
Наследственная изменчивость: особенности и значение. Наследственная изменчивость примеры растений
особенности и значение :: SYL.ru
В нашей статье речь пойдет об уникальном свойстве всех живых организмов, которое обеспечило возникновение огромного количества видов живых существ. Это наследственная изменчивость. Что это такое, каковы ее особенности и механизм осуществления? На эти и многие другие вопросы вы сейчас найдете ответы.
Что изучает генетика
Сравнительно молодая наука генетика в 19-м веке открыла человечеству многие тайны его происхождения и развития. А предметом ее изучения являются только два свойства живых организмов: наследственность и изменчивость. Благодаря первому обеспечивается преемственность поколений и осуществляется точная передача генетической информации в целом ряду поколений. А вот изменчивость обеспечивает возникновение новых признаков.
Значение изменчивости
Зачем же организму приобретать эти новые признаки? Ответ достаточно прост: для возможности адаптации. На фото ниже перед вами представители нескольких рас одного биологического вида - Человек Разумный. Их морфологические различия на данном этапе не имеют, естественно, никакого приспособительного значения. А вот их далеким предкам новые черты помогали выжить в тяжелых условиях. Так, представители монголоидной расы имеют узкий разрез глаз, поскольку в степях часто были пыльные бури. А негроиды имеют темную кожу в качестве защиты от палящих солнечных лучей.
Виды изменчивости
Изменчивостью называют свойство организмов приобретать новые признаки в процессе их исторического и индивидуального развития. Она бывает двух видов. Это модификационная и наследственная изменчивость. Их объединяет ряд признаков. Например, неизбежно возникают изменения во внешнем строении организмов. Но вот по продолжительности существования модификаций и степени действия они абсолютно отличаются.
Модификационная изменчивость
Этот вид изменчивости является ненаследственным. Он не закрепляется в генотипе, не носит постоянный характер и возникает под воздействием изменений условий окружающей среды. Ярким примером модификационной изменчивости может служить известный опыт с кроликом. Ему сбривали небольшой участок серой шерсти. А на голый участок кожи прикладывали лед. Через некоторое время на этом месте вырастала шерсть белого цвета, которую также сбривали. Но лед в этом случае не прикладывали. В результате на данном участке снова вырастали волосы темного цвета.
Наследственная изменчивость
Данный вид изменчивости носит постоянный характер, поскольку затрагивает структуру генотипа до уровня нуклеотидов ДНК. При этом новые признаки передаются новым поколениям. Наследственная изменчивость, в свою очередь, также бывает двух типов: комбинативная и мутационная. Первая возникает в случае появления нового сочетания генетического материала. Ее самым простым примером служит слияние гамет в ходе полового размножения. В результате организм, получая по половине генетической информации от мужского и женского организма, приобретает новые признаки.
Второй вид - это мутационная наследственная изменчивость. Она заключается в возникновении резких ненаправленных изменений генотипа под воздействием различных факторов. Ими могут быть ионизирующее и ультрафиолетовое излучение, высокая температура, азотсодержащие химические вещества и другие.
В зависимости от уровня структуры генетического аппарата, в котором происходят изменения, различают несколько типов таких наследственных модификаций. При геномных изменяется число хромосом в общем наборе. Это ведет к анатомическим и морфологическим изменениям в организме. Так, появление третьей хромосомы в 21-й паре вызывает болезнь Дауна. При хромосомных мутациях возникает перестройка этой структуры. Они встречаются гораздо реже, чем геномные. Участки хромосом могут дублироваться или отсутствовать, перекручиваться, изменять свое положение. А вот генные мутации, которые также называют точечными, нарушают последовательность мономеров в структуре нуклеиновых кислот.
Независимо от вида мутаций, все они, как правило, не несут для организма полезных признаков. Поэтому человек учится управлять ими искусствено. Так, в селекции часло используется полиплоидия - кратное увеличение числа хромосом в наборе. В результате растение становится более мощным и дает крупные плоды в большом количестве. Никого уже не удивишь инжирным персиком и другими вкусными растительными гибридами. А ведь они являются результатом искусственно проведенной наследственной изменчивости.
Наследственная изменчивость в процессе эволюции
Развитие генетики помогло сделать значительный шаг вперед и в развитии эволюционного учения. Тот факт, что человека и обезьяну отличает лишь одна пара хромосом, стал существенным доказательством теории Дарвина. У растений и животных в историческом развитии можно проследить наследование прогрессивных черт, которые передавались и закреплялись в генотипе. К примеру, водоросли вышли на сушу благодаря тому, что в генотипе закрепился признак наличия механической и проводящей тканей. Каждое последующее поколение оставляло для себя только нужные, полезные признаки, которые корректировались в зависимости от условий обитания и окружающей среды. Так появились господствующие виды растений и животных, обладающие самыми прогрессивными чертами строения.
Итак, наследственная изменчивость - это способность организмов приобретать новые признаки, которые закрепляются в генотипе. Такие изменения носят продолжительный характер, не исчезают при изменении условий среды и передаются по наследству.
www.syl.ru
Наследственность и изменчивость древесных растений
Наследственность и изменчивость древесных растений
Генетика – это наука, которая изучает механизмы и закономерности наследственности и изменчивости организмов, а также методы управления этими процессами.
Наследственностью называют свойство организмов обеспечивать материальную и функциональную преемственность поколений, определенный план строения и характер их индивидуального развития, а также норму реакции на условия внешней среды.
Изменчивостью называют различия признаков и свойств между двумя или группой особей, предками и потомками одного и того же или разных видов растений и животных.
Наследственность и изменчивость изучается на разных уровнях организации живой материи: молекулярном, хромосомном, клеточном, организменномипопуляционном. Первые уровни будут более подробно рассмотрены далее.
Что же касается организменногоипопуляционного уровня, то изучаемая на них изменчивость может быть разделена еще на несколько групп: метамерная изменчивость (различия отдельных частей организма), индивидуальная изменчивость (различия у отдельных особей), групповая, или внутрипопуляционная изменчивость (различия в группе особей одного вида), межпопуляционная изменчивость и надвидовая изменчивость.
При изучении изменчивости выделяют качественные и количественные признаки. По характеру изменения признаков различают прерывистую и непрерывную (клинальную) изменчивость. Эти типы изменчивости отмечают при рассмотрении признака во времени и пространстве. Примером прерывистой изменчивости во времени могут служить возрастные изменения светолюбия сосны обыкновенной. Примером прерывистой изменчивости в пространстве могут служить изменения, вызванные резким перепадом экологических и климатических условий, получившие название экологической и географической изменчивости. Непрерывную изменчивость можно наблюдать, например, на приросте запаса древесины в лесу. Непрерывно падает бонитет древостоя в одном и том же типе леса в направлении с юго-запада на северо-восток.
С.А. Мамаев предлагает делить изменчивость древесных растений на два типа: внутривидовую и внутриорганизменную, или эндогенную. Внутривидовая изменчивость подразделяется на несколько форм: индивидуальную, половую, хронографическую (сезонную и возрастную), экологическую, географическую и гибридогенную.
В зависимости от того передаются ли изменения потомству, изменчивость делят на две категории: наследственную (возникающую в результате мутаций и рекомбинаций) и ненаследственную, или модификационную.
5.Методы изучения наследственности.
Анализ наследственности осуществляется в основном на трех уровнях организации живой материи (молекулярно-клеточном, организменном и популяционном, или групповом). Для изучения наследственности на каждом из этих уровней используется своя группа методов.
К первой группе – относятся цитологический, биохимический и молекулярно-генетический методы, которые служат для изучения структуры материальных носителей наследственности. Цитологический метод направлен на изучение структур и органелл клетки. Биохимический метод используется для изучения структуры химических компонентов клетки и метаболических процессов, происходящих в клетках на различных этапах ее развития. Из биохимического метода позднее, в качестве самостоятельного, выделился молекулярно-генетический метод, объектом анализа которого служат изменения в структуре нуклеиновых кислот.
Ко второй группе – относятся гибридологический и генеалогический методы, с помощью которых определяются закономерности наследования того или иного признака или группы признаков. Гибридологический метод(классический метод генетического анализа) изучает характер наследования признаков по анализу потомков (при скрещивании особей, различающихся по контрастным (альтернативным) признакам). Генеалогический метод изучает характер наследования признаков по анализу предков.
Характерной особенностью третьей группы методов изучения наследственности, к которой относятся приемы популяционно-статистического анализа, является изучение степени влияния генов и факторов внешней среды на развитие признаков и свойств организмов. Сущность этого метода состоит в том, что на достаточно репрезентативной выборке особей одного вида или сорта, определяется средняя величина признака и другие статистические параметры. Основная задача метода – определение коэффициента наследуемости с целью оценки пригодности изучаемого признака для целей селекции.
10.
Кроссинго́вер — явление обмена участками гомологичных хромосом во время конъюгации при мейозе. Помимо мейотического описан также митотический кроссинговер.
Биологическое значение кроссинговера.
Благодаря сцепленному наследованию удачные сочетания аллелей оказываются относительно устойчивыми. В результате образуются группы генов, каждая из которых функционирует как единый суперген, контролирующий несколько признаков. В то же время, в ходе кроссинговера возникают рекомбинации – т.е. новые комбинации аллелей. Таким образом, кроссинговер повышает комбинативную изменчивость организмов.
Это означает, что…
а) в ходе естественного отбора в одних хромосомах происходит накопление «полезных» аллелей (и носители таких хромосом получают преимущество в борьбе за существование), а в других хромосомах скапливаются нежелательные аллели (и носители таких хромосом выбывают из игры – элиминируются из популяций)
б) в ходе искусственного отбора в одних хромосомах накапливаются аллели хозяйственно-ценных признаков (и носители таких хромосом сохраняются селекционером), а в других хромосомах скапливаются нежелательные аллели (и носители таких хромосом выбраковываются).
11.
12.
Моногибридное скрещивание – это скрещивание, при котором родительские формы различаются по одной паре альтернативных признаков.
Особи, которые не дают в потомстве ращепления и сохраняют свои признаки в «чистом» виде, называют гомозиготными, а те, у которых в потомстве происходит ращепление,- гетерозиготными.
Совокупность всех признаков организма, начиная с внешних и кончая особенностями строения функционирования клеток, тканей и органов, называется фенотипом.
Признаки и свойства организма проявляются под контролем наследственных факторов, т.е. генов. Совокупность всех генов организма называют генотипом.Алле́ли — различные формы одного и того же гена, расположенные в одинаковых участках гомологичных хромосом и определяющие альтернативные варианты развития одного и того же признака.Домина́нтность (доминирование) — форма взаимоотношений между аллелями одного гена, при которой один из них (доминантный) подавляет (маскирует) проявление другого (рецессивного) и таким образом определяет проявление признака как у доминантных гомозигот, так и у гетерозигот.
РЕЦЕССИВНОСТЬ форма взаимоотношений двух аллельных генов, при которой один из них (рецессивный) оказывает менее сильное влияние на соответствующий признак, чем другой (доминантный). Моногибридное скрещивание. Первый закон Менделя.
В опытах Менделя при скрещивании сортов гороха, которые имели желтые и зеленые семена, все потомство (т.е. гибриды первого поколения) оказалось с желтым семенами. При этом не имело значения, из какого именно семена (желтого или зеленого) выросли материнские (отцовские) растения. Итак, оба родителя в равной степени способны передавать свои признаки потомству. Аналогичные результаты были обнаружены и в опытах, в которых во внимание брались другие признаки. Так, при скрещивании растений с гладкими и морщинистым семенами все потомство имело гладкие семена. При скрещивании растений с пурпурными и белыми цветками у всех гибридов оказались лишь пурпурные лепестки цветков и т. д. Обнаруженная закономерность получила название первый закон Менделя, или закон единообразия гибридов первого поколения. Состояние (аллель) признака, проявляющегося в первом поколении, получило название доминантного, а состояние (аллель), которое в первом поколении гибридов не проявляется, называется рецессивным. «Задатки» признаков (по современной терминологии - гены) Г. Мендель предложил обозначать буквами латинского алфавита. Состояния, принадлежащие к одной паре признаков, обозначают одной и той же буквой, но доминантный аллель - большой, а рецессивный - маленькой.
13.
Наследование при дигибридном скрещивании. При моногибридном скрещивании родительские формы отличаются по одной паре признаков или аллелей гена. Совершенно очевидно, что в большинстве случаев организмы различаются по многим генам. Чтобы проанализировать наследование нескольких признаков, необходимо разложить эти сложные явления на более простые составные элементы, а затем представить весь процесс в целом. Так поступал в своей работе Г. Мендель. Он изучил наследование каждой пары признаков в отдельности, не обращая внимания на другие пары, а затем сопоставил и объединил все эти наблюдения.
Гибриды, полученные от скрещивания организмов, различающихся двумя парами альтернативных признаков, были названы дигетерозиготами, тремя парами — тригетерозиготами,, многими признаками — полигетерозиготами, а скрещивания соответственно ди-, три- и полигибридными. Для дигибридного скрещивания Г. Мендель взял гомозиготные растения гороха, различающиеся одновременно по двум парам признаков. Материнское растение имело гладкие семена желтой окраски, отцовские — морщинистые зеленые семена. Гибрид первого поколения этого скрещивания имеет гладкие и желтые семена. Следовательно, гладкая форма семени доминирует над морщинистой, а желтая — над зеленой. Обозначим аллели гладкой формой А, морщинистой а, аллели желтой окраски В, зеленой b . Гены, определяющие развитие разных пар признаков, называются неаллельными. В данном случае гены формы семени А и а неаллельны генам окраски b и В. Неаллельные гены обозначают разными буквами алфавита.
Родительские растения имели генотипы ААВВ и aabb и образовывали гаметы соответственно АВ и ab. В этом случае генотип гибрида f1 будет АаВЬ, т. е. он является дигетерозиготным.Таким образом, неаллельные гены при образовании гамет свободно комбинируются между собой, благодаря чему образуются новые комбинации генов (Ab и ab) по сравнению с родительскими формами и новые комбинации признаков у гибридов рь — гладкие зеленые семена и морщинистые желтые. Появление новых комбинаций признаков в результате скрещивания получило название комбинативной изменчивости. Комбинативная изменчивость играет большую роль в эволюции, так как она дает новые сочетания приспособительных признаков, возникающих при скрещивании. Она постоянно используется и в селекции для улучшения пород животных и сортов растений путем скрещиваний.
14.
Мутация –это внезапно наследуемые изменения генетического материала которые могут возникнуть спонтанно либо могут быть индуцированны внешним воздействием на организм. Процесс возникновения мутации называется мутагенезом.
Существует следующая классификация мутаций:
· По характеру изменения генома мутации подразделяются:
Генные
Хромосомные
Геномные (изменение числа хромосом)
· По происхождению:
В спонтанные (без видимых причин)
Индуцированные (вызваны искусственно)
· По проявлению:
Доминантные
Рецессивные
Основное внимание при изучении генных мутаций уделяют нуклеотидам ДНК.
Генные мутации подразделяют на следующие группы:
Ø Транзиция - замена азотистых оснований: пуриновых на пуриновые, пиримидиновых на пиримидиновые. При этом изменяется только тот кодон, в котором произошли танзиции.
Ø Трансверсия - замена пуриновых оснований на пиримидиновые или пиримидиновые на пуриновые.
Ø Вставки – происходит вставка линий пары нуклеотидов
Ø Выпадение – выпадение пары нуклеотидов
Генные мутации обычно затрачивают единичные гены, по этому они образуются наиболее часто. С ними связан большинство изменений: морфологических, биохимических, физиологических признаков организма.
16.
По причинам вызывающим мутации их подразделяют на спонтанные и индуцированные.
Спонтанные мутации происходят под действием естественных мутагенных факторов, без вмешательства человека.
Индуцированные мутации - результат направленного воздействия определенных мутагенных факторов.
18.
Под полиплоидией понимают кратное увеличение основного числа хромосом в ядре, а всякое отклонение числа хромосом от нормального диплоидного в сторону как увеличения, так и уменьшения, а также кратное и некратное называют гетерополиплоидией или гетероплоидией.
В 1916 г. Г. Винклер, изучая прививки паслена на томат, обнаружил в местах соединения привоя и подвоя клетки с увеличенным набором хромосом. Ученый назвал это явление полиплоидией (от греч. poly — многократный и plooseidos — вид).
Интерес к полиплоидии еще больше возрос в 40-х годах XX в., когда американские исследователи Блексли и Эйвери, а также Эйгсти, Небель и Раттл (1937) провели многочисленные успешные опыты по обработке семян и растений колхицином с целью получения полиплоидов и разработали основные способы удвоения числа хромосом в клетках растений.
Механизм действия колхицина на делящиеся клетки состоит в том, что он блокирует веретено деления в метафазе, в результате чего дочерние хромосомы не расходятся к полюсам, а остаются в центре материнской клетки (С-митоз или K-митоз).
Экспериментально возникшая в 30-х годах XX в. полиплоидия стала играть огромную роль в селекции сельскохозяйственных растений, а также в генетической инженерии.
20.
Популяция – совокупность особей одного вида, длительно населяющих определ. террит. и имеющих сходный генофонд в следствие свободного скрещивания между собой.
Обширность ареола, население популяций, зависит от многих факторов
Одним из факторов явл. подвижность особей.
Совокупность всех генов в популяции наз. генофондом. Генофонды популяций объед. и образуют генофонд вида.
В популяции могут сущ-ть чистые линии – гомозиготные организмы. Поэтому отбор может идти только в полиморфных популяциях.
Все популяции можно разделить на 3 категории, в зависимости от степени их изоляции, от регулярности и постоянства связей м/у особями разных популяций.
1. географические- разобщены географ факторами(горы, реки)
2. экологические- разобщены м/ду соб вследствие действия эколог факторов(сезонность)
3. элементарные- изоляция в таких популяциях носит эпизодический характер.В этих попул-х осущ-ся регулярн и частые миграции особей из одной популяции в другую.
Наследственность популяции- распределение в ней генотипов и фенотипов аллелей.Генетич состав попул относительно постоянен.
21.
Основные положения хромосомной теории наследственности
Сформулировал Морган: 1. главным клеточными структурами ответственными за передачу наследственной информации, явл хромосомы кот содержат гены – носители генетической информации. 2. в хромосомах гены расположены линейно, в пределах одной хромосомы гены образуют одну группу сцепления. Число групп сцепления равно гаплоидному числу хромосом. 3. В мейозе между гомологичными хромосомами мажет происходить кроссинговер, что явл основой биологического разнообразия видов и базой для естественного отбора организмов.
В ядре наход хромосомы. Различ два вида хромосом гаплоидный и диплоидный. На стадии метафазы и ранней анафазы митоза и мейоза легче всего опред форму и размер хромосом. Совок всех морф признаков назыв кариотипом. Форма каждой хромосомы определяется положением первичной перетяжки, где располагается центромера. Ген – элементарная единица наследственности. Ген часть хромосомы ответственная за проявления отдельного признака организма. Гены отдельн признаков лежат в опред местах хромосом – локусах. Хромосомы в клетках парные. Парные гены назыв аллеями. Гены классифицируют по признакам которыми они управляют. Аддитивные гены имеют слабый эффект контролируют один и тот же признак. Гены доминантные вызывает выраженность признака. Рецессивный ген – это ген действ кот проявляется очень слабо или совсем не проявляется. Эпистатические гены если один ген по отношению к другоме доминантен и если эти гены не аллельны.
Гетерозис.
Гетерозис — увеличение жизнеспособности гибридов вследствие унаследования определённого набора аллелей различных генов от своих разнородных родителей. Это явление противоположно результатам инбридинга, или близкородственного скрещивания, приводящего к гомозиготности. Увеличение жизнеспособности гибридов первого поколения в результате гетерозиса связывают с переходом генов в гетерозиготное состояние, приэтом рецессивные летальные и полулетальные аллели, снижающие жизнеспособность гибридов, не проявляются. Также в результате гетерозиготации могут образовываться несколько аллельных вариантов фермента, действующих в сумме более эффективно, чем поодиночке (в гомозиготном состоянии). Механизм действия гетерозиса ещё не окончательно выяснен. Явление гетерозиса зависит от степени родства между родительскими особями: чем более отдалёнными родственниками являются родительские особи, тем в большей степени проявляется эффект гетерозиса у гибридов первого поколения.
Явление гетерозиса наблюдалось ещё до открытия законов Менделя И. Г. Кёльрейтером, термин «гетерозис» (в переводе с греческого языка — изменение, превращение), в 1908 Г. Шулл описал гетерозис у кукурузы.
У растений (по А. Густафсону) выделяют три формы гетерозиса: т. н. репродуктивный гетерозис, в результате которого повышается плодородность гибридов и урожайность, соматический гетерозис, увеличивающий линейные размеры гибридного растения и его массу, и приспособительный гетерозис (называемый также адаптивным), повышающий приспособленность гибридов к действию неблагоприятных факторов окружающей среды.
Анализ эффекта гетерозиса с позиций открытых в начале XX в. основных генетических закономерностей гетерозиса:
1) гетерозиготным состоянием гибридов по многим генам;
2) возникновением взаимодействия доминантных благоприятных генов; 3) сверхдоминированием — гетерозигота превосходит гомозиготу. В доказательство первого положения приводятся примеры получения межлинейных гибридов. При скрещивании гомозиготных инбредных линий гибриды первого поколения гетерозиготны по многим генам. При этом действие вредных рецессивных мутантных аллелей подавляется доминантными аллелями обоих родителей.
Большой хозяйственный интерес представляют пути закрепления гетерозиса у древесных растений. Сохранение эффекта гетерозиса осуществляется вегетативным размножением уникальных гибридов. Мы живем в век невиданных темпов научно-технического прогресса, обусловленного интенсивным развитием и огромными достижениями современной науки. Среди наук, определяющих научно-технический прогресс, наряду с физикой, химией, электроникой и другими науками, все большее значение приобретают биологические, в особенности молекулярная биология, включающая молекулярную генетику и биохимию. Эти науки за последние 30 лет достигли выдающихся результатов в познании живого.
ГЕНЕТИКА ПОПУЛЯЦИЙ
Все виды живых организмов на Земле представлены определенными группами особей, которые называются популяциями. Популяция — это элементарная единица эволюции. Действительно, вид, занимающий часто огромный ареал, содержащий многообразие форм, различных в каждом географическом районе, оказывается слишком громоздкой системой для однозначного ответа на действие естественного отбора, имеющего различные направления, характер и силу в разных районах. В то же время отдельный организм (особь) также не может быть единицей эволюции, так как содержит лишь незначительную часть генетической информации вида, а с его смертью утрачивается и эта доля информации. Таким образом, единицей эволюции может быть только достаточно большая группа особей вида, которая будет репрезентативной (представительной) и в то же время достаточно малой, чтобы быть подвижной системой, чутко реагирующей, на воздействие естественного отбора. Такой группой особей оказалась популяция.
Впервые термин популяция был предложен 3. Л. Иоганнсеном 1907 г. в работе «Чистые линии и популяции». В его трактовке популяция — это группа особей, отличающихся от чистой линии тем, что каждый признак в ней представлен многими аллелями основного, определяющего его гена. В чистой линии признак представлен моноаллельно. Другими словами, гетерозиготность признака отличает популяцию от гомозиготности его в чистой линии.
Современный этап экспериментального изучения популяции начался с выхода в 1926 г. работы С. С. Четверикова «О некоторых моментах эволюционного процесса с точки зрения современной генетики». Учение о популяциях как раздел общей биологии еще только развивается и не достигло уровня законченного исследования. Однако значение этого раздела трудно переоценить, так как учение о популяциях имеет прикладное значение. Учение о популяциях включает вопросы: динамики численности вида, микроэволюционных процессов и пусковых механизмов в эволюции, систематики и таксономии и, наконец, что особенно важно для лесоводства, вопросы, связанные с изучением генетики объектов, обычный генетический анализ которых сильно затруднен.
24.
Структура популяций
Показатели структуры популяций. Как первая надорганизмен-ная биологическая система, популяция обладает определенной структурой и свойствами. Структуру популяции отражают такие ее показатели, как численность и распределение особей в пространстве, соотношение групп по полу и возрасту, их морфологические, поведенческие и другие особенности.
Численность — общее количество особей в популяции. Если рождаемость перестанет покрывать естественную убыль, и оставшиеся особи в течение сравнительно короткого времени вымрут.
Плотность — число особей на единицу площади или объема. При увеличении численности плотность популяции, как правило, возрастает; она остается прежней лишь в случае ее расселения и расширения ареала.
Пространственная структура популяции характеризуется особенностями размещения особей на занимаемой территории. Она определяется свойствами местообитания и биологическими особенностями вида. Наряду со случайным и равномерным распределением в природе наиболее часто встречается групповое распределение.
Половая структура отражает определенное соотношение мужских и женских особей в популяции.
Возрастная структура отражает соотношение различных возрастных групп в популяциях, зависящее от продолжительности жизни, времени наступления половой зрелости, числа потомков в помете, количества потомств за сезон и др. Если какая-либо возрастная группа сокращается либо увеличивается, это сказывается на общей численности популяции.
Экологическая структура свидетельствует об отношении различных групп организмов к условиям окружающей среды.
Внутривидовой полиморфизм обусловливает уникальность сочетания аллелей у разных особей. Причиной полиморфизма служат половое размножение, которое обеспечивает комбинативную изменчивость, и мутации, изменяющие субстрат наследственности. Поддержание внутривидового полиморфизма обеспечивает устойчивость вида и гарантирует его существование в различных условиях среды.
Полиморфи́зм в биологии — способность некоторых организмов существовать в состояниях с различной внутренней структурой или в разных внешних формах.
Внешний (а также внутренний, например, биохимический) полиморфизм может быть обусловлен внутривидовыми генетическими различиями. С другой стороны, возможен полиморфизм, при котором организмы с практически идентичным геномом в зависимости от внешних условий приобретают различные фенотипические формы.
27.
Харди — Вайнберга закон, закон популяционной генетики, устанавливающий соотношение между частотами генов и генотипов в популяции со свободным скрещиванием. Сформулирован в 1908 независимо английским математиком Г. Харди и немецким врачом В. Вайнбергом. Закон утверждает, что если численность популяции диплоидных организмов настолько велика, что можно пренебречь случайными флуктуациями частот генов ), если в ней отсутствуют мутации, миграция и отбор (по изучаемому гену), то частоты генотипов AA, Aa и aa в популяции остаются одинаковыми из поколения в поколение (после первого) и удовлетворяют соотношениям Харди ‒ Вайнберга:p2(AA): 2pq (Aa): q2(aa), где А и а ‒ аллели несцепленного с полом гена, p ‒ частота аллеля А,q ‒ частота аллеля а. Х. ‒ В. з. распространяется и на случай многоаллельного гена. В популяциях полиплоидных организмов (а также в популяциях диплоидов ‒ для генов, сцепленных с полом) соответствующие соотношения устанавливаются лишь через большое число поколений. Если в популяции выполняются соотношения Х. ‒ В. з., то это не свидетельствует ещё об отсутствии популяционно-генетических процессов. Например, скрещивание близкородственных особей (инбридинг), способствующее увеличению доли гомозигот в популяции, в сочетании с отбором против гомозигот может привести к частотам генотипов, удовлетворяющим соотношениям Х. ‒ В. з. Сопоставление фактически наблюдаемых частот генотипов с теоретически ожидаемыми по Х. ‒ В. з. в ряде случаев позволяет оценить частоты аллелей, вычленить влияющие на них факторы и получить количественные характеристики отбора, неслучайности скрещивания, миграции, случайных флуктуаций и т.п. Представление о генетическом равновесии в популяциях, впервые нашедшем выражение в Х. ‒ В. з., составляет основу современной концепции о взаимодействии популяционно-генетических процессов.
28.
Генная инженерия — понятие более узкое, чем генетическая инженерия, и имеет отношение только к отдельному гену или генам. В ее задачу входят выделение, конструирование и клонирование новых рекомбинантных генов, или молекул ДНК, создание банков генов. В то же время генетическая инженерия — более широкое понятие. Она изучает проблемы направленного конструирования с помощью методов генной инженерии новых живых существ с заданными наследственными признаками и свойствами.
Техника выращивания в искусственных условиях клеток высших организмов резко расширила возможности генетической инженерии растений и животных, поскольку клетки, и в особенности «голые» клетки растений (протопласты), являются удобными реципиентами для введения чужеродного генетического материала.
Вместе с тем культивирование клеток и тканей в пробирках приобрело самостоятельное практическое значение. Оказалось, что в процессе культивирования с клетками можно осуществлять различного рода манипуляции, в результате которых можно получать клетки, а у растений даже целые организмы с новыми наследственными свойствами (гибриды), клетки, способные продуцировать, в огромных количествах важные для человека вещества, моноклональ-ные антитела, размножать (клонировать) ценные генотипы и т. д. Это направление биологии получило название клеточной инженерии. Генетическая и клеточная инженерия направлены на решение общей задачи — осуществление контролируемых биологических манипуляций, связанных с генами, хромосомами, геномами, клетками, протопластами и органеллами клеток с целью создания новых генетических программ (генотипов).
Генетическая инженерия — наука совсем молодая. Формально датой ее рождения считают 1972 год, когда американский ученый профессор Берг с сотрудниками создали первую химерную (гибридную) молекулу ДНК.
29.
Генофонд лесных древесных пород и его сохранение.
Для успешного долговременного селекционного улучшения лесных древесных пород необходимо иметь широкую генетическую базу или значительный генофонд, поэтому очень важно заботиться о сохранении уже существующего генетического потенциала популяций. В любой селекционной программе необходимо предусматривать консервацию генов и генных комплексов, при этом необходимо не допускать сужение генетической базы или возникновения эффекта инбредной депрессии.
консервация генов имеет большое значение и его общая цель – сохранение.
Изучите методы консервирования генетических ресурсов:
- консервация генофонда in situ;
- консервация ex situ;
Консервация in situ, или консервация деревьев и насаждений в естественных популяциях, т. е. в местах их извечного проживания.
Консервация ex situ, или сохранение генов, генных комплексов или генотипов в искусственных условиях, т. е. не в месте их естественного проживания.
Следует также заметить, что одни группы растении могут сохраняться только in situ, другие — только ex situ, третьи — обоими методами.
- консервация географических происхождений, экотипов и популяций;
- консервация признаков отдельных деревьев.
Изучите формы выделения и сохранения ценного генофонда лесных древесных пород в России:
- лесные генетические резерваты;
- отбор и сохранение отдельных ценным насаждений и деревьев.
Имейте представление о мировой политике в области сохранения биоразнообразия и консервации генофонда.
Генофонд — это совокупность генов одной популяции, в пределах которой они характеризуются определенной частотой встречаемости. целью селекции является использование существующей изменчивости для получения большего числа деревьев с желательными характеристиками роста, качества или устойчивости. При этом генетическая база обычно ограничивается или сужается для хозяйственно важных характеристик. В начале любой генетико-селекционной программы, особенно ориентированной на отбор, а также в начале любой хозяйственной деятельности, которая ведет к изменениям в составе генофонда, необходимо позаботиться о сохранении уже существующего генетического потенциала популяций..
Причины потерь генетических ресурсов могут быть самыми разными, например: инвазия насекомых, эпизоотия, вырубка, очистка для сельскохозяйственного пользования, экспансия городов, пожары, штормы и др. Любое действие, которое разрушает леса или часть леса, может вести к опасной в генетическом отношении ситуации. Особенно эта ситуация пагубна для популяций с ограниченным ареалом. Значительна опасность исчезновения генофонда в экстремальных условиях, в которых изведение лесов не компенсируется их восстановлением. К тяжелым последствиям ведет не только полное исчезновение вида, но и просто истощение его генетического потенциала, так как в этом случае возможна потеря устойчивости и наиболее ценных комплексов генов.
31.
Селе́кция —отрасль сельского хозяйства, занимающуюся выведением новых сортов и гибридов сельскохозяйственных культур и пород животных. Лесная (от лат. selectio - выбор отбор), наука о методах отбора в естественных популяциях или искусственного получения форм и сортов древесных и кустарниковых растений, имеющих хозяйственную ценность. Селекция как наука оформилась лишь в последние десятилетия. В прошлом она была больше искусством, чем наукой. Селекция разрабатывает способы воздействия на растения и животных с целью изменения их наследственных качеств в нужном для человека направлении.
История:Первоначально в основе селекции лежал искусственный отбор, когда человек отбирает растения или животных с интересующими его признаками. До XVI—XVII веков отбор происходил бессознательно. Только в последнее столетие человек, еще не зная законов генетики, стал использовать отбор сознательно или целенаправленно, скрещивая те растения, которые удовлетворяли его в наибольшей степени. История включает 3 периода: додарвинский, дарвинский, современный. Додарвинский период в истории развития практической селекции включает в свою очередь этапы примитивной, народной и промышленной селекции. Примитивная селекция у древних народов развивалась очень медленно, ее успехи носили случайный характер. В эпоху неолита, т.е. за 10000 лет до н.э., люди уже возделывали многие растения. Дикие виды растений вводились в культуру путем отбора лучших форм растений. В конце XVIII — начале XIX вв. с развитием производительных сил общества возникает промышленная селекция. Выведение новых пород животных и сортов растений, проводившееся в широких масштабах многими заводами и фирмами, приняло промышленный характер. В результате Ч. Дарвин создал теорию селекции — учение об искусственном отборе. Историческое значение достижений практической селекции заключалось в накоплении ценного фактического материала, позволившего ему открыть творческий характер искусственного отбора в эволюции культурных растений. Изменчивость, наследственность и отбор были названы им факторами — предпосылками селекции. Учение об изменчивости живых форм становится предметом науки. С 1900 г начался современный этап. Генетика оформилась как самостоятельная наука.
32.
Селекция лесная является частью селекции - прикладного раздела генетики, который разрабатывает теорию, методы создания и совершенствования сортов растений. Основные этапы лесной селекции: 1) изучение сортового и видового разнообразия древесных растений, являющихся объектами селекционной работы; 2) анализ закономерностей наследования при гибридизации и мутационном процессе, основанный на данных частной генетики; 3) исследование роли среды в развитии признаков и свойств селектируемых видов; 4) разработка систем искусственного отбора, способствующих закреплению и усилению желательных признаков. Лесная селекция является базой для развития лесного семеноводства на генетико-селекционной основе. Селекцию древесных растений проводят на повышение продуктивности, улучшение качественных показателей растений и усиление устойчивости (к патогенам насекомым - вредителям леса).
33.
Задачи современной селекции вытекают из ее определения — это выведение новых и совершенствование уже существующих сортов растений, пород животныхимикроорганизмов. Сортом, породой называют устойчивую группу (популяцию) живых организмов, искусственно созданную человеком и имеющую определенные наследственные особенности.
Цель селекции заключаются в изучении изменчивости и наследственности хозяйственно-ценных признаков и кустарников. Селекцию древесных растений проводят на повышение продуктивности, улучшение качественных показателей растений и усиление устойчивости (к патогенам насекомым - вредителям леса). Задача этой науки заключается в разработке селекционно-генетических методов повышения продуктивности и улучшения качества создаваемых лесов. Основные методы селекции лесной - искусственный мутагенез, отбор (массовый, индивидуальный, групповой) и гибридизация, основанными на достижениях генетики: методом выведения самоопыленных линий и последующего получения линейных гибридов, методом экспериментальной полиплоидии, методом экспериментального мутагенеза.
34.
Географическая изменчивость
Не нашли то, что искали? Воспользуйтесь поиском гугл на сайте:
zdamsam.ru
Наследственная изменчивость
Наследственная изменчивость
Комбинативная изменчивость. Наследственную, или геноти-пическую, изменчивость подразделяют на комбинативную и мутационную.
Комбинативной называют изменчивость, в основе которой лежит образование рекомбинаций, т. е. таких комбинаций генов, которых не было у родителей.
В основе комбинативной изменчивости лежит половое размножение организмов, вследствие которого возникает огромное разнообразие генотипов. Практически неограниченными источниками генетической изменчивости служат три процесса:
- Независимое расхождение гомологичных хромосом в первом мейотическом делении. Именно независимое комбинирование хромосом при мейозе является основой третьего закона Менделя. Появление зеленых гладких и желтых морщинистых семян гороха во втором поколении от скрещивания растений с желтыми гладкими и зелеными морщинистыми семенами — пример комбинативной изменчивости.
- Взаимный обмен участками гомологичных хромосом, или кроссинговер (см. рис. 3.10). Он создает новые группы сцепления, т. е. служит важным источником генетической рекомбинации аллелей. Рекомбинантные хромосомы, оказавшись в зиготе, способствуют появлению признаков, нетипичных для каждого из родителей.
- Случайное сочетание гамет при оплодотворении.
Эти источники комбинативной изменчивости действуют независимо и одновременно, обеспечивая при этом постоянную «перетасовку» генов, что приводит к появлению организмов с другими генотипом и фенотипом (сами гены при этом не изменяются). Однако новые комбинации генов довольно легко распадаются при передаче из поколения в поколение.
Комбинативная изменчивость является важнейшим источником всего колоссального наследственного разнообразия, характерного для живых организмов. Однако перечисленные источники изменчивости не порождают существенных для выживания стабильных изменений в генотипе, которые необходимы, согласно эволюционной теории, для возникновения новых видов. Такие изменения возникают в результате мутаций.
Мутационная изменчивость. Мутационной называется изменчивость самого генотипа. Мутации — это внезапные наследуемые изменения генетического материала, приводящие к изменению тех или иных признаков организма.
Основные положения мутационной теории разработаны Г. Де Фризом в 1901—1903 гг. и сводятся к следующему:
- Мутации возникают внезапно, скачкообразно, как дискретные изменения признаков.
- В отличие от ненаследственных изменений мутации представляют собой качественные изменения, которые передаются из поколения в поколение.
- Мутации проявляются по-разному и могут быть как полезными, так и вредными, как доминантными, так и рецессивными.
- Вероятность обнаружения мутаций зависит от числа исследованных особей.
- Сходные мутации могут возникать повторно.
- Мутации ненаправленны (спонтанны), т. е. мутировать может любой участок хромосомы, вызывая изменения как незначительных, так и жизненно важных признаков.
Почти любое изменение в структуре или количестве хромосом, при котором клетка сохраняет способность к самовоспроизведению, обусловливает наследственное изменение признаков организма. По характеру изменения генома, т. е. совокупности генов, заключенных в гаплоидном наборе хромосом, различают генные, хромосомные и геномные мутации.
Генные, или точковые, мутации— результат изменения нуклеотидной последовательности в молекуле ДНК в пределах одного гена. Такое изменение в гене воспроизводится при транскрипции в структуре иРНК; оно приводит к изменению последовательности аминокислот в полипептидной цепи, образующейся при трансляции на рибосомах. В результате синтезируется другой белок, что ведет к изменению соответствующего признака организма. Это наиболее распространенный вид мутаций и важнейший источник наследственной изменчивости организмов.
Существуют разные типы генных мутаций, связанных с добавлением, выпадением или перестановкой нуклеотидов в гене. Это дупликации (повторение участка гена), вставки (появление в последовательности лишней пары нуклеотидов), делеции ("выпадение одной или более пар нуклеотидов), замены нуклеотид-ных пар (AT -> ГЦ; AT ->; ЦГ; или AT -> ТА), инверсии (переворот участка гена на 180°).
Эффекты генных мутаций чрезвычайно разнообразны. Большая часть из них фенотипически не проявляется, поскольку они рецессивны. Это очень важно для существования вида, так как в большинстве своем вновь возникающие мутации оказываются вредными. Однако их рецессивный характер позволяет им длительное время сохраняться у особей вида в гетерозиготном состоянии без вреда для организма и проявиться в будущем при переходе в гомозиготное состояние.
Вместе с тем известен ряд случаев, когда изменение лишь одного основания в определенном гене оказывает заметное влияние на фенотип. Одним из примеров служит такая генетическая аномалия, как серповидноклеточная анемия. Рецессивный аллель, вызывающий в гомозиготном состоянии это наследственное заболевание, выражается в замене всего одного аминокислотного остатка в (B-цепи молекулы гемоглобина (глутаминовая кислота —» —> валин). Это приводит к тому, что в крови эритроциты с таким гемоглобином деформируются (из округлых становятся серповидными) и быстро разрушаются. При этом развивается острая анемия и наблюдается снижение количества кислорода, переносимого кровью. Анемия вызывает физическую слабость, нарушения деятельности сердца и почек и может привести к ранней смерти людей, гомозиготных по мутантному аллелю.
Хромосомные мутации (перестройки, или аберрации) — это изменения в структуре хромосом, которые можно выявить и изучить под световым микроскопом.
Известны перестройки разных типов (рис. 3.13):
- нехватка, или дефишенси, — потеря концевых участков хромосомы;
- делеция — выпадение участка хромосомы в средней ее части;
- дупликация — двух- или многократное повторение генов, локализованных в определенном участке хромосомы;
- инверсия — поворот участка хромосомы на 180°, в результате чего в этом участке гены расположены в последовательности, обратной по сравнению с обычной;
- транслокация — изменение положения какого-либо участка хромосомы в хромосомном наборе. К наиболее распространенному типу транслокаций относятся реципрокные, при которых происходит обмен участками между двумя негомологичными хромосомами. Участок хромосомы может изменить свое положение и без реципрокного обмена, оставаясь в той же хромосоме или включаясь в какую-то другую.
При дефишенси, делециях и дупликациях изменяется количество генетического материала. Степень фенотипического изменения зависит от того, насколько велики соответствующие участки хромосом и содержат ли они важные гены. Примеры дефишенси известны у многих организмов, включая человека. Тяжелое наследственное заболевание —синдром «кошачьего крика» (назван так по характеру звуков, издаваемых больными младенцами), обусловлен гетерозиготностью по дефишенси в 5-й хромосоме. Этот синдром сопровождается сильным нарушением роста и умственной отсталостью. Обычно дети с таким синдромом рано умирают, но некоторые доживают до зрелого возраста.
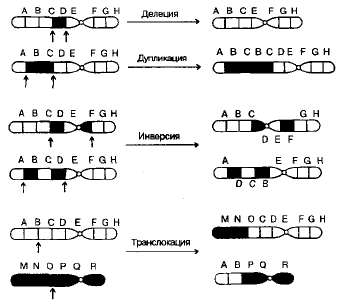
3.13. Хромосомные перестройки, изменяющие расположение генов в хромосомах.
Геномные мутации — изменение числа хромосом в геноме клеток организма. Это явление происходит в двух направлениях: в сторону увеличения числа целых гаплоидных наборов (полиплоидия) и в сторону потери или включения отдельных хромосом (анеуплоидия).
Полиплоидия — кратное увеличение гаплоидного набора хромосом. Клетки с разным числом гаплоидных наборов хромосом называются триплоидными (Зn), тетраплоидными (4n), гексанло-идными (6n), октаплоидными (8n) и т. д.
Чаще всего полиплоиды образуются при нарушении порядка расхождения хромосом к полюсам клетки при мейозе или митозе. Это может быть вызвано действием физических и химических факторов. Химические вещества типа колхицина подавляют образование митотического веретена в клетках, приступивших к делению, в результате чего удвоенные хромосомы не расходятся и клетка оказывается тетрагшоидной.Для многих растений известны так называемые полиплоидные ряды. Они включают формы от 2 до 10n и более. Например, полиплоидный ряд из наборов в 12, 24, 36, 48, 60, 72, 96, 108 и 144 хромосомы составляют представители рода паслен (Solanum). Род пшеница (Triticum) представляет ряд, члены которого имеют 34, 28 и 42 хромосомы.
Полиплоидия приводит к изменению признаков организма и поэтому является важным источником изменчивости в эволюции и селекции, особенно у растений. Это связано с тем, что у растительных организмов весьма широко распространены гермафродитизм (самоопыление), апомиксис (партеногенез) и вегетативное размножение. Поэтому около трети видов растений, распространенных на нашей планете, — полиплоиды, а в резко континентальных условиях высокогорного Памира произрастает до 85% полиплоидов. Почти все культурные растения тоже полиплоиды, у которых, в отличие от их диких сородичей, более крупные цветки, плоды и семена, а в запасающих органах (стебель, клубни) накапливается больше питательных веществ. Полиплоиды легче приспосабливаются к неблагоприятным условиям жизни, легче переносят низкие температуры и засуху. Именно поэтому они широко распространены в северных и высокогорных районах.
В основе резкого увеличения продуктивности полиплоидных форм культурных растений лежит явление полимерии (см. § 3.3).
Анеуплоидия, или гетероплодия, — явление, при котором клетки организма содержат измененное число хромосом, не кратное гаплоидному набору. Анеуплоиды возникают тогда, когда не расходятся или теряются отдельные гомологичные хромосомы в митозе и мейозе. В результате нерасхождения хромосом при гамето-генезе могут возникать половые клетки с лишними хромосомами, и тогда при последующем слиянии с нормальными гаплоидными гаметами они образуют зиготу 2n + 1 (трисомик) по определенной хромосоме. Если в гамете оказалось меньше на одну хромосому, то последующее оплодотворение приводит к образованию зиготы 1n - 1 (моносомик) по какой-либо из хромосом. Кроме того, встречаются формы 2n - 2, или нуллисомики, так как отсутствует пара гомологичных хромосом, и 2n + х, или полисомики.
Анеуплоиды встречаются как у растений и животных, так и у человека. Анеуплоидные растения обладают низкой жизнеспособностью и плодовитостью, а у человека это явление нередко приводит к бесплодию и в этих случаях не наследуется. У детей, родившихся от матерей старше 38 лет, вероятность анеуплоидии повышена (до 2,5%). Кроме того, случаи анеуплоидии у человека вызывают хромосомные болезни.
У раздельнополых животных как в естественных, так и в искусственных условиях полиплоидия встречается крайне редко. Это обусловлено тем, что полиплоидия, вызывая изменение соотношения половых хромосом и аутосом, приводит к нарушению конъюгации гомологичных хромосом и тем самым затрудняет определение пола. В результате такие формы оказываются бесплодными и маложизнеспособными.
Спонтанные и индуцированные мутации. Спонтанными называют мутации, возникающие под влиянием неизвестных природных факторов, чаще всего как результат ошибок при воспроизведении генетического материала (ДНК или РНК). Частота спонтанного мутирования у каждого вида генетически обусловлена и поддерживается на определенном уровне.
Индуцированный мутагенез — это искусственное получение мутаций с помощью физических и химических мутагенов. Резкое увеличение частоты мутаций (в сотни раз) происходит под воздействием всех видов ионизирующих излучений (гамма- и рентгеновские лучи, протоны, нейтроны и др.), ультрафиолетового излучения, высоких и низких температур. К химическим мутагенам относятся такие вещества, как формалин, азотистый иприт, колхицин, кофеин, некоторые компоненты табака, лекарственных препаратов, пищевых консервантов и пестицидов. Биологическими мутагенами являются вирусы и токсины ряда плесневых грибов.
В настоящее время ведутся работы по созданию методов направленного воздействия различных мутагенов на конкретные гены. Такие исследования очень важны, поскольку искусственное получение мутаций нужных генов может иметь большое практическое значение для селекции растений, животных и микроорганизмов.
Закон гомологических рядов в наследственной изменчивости. Крупнейшим обобщением работ по изучению изменчивости в начале XX в. стал закон гомологических рядов в наследственной изменчивости. Он был сформулирован выдающимся русским ученым Н. И. Вавиловым в 1920 г. Сущность закона заключается в следующем: виды и роды, генетически близкие, связанные друг с другом единством происхождения, характеризуются сходными рядами наследственной изменчивости. Зная, какие формы изменчивости встречаются у одного вида, можно предвидеть нахождение аналогичных форм у родственного ему вида.
В основе закона гомологических рядов фенотипической изменчивости у родственных видов лежит представление о единстве их происхождения от одного предка в процессе естественного отбора. Поскольку общие предки имели специфический набор генов, то их потомки должны обладать примерно таким же набором.
Более того, у родственных видов, имеющих общее происхождение, возникают и сходные мутации. Это означает, что у представителей разных семейств и классов растений и животных со сходным набором генов можно встретить параллелизм — гомологические ряды мутаций по морфологическим, физиологическим и биохимическим признакам и свойствам. Так, у разных классов позвоночных встречаются сходные мутации: альбинизм и отсутствие перьев у птиц, альбинизм и бесшерстность у млекопитающих, гемофилия у многих млекопитающих и человека. У растений наследственная изменчивость отмечена по таким признакам, как пленчатое или голое зерно, остистый или безостый колос и др.
Закон гомологических рядов, отражая общую закономерность мутационного процесса и формообразования организмов, представляет широкие возможности для его практического использования в сельскохозяйственном производстве, селекции, медицине. Знание характера изменчивости нескольких родственныхх видов дает возможность поиска признака, который отсутствует у одного из них, но характерен для других. Таким путем были собраны и изучены голозерные формы злаков, односемянные сорта сахарной свеклы, не нуждающиеся в прорывке, что особенно важно при механизированной обработке почв. Медицинская наука в качестве моделей для изучения болезней человека получила возможность использовать животных с гомологическими заболеваниями: это сахарный диабет крыс; врожденная глухота мышей, собак, морских свинок; катаракта глаз мышей, крыс, собак и др.
Закон гомологических рядов позволяет также предвидеть возможность появления мутаций, еще неизвестных науке, которые могут использоваться в селекции для создания новых ценных для хозяйства форм.
sbio.info
Изменчивость: наследственная и ненаследственная | Биология. Реферат, доклад, сообщение, краткое содержание, конспект, сочинение, ГДЗ, тест, книга
Вопрос 1. Какие виды изменчивости вам известны?
Существует два основных вида изменчивости — ненаследственная и наследственная.
Ненаследственная (фенотипическая или модификационная) изменчивость — это процесс появления новых признаков под влиянием факторов внешней среды, не затрагивающих генотип. В качестве примера можно привести дуб, листья которого в процессе развития приобрели разную площадь в зависимости от освещенности (маленькую — при яркой освещенности, большую — при слабой).
Наследственная изменчивость связана с изменениями генотипа; признаки и свойства, приобретенные вследствие этого, передаются следующим поколениям.
Существует два типа наследственной изменчивости — комбинативная и мутационная.
Комбинативная изменчивость заключается в появлении новых признаков в результате образования новых комбинаций генов родителей в генотипах потомков. Комбинативную изменчивость обеспечивают случайное расхождение гомологичных хромосом в мейозе, обмен участками гомологичных хромосом в профазе I мейоза, случайная встреча гамет при оплодотворении, случайный выбор родительских пар.
Мутационная изменчивость обусловлена изменениями генов и хромосом.
Вопрос 2. Что такое норма реакции?
Норма реакции (иначе — пределы модификационной изменчивости) — это пределы, в которых возможно изменение признака при определенном генотипе. Норма реакции может быть как очень широкой (вес человека), так и очень узкой (группа крови). Обычно узкой нормой реакции обладают признаки, обеспечивающие жизненно важные качества организма. Важно также то, что от родителей потомству передается не жестко запрограммированное значение того или иного признака, а его норма реакции.
Вопрос 3. Почему фенотипическая изменчивость не передается по наследству?
Фенотипическая изменчивость не затрагивает генотип, обеспечивая лишь то или иное проявление заложенных в нем признаков. Она обычно предсказуема и у разных особей одного вида проходит однонаправленно. Например, если пшеничное поле не получает достаточно влаги, то у всех его растений плохо формируется колос. Генотип у особей в этом случае остается неизменным, поэтому передачи информации о модификациях потомству не происходит. Следовательно, фенотипическая изменчивость не наследуется.
Вопрос 4. Что такое мутации? Охарактеризуйте основные свойства мутаций.
Мутации — это внезапные естественные или вызванные искусственно изменения генетического материала, приводящие к изменению тех или иных фенотипических признаков и свойств организма. Основные свойства мутаций:
- спонтанность — мутации возникают случайно;
- неспецифичность — могут возникать в любом участке генома;
- скачкообразность — вызывают новые качественные изменения;
- ненаправленность — возникшие изменения генотипа и фенотипа могут быть как биологически вредными, так и полезными.
Вопрос 5. Приведите классификацию мутаций по уровню изменений наследственного материала.
Различают три основных типа мутаций: Материал с сайта //iEssay.ru
- генные мутации вызывают изменения в отдельных генах, нарушая порядок и число нуклеотидов в цепи ДНК. Это приводит к синтезу измененного (как правило, дефектного) белка. Следствием генных мутаций являются такие заболевания, как фенилкетонурия и мышечная дистрофия Дюшена;
- хромосомные мутации затрагивают значительный участок хромосомы, вызывая нарушения сразу в нескольких (иногда — многих) генах. Описаны случаи потери участка хромосомы, его переворота, перемещения, удвоения и т. п.;
- геномные мутации приводят к изменению числа хромосом в кариотипе. Они возникают в результате нарушения расхождения гомологичных хромосом. Примером может служить синдром Дауна, который возникает при появлении лишней 21-й хромосомы. При этом общее число хромосом становится равным 47. Другим примером геномных мутаций является формирование полиплоидных растений (чаще всего тетраплоидных).
Вопрос 6. Назовите основные группы мутагенных факторов. Приведите примеры мутагенов, относящихся к каждой группе.
Мутагенные факторы можно разделить на три группы:
- физические мутагены — все типы ионизирующих излучений (у-лучи, рентгеновские лучи), ультрафиолетовое излучение, высокая и низкая температура;
- химические мутагены — аналоги нуклеиновых кислот, перекиси, соли тяжелых металлов (свинца, ртути), азотистая кислота, многие органические соединения;
- биологические мутагены — чужеродная ДНК и вирусы, которые, встраиваясь в ДНК хозяина, нарушают работу генов.
- объясните почему фенотипическая изменчивость не передается по наследству
- примеры наследственной изменсивости
- пример ненаследственной изменчивости у растений
- реферат на тему ненаследственная изменчивость
- тест наследственная и ненаследственная изменчивость
iessay.ru
Наследственная изменчивость
Наследственная изменчивость затрагивает генотип и передается по наследству. Она бывает комбинативной и мутационной.
Комбинативная изменчивость — появление новых сочетаний признаков вследствие перекомбинации генов. Основой комбинативной изменчивости является половой процесс; случайная комбинация негомологичных хромосом в мейозе и, как следствие, независимое наследование признаков; рекомбинация генов в результате кроссинговера. Комбинативная изменчивость определяет разнообразие особей и необходима для вида в его приспособлении к условиям среды.
В основе комбинативной изменчивости лежит половое размножение организмов, вследствие которого возникает огромное разнообразие генотипов. Практически неограниченными источниками генетической изменчивости служат три процесса:
► Независимое расхождение гомологичных хромосом в первом мейотическом делении. Именно независимое комбинирование хромосом при мейозе является основой третьего закона Менделя. Появление зеленых гладких и желтых морщинистых семян гороха во втором поколении от скрещивания растений с желтыми гладкими и зелеными морщинистыми семенами — пример комбинативной изменчивости.
► Взаимный обмен участками гомологичных хромосом, или кроссинговер. Он создает новые группы сцепления, т. е. служит важным источником генетической рекомбинации аллелей. Рекомбинантные хромосомы, оказавшись в зиготе, способствуют появлению признаков, нетипичных для каждого из родителей.
► Случайное сочетание гамет при оплодотворении.
Эти источники комбинативной изменчивости действуют независимо и одновременно, обеспечивая при этом постоянную «перетасовку» генов, что приводит к появлению организмов с другими генотипом и фенотипом (сами гены при этом не изменяются). Однако новые комбинации генов довольно легко распадаются при передаче из поколения в поколение.
Комбинативная изменчивость является важнейшим источником всего колоссального наследственного разнообразия, характерного для живых организмов. Однако перечисленные источники изменчивости не порождают существенных для выживания стабильных изменений в генотипе, которые необходимы, согласно эволюционной теории, для возникновения новых видов. Такие изменения возникают в результате мутаций.
Мутационная изменчивость — наследственные изменения генотипического материала хромосом и генов. Мутации имеют ряд характерных особенностей.
1. Затрагивают генотип и наследуются.
2. Носят скачкообразный и индивидуальный характер. Возникают у единичных особей в популяции.
3. Неадекватны условиям среды и могут быть нейтральными, полезными, чаще вредными.
4. Могут привести к образованию новых признаков, популяций или гибели организма.
В основе любых мутаций лежит появление новых типов белков.
Почти любое изменение в структуре или количестве хромосом, при котором клетка сохраняет способность к самовоспроизведению, обусловливает наследственное изменение признаков организма. По характеру изменения генома, т. е. совокупности генов, заключенных в гаплоидном наборе хромосом, различают генные, хромосомные и геномные мутации.
Генные, или точковые, мутации— результат изменения нуклеотидной последовательности в молекуле ДНК в пределах одного гена. Такое изменение в гене воспроизводится при транскрипции в структуре иРНК; оно приводит к изменению последовательности аминокислот в полипептидной цепи, образующейся при трансляции на рибосомах. В результате синтезируется другой белок, что ведет к изменению соответствующего признака организма. Это наиболее распространенный вид мутаций и важнейший источник наследственной изменчивости организмов.
Существуют разные типы генных мутаций, связанных с добавлением, выпадением или перестановкой нуклеотидов в гене. Это: - дупликации (повторение участка гена), - вставки (появление в последовательности лишней пары нуклеотидов), - делеции ("выпадение одной или более пар нуклеотидов), - замены нуклеотидных пар, - инверсии (переворот участка гена на 180°).
Эффекты генных мутаций чрезвычайно разнообразны. Большая часть из них фенотипически не проявляется, поскольку они рецессивны. Это очень важно для существования вида, так как в большинстве своем вновь возникающие мутации оказываются вредными. Однако их рецессивный характер позволяет им длительное время сохраняться у особей вида в гетерозиготном состоянии без вреда для организма и проявиться в будущем при переходе в гомозиготное состояние.
Хромосомные мутации (перестройки, или аберрации) — это изменения в структуре хромосом, которые можно выявить и изучить под световым микроскопом.
Известны перестройки разных типов:
- нехватка, или дефишенси, — потеря концевых участков хромосомы; - делеция — выпадение участка хромосомы в средней ее части; - дупликация — двух- или многократное повторение генов, локализованных в определенном участке хромосомы; - инверсия — поворот участка хромосомы на 180°, в результате чего в этом участке гены расположены в последовательности, обратной по сравнению с обычной; - транслокация — изменение положения какого-либо участка хромосомы в хромосомном наборе. К наиболее распространенному типу транслокаций относятся реципрокные, при которых происходит обмен участками между двумя негомологичными хромосомами. Участок хромосомы может изменить свое положение и без реципрокного обмена, оставаясь в той же хромосоме или включаясь в какую-то другую.
Геномные мутации — изменение числа хромосом в геноме клеток организма. Это явление происходит в двух направлениях: в сторону увеличения числа целых гаплоидных наборов (полиплоидия) и в сторону потери или включения отдельных хромосом (анеуплоидия).
Полиплоидия — кратное увеличение гаплоидного набора хромосом. Клетки с разным числом гаплоидных наборов хромосом называются триплоидными (Зn), тетраплоидными (4n), гексанло-идными (6n), октаплоидными (8n) и т. д.
Чаще всего полиплоиды образуются при нарушении порядка расхождения хромосом к полюсам клетки при мейозе или митозе. Это может быть вызвано действием физических и химических факторов. Химические вещества типа колхицина подавляют образование митотического веретена в клетках, приступивших к делению, в результате чего удвоенные хромосомы не расходятся и клетка оказывается тетрагшоидной.
Полиплоидия приводит к изменению признаков организма и поэтому является важным источником изменчивости в эволюции и селекции, особенно у растений. Это связано с тем, что у растительных организмов весьма широко распространены гермафродитизм (самоопыление), апомиксис (партеногенез) и вегетативное размножение. Поэтому около трети видов растений, распространенных на нашей планете, — полиплоиды, а в резко континентальных условиях высокогорного Памира произрастает до 85% полиплоидов. Почти все культурные растения тоже полиплоиды, у которых, в отличие от их диких сородичей, более крупные цветки, плоды и семена, а в запасающих органах (стебель, клубни) накапливается больше питательных веществ. Полиплоиды легче приспосабливаются к неблагоприятным условиям жизни, легче переносят низкие температуры и засуху. Именно поэтому они широко распространены в северных и высокогорных районах.
Анеуплоидия, или гетероплодия, — явление, при котором клетки организма содержат измененное число хромосом, не кратное гаплоидному набору. Анеуплоиды возникают тогда, когда не расходятся или теряются отдельные гомологичные хромосомы в митозе и мейозе. В результате нерасхождения хромосом при гаметогенезе могут возникать половые клетки с лишними хромосомами, и тогда при последующем слиянии с нормальными гаплоидными гаметами они образуют зиготу 2n + 1 (трисомик) по определенной хромосоме. Если в гамете оказалось меньше на одну хромосому, то последующее оплодотворение приводит к образованию зиготы 1n - 1 (моносомик) по какой-либо из хромосом. Кроме того, встречаются формы 2n - 2, или нуллисомики, так как отсутствует пара гомологичных хромосом, и 2n + х, или полисомики.
Анеуплоиды встречаются как у растений и животных, так и у человека. Анеуплоидные растения обладают низкой жизнеспособностью и плодовитостью, а у человека это явление нередко приводит к бесплодию и в этих случаях не наследуется. У детей, родившихся от матерей старше 38 лет, вероятность анеуплоидии повышена (до 2,5%). Кроме того, случаи анеуплоидии у человека вызывают хромосомные болезни.
biofile.ru
Наследственная изменчивость.
Поиск Лекций1.Ненаследственная изменчивость
1. Мутационная изменчивость: генная, геномная и хромосомная.
2. Комбинационная изменчивость.
3. Значение наследственной изменчивости в эволюции.
4. Методы изучения изменчивости растений
Ненаследственная изменчивость
Каждая особь с момента возникновения растет и развивается по определенной наследственной программе, т.е. в соответствии со своим генотипом. Но реализация этой наследственной программы, выражающаяся в признаках и свойствах конкретного фенотипа, во многом зависит от условий внешней среды.
Выше уже отмечалось, что наследственные возможности организма в процессе его жизни в определенных условиях никогда полностью не реализуется. Благодаря этому организм, не изменяя своей наследственности (генотипа) способен существовать при известных колебаниях условий внешней среды в относительно разнообразной обстановке. Но в каждом отдельном случае он будет приобретать специфический облик, определяемый разным характером реализации генотипа в различных условиях существования. Следовательно, здесь мы имеем дело с изменчивостью только фенотипа, генотип организма не изменяется. Поэтому такая изменчивость характерна только для конкретной особи и по наследству потомкам не передается.
Рассмотрим некоторые примеры. Если с тополя срезать два черенка и вырастить из одного саженец на бедной песчаной почве, а из другого на плодородной суглинистой почве то, несмотря на их, совершенно одинаковую наследственность, вследствие разных условий почвенного питания, рост и размеры этих деревцев будут разными. Теперь вновь срежем с них черенки одинакового диаметра и вырастим деревца в однородных почвенных условиях. Получим растения, которые мало чем будут отличаться друг от друга.
Таким образом, наблюдаемая в природе широкая внутривидовая изменчивость и индивидуальность во внешнем облике организмов объясняется в известной мере разнообразием условий их существования и вследствие этого разным характером реализации генотипов в конкретных фенотипах. Изменения в организмах, обусловленные только этими причинами, по наследству не передаются, так как наследственная программа остается неизменной. Такая изменчивость является ненаследственной, фенотипической или модификационной, а сами изменения носят название модификаций.
Вполне понятно, что эта форма изменчивости не может служить источником исходного материала для последующего естественного и искусственного отборов. Поэтому она имеет весьма ограниченное значение в эволюционном процессе и в селекции.
Наследственная изменчивость
Для эволюционного процесса имеет значение только наследственная изменчивость, которая, возникнув у отдельной особи, передается по наследству. Только благодаря такой изменчивости из примитивных одноклеточных организмов за два миллиарда лет на земле возник человек и окружающая его живая природа.
Чем же обуславливается наследственная изменчивость и почему она передается в ряду поколений?
Для того, чтобы разобраться с этим очень важным вопросом, нам еще раз нужно вспомнить, в чем сущность наследственности.
Отмечая исключительную роль белков в организме, мы говорим, что видовая специфичность, свойства, признаки и особенности развития особи определяется, прежде всего, тем, какие белки и в какой последовательности в ней синтезируются. Следовательно, наследственность в ряду поколений обеспечивается благодаря тому, что у потомков с момента зарождения и на протяжении всей их жизни белки синтезируются по той же программе, что и у их родителей.
Мы знаем, что программа синтеза белков содержится в каждой клетке живого существа, она записана на ДНК хромосом в строгой последовательности нуклеотидов и представлена совокупностью генов. Эта программа в процессе клеточного деления воспроизводится благодаря редупликации ДНК и передается в ряду поколений. В этом случае гены остаются постоянными, наследственность не изменяется, а изменчивость особей может быть только фенотипической, когда под влиянием изменившихся условий среды изменяется реализация генотипа.
Но так ли абсолютно устойчива структура ДНК и генов? Так ли постоянна наследственная программа? Оказывается, нет.
Установлены различные причины, вследствие которых происходит изменение и нарушение наследственной программы. Это прежде перестройка ДНК и хромосом. Достаточно измениться очередности нуклеотидов в ДНК хотя бы за счет одного из них, как это сразу же сказывается на белковом синтезе, что в свою очередь, вызовет изменения в организме. Все изменения в структуре ДНК и хромосом объединяются под общим названием мутации.Внезапное и скачкообразное изменение наследственной программы называется мутацией.
В зависимости от характера изменений, происходящих в хромосомах генотипа, мутации подразделяются на генные мутации, хромосомные мутации и геномные мутации.
В случае генных мутаций, которые называются также точковыми, изменения происходят в молекулярной структуре гена. Они вызываются нарушением очередности нуклеотидов в ДНК вследствие вставок, выпадения или замены отдельных нуклеотидов. В результате происходит изменение считывания наследственной программы с ДНК, что приводит к изменению очередности аминокислот или их состава в полипептидных цепочках белков и, в конечном итоге, к возникновению мутаций.
Генные мутации имеют наибольшее значение в эволюционном процессе и представляют большой интерес для селекции.
Хромосомные мутации обуславливаются перестройками хромосом и нарушением их структуры, происходящими обычно при клеточном делении.
В зависимости от характера возникающих перестроек различают нехватки, делеции, дупликации, инверсии и транслокации хромосом.
Нехватка - теряется концевая часть хромосомы и хромосома укорачивается.
Делеция – теряется средняя часть хромосомы.
Дупликация – происходит удвоение какого-либо участка хромосом.
Инверсия – хромосомы разрываются и срастаются вновь другими концами.
Транслокация – взаимный обмен частями негомологичных хромосом.
Изменчивость организмов при хромосомных перестройках в большинстве случаев связана не с нарушением структуры генов, а с изменением их взаимного расположение в хромосоме. Было установлено, что действие гена зависит от его положения в хромосоме и соседства с другими генами. Это явление было названо «эффектом положения» Перестройки хромосом вызывают существенные изменения наследственной программы, приводят к значительному нарушению белкового синтеза и часто гибели для организма. При незначительных перестройках хромосом у организма возникают новые свойства и признаки.
Многие исследователи считают хромосомные мутации наиболее широко распространенными, особенно при искусственном получении мутаций.
Геномные мутации – это изменения числа хромосом в клетке, возникающие чаще всего в результате нарушений клеточного деления. При этом может быть уменьшение или увеличение числа хромосом полными гаплоидными наборами и тогда возникают гаплоиды и полиплоиды, ил за счет отдельных хромосом в диплоидном наборе и образуются гетероплоиды.
Вызывая изменения наследственной программы, а следовательно и белкового синтеза, мутации приводят к изменчивости организмов, которая называется мутационной изменчивостью.
Все изменения, происходящие в структуре ДНК, копируются в ее новых молекулах при редупликации, которые передаются во вновь образующиеся соматические клетки и половые клетки. В результате потомство получает уже измененную наследственную программу. Вот поэтому изменения, возникшие в одном организме вследствие нарушения наследственной программы, наследуются потомками в ряду поколений.
Мутационная изменчивость характеризуются рядом свойств. Мутации: 1) могут происходить у любого организма, 2) на любой стадии его развития, 3) в различных тканях и клетках, 4) они возникают внезапно, без всяких переходов, 5) являются устойчивыми в ряду поколений.
Мутации происходят под влиянием как внешних, так и внутренних факторов. Однако мутации всегда случайны, разнонаправлены и не соответствуют факторам, их вызывавшим. Для организма они могут быть и полезными и вредными, но чаще всего они вредны, т.к. нарушают генный баланс, слаженность генотипа. Более того, мутации могут привести организм к гибели и тогда они называются летальными.
В естественных условиях мутации возникают редко, но поскольку каждый вид состоит из миллионов особей, они имеют большее значение для эволюции вида.
Факты мутационной изменчивости были известны уже давно. В частности Ч.Дарвин упоминает о некоторых случаях мутационной изменчивости. Однако первая серьезная работа в области изучения мутаций была выполнена лишь в конце Х1Х века голландским ученым де Фризом, изложенная впоследствии в его классическом труде «Мутационная теория». Кстати, термин «мутации» был предложен также де Фризом. Под мутацией он понимал явление внезапного скачкообразного изменения наследственного признака.
Таким образом, наследственная изменчивость обуславливается нарушением наследственной программы, изменением генотипа организма. Но только ли за счет мутаций изменяется генотип.
Изменчивость возникающая в процессе полового размножения называется комбинационной.
Этапы возникновения комбинационной изменчивости:
а профазе 1 в результате кроссинговера;
в анофазе 1 при независимом расхождении гамологичных хромосом каждой пары (материнских и отцовских) к различным полюсам клетки;
при оплодотворении может происходить случайное сочетание половых клеток.
При комбинационной изменчивости происходит новая комбинация генов. Сами гены, их молекулярная структура не изменяются. Изменяются лишь их сочетания и характер взаимодействия в генотипе
Комбинационная изменчивость связана только с новыми комбинациями и рекомбинациями генов, и дает огромное разнообразие форм
Генные мутации создают новые наследственные единицы-гены и, тем самым представляет естественному отбору исходный материал. Именно генные мутации вызывают ту самую неопределенную изменчивость, которой Дарвин придавал главное значение в эволюции
Естественный отбор оценивает качество мутаций. Он сохраняет те формы, которые в результате мутаций оказались более приспособленными к данным условиям и уничтожает формы с мутациями, снижающими их приспособленность
Методы изучения изменчивости.
В основу методов изучения генетической изменчивости положено определение степени влияния наследственности и среды в проявлении фенотипа. Наследственная изменчивость всегда проявляется в фенотипической форме, т.е. в сложном взаимодействии генотипов и среды обитания. Применительно к селекционной практике методы изучения различных форм внутривидовой изменчивости должны быть направлены на то, чтобы определить, что в данном фенотипе или группе фенотипов, обусловлено внешней средой и что влиянием факторов наследственности. При изучении внутривидовой изменчивости применяются статистические методы обработки количественных признаков отдельных выборок групп особей, относящихся к разным видам, подвидам или сортам. Примером этого может служить использование коэффициента вариации С, % при сравнительном изучении изменчивости. Для характеристики вариационных рядов у древесных растений широко применяется шкала уровней изменчивости признаков , составленная С.А. Мамаевым.
Уровень изменчивости: очень высокий - С, % -более 40; высокий - С, % -21-40; средний - С, % - 13-20; низкий - С, % -7-12; очень низкий - С, % - менее 7.
Исследование изменчивости древесных растений проводятся последовательно в три этапа.
На первом этапе дается оценка характера и степени варьирования признаков в пределах организма, т.е. дается характеристика эндогенной изменчивости.
На втором этапе производится оценка различных форм внутривидовой изменчивости - индивидуальная, половая экологическая.
На третьем этапе изучается межпопуляционная изменчивость, к которой относится географическая изменчивость.
Математические методы изучения наследственной изменчивости древесных растений направлены на определение наследственных и ненаследственных факторов в изменчивости среднего уровня признака. Для этой цели часто используется дисперсионный анализ. При использовании дисперсионного анализа в целях анализа изменчивости определяется суммарное (аддитивное) действие случайных и факториальных варианс на общую вариансу и выражается уравнением

 ,
,
где  - общая фенотипическая варианса,
- общая фенотипическая варианса,  - наследственная (генотипическая) варианса, характеризующая генетическую изменчивость,
- наследственная (генотипическая) варианса, характеризующая генетическую изменчивость,  - ненаследственная варианса, которая характеризует изменения, вызванные внешними условиями.
- ненаследственная варианса, которая характеризует изменения, вызванные внешними условиями.
Для характеристики доли генетической изменчивости в общей фенотипической пользуются не абсолютной величиной генетической вариансы, а относительной, как показателем удельного веса. Этот показатель называется коэффициентом наследуемости Н2.

Наиболее простой и быстрый способ определения меры генетической изменчивости в отдельной выборке - это метод испытания клонового потомства. В разных условиях высаживают вегетативно размноженные деревья (клоны). Изменчивость признака между клонами разных деревьев определяют генетическую вариансу , а изменчивость клонового потомства в разных условиях дает паратипическую или ненаследственную вариансу .
poisk-ru.ru
Доказательство на примере культурных растений семейства Злаковые закона гомологических рядов в наследственной изменчивости Н.И. Вавилова
Доказательство на примере культурных растений семейства Злаковые закона гомологических рядов в наследственной изменчивости Н.И. Вавилова
«Виды и роды, генетически близкие, характеризуются сходными рядами наследственной изменчивости с такой правильностью, что, зная ряд форм в пределах одного вида, можно предвидеть нахождение параллельных форм у других видов и родов. Чем ближе генетически расположены в общей системе роды и виды, тем полнее сходство в рядах их изменчивости. Целые семейства растений в общем характеризуются определенным циклом изменчивости, проходящей через все роды и виды, составляющие семейство».
Зако́н гомологи́ческих рядов — закон наследственной изменчивости, сформулированный Н.И. Вавиловым в 1920 году. Закон устанавливает параллелизм в изменчивости родственных групп растений, в основе которого лежит гомология генов (их одинаковое молекулярное строение) и гомология (сходство) в порядке их расположения в хромосомах у родственных видов.
Н.И. Вавилов обнародовал этот закон в докладе «Закон гомологических рядов в наследственной изменчивости» в день открытия в Саратове III Всероссийского селекционного съезда 4 июня 1920 года. Позднее ботаник-физиолог профессор Заленский сказал о законе Вавилова:
Главная ценность закона гомологических рядов в наследственной изменчивости состоит в его прогностической сущности, позволяющей вести целенаправленный поиск гендоноров адаптивно значимых и хозяйственно ценных признаков.
На основе закона растениеводы и животноводы могут целенаправленно искать и находить нужные признаки и варианты у различных видов в мировом многообразии форм как культурных растений и домашних животных, так и у их диких родичей. Например, если черная окраска зерна встречается у ржи, пшеницы и других злаков, то растения с такой же окраской зерна могут быть получены у овса и проса. Так Н.И. Вавилов предсказал наличие безалкалоидного (неядовитого) люпина, учитывая существование безалкалоидных форм в родах гороха, фасоли и других растений семейства бобовых.
Цель: влияние климатических факторов (солнечный свет и вода) на злаковые культуры, произрастающие на одном участке (биотоп).
Задачи: наблюдать за воздействием абиотических факторов за культурами одного семейства.
Гипотеза: если на культуры одного семейства (злаковые) воздействуют одинаковые абиотические факторы, то результативность будет одинаковой.
В исследовании использовали 4 вида злаковых растений: рожь, ячмень, овес, пшеница.
Наблюдение:Первые всходы
17.05-20.05
В зависимости от вида
-Зерновка овса, словно панцирем, плотно прикрыта цветковыми чешуями, и они оказывают воздействие на ход прорастания:появляющееся колеоптиле не в силах пробить этот пленчатый панцирь и проходит вдоль него, пока не найдет выход у верхушки зерновки, где между соприкасающимися чешуями имеется щель; выйдя из нее, оно направляется кверху.
Продолжающее расти колеоптиле пронзает почву и выходит на дневную поверхность;
-Ячмень:начинается с прорастания семян, заканчивается появлением всходов и образованием третьего листа. Общая продолжительность ее 15-30 дней.
Вначале развивается главный побег и в пазухах зародышевых листьев закладываются почки боковых побегов. Главный зародышевый корешок к концу фазы всходов проникает в почву на глубину до 40 см.
-Пшеница: При температуре почвы на глубине заделки семян 15 °С всходы появляются на 7-ой день (19.05)
-Рожь: для прорастания зародышевых корней требуется 3,8°С, а для появления первого листа требуется более высокая температура. Первым трогается в рост главный корешок и углубляясь в почву поглощает воду и минеральные вещества почвенного раствора. Всходы на 6-8 день.
Кущение: Кущение - это биологический процесс, имеющий важное значение в жизни растений, так как в фазу кущения закладываются органы, определяющие урожай.
С 20.05
-У ржи кущение начинается, когда образуется 4-й лист. В это время появляется первый боковой побег из почки в пазухе первого зародышевого листа. Затем появляются следующие побеги из почек в пазухах следующих зародышевых листьев. Одновременно развиваются узловые корни. Каждый новый побег имеет свои корни. Таким путем формируется узел кущения, который располагается близко к поверхности почвы. Узел кущения - важная часть растения, так как в нем формируются боковые побеги и узловые корни. Степень кущения зависит от многих факторов - плодородия почвы, погодных условий, сроков посева, норм высева и глубины заделки семян, а также от биологических особенностей сорта. Кущение продолжается 35-37 дней.
-Кущение яровой пшеницы лучше проходит при температуре 10-12°С.
Недостаток влаги в почве в этот период увеличивает количество бесплодных колосков
-овес:Эта фаза начинается обычно через 10- 15 дней после появления всходов, в момент развития 3- 4-го листа. Число всех стеблей на одно растение обозначается как общая кустистость, а число стеблей с нормально развитой метелкой - как продуктивная. Последняя обычно составляет 2-4 стебля. В разреженных посевах кустистость возрастает. Сорта голозерного овса кустятся сильнее пленчатого. В фазе полного кущения у сортов овса различают следующие формы куста: прямостоячую (сомкнутую), распластанную (лежачую) и промежуточную.
- Кустится ячмень сильнее, чем яровая пшеница и овес. При появлении третьего листа ячмень начинает куститься, то есть образовывать боковые побеги. Главный узел кущения расположен в почве на глубине 1 — 3 см, важно, чтобы узел кущения находился в увлажненном слое почвы.
выход в трубку
С 10-24.06
- Ячмень сильно страдает от быстрого наступления высокой температуры в фазе выхода в трубку, когда формируется продуктивность колоса. В период выхода в трубку — колошения наиболее благоприятна среднесуточная температура 20—22°С, Фаза выхода в трубку наступает примерно через 3—4 недели после появления полных всходов.
- пшеница:Период от выхода в трубку до колошения — очень важный этап в развитии зерновых хлебов. В это время усиленно растут листья и соломина, формируется колос. Растение испытывает повышенную потребность во влаге и питательных веществах. В фазе выхода в трубку у яровой пшеницы можно легко различить узлы, междоузлия, язычки и ушки. Стеблевой узел представляет перехват на тех частях стебля, где имеются сплошные перегородки. Междоузлием является часть стебля между двумя узлами. Междоузлия бывают полые, реже выполненные вверху рыхлой тканью. Над стеблевым узлом располагается листовой узел — небольшое кольцевое утолщение, с помощью которого лист прикреплен к стеблю. Лист плотно охватывает междоузлие, что придает стеблю прочность и защищает его нежные растущие части от внешних повреждений. В месте перехода листового влагалища в листовую пластинку находится язычок, представляющий тонкую пленку, прилегающую к стеблю. Около язычка по краям листового влагалища помещаются два полулунных рожка, или ушка, которые способствуют закреплению влагалища листа на стебле.
- Овес:Начинается фаза через 10-15 дней после кущения и означает начало образования соломины. На практике можно определить прощупыванием узла на стебле от поверхности почвы. Обычно с этого момента начинается быстрый рост надземных органов и корней, который продолжается до начала цветения; позднее процессы роста замедляются и постепенно затухают. После выхода в трубку появляются органы полового размножения - цветки, собранные в колоски и в соцветие - метелку.
-Рожь: Вытягивание первого междоузлия на 5-6 см и появление нижнего стеблевого узла на поверхности почвы принято считать началом фазы выхода в трубку. С этого момента начинается быстрый рост стеблей и развитие колоса, заключенного внутри трубки листовых влагалищ. В это время заканчивается образование основной массы листьев. К концу фазы растения достигают высоты, соответствующей особенностям данного сорта.
колошение
С 10.07
- Рожь: Фаза отмечается при выдвижении колоса из влагалища верхнего листа. Листья раздвигаются в результате вытягивания междоузлий, листья увеличиваются в размере. Стебель укрепляется механически и выдерживает увеличивающуюся массу листьев и колоса. Колошение начинается в среднем через 40-50 дней после весеннего возобновления вегетации. Период от кущения до колошения у яровой ржи- 25-30 дней.
С 10.07
-Овес: определяют по появлению первого колоска из влагалища верхнего листа.
С 15.07
-Пшеница: Фаза колошения начинается выходом из влагалища последнего листа. Колошение у яровой пшеницы наступает через 50-60 дней после посева и длится 10-12 дней. Эта фаза продолжается 1-4 дня в зависимости от сорта и погодных условий. Наиболее благоприятная температура в этот период 20-25 ºС. В период выхода в трубку и колошения происходит самый интенсивный рост вегетативной массы растений, а также расходуется большое количество влаги (50-60 % потребляемого количества за вегетацию)
С 13.07
-Ячмень: Фаза колошения наступает, когда колос наполовину выходит из влагалища листа. Обычно это случается на 38-42 день после появления всходов. К наступлению колошения ячмень полностью сформировывает генеративные органы— пыльники и пестик с рыльцами.
цветение
С 13.07
-Рожь: При созревании генеративных органов цветковые чешуи раздвигаются вследствии набухания лодикул. При этом наружу выбрасываются тычинки и рыльце пестика. В каждом пыльнике у ржи до 20 тыс. пыльцевых зерен.. Цветение яровой ржи наступает через 3-6 дней после колошения и продолжается 6-7 дней. Цветение начинается со средней части колоса. Пыльца переносится ветром на рыльце другого цветка и прорастает, проникает в завязь и сливается с яйцеклеткой. На рыльце своего цветка пыльца плохо прорастает. Для цветения требуется 12,5°С. Цветение каждого цветка продолжается 12-25 минут.
С 15.07
-Овес: Овес считается самоопылителем, однако у гексаплоидных видов нередки случаи перекрестного опыления. Цветение овса начинается одновременно с выходом метелки из влагалища с растрескиванием пыльников самых верхних ее колосков и концов отдельных веточек. Затем цветение последовательно переходит к основанию веточек и мутовок метелки. В колоске оно начинается с нижнего цветка и идет в восходящем порядке, поэтому колоски в метелке разновозрастные: если в нижней части метелки еще наблюдается цветение, в верхушечных колосках происходит налив зерна.
С 25.07
-Пшеница: Цветение яровой пшеницы протекает интенсивнее в утренние и вечерние часы. Продолжительность цветения одного колоса 3-5 дней, а всего поля обычно 5-7 дней. Яровая пшеница относится к самоопыляющимся культурам, но не исключено и перекрестное опыление. Недостаточная влажность и повышенная температура воздуха в это время снижают степень оплодотворения цветков, что уменьшает число зерен в колосе
С 20.07
-Ячмень: Цветение — важный период в жизни растений, так как к концу его прекращается рост вегетативных органов, а в большинстве случаев — и рост корневой системы. Ячмень — самоопыляющееся растение, однако в ряде случаев опыляется и перекрестно. В каждом развитом цветке у него находятся мужские и женские органы. Цветение ячменя чаще всего совпадает с началом колошения и реже (через 1—3 дня) после него. В засушливые годы ячмень цветет во влагалище листа.
Цветение начинается со средних колосков и одновременно распространяется на верхние и нижние части колоса. Наиболее интенсивное цветение и оплодотворение наблюдается в утренние часы. Весь процесс оплодотворения продолжается 6—8 ч.
созревание
С 20.05
-Рожь:Поскольку колошение происходит не одновременно, то зерно от разных побегов формируется при различных условиях. В пределах каждого колоса зерно также не однородно. Кроме внешних условий на зерно влияют внутренние условия, поэтому зерно отличается по морфологии, биохимии. Скорость созревания зерновки зависит от погодных условий. Сильное влияние оказывают ветры, температура воздуха и влажность. При молочной спелости зерно имеет зеленую окраску, содержит 60-40% воды, в которой растворены органические вещества. При восковой спелости растение желтеет. Зерно становится желтым, имеет консистенцию воска. В эту фазу зерно отделяется от материнского растения. Влаги в зерне 20-40%. При полной спелости растение полностью желтеет, зерно твердеет, имеет 14-16% влаги. В период созревания в зерне увеличивается содержание крахмала, в стебле уменьшается содержание сахара. В зерне до конца молочной спелости увеличивается содержание азота. В начале созревания в зерне активны различные ферменты, но в стадии восковой спелости зерно устойчиво на влияние ферментов. При полной спелости зерно имеет слабую ферментативную активность. В молочной и восковой спелости в зерне много аскорбиновой кислоты, каротина и каротиноидов. При полной спелости их количество резко уменьшается. В период восковой спелости в зерновке накапливаются все жизненно необходимые вещества, обеспечивающие нормальную всхожесть.
С 22.07
-Овес: При послеуборочном дозревании в зерне протекают сложные биохимические процессы. Содержание азотистых веществ и крахмала в зерне по мере хранения увеличивается, количество сахаров уменьшается. Семена при хранении от - 1 до 25 °С имеют малорастворимые белки, которые распадаются до аминокислот, тормозится дыхание. При покое у семян ржи влажность должна быть 10%. Если влажность увеличивается дыхание усиливается, что приводит к самосогреванию зерна.
С 28.07
-пшеница: созревание зерна наступает после оплодотворения завязи, когда начинается приток в нее питательных веществ и постепенное ее разрастание. Различают три фазы созревания: молочную, восковую и полное созревание.
С 22.07
-Ячмень: Формирование зерна у ярового ячменя продолжается 10-11 дней. В конце этого этапа зерно достигает окончательной длины, но остается щуплым. Содержание влаги в нем снижается с 80 до 65%. Растение и зерно имеют зеленый цвет. Налив зерна длится 11-14 дней, от начала молочной до восковой спелости. Во время налива происходит максимальное накопление в зерне минеральных и органических веществ, при снижении влаги до 40%. При сдавливании зерна в молочной спелости выделяется белая жидкость.
Сбор семян
-Рожь: При послеуборочном дозревании в зерне протекают сложные биохимические процессы. Содержание азотистых веществ и крахмала в зерне по мере хранения увеличивается, количество сахаров уменьшается. Семена при хранении от - 1 до 25 °С имеют малорастворимые белки, которые распадаются до аминокислот, тормозится дыхание. При покое у семян ржи влажность должна быть 10%. Если влажность увеличивается дыхание усиливается, что приводит к самосогреванию зерна.
- Овес:
-Пшеница: Молочная спелость наступает через 8-18 дней после начала цветения. В эту фазу зерно достигает нормальной длины, при надавливании из него выступает белая, густой консистенции жидкость. Влажность зерна при этом высокая – 72-47 %.
Восковая спелость наступает через 10-14 дней после молочной. Зерно приобретает желтоватую окраску, его содержимое, как воск, хорошо режется ногтем. В этой фазе спелости в зерновке содержится 32-25 % воды.
Полная спелость характеризуется потерей воды в зерне до 18-15 %, оно приобретает характерную для сорта окраску, твердость, ногтем не режется.
-Ячмень:
Подводя итоги рассмотренным закономерностям, мы приходим к следующим положениям.
Виды и роды, генетически близкие, характеризуются сходными рядами наследственной изменчивости с такой правильностью, что, зная ряд форм в пределах одного вида, можно предвидеть нахождение параллельных форм у других видов и родов. Чем ближе генетически расположены в общей системе роды и линнеоны, тем полнее сходство в рядах их изменчивости.
Гомологическая изменчивость обусловлена генетическим родством видов и родов, проявляется лучше всего в переделах одного семейства, тогда как в аналогичной и общей изменчивости генетическое родство не играет роли.
Позже было установлено, что этот закон распространяется на животные и микроорганизмы. Так, альбинизм встречается у всех классов позвоночных животных, коротколапость наблюдается у всех пород крупного рогатого скота, овец и собак.
Подготовка почвы к посадке 


Посадка семян и полив



Первые всходы и кущение


Созревание





infourok.ru










