных исследований на эту тему больше не проводилось, хотя изменился и климат, и ассортимент растений, накопился колоссальный практический опыт. Время от времени подобные карты пытаются составить отдельные хозяйства, но из-за нехватки ресурсов их работа ограничивается отдельными районами.
Как повысить морозостойкость растений
В заключение хочется дать несколько советов из практики садоводства. Многие культуры, рекомендованные для более благоприятных климатических зон, при должном уходе вполне возможно выращивать и у нас. Для повышения зимостойкости отдельных видов рекомендуется:
- Защищать от морозов корневую систему. Это можно сделать с помощью хвойного лапника, сухих опавших листьев, торфа, опилок, промышленного укрывного материала. Хорошо зарекомендовал себя и пенопласт, обладающий высокими теплоизолирующими свойствами.
- Укрывать не только корни, но и надземную часть растения. Конечно, крону мощных древесных культур защитить сложно (хотя некоторые садоводы и умудряются обматывать нижнюю часть стволов тканью или полиэтиленом), а вот лианоподобные виды и растения с гибкими стеблями следует аккуратно уложить на землю и укрыть.
[!] Особенно важно защищать молодые культуры, возраст которых – 2-3 года. Их корневая система еще не до конца сформирована и располагается близко к поверхности земли.
[!] Зимние укрытия следует устраивать только после наступления первых морозов. На более ранних сроках в них могут поселиться мыши и другие грызуны.
- Своевременно вносить удобрения и подкормки. Это повысит выносливость, а значит, и сопротивляемость растения к холодам.
- Мульчировать почву. Мульчирование необходимо не только для защиты от морозов, но и для сохранения влаги в земле.
- Обращать внимание на структуру почвы и её кислотность, при необходимости обогащать субстрат.
Конечно, придется приложить немало усилий, но и результат того стоит: какого садовода не порадует редкий экзот на участке. Ну, а от российских ученых мы будем ждать новых исследований в этой необычайно интересной области.
wikibotanika.ru
Морозоустойчивость растений
Морозоустойчивость- это способность переносить температуру ниже О °С, низкие отрицательные температуры. Морозоустойчивые растения способны предотвращать или уменьшать действие низких отрицательных температур. Морозы в зимний период с температурой ниже -20 °С обычны для значительной части территории России. Воздействию морозов подвергаются однолетние, двулетние и многолетние растения. Растения переносят условия зимы в различные периоды онтогенеза. У однолетних культур зимуют семена (яровые растения), раскустившиеся растения (озимые), у двулетних и многолетних - клубни, корнеплоды, луковицы, корневища, взрослые растения. Способность озимых, многолетних травянистых и древесных плодовых культур перезимовывать обусловливается их достаточно высокой морозоустойчивостью. Ткани зимостойких растений могут замерзать, однако растения не погибают. Большой вклад в изучение физиологических основ морозоустойчивости внесли Я. А. Максимов (1952), Г. А. Самыгин (1974), И. И. Туманов (1979) и другие отечественные исследователи.
Замерзание растительных клеток и тканей и происходящие при этом процессы. Способность растении переносить отрицательные температуры определяется наследственной основой данного вида растений, однако морозоустойчивость одного я того же растения зависят от условии, предшествующих наступлению морозов, влияющих на характер льдообразования. Лед может образовываться как в протопласте клетки, так и в межклеточном пространстве. Не всякое образование льда приводит клетки растения к гибели.
Постепенное снижение температуры со скоростью 0,5-1 °С/ч приводит к образованию кристаллов льда прежде всего в межклеточниках и первоначально не вызывают гибели клеток. Однако последствия этого процесса могут быть губительными для клетки. Образование льда в протопласте клетки, как правило, происходит при быстром понижении температуры. Происходит коагуляция белков протоплазмы, кристаллами образовавшегося в цитозоле льда повреждаются клеточные структуры, клетки погибают. Убитые морозом растения после оттаивания теряют тургор, из их мясистых тканей вытекает вода.
Условия и причины вымерзания растений. Образующийся при медленном промерзании в межклеточниках и клеточных стенках лед оттягивает воду из клеток; клеточный сок становится концентрированным, изменяется рН среды. Выкристаллизовавшийся лед действует как сухой воздух, иссушая клетки и сильно изменяя их осмотические свойства. Кроме того, цитоплазма подвергается сжатию кристаллами льда. Образующиеся кристаллы льда вытесняют воздух из межклеточников, поэтому замерзшие листья становятся прозрачными.
Если льда образуется немного и клетки не были механически повреждены его кристаллами, то при последующем оттаивания такие растения могут сохранить жизнеспособность. Так, в листьях капусты при температуре -5...-6 °С образуется некоторое количество льда в межклеточниках. Однако при последующем медленном оттаивании межклеточники заполняются водой, которая поглощается клетками, и листья возвращаются в нормальное состояние.
Однако клетки, ткани и растения могут погибнуть от мороза. Основными причинами гибели меток растений при низких отрицательных температурах и льдообразовании являются чрезмерное обезвоживание клеток или механическое давление, сжатие клеток кристаллами льда, повреждающее тонкие клеточные структуры. Оба эти фактора могут действовать одновременно.
Летальность действия мороза определяется несколькими обстоятельствами. Последствия воздействия низких отрицательных температур зависят от оводненности тканей растения. Насыщенные водой ткани легко повреждаются, сухие же семена могут выносить глубокие низкие температуры (до -196 °С). Низкое содержание воды предохраняет от образования льда в растениях при промораживании. Разные растения, их клетки имеют свой критический предел обезвоживания и сжатия.
Гибель клеток, тканей и растений под действием морозов обусловливается необратимыми изменениями, происходящими в протопласте клеток: его коагуляцией, денатурацией коллоидов протопласта, механическим давлением льда, повреждающим поверхностные структуры цитоплазмы, кристаллами льда, нарушающими мембраны и проникающими внутрь клетки. Вредное влияние оказывает повышение концентрации и изменение рН клеточного сока, сопровождающие обезвоживание клеток.
Действие льда, особенно при длительном влиянии низких температур, сходно с обезвоживанием клеток при засухе. Признаками повреждения клеток морозом являются потеря ими тургора, инфильтрация межклеточников водой и выход ионов из клеток. Выход ионов К и сахаров из клеток, по-видимому, связан с повреждением мембранных систем их активного транспорта. Поврежденные растения при переносе в теплое место имеют вид ошпаренных, утрачивают тургор, листья быстро буреют и засыхают. При оттаивании мороженых клубней картофеля, корнеплодов кормовой и сахарной свеклы вода легко вытекает из тканей. Важно отметить, что состояние переохлаждения (без образования льда) растения переносят без вреда; при тех же температурах, но с образованием льда в тканях растения гибнут.
Растения по-разному реагируют на образование льда в тканях: клубни картофеля и георгина быстро погибают, капуста и лук переносят лишь умеренное промораживание, рожь и пшеница выдерживают на уровне узла кущения морозы до -15...-20 °С.
У устойчивых к морозу растений имеются защитные механизмы, в основе которых лежат определенные физико-химические изменения. Морозоустойчивые растения обладают приспособлениями, уменьшающими обезвоживание клеток.
При понижении температуры у таких растений отмечаются повышение содержания сахаров и других веществ, защищающих ткани (криопротекторы), это прежде всего гидрофильные белки, моно- и олигосахариды;
снижение оводненности клеток;
увеличение количества полярных липидов и снижение насыщенности их жирнокислотных остатков,
увеличение количества защитных белков.
На степень морозоустойчивости растений большое влияние (оказывают сахара, регуляторы роста и другие вещества, образующиеся в клетках. В зимующих растениях в цитоплазме накапливаются сахара, а содержание крахмала снижается. Влияние сахаров на повышение морозоустойчивости растений многосторонне (предохраняет от замерзания большой объем внутриклеточной воды, заметно уменьшает количество образующегося льда).
Сахара защищают белковые соединения от коагуляции при вымораживании; они образуют гидрофильные связи с белками цитоплазмы, предохраняя их от возможной денатурации, повышают осмотическое давление и снижают температуру замерзания цитозоля. В результате накопления сахаров содержание прочносвязанной воды увеличивается, а свободной уменьшается. Особое значение имеет защитное влияние сахаров на белки. сосредоточенные в поверхностных мембранах клетки. Сахара увеличивают водоудерживающую способность коллоидов протоплазмы клеток; связанная с коллоидами вода в виде гидратных оболочек биополимеров при низких температурах не замерзает и не транспортируется.
Криопротекторами являются также молекулы гемицеллюлоз (ксиланы, арабиноксиланы), выделяемые цитоплазмой в клеточную стенку, обволакивающие растущие кристаллы льда, что предотвращает образование крупных кристаллов, повреждающих клетку. Так клетки защищаются как от внутриклеточного льда, так и от чрезмерного обезвоживания.
Значительное количество защитных белков и модификации молекул липидов увеличивают структурированность клеток. У большинства растений возрастает синтез водорастворимых белков. Белковые вещества, частично гидролизуясь, увеличивают содержание свободных АК.
В тканях морозоустойчивых растений в конце лета и осенью накапливаются в достаточно количестве запасные вещества (прежде всего сахара), которые используются весной при возобновлении роста, обеспечивая потребности растении в строительном материале и энергии. Необходимо также учитывать устойчивость растении к болезням, вероятность развития которых увеличивается при повреждении тканей морозом.
Закаливание растений, его фазы. Морозоустойчивость - не постоянное свойство растений. Она зависит от физиологического состояния растении и условии внешней среды. Растения, выращенные при относительно низких положительных температурах, более устойчивы, чем выращенные при относительно высоких осенних температурах. Свойство морозоустойчивости формируется в процессе онтогенеза растения под влиянием определенных условии среды в соответствии с генотипом растения и связано с резким снижением темпов роста, переходом растения в состояние покоя.
Жизненный цикл развития озимых, двуручек, двулетних и многолетних растении контролируется сезонным ритмом светового и температурного периодов. В отличие от яровых однолетних растении они начинают готовиться к перенесению неблагоприятных зимних условии с момента остановки роста и затем в течение осени во время наступления пониженных температур.
Повышение морозоустойчивости растении тесно связано с закаливанием - постепенной подготовкой к воздействию низких зимних температур. Закаливание - это обратимая физиологическая устойчивость к неблагоприятным воздействия среды.
Способностью к закаливанию обладают не все растения. Растения южного происхождения не способны переносить морозы. Способность к закаливанию у древесных и зимующих травянистых растений северных широт, переживающих значительное понижение температуры в зимний период, в период летней вегетации отсутствует и проявляется только во время наступления осенних пониженных температур (если растение к этому времени прошло необходимый цикл развития). Процесс закалки приурочен лишь к определенным этапам развития растений. Для приобретения способности к закаливанию растения должны закончить процессы роста.
Разные органы растении имеют неодинаковую способность к закаливанию, например, листья листопадных деревьев (яблоня, груша, вишня) не обладают способностью к закаливанию, цветочные почки закаливаются хуже, чем листовые. У вегетирующих растений легко вымерзают растущие и не закончившие рост органы. Выносливость растении к низким температурам в этот период незначительная.
Эффект закаливания может не проявляться, если какая-либо причина (засуха, поздний посев, посадка и др.) произошла задержка развития растений. Так, если в течение дета у плодовых растении процессы роста из-за летней засухи не успели закончиться, то зимой это может привести к гибели растений. Дело в том, что засуха, приостанавливая рост летом, не позволяет растениям завершить его к осени. Одновременно при закалке должен произойти отток различные веществ из надземных органов в подземные зимующие (корневые системы, корневища, луковицы, клубни). По этой же причине закаливание травянистых и древесных растений ухудшает избыточное азотное питание, удлиняющее период роста до поздней осени, в результате растения не способны пройти процессы закаливания и погибнут даже при небольших морозах.
Яровые злаки при озимом посеве по сравнению с озимыми растут при более пониженных положительных температурах, в весенний период почти не снижают темпов роста и не способны к закаливанию. Большую роль в закаливании играют условия внешней среды.
На озимых культурах убедительно покатана необходимость света для процесса закаливания. Сокращение фотопериода служит для растений сигналом к прекращению роста и накопления ингибиторов в растениях. Вероятно, с этих процессов начинается формирование морозоустойчивости у растений.
Растения, выращенные при несоответствующем фотопериоде не успевают завершить летний рост и не способны к закаливанию. Установлено, что длинный день способствует образованию в листьях черной смородины фитогормонов - стимуляторов роста, а короткий - накоплению ингибиторов. В естественные условиях к закаливанию способен лишь организм в целом, при обязательном наличии корневой системы. По-видимому, в корнях вырабатываются вещества, повышающие устойчивость растения к морозу.
Фазы закаливания. По И. И. Туманову (1979) процесс закаливания растений требует определенного комплекса внешние условий и проходит в две фазы, которым предшествуют замедление роста и переход растении в состояние покоя. Прекращение роста и переход в состояние покоя - необходимые условия прохождения первой фазы закаливания. Однако само по себе оно лишь немного повышает морозоустойчивость растения. У травянистых растений переход в состояние покоя происходит в период первой фазы закаливания. У древесных покой наступает в начале осени, до прохождения первой фазы закаливания.
При переходе в состояние покоя изменяется баланс фитогормонов: уменьшается содержание ауксина и гиббереллинов и увеличивается содержание абсцизовой кислоты, которая, ослабляя и ингибируя ростовые процессы, обусловливает наступление периода покоя. Поэтому обработка растений озимой пшеницы, люцерны и других культур в этот период ингибиторами роста (например, хлорхолинхлоридом - ССС или трийодбензойной кислотой) повышает устойчивость растении к низким температурам.
Первая фаза закаливания проходит на свету и при низких положительных температурах в ночное время (днем около 10 °С, ночью около 2 °С), останавливающих рост, и умеренной влажности почвы. Озимые злаки проходят первую фазу на свету при среднесуточной температуре 0,5-2 °С за 6-9 дней, древесные за 30 дней. В эту фазу продолжается дальнейшее замедление и даже происходит полная остановка ростовых процессов.
Свет в этой фазе необходим не только для фотосинтеза, но и для поддержания ультраструктур клетки. В таких условиях за счет фотосинтеза образуются сахара, а понижение температуры в ночное время значительно снижает их расход на дыхание и рост. В результате в клетках растений накапливаются сахароза, другие олигосахариды, растворимые белки и т. д., в мембранах возрастает содержание ненасыщенных жирных кислот, снижается точка замерзания цитоплазмы, отмечается некоторое уменьшение внутриклеточной воды.
Благоприятные условия для прохождения первой фазы закаливания озимых растений складываются при солнечной и прохладной (дневная температура до 10 °С) погоде, способствующей накоплению в тканях растений углеводов и других защитных веществ. В естественных условиях оптимальный срок первой фазы закаливания озимых злаков до двух недель. За это время количество сахаров в растениях возрастает до 70 % на сухую массу или до 22 % на сырую массу, т. е. близко содержанию сахаров в корнеплодах лучших сортов сахарной свеклы.
Растения озимой пшеницы можно закалить и в темноте при 2 °С, если их корни или узлы кущения погрузить в раствор сахарозы. Такие растения выдерживают морозы до -20 °С (И. И. Туманов, 1979). Накапливающиеся в процессе закаливания сахара локализуются в клеточном соке, цитоплазме, клеточных органеллах, особенно в хлоропластах. При закаливании растений высокоморозоустойчивого сорта озимой пшеницы при температуре, близкой к О °С, количество сахаров в хлоропластах листьев увеличивалось в 2,5 раза, благодаря чему хлоропласты продолжали функционировать. Повышение содержания сахаров в хлоропластах коррелирует с морозоустойчивостью растений.
В хлоропластах содержатся те же формы сахаров что и в листьях: фруктоза, глюкоза, сахароза, олигосахара (Т. И. Трунова, 1970). Имеются данные, что при накоплении сахаров процесс фотофосфорилирования продолжается даже при отрицательных температурах. Более морозоустойчивые виды и сорта растений лучше накапливают сахар именно при сочетании пониженной температуры и умеренной влажности почвы. Дело в том, что в первой фазе закаливания происходит уменьшение содержания свободной воды, а излишняя влажность почвы при дождливой осени затрудняет этот процесс, повышается вероятность в последующем образования внутриклеточного льда и гибели растений.
Метаболические изменения, наблюдаемые во время первой фазы, могут быть вызваны изменением гормонального и энергетического балансов, что определяет синтез и активацию специфических ферментов, свойства веточных мембран закаленных тканей. Накапливающаяся в тканях абсцизовая кислота увеличивает проницаемость мембран для воды, водоотдачу теток. К. концу первой фазы закаливания все зимующие растения переходят в состояние покоя. Однако процессы закалки, перестройки процессов обмена веществ продолжаются.
Вторая фаза закаливания не требует света и начинается сразу же после первой фазы при температуре немного ниже О °С. Для травянистых растений она может протекать и под снегом. Длится она около двух недель при постепенном снижении температуры до -10...-20 °С и ниже со скоростью 2-3 °С в сутки, что приводит к частичной потере воды ветками, освобождению клеток тканей от избыточного содержания воды иди витрификации (переходу воды в стеклообразное состояние). Явление витрификации воды в растительных клетках наступает при резком охлаждении (ниже -20 °С). Стеклообразная растительная ткань долго сохраняет свою жизнеспособность.
При постепенном понижении температуры в межклеточниках образуется лед и начинают функционировать механизмы, предохраняющие подготовленные в первой фазе закаливания растения от чрезмерного обезвоживания. Накопившиеся в первой фазе закаливания сахара изменяют устойчивость биоколлоидов цитоплазмы к низким температурам, возрастает относительное количество коллоидно-связанной воды.
Вторая фаза обеспечивает отток из цитозоля клеток почти всей воды, которая может замерзнуть при отрицательной температуре. При критических температурах отток воды из клеток значительно ухудшается, появляется много переохлажденной воды, которая затем замерзает внутри протопласта и может привести к гибели клеток. Следовательно, чем менее морозоустойчиво растение, тем медленнее должна протекать вторая фаза закаливания.
Действующими факторами второй фазы закаливания являются обезвоживание, вызывающее сближение молекул в цитозоле, вязкость которого соответственно увеличивается; низкая температура, уменьшающая тепловое движение молекул в протопласте. В результате во второй фазе закаливания происходит перестройка белков цитоплазмы, накапливаются низкомолекулярные водорастворимые белки, более устойчивые к обезвоживанию, синтезируются специфические белки. Содержание незамерзающей (связанной) воды в тканях зимостойкой пшеницы почти в 3 раза выше по сравнению с незимостойкой.
Перестройка цитоплазмы увеличивает проницаемость ее для воды, способствует более быстрому оттоку воды в межклеточники, что снижает опасность внутриклеточного льдообразования. При обезвоживании, происходящем под влиянием льдообразования, наблюдаются сближение и деформация белковых молекул. связи между которыми могут рваться и не восстанавливаются, что пагубно для клетки. Очевидно, при таких условиях происходит быстрое смещение структурных частиц по отношению друг к другу, что приводит к разрушению субмикроскопической структуры протопласта (И. И. Туманов).
Цитоплазма закаленных растений более устойчива к механическому давлению. Поэтому важно наличие у молекул белков сульфгидрильных и других гидрофильных группировок, которые способствуют удержанию воды, препятствуют слишком сильному сближению молекул белка. Между содержанием сульфгидрильных групп и морозоустойчивостью клеток растений установлена положительная связь. Благодаря изменению свойств молекул белков и межмолекулярных связей в процессе закаливания постепенное обезвоживание приводит к переходу цитоплазмы из состояния золя в гель.
Первая фаза закаливания повышает морозоустойчивость растений с -5 до -12 °С, вторая увеличивает морозоустойчивость, например, у пшеницы до -18...-20 °С, у ржи -до -20...-25 °С. Растения, находящиеся в глубоком органическом покое, отличаются способностью к закаливанию и выдерживают промораживание до -195 °С. Так, черная смородина после наступления состояния глубокого покоя и завершения первой фазы закаливания переносила охлаждение до -253 °С (И. И. Туманов. 1979).
Не у всех растений процесс закаливания проходит в две фазы.
У древесных растений, имеющих в тканях достаточное количество сахаров, сразу же протекают процессы, свойственные второй фазе закаливания. Однако не все растения способны к закаливанию. Теплолюбивые растения (хлопчатник, рис, бахчевые культуры) при длительном пребывании при температурах немного выше О °С не только не становятся устойчивыми, но еще сильнее повреждаются или даже погибают, так как в них накапливаются ядовитые вещества, усиливающие губительное действие на растения низких температур.
Обратимость процессов закаливания. Морозоустойчивость представляет собой процесс, а не постоянное свойство растений. Процесс закаливания обратим, при этом морозоустойчивость растений снижается. Развитие процесса закаливания в значительной степени зависит от условий его протекания. Особенно заметное влияние на морозоустойчивость оказывают условия закаливания растений в осенний период, определяемые в первую очередь соотношением числа ясных дней с пониженными положительными температурами ночью и числа пасмурных, дождливых дней с относительно сближенными высокими температурами днем и ночью. Чем это отношение выше, тем лучше условия для закалки (табл.).
Критические температуры (°С) повреждения растений озимой пшеницы при разных условиях закалки
| Регион | Условия закалки | ||
| хорошие | средние | плохие | |
| Украина, Северный Кавказ Белоруссия | -20 | -16 | -14 |
| Центрально-Черноземная зона, сев. районы Нечерноземной зоны | -22 | -18 | -16 |
| Поволжье, Западная Сибирь, Северный Казахстан | -25 | -20 | -18 |
У хорошо закаленных растений благодаря высокой концентрации клеточного сока, пониженному содержанию воды образуется значительно меньше кристаллов льда, причем не в клетке а в межклеточниках. Такие растения погибают только при очень, сильных морозах. При закаливании происходят обратимые физиологические изменения. При неустойчивой осенней и зимней погоде приобретенная в процессе закалки морозоустойчивость снижается. Наблюдается тесная связь между морозоустойчивостью растений и ростовыми процессами. Переход к состоянию покоя всегда сопровождается повышением устойчивости, а от достояния покоя к росту - снижением. В связи с этим морозоустойчивость одного и того же вида растений довольно сильно меняется в течение года: летом она минимальная (растения могут погибнуть при температурах намного выше тех, которые они выдерживают зимой), осенью увеличивается, а в конце зимы и в начале весны опять снижается. Повышение температуры весной сопровождается противоположными закаливанию физиолого-биохимическими изменениями - происходит процесс раззакаливания растений. Весной растения часто гибнут даже от небольших заморозков.
Способы повышения морозоустойчивости. Основа решения этой задачи -селекция морозоустойчивых сортов растений, хорошо адаптирующихся к климатическим условиям данного региона. Следует еще раз отметить, что процесс закаливания представляет собой временную адаптацию цитоплазмы, определяющую степень устойчивости к последующим повреждениям низкими температурами. Морозоустойчивость же формируется в соответствии с генотипом в процессе онтогенеза под влиянием определенных условий внешней среды и связана с наступлением периода покоя, его глубиной и длительностью.
Агротехника конкретного вида растений (срок и способ посева и др.) должна максимально способствовать формированию в процессе закалки реализации возможной генетически детерминированной морозоустойчивости сорта. В северных и центральных районах России с неустойчивой весной и частым возвращением весенних заморозков более устойчивы и урожайны сорта озимых хлебов и плодовых многолетних культур с более глубоким зимним покоем, с поздним сроком возобновления весенней вегетации (***). Наоборот, в районах с устойчивым нарастанием положительных температур весной преимущество имеют рано вегетирующие виды и сорта растений.
Морозоустойчивость сортов озимой пшеницы определяется не только количеством сахаров, накопленных с осени, но и их экономным расходованием в течение зимы. У растений зимостойких сортов озимой пшеницы в зимний период с понижением температуры содержание моносахаридов (глюкоза, фруктоза) увеличивается за счет расщепления сахарозы на глюкозу и фруктозу, что снижает точку замерзания клеточного сока. Узел кущения злаков, корневая шейка бобовых - своеобразная кладовая энергетических ресурсов растения в зимний период и орган побегообразования весной.
Морозоустойчивость растений озимой пшеницы положительно коррелирует с содержанием сахаров в узлах кущения. В хороших посевах озимой пшеницы в листьях в декабре содержание растворимых углеводов достигает 18-24 % (на сухое вещество), а в узлах кущения - 39-42 %. В опытах более морозоустойчивый сорт озимой пшеницы Мироновская 808 расходовал за зиму всего 10 % углеводов, а менее устойчивый сорт Безостая 1-23 % углеводов. Растения, закладывающие узлы кущения глубоко (3-4 см), как правило, более морозоустойчивы, чем те, у которых узел кущения находится близко к поверхности (1-2 см). Глубина залегания узла кущения и мощность его развития зависят от качества семян, способа посева, обработки почвы.
На морозоустойчивость существенное влияние оказывают условия почвенного питания, особенно в осенний период. Устойчивость растений к морозу возрастает на постоянно известкуемых почвах при внесении под посев озимых калийно-фосфорных одобрений, тогда как избыточные азотные удобрения, способствуя процессам роста, делают растения озимых более чувствительными к морозам. На морозоустойчивость, как и на холодостойкость растений, положительное влияние оказывают микроэлементы (кобальт, цинк, молибден, медь, ванадий и др.)Например, цинк повышает содержание связанной воды, усиливает накопление сахаров, молибден способствует увеличению содержания общего и белкового азота.
Методы изучения морозоустойчивости растений. И. И. Тумановым с сотрудниками предложены лабораторные методы ускоренного определения морозоустойчивости различных культурных растений. Испытуемые растения после закаливания подвергают воздействию критических низких температур в холодильных камерах, что дает возможность выявить невымерзающие растения. Такая ускоренная оценка морозоустойчивости имеет большое преимущество перед обычным полевым способом оценки, так как последний требует много времени (иногда нескольких лет).
Другие надежные и удобные в исполнении лабораторные методы определения морозоустойчивости основаны на измерении вязкости цитоплазмы в клетках тканей исследуемых органов, определении электропроводности и др. Определение морозоустойчивости культурных растений мирового ассортимента показало, что страны СНГ обладают самыми устойчивыми их формами. Наиболее устойчивые сорта озимой пшеницы выведены опытными учреждениями юго-востока и северо-востока России, где природная обстановка благоприятствует выведению морозоустойчивых форм.
studfiles.net
Морозоустойчивость растений
Морозоустойчивость - способность растений переносить температуру ниже 0 °С, низкие отрицательные температуры. Морозоустойчивые растения способны предотвращать или уменьшать действие низких отрицательных температур. Морозы в зимний период с температурой ниже -20 °С обычны для значительной части территории России. Воздействию морозов подвергаются однолетние, двулетние и многолетние растения. Растения переносят условия зимы в различные периоды онтогенеза. У однолетних культур зимуют семена (яровые растения), раскустившиеся растения (озимые), у двулетних и многолетних - клубни, корнеплоды, луковицы, корневища, взрослые растения. Способность озимых, многолетних травянистых и древесных плодовых культур перезимовывать обусловливается их достаточно высокой морозоустойчивостью. Ткани этих растений могут замерзать, однако растения не погибают. Большой вклад в изучение физиологических основ морозоустойчивости внесли Н. А. Максимов (1952), Г. А. Самыгин (1974), И. И. Туманов (1979) и другие отечественные исследователи.
 Замерзание растительных клеток и тканей и происходящие при этом процессы. Способность растений переносить отрицательные температуры определяется наследственной основой данного вида растений, однако морозоустойчивость одного и того же растения зависит от условий, предшествующих наступлению морозов, влияющих на характер льдообразования. Лед может образовываться как в протопласте клетки, так и в межклеточном пространстве. Не всякое образование льда приводит клетки растения к гибели.
Замерзание растительных клеток и тканей и происходящие при этом процессы. Способность растений переносить отрицательные температуры определяется наследственной основой данного вида растений, однако морозоустойчивость одного и того же растения зависит от условий, предшествующих наступлению морозов, влияющих на характер льдообразования. Лед может образовываться как в протопласте клетки, так и в межклеточном пространстве. Не всякое образование льда приводит клетки растения к гибели. Постепенное снижение температуры со скоростью 0,5-1 °С/ч приводит к образованию кристаллов льда прежде всего в межклеточниках и первоначально не вызывают гибели клеток. Однако последствия этого процесса могут быть губительными для клетки. Образование льда в протопласте клетки, как правило, происходит при быстром понижении температуры. Происходит коагуляция белков протоплазмы, кристаллами образовавшегося в цитозоле льда повреждаются клеточные структуры, клетки погибают. Убитые морозом растения после оттаивания теряют тургор, из их мясистых тканей вытекает вода. Условия и причины вымерзания растений.

Образующийся при медленном промерзании в межклеточниках и клеточных стенках лед оттягивает воду из клеток; клеточный сок становится концентрированным, изменяется рН среды. Выкристаллизовавшийся лед действует как сухой воздух, иссушая клетки и сильно изменяя их осмотические свойства. Кроме того, цитоплазма подвергается сжатию кристаллами льда. Образующиеся кристаллы льда вытесняют воздух из межклеточников, поэтому замерзшие листья становятся прозрачными. Если льда образуется немного и клетки не были механически повреждены его кристаллами, то при последующем оттаивании такие растения могут сохранить жизнеспособность. Так, в листьях капусты при температуре -5...-6 °С образуется некоторое количество льда в межклеточниках. Однако при последующем медленном оттаивании межклеточники заполняются водой, которая поглощается клетками, и листья возвращаются в нормальное состояние.
Однако клетки, ткани и растения могут погибнуть от мороза. Основными причинами гибели клеток растений при низких отрицательных температурах и льдообразовании являются чрезмерное обезвоживание клеток или механическое давление, сжатие клеток кристаллами льда, повреждающее тонкие клеточные структуры. Оба эти фактора могут действовать одновременно. Летальность действия мороза определяется несколькими обстоятельствами. Последствия воздействия низких отрицательных температур зависят от оводненности тканей растения. Насыщенные водой ткани легко повреждаются, сухие же семена могут выносить глубокие низкие температуры (до -196 °С). Низкое содержание воды предохраняет от образования льда в растениях при промораживании. Разные растения, их клетки имеют свой критический предел обезвоживания и сжатия, превышение которого, а не только снижение температуры приводит к их гибели. Гибель клеток, тканей и растений под действием морозов обусловливается необратимыми изменениями, происходящими в протопласте клеток: его коагуляцией, денатурацией коллоидов протопласта, механическим давлением льда, повреждающим поверхностные структуры цитоплазмы, кристаллами льда, нарушающими мембраны и проникающими внутрь клетки. Вредное влияние оказывает повышение концентрации и изменение рН клеточного сока, сопровождающие обезвоживание клеток.
Действие льда, особенно при длительном влиянии низких температур, сходно с обезвоживанием клеток при засухе. Признаками повреждения клеток морозом являются потеря ими тур-гора, инфильтрация межклеточников водой и выход ионов из клеток. Выход ионов К+ и Сахаров из клеток, по-видимому, связан с повреждением мембранных систем их активного транспорта. Поврежденные растения при переносе в теплое место имеют вид ошпаренных, утрачивают тургор, листья быстро буреют и засыхают. При оттаивании мороженых клубней картофеля, корнеплодов кормовой и сахарной свеклы вода легко вытекает из тканей. Важно отметить, что состояние переохлаждения (без образования льда) растения переносят без вреда; при тех же температурах, но с образованием льда в тканях растения гибнут.
Растения по-разному реагируют на образование льда в тканях: клубни картофеля и георгина быстро погибают, капуста и лук переносят лишь умеренное промораживание, рожь и пшеница выдерживают на уровне узла кущения морозы до -15...-20 °С. У устойчивых к морозу растений имеются защитные механизмы, в основе которых лежат определенные физико-химические изменения. Морозоустойчивые растения обладают приспособлениями, уменьшающими обезвоживание клеток. При понижении температуры у таких растений отмечаются повышение содержания Сахаров и других веществ, защищающих ткани (криопротекторы), это прежде всего гидрофильные белки, моно- и олигосахариды; снижение оводненности клеток; увеличение количества полярных липидов и снижение насыщенности их жирнокислотных остатков; увеличение количества защитных белков.
На степень морозоустойчивости растений большое влияние оказывают сахара, регуляторы роста и другие вещества, образующиеся в клетках. В зимующих растениях в цитоплазме накапливаются сахара, а содержание крахмала снижается. Влияние сахаров на повышение морозоустойчивости растений многосторонне. Накопление Сахаров предохраняет от замерзания большой объем внутриклеточной воды, заметно уменьшает количество образующегося льда.
Сахара защищают белковые соединения от коагуляции при вымораживании; они образуют гидрофильные связи с белками цитоплазмы, предохраняя их от возможной денатурации, повышают осмотическое давление и снижают температуру замерзания цитозоля. В результате накопления Сахаров содержание прочносвязанной воды увеличивается, а свободной уменьшается. Особое значение имеет защитное влияние Сахаров на белки, сосредоточенные в поверхностных мембранах клетки. Сахара увеличивают водоудерживающую способность коллоидов протоплазмы клеток; связанная с коллоидами вода в виде гидратных оболочек биополимеров при низких, температурах не замерзает и не транспортируется, оставаясь в клетке.
Криопротекторами являются также молекулы гемицеллюлоз (ксиланы, арабиноксиланы), выделяемые цитоплазмой в клеточную стенку, обволакивающие растущие кристаллы льда, что предотвращает образование крупных кристаллов, повреждающих клетку. Так клетки защищаются как от внутриклеточного льда, так и от чрезмерного обезвоживания. Значительное количество защитных белков и модификации молекул липидов увеличивают структурированность клеток. У большинства растений возрастает синтез водорастворимых белков. Белковые вещества, частично гидролизуясь, увеличивают содержание свободных аминокислот. В тканях морозоустойчивых растений в конце лета и осенью накапливаются в достаточном количестве запасные вещества (прежде всего сахара), которые используются весной при возобновлении роста, обеспечивая потребности растений в строительном материале и энергии. Необходимо также учитывать устойчивость растений к болезням, вероятность развития которых увеличивается при повреждении тканей морозом.
Морозоустойчивость - не постоянное свойство растений. Она зависит от физиологического состояния растений и условий внешней среды. Растения, выращенные при относительно низких положительных температурах, более устойчивы, чем выращенные при относительно высоких осенних температурах. Свойство морозоустойчивости формируется в процессе онтогенеза растения под влиянием определенных условий среды в соответствии с генотипом растения, связано с резким снижением темпов роста, переходом растения в состояние покоя.
Жизненный цикл развития озимых, двуручек, двулетних и многолетних растений контролируется сезонным ритмом светового и температурного периодов. В отличие от яровых однолетних растений они начинают готовиться к перенесению неблагоприятных зимних условий с момента остановки роста и затем в течение осени во время наступления пониженных температур. Повышение морозоустойчивости растений тесно связано с закаливанием - постепенной подготовкой растений к воздействию низких зимних температур. Закаливание - это обратимая физиологическая устойчивость к неблагоприятным воздействиям среды.
Способностью к закаливанию обладают не все растения. Растения южного происхождения не способны переносить морозы. Способность к закаливанию у древесных и зимующих травянистых растений северных широт, переживающих значительное понижение температуры в зимний период, в период летней вегетации отсутствует и проявляется только во время наступления осенних пониженных температур (если растение к этому времени прошло необходимый цикл развития). Процесс закалки приурочен лишь к определенным этапам развития растений. Для приобретения способности к закаливанию растения должны закончить процессы роста.
Разные органы растений имеют неодинаковую способность к закаливанию, например, листья листопадных деревьев (яблоня, груша, вишня) не обладают способностью к закаливанию; цветочные почки закаливаются хуже, чем листовые. У вегетирующих растений легко вымерзают растущие и не закончившие рост органы. Выносливость растений к низким температурам в этот период незначительная.
Эффект закаливания может не проявиться, если по каким-либо причинам (засуха, поздний посев, посадки и др.) произошла задержка развития растений. Так, если в течение лета у плодовых растений процессы роста из-за летней засухи не успели закончиться, то зимой это может привести к гибели растений. Дело в том, что засуха, приостанавливая рост летом, не позволяет растениям завершить его к осени. Одновременно при закалке должен произойти отток различных веществ из надземных органов в подземные зимующие (корневые системы, корневища, луковицы, клубни). По этой же причине закалку травянистых и древесных растений ухудшает избыточное азотное питание, удлиняющее период роста до поздней осени, в результате растения не способны пройти процессы закаливания и гибнут даже при небольших морозах.
Яровые злаки при озимом посеве по сравнению с озимыми растут при более пониженных положительных температурах, в осенний период почти не снижают темпов роста и не способны к закаливанию. Большую роль в закаливании играют условия внешней среды. Так, на озимых культурах убедительно показана необходимость света для процесса закаливания. Сокращение фотопериода служит для растений сигналом к прекращению роста и стимулом для накопления ингибиторов в растениях. Вероятно, с этих процессов начинается формирование морозоустойчивости у растений.
Растения, выращенные при несоответствующем фотопериоде, не успевают завершить летний рост и не способны к закаливанию. Установлено, что длинный день способствует образованию в листьях черной смородины фитогормонов стимуляторов роста, а короткий - накоплению ингибиторов. В естественных условиях к закаливанию способен лишь организм в целом, при обязательном наличии корневой системы. По-видимому, в корнях вырабатываются вещества, повышающие устойчивость растения к морозу. Процесс закаливания растений требует определенного комплекса внешних условий и проходит в две фазы, которым предшествуют замедление роста и переход растений в состояние покоя. Прекращение роста и переход в состояние покоя - необходимые условия прохождения первой фазы закаливания. Однако само по себе оно лишь немного повышает морозоустойчивость растения. У травянистых растений переход в состояние покоя происходит в период первой фазы закаливания. У древесных покой наступает в начале осени, до прохождения первой фазы закаливания.
При переходе в состояние покоя изменяется баланс фитогормонов: уменьшается содержание ауксина и гиббереллинов и увеличивается содержание абсцизовой кислоты, которая, ослабляя и ингибируя ростовые процессы, обусловливает наступление периода покоя. Поэтому обработка растений озимой пшеницы, люцерны и других культур в этот период ингибиторами роста (например, хлор-холинхлоридом - ССС или трииодбензойной кислотой) повышает устойчивость растений к низким температурам.
Первая фаза закаливания проходит на свету и при низких положительных температурах в ночное время (днем около 10 °С, ночью около 2 °С), останавливающих рост, и умеренной влажности почвы. Озимые злаки проходят первую фазу на свету при среднесуточной температуре 0,5-2 °С за 6-9 дней, древесные - за 30 дней. В эту фазу продолжается дальнейшее замедление и даже происходит полная остановка ростовых процессов.
Свет в этой фазе необходим не только для фотосинтеза, но и для поддержания ультраструктур клетки. В таких условиях за счет фотосинтеза образуются сахара, а понижение температуры в ночное время значительно снижает их расход на дыхание и процессы роста. В результате в клетках растений накапливаются сахароза, другие олигосахариды, растворимые белки и т. д., в мембранах возрастает содержание ненасыщенных жирных кислот, снижается точка замерзания цитоплазмы, отмечается некоторое уменьшение внутриклеточной воды.
Благоприятные условия для прохождения первой фазы закаливания озимых растений складываются при солнечной и прохладной (дневная температура до 10 °С) погоде, способствующей накоплению в тканях растений углеводов и других защитных веществ. В естественных условиях оптимальный срок первой фазы закаливания озимых злаков до двух недель. За это время количество сахаров в растениях возрастает до 70 % на сухую массу или до 22 % на сырую массу, т. е. близко содержанию Сахаров в корнеплодах лучших сортов сахарной свеклы.
Растения озимой пшеницы можно закалить и в темноте при 2 °С, если их корни или узлы кущения погрузить в раствор сахарозы. Такие растения выдерживают морозы до -20 °С. Накапливающиеся в процессе закаливания сахара локализуются в клеточном соке, цитоплазме, клеточных органеллах, особенно в хлоропластах. При закаливании растений высоко морозоустойчивого сорта озимой пшеницы при температуре, близкой к О °С, количество Сахаров в хлоропластах листьев увеличивалось в 2,5 раза, благодаря чему хлоропласты продолжали функционировать. Повышение содержания сахаров в хлоропластах коррелирует с морозоустойчивостью растений.
В хлоропластах содержатся те же формы сахаров, что и в листьях: фруктоза, глюкоза, сахароза, олигосахар. Имеются данные, что при накоплении сахаров процесс фотофосфорилирования продолжается даже при отрицательных температурах. Более морозоустойчивые виды и сорта растений лучше накапливают сахар именно при сочетании пониженной температуры и умеренной влажности почвы. Дело в том, что в первой фазе закаливания происходит уменьшение содержания свободной воды, а излишняя влажность почвы при дождливой осени затрудняет этот процесс, повышается вероятность в последующем образования внутриклеточного льда и гибели растений.
Метаболические изменения, наблюдаемые во время первой фазы, могут быть вызваны изменением гормонального и энергетического балансов, что определяет синтез и активацию специфических ферментов, свойства клеточных мембран закаленных тканей. Накапливающаяся в тканях абсцизовая кислота увеличивает проницаемость мембран для воды, водоотдачу клеток. К концу первой фазы закаливания все зимующие растения переходят в состояние покоя. Однако процессы закалки, перестройки процессов обмена веществ продолжаются. Вторая фаза закаливания не требует света и начинается сразу же после первой фазы при температуре немного ниже 0°С. Для травянистых растений она может протекать и под снегом. Длится она около двух недель при постепенном снижении температуры до -10...-20 °С и ниже со скоростью 2-3 °С в сутки, что приводит к частичной потере воды клетками, освобождению клеток тканей от избыточного содержания воды или витрификации (переходу воды в стеклообразное состояние). Явление витрификации воды в растительных клетках наступает при резком охлаждении (ниже -20 °С). Стеклообразная растительная ткань долго сохраняет свою жизнеспособность. При постепенном понижении температуры в межклеточниках образуется лед и начинают функционировать механизмы, предохраняющие подготовленные в первой фазе закаливания растения от чрезмерного обезвоживания. Накопившиеся в первой фазе закаливания сахара изменяют устойчивость биоколлоидов цитоплазмы к низким температурам, возрастает относительное количество коллоидно-связанной воды.
Вторая фаза обеспечивает отток из цитозоля клеток почти всей воды, которая может замерзнуть при отрицательной температуре. При критических температурах отток воды из клеток значительно ухудшается, появляется много переохлажденной воды, которая затем замерзает внутри протопласта и может привести к гибели клеток. Следовательно, чем менее морозоустойчиво растение, тем медленнее должна протекать вторая фаза закаливания.
Действующими факторами второй фазы закаливания являются обезвоживание, вызывающее сближение молекул в цитозоле, вязкость которого соответственно увеличивается; низкая температура, уменьшающая тепловое движение молекул в протопласте. В результате во второй фазе закаливания происходит перестройка белков цитоплазмы, накапливаются низкомолекулярные водорастворимые белки, более устойчивые к обезвоживанию, синтезируются специфические белки. Содержание незамерзающей (связанной) воды в тканях зимостойкой пшеницы почти в 3 раза выше по сравнению с незимостойкой.
Перестройка цитоплазмы увеличивает проницаемость ее для воды, способствует более быстрому оттоку воды в межклеточники, что снижает опасность внутриклеточного льдообразования. При обезвоживании, происходящем под влиянием льдообразования, наблюдаются сближение и деформация белковых молекул, связи между которыми могут рваться и не восстанавливаются, что пагубно для клетки. Очевидно, при таких условиях происходит быстрое смещение структурных частиц по отношению друг к другу, что приводит к разрушению субмикроскопической структуры протопласта.
Цитоплазма закаленных растений более устойчива к механическому давлению. Поэтому важно наличие у молекул белков сульфгидрильных и других гидрофильных группировок, которые способствуют удержанию воды, препятствуют слишком сильному сближению молекул белка. Между содержанием сульфгидрильных групп и морозоустойчивостью клеток растений установлена положительная связь. Благодаря изменению свойств молекул белков и межмолекулярных связей в процессе закаливания постепенное обезвоживание приводит к переходу цитоплазмы из состояния золя в гель.
Первая фаза закаливания повышает морозоустойчивость растений с -5 до -12 °С, вторая увеличивает морозоустойчивость, например, у пшеницы до -18...-20 °С, у ржи - до -20...-25 °С. Растения, находящиеся в глубоком органическом покое, отличаются способностью к закаливанию и выдерживают промораживание до -195 °С. Так, черная смородина после наступления состояния глубокого покоя и завершения первой фазы закаливания переносила охлаждение до -253 °С.
Не у всех растений процесс закаливания проходит в две фазы. У древесных растений, имеющих в тканях достаточное количество Сахаров, сразу же протекают процессы, свойственные второй фазе закаливания. Однако не все растения способны к закаливанию. Теплолюбивые растения (хлопчатник, рис, бахчевые культуры) при длительном пребывании при температурах немного выше О °С не только не становятся устойчивыми, но еще сильнее повреждаются или даже погибают, так как в них накапливаются ядовитые вещества, усиливающие губительное действие на растения низких температур. В период прохождения фаз закаливания формируется морозоустойчивость растений. Морозоустойчивость представляет собой процесс, а не постоянное свойство растений. Процесс закаливания обратим, при этом морозоустойчивость растений снижается. Развитие процесса закаливания в значительной степени зависит от условий его протекания. Особенно заметное влияние на морозоустойчивость оказывают условия закаливания растений в осенний период, определяемые в первую очередь соотношением числа ясных дней с пониженными положительными температурами ночью и числа пасмурных, дождливых дней с относительно сближенными высокими температурами днем и ночью. Чем это отношение выше, тем лучше условия для закалки.
Критические температуры повреждения растений озимой пшеницы при разных условиях закалки
У хорошо закаленных растений благодаря высокой концентрации клеточного сока, пониженному содержанию воды образуется значительно меньше кристаллов льда, причем не в клетке, а в межклеточниках. Такие растения погибают только при очень сильных морозах. При закаливании происходят обратимые физиологические изменения. При неустойчивой осенней и зимней погоде приобретенная в процессе закалки морозоустойчивость снижается. Наблюдается тесная связь между морозоустойчивостью растений и ростовыми процессами. Переход к состоянию покоя всегда сопровождается повышением устойчивости, а от состояния покоя к росту - снижением. В связи с этим морозоустойчивость одного и того же вида растений довольно сильно меняется в течение года: летом она минимальная (растения могут погибнуть при температурах намного выше тех, которые они выдерживают зимой), осенью увеличивается, а в конце зимы и в начале весны опять снижается. Повышение температуры весной сопровождается противоположными закаливанию физиологобиохимическими. изменениями - происходит процесс закаливания растений. Весной растения часто гибнут даже от небольших заморозков.
Основа решения этой задачи - селекция морозоустойчивых сортов растений, хорошо адаптирующихся к климатическим условиям данного региона. Следует еще раз отметить, что процесс закаливания представляет собой временную адаптацию цитоплазмы, определяющую степень устойчивости к последующим повреждениям низкими температурами. Морозоустойчивость же формируется в соответствии с генотипом в процессе онтогенеза под влиянием определенных условий внешней среды и связана с наступлением периода покоя, его глубиной и длительностью.
biofile.ru
Влияние холода на растения и приспособления к нему
О пределе холода, который способны вынести растения в естественных условиях, дают представление величины предельно низких температур на земном шаре. Там, где зарегистрирована самая низкая температура (—90°С, станция «Восток» в Антарктиде), растительность отсутствует; а в районах, где живут растения, отмечена температура —68°С (Оймякон в Якутии, область таежных лесов из лиственницы — Larix dahurica).
Растительный покров обширных территорий земного шара (умеренные и арктические области, высокогорья) ежегодно в течение нескольких месяцев подвергается действию низких температур. Кроме того, в отдельных районах и в более теплые сезоны растения могут испытывать кратковременные воздействия пониженных температур (ночные и утренние заморозки). Наконец, есть местообитания, где вся жизнь растений проходит на весьма пониженном температурном фоне (арктические снежные и морские водоросли, приснежная—нивальная растительность в высокогорьях). Неудивительно, что естественный отбор выработал у растений ряд защитных приспособлений к неблагоприятному действию холода.
Кроме непосредственного влияния низкой температуры на растения под действием холода возникают и другие неблагоприятные явления. Например, уплотнение и растрескивание замерзшей почвы приводит к разрыву и механическому повреждению корней, образование ледяной корки на поверхности почвы ухудшает аэрацию и дыхание корней. Под толстым и долго лежащим снежным покровом при температуре около 0°С наблюдается зимнее «выпревание», истощение и гибель растений в связи с расходом резервных веществ на дыхание, грибными заболеваниями («снежная плесень») и т. д., а в случае избыточно увлажненной почвы для растений опасно также зимнее «вымокание». В тундре и северной тайге распространено явление морозного «выпирания» растений, которое вызывается неравномерным замерзанием и расширением почвенной влаги. При этом возникают силы, выталкивающие растение из почвы, в результате чего происходит «выпучивание» целых дернин, оголение и обрывы корней и т. д. вплоть до повала небольших деревьев. Поэтому кроме собственно холодостойкости (или морозостойкости) — способности переносить прямое действие низких температур, различают еще зимостойкость растений - способность к перенесению всех перечисленных выше неблагоприятных зимних условий.
Особо следует остановиться на том, как влияет на растения низкая температура почвы. Холодные почвы в сочетании с умеренно-теплым режимом воздушной среды растений (а иногда и со значительным нагреванием надземных частей растений) — явление нередкое. Таковы условия жизни растений на болотах и заболоченных лугах с тяжелыми почвами, в некоторых тундровых и. высокогорных местообитаниях и в обширных областях вечной мерзлоты (около 20% всей суши), где в период вегетации оттаивает лишь неглубокий, так называемый «деятельный» слой почвы. В условиях пониженных температур почвы после снеготаяния (0—10°С) проходит значительная часть вегетации ранневесенних лесных растений — «подснежников». Наконец, кратковременные периоды резкого несоответствия холодных почв и прогретого воздуха испытывают ранней весной многие растения умеренного климата (в том числе и древесные породы).
Еще в прошлом веке немецкий физиолог Ю. Сакс показал, что при охлаждении почвы до околонулевых температур (обкладывание горшка льдом) может наступить завядание даже обильно политых растений, поскольку при низких температурах корни не способны интенсивно всасывать воду. На этом основании в экологии распространилось мнение о «физиологической сухости» местообитаний с холодными почвами (т. е. недоступности влаги растениям при ее физическом обилии). При этом упускали из виду, что Сакс и другие физиологи свои опыты производили с достаточно теплолюбивыми растениями (огурцы, тыква, салат и др.) и что в природных холодных местообитаниях растения, для которых низкие температуры почв служат естественным фоном, возможно, реагируют на них совсем иначе. Действительно, современные исследования показали, что у большинства растений тундр, болот, у ранневесенних лесных эфемероидов отсутствуют те явления угнетения (затруднение всасывания воды, расстройства водного режима и т. д.), которые могли бы быть вызваны «физиологической сухостью» холодных почв. Это же показано и для многих растений в областях вечной мерзлоты. Вместе с тем нельзя полностью отрицать угнетающее влияние низких температур на всасывание влаги и другие стороны жизнедеятельности корней (дыхание, рост и др.), а также на активность почвенной микрофлоры. Оно без сомнения имеет значение в комплексе трудных условий для жизни растений в холодных местообитаниях. «Физиологическая сухость», «физиологическая засуха» из-за низкой температуры почв возможны в жизни растений в наиболее трудных условиях, например при выращивании на холодных почвах теплолюбивых растений или ранней весной для древесных пород, когда еще необлиственные ветви сильно нагреваются (до 30—35°С) и увеличивают потерю влаги, а интенсивная работа корневых систем еще не началась.
Каких-либо специальных морфологических приспособлений, защищающих от холода, у растений нет, скорее можно говорить о защите от всего комплекса неблагоприятных условий в холодных местообитаниях, включающего сильные ветры, возможность иссушения и т. д. У растений холодных областей (или у переносящих холодные зимы) часто встречаются такие защитные морфологические особенности, как опушение почечных чешуи, зимнее засмоление почек (у хвойных), утолщенный пробковый слой, толстая кутикула, опушение листьев и т. д. Однако их защитное действие имело бы смысл лишь для сохранения собственного тепла гомеотермных организмов, для растений же эти черты, хотя и способствуют терморегуляции (уменьшение лучеиспускания), в основном важны как защита от иссушения. В растительном мире есть интересные примеры адаптации, направленных на сохранение (хотя и кратковременное) тепла в отдельных частях растения. В высокогорьях Восточной Африки и Южной Америки у гигантских «розеточных» деревьев из родов Senecio, Lobelia, Espeletia и других от частых ночных морозов существует такая защита: ночью листья розетки закрываются, защищая наиболее уязвимые части — растущие верхушки. У некоторых видов листья опушены снаружи, у других в розетке скапливается выделяемая растением вода; ночью замерзает лишь поверхностный слой, а конусы нарастания оказываются защищенными от мороза в своеобразной «ванне».
Среди морфологических адаптации растений к жизни в холодных местообитаниях важное значение имеют небольшие размеры и особые формы роста. Не только многие травянистые многолетники, но также кустарники и кустарнички полярных и высокогорных областей имеют высоту не более нескольких сантиметров, сильно сближенные междоузлия, очень мелкие листья (явление нанизма или карликовости). Кроме хорошо известного примера — карликовой березки ( Betula папа), можно назвать карликовые ивы ( Sahx polaris, S. arctica, S. herbacea) и многие другие. Обычно высота этих растений соответствует глубине снежного покрова, под которым зимуют растения, так как все части, выступающие над снегом, гибнут от замерзания и высыхания. Очевидно, в образовании карликовых форм в холодных местообитаниях немалую роль играют и бедность почвенного питания в результате подавления активности микробов, и торможение фотосинтеза низкими температурами. Но независимо от способа образования карликовые формы дают известное преимущество растениям в приспособлении к низким температурам: они располагаются в припочвенной экологической микронише, наиболее прогреваемой летом, а зимой хорошо защищены снежным покровом и получают дополнительный (хотя и небольшой) приток тепла из глубины почвы.
Другая адаптивная особенность формы роста — переход сравнительно крупных растений (кустарников и даже деревьев) от ортотропного (вертикального) к плагиотропному (горизонтальному) росту и образование стелющихся форм— стланцев, стлаников, стланичков. Такие формы способны образовывать кедровый стланик ( Pinus pumila), можжевельник ( Juniperus sibirica, J. communis, J. turkestanica), рябина и др. Ветви стланцев распластаны по земле и приподнимаются не выше обычной глубины снежного покрова. Иногда это результат отмирания ствола и разрастания нижних ветвей (например, у ели), иногда это рост дерева как бы «лежа на боку» с плагиотропным, укоренившимся во многих местах стволом и приподнимающимися ветвями (кедровый стланик). Интересная особенность некоторых древесных и кустарниковых стлаников — постоянное отмирание старой части ствола и нарастание «верхушки», в результате чего трудно определить возраст особи.
Стланики распространены в высокогорных и полярных областях, в условиях, которых уже не выдерживают древесные породы (например, на верхней границе леса). Своеобразные «стланиковые» формы в крайних условиях встречаются и у кустарничков, и даже у видов лишайников, обычно имеющих прямостоячий кустистый рост: на скалах Антарктиды они образуют стелющиеся слоевища,
В зависимости от условий возможны видоизменения роста одного и того же вида. Но есть виды, целиком перешедшие к форме стланика, например горный сосновый стланик, произрастающий в Альпах и Карпатах — Pinus mughus, выделенный в качестве самостоятельного вида из сосны горной — Pinus montana.
К числу форм роста, способствующих выживанию растений в холодных местообитаниях, принадлежит еще одна чрезвычайно своеобразная — подушковидная. Форма растения-подушки образуется в результате усиленного ветвления и крайне замедленного роста скелетных осей и побегов. Мелкие ксерофильные листья и цветки расположены по периферии подушки. Между отдельными ветвями скапливаются мелкозем, пыль, мелкие камни. В результате некоторые виды растений-подушек приобретают большую компактность и необычайную плотность: по таким растениям можно ходить, как по твердой почве. Таковы Silene acaulis. Gypsophila aretioides, Androsace helvetica, Acantholimon diapensioides. Издали их трудно отличить от валунов. Менее плотны колючие подушки из родов Eurotia, Saxifraga.
Растения-подушки бывают разных размеров (до 1 м в поперечнике) и разнообразных очертаний: полушаровидные, плоские, вогнутые, иногда довольно причудливых форм (в Австралии и Новой Зеландии их называют «растительными овцами»).
Благодаря компактной структуре растения-подушки успешно противостоят холодным ветрам. Поверхность их нагревается почти так же, как и поверхность почвы, а колебания температуры внутри менее выражены, чем в окружающей среде. Отмечены случаи значительного повышения температуры внутри подушки; например, у наиболее распространенного вида высокогорий Центрального Тянь-Шаня Dryadanthe tetrandra при температуре воздуха 10°С внутри подушки температура доходила до 23°С благодаря аккумуляции тепла в этом своеобразном «парнике». В связи с медленным ростом растения-подушки по долговечности вполне сравнимы с деревьями. Так, на Памире подушка Acantholimon hedini диаметром 3 см имела возраст 10—12 лет, при 10 см — 30—35 лет, а возраст крупных подушек достигал не одной сотни лет.
В пределах общей формы растений-подушек существует экологическое разнообразие: например в горах, окружающих Средиземное море, распространены менее компактные по строению ксерофильные «колючие подушки», которые не встречаются высоко в горах, так как малоустойчивы к холоду, но зато очень устойчивы к засухе. Рыхлое строение подушки здесь оказывается более выгодным для растения, чем компактное, так как в условиях летней засухи и сильной инсоляции снижает опасность перегрева ее поверхности. Температура поверхности средиземноморских подушек обычно ниже температуры воздуха благодаря сильной транспирации, а внутри подушки создается особый микроклимат; например, влажность воздуха держится на уровне 70—80% при влажности наружного воздуха 30%. Таким образом, здесь форма подушки — это приспособление к совсем иному комплексу факторов, отсюда и ее иная «конструкция».
Среди других особенностей роста, помогающих растениям преодолевать действие холода, следует еще упомянуть различные приспособления, направленные на углубление зимующих частей растений в почву. Это развитие контрактильных (сократительных) корней — толстых и мясистых, с сильноразвитой механической тканью. Осенью они высыхают и сильно сокращаются в длину (что хорошо заметно по поперечной морщинистости), при этом возникают силы, втягивающие в почву зимующие почки возобновления, луковицы, корни, корневища.
Контрактильные корни встречаются у многих растений высокогорий, тундр и других холодных местообитаний. Они позволяют, в частности, успешно противостоять морозному выпиранию растений из почвы. В последнем случае они не только втягивают почку возобновления, но и ориентируют ее перпендикулярно поверхности, если растение повалено. Глубина втягивания контрактильными корнями варьирует от сантиметра до нескольких десятков сантиметров в зависимости от особенностей растения и механического состава почвы.
Адаптивное изменение формы как защита от холода — явление, ограниченное в основном холодными районами. Между тем действие холода испытывают и растения более умеренных областей. Гораздо более универсальны физиологические способы защиты. Они направлены прежде всего на снижение точки замерзания клеточного сока, предохранение воды от вымерзания и т. д. Отсюда такие особенности холодостойких растений, как повышение концентрации клеточного сока, главным образом за счет растворимых углеводов. Известно, что при осеннем повышении холодостойкости («закаливании») крахмал превращается в растворимые сахара. Другая черта холодостойких растений — повышение доли коллоидно-связанной воды в общем водном запасе.
При медленном снижении температуры растения могут вынести охлаждение ниже точки замерзания клеточного сока в состоянии переохлаждения (без образования льда). Как показывают эксперименты, уровень точек переохлаждения и замерзания тесно связан с температурными условиями обитания. Однако у растений состояние переохлаждения возможно лишь при небольшом холоде (несколько градусов ниже нуля). Гораздо более действенным этот путь адаптации оказывается у других пойкилотермных организмов— насекомых, у которых роль антифризов играют глицерин, трегалоза и другие защитные вещества (открыто зимующие насекомые могут вынести переохлаждение клеточных соков без замерзания до — 30°С).
Многие растения способны сохранять жизнеспособность и в промерзшем состоянии. Есть виды, замерзающие осенью в фазе цветения и продолжающие цвести после оттаивания весной (мокрица — Stellaria media, маргаритка— Bellis perennis, арктический хрен — Cochlearia fenestrata и др.). Ранневесенние лесные эфемероиды («подснежники») в течение короткой вегетации неоднократно переносят весенние ночные заморозки: цветки и листья промерзают до стекловидно-хрупкого состояния и покрываются инеем, но уже через 2—3 ч после восхода солнца оттаивают и возвращаются в обычное состояние. Хорошо известна способность мхов и лишайников переносить длительное промерзание зимой в состоянии анабиоза. В одном из опытов лишайник Cladonia замораживали при —15°С на 110 недель (более двух лет!).
После оттаивания лишайник оказался живым и вполне жизнеспособным, у него возобновились фотосинтез и рост. Очевидно, у лишайников в крайне холодных условиях существования периоды такого анабиоза очень длительны, а рост и активная жизнедеятельность осуществляются лишь в короткие благоприятные периоды (причем не каждый год). Такое частое прерывание активной жизни на долгие сроки, по-видимому, объясняет колоссальный возраст многих лишайников, определенный радиоуглеродным методом (до 1300 лет у Rhizocaгрon geographicum и Альпах, до 4500 лет у лишайников в Западной Гренландии).
Анабиоз — «крайняя мера» в борьбе растения с холодом, приводящая к приостановке жизненных процессов и резкому снижению продуктивности. Гораздо большее значение в адаптации растений к холоду имеет возможность сохранения нормальной жизнедеятельности путем снижения температурных оптимумов физиологических процессов и нижних температурных границ, при которых эти процессы возможны. Как видно на примере оптимальных температур для фотосинтеза и его нижних температурных порогов, эти явления хорошо выражены у растений холодных местообитаний. Так, у альпийских и антарктических лишайников для фотосинтеза оптимальна температура около 5°С; заметный фотосинтез удается обнаружить у них даже при —10°С. При сравнительно низких температурах лежит оптимум фотосинтеза у арктических растений, высокогорных видов, ранневесенних эфемероидов. Зимой при отрицательных температурах способны к фотосинтезу многие хвойные древесные породы. У одного и того же вида температурные оптимумы фотосинтеза связаны с изменением условий: так, у альпийских и арктических популяций травянистых многолетников — Оху ria digyna, Thalictrum alpinum и других видов они более низкие, чем у равнинных. Показательно в этом отношении и сезонное смещение оптимума по мере повышения температуры от весны к лету и снижения от лета к осени и зиме.
При низких температурах для растений чрезвычайно важно сохранить достаточный уровень дыхания — энергетической основы роста и репарации возможных повреждений холодом. На примере ряда растений памирских высокогорий показано, что в этих условиях довольно интенсивное дыхание сохраняется после действия температуры от —6 до —10°С.
Еще один пример устойчивости физиологических процессов к холоду— зимний и предвесенний подснежный рост у растений тундр, высокогорий и других холодных местообитаний с коротким вегетационным периодом, обусловленным заблаговременной подготовкой. Это явление чрезвычайно ярко выражено у эфемероидов лесостепных дубовых лесов (пролески — Scilla sibirica, хохлатки — Corydalis halleri, гусиного лука — Gagea lutea, чистяка — Ficariaverna и др.), у которых уже в начале зимы начинается рост побегов со сформированными внутри бутонами (вначале в промерзшей почве, а затем над почвой, внутри снежного покрова. Не прекращается у них зимой и формирование генеративных органов. По мере приближения сроков снеготаяния скорость подснежного роста заметно возрастает. В пору раннего «предвесенья», когда лес кажется еще совсем безжизненным, под снеговым покровом над почвой уже возвышаются тысячи ростков пролески и гусиного лука, достигающих к этому времени 2—7 см высоты и готовых начать цветение, как только сойдет снег. Образование хлорофилла у ранневесенних эфемероидов также начинается при низких температурах порядка 0°С, еще под снегом.
Экологические различия холодостойкости растений
В экологии и экологической физиологии в качестве одного из показателей устойчивости к холоду используется способность растения переносить низкую температуру в экспериментальных условиях в течение определенного срока. Накоплено много данных, позволяющих сравнивать растения различных по температурным условиям местообитаний. Однако эти данные не всегда строго сравнимы, поскольку температура, которую способно вынести растение, в числе прочих причин зависит и от продолжительности ее действия (так, небольшой холод порядка —3—5°С умеренно теплолюбивое растение способно вынести в течение нескольких часов, но та же температура может оказаться губительной, если будет действовать несколько суток), В большинстве экспериментальных работ принято охлаждение растений в течение суток или близкого срока.
Как видно из нижеследующих данных и, холодостойкость растений весьма различна и зависит от условий, в которых они обитают.
Один из крайних примеров холодостойкости — так называемый «криопланктон». Это снежные водоросли, живущие в поверхностных слоях снега и льда и при массовом размножении вызывающие его окрашивание («красный снег», «зеленый снег» и т. д.). В активных фазах они развиваются при 0°С (летом на оттаявшей поверхности снега и льда). Пределы устойчивости к низким температурам от —36°С у Chlamydomonas nivalis до —40, —60°С у Pediastrutn boryanum, Hormidium flaccidum. Столь же велика холодостойкость фитопланктона полярных морей, нередко зимующего в корке льда.
Большой холодостойкостью отличаются альпийские карликовые кустарнички — Rhododendron ferrugineum, Erica carnea и др. (—28, —36°С), хвойные древесные породы: так, для сосны Pinus strobus в Тирольских Альпах в экспериментах отмечена рекордная температура: —78°С.
Совсем небольшая холодостойкость у растений тропических и субтропических областей, где они не испытывают действия низких температур (за исключением высокогорий). Так, для водорослей тропических морей (особенно мелководных районов) нижняя температурная граница лежит в пределах 5—14°С (вспомним, что для водорослей арктических морей верхняя граница составляет 16°С). Саженцы тропических древесных пород гибнут при 3—5°С. У многих тропических термофильных растений, например декоративных оранжерейных видов из родов Gloxinia, Coleus, Achimenes и др., понижение температуры до нескольких градусов выше нуля вызывает явления «простуды»: при отсутствии видимых повреждений через некоторое время останавливается рост, опадают листья, растения завядают, а затем и гибнут. Известно это явление и для теплолюбивых культурных растений (огурцов, томатов, фасоли).
Очень невелика устойчивость к холоду у термофильных плесневых грибов из родов Mucor, Thermoascus, Anixia и др. Они гибнут за три дня при температуре 5—6°С и даже температуру 15—17°С не могут выносить дольше 15—20 дней.
В зависимости от степени и специфического характера холодостойкости можно выделить следующие группы растений.
Нехолодостойкие растения
К этой группе относятся все те растения, которые серьезно повреждаются уже при температурах выше точки замерзания: водоросли теплых морей, некоторые грибы и многие листостебельные растения тропических дождевых лесов.
Неморозостойкие растения
Эти растения хотя и переносят низкие температуры, но вымерзают, как только в тканях начинает образовываться лед. Неморозостойкие растения защищены от повреждения только средствами, замедляющими замерзание. В более холодное время года у них повышается концентрация осмотически активных веществ в клеточном соке и в протоплазме, а также переохлаждаемость, что предотвращает или замедляет образование льда при температурах примерно до —7°С, а при постоянном переохлаждении и до более низких температур. В период вегетации все листостебельные растения неморозостойки. В течение всего года чувствительны к образованию льда глубоководные водоросли, холодных морей и некоторые пресноводные водоросли, тропические и субтропические древесные растения и различные виды из умеренно-теплых районов.
Льдоустойчивые» растения
В холодное время года эти растения переносят внеклеточное замерзание воды и связанное с ним обезвоживание. Устойчивыми к образованию льда становятся некоторые пресноводные водоросли и водоросли приливной зоны, наземные водоросли, мхи всех климатических зон (даже тропической) и многолетние наземные растения областей с холодной зимой. Некоторые водоросли, многие лишайники и различные древесные растения способны чрезвычайно сильно закаливаться; тогда они остаются без повреждений и после продолжительных суровых морозов, и их можно охлаждать даже до температуры жидкого азота.
biofile.ru
Зимостойкость и морозостойкость садовых растений
Реклама
Тема зимней выносливости растений тем более не оставляет равнодушными специалисте – питомниководов, ландшафтных дизайнеров и профессиональных садовников.
Это обусловливается учащением аномальных климатических явлений: зимних оттепелей, ранних морозов при отсутствии снежного покрова, резких перепадов температуры с установлением ее новых максимумов и минимумов для данного региона, или из зарубежных питомников теплолюбивые виды, не переносящие наших зимних условий, а еще чаще – растения так называемого дополнительного ассортимента, объединяемые в своеобразную группу риска.
Однако Нечерноземье, несмотря на происходящие климатические изменения, остается зоной рискованного земледелия и садоводства.
Читайте также: Сорта морозостойких плодовых деревьев
ГРУППА РИСКА
Вызывает вопросы и оценка выживаемости зимой довольно многих растений, предлагаемых на рынке. Не секрет, что далеко не весь ассортимент отвечает требованиям зимостойкости, предъявляемым к посадочному материалу в наших климатических условиях.
Довольно часто в садовых центрах можно увидеть завозимые из южных регионов страны или из зарубежных питомников теплолюбивые виды, не переносящие наших зимних условий, а еще чаще -растения так называемого дополнительного ассортимента, объединяемые в своеобразную группу риска.
Однако Нечерноземье, несмотря на происходящие климатические изменения, остается зоной рискованного земледелия и садоводства. Не следует забывать и о таком явлении, как фотопериодизм растений.
Привезенные из теплых районов виды и сорта формируются в условиях короткого светового дня. При выращивании их в наших широтах с длинным днем они запаздывают в своем развитии и не заканчивают рост при наступлении холодных дней. В результате – не успевают подготовиться к зиме и поэтому чаще вымерзают. Проблемы зимовки у привезенных из других регионов растений могут быть и из-за несоответствия сроков наступления климатических сезонов.
В числе тех растений, о которых шла речь выше, можно назвать глицинии – китайскую и г. обильноцветущую, тую восточную и т. складчатую, падуб остролистный, фотинию Фразера, кииарисовик Лавсона, кампсис укореняющийся, самшит вечнозеленый, клены веерные, пиерис японский или аукубу японскую.
В условиях средней полосы России эти растения будут постоянно или периодически подмерзать зимой (или вымерзать выше уровня снега), и даже в самых благоприятных микрообитаниях они могут отличаться замедленным ростом, скудным цветением и не достигать максимальной декоративности, как например: гортензия крупнолистная, кольквиция прелестная, большинство завозимых магнолий. Некоторые из растений в большей степени страдают даже не от низких отрицательных температур, а от частого чередования морозов и оттепелей и других особенностей зимовки.
РАЙОНИРОВАНИЕ
Работы подревокультурному районированию европейской части России для задач декоративного садоводства были начаты еще в начале прошлого века Э.Л. Вольфом, затем этими вопросами занимался И.И. Галактионов в Академии коммунального хозяйства РСФСР. В конце 70-х годов эту работу в масштабах всей территории СССР проводил А.И. Колесников. Над схемой климатического районирования территории нашей страны работал также Б.П. Алисов. В ГБС РАН в это же время П.И. Лапиным был разработан метод интегральной оценки жизнеспособности и перспективности интродуцированных видов на основе семи разных показателей, в том числе и зимостойкости растений, определяемой по специальной шкале.
Сейчас об этих разработках знают в основном узкие специалисты, и на практике при подборе ассортимента посадочного материала для проектов озеленения они не учитываются. К тому же за прошедшие десятки лет их актуальность перестала быть очевидной, ведь климатические условия на территории Евразии существенно изменились. Наряду с регистрируемыми глобальными изменениями климата растет неравномерность регионального проявления различных климатических явлений.
ОЦЕНКА ЗИМОСТОЙКОСТИ РАСТЕНИЙ
В отсутствии общепризнанных отечественных критериев для оценки зимостойкости растений в практической работе стало принято опираться на карту зон морозоустойчивости растений (USDA Hardiness Zones and Average Annual Minimum Temperature Range), разработанную Департаментом сельского хозяйства США и адаптированную для территории Европы. В качестве основного фактора, лимитирующего вегетацию растений, были использованы величины многолетних средних минимальных температур зимнего периода.

Территория центральной и северо-западной части России, в соответствии с принятой в Европе 11-зонной картой, главным образом попадает в зоны USDA 3-5 с температурными минимумами от -39,9 °С до-23,4 °С.
В зарубежных каталогах и справочниках для каждого растения указываются значения зон морозоустойчивости. При этом подразумевается, что отнесение того или иного растения к конкретному диапазону зон дает гарантию его успешной вегетации в этих условиях. Однако следует сразу отметить, что механическое использование данных карты при посадках приводит к существенным ошибкам, поскольку степень морозоустойчивости растения далеко не всегда является достаточным основанием для оценки возможности его успешного выживания зимой на той или иной территории.
Кроме того, для преобладающей части азиатской территории нашей страны карты зон просто не существует.
Гораздо объективнее успешность вегетации растений оценивается по их зимостойкости. Зимостойкость – это способность растений противостоять тяжелым условиям зимнего и ранневесеннего периодов. Данный термин имеет широкое наполнение, он включает в себя не только собственно морозоустойчивость, то есть способность того или иного вида или сорта выдерживать низкие температуры без повреждения тканей, но и воздействие многих других факторов, например выпревания растений поздней зимой во время оттепелей. В результате выпревания происходит прерывание вынужденного покоя растений и их углеводное истощение.
Кроме того, это влияние вымокания и выпирания растений из почвы, воздействие поздних весенних заморозков, резких скачков температуры с переходом через 0°С, негативная роль ледяной корки и других неблагоприятных зимних факторов. Зимостойкость растений меняется с возрастом, в зависимости от характера микроклимата, почвенно-экологических особенностей, в частности влажности почвы и обеспеченности ее калием и фосфором, а также условий вегетации. Нелишне также напомнить что привитые на штамб растения и пестролистные формы обычно сильнее страдают во время зимы, так же, как молодые и недавно пересаженные растения. Практическое использование зональной карты осложняется также тем, что в пределах каждой зоны условия вегетации и зимовки растений могут существенным образом изменяться в зависимости от ряда местных факторов.
Среди них – высота местности над уровнем моря, положение участка в рельефе, экспозиция склона, направление, частота, периодичность и скорость преобладающих ветров, наличие и высота снежного покрова, продолжительность сильных морозов и зимняя температура почвы. Весьма важными для успешной зимовки растений являются условия их влаго- и теплообеспеченности в предшествующий вегетационный период, сумма активных температур и продолжительность периода вегетации.
Большую роль могут играть условия произрастания: близость крупных городов и промышленных центров, так называемых островов тепла, обширных водных пространств, смягчающих резкие перепады температур, а также защищенность участка с посадками растений. Это может быть стена леса, забор, строения или складки местности.
Большое значение имеет также ь надлежащий уход за культурами: своевременный полив, достаточное минеральное питание, борьба с болезнями и вредителями, зимнее укрытие, если это необходимо. С учетом всех этих обстоятельств многие растения из «группы риска»> могут достаточно успешно расти в защищенных экотопах. Кроме того, они могут использоваться в озеленении в качестве порослевой культуры, чему есть масса примеров.
Читайте также: Морозоустойчивые многолетние цветы
СПОРНЫЕ МОМЕНТЫ
Вполне вероятные в средней полосе 30-градусные морозы уже превышают планку, устанавливаемую для растений зоны USDA 4, а к их числу относят такие привычные для нас виды, как липа мелколистная, ясень обыкновенный, клен остролистный, к. серебристый и к. ясенелистныи, конский каштан обыкновенный, калина обыкновенная, лиственница европейская, сосна горная, многие рябины, яблони, спиреи, кизильники и сирени.
Породы, относящиеся к зоне 5 морозоустойчивости, будут в условиях суровой зимы испытывать еще больший температурный стресс. А это опять же вовсе не редкие экзоты. а хорошие всем знакомые можжевельники многих видов и сортов, туя западная, ель канадская, пихта корейская, вязы голый и мелколистный.
И даже – дубы черешчатый и красный, бузина черная, многие ивы, боярышники, чубушники, гортензии, форзиции и барбарисы, большинство плодовых деревьев и ягодных кустарников. Сказанное выше справедливо в том случае, если отнесение растении к тем или иным зонам морозоустойчивости не вызывает сомнений, но так бывает не всегда. Например, катальпу бигнониевидную в разных каталогах относят к зонам USDA 6Ь и даже 7, хотя известно немало случаев ее успешного произрастания в Москве.
То же самое касается таких растений, как спирея острозазубренная, с. Тунберга, шелковица белая или барвинок малый. Очевидно, что и характеру градации зон, и к соответствию им тех или иных пород с точки зрения их зимней выносливости возникают вопросы.

В ЗАКЛЮЧЕНИЕ
Проблема выбора ассортимента зимостойких декоративных культур, как видим, является достаточно сложной и комплексной. Очевидно, на данном этапе за неимением других подходящих вариантов пока остается целесообразным пользоваться имеющейся картой зон морозоустойчивости, которая намечает определенные ориентиры для практиков – специалистов в области озеленения и опытных садоводов-любителей.
Однако к использованию карты следует подходить критически, опираясь на опыт выращивания конкретных растений в данном регионе. Кроме того, назрела необходимость в создании современной системы древокультурного районирования, охватывающего всю территорию России, на основании прежних разработок и фактических данных, отражающих сложившиеся местные особенности экотопических условий и современные данные по изменению климата.
© Автор: Андрей Лысиков Фото: автора
Реклама
Ниже другие записи по теме "Дача и сад - своими руками"
Подпишитесь на обновления в наших группах.



Будем друзьями!
vsaduidoma.com









