
Исследовательская работа Анатомические и морфологические особенности растений как основа, определяющая декоративность. Морфологические особенности растений
Исследовательская работа Анатомические и морфологические особенности растений как основа, определяющая декоративность
Министерство образования РЕСПУБЛИКи БАШКОРТОСТАН
казённое учреждение «управление образования»
ГОРОДСКОГО ОКРУГА ГОРОД КУМЕРТАУ РЕСПУБЛИКИ БАШКОРТОСТАН
Муниципальное бюджетное общеобразовательное учреждение
Гимназия № 1 им. Н.Т. Антошкина
городского округа город Кумертау Республики Башкортостан
выполнил: Толстов Артем
ученик 7а класса,
МБОУ Гимназии №1
Научный руководитель:
Янсиярова Гульназ Флюровна,
учитель биологии
МБОУ Гимназии №1
КУМЕРТАУ
2017
Оглавление
Введение………………………………………………………………………..2
-
История изучения………………………………………………………..3
-
Законы красоты………………………………………………………….3
-
Зрительное восприятие………………………………………………….6
-
Методика исследования…………………………………………………7
-
Гистологические особенности комнатных растений……………………8
-
Морфологические особенности растений ………………………………8
-
Значение цветов в эволюции живых организмов…………..…….………8
-
Выводы………………………………………………………….………9
-
Список литературы……………………………………………..…….….10
-
Приложение………………...…………………………………...……….11
ВВЕДЕНИЕ
Актуальность исследования. Красота природы очевидна. Цветы принято дарить, восхищаться ими. Они сопровождают нас всю жизнь, наполняя мгновения радостью и умиротворением. Все народы мира воспринимают, осознают и восхищаются красотой цветов, но механизм восприятия остается не до конца изученным и вызывает множество вопросов. Почему цветы красивые для нас? В чем их секрет?
Объект следования: комнатные и дикорастущие растения.
Цель исследования: выяснить, почему те растения, которые нас окружают, воспринимаются нами красивыми.
Задачи исследования:
-
Выяснить, что определяется в качестве красивого при восприятии окружающих человека растений и был ли до нас уже исследован этот вопрос?
-
Существуют ли законы красоты растений ?
Оборудование и программное обеспечение:
Фотоаппарат, микроскоп Альтами, видеофотокамера Альтами, цифровой USB микроскоп со встроенной камерой, биохимическая лаборатория; программа для создания, хранения и записи данных электронных микроскопов Digital Imager, программа обработки изображений Point.
История изучения
Одно из первых описаний тонкой структуры растений было дано в книге английского ученого Роберта Гука «Микрография или некоторые физиологические описания мельчайших телец при помощи увеличительных стекол» (1665). Гук описал некоторые растительные ткани и заметил их клеточное строение. Итальянский ученый М. Мальпиги во второй половине XVII в. тщательно описал микроструктуру листьев, стеблей и корней. Особенно детально он изучил строение стебля (коры, древесины и сердцевины). Почти одновременно с Мальпиги исследовал строение растений и английский натуралист Неэмия Грю, автор «Анатомии растений» (1682). Он сделал много тонких и тщательных наблюдений, установил понятие «ткань», описал строение разных тканей растения (Королькова, 2008; Серебрякова, 2006).
Большинство исследований растений длительное время не затрагивало тему «красоты цветка», а было направлено на строение, лекарственные свойства, физиологические процессы. Много книг посвящено систематике растений. В наше время, в связи с развитием науки и техники, мы можем уверенно сказать, что красота цветов, трав, деревьев, кустарников - вполне закономерный, длительно эволюционно формировавшийся процесс (Еленевский,2006; Иллюстрированное…, 2004; Иллюстрированный, 2002-2004; Маевский, 2006; Миркин, 2002; Определитель, 1995).
Законы красоты
Множество примеров воспевания прекрасного в окружающем мире встречается в литературе (С. Маршак «Ландыш», И. Северянин «Поэма о незабудках», И. Бунин «Полевые цветы», С. Смирнова-Понамарёва «Шиповник», В. Жуковский «О дивной розе…»Л. Кузьминская «Фиалки», В. Ивченко «Васильки», И. Виноградский «Крокусы», Вс. Рождественский «Астры»), живописи (Винсент Ван Гог «Ваза с розовыми цветами», Альбрехт Дюрер «Пучок первоцветов», Э. Мане «Голубые водяные лилии», «Сирень в вазе», Энди Уорхол «Цветы»), музыке (Антонов Юрий «Сирень», Данцевич Александр «Черёмуха», Зыкина Людмила «Тонкая рябина», Кадышева Надежда «Подари берёзка», Казаченко Вадим «Белая акация», Ротару София «Лаванда» и др.).
Наблюдая за исследуемыми объектами, мы заметили, что в глаза бросается интересная форма растений. Если речь идет о деревьях и кустарниках, принято говорить о форме кроны, травянистые многолетники "щеголяют" формой куста. Так, разные сорта рябины могут "похвастаться" яйцевидными, пирамидальными и плакучими кронами, клены - раскидистыми и шаровидными, сосны - зонтиковидными, подушковидными и пирамидальными (art-ginkgo.com). Среди травянистых многолетников есть немало видов и сортов с компактными кустами, привлекательные не только в цвету. Плотные кроны (у многих хвойных) придают композициям основательность, прочность, прозрачные, кружевные, (сосна, береза) - воздушность и легкость. Необыкновенно привлекательна графика ветвления побегов. Размер листьев определяет характер поверхности кроны растения - ее фактуру. У видов с крупными листьями она рыхлая или грубофактурная. Грубая фактура кроны интересна сама по себе, так же как и бархатистая, мелкофактурная поверхность крон некоторых хвойных. Бархатистая фактура кроны ценится в качестве наиболее выигрышного фона, на котором выступают мельчайшие детали строения других растений, оттенков цвета (art-ginkgo.com). Есть своя собственная фактура поверхности и у листа - глянцевая, матовая, рельефная от сети выступающих или вдавленных жилок, кожистая и плотная… На первый взгляд все листья зеленые. Однако разнообразие оттенков (текстуры листьев) этого цвета в природе так же велико, как и разнообразие самих растений. У растений, приспособленных к яркому солнечному свету, листва или хвоя имеет сизый, голубой оттенок. Лесные виды, приспособленные к недостатку света, частенько "предпочитают" желто-зеленый оттенок зелени. Сезонная расцветка листьев - одно из красивейших явлений в растительном мире, по масштабу и колориту значительно превосходящее цветение. Не только осенью, но уже ранней весной можно любоваться разнообразными оттенками листвы. Если есть в разнообразии растительных форм та, в которой природа не знает ограничений,- это, несомненно, форма листа. Существуют композиционные приемы, универсальные для всех видов искусств. Их применяют и для составления растительных композиций. В составлении групп из растений используют прием контраста - сочетание противоположных признаков (светлое и темное, "тяжелое" и "легкое", вертикаль и горизонталь). А в результате эти признаки зрительно усиливаются. Если контраст основан на различиях, то нюанс - на сходстве. Нюанс позволяет обыгрывать полутона и оттенки, небольшую разницу в форме, размерах листьев и цветков, графике ветвления. Метод повтора одного и того же (ритм) – повторяющиеся одинакового размера, цвета и формы части растений или целые растения( http://home.onego.ru/~otsoppe/book48.htm ).
Большинство попыток к пониманию законов красоты связано с математическими исследованиями (Зенкевич, 2005; Коксетер, 2003; Демьянов, 1986) (рис. 8). Симметрия - закономерное расположение подобных частей тела или форм живого организма, совокупности живых организмов относительно центра или оси симметрии. Асимметрия — отсутствие симметрии. Иногда этот термин используется для описания организмов, лишённых симметрии первично, в противоположность диссимметрии — вторичной утрате симметрии или отдельных её элементов. Понятия симметрии и асимметрии альтернативны. Чем более симметричен организм, тем менее он асимметричен и наоборот. В природе и, в частности, в живой природе симметрия не абсолютна и всегда содержит некоторую степень асимметрии.
Среди элементов симметрии различают следующие:
-
плоскость симметрии — плоскость, делящая объект на две равные половины;
-
ось симметрии — прямая линия, при повороте вокруг которой на некоторых угол, меньший 360, объект совпадает сам с собой;
-
центр симметрии — точка. делящая пополам все прямые линии, соединяющие подобные точки объекта.
Обычно через центр симметрии проходят оси симметрии, а через ось симметрии — плоскости симметрии, однако существуют тела и фигуры, у которых при наличии центра симметрии нет ни осей, ни плоскостей симметрии, а при наличии оси симметрии отсутствуют плоскости симметрии.
У биологических объектов встречаются следующие типы симметрии: 1. сферическая симметрия — симметричность относительно вращений в трёхмерном пространстве на произвольные углы, 2.аксиальная симметрия — симметричность относительно поворотов на произвольный угол вокруг какой-либо оси 3. симметрия вращения 4. двусторонняя симметрия — симметричность относительно плоскости симметрии, 5. трансляционная симметрия — симметричность относительно сдвигов пространства в каком-либо направлении на некоторое расстояние), 6. триаксиальная асимметрия — отсутствие симметрии по всем трём пространственным осям, 7. Существует и другая классификация симметрии, отражающая эволюцию растений (http://www.muldyr.ru/a/a/simmetriya_biologiya_-_tipyi_simmetriy) (прил.2).
В любом растении можно найти какую-то его часть обладающую осевой (двусторонней), центральной (лучевой) или винтовой симметрией (реже выделяются другие виды симметрии). Центральная (лучевой) симметрия наиболее характерна для плодов растений и некоторых цветов. Рассмотрим разрез любой ягоды. В разрезе она представляет собой окружность, а окружность имеет центр симметрии. Для цветов характерна и поворотная симметрия. Цветок можно, например, повернуть вокруг некоторой прямой на угол, равный 360о/5 (или кратный ему), и он совместится сам с собой. Осевая (двусторонняя) характерна для листьев, семян, некоторых плодов и др. Стебель растения обладает винтовой осью симметрии. У подсолнечника каждый листок появляется после поворота на 72о. Листья на стебле располагаются по спирали так, чтобы, чтобы, не мешая друг другу, воспринимать солнечный цвет. Симметрия форм цветов придаёт им красоту. В подавляющем большинстве случаев листья могут быть разделены на две более или менее симметричные половинки, т.е. они обладают одной плоскостью симметрии, проходящей вдоль черешка плоскостью симметрии, проходящей вдоль черешка и по середине пластинки. Сложный лист отличается от простого тем, что на черешке имеется несколько пластинок. Несмотря на их разнообразие, все они обладают одной плоскостью симметрии. Листья также подразделяются на «правшей» и «левшей». (Шафрановский, 1985; Заренков, 2009).
Зрительное восприятие
Большое значение в восприятии изображения растений имеет зрение, представляющее собой процесс психофизиологической обработки изображения объектов окружающего мира, осуществляемый зрительной системой, и позволяющий получать представление о величине, форме (перспективе) и цвете предметов, их взаимном расположении и расстоянии между ними. Из-за большого числа этапов процесса зрительного восприятия его отдельные характеристики рассматриваются с точки зрения разных наук — оптики (в том числе биофизики), психологии, физиологии, химии (биохимии) (прил. 3).
В глазу человека содержатся два типа светочувствительных клеток (фоторецепторов): высокочувствительные палочки и менее чувствительные колбочки. Палочки функционируют в условиях относительно низкой освещённости и отвечают за действие механизма ночного зрения, однако при этом они обеспечивают только нейтральное в цветовом отношении восприятие действительности, ограниченное участием белого, серого и чёрного цветов. Колбочки работают при более высоких уровнях освещённости, чем палочки. Они ответственны за механизм дневного зрения, отличительной особенностью которого является способность обеспечения цветового зрения.
Возникает вопрос: только ли люди видят красоту окружающих растений? Оказывается, приматов (в том числе и человека) мутация вызвала появление дополнительного типа колбочек — цветовых рецепторов. Это было вызвано расширением экологической ниши млекопитающих, переходом части видов к дневному образу жизни, в том числе на деревьях. Мутация была вызвана появлением изменённой копии гена, отвечающего за восприятие средней, зелёночувствительной области спектра. Она обеспечила лучшее распознавание объектов «дневного мира» — плодов, цветов, листьев. В сетчатке глаза человека есть три вида колбочек, максимумы чувствительности которых приходятся на красный, зелёный и синий участки спектра(прил. 3). Ещё в 1970-х годах было показано, что распределение типов колбочек в сетчатке неравномерно: «синие» колбочки находятся ближе к периферии, в то время как «красные» и «зеленые» распределены случайным образом, что было подтверждено более детальными исследованиями в начале XXI века. Соответствие типов колбочек трём «основным» цветам обеспечивает распознавание тысяч цветов и оттенков. Кривые спектральной чувствительности трёх видов колбочек частично перекрываются, что способствует явлению метамерии. Очень сильный свет возбуждает все 3 типа рецепторов, и потому воспринимается, как излучение слепяще-белого цвета (эффект метамерии лежит в основе восприятия белых лепестков)(Пэдхем, 1978).
МЕТОДЫ ИССЛЕДОВАНИЯ
Мною исследованы растения цветочных магазинов, домашние, школьные и дикорастущие формы улиц, садов, парков и скверов. Работа выполнялась периодически в течение 1 года. Нами созданы коллекции снимков и гербарные материалы. Наиболее важные с точки зрения исследования практические результаты и материалы мы приводим в работе. Нами использовался оптический микроскоп Альтами, usb цифровой микроскоп и камеру (сенсор 1\ 4 дюйма CMOS, увеличение 200х, разрешение 1280х1024 пикс.) В данной работе выявлялись морфологические особенности исследуемых объектов с точки зрения законов красоты, мы постарались ответить на вопрос «Почему те растения, которые нас окружают, воспринимаются нами красивыми?» Исследования внешнего строения трав, кустарников и деревьев проводились в рамках антропогенных территорий города Кумертау.
Гистологические особенности комнатных растений
Изучение растений на тканевом уровне позволило мне понять причины морфологических особенностей растений. Большое значение имеют различные красящие вещества – пигменты, специфичные для каждого вида растений. Наиболее распространены антоцианы. В зависимости от реакций, которые он претерпевает в клеточном соке с солями, дубильными веществами, кислотами, он и придает различную окраску клеточному соку. Не следует смешивать антоциан с хлорофиллом, каротином, ксантофиллом и другими пигментами пластид (Желнин, 2006). Кроме привлечения насекомых, антоциан обладает защитным действием от низких температур, вредных короткосветовых волн. Из желтых пигментов в клеточном соке встречается антохлор. Мною были рассмотрены образцы растений и то, что я увидел представлено в приложении (прил. 4,7)
МОРФОЛОГИЧЕСКИЕ ОСОБЕННОСТИ РАСТЕНИЙ
Нам удалось описать внешнее строение всех изученных объектов, выделить существенные морфологические признаки, указывающие на наличие законов красоты (результаты в прил. 5, 6, 7, 8).
Значение цветов в эволюции живых организмов
Я думаю, что в природе все закономерно и красота цветов имеет большее, чем мы думаем значение. Чтобы убедиться в этом я стал искать литературу по этому вопросу и вот что нашел. Оказывается, секрет теории о красоте растений связан с науками психологией и эволюцией. Идея этой теории заключается в том, что: красота - это простой биологический фактор, конечная цель которого привлекать особей к размножению. И процесс этот задуман природой не для развлечения, а для продолжения рода.
В природе для роста и выживания требуется достаточное количество ресурсов. Деревьям и растениям необходимы солнечный свет, вода и питательные вещества. Животные нуждаются в достаточном количестве пищи и возможности размножения. При этом возникает жесткая конкуренция. Животные, расположенные выше по социальной лестнице, имеют больше возможностей для выживания. Особи, не способные бороться за пищу, в конечном счете, погибают. Благодаря естественному отбору из поколения в поколение передаются гены, несущие в себе качества выживших особей, такие как адаптация к условиям обитания. Постепенно эти качества становятся частью генетического кода этого вида (Желнин, 2006).Так частью генетического рода людей, как и большинства животных и насекомых, стала врожденная способность к восприятию красоты цветов. Наше восприятие красоты цветов является инстинктивным ответом на определенные цветовые оттенки, которые первоначально помогали нашим примитивным предкам находить пищу. В свою очередь цветы, таким образом, передают информацию о скором созревании плода. Растения используют цветы для привлечения животных и насекомых к помощи в размножении через опыление и распространение семян. Цветы становятся наиболее яркими, когда они созрели и готовы к опылению. Контрастные цвета делают цветок более заметным для насекомых и животных. Наличие цветовой двусторонней симметрии в цветке говорит о его большей развитости. Цветы дают возможность определенным видам растений, готовым к опылению, не затеряться в море зелени, привлечь насекомых или животных, способных помочь в размножении.
ВЫВОДЫ
-
Нами были исследованы различные виды растений. Мы пришли к выводу, что законы красоты существуют и действуют на всех исследованных объектах. Следовательно, они в той или иной степени присущи всем растениям. Исследовав литературные источники, мы пришли к выводу, что секрет красоты в упорядоченности, соответствии законам и правилам красоты. Они работают на внутриклеточном, клеточном, тканевом и организменном уровнях. Механизмы красоты, проявляющиеся на микроскопическом уровне, определяют внешнюю привлекательность цветка. Даже процессы смерти в клетках и тканях растений следуют определенным механизмам и подчиняются законам красоты (пример с лепестками желтой хризантемы).
-
Частью генетического кода людей, как и большинства животных и насекомых, стала врожденная способность к восприятию красоты цветов.
Материалы по данному исследованию могут быть использованы на занятиях по ботанике, общей биологии, могут служить основой к созданию ландшафтных композициях в озеленении города.
Список литературы
-
Ботаника с основами фитоценологии. Анатомия и морфология растений. Серебрякова Т.И. — М: Академкнига, 2006. — 543 с.
-
Ботаника. Систематика высших, или наземных, растений. Еленевский А.Г., Соловьева М.П., Тихомиров В.Н. — М: Академия, 2006. — 464 с.
-
Демьянов В. П. Геометрия и Марсельеза. – М.: Знание, 1986
-
Желнин Ю.Ю., Лихопуд Э.В. «Пигменты листа. Их свойства и значение.», Ж. «Биология в школе», № 4, с. 42-43, 2006 г.
-
Заренков Н.А. Биосимметрика. М.: Книжный дом «ЛИБРОКОМ», 2009. - 320 с.
-
Зенкевич И. Г. Эстетика урока математики. – М.: Дрофа.2005.
-
Иллюстрированное руководство для ботанических практик и экскурсий в Средней России. В.Э. Скворцов. — М: Т-во науч. изд. КМК, 2004. — 506 с.
-
Иллюстрированный определитель растений средней России. И.А. Губонов, К.В. Киселева, В.С. Новиков, В.Н. Тихомиров. — Т. 1-3. — М.: Т-во науч. изд. КМК, 2002-2004.
-
Коксетер Г. С. Новые встречи с геометрией. – М.: Наука, 2003.
-
Королькова Е.О. Методические рекомендации к лабораторным занятиям по анатомии и морфологии растений: Учебное пособие к лабораторно-практическим занятиям по ботанике. — М.: МАКС Пресс, 2008. — 22 с.
-
Маевский П.Ф. Флора Средней полосы Европейской части России. 10-е изд. — М: Т -во науч. изд. КМК, 2006. — 600 с.
-
Маевский П.Ф. Флора Средней полосы Европейской части России. 10-е изд. — М: Т -во науч. изд. КМК, 2006. — 600 с.
-
Миркин Б.М., Наумова Л.Г., Мулдашев А.А. Высшие растения: краткий курс систематики с основами науки о растительности. —М.: Логос, 2002. — 256 с.
-
Определитель сосудистых растений центра европейской России. И.А. Губанов, К.В. Киселева, В.С. Новиков, В.Н. Тихомиров. 2-е изд., дополн. и перераб. — М.: Аргус, 1995. — 560 с.
-
Практикум по анатомии и морфологии растений Викторов В.П., Гуленкова М.А., Дорохина Л.Н. и др. Под ред. Л.Н. Дорохиной — М: Академия, 2004. — 176 с.
-
Ч. Пэдхем, Дж. Сондерс. Восприятие света и цвета.- М.: «Мир», 1978.— С. 19.
-
Ченцов Ю.С. Введение в клеточную биологию. — М: ИКЦ «Академкнига», 2004. — 495 с.
-
Шафрановский И.И. Симметрия в природе. Ленинград, "Недра", 1985. - 168 с.
-
http://home.onego.ru/~otsoppe/book48.htm //Красота по правилам Из приложения "Цветы в саду и дома" журнала "Приусадебное хозяйство" 1/02.
-
http://biofile.ru // Исследования в области ботаники и зоологии в Новое Время.
-
art-ginkgo.com//Правила составления растительных композиций.
ПРИЛОЖЕНИЕ
Приложение 1
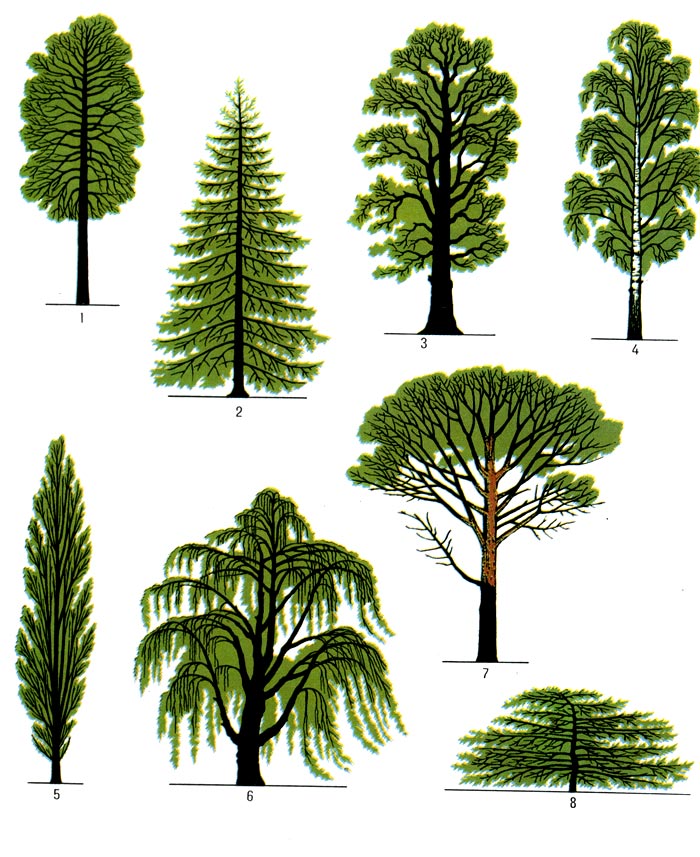
Рис. 1 Крона деревьев и кустарников 1- липы, 2 - ели, 3 - дуба, 4 - берёзы, 5 - тополя пирамидального, 6 – ивы плакучей, 7 - клёна, 8 – можжевельника казацкого. Примеры графичности крон.

Рис.2 Кроны деревьев. Графика побегов. Вид снизу. Парк им. Гагарина г. Кумертау.



Рис.3 Береза, клен, дуб – примеры разреженных, грубофактурных крон с различной текстурой и фактурой листьев. Сезонная расцветка. В цветах встречается контраст и нюанс.



Рис. 4 Фактура листьев Рис. 5 Текстура листьев


Рис. 6 Примеры нюанса цвета

Рис. 7 Сочетание нюанса и контраста
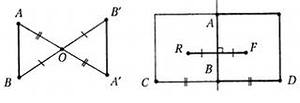
Рис.8 Центральная симметрия
Приложение 2
Примула (Primulaceae), Нарцисс (Amaryllidaceae), Pyrola (Ericaceae)
Pyrola (Ericaceae)
Дисимметрия
Две
(дисимметричные)
Билатеральная симметрия
Dicentra(Fumariaceae)

Зигоморфия
Одна (моносимметричные)
Билатеральная, нерегулярная, медиальная зигоморфия
медиальная зигоморфия или билатеральная симметрия
Salvia (Lamiaceae), Орхидея (Orchidaceae), Scrophularia (Scrophulariaceae)

трансверс (верх-низ) зигоморфия
Fumaria и Corydalis (Fumariaceae)

диагональная зигоморфия
облигатная зигоморфия
Aesculus (Hippocastanaceae) находят у Malpighiaceae, Sapindaceae

Приобретённая асимметрия
нет
Нерегулярная, асимметрия
-новая асимметрия
Нерегулярная, асимметрия
Centranthus (Valerianaceae), находят у Cannaceae, Fabaceae, Marantaceae, Zingiberaceae

- энантиоморфия (моно-энантиоморфия, диэнантиоморфия
Энантиостилия, неравнолатеральная
Cassia (Caeasalpinaceae), Cyanella (Tecophilaeceae), Monochoria (Pontederiaceae), Solanum (Solanaceae), Barberetta и Wachendorffia (Haemodoraceae)
Приложение 3
Восприятие цветов
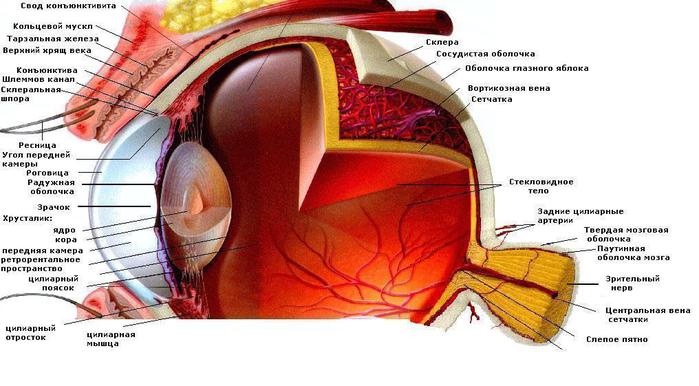
Рис. 9 Строение глаза
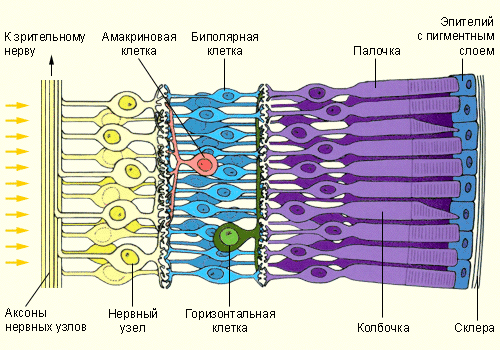
Рис.10 Колбочки и палочки в основе строения глаза
Восприятие цветов - это способность зрительной системы различать цвета объектов в поле зрения. Для полноценного восприятия цветов нужна согласованная работа чувствительного аппарата зрительной системы. Механизм цветовосприятия по общепринятой, но уже устаревшей теории о трех компонентах состоит в том, что в зрительной системе имеются три датчика, восприимчивых к трем основным цветам: красному, желтому и синему. Поэтому правильное восприятие цветов называется трихромазией. При определенном смешении трех главных цветов возникает видимость белого цвета. При нарушении работы одного или двух датчиков основных цветов правильного смешения цветов не наблюдается и возникают нарушения цветовосприятия. Цвета отличаются друг от друга кроме всего прочего и по степени яркости. Самым ярким является желтый цвет, за ним следуют оранжевый, красный, зеленый, синий, фиолетовый цвет. Это яркость цвета не абсолютная, а субъективная. Так как она связана с особенностями восприятия данных цветов именно человеком. Маленькое желтое пятно на темной фотографии воспринимается зрением мощнее, чем большое фиолетовое. Например, все знают, что белая одежда зрительно полнит, а черная наоборот - стройнит. Цвет бывает теплым (например, красный, оранжевый, желтый) и холодным (скорее всего это зеленый, синий, пурпурный).
Необходимо еще учитывать и то, что зрение человека имеет еще и такое интересное свойство, как адаптивность. Оно автоматом настраивается на освещенность объекта. Заскочив в достаточно темное помещение в яркий полдень, некоторое время вы почти ничего не будете видеть) (Пэдхем, 1978).
Приложение 4
Гистологические особенности исследованных растений

Рис. 11 Лилия белая (Lílium) (свежий препарат 160х). Чередуются клетки причудливой «амебоидной»формы («ритм»).

Рис. 12 Лист лилии (Lílium) (160х). Видны устьица одинакового размера и на равном расстоянии («ритм»). Две половинки устьица – замыкающие клетки – симметричны.
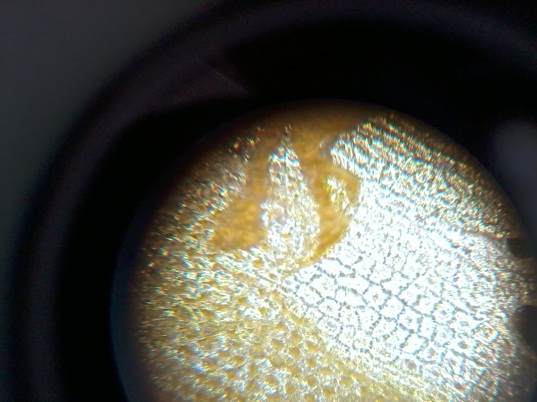
Рис. 13 Хризантема желтая (Chrysánthemum) (64х). Чередуются полосы квадратных, ромбовидных, овальных, трапецевидных и др. клеток (ритм). Нюанс проявляется в переходах оттенков с более светлых к темным.
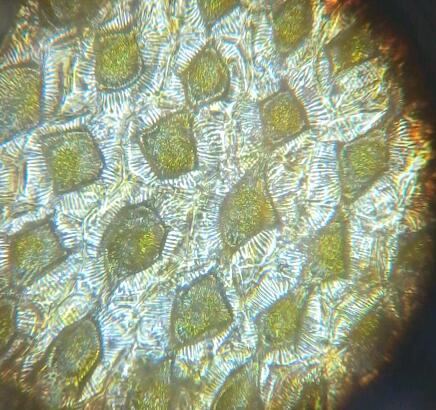
Рис. 14 Хризантема желтая (Chrysánthemum) (640х). При высыхании лепестков наблюдался механиз сходный с плазмолизом: отставание протопласта от клеточной стенки. При этом получались ромбовидной формы структуры, повторяющиеся из клетки в клетку и содержащие в центре ядро.
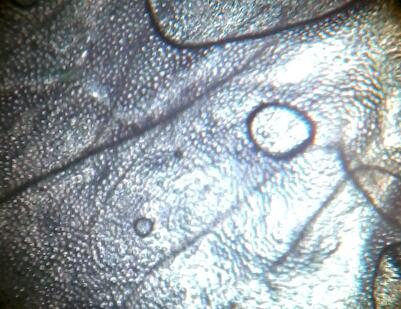
Рис. 15 Гвоздика белая (Diánthus) (64х). Мелкие клетки дают яркую белую окраску.

Рис. 16 Хризантема белая(Chrysánthemum) отличается наличием участков, содержащих пигменты. Переход от части с пигментами и без постепенный и соответствует «нюансу».
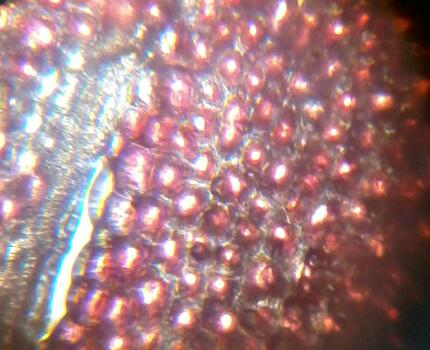
Рис. 17 Гвоздика красная (Diánthus) (160х). Клетки за счет пигментов клеточного сока переливаются словно россыпь драгоценных камней. Пвторение клеток – «ритм».
infourok.ru
1. Особенности морфологической эволюции фототрофных растений. Типы морфологической организации растений. Основные вегетативные органы цветковых растений.
И у одноклеточных зеленых водорослей, и у высших растений клетка сохраняет одни и те же основные черты организации. Для нее характерна наружная сплошная упругая полисахаридная оболочка, образующая опорную систему, наружный скелет клетки. Возникновение такой оболочки явилось следствием фототрофного питания, при котором в клетке образуется избыток продуктов фотосинтеза — растворимых углеводов. Эти вещества выводятся из клетки и превращаются в нерастворимые углеводы оболочки. Вода, С02 и различные минеральные соли, равномерно рассеянные вокруг растения в почве и атмосфере, проникают через оболочку. Вещества, выделяемые клеткой, также проходят через клеточную оболочку в виде истинных растворов. Таким образом, наличие сплошной оболочки, одевающей клетку, соответствует основному признаку обмена веществ у растений — поглощению и выделению веществ в растворенном виде.
Поскольку элементы питания равномерно рассеяны в окружающей среде, растения в ходе эволюции постепенно потеряли подвижность и перешли к прикрепленному образу жизни. Отдельные особи в течение всей жизни остаются на одном месте, а передвижение чаще всего связано с постепенным нарастанием и отмиранием более старых органов или с размножением.Элементы питания, равномерно распределенные во внешней среде, поглощаются через наружную поверхность растения. Чем больше поверхность соприкосновения, тем интенсивнее идет обмен веществ. Поэтому в процессе эволюции наблюдается тенденция к увеличению поверхности
2. Соцветия. Общая характеристика. Простые и сложные соцветия.
Соцве́тие (лат. inflorescentia) — часть системы побегов покрытосеменного растения, несущая цветки и в связи с этим разнообразно видоизменённая. Соцветия обычно более или менее четко отграничены от вегетативной части растения.
По наличию и характеру прицветных листьев (прицветников):
Фрондозные (лат. frondis — листва, листья, зелень), или олиственные — соцветия, в которых прицветники имеют хорошо развитые пластинки (например, фуксия, фиалка трёхцветная, вербейник монетчатый).
Брактеозные — соцветия, в которых прицветники представлены чешуевидными листьями верховой формации — брактеями (например, ландыш, сирень, вишня).
Эбрактеозные, или голые — соцветия, в которых прицветники редуцированы (например, дикая редька, пастушья сумка и другие капустные (крестоцветные).
По степени разветвления:
Простые — соцветия, в которых на главной оси располагаются одиночные цветки и, таким образом, ветвление не превышает двух порядков (например, гиацинт, черёмуха, подорожник и др.).
Сложные — соцветия, в которых на главной оси располагаются частные (парциальные) соцветия, то есть ветвление достигает трёх, четырёх и более порядков (например, сирень, бирючина, калина и др.).
По типу нарастания и направлению раскрывания цветков:
Рацемозные, или ботрические (от лат. racēmus и греч. ботрион — кисть, гроздь) — соцветия, характеризующиеся моноподиальным типом нарастания осей и акропетальным (то есть направленным от основания оси к её верхушке) раскрыванием цветков (например, иван-чай, пастушья сумка и др.)
Цимозные (от лат. cyma — полузонтик) — соцветия, характеризующиеся симподиальным типом нарастания осей и базипетальным (то есть направленным от верхушки оси к её основанию) раскрыванием цветков (например, медуница).
По характеру поведения апикальных меристем:
Закрытые, или определённые — соцветия, в которых апикальные (верхушечные) меристемы осей расходуются на образование верхушечного цветка (все цимозные соцветия, а также рацемозные некоторых растений: хохлаток, толстянок, колокольчиков и др.).
Открытые, или неопределённые — соцветия, в которых апикальные меристемы осей остаются в вегетативном состоянии (ландыш, гиацинт, грушанка и др.).
Простые соцветия[править | править исходный текст]
Как уже было сказано выше, простыми называются соцветия, в которых все цветки располагаются только на главной оси. Обычно соцветия этой группы являются рацемозными.Сложными называются соцветия, в которых на главной оси располагаются не одиночные цветки, а парциальные (частные) соцветия.
5БИЛЕТ
studfiles.net
Биологические и морфологические особенности культуры
Гречиха (Fagopyrum sagittatum Libib. (Fagopyrum esculentum Moench) — однолетнее растение семейства гречишных (Polygonaceae), включающая несколько видов. Другие названия: черный рис, черная пшеница.
Требования к температуре.
Гречиха относится к теплолюбивым культурам. Семена ее начинают прорастать при температуре почвы 7- 8 °С, однако развитие проростков лучше идет при температуре от 15 до 30 °С. Всходы ее чувствительны к заморозкам и повреждаются при температуре воздуха -2...-3 °С, при -4 °С растения полностью погибают. При температуре ниже 12-13 °С гречиха растет плохо.
В период цветения — плодообразования гречиха очень восприимчива к повышенной температуре, которая отрицательно влияет на образование плодов, особенно при недостатке влаги. Количество оплодотворенных завязей редко превышает 20 %. Жара и засуха, дожди и туманы, ветры, резкие колебания температуры нарушают опыление, налив семян и приводят к снижению урожая. Оптимальная температура воздуха в этот период должна составлять 17-25 °С при относительной влажности не менее 50 %. Температура выше 30 °С и относительная влажность воздуха менее 30 % приводят к ухудшению опыления и массовому отмиранию завязей. Отрицательно влияют на образование плодов и температуры ниже 12-14 °С. При высокой агротехнике и организации пчелоопыления количество завязавшихся плодов значительно увеличивается, и созревание семян происходит дружно.
Требования к влаге.
Гречиха — влаголюбивая культура. Транспирационный коэффициент варьирует от 480 до 600. На образование единицы органического вещества она расходует в 3 раза больше влаги, чем просо и в 2 раза больше, чем пшеница и ячмень. Период цветение — налив плодов — критический период по отношению к влаге. Максимальное потребление влаги (50-60 %) происходит в первые 2-3 декады периода цветения-плодообразования. М. Е Николаев, изучая влияние различной влажности почвы на скорость роста гречихи, установил, что самая низкая скорость роста растений гречихи (13,2 мм в сутки) была при влажности почвы 30 % от полной влагоемкости. Затем с повышением влажности абсолютные приросты растений в высоту увеличивались и достигали максимальных величин (39,2-40,0 мм/сут) при влажности почвы 70-80 %. Другими словами, скорость роста при данных условиях влажности была в 3 раза выше по сравнению с изначальной. Дальнейшее возрастание влажности почвы до 90 % нарушало ход ростовых процессов и резко снижало рост, но максимальное развитие корневой системы наблюдалось при 60 % от полной полевой влагоемкости.
Требования к плодородию почвы.
Выбор участка имеет порой решающее значение. Для возделывания гречихи малопригодны бугристые или низинные места. На буграх растения страдают от недостатка влаги, а в низких местах — от ее избытка. Кроме того, в понижениях скапливается холодный воздух, который, переохлаждаясь, образует туман, влияющий отрицательно на цветение и плодообразование гречихи. Культура быстро развивается на легких, плодородных, хорошо аэрируемых почвах. На участках, расположенных вблизи водоемов, окруженных лесом, а также защищенных от действия иссушающих ветров, гречиха лучше завязывает плоды, формирует более высокий урожай. Она переносит повышенную кислотность, однако лучшие урожаи дает на слабокислых и близких к нейтральным почвах. Оптимальные агрохимические показатели почв: pH в КСI — 5-7, содержание гумуса не менее 1,6 %, подвижных форм фосфора и калия — не менее 100 мг/кг почвы.
Корневая система гречихи характеризуется слабым развитием, но очень высокой физиологической активностью. По массе корней на единицу площади она уступает пшенице в 2,4 раза, ячменю — в 1,6 раза, а по поглотительной способности, наоборот, превосходит в 2,7 раза пшеницу и в 5,5 раза ячмень. Гречиха лучше развивается на почвах с хорошей аэрацией, влагоемкостью и плодородных. Такими свойствами в наибольшей мере обладают легко- и среднесуглинистые, хорошо окультуренные почвы. Культура переносит повышенную кислотность, однако лучшие урожаи дает на почвах слабокислых и близких к нейтральным.
Лучшие почвы для гречихи — дерново-подзолистые легкосуглинистые и связно-супесчаные, подстилаемые с глубины до 1 м моренным суглинком, хорошо аэрируемые. На песчаных почвах высевать гречиху не следует, так как она наиболее требовательная к влаге культура среди яровых Она также плохо растет на заплывающих холодных тяжелых почвах. В низких местах, где посевы могут пострадать от избытка влаги, заморозков, туманов, гречиху высевать нельзя. При соответствующих мероприятиях (известкование, удобрение и т. д.) гречиха может произрастать и на недостаточно окультуренных почвах: кислых, болотных и малоплодородных, вышедших из-под леса, на осушенных торфяниках. Малопригодны для нее и возвышенности. На тучных и переудобренных почвах при достаточном увлажнении гречиха развивает большую вегетативную массу в ущерб образованию плодов.
studfiles.net
МОРФОЛОГИЧЕСКИЕ ОСОБЕННОСТИ СТЕБЛЕЙ
Морфологическое строение стеблей покрытосеменных растений характеризуется значительно большим разнообразием, чем анатомическое. По характеру и направлению роста можно выделить стебли прямостоячие, приподнимающиеся, вьющиеся, цепляющиеся, ползучие, стелющиеся, укороченные.
Прямостоячие стебли присущи подавляющему большинству растений. Они имеют достаточно хорошо развитые механические ткани, что позволяет им самостоятельно поддерживать вертикальное отрицательно геотропичное положекние.
У растений с приподнимающимся стеблем (Trifolium pratense – клевер луговой) основание дуговодно изогнуто, а верхушка стебля располагается вертикально.
Вьющиеся стебли отличаются способностью к круговым (нутационным) движениям, что позволяет растениям обвиваться вокруг различных опор и принимать вертикальное положение. Круговые движения связаны с перемещением вокруг оси органа зон усиленного роста. Вьющиеся растения очень богаты гибереллином, и можно считать, что этот фитогормон причастен к осуществлению обвивания. Есть растение с правовращающимися (по часовой стрелке – жимолость-каприфоль – Lonicera сaprifolium; хмель обыкновенный – Humulus lupalus) и левовращающимися (против часовой стрелки – Ипомея пурпуровая – Ipomea purpurea, фасоль огненно-красная – Phaseolus coccineus, вьюнок полевой – Convolvulus arvensis) стеблями. В большинстве случаев растения обладают способностью к левовращению. Правое вращение стеблей отмечено у немногих видов. При помощи различных приспособлений поддерживают стебель в вертикальном положении цепляющиеся растения. У виноградов амурского (Vitis amurensis), винограда девичьего пятилисточкового (Partenocyssus quinquefolia) и других представителей семейства виноградных стебель цепляется за опору при помощи усиков. Так же цепляются за опору и многие представители семейства бобовые (горошек тонколистный – Vicia tenuifolia, чина клубненосная – Lathyrus tuberosus). У плюща обыкновенного (Hedera helix) на концах придаточных корней, которые в большом количестве образуются на стебле, возникают особые присоски. При помощи таких корней-присосок плющ прицепляется к стволам деревьев, а в южных районах, где его используют для вертикального озеленения – к стенам зданий.
При помощи черешков листьев, которые обвиваются вокруг опоры, поднимаются на высоту двух – четырех метров стебли многих видов ломоноса (Clematis).
Прямостоячие, приподнимающиеся, вьющиеся и цепляющиеся стебли являются ортотропными, они в конечном счете принимают вертикальное положение. Однако есть и плагиотропные стебли, располагающиеся горизонтально. К ним относятся ползучие и стелющиеся стебли.
Ползучими называются стебли, у которых в узлах образуются придаточные корни, при их помощи растение прикрепляется к субстрату и захватывает территорию – «ползет». Такие стебли характерны для лютика ползучего (Ranunculus repens), живучки ползучей (Ajuga reptans), земляники лесной (Fragaria vesca). Иногда ползучие стебли земляники называют усами.
В отличие от ползучих стелющиеся стебли не образуют придаточных корней и их олиственные или безлистные стебли распростерты по субстрату (ястребиночка обыкновенная – Pilosella officinarum) стелющиеся стебли встречаются как среди травянистых, так и среди деревянистых растений (кизильник горизонтальный – Cotoneaster saxatile f. horizontale). Предполагают, что у древесных и кустарниковых растений стелющиеся стебли являются результатом приспособления к неблагоприятным условиям. Много видов со стелющимися стеблями произрастает в субарктических и арктических областях, в горах, а также на океанических подбережьях. У травянистых растений стелющиеся стебли часто образуются при сильном затенении или избыточном увлажнении (лисохвост коленчатый – Alopecurus geniculatus, л. равный – G. aequalis; полевица побегообразующая – Agrostis stolonizans).
Особым типом стеблей является укороченный стебель. Укороченный стебель имеет сильно сближенные узлы и погружен в почву, поэтому над субстратом образуется розетка листьев (одуванчик лекарственный – Taraxacum officinale, подорожник большой – Plantago major и др.). Цветоносные стебли (цветоносы) у таких растений безлистные, прямостоячие.
Различаются стебли и по поперечному сечению. Наиболее типичным является круглый стебель. У злаков полый в междоузлиях круглый стебель называется соломиной. Стебли могут быть трехгранными (виды рода Carex – осока), четырехгранными (семейство Labiatae – Губоцветные), сплюснутыми (мятлик сплюснутый – Poa compressa, опунция – Opuntia), ребристыми (гладыш прусский – Laserpitium prutenicum, хвощ полевой – Equisetum arvense). Своеобразное строение имеет крылатый стебель (Lathyrus sylvestris – чина лесная, чертополох курчавый –- Carduus crispus). У этих растений вдоль круглого стебля на всем протяжении или участками прикреплены листовидные образования. Поперечное сечение такого стебля похоже на рыбий позвонок.
Морфологические особенности стебля имеют важное диагностическое значение и используются при идентификации видов покрытосеменных растений.
Указанные морфологические особенности характерны для стеблей, выполняющих типичные функции. При выполнении дополнительных функций стебель метаморфизируется, а вместе с ним метаморфизируются и листья. Таким образом, следует говорить не просто о метаморфозе стебля, а о метаморфозе побега в целом.
МЕТАМОРФОЗЫ ПОБЕГА
Метаморфозы побега более разнообразны, чем метаморфозы корня. Довольно часто метаморфизированные образования имеют сходное строение, поэтому в связи с понятием метаморфозов органов возникли понятия об аналогичных и гомологичных органах.
Аналогичными называются органы, которые имеют различное происхождение, но выполняют одинаковые функции и имеют сходный внешний вид (морфологическое строение). Например, колючки, усики стеблевого и листового происхождения. Определить в таком случае происхождение метаморфизированного образования можно по расположению их на стебле. Метаморфозированные образования стеблевого (побегового) происхождения выходят из пазухи листа. Метаморфозы листового происхождения располагаются на стеблевом узле.
Гомологичными называются органы, имеющие одинаковое происхождение, но выполняющие разные функции, вследствии чего имеют различное строение. Гомологичны листу колючки листового происхождения (барбарис обыкновенный – Berberis vulgaris) усики листового происхождения (горох посевной – Pisum sativum, чина луговая – Lathyrus pratensis), сухие чешуи на корневищах (пырей ползучий – Elytrigia repens, осока мохнатая – Carex hirta), сочные чешуи луковиц (лук репчатый – Allium cepa, нарцисс гибридный – Narcissus hybridus) и др.
К метаморфозам побега относятся корневища, луковицы, клубни, столоны, кладодии (или филлокладии), колючки и усики.
Корневище (rhizoma) – подземный метаморфизированный побег, у которого в отличие от типичного побега листья редуцированы и превращены в сухие (осока мохнатая – Carex hirta) или сочные (Петров крест обыкновенный – Lathraea squamaria) чешуи.
По особенностям формирования различают корневища эпигеогенные (копытень европейский – Asarum europaeum, представители родов Манжетка – Alchemilla, Гравилат – Geum, Медуница – Pulmonaria) и гипогеогенные (ландыш майский – Convallaria majalis, грушанка круглолистная – Pyrola rotundifolia и др.)
По направлению роста различают корневища плагиотропные – горизонтально распростертые (гравилат речной – Geum rivale) и ортотропные (вех ядовитый – Cicuta virosa) – растущие вертикально по направлению силы тяжести Земли.
Корневища могут ветвиться моноподиально (вороний глаз четырехлистный – Paris quadrifolia) и симподиально (купена лекарственная – Polygonatum odoratum,
По консистенции корневища бывают сухими (пырей ползучий) и сочными (аир обыкновенный – Acorus calamus, ирис безлистный – Iris aphylla).
Луковица (bulbus) – метаморфизированный побег, большая часть которого образована метаморфизированными листьями – сочными чешуями.
В зависимости от места образования на растении луковицы бывают подземными и надземными. Надземные луковицы имеют небольшие размеры, их часто называют бульбочками, или луковичками. Луковички могут формироваться в пазухах листа (лилия тигровая – Lilium tigrinus, зубянка луковичная – Dentaria bulbifera) или в соцветии (чеснок – Allium sativum, мятлик луковичный – Poa bulbosa, лук огородный – Allium oleraceum).
По особенностям расположения чешуй различают луковицы туникатные и черепитчатые. Туникатные луковицы образованы сросшимися концентрически расположенными на донце – уплощенном стебле – чешуями. Число чешуй в луковице варьирует от одной (чеснок – Allium sativum) до многих (лук репчатый – Allium cepa). По степени сложности луковицы бывают простыми и сложными. В сложной луковице (чеснок) под общей сухой чешуей располагается много луковичек.
Поскольку луковицы являются метаморфизированными побегами их можно различать и по типам ветвления. У симподиальных луковиц (тюльпан гибридный – Tulipa hybrida, рябчик королевский – Fritillaria imperialis, лук репчатый – Allium cepa) цветонос формируется из верхушечной почки, а возобновление (образование дочерних луковиц) – из пазушных.
У моноподиальных луковиц (подснежник снежнобелый – Galanthus nivalis, нарцисс гибридный – Narcissus hybridus) возобновление идет из верхушечной почки, а цветоносы – из пазушных.
По продолжительности жизни луковицы бывают многолетними (гипеаструм гибридный – Hyppeastrum hybridum, нарцисс) и однолетними (тюльпан, лук). Однолетние луковицы ежегодно отмирают, а вместо отмершей (материнской) луковицы образуется замещающая луковица и луковицы-детки. Растения, у которых луковицы ежегодно отмирают, следует считать не многолетниками, а вегетативными малолетниками.
Вегетативными малолетниками являются и клубнелуковичные растения (шпажник черепитчатый – Gladiolus imbricatus, шафран посевной – Crocus sativus).
Клубнелуковица (bulbotuber) в отличие от луковицы формируется преимущественно за счет разрастания и уплощения стебля. Листья на клубнелуковицах чешуйчатые и надежно прикрывают верхушечную и пазушные почки. Клубнелуковицы, как и луковицы, могут быть однолетними (шафран, гладиолус гибридный – Gladiolus hybridus), так и многолетними (безвременник осенний – Colchicum.autumnale).
Клубни (tuber) определяют как метаморфизированные подземные побеги.
Однако у некоторых растений клубни или мелкие образования – клубеньки возникают в надземной части растения. Мощный надземный клубень образуется в результате разрастания стебля у капусты кольраби (Brassica oleracea var. longeloides).
У ряда тропических эпифитных орхидей основание стебля сильно разрастается и превращается в туберидий (кеттлея – Cattleya, максиллярия – Maxillaria).
У горца живородящего (Polygonum vivipara) клубеньки образуются в соцветиях, а у чистяка весеннего (Ficaria verna) – в пазухах листьев.
Подземные клубни могут иметь различное происхождение. Если у картофеля (Solanum tuberosum), топинамбура (Helianthus tuberosus) это метаморфизированные побеги, о чем свидетельствует наличие почек (верхушечных и пазушных), стеблевых узлов (бровка), междоузлий, редуцированных до едва заметных чешуек листьев, то у цикламена индийского (Cyclamen indica) подземный клубень образован только частью стебля – гипокотилем.
Столоны – безлистные образования, фактически единственное сильно удлиненное междоузлие подземного побега, несущее на конце клубень (картофель) или луковицу (лук угловатый – Allium angulusum). Столоны обычно плагиотропы, однако у дикорастущих тюльпанов и у культурных, если их ежегодно не выкапывают, образуются ортотропные столоны, и дочерние луковицы постепенно сильно заглубляются. Это приводит к измельчению луковиц и к быстрому вырождению сорта.
У растений засушливых местообитаний побеги могут превращаться в кладодии (clados) – листоподобно уплощенные образования (Ruscus – иглица), или филлокладии (Asparagus – спаржа). Термины кладодии и филлокладии некоторые ботаники рассматривают как синонимы, другие придают им самостоятельное значение и указывают, что для кладодий характерен длительный рост, а рост филлокладий ограничен. И кладодии, и филлокладии повернуты ребром к солнцу, что имеет важное приспособительное значение (уменьшение испарения, защита от перегревания). Кладодии и филлокладии всегда располагаются в пазухах чешуйчатых «листьев», что подтверждает их побеговое (стеблевое) происхождение. Об этом же свидетельствует образование цветков. У иглиц на многолетних кладодиях находятся довольно длинные цветоносы, на концах которых ежегодно формируются цветки.
Колючки стеблевого происхождения характерны для ряда деревьев (Pyrus communis – груша обыкновенная) и кустарников (Crataegus monogina – боярышник однопестичный). У формирующихся, молодых, колючек на стебле видны зачаточные, недоразвитые листья, которые расположены так же, как и типичные листья. С возрастом колючка одревесневает, а зачаточные листья абортируются, т. е. полностью исчезают. Колючки выполняют защитную функцию.
Усики стеблевого происхождения или выходят из пазухи листа, или заканчивают каждый метамер симподиально ветвящегося стебля (огурец посевной – Cucumis sativus, виноград культурный – Vitis vinifera).Усики могут быть простыми или разветвленными, основная их функция опорная.
Таким образом, метаморфозы побега разнообразны. Метаморфизированные побеги выполняют различные функции, в том числе функции сохранения и размножения вида (клубни, корневища, луковицы, клубнелуковицы).
Похожие статьи:
poznayka.org
Презентация на тему "Морфологические особенности растений различных видов"
Презентация на тему: Морфологические особенности растений различных видовСкачать эту презентацию
Скачать эту презентацию
№ слайда 1 Описание слайда:
Описание слайда: Лабораторная работа Тема: «Морфологические особенности растений различных видов»
№ слайда 2 Описание слайда:
Описание слайда: Цель работы : -закрепить понятие морфологического критерия вида; -учиться составлять описательную характеристику растений.
№ слайда 3 Описание слайда:
Описание слайда: Оборудование: Живые растения и гербарные материалы растений разных видов: лютик едкий, лютик ползучий, полынь приморская, полынь горькая, крапива двудомная (жгучая), глухая крапива (яснотка белая)
№ слайда 4 Наблюдения и выводы запишите в тетрадь.") Описание слайда:
Описание слайда: Ход работы Смотри стр.292 (учебник) Наблюдения и выводы запишите в тетрадь.
№ слайда 5 Описание слайда:
Описание слайда: Лютик едкий Домен: Эукариоты Царство: Растения Отдел: Цветковые Класс: Двудольные Порядок: Лютикоцветные Семейство: Лютиковые Род: Лютик Вид: Лютик едкий
№ слайда 6 Описание слайда:
Описание слайда: Лютик едкий — многолетнее травянистое растение, достигает в высоту 20—50 см. Листья — нижние — длинночерешковые, длиной 5—10 см, пятиугольные, пальчатораздельные; верхние — сидячие, трёхраздельные с линейными, зубчатыми долями. Цветки — ярко-жёлтого цвета, достигают 2 см в диаметре, одиночные или собраны всоцветие полузонтик. Чашелистиков пять;лепестков — пять; множество тычинок и пестиков. Цветёт в июне. Плод — многоорешек плод - Народное название: куриная слепота. цветок
№ слайда 7 Описание слайда:
Описание слайда: Лютик ползучий Царство:Растения Отдел:Покрытосеменные Класс:Двудольные Порядок:ЛютикоцветныеСемейство:ЛютиковыеРод:Лютик Вид: Лютик ползучий
№ слайда 8 Описание слайда:
Описание слайда: Лютик ползучий — многолетнее травянистое растение высотой 15-40 см с коротким корневищем. Стебель восходящий или стелющийся, укореняющийся в узлах. Прикорневые листья черешковые, тройчатые, состоящие из ромбовидно-яйцевидных, глубоко тройчатораздельных, неравнозубчатых листочков. Верхние листья сидячие, ланцетные, тройчатораздельные.Цветки обоеполые, правильные, с пятью оттопыренными чашелистиками, пятью золотисто-жёлтыми лепестками, многочисленными тычинками и пестиками. Цветки одиночные верхушечные или собраны в полузонтик. Плод — многоорешек с короткими носиками у орешков. Растёт на влажных, затенённых, наносных почвах: по берегам рек и озёр, на влажных лугах, вкустарниковых зарослях, на лесных болотах, по полям и огородам.
№ слайда 9 Описание слайда:
Описание слайда: Лютик ползучий
№ слайда 10 Описание слайда:
Описание слайда: Лютик азиатский, махровый • однолетние и многолетние корневищные или корнеклубневые растения; • стебли прямостоячие, восходящие или распростертые, часто укореняющиеся в узлах; • листья цельные, лопатообразные, пальчато- или перистораздельные, расположены очередно; • цветки одиночные или собраны в соцветие, желтые, реже белые или красные, простые или махровые; • плод - многоорешек; • семена голые или волосистые, плоские или выпуклые
№ слайда 11 Описание слайда:
Описание слайда: Внимание! Любые виды этого неприхотливого растения ядовиты, хотя содержат и массу полезных веществ, из-за чего с осторожностью используется в народной медицине. Самые распространенные представители этого семейства – лютик едкий, лютик ползучий, лютик ядовитый.
№ слайда 12 Описание слайда:
Описание слайда: Крапива двудомная или жгучая
№ слайда 13 - многолетнее травянистое жгучее растение с") Описание слайда:
Описание слайда: Крапива двудомная (Urtica dioica L.) - многолетнее травянистое жгучее растение семейства крапивных (Urtiсасеае) с длинным ползучим корневищем. Стебель прямостоячий, высотой 90—120 см, с супротивносидячими яйцевидноланцетными черешковыми листьями длиной 8—17 см. Цветки мелкие, зеленые, однополые, собраны в колосовидные повисающие соцветия. Плод — яйцевидный или эллиптический, желтовато-серого цвета орешек 1,2—1,5 мм длины. Цветет с июня до сентября. Крапива как сорное растение распространена повсеместно в Европейской части России, на Кавказе и в Западной Сибири, встречается в Восточной Сибири, на Дальнем Востоке и в Средней Азии
№ слайда 14 Описание слайда:
Описание слайда: Глухая крапива, яснотка белая Царство:Растения Отдел:Цветковые растения Класс:Двудольные Порядок:Ясноткоцветные Семейство:Яснотковые Род:Яснотка Вид:Яснотка белая
№ слайда 15, многолетнее травянистое растение с") Описание слайда:
Описание слайда: Глухая крапива, яснотка белая (Lamium album), многолетнее травянистое растение семейства губоцветных. По форме листьев похожа на крапиву, но лишена жгучих волосков. Цветки белые, в ложных мутовках. Корневище ползучее горизонтальное. Стебли четырёхгранные, прямостоячие, до 60 см высотой. Листья супротивные. Черешки листьев до 4 см длиной (у верхушечных листьев много короче — до 1 см). Листовые пластинки яйцевидные или сердцевидные, морщинистые, до 10 см длиной, остропильчатые. Всё растение коротко опушено. Растет в умеренной зоне Северного полушария.
№ слайда 16 Описание слайда:
Описание слайда: Полынь горькая Домен: Эукариоты Царство: Растения Отдел: Цветковые Класс: Двудольные Порядок: Астроцветные Семейство: Астровые Род: Полынь Вид: Полынь горькая
№ слайда 17 Описание слайда:
Описание слайда: Полынь горькая Высота растения 50—125 см, нередко растёт как полукустарник, со стержневым ветвистым корнем и прямостоячими побегами, с серебристо-войлочным опушением. Стебли прямые, слаборебристые, в верхней части ветвистые. Нижние листья длинночерешковые, дважды-трижды перисто-рассечённые, средние — короткочерешковые, дважды перисто-рассечённые, верхние — почти сидячие, перистые или дважды тройчато-раздельные; дольки всех листьев линейно-продолговатые, тупо заострённые. Цветки все трубчатые, жёлтые; краевые — пестичные, срединные — обоеполые. Корзинки шаровидные, 2,5—3,5 мм в диаметре, в нешироком метельчатом соцветии. Обёртка корзинок черепитчатая, листочки широко-плёнчатые. Цветоложе выпуклое, волосистое. Цветение в европейской части России в июне—июле. Плод — буроватая заостренная семянка около 1 мм длиной, продолговато-клиновидная, тонко-бороздчатая, на верхушке с округлой, слегка выпуклой площадкой.
№ слайда 18 Описание слайда:
Описание слайда: Полынь приморская Многолетнее травянистое растение или полукустарник высотой 20-100 см, бело-опушенное, сильно пахнущее, с деревянистым корнем. Стебли многочисленные, восходящие, в верхней части ветвистые, образующие небольшую дернину. Листья с обеих сторон беловатоопушенные, очередные, перисторассеченные, с узкими линейными долями, длиннее корзинок; нижние листья черешковые. Соцветие раскидисто-метельчатое, с отогнутыми книзу цветоносами. Корзиночки поникшие, расположенные метелками на концах веточек стеблей; они очень мелкие, эллиптически-овальные, 1-2 мм длинной, зеленовато-серые в нерасцветшем состоянии, содержат 3-5 цветочков. Трубчатые цветки красноватые или желтоватые. Цветет в июле - октябре. Растет в степях, на сухой солонцеватой почве и меловых отложениях в степной и полупустынных зонах России и по побережью Черного, Азовского и Балтийского морей.
№ слайда 19 Описание слайда:
Описание слайда: Полынь приморская
ppt4web.ru
Лекция 7. ЭКОЛОГО-МОРФОЛОГИЧЕСКАЯ КЛАССИФИКАЦИЯ ЖИЗНЕННЫХ ФОРМ РАСТЕНИЙ ДАЛЬНЕГО ВОСТОКА по А.Г. Крылову (1983)
Москалюк Т.А.
Список литературы
Крылов А.Г. Жизненные формы лесных фитоценозов. Л.: Наука, 1984. 184 с.
Вопросы
1. Методологические принципы построения классификации
2. Жизненные формы деревьев
3. Жизненные формы кустарников
4. Жизненные формы трав и кустарничков
1. Методологические принципы построения классификации
Для растений Дальнего Востока одна из классификаций жизненных форм разработана А.Б. Безделевым и Т.А. Безделевой. Другой вариант разработан А.Г. Крыловым (1983). При выборе признаков для анализа, сравнения и классификации видов лесных растений в системе жизненных он принял несколько условий:
- - набор признаков должен разносторонне характеризовать экологически значимые морфологические свойства видов;
- - для удобства оперирования количество признаков не должно быть слишком большим;
- - признаки должны быть универсальными для сопоставления растений различных форм роста;
- - экологические свойства видов должны учитываться наряду с морфологическими особенностями.
Особое внимание было обращено на признаки листьев, так как именно облиственные растения во время вегетационного периода создают характерную структуру растительного сообщества, а листовой аппарат фитоценоза, ответственный за его газообмен с воздушной средой, является главным источником продуктов фотосинтеза и потребителем элементов минерального питания.
В результате был выбран следующий набор морфологических и экологических признаков видов лесных растений.
Основная форма роста – качественный признак принадлежности к деревьям, кустарникам, кустарничкам, лианам, травам.
Характер вегетативного разрастания – для кустарников, кустарничков и трав. Признак преимущественно качественный.
Надземные размеры растений. Для основных форм роста – свои количественные градации высоты растений.
Продолжительность жизни и размеры листьев.
Консистенция листовой пластинки.
Теневыносливость (экологический признак).
Отношение растений к влаге (экологический признак).
Отношение растений к плодородию почв (экологический признак).
2. Жизненные формы деревьев
Выделены следующие "группы" жизненных форм одноствольных термопериодических летнеактивных деревьев, большинство из которых уже давно и широко используется лесоводами и геоботаниками:
- - Низкие (ценотически подчиненные) деревья
- - Широколиственные деревья
- - Мелколиственные деревья
- - Пойменные тополя (широколистные пойменные)
- - Пойменные древесные ивы (узколистные пойменные)
- - Летнехвойные деревья
- - Темнохвойные деревья
- - Светлохвойные (боровые) деревья
Между частью групп трудно провести границы, но по формам организации типичных представителей они хорошо различаются. Группы с немногочисленными или экологически сходными представителями можно не дифференцировать. Наиболее сложны и богаты представителями в Приморье широколиственные и низкие (ценотически подчиненные) деревья. Среди первых можно выделить мелколиственно-широколиственные деревья, собственно широколиственные деревья и плотно-широколиственные ксеромезофильные деревья. При дальнейшей дифференциации групп жизненных форм широколиственных деревьев целесообразно учитывать их различия по высоте и по отношению к климатическим и почвенным условиям.
Обращает на себя внимание обособленное положение дуба монгольского. Он обладает умеренным сходством с липой маньчжурской, тополем Максимовича, березой маньчжурской и лиственницей ольгинской. По морфологическим особенностям дуб – явно "широколиственное" дерево. Однако его засухоустойчивость, светолюбие и относительная неприхотливость к почвенным условиям заметно выделяют его из большой группы мегатрофных, теневыносливых и собственно мезофильных широколиственных пород.
Среди низких лиственных деревьев выделяются собственно лесные (ценотически подчиненные) деревья и растения производных (серийных) лесных группировок, существенно отличающиеся экологически и биоморфологически (от Alnus hirsuta до Padus maackii).
По эколого-морфологическим признакам очень тесную группу образуют мелколиственные деревья, хотя они могут существенно отличаться по эколого-ценотическим особенностям в субальпийско-подгольцовых (Betula lanata), таежно-неморальных (В. соstata), дубравных (В. davurica) ценозах и серийных группировках (В. mandshurica и Populus davidiana).
Биоморфологически сходные главнейшие виды хвойных в Приморье – Pinus koraiensis и Picea ajanensis, составляют ядро темнохвойных пород. К ним примыкает Abies nephrolepis, Taxus cuspidata и Abies holophylla.
Лиственница экологически может быть сближена также с таежными лесообразователями, но по характеру приспособленности к среде и по совокупности биоморфологических черт она обнаруживает еще большее сходство с березами. Особняком среди хвойных стоит сосна густоцветковая, или могильная (Pinus funebris), по экологическим особенностям (светолюбие, ксерофитизм) приближающаяся к дубу монгольскому.
Более полно жизненные формы древесных растений Дальнего Востока приведены в схеме ниже.
3. Жизненные формы кустарников
Свыше 30 видов кустарников дальневосточных лесов и смежной с лесами болотной, подгольцовой и лесотундровой растительности были сопоставлены по ряду морфологических и экологических признаков.
По размерам А.Г. Крылов различает высокие (выше 2 м), средней высоты (1-2 м) и низкие кустарники (ниже 1 м). По характеру вегетативного разрастания – аэроксильные (надземное образование новых побеговых систем) и геоксильные (подземное) кустарники.
Так же как и для характеристики трав, для кустарников использованы морфолого-экологические признаки листьев и принадлежность к экологическим группам в отношении влажности и богатства почв. Анализ экологического и ценотического значения отдельных морфологических признаков кустарников приводит к выводу о неравноценности отдельных признаков в разных группах растений.
Заметно обособлены кедровый стланик, полулистопадные олиготрофные рододендроны (Rhododendron dauricum, R. mucronulatum), вечнозеленые олиготрофные приземистые кустарники.
Аэроксильные Philadelphus tenuifolius, Lonicera gibbiflora, L. praeflorens, Acer barbinerve и геоксильные Eleutherococcus senticosus, Corylus mandshurica, Euonymus pauciflora, E. macroptera тоже могут быть объединены по совокупности экологических и морфологических свойств в эколого-ценотическую группу неморальных кустарников.
А.Г. Крылов, вслед за И.Г. Серебряковым (1964), рассматривает кустарники как тип жизненных форм, включающий деревянистые растения средних размеров – высотой от 0,5 до 4(6) м. Одна из отличительных черт биологии большинства кустарников – смена побеговых систем в онтогенезе (возрастном цикле). Она не свойственна большинству деревьев. За счет этой биологической особенности у кустарников постоянно обновляются надземные оси, что при ограниченных размерах этих растений и при недолговечности их осевых побегов позволяет им неограниченно долго сохранять свою роль в сообществе.
Однако этот признак не может быть безоговорочно принят как главная отличительная особенность всех кустарников, так как в условиях, неблагоприятных для активного вегетативного развития, деревянистые растения средних размеров (кустарники) могут длительное время развиваться без смены побеговых систем.
Причислять на этом основании растения скромных размеров (например, лещины) к деревьям вряд ли имеет смысл. Здесь мы имеем дело с кустарниками низкой вегетативной активности. С другой стороны, не следует причислять к кустарникам многоствольные деревья, способные неоднократно давать, порослевые побеги и сменой побеговых систем напоминающие многие кустарники. От кустарников их отличает значительная долговечность надземных осей (стволов) и большие надземные размеры стволов и крон взрослых растений.
При выделении классов жизненных форм кустарников выделены, как обычно, лиственные и хвойные кустарники. Среди лиственных кустарников на уровне подкласса целесообразно разделить вечнозеленые и листопадные растения. В подклассе листопадных кустарников выделены три группы жизненных форм, различающиеся совокупностью признаков. В названиях групп отражены их экологические различия. Биоморфологические особенности, скоррелированные с более узкой адаптацией к почвенно-гидрологическим, а отчасти к климатическим условиям, позволили дифференцировать группы жизненных форм на подгруппы. Группы и подгруппы жизненных форм кустарников выделяются по комплексу эколого-морфологических признаков и представляют собой достаточно естественные совокупности сходно приспособленных растений.
Лещина разнолистная – пример адаптации лесного широколистного кустарника к условиям периодического дефицита влаги (зона дубовых лесов). По сравнению со своими сородичами из неморальных лесов она обладает более плотными светолюбивыми листьями и способностью произрастать на сухих местообитаниях, при оптимуме – на свежих местообитаниях. К сходным условиям адаптирована другой вид подлеска дубняков – леспедеца двуцветная. Биоморфологически она отличается довольно мелкими листьями, быстрым нарастанием тонких и длинных побегов, частично отмирающих в зимний период, поздним и продолжительным цветением. Близкую жизненную форму имеет секуринега полукустарниковая. Однако она никогда не образует таких обильных зарослей на большой площади, как леспедеца.
В схеме 2 приводится классификация жизненных форм кустарников, интересующих нас как спутники лесообразующих пород лесных фитоценозов.
4. Жизненные формы трав и кустарничков
В качестве опыта А.Г. Крыловым (1983) построена классификация трав и кустарничков, доминирующих в покрове лесов Приморья. Сходные жизненные формы или те же виды растений распространены в различных регионах умеренного пояса. Поэтому данная классификация жизненных форм имеет не только методическое значение. Она может использоваться при анализе синузиальной структуры нижних ярусов фитоценозов лесной, бореальной (таежные леса) и неморальной (широколиственные леса) растительности.
Группы, выделенные по характеристикам листового аппарата. Для количественного сопоставления размеров листьев разных групп травянистых растений, имеющего экологическое значение, использованы придержки размеров листьев, применяемые для целей систематики (табл. 1).
Таблица 1
Качественные и количественные характеристики формы и размеров листьев в разных группах травянистых растений
| Качественная характеристика трав по размерам листьев | Значения количественных признаков в разных группах травянистых растений | ||
| "разнотравье" | злаки и осоки | папоротники | |
| Нитевиднолистные | – | Ширина листа меньше 1 мм | – |
| Мелко- или узколистные | Ширина обычного листа меньше 2 см | Ширина листа 1-5 мм | – |
| С листьями среднего размера | Ширина листа от 2 до 5 см | Ширина листа 5-10 мм | Ширина сливающихся долей листьев меньше 2 см |
| Широко- или крупнолистные | Ширина листовой пластинки больше 5 см | Ширина листа больше 10 мм | Ширина сливающихся долей листьев больше 2 см |
По форме листьев злаки и осоки существенно отличаются от "разнотравья", иной характер фотосинтетического аппарата имеют хвощи и папоротники.
По продолжительности жизни листьев различаются растения с однолетними листьями, с перезимовывающими листьями и с дву-многолетними листьями ("вечнозеленые").
По теневыносливости листового аппарата можно различать растения светолюбивые, средне-теневыносливые и глубоко-теневыносливые. По консистенции листа выделяются травы с нежными листьями, с обычными листьями, жестколистные и толстолистные.
По размерам надземных вегетативных органов различаются травы крупные (выше 0,5 м), средние (0,15-0,5 м) и низкие (меньше 0,15 м).
По характеру побегов и способности к вегетативному разрастанию травы можно подразделить на вегетативно малоподвижные, дерновинные, длиннокорневищные и ползучие. Все кустарнички во флоре Дальнего Востока – вегетативно подвижны.
Сопоставление сходства по всем признакам позволило выделить четко очерченные экологические группы и дать им среднюю групповую характеристику по тем же признакам. Для сопоставления использовались также обычные качественные градации по отношению растений к влажности и пищевому режиму почв.
Эколого-морфологические классы. По совокупности признаков с дополнительным учетом эколого-ценотических и биологических особенностей видов выделены наиболее четкие группы жизненных форм. В одну группу выделились высокотравье, крупнотравье и крупные папоротники. Можно также отметить близость к лесо-луговому разнотравью вегетативно малоподвижного широкотравья и крайних по отношению к влажности почв "разнотравных" видов. Отчетливо обособились рыхлодерновинные лесные осоки, таежное мелкотравье, длиннокорневищные папоротники и таежные кустарнички.
1. Высокие и крупные гигромезофильные и мезофильные травы и папоротники („крупнотравье", высокотравье) – результат адаптации к богатым влажным почвам в условиях лугов или разреженных лесов.
Формируемый травяной покров отличается мощным развитием, высокой сомкнутостью и значительной устойчивостью, в связи с чем большинство особей в течение ряда лет занимает лишь свое место. Хорошее развитие получают как подземные, так и надземные органы. У многих представителей ортотропные высокие стебли и крупные листья.
2. Вегетативно малоподвижные мезофильные травы средних размеров ("разнотравье"), приспособленные к сообитанию в сомкнутом травостое на суходольных, довольно богатых почвах. Меньшее плодородие и некоторая сухость почв объясняют уменьшение размеров растений. В класс разнотравья входит и широкотравье.
3. Длиннокорневищные крупные и средних размеров травы местообитаний с неустойчивыми почвенными условиями или переменной ценосредой (например, под пологом леса). Почвенные условия достаточно благоприятны, о чем свидетельствуют значительные размеры растений, но пригодность местообитаний для длительного существования растений на одном месте постоянно нарушается.
Группа включает растения серийных луговых, лугово-болотных группировок или травяного покрова лесов на среднебогатых почвах. Данная группа объединяет очень пестрый набор жизненных форм.
4. Рыхлодерновинные невысокие травы ("осочки") – доминанты травяного покрова на свежих, часто суховатых почвах. Конкуренция со стороны других жизненных форм трав ослаблена слишком дренированными почвами либо снята неблагоприятной лесной ценосредой.
5. Вегетативно подвижное лесное мелкотравье – травы темнохвойной тайги. Развиваются в условиях неблагоприятной световой и почвенной среды, сильно преобразованной сомкнутым древостоем.
С недостатком света и элементов минерального питания коррелируют малые размеры растений, а вегетативная подвижность – приспособление к изменяющимся микросредам в приземном ярусе.
6. Мезофильные вегетативно подвижные кустарнички – иная форма, свойственная напочвенному покрову тайги.
 | В экологических группах может наблюдаться большая или меньшая вариабельность особенностей видов. Выделенные „основные группы" жизненных форм трав можно включить в экологическую систему на правах классов. Далее в классах выявляются более однородные группы жизненных форм – либо стержневые для всего класса, либо обладающие своеобразными эколого-морфологическими чертами, либо представляющие собой переходные звенья от одного класса к другому. Морфологические особенности вышеуказанных групп жизненных форм травянистых лесных растений можно проиллюстрировать схематически. На рис. 1 показаны основные габитуальные отличия классов жизненных форм крупнотравья и разнотравья. Общие отличительные черты крупнотравья – большая величина надземной части растения и крупные листья. В пределах класса осмунд выделяется плотными вайями и мощными длинными корневищами, внешне собственно крупнотравье и высокотравье разнятся лишь размерами. В той и другой группе отдельные представители могут обладать или не обладать вегетативной подвижностью. В классе разнотравья центральное место принадлежит мезофильному лесному и луговому разнотравью: средние размеры растения, средние размеры листьев, факультативная вегетативная подвижность. Габитуально сходное с крупнотравьем крупное ксеромезо-фильное разнотравье экологически гораздо ближе к разнотравью. Специализация к условиям сухих или сырых почв у разнотравья не сопряжена с резкими габитуальными отличиями. К сухим почвам хорошо приспособлена группа плотнолистного разнотравья. Гигрофильное разнотравье характеризуется уменьшением ширины листьев (Sanguisorba parviflora, Pedicularis resupinata). Изображение строения широкотравья трех групп жизненных форм (см. рис. 1) показывает одновременно их сходство (среднего и крупного размера листья, одноярусное расположение листового аппарата) и различия (вегетативная подвижность и неподвижность, надземные размеры растений). Таким образом, если по комплексу количественных и качественных признаков разнотравье и широкотравье различить почти невозможно, то исконно неморально-лесные растения (широкотравье) имеют вполне отчетливые габитуальные различия. |
 | Длиннокорневищные злаки и осоки (рис. 2) представлены 7 схемами строения вегетативных органов растений, соответствующих группам жизненных форм различной экологической специализации. Мезофильные лесные осоки – экологический аналог длинно-корневищного широкотравья. На сырых почвах имеются крупные злаки (Calamagrostis langsdorffii), осоки (Carex dispalata), сопоставимые по размерам с гигрофильным крупнотравьем. Экологическим аналогом ксеро-мезофильного разнотравья является группа плотнолистных крупных ксеро-мезофильных злаков (Spodiopogon sibiricus, Arundinella anomala). В том же направлении адаптирован Pteridium aquilinum. Мезофильные длиннокорневищные травы – характернейшие растения лугов и лесов в условиях поймы, а также серийных ценозов на богатых почвах. Интересно отметить, что идентичную с узколистной крапивой жизненную форму имеют бор развесистый, многие уремные растения, например, котовник маньчжурский. Длиннокорневищные луговые и лесные злаки образуют свою естественную группу жизненных форм. С группой длиннокорневищных мезофильных злаков связаны лесные рыхлодерновинные вейники ксеромезофильной (Calamagrostis arundinacea) и гигромезофильной ( С. obtusata) специализации. |
 | С длиннокорневищными мезофильными лесными папоротниками имеют большое эколого-морфологическое сходство вегетативно подвижное широкотравье, мезофильные лесные осоки и мезофильное лесное мелкотравье (рис. 3). Эти папоротники не только родственны крупным лесным папоротникам, но и имеют с ними много общего в экологических свойствах, что наглядно выражается в частом совместном произрастании тех и других в растительных сообществах. В благоприятных для роста и развития условиях надземные размеры длинно-корневищных папоротников близки к размерам крупных папоротников. На обедненных почвах и в условиях сильного угнетения под пологом сомкнутого древостоя длинно-корневищные папоротники по размерам и жизненной форме приближаются к мелкотравью. Дерновинные "осочки" и ксеромезофильные ирисы представлены тремя близкими группами жизненных форм (см. рис. 3). На схеме во всех группах показан один тип вегетативного разрастания особи. Среди мелких "осочек" у Сагех ussuriensis – иной тип разрастания. За счет побегов на укороченных корневищах формируются небольшие плотные дернинки, от которых отходят длинные корневища, дающие начало новым дернинкам (парциальным кустам). В результате особь или клон уссурийской осоки может иметь обширную рыхлую дерновину, состоящую из множества дерновинок, соединенных длинными корневищами. Но роль в сообществе всех этих осок и ирис одинакова. Представленные на рис. 3 схемы строения мелкотравья показывают достаточно широкую амплитуду приспособленности к условиям увлажнения у растений данного класса жизненных форм: от ксерофильных боровых растений до таежно-болотных гигрофитов. Стержневой группой жизненных форм здесь является вегетативно подвижное лесное мелкотравье, характерное для темнохвойной тайги, иных бореальных лесов и неморальных сообществ. |
Ниже приведен перечень всех классов и групп классификации жизненных форм для трав и кустарничков А.Г. Крылова с указанием видов растений по каждой группе.
Эколого-морфологическая классификация деревьев Тип Деревья
Класс Лиственные деревья
Подкласс Высокие деревья
Группа Широколиственные деревья
Подгруппа Типичные мезофильные широколиственные деревья – Fraxinus mandshurica, Juglans mandshurica, Kalopanax septemlobus, Phellodendron amurense, Тiliа mandshurica, Ulmus laciniata
Подгруппа Мелколиственно-широколиственные деревья – Acer mandshuricum, A. mono, Betula costata, Tilia amurensis, T. sibirica, Ulmus japonica
Подгруппа Плотношироколиственные ксеромезофильные деревья – Quercus mongolica
Группа Мелколиственные деревья – Betula davurica, В. ermanii, В. lanata, В. mandshurica, В. pendula, В. pubescens, Populus davidiana, P. tremula
Группа Широколистные пойменные деревья – Populus koreana, P. laurifolia, P. maximowiczii
Группа Узколистные пойменные деревья – Chosenia arbutifolia, Salix rorida, S. cardiophylla
Подкласс Низкие деревья
Группа Типичные мезофильные лесные деревья – Acer pseudo-sieboldianum, A. tegmentosum, A. ukurunduense, Carpinus cordata, Syringa amurensis, Sorbus sibirica
Группа Гигрофильные широколистные деревья – Alnus hirsuta
Группа Мезофильные деревья серийных сообществ – Padus maackii
Класс Хвойные деревья
Подкласс Высокие деревья
Группа Темнохвойные деревья
Подгруппа бореальные темнохвойные деревья – Abies nephrolepis, A. sibirica, Picea ajanensis, P. koraiensis, P. obovata, Pinus sibirica
Подгруппа Неморальные темнохвойные деревья – Abies holophylla, Pinus koraiensis
Группа Светлохвойные деревья – Pinus funebris, P. sylvestris
Группа Летнехвойные деревья – Larix gmelinii, L. olgensis, L. sibirica
Подкласс Низкие деревья
Группа Теневыносливые лесные вечнозеленые деревья – Taxus cuspidate.
Эколого-морфологическая классификация кустарников
Тип Кустарники
Класс Лиственные кустарники
Подкласс Листопадные кустарники
Группа Мезо- и эвтрофные кустарники
Подгруппа Ксеромезофильные широколистные кустарники
Дубравные – Corylus heterophylla
Подгруппа Ксеромезофильные мелколистные кустарники
Дубравные – Lespedeza bicolor
Подгруппа Мезофильные высокие аэроксильные кустарники
Уремные – Viburnum opulus, V. sargentii
Неморальные – Acer barbinerve
Таежно-подгольцовые – Duschekia fruticosa
Бореально-лесные – Caragana arborescens
Подгруппа Мезофильные высокие геоксильные кустарники
Неморальные – Corylus mandshurica, Eleutherococcus senticosus
Подгруппа Мезофильные лесные ,обычные* кустарники
Неморальные – Euonymus macroptera, E. pauciflora, Lonicera gibbiflora, L. praeflorens, Philadelphus tenuifolius
Бореально-лесные – bonicera altaica, L. edulis, Ribes hispidulum, Rosa acicularis, Spiraea chamaedryfolia, S. media, S. ussuriensis
Бореально-уремные – Ribes nigrum, Sorbaria sorbifolia, Swida alba
Подгруппа Мезогигрофильные кустарники
Лесо-болотные – Spiraea salicifolia
Подгруппа Мезофильные пионерные кустарники
Лесо-уремные – Rubus idaeus, R. sachalinensis
Подгруппа Мезофильные лесные приземистые кустарники
Таежные – Spiraea betulifolia
Неморальные – Ribes maximoviczianum
Подгольцово-лесные – Cotoneaster uniflorus
Группа Олиготрофные мелколистные кустарники
Подгруппа Гигрофильные высокие
Мохово-болотные – Betula ovalifolia
Подгруппа Психрофильные высокие
Таежно-подгольцовые – Betula divaricata
Подгруппа Гигрофильные приземистые
Мохово-болотные – Myrica tomentosa
Подгруппа психрофильные приземистые
Лесотундровые – Betula rotundifoHa
Группа Олиготрофные полулистопадные кустарники
Неморально-боровые – Rhododendron mucronulatum
Горно-боровые – Rhododendron dauricum
Подкласс Вечнозеленые кустарники
Группа Мезотрофные крупные кустарники
Горно-неморальные – Rhododendron fauriei
Подгруппа Психрофильные кустарники
Подгольцовые – Rhododendron aureum
Подгруппа Гигрофильные кустарники
Болотно-таежные – Ledum hypoleucum
Мохово-болотные – Andromeda polifolia, Chamaedaphne calyculata
Класс Хвойные кустарники
Группа Стланики
Подгольцовый стланик – Pinus pumila
Группа – Вегетативно малоподвижные приземистые кустарники
Лесотундровые кустарники – Juniperus pseudosabina, J. sibirica
Классификация жизненных форм трав и кустарничков
Отдел А. Травы
Тип. Травы-автотрофы
Класс I. Крупнотравье
Группа 1. Мезотрофное мезогигрофипьное плотнолистное крупнотравье
Болотно-лесной папоротник – Osmundastrum asiaticum
Группа 2. Мезогигрофильное высокотравье
Лесо-луговое высокотравье – Alfredia cernua, Cirsium heterophyllum, Delphinium elatum
Лугово-уремное высокотравье – Cacalia hastata
Группа 3. Типичное крупнотравье
Субальпийско-луговое крупнотравье – Rhaponticum sp., Phlomis alpina
Луговое крупнотравье – Angelica cincta, Anthriscus sylvestris, Filipendula palmata, F. ulmaria, Heracleum midendorffii, Ligularia fischeri, Polemonium caeruleum, Saussureа latifolia, Senecio cannabifolius, Veratrum lobelianurn.
Лесное крупнотравье – Aconitum septentrionale, Angelica sylvestris, Bupleurum longifolium, Crepis lyrata, C. sibirica, Euphorbia pilo-sa, Heracleum dissectum, Paeonia anomala
Группа 4. Крупные лесные папоротники
Таежные и бореально-лесные папоротники – Athyrium filix-femina, Dryopteris austriaca
Неморальные папоротники – Thorium rubies, Dryopteris bushing, D. coreano-montana, D. filix-mass
Группа 5. Крупные лесные травы
Бореально-лесные травы – Bathyrus gmelinii
Уремно-неморальные травы – Aconitum arcuatum
Класс II. Разнотравье
Группа 1. Крупное ксеромезофильное разнотравье
Луговое крупное разнотравье – Aconitum barbatum, Ligularia glauca
Дубравное крупное разнотравье – Bupleurum longiradiatum, Cimicifuga dahurica, Doellingeria scabra, Hemerocalis middendorfii, Synurus deltoides
Группа 2. Типичное мезофильное разнотравье
Луговое разнотравье – Cimicifuga simplex, Geranium vlassovianurn, Lysimachia davurica, Sanguisorba officinalis, Serratula wolffii, Thalictrum simplex, Trollius asiaticus
Степисто-луговое разнотравье – Adenophora lilifolia, Campanula glomerata, Dracocephalum argunense, D. ruyschiana, Patrinia scabiosifolia
Альпийско-луговое разнотравье – Aquilegia glandulosa, Doronicum altaicum, Dracocephalum altaiense, Sanguisorba aipina
Бореально-лесное разнотравье – Artemisia maximovicziana, Bupleurum multinerve, Galium boreale, Geranium altaiflorum, G. pseudosibiricum, G. sylvaticum, Lamium album, Lathyrus frolovii, L. humilis, L. komarovii, L. vernus, Potentilla fragarioides, Pulmonaria dacica, Ranunculus propinquis, Saussurea controversa, Solidago virgaurea, Thalictrum minus, Trifolium lupinaster, Vicia unijuga
Неморальное разнотравье – Aruncus asiaticus, Thalictrum contortum
Дубравное разнотравье – Adenophora pereskiifolia, Artemisia stolonifera, Galium platygalium, Geranium maximowichii
Боровое разнотравье – Hieracium umbellatum
Гpуппa 3. Ксеромезофильное плотнолистное разнотравье
Плотнолистное дубравное разнотравье – Atractylodes ovata, Dictamnus dasycarpus
Плотнолистное степисто-луговое разнотравье – Platycodon grandiflorus, Veratrum nigrum
Группа 4. Гигрофильное разнотравье
Луговое гигрофильное разнотравье – Pedicularis resupinata, Sanguisorba parviflora, Trollius chinensis
Группа 5. Ксерофильное низкое разнотравье
Боровое гемиэфемероидное разнотравье – Pulsatilla patens
Группа 6. Вегетативно-малоподвижное теневое широкотравье
Типичное неморальное широкотравье – Actaea acuminata, A. spicata, Hylomecon vernalis, Osniorhiza aristata, Saussurea subtriangulata, Smilacina hirta, Stachys sylvatica, Thalictrum filamentosum, Valeriana fauriei.
Группа 7. Вегетативно подвижное широкотравье
Неморальное вегетативно подвижное широкотравье – Anemonoides udensis, Cardamine leucantha, Galium odoratum, Paris hexaphylla. Дубравное широкотравье – Aegopodium podagraria, Chloranthus japonicus, Convallaria keiskei
Бореально-лесное широкотравье – Paris quadrifolia, Polygonatum odoratum
Группа 8. Низкое широкотравье
Неморальное низкое широкотравье – Asarum europaeum, A. sieboldii, Hepatica asiatica
Класс III. Длиннокорневищные травы
Группа 1. Крупные гигрофильные злаки
Луговые гигрофильные злаки – Calamagrostis langsdorffii
Группа 2. Крупные гигрофильные осоки
Болотно-уремные осоки – Carex dispalata
Группа 3. Ксеромезофильные злаки
Степисто-луговые злаки – Arundinella anomala, Spodiopogon sibiricus
Группа 4. Ксеромезофильные папоротники
Лесные папоротники – Pteridium aquilinum
Группа 5. Мегатрофные крупные травы
Сорно-уремные травы – Urtica angustifolia, U. dioica
Группа 6. Мезофильные злаки
Луговые корневищные злаки – Elytrigia repens, Helictotrichon pubescens, Trisetum sibiricum
Бореально-лесные корневищные злаки – Milium effusum, Poa sibirica Неморальные злаки – Brachypodium sylvaticum, Pestuca altissima, Neomolinia mandshurica
Группа 7. Лесные меэофильные папоротники
Таежные папоротники – Diplazium sibiricum, Dryopteris amurensis, Gymnocarpium dryopteris, G. robertianum, Phegopteris connectilis Неморальные папоротники – Adiantum pedatum, Athyrium spinulosum, Deparia pycnosora, Polystichum tripteron
Группа 8. Лесные мезотрофные травы
Болотно-таежные травы – Angelica maximowiczii, Smilacina davurica
Бореально-лесные хвощи – Equisetum pratense, E. sylvaticum
Группа 9. Мегатрофные лесные осоки
Неморальные осоки – Carex campylorhina, С. siderosticta
Таежные осоки – Carex xyphium
Группа 10. Мезотрофные гигрофильные осоки
Таежно-болотные осоки – Carex globularis
Группа 11. Рыхлодерновинные гигромезофильные лесные злаки
Таежные злаки – Calamagrostis obtusata
Группа 12. Рыхлодерновинные ксеромезофильные лесные злаки
Бореально-лесные злаки – Calamagrostis arundinacea
Дубравные злаки – Calamagrostis brachytricha
Класс IV. Рыхлодерновинные ксеромезофильные узколистные травы ("осочки")
Группа 1. Мелкие "осочки"
Дубравные мелкие "осочки" – Carex nanella
Неморальные мелкие "осочки" – Carex ussuriensis
Группа 2. "Осочки" средних размеров
Неморальные "осочки" – Carex: quadriflora
Дубравные "осочки" – Carex hypochlora, С. lanceolata, С. reventa
Бореально-лесные "осочки" – Carex macroura, С. subebracteata, С. van-hueurсkii
Лесостепные "осочки" – Carex pediformis
Группа 3. Плотнолистные травы ("ирисы")
Бореально-лесные ирисы – Iris ruthenica, I. uniflora
Класс V. Вегетативно подвижное мелкотравье
Группа 1. Гигромезофильное мелкотравье
Таежно-болотное мелкотравье – Rubus arcticus, Smilacina trifolia
Группа 2. Теневое мезофильное мелкотравье
Таежное мелкотравье – Chamaepericlymenum canadense, Maianthemum bifolium, Pyrola incarnata, Oxalis acetosella, Trientalis europaea
Неморальное мелкотравье – Galium dahuricum, Trigonotis radicans,
Бореально-лесное мелкотравье – Cruciata krylovii, Myosotis krylovii, Viola biflora
Группа З. Гемиэфемероидное мелкотравье
Неморальное гемиэфемероидное мелкотравье – Jeffersonia dubia
Группа 4. Ксерофильное мелкотравье
Боровое мелкотравье – Antennaria dioica
Класс VI. Одно-двулетние травы
Группа 1. Мезофильные мегатрофные травы
Уремные однолетники – Impatiens noli-tangere
Группа 2. Ксерофильные мезотрофные травы
Дубравные одно-двулетники – Campanula punctata, Melampyrum roseum
Класс VII. Эфемероиды
Группа 1. Мезофильные типичные эфемероиды
Неморальные эфемероиды – Adonis amurensis, Anemonoides amurensis, Corydalis ambigua
Бореально-лесные эфемероиды – Anemonoides altaica, Corydalis bracteata, Erythronium sibiricum
Класс VIII. Ползучие толстолистные травы.
Скально-таежные баданы – Bergenia crassifolia, В. pacifica
Отдел Б. Кустарнички
Класс I. Вечнозеленые кустарнички
Группа 1. Ксеромезофильные олиготрофные кустарнички
Боровые кустарнички – Arctostaphylos uva-ursi, Chimaphila umbellata, Diphasiastrum complanatum, Pyrola chlorantha, Vaccinium vitis-idaea
Группа 2. Мезофильные мезотрофные кустарнички
Таежные кустарнички – Linnaea borealis, Lycopodium annotinum, 1л. clavatum, Orthilia obtusata, O. secunda
Неморальные кустарнички – Chimaphila japonica
Группа З. Гигрофильные олиготрофные кустарнички
Таежно-болотные кустарнички – Ledum palustre
Болотно-лиственнично-таежные кустарнички – Ledum hypoleucum
Группа 4. Психрофильные олиготрофные кустарнички
Лесотундровые кустарнички – Empetrum sibiricum
Класс II. Листопадные кустарнички
Группа 1. Мезофильные олигомезотрофные кустарнички
Таежные кустарнички – Vaccinium myrtillus
Группа 2. Гигрофильные мезоолиготрофные кустарнички
Таежно-болотные кустарнички – Vaccinium uliginosum
Группа 3. Психрофильные мезоолиготрофные кустарнички
Лесотундровые кустарнички – Arctous alpina
botsad.ru










