Макро- и микроэлементы, необходимые для растений, и их физиологическая роль. Микроэлементы и микроэлементы для растений
Элементы для растений или что такое NPK, макроэлементы и микроэлементы.
Уважаемые дачники, мы понимаем с Вами, что почва как среда укоренения и питания растений должна располагать всеми необходимыми для жизни и развития растений веществами и содержать нужные элементы.
Только из земли как из источника и носителя питательных веществ растения черпают силы для роста, цветения и полноценного плодоношения. Однако, получая питательные вещества из почвы, растения постепенно исчерпывают их запас, который при интенсивном возделывании культур не восполняется естественным образом. Поэтому, чтобы поддерживать в почве уровень содержания питательных веществ, достаточный для жизнеобеспечения растений, и предотвратить ее истощение, необходимо компенсировать естественные процессы выноса питательных веществ растениями и вымывания их в нижние слои почвы путем внесения удобрений, восполняя тем самым израсходованный запас ценных элементов.
Элементы для растений. Знакомство
Удобрения играют важную роль в питании растений: они непосредственно воздействуют на режим питания, улучшают свойства почвы, способствуют переводу питательных веществ в доступные для растений формы. Самый сложный вопрос в практике внесения удобрений — определить, где проходит грань между дефицитом питательных веществ, когда растению грозит минеральное голодание, и переизбытком минеральных элементов, влекущим за собой опасное перенасыщение почвы и растений. От установления этой границы зависит оптимальное дозирование удобрений. Задача создания баланса питательных веществ в почве очень сложна. Поэтому, в подготовленных мною материалах, мы постараемся разобраться, что надо знать для определения оптимального уровня питательных веществ и микроэлементов содержащихся в почве, учитывать уровень ее кислотности, от которой во многом зависит степень усвояемости отдельных веществ растениями, а также потребности растений в питательных веществах разных групп. К тому же в течение вегетационного периода потребность растения в определенных питательных веществах меняется в зависимости от того, какие органы растения усиленно растут в данный промежуток времени. Листовой массе и молодым побегам требуется азот, кальций и калий, корневой системе — фосфор и магний, цветам — азот и фосфор. Поэтому изменяющиеся потребности растения в элементах питания на разных стадиях развития также следует учитывать при определении нормы внесения удобрений.
Исходя из выше сказанного, рассмотрим тему удобрений, поэтапно и разберемся для начала, что такое (NPK), макро- и микроэлементы.
Элементы для растений. Что такое NPK
Химические элементы, необходимые растениям, находятся в почве в составе различных соединений. К их числу относятся азот, фосфор, калий, кальций, магний, сера, марганец, бор, медь, цинк, железо и др. Но не все питательные вещества требуются растениям в одинаковых количествах. Одних, например азота, фосфора и калия, требуется больше – их называют макроэлементами; других (магний, серы, кальция) – гораздо меньше; а некоторые (бор, медь, цинк, молибден и др.) нужны в самых незначительных количествах – их называют микроэлементами.
Азот (N) – один из самых необходимых растениям элементов питания. Он входит в состав аминокислот, всех простых и сложных белков, которые являются главной составной частью цитоплазмы растительных клеток, играющих исключительно важную; роль в обмене и передаче наследственных свойств. Азот содержится также в хлорофилле, фосфатидах, алкалоидах, ферментах и во многих других органических веществах растительных клеток. Основная масса азота сосредоточена в органическом веществе почвы, прежде всего в гумусе. Однако получать высокие урожаи только за счет природных запасов азота очень трудно. Поэтому растения требуют пополнения запасов почвенного азота из других источников, прежде всего – внесением органических и минеральных азотных удобрений.
Фосфор (P) входит в состав нуклеиновых кислот и нуклеопротеидов, которые участвуют в построении цитоплазмы и ядра клеток. Он содержится в фитине (запасном веществе семени), который используется как источник фосфора во время прорастания, а также в фосфатидах, сахарофосфатах, витаминах и многих ферментах. В растительной клетке фосфор играет исключительно важную роль в энергетическом обмене, участвует в разнообразных процессах обмена веществ, деления и размножения. В почве фосфор находится в форме органических и минеральных соединений в виде солей кальция, магния, алюминия и железа. Минеральный фосфор, который требуется для питания растений, представлен в основном малодоступными для растений формами. Поэтому внесение фосфорных удобрений эффективно на большинстве почв. Улучшение питания фосфором увеличивает урожайность растений, содержание сахара, крахмала и жира, повышает их зимостойкость.
Калий (K). Его физиологические функции в растительном организме разнообразны. Он нормализует обмен веществ в клетках, повышает устойчивость растений к засухе, положительно влияет на интенсивность фотосинтеза, окислительных процессов, на углеводный и азотный обмен. Повышая активность ферментов, участвующих в углеводном обмене, калий способствует накоплению сахаров, повышает устойчивость культур к заболеваниям. В почве основное количество калия содержится в виде труднодоступных минеральных соединений. Особенно бедны калием легкие почвы. Недостаток калия восполняется внесением органических и минеральных удобрений.
Элементы для растений. Макроэлементы
Для своего роста и развития растения нуждаются также в таких макроэлементах, как магний, кальций и сера.
Кальций имеет большое значение в создании благоприятных для растений физических и биологических свойств почвы.
Магний входит в состав хлорофилла, участвует в образовании углеводов. Недостаток кальция проявляется чаще всего на кислых почвах, магния – на легких кислых почвах.
Сера входит в состав белков, растительных масел, ферментов и витаминов. Под влиянием серы повышается устойчивость растений к низким температурам, засухе, болезням.
Элементы для растений. Микроэлементы
Микроэлементы играют важную роль в питании растений. Они принимают участие в окислительно-восстановительных процессах, углеводном и азотном обмене, образовании хлорофилла, входят в состав многих ферментов и витаминов, влияют на проницаемость клеточных мембран и поступление элементов питания в растения. Недостаток, как и избыток, микроэлементов снижает урожайность и качество сельскохозяйственных культур, может вызвать заболевания человека и домашних животных. Наиболее изучено влияние таких микроэлементов, как бор, медь, цинк, марганец, молибден, меньше кобальт, йод, селен.
Материал подготовил: специалист по садоводству Буйновский О.И.
sadovniki.org
Макро-, мезо-, микроэлементы: источники, взаимодействие, потребности растений
31.01.2017
По оценкам разных исследователей, для питания растений необходимо от 68 до 84 элементов периодической системы Д. И. Менделеева. Роль далеко не всех их изучена досконально. Тем не менее, общепризнано, что определенная часть найденных в растениях и почве элементов является совершенно необходимой для нормального роста и развития растений, получения хороших урожаев.
Все элементы, участвующие в минеральном питании растений, принято классифицировать в зависимости от их содержания в растениях и в почве. Обычно их разделяют на макроэлементы и микроэлементы. По этой классификации, элементы, содержание которых в перерасчете на сухое вещество составляет от сотых долей процента до нескольких десятков процентов, являются макроэлементами. Те элементы, содержание не превышает тысячных долей процента, относят к микроэлементам.
В настоящее время эта классификация дополнена. Часть элементов сейчас относят к мезоэлементам, т.е., по сути, они образуют группу, промежуточную между макро- и микроэлементами. Кроме того, иногда выделяют ультрамикроэлементы. Это те элементы, содержание которых в растениях ничтожно мало, а физиологическая роль и влияние практически не изучены.

Если придерживаться уточненной классификации, то к макроэлементам относятся азот, фосфор и калий, к мезоэлементам – сера, кальций, магний, к микроэлементам – бор, молибден, цинк, медь, кобальт, марганец, барий, кремний, хлор, натрий, титан, серебро, ванадий, железо, никель, селен, литий, йод, алюминий.
Приведенная классификация, как и любая другая, достаточно условна, и те или иные элементы в работах разных авторов порой попадают в разные группы. Кроме того, в тканях некоторых видов растений отдельные микроэлементы содержатся в количествах, характерных для макроэлементов. Тем не менее, для практических целей, т.е. организации минерального питания растений в хозяйственных условиях, эта классификация достаточно удобна и позволяет адекватно оценить роль тех или других элементов в получении урожая, правильно подобрать методы восполнения их недостатка в почве.
Макроэлементы и мезоэлементы необходимы растению в достаточно больших количествах, потому что являются «строительным материалом», в первую очередь, для белков. Микроэлементы входят в состав ферментов, витаминов и т.п. Нормальное развитие и функционирование как отдельных клеток, так и всего растительного организма невозможно без оптимального обеспечения элементами всех этих групп.
Отсутствие или недостаток любого из элементов, необходимых для роста и размножения, вызывает вполне определенные симптомы голодания. Однако, поступая в повышенных дозах, как макро, так и микроэлементы становятся токсичными для растений и употребляющих их людей и животных.
Питательные вещества при корневом питании растения получают из почвы. Основным источником поступления микроэлементов в почву являются материнские почвообразующие породы. При этом почвы очень различаются по содержанию микроэлементов. Так, в моренных лессовидных суглинках содержание кобальта, хрома, стронция в 2 – 2,5 раза больше, а никеля, ванадия, титана, бария, бора, марганца – в 3 – 4 раза больше, чем в песках. Торфяно-болотные почвы бедны микроэлементами. При этом, содержание микроэлементов в почве увеличивается по мере накопления в ней органических веществ. То есть, при внесении навоза, компоста и других органических удобрений, почва обогащается не только макро-, но и микроэлементами.
Растворимость микроэлементов в почвах имеет большое значение для их биологической доступности и способности к перемещению. Тяжелые почвы (как щелочные, так и нейтральные) хорошо удерживают микроэлементы и поэтому медленно поставляют их растениям, что может приводить к нехватке некоторых элементов. Легкие почвы, наоборот, могут быть источником легкодоступных микроэлементов, но при этом их запас быстрее истощается. Поэтому при оценке обеспеченности почв микроэлементами важно учитывать не только их валовое содержание, но и наличие подвижных форм. Причем, разница между этими двумя значениями может быть весьма существенной. Например, бор в подвижной форме составляет лишь 2 – 4% от валового содержания этого микроэлемента, медь, молибден, кобальт, цинк – 10 – 15%.
Обеспеченность почвы микроэлементами меняется в течение вегетационного периода, а также зависит от интенсивности осадков, испарения влаги из почвы и т.д. В зависимости от этих факторов, концентрации микроэлементов в почвенных растворах могут изменяться более чем в 10 раз. Это необходимо учитывать при проведении анализов почвы. При этом концентрации макроэлементов, хотя также зависят от упомянутых факторов, изменяются в меньшей степени.
Перенос растворенных элементов в почве может происходить двумя путями: через почвенный раствор (диффузия) и вместе с движущимся почвенным раствором (вымывание). В зависимости от климата, этот процесс имеет свои особенности. Так, в прохладном влажном климате вымывание микроэлементов вниз по профилю почвы проявляется сильнее, чем их накопление. А в теплом сухом климате более характерно восходящее движение микроэлементов.
Состояние и доступность микроэлементов в почве зависит от ее кислотности. Так, цинк, марганец, медь, железо, кобальт, бор легко выщелачиваются в кислых почвах. Но если pH почвы поднимается выше 7, эти элементы образуют довольно устойчивые соединения. Молибден и селен, наоборот, мобилизуются в щелочных почвах, а в кислых становятся практически нерастворимыми.

Уровень содержания элементов также связан с биологической активностью почв. Низкая концентрация микроэлементов стимулирует увеличение бактерий в почве, а повышенное их содержание оказывает негативное влияние на почвенную микробиоту. Причем, наиболее токсичны микроэлементы для бактерий, фиксирующих свободный азот. В биомассе микроорганизмов микроэлементы могут накапливаться в таких больших концентрациях, что это влияет на уровень их содержания в почве в целом. При этом, связанные микроорганизмами микроэлементы становятся менее доступными для растений. Также менее доступны для растений элементы, фиксированные на оксидах, тогда как адсорбированные на глинистых минералах – наиболее доступные.
В целом, в почвах более половины общего содержания микроэлементов удерживается органическим веществом. Например, на торфяниках у растений нередко проявляются симптомы дефицита цинка, меди, молибдена, марганца. Причина этого – сильное удержание этих элементов нерастворимыми гуминовыми кислотами.
Степень поглощения растениями микроэлементов и интенсивность их роста в значительной степени зависит от наличия в почве макроэлементов – азота, фосфора и калия. Так, повышение уровня азотного питания увеличивает поступление в растения фосфора, калия, кальция, магния, меди, марганца и цинка. Но при избытке азота наблюдается обратная закономерность. Избыточные дозы фосфора снижают поступление в растение меди, железа и марганца. В присутствии фосфатов уменьшается поглощение растениями цинка. Калий может снижать поступление кальция и магния.
Микроэлементы, в свою очередь, влияют на поступление в растения макроэлементов. Так, поступление азота в растения снижается при дефиците железа, марганца и цинка. Положительно влияют на поглощение азота молибден и кобальт. Поглощение растениями фосфора увеличивается при наличии меди, цинка, кальция и молибдена, но уменьшается под влиянием магния и железа. Поступление в растения калия снижается под влиянием меди, марганца, никеля, цинка, молибдена, железа и бора, а возрастает при наличии хлора.
Описанные явления антагонизма и синергизма ионов очень сильно зависят от других факторов – температуры, вида растений, реакции среды, концентрации питательных веществ.
Интенсивность поглощения питательных веществ растениями также сильно зависит от температуры окружающей среды. Оптимальной для этого является температура + 25 — + 30 °С. Если температура поднимается выше + 35 °С либо падает ниже + 10 — + 12 °С, поглощение питательных веществ растениями замедляется, а потом и вовсе приостанавливается до наступления благоприятных условий.
Общеизвестный факт – на одной и той же почве, при одинаковом содержании в ней макро- и микроэлементов растения разных видов чувствуют себя по-разному. Связано это с их неодинаковыми потребностями в элементах питания. Причем, эти потребности различаются даже в те или иные периоды развития одного и того же растения. Например, для питания проростка гораздо важнее резерв микроэлементов в семени, чем их содержание в почве. Но для всех растений и периодов их развития является справедливым правило незаменимости элементов, согласно которому ни один из питательных элементов не может быть заменен другим. Поэтому при недостатке любого макро- или микроэлемента нет смысла пытаться увеличить урожай за счет внесения других элементов. Отсюда же следует, что для успешного восполнения нехватки питательных веществ нужно точно знать, каких именно элементов недостаточно.
Особенно чувствительны к недостатку или избытку питательных элементов молодые растения. В то же время, есть элементы, которые более необходимы растениям именно на первых этапах развития. Например, это относится к фосфору. В фазе активного роста сначала растения больше нуждаются в азоте, но со временем происходит увеличение потребности в калии. В период образования бутонов и цветения особенно важны фосфор и азот, а также бор.
Разные виды сельскохозяйственных культур довольно сильно различаются по чувствительности к дефициту микроэлементов (см. таблицу).



Для практических целей также важным является показатель выноса питательных веществ с урожаем. Относительное содержание элементов минерального питания в основной и побочной продукции разных сельскохозяйственных культур определяется, прежде всего, их видовыми особенностями, а также от сорта и условий выращивания. В частности, капуста, картофель, сахарная свекла, подсолнечник, кормовые корнеплоды для создания более высокого урожая потребляют гораздо больше питательных веществ, чем зерновые. Вынос питательных веществ из почвы возрастает с увеличением урожая. Тем не менее, затраты питательных веществ на единицу продукции при этом уменьшаются.

Все перечисленные особенности следует учитывать, разрабатывая стратегию и текущие планы обеспечения растений в определенном хозяйстве питательными элементами. В то же время, необходимо помнить и о том, что урожай предназначен потребителям. А конечные потребители сельскохозяйственной продукции – люди. И, например, недостаток микроэлементов в плодах растений может отрицательно влиять на здоровье потребителей, как и избыток тех или иных веществ.
agrostory.com
Макро- и микроэлементы для водных растений. Биокатализаторы
Растение извлекает из внешней среды самые разнообразные, необходимые ему для жизни вещества. Питание растений — это получение из внешней среды макро- и микроэлементов, биокатализаторов и переработка их в растительном организме.
Потребность растений в макро- и микроэлементах очень высока.У каждого вида она своя. Данную потребность устанавливают, изучая их химический состав, а также опытным путем, добавляя в водную среду искусственные смеси.Этими методами было установлено, что жизненно необходимыми для успешного роста и размножения растений являются 15 минеральных веществ и три органических: углерод, кислород и водород.
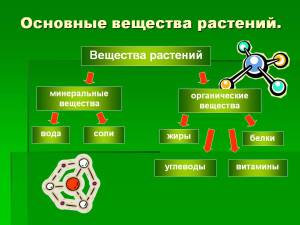
Органические вещества растения получают как СО2, О2 и Н2О.Из пятнадцати же минеральных 7 нужны в относительно больших количествах — это макроэлементы, а 8 — в очень небольших — это микроэлементы.Макроэлементы: фосфор, азот, кальций, калий, сера, магний, железо.Микроэлементы: марганец, бор, цинк, медь, молибден, кобальт, йод, ванадий.
У наземных растений есть и еще один макроэлемент: кремний. Он обеспечивает «скелет» растения, помогая стеблю находиться в прямостоячем состоянии, однако в водной среде поддерживать прямостоячее состояние стебля гораздо проще, поэтому гидрофиты потребляют кремний в значительно меньших количествах. Потребление кремния у водного или болотного растения значительно возрастает, если его стебель «выходит» наружу и начинает расти там.
Есть в тканях растений и очень небольшое количество радиоактивных веществ:  торий, уран, радий и др.Часть специалистов также считает, что растениям еще необходим натрий, но это пока не доказано: установлено лишь, что этот элемент необходим некоторым видам наземных растений.
торий, уран, радий и др.Часть специалистов также считает, что растениям еще необходим натрий, но это пока не доказано: установлено лишь, что этот элемент необходим некоторым видам наземных растений.
Макроэлементы формируют минеральное питание растений. Это основа роста и благополучия гидрофитов.Они напрямую влияют на рост и развитие растений. В практике зоологических хозяйств и ботанических садов чаще всего вносят азотные, фосфатные и калийные удобрения, а также органические удобрения: торф, навоз (является источником сразу азота, калия и фосфора), компост.Как видим, макроэлементы — часто это органические удобрения.Они, действительно, нужны растениям в большом количестве, но и «перебарщивать» с их внесением не стоит: зачастую в аквариуме, особенно в старом, наблюдается избыток азотистых и фосфорных элементов, что само по себе вредно. А вот калий, железо часто в недостатке. Рекомендуются регулярные тесты по этим элементам и их внесение.
 Внесение большого объема макроэлементов в биотоп, особенно в сочетании с повышением температуры может вызвать бурный рост гидрофитов, что не всегда хорошо. Например, в травниках, которые и так уже густо засажены, это приведет к серьезному нарушению задуманных автором композиций растений. Поэтому некоторые профессиональные создатели травников вносят макроэлементы в очень незначительных дозах, а температуру поддерживают на уровне 15 — 20 (!!!) градусов.Еще одна серьезная и широко распространенная неприятность, происходящая при избытке макроэлементов в отечественных биотопах — вспышка водорослей.
Внесение большого объема макроэлементов в биотоп, особенно в сочетании с повышением температуры может вызвать бурный рост гидрофитов, что не всегда хорошо. Например, в травниках, которые и так уже густо засажены, это приведет к серьезному нарушению задуманных автором композиций растений. Поэтому некоторые профессиональные создатели травников вносят макроэлементы в очень незначительных дозах, а температуру поддерживают на уровне 15 — 20 (!!!) градусов.Еще одна серьезная и широко распространенная неприятность, происходящая при избытке макроэлементов в отечественных биотопах — вспышка водорослей.
Помимо высоких концентраций макроэлементов богатые перегноем субстраты содержат также и биотические вещества-биокатализаторы: витамины, ферменты, ауксины, некоторые аминокислоты и другие биотические элементы.Более всего их содержится в черноземах, торфе, богатой разложившимися органическими остатками почве, дрожжах.
Растение само в состоянии синтезировать биокатализаторы из элементов, находящихся в почве/грунте.Однако в условиях обедненного веществами искусственного грунта этот синтез может и не состояться или пройти в недостаточных количествах. И тогда необходимо искусственное внесение биокатализаторов в грунт.Лучшими биокатализаторами являются: водорастворимые витамины группы В: рибофлавин, тиамин В1, биотин, В12, перидоксин В6; никотиновая кислота РР, инозит, параамино-бензойная, фолиевая, пантотеновая и аскорбиновая кислоты; жирорастворимые витамины группы А и D; аминокислоты; ферменты; пуриновые и пиримидиновые основания; антибиотики; ауксин и гетероауксин.Недостаток биотических веществ может вызвать у растений отставание в развитии, и даже заболевания.
Микроэлементы нужны растениям в значительно меньших дозах, однако от этого их роль не уменьшается. Эти вещества получают как из органических соединений, так и из минералов. Морские растения извлекают из воды много йода. Без микроэлементов невозможен рост и цветение гидрофитов, а также они будут желтеть и болеть. Каждый из этих элементов играет в жизни растения важную роль и обычно не может быть заменен другим: при недостатке какого-либо элемента в емкости, гидрофит может быть неспособным усваивать другие элементы.
Как правило, микроэлементы чаще всего и вносятся в биотоп периодически (исключение — железо и калий вносятся периодически из макроэлементов), поскольку аквариумная среда сама не в состоянии их произвести.Начинающему любителю можно рекомендовать покупные комплексы микроэлементов, так как самому ему подобрать точный состав и дозировку будет достаточно сложно, а небольшой избыток микроэлементов в биотопе, в целом, невреден.

www.aqualover.ru
Микроэлементы и макроэлементы для овощей
Для нормального роста и развития растению нужны 9 главных питательных элементов, которые называются биогенными - необходимыми для жизни. Это кислород (О), водород (Н), углерод (С), азот (N), фосфор (Р), калий (К), кальций (Са), магний (Мg), сера (S), железо (Fe) .
Если в пище растения не хватает хотя бы одного из этих элементов, то оно растет только до тех пор, пока не иссякнут его собственные запасы этого элемента, после чего погибает. Кроме этих главных, так называемых макроэлементов в растении есть и другие в незначительном количестве, в рассеянном состоянии, из-за чего их называют рассеянными или микроэлементами. Это бор (В), натрий (Na), кремний (СИ), цинк (Zn), медь (Сu) и др. Часть питательных веществ - углерод и кислород - растения получают из воздуха, а другую, большую - азот, фосфор, калий, кальций, магний и др. - из почвы. При этом они воспринимают необходимые элементы не в чистом виде, а в виде различных химических соединений. Например, фосфор потребляется ими в форме фосфорной кислоты, азот - в форме аммиака или азотистой кислоты. Все эти компоненты в пище растений должны быть в необходимом равновесии. Так, согласно закону Либиха, если в почве не хватает хотя бы одного из главных биогенных элементов (N, Р, К, Са), то растение и остальные из элементов сможет потребить только ограниченно, даже если они в почве в необходимом количестве. Кислород, углерод, водород - это основные строительные материалы. Кислород и углерод растения берут из воздуха. Садовник практически не может влиять на их потребление, разве что только своевременно смывать с растений пыль. Водород растения получают, разлагая воду в процессе фотосинтеза.Азот - один из важнейших для растения элементов. Потребляется он в форме аммиака или азотной кислоты, только бобовые растения обладают способностью с помощью клубеньковых бактерий усваивать азот в чистом виде из атмосферы. Потребность растений в нем велика. Так, плодоносящая яблоня берет из почвы до 7 кг этого элемента в год. Поэтому необходимо поставлять растение азотом регулярно, но одновременно в определенном соотношении с другими питательными веществами. Слишком большие нормы азотных удобрений вызывают буйство вегетативных органов (листья), в результате чего уменьшается сопротивляемость растения болезням. От избытка азота плоды вырастают бледными, теряют свой аромат и плохо хранятся. Передозировка этого элемента приводит и к тому, что на деревьях достаточно долго держится зеленые листья и они бывают плохо подготовленными к зимнему сну. Осенью азотные соединения переходят из листьев в древесную часть растения, где хранятся как запас на весну. В начале вегетации потребление азота бывает повышенным, поскольку он идет на образование новых побегов, листьях и корней. Азот из деревянных элементов быстро расходуется и его запас нужно пополнять, подпитывая растение быстродействующими азотными удобрениями (селитра). Именно поэтому весной фруктовые деревья и ягодники следует удобрять как можно раньше. Недостаток азота в почве проявляется карликовым ростом, бледно зеленым цветом листьев и их преждевременным опадением. Фруктовым деревьям, особенно молодым, щедрая подкормка азотом необходимо.
Кислород, углерод, водород - это основные строительные материалы. Кислород и углерод растения берут из воздуха. Садовник практически не может влиять на их потребление, разве что только своевременно смывать с растений пыль. Водород растения получают, разлагая воду в процессе фотосинтеза.Азот - один из важнейших для растения элементов. Потребляется он в форме аммиака или азотной кислоты, только бобовые растения обладают способностью с помощью клубеньковых бактерий усваивать азот в чистом виде из атмосферы. Потребность растений в нем велика. Так, плодоносящая яблоня берет из почвы до 7 кг этого элемента в год. Поэтому необходимо поставлять растение азотом регулярно, но одновременно в определенном соотношении с другими питательными веществами. Слишком большие нормы азотных удобрений вызывают буйство вегетативных органов (листья), в результате чего уменьшается сопротивляемость растения болезням. От избытка азота плоды вырастают бледными, теряют свой аромат и плохо хранятся. Передозировка этого элемента приводит и к тому, что на деревьях достаточно долго держится зеленые листья и они бывают плохо подготовленными к зимнему сну. Осенью азотные соединения переходят из листьев в древесную часть растения, где хранятся как запас на весну. В начале вегетации потребление азота бывает повышенным, поскольку он идет на образование новых побегов, листьях и корней. Азот из деревянных элементов быстро расходуется и его запас нужно пополнять, подпитывая растение быстродействующими азотными удобрениями (селитра). Именно поэтому весной фруктовые деревья и ягодники следует удобрять как можно раньше. Недостаток азота в почве проявляется карликовым ростом, бледно зеленым цветом листьев и их преждевременным опадением. Фруктовым деревьям, особенно молодым, щедрая подкормка азотом необходимо. Многолетники (фруктовые и декоративные древесные породы, многолетние цветы) надо удобрять азотом только до конца июля, чтобы древесина успела как следует созреть и была хорошо подготовлена к зиме. Большая часть лиственных овощных культур требует щедрого удобрения азотом в течение всего вегетационного периода. Корнеплоды требуют такого удобрения только сначала, позже подкормка азотом делает их менее способными к хранению. Что же касается декоративных растений, то избыток азота в них проявляется увеличением листьях за счет ухудшения цветения (цветы не имеют также ярких цветов).Фосфор особенно важен для вызревания плодов. Растение более всего нуждается в нем во время образования завязей. В отличие от азота, фосфор сокращает вегетационный период. Количество азота и фосфора в питании растения должна быть в определенном соотношении, иначе при недостатке последнего будет особенно сильно проявляться действие азота, будто его был избыток. У деревьев недостаток фосфора тормозит рост черенков, размеры листьев уменьшаются, и на них проступают красные прожилки. Овощи в таких случаях без вкуса, морковь бледная. В цветочных растений при недостатке фосфора образуется меньше цветов, а их цвета теряют выразительность.Калий - элемент, который усиливает морозостойкость растений, способствует их общему хорошему состоянию. Растения, особенно молодые, требуют много калия. Чем больше они потребляют азота, тем больше нужно им калия. Его недостача не проявляется так заметно, как других основных питательных элементов, но растения, которым не хватает калия, плохо переносят засуху, не выдерживают морозов и вообще часто погибают из-за неблагоприятных погодных условий.Кальций, в отличие от других питательных элементов, растение накапливает в своем организме постоянно. Оно использует его как строительный материал, главным образом при образовании древесной ткани и корней. Особенно нужен кальций косточковым растениям, для которых он, наряду с кремнием, - главный компонент при формировании косточек. Когда, например, культивируют абрикосовые и персиковые деревья на кислых почвах, где мало кальция, то сталкиваются с тем, что косточки в плодах или вообще не образуются, или слабая косточка трескается, плод будто разломился, и, как правило, быстро загнивает. При недостатке кальция страдают и корни: медленно растут, образуют мало побочных корешков и корневых волосков. Недостаток кальция наблюдается на кислых почвах, часто замокших и слабовоздушных. Такие почвы следует известковать; помогает также осушения и рыхление. Избыток кальция мешает растениям потреблять фосфор, железо, магний и другие элементы. В таких случаях в почву следует вносить торф. На недостаток и избыток кальция растения реагируют одинаково - хлорозом, то есть бледностью листовой ткани.Магний нужен растению для образования хлорофилла. Он - существенный компонент листовой зелени. При его недостатке листьях тускнеют и даже желтеет. По сравнению с другими питательными элементами, магния растению нужно не очень много. В большинстве случаев потребность в нем удовлетворяет естественное наличие в нем в почве. Недостаток этого элемента, как правило, проявляется тогда, когда есть избыток кальция, ограничивающее усвоение магния растением. Внешне это дает о себе знать типичным хлорозом, а у яблонь - коричневой пятнистостью листьев. Если уменьшить содержание кальция в почве, то войдет в норму и потребление растением магния, который очень важен для цветения, плодоношения, созревания семян. В семенах магния и фосфора очень много, а в зеленом семени его даже в 3 раза больше, чем кальция.Сера. Потребность в сере для различных растений разная. Фруктовым деревьям она нужна в незначительном количестве. Овощные культуры требуют ее больше. Очень требовательны по ее содержанию в почве сельдерей, лук, чеснок, помидоры. Поэтому эти культуры необходимо удобрять сульфатами, в которых, наряду с другими питательными веществами, есть и сера.Железо необходимо для образования хлорофилла и течения других жизненно важных процессов. Однако сами растения используют сравнительно немного, и поэтому, как правило, им хватает естественного содержания этого элемента в почве. Недостаток железа проявляется на зеленых листьях: растения блекнут. Иногда это случается из-за избытка кальция в почве. Недостаток железа можно пополнить, особенно в фруктовом саду, поливая в нем однопроцентным раствором железного купороса.Рассеянные элементы (микроэлементы). Из числа таких элементов особенно большое значение имеет бор, прежде всего для бобовых и картофеля, а также для общего развития остальных растений. Его недостаток в древесных растений проявляется засыханием верхушек, слабым цветением, появлением небольших кожистых скрученных листьев. С нехваткой бора в почве связаны и такие заболевания растений, как черная гниль яблок и камьянистость груш.Кремний имеет значение как строительный материал. Вместе с кальцием он участвует в образовании косточек и деревянных элементов. Что касается хлора, то некоторые растения нуждаются в нем (например, сельдерей), но для большинства растений он противопоказан, поскольку ухудшает хозяйственно полезные признаки. Наиболее уязвимыми бывают виноградная лоза, ягодные культуры, картофель.Медь имеет для растений такое же значение, как и железо; если ее не хватает, на листьях появляются белые пятна.Молибден нужен растениям в минимальных количествах, и поэтому его недостаток у большинства из них практически не проявляется, за исключением цветной капусты, в которой в таком случае не образуется головка. Микроэлементов растениям нужно совсем немного и бывает достаточно их естественного содержания в почве. Однако в саду, где в результате интенсивного выращивания различных культур расход питательных веществ повышен, целесообразно время от времени их пополнять, внося удобрения, в которых есть микроэлементы, или обогащая землю их концентратом.
Многолетники (фруктовые и декоративные древесные породы, многолетние цветы) надо удобрять азотом только до конца июля, чтобы древесина успела как следует созреть и была хорошо подготовлена к зиме. Большая часть лиственных овощных культур требует щедрого удобрения азотом в течение всего вегетационного периода. Корнеплоды требуют такого удобрения только сначала, позже подкормка азотом делает их менее способными к хранению. Что же касается декоративных растений, то избыток азота в них проявляется увеличением листьях за счет ухудшения цветения (цветы не имеют также ярких цветов).Фосфор особенно важен для вызревания плодов. Растение более всего нуждается в нем во время образования завязей. В отличие от азота, фосфор сокращает вегетационный период. Количество азота и фосфора в питании растения должна быть в определенном соотношении, иначе при недостатке последнего будет особенно сильно проявляться действие азота, будто его был избыток. У деревьев недостаток фосфора тормозит рост черенков, размеры листьев уменьшаются, и на них проступают красные прожилки. Овощи в таких случаях без вкуса, морковь бледная. В цветочных растений при недостатке фосфора образуется меньше цветов, а их цвета теряют выразительность.Калий - элемент, который усиливает морозостойкость растений, способствует их общему хорошему состоянию. Растения, особенно молодые, требуют много калия. Чем больше они потребляют азота, тем больше нужно им калия. Его недостача не проявляется так заметно, как других основных питательных элементов, но растения, которым не хватает калия, плохо переносят засуху, не выдерживают морозов и вообще часто погибают из-за неблагоприятных погодных условий.Кальций, в отличие от других питательных элементов, растение накапливает в своем организме постоянно. Оно использует его как строительный материал, главным образом при образовании древесной ткани и корней. Особенно нужен кальций косточковым растениям, для которых он, наряду с кремнием, - главный компонент при формировании косточек. Когда, например, культивируют абрикосовые и персиковые деревья на кислых почвах, где мало кальция, то сталкиваются с тем, что косточки в плодах или вообще не образуются, или слабая косточка трескается, плод будто разломился, и, как правило, быстро загнивает. При недостатке кальция страдают и корни: медленно растут, образуют мало побочных корешков и корневых волосков. Недостаток кальция наблюдается на кислых почвах, часто замокших и слабовоздушных. Такие почвы следует известковать; помогает также осушения и рыхление. Избыток кальция мешает растениям потреблять фосфор, железо, магний и другие элементы. В таких случаях в почву следует вносить торф. На недостаток и избыток кальция растения реагируют одинаково - хлорозом, то есть бледностью листовой ткани.Магний нужен растению для образования хлорофилла. Он - существенный компонент листовой зелени. При его недостатке листьях тускнеют и даже желтеет. По сравнению с другими питательными элементами, магния растению нужно не очень много. В большинстве случаев потребность в нем удовлетворяет естественное наличие в нем в почве. Недостаток этого элемента, как правило, проявляется тогда, когда есть избыток кальция, ограничивающее усвоение магния растением. Внешне это дает о себе знать типичным хлорозом, а у яблонь - коричневой пятнистостью листьев. Если уменьшить содержание кальция в почве, то войдет в норму и потребление растением магния, который очень важен для цветения, плодоношения, созревания семян. В семенах магния и фосфора очень много, а в зеленом семени его даже в 3 раза больше, чем кальция.Сера. Потребность в сере для различных растений разная. Фруктовым деревьям она нужна в незначительном количестве. Овощные культуры требуют ее больше. Очень требовательны по ее содержанию в почве сельдерей, лук, чеснок, помидоры. Поэтому эти культуры необходимо удобрять сульфатами, в которых, наряду с другими питательными веществами, есть и сера.Железо необходимо для образования хлорофилла и течения других жизненно важных процессов. Однако сами растения используют сравнительно немного, и поэтому, как правило, им хватает естественного содержания этого элемента в почве. Недостаток железа проявляется на зеленых листьях: растения блекнут. Иногда это случается из-за избытка кальция в почве. Недостаток железа можно пополнить, особенно в фруктовом саду, поливая в нем однопроцентным раствором железного купороса.Рассеянные элементы (микроэлементы). Из числа таких элементов особенно большое значение имеет бор, прежде всего для бобовых и картофеля, а также для общего развития остальных растений. Его недостаток в древесных растений проявляется засыханием верхушек, слабым цветением, появлением небольших кожистых скрученных листьев. С нехваткой бора в почве связаны и такие заболевания растений, как черная гниль яблок и камьянистость груш.Кремний имеет значение как строительный материал. Вместе с кальцием он участвует в образовании косточек и деревянных элементов. Что касается хлора, то некоторые растения нуждаются в нем (например, сельдерей), но для большинства растений он противопоказан, поскольку ухудшает хозяйственно полезные признаки. Наиболее уязвимыми бывают виноградная лоза, ягодные культуры, картофель.Медь имеет для растений такое же значение, как и железо; если ее не хватает, на листьях появляются белые пятна.Молибден нужен растениям в минимальных количествах, и поэтому его недостаток у большинства из них практически не проявляется, за исключением цветной капусты, в которой в таком случае не образуется головка. Микроэлементов растениям нужно совсем немного и бывает достаточно их естественного содержания в почве. Однако в саду, где в результате интенсивного выращивания различных культур расход питательных веществ повышен, целесообразно время от времени их пополнять, внося удобрения, в которых есть микроэлементы, или обогащая землю их концентратом.www.sotka.info
Значение микроэлементов для растений. Основные микроэлементы потребляются растениями в очень малых количествах, измеряемых тысячными и даже стотысячными долями процента, но незаменимы для их развития. Каждый из перечисленных микроэлементов играет собственную роль, но в ряде случаев они могут заменять друг друга. Основные микроэлементы потребляются растениями в очень малых количествах, измеряемых тысячными и даже стотысячными долями процента, но незаменимы для их развития. Каждый из перечисленных микроэлементов играет собственную роль, но в ряде случаев они могут заменять друг друга.
Внесение органических удобрений очень важно на почвах с низким содержанием микроэлементов. Это торфяные, дерново-подзолистые, легкие по механическому составу почвы и почвы с низким содержанием гумуса. На этих полях опрыскивание растений гуматом надо сочетать с внесением гуматов в почву. От содержания гумуса и pH почвы зависит подвижность микроэлементов, а содержание бора и меди зависит еще и от увлажнения почвы. Гуминовые соединения способны усиливать защитные функции растительного организма. Защитное действие гуматов с наибольшей силой проявляется в экстремальных условиях (высокая или низкая температура, засуха или переувлажнение, недостаточное количество света и кислорода в почве, накопление ядохимикатов). Есть факты выживания растений кукурузы, картофеля в условиях низкой температуры (до -12 градусов С). Гуматы ослабляют или полностью нейтрализуют токсическое и мутагенное действие пестицидов. Под влиянием гуматов растения лучше переносят избыточные дозы удобрений, особенно азотных и повышенные дозы пестицидов. Спектр сельскохозяйственный культур, на которых отмечено повышение выхода продукции после обработки гуматом включает зерновые, картофель, кукурузу, овощи, подсолнечник, сахарную свеклу, плодово-ягодные культуры, виноград, цитрусовые , цветочно-декоративные растения. Благоприятно влияют гуматы не только на количественные показатели роста, но и на качество растительной продукции. Под их влиянием в растениях возрастает содержание витамина С, каротина, рибофлавина, неоцина. В зависимости от культуры прирост составляет от 25 до100%. Увеличивается также содержание белка, крахмала, нуклеиновых кислот, сахаров, что благоприятно сказывается на качестве сельхозпродукции. Роль микроэлементов в питание растений. Изучение минерального питания растительных организмов включает в себя знакомство и с микроэлементами. Химические элементы, которые, входя в состав организмов растений, принимают участие в процессах обмена веществ и обладают выраженной биологической ролью, получили название биогенных элементов. К числу биоэлементов относятся: азот, водород, железо, йод, калий, кальций, кислород, кобальт, кремний, магний, марганец, медь, молибден, натрий, сера, стронций, углерод, фосфор, фтор, хлор, цинк. Помимо общего благоприятного влияния на процессы роста и развития, установлено специфическое воздействие ряда микроэлементов на важнейшие физиологические процессы — например, фотосинтез у растений. Влияние основных микроэлементов на растенияКобальт.Внесение 300 г сернокислого кобальта на 1 га значительно повышает урожай винограда: вес ягод увеличивается на 35%, сахаристость — на 14%, кислотность снижается на 10%. Известкование почв снижает усвояемость растениями кобальта. Так же влияет избыток марганца и железа в почвах; наоборот, фосфор усиливает поступление кобальта в растения.Марганец.Среднее содержание марганца в растениях равно 0,001 %. Марганец служит катализатором процессов дыхания растений, принимает участие в процессе фотосинтеза. Исходя из высокого окислительно-восстановителыюго потенциала марганца можно думать, что марганец играет такую же роль для растительных клеток, как железо — для животных.Благотворное влияние марганца на рост и развитие растений очевидно; так, И. В. Мичурин подметил, что у гибридных сеянцев миндаля под влиянием марганца срок первого плодоношения ускоряется на 6 лет. Этот факт явился первым описанным в литературе случаем замечательного ускорения роста и созревания растений под влиянием микроэлементов. При недостатке марганца в почвах (низком содержании либо неблагоприятных условиях для усвоения его растениями) возникают заболевания растений, характеризующиеся в общем появлением на листьях растений хлоротичных пятен, которые в дальнейшем переходят в очаги некроза (отмирания). Обычно при этом заболевании происходит задержка роста растений и их гибель. У различных видов растений заболевание марганцевой недостаточностью имеет свои специфические проявления и получило соответственные названия. Болезни плодовых растений проявляются в хлорозе листьев (у главной жилки), преимущественно старых (недостаточность железа проявляется главным образом на молодых листьях). Отмирают ветви, светлеют плоды. Сильнее всего поражается груша; вишня и яблоня — меньше.Встречается также серая пятнистость клубники и другие заболевания.Избыток марганца, так же как и его недостаток, неблагоприятно сказывается на растениях.Медь.Медь необходима для жизнедеятельности растительных организмов. Почти вся медь листьев сосредоточена в хлоропластах и тесно связана с процессами фотосинтеза; она участвует в синтезе таких сложных органических соединений, как антоциан, железопорфирины и хлорофилл; медь стабилизирует хлорофилл, предохраняет его от разрушения.Медь входит в качестве структурного компонента в состав соединения с белком (медьпротеида, содержащего 0,3% меди), образуя окислительный фермент полифенолоксидазу. Этот фермент впервые был обнаружен в клубнях картофеля, шампиньонах, а в дальнейшем в составе большинства распространенных растений. Медь способствует синтезу в растениях железосодержащих ферментов, в частности пероксидазы. Установлено положительное влияние меди на синтез белков в растениях и благодаря этому — на водоудерживающую способность растительных тканей. Напротив, при недостатке меди гидрофильность коллоидов тканей уменьшается. Очевидно, вследствие этого медь в виде удобрений имеет значение для придания растениям засухо- и морозоустойчивости, а также, возможно, устойчивости к бактериальным заболеваниям. Болезни недостаточности меди у растений: - экзантема, или суховершинность плодовых деревьев. Поражает цитрусовые , а также яблони, груши, сливы и маслины. У цитрусовых листья достигают больших размеров, молодые побеги изгибаются, на них развиваются вздутия, затем трещины. Пораженные побеги теряют листья и высыхают. Крона деревьев приобретает кустовидную форму. Плоды мелкие с бурыми пятнами и бородавками. Листья имеют сначала ярко-зеленый цвет, а в дальнейшем появляется пятнистость и хлороз. У яблонь заболевание проявляется в отмирании верхушек побегов — наступает увядание и свертывание листьев. Края листьев становятся как бы обожженными. У персиков наступает гибель побегов, ухудшается цветение и завязывание плодов; на листьях появляются крупные хлоротичные пятна. Цинк. Цинк является составным компонентом фермента карбоангидразы. Входя в состав карбоангидразы, цинк влияет на важнейшую фотохимическую реакцию «темновой» утилизации углекислого газа растениями и на процесс выделения СО2, т. е. на процесс дыхания растений. Растения, развивающиеся в условиях недостаточности цинка, бедны хлорофиллом; напротив, листья, богатые хлорофиллом, содержат максимальные количества цинка. В зеленых листьях цинк, возможно, связан с порфиринами. Под влиянием цинка происходит увеличение содержания витамина С, каротина, углеводов и белков в ряде видов растений, цинк усиливает рост корневой системы и положительно сказывается на морозоустойчивости, а также жаро-, засухо- и солеустойчивости растений. Соединения цинка имеют большое значение для процессов плодоношения. Горох, сорго и бобы в водных культурах не дают семян при концентрации цинка в среде 0,005 мг на 1 л и ниже. С повышением концентрации цинка в питательной смеси соответственно число семян увеличивается.
БОР.Бор ускоряет ряд процессов в растении, особенно положительно влияет на прорастание пыльцы и рост пыльцевых трубок в цветках. Недостаток его чаще сказывается на карбонатных, заболоченных, а также произвесткованных почвах, особенно в засушливые годы. Дефицит бора усугубляется при внесении азотных удобрений в избыточном количестве и извести.Борное голодание сопровождается нарушением углеводного и белкового обмена, вследствие чего в тканях накапливаются сахара и аммиачный азот. Это выражается в отмирании верхушечных почек и мелких корешков, осыпании завязи, изменении окраски листьев в красную или и бронзовую, в появлении пятен на плодах. Недостаток бора растения могут испытывать при низком содержании его в почве, а также вследствие способности переходить в труднодоступные для растений соединения.У малины при недостатке бора листья удлиненные, тонкие, с глубокими вырезами, при слабой недостаточности изогнутые, с неровной поверхностью, закручены. Земляника отличается низкорослостью, листья имеют чашевидную форму, сморщенные и коричневые по краям. Усы и корни растут плохо.Как недостаток, так и избыток бора вредны для растений. Избыток проявляется в задержке роста растения и появлении по краям старых листьев сначала желтой каемки, затем эти ткани отмирают, края закручиваются вверх или вниз. Листья приобретают куполообразную форму, а потом засыхают и опадают. Урожайность растений низкая. Особенно чувствительны к избытку бора картофель и огурцы.. Птичий помет также богат микроэлементами: в 100 г сухого вещества содержится марганца 15—38 мг, цинка — 12— 39, кобальта 1-1,3, меди — 1 —1,3, — 0,5, железа — 367—900 мг. Значительная часть элементов питания в помете (азота около 50 %, фосфора — 4 и калия — 60 %) находится в водорастворимой форме. Добавление воды в помет заметно снижает его удобрительную ценность. Поэтому сейчас все стали больше переходить на органику. |
serezha-pankin1.narod2.ru
Макро- и микроэлементы, необходимые для растений, и их физиологическая роль.
Анализ обнаруживает в растениях почти все элементы периодической системы Менделеева. Главные из них – микро- и макроэлементы.
Макроэлементы.
Их содержание в растениях колеблется от 0,01-10% от сырой массы растения. Необходимые макроэлементы растений: азот, фосфор, калий, кальций, магний, сера, натрий, кремний, железо, селен С помощью гравийных и водных культур была показана их необходимость для растений и были разработаны специальные смеси для водных культур.
Азот.
Входит в состав белков, нуклеиновых кислот, АТФ, АДФ, коферментов, хлорофиллов, цитохромов, некоторых липидов, многих витаминов, гормонов роста растений. Азот является составной частью важнейших для жизни веществ. Он непосредственно влияет на рост растений.
Фосфор.
Входит в состав ДНК, РНК, АТФ, коферментов, фосфолипидов, сахарофосфатов, белков, многих других промежуточных продуктов метаболизма. Фосфорсодержащие вещества занимают центральное место в конструктивном и энергетическом обмене. Важна роль фосфора в фотосинтезе и дыхании. Кроме того энергия при фотосинтетическом и окислительном фосфорилировании запасается в макроэргических фосфатных связях АТФ. Фосфор важен для цветения и плодоношения растений.
Калий.
Не входит в состав органического вещества, регулирует состояние цитоплазмы клеток растений, повышая ее проницаемость и уменьшая вязкость, находится в клеточном соке, принимает активное участие в осмотических явлениях клеток, движении устьиц, усиливает биосинтез крахмала, ускоряет процессы фотосинтетического фосфорилирования, отток ассимилятов. Основная роль калия – регуляторная – принимает участие в процессах обмена веществ в растении.
Сера.
Содержится во всех белках, входит в состав аминокислот (метионина, цистеина, цистина), содержится в витаминах (тиамин, биотин), липоевой кислоте, сульфолипидах, коферменте А, чесночных и горчичных маслах. Дисульфидные группы участвуют в образовании третичной структуры белков, а сульфгидрильные - в образовании ферментов с участием НАД и ФАД. Сера играет важную роль в белковом и липидном обменах, в энергетике растений, важна для поддержания структуры мембран тилакоидов хлоропластов.
Кальций.
Содержится в растениях в органических веществах и в ионной форме, входит в состав клеточной стенки растений, в состав хромосом, мембран, стабилизируя их структуру. В свободном виде выступает в качестве антагониста калия – повышает вязкость и снижает проницаемость цитоплазмы, нейтрализует избыток органических кислот в клетках, поддерживает жизнедеятельность меристем.
Магний.
Находится в составе молекулы хлорофилла и хелатов, играет роль в стабилизации структуры рибосом, регулирует состояние цитоплазмы, повышая вязкость и понижая проницаемость цитоплазмы, является кофактором многих ферментов.
Натрий.
Для некоторых групп растений (галофитов) засоленных мест обитания имеет важное значение. Для большинства растений не нужен.
Кремний.
В больших количествах содержится в листьях некоторых древесных пород (в хвое ели), входит в состав клеточных стенок древесины, панциря диатомовых водорослей. Многие растения обходятся без него.
2.Микроэлементы.
Их содержание в растениях достигает 0,00001-0,001% сырой массы. Являются абсолютно необходимыми для жизнедеятельности растений: медь, цинк, бор, марганец, молибден и некоторые другие. Их действие строго специфично, при исключении хотя бы одного из них из питания растений нарушаются процессы жизнедеятельности, невозможна замена одного на другой.
Железо.
Содержится в количестве 0,08%. В качестве кофактора входит в состав ферментов, участвующих в синтезе хлорофилла, входит в состав оксиредуктаз, в ферментный комплекс нитрогеназы, то есть участвует в азотфиксации, содержится в молекулах цитохромов, ферредоксина, участвует в процессе переноса электронов.
Медь.
Встречается в составе ферментов, участвующих в биосинтезе хлорофилла, входит в состав ферментов оксидаз, участвующих в дыхании, в состав белка пластоцианина, активирует нитроредуктазу, то есть участвует в азотном обмене. Недостаток меди вызывает задержку роста и цветения.
Цинк.
Играет важную роль в белковом обмене, входя в состав пептидогидролаз, принимает участие в синтезе индолилуксукной кислоты (гормон растений), влияет на синтез аминокислоты триптофана, активирует ряд ферментов гликолиза и реакции ПФП.
Бор.
Имеет широкий спектр действия. Влияет на деление клеток, способствуя росту корневых и надземных частей растений, участвует в прорастании пыльцы и росте завязи, способствует оттоку углеводов из хлоропластов, повышает эластичность клеточной стенки и засухоустойчивость растений.
Марганец.
В качестве кофактора некоторых ферментов катализирует реакции фотосинтеза и дыхания, участвует в процессе восстановления нитратов, обмене железа, поддерживает структуру мембран тилакоидов, активирует ферменты цикла Кребса, участвует в синтезе м-РНК в ядре.
Молибден.
Играет важную роль в азотном обмене, участвует в процессе азотфиксации, в реакциях биосинтеза белка, аскорбиновой кислоты, способствует лучшему усвоению кальция, росту корневых систем растений. При недостатке молибдена, рост растений тормозится.
Кроме перечисленных микроэлементов важную роль играют в метаболизме растений селен, иод, ванадий, титан, никель.
10. Ядро, строение и функции
Ядро– обязательная часть эукариотической клетки. Это место хранения и воспроизведения наследственной информации. Ядро также служит центром управления обменом веществ и почти всех процессов, происходящих в клетке. Чаще всего в клетках имеется лишь одно ядро, редко — два или несколько. Форма его чаще всего шаровидная или эллипсоидальная. В молодых, особенно меристематических, клетках оно занимает центральное положение, но позднее обычно смещается к оболочке, оттесняемое растущей вакуолью. Снаружи ядро покрыто двойной мембраной – ядерной оболочкой, пронизанной порами, на краях которых наружная мембрана переходит во внутреннюю. Внутреннее содержимое ядра – кариоплазма с погруженными в нее хроматином и ядрышками, и рибосомами. В процессе клеточного деления хроматин все более уплотняется и в конце концов собирается в хромосомы. По химическому составу ядро отличается высоким содержанием ДНК. Основная масса ДНК сосредоточена в Хроматине — особых нуклеопротеидных нитях, рассеянных по всему ядру. В ядре заметно одно или несколько Ядрышек. Подобно хроматину, ядрышки не имеют мембраны и свободно лежат в кариоплазме, состоя в основном из белка. Они содержат РНК и имеют большую плотность, чем ядро. Основная функция ядрышек — синтез некоторых форм РНК и формирование предшественников рибосом
11. С-4 путь фотосинтеза (цикл Хетча-Слека)
С4-путь фотосинтеза (цикл Хетча-Слэка-Карпилова)
Ряд растений, происходящих из тропиков (например,кукуруза, сорго, сахарный тростник), имеют другой типфиксации СО2, принципиально отличающийся от циклаКальвина. Этот путь ассимиляции СО2 был назван С4-путёмфотосинтеза или циклом Хетча-Слэка-Карпилова, по имениавстралийских учёных: М. Хетча, К. Слэка и русскогоисследователя Ю.С. Карпилова, впервые описавших этотцикл а растения с таким путём фотосинтезаназывают С4-растениями.
С4-растения имеют особое анатомическое строениелистьев. У них хорошо развита проводящая система, сосудистые пучки окружены крупными клеткамипаренхимы (клетки обкладки пучков). Листья этих растенийсодержат два разных типа хлоропластов: хлоропластыобычного вида - в клетках мезофилла и большое количествокрупных хлоропластов, часто не имеющих гран, - в клетках,окружающих проводящие пучки (обкладка).
В мелких клетках мезофилла листа происходитпервичное акцептирование СО2 по С4-пути, характернойособенностью которого является то, что СО2присоединяется к фосфоенолпировиноградной кислоте(ФЕП) - трёхуглеродному соединению, содержащемумакроэргическую связь. В результате этогокарбоксилирования образуется четырёхуглероднаящавелевоуксусная кислота (ЩУК), что и позволило назватьэтот путь С4-путь фотосинтеза.
Реакцию карбоксилирования катализирует ферментфосфоенолпируваткарбоксилаза (ФЕП-карбоксилаза), которая может работать при гораздо более низкихконцентрациях СО2, чем рибулозодифосфаткарбоксилаза вцикле Кальвина). Затем образующиеся четырёхуглеродныекислоты диффундируют из клеток мезофилла в клеткиобкладки, где распадаются на пировиноградную кислоту(ПВК) и СО2 с образованием восстановленного НАДФ·Н2.
Освобождающийся СО2 поступает в цикл Кальвина иприсоединяется к рибулозодифосфату, а пировинограднаякислота (ПВК) возвращается в клетки мезофилла, где сучастием АТФ превращается в ФЕП и цикл повторяется.
Таким образом, С4-путь является как быдополнительным насосом (помпой), поставляющим
добавочные порции СО2, повышает его концентрацию в
растении, ибо концентрация СО2 в ассимиляционной ткани
ниже, чем в воздухе является лимитирующим фактором в
процессе фотосинтеза.
Растения С4-типа являются одними из наиболеепродуктивных культур. Так, при оптимальных условиях, интенсивность фотосинтеза у кукурузы (С4-растение) в 2-2,5 раза выше, чем у пшеницы (С3-растение).
Хлоропласты разных типов клеток характеризуются иразным типом фосфорилирования. В клетках мезофилла восновном идёт нециклическое фосфорилирование иобразуется НАДФ·Н, необходимый для цикла Кальвина, идущего в клетках обкладки. В хлоропластах клетокобкладки осуществляется циклическое фосфорилирование.
К этому можно добавить, что положение клетокобкладки создаёт условия для передачи конечныхпродуктов фотосинтеза (в частности, сахарозы) непосредственно в ситовидные трубки флоэмы, по которымэти продукты могут затем транспортироваться в другиечасти растения.
С4-растения имеют ещё ряд преимуществ. У них почти
отсутствует фотодыхание, растения же С3-типахарактеризуются высокой интенсивностью этого процесса.
Под фотодыханием понимают процесс поглощения О2 ивыделения СО2 активируемый светом, при котором может"сжигаться" до 50% органических веществ, образующихся в
процессе фотосинтеза. При этом не происходит синтезАТФ.
12. Рост растений, типы роста
У многоклеточных растений в отличие от животных рост (за исключением ранних стадий развития зародыша) происходит только в определенных участках, которые называются меристемами.
Меристемы – это зоны в растительном организме, где происходит регулярное размножение растительных клеток. Эти зоны расположены апикально, т. е. на вершине растущего органа (в главных и боковых побегах и корнях), базипетально (в листьях и междоузлиях), или интеркалярно, например, над узлами в саломине злаков. Интеркалярная – это вставочная меристема (табл. 6.1).
Между листом и стеблем в пазухах листьев, закладываются пазушные почки. Пазушные почки, которые длительное время не дают побегов, называют спящими; при определенных условиях они пробуждаются и из них развиваются побеги.
Таблица 6.1. Типы меристем и их функции
| Типы меристем | Местонахождение | Роль |
| Апикальная | В кончиках корней и побегов | Обеспечивает первичный рост; образует первичное тело растения |
| Латеральная | В более старых частях растений, лежит параллельно длинной оси органа (пробковый камбий) | Обеспечивает вторичный рост. Васкулярный камбий дает начало вторичным проводящим тканям; образуется перидерма, которая замещает эпидермис и содержит пробку |
| Интеркалярная | Между участками постоянных тканей, например в узлах многих однодольных | Делает возможным рост в длину в промежуточных участках. Это характерно для растений, у которых апикальные участки повреждаются (объедание животными злаков и т. п.) |
Латеральная (камбий) меристема лежит (параллельно) вдоль длинной оси органа (например, пробковый камбий) и обеспечивает утолщение.
Внутренние физиолого-биохимические реакции, обеспечивают координированный ход ростового процесса на всех этапах жизни, определяют механизмы роста. Различают первичные и вторичные механизмы роста.
К первичным механизмам роста относят физиолого-биохимические реакции, которые лежат в основе начальных этапов ростового процесса (лаг-фаза) и фазы ускоренного роста (логарифмическая фаза). К этим же механизмам относят электрофизиологические, гормональные и генетические реакции, которые запускают и поддерживают нормальный ход роста клеток, тканей и органов.
Вторичные механизмы роста – это физиолого-биохимические реакции, которые участвуют в нормальном ходе роста (лог-фаза и фаза замедления роста) и происходят в процессе жизнедеятельности растений. К ним относят корреляции между органами, донор-акцепторные связи, метаболическая координация между ростом и другими физиологическими процессами (фотосинтез, транспорт, запасание веществ, стресс).
Таким образом, чаще всего существуют два типа роста: первичный и вторичный. В результате первичного роста может образоваться целое растение (для большинства однолетних и травянистых двудольных это единственный тип роста). В нем участвует апикальная, а иногда и интерполярная меристемы.
У некоторых растениях за первичным ростом идет вторичный рост, в котором участвуют латеральные меристемы. Он в большей мере характерен для кустарников и деревьев. У ряда травянистых растений наблюдается вторичное утолщение стебля, например, развитие дополнительных проводящих пучков у подсолнечника.
Кроме этого, различают еще диффузионный рост. Это рост во время деления всех клеток.
13. Пластиды, строение, функции
Пластиды являются основными цитоплазматическими органеллами клеток автотрофных растений. Название происходит от греческого слова «plastos», что в переводе означает «вылепленный».
Главная функция пластид – синтез органических веществ, благодаря наличию собственных ДНК и РНК и структур белкового синтеза. В пластидах также содержатся пигменты, обусловливающие их цвет. Все виды данных органелл имеют сложное внутреннее строение. Снаружи пластиду покрывают две элементарные мембраны, имеется система внутренних мембран, погруженных в строму или матрикс.
Классификация пластид по окраске и выполняемой функции подразумевает деление этих органоидов на три типа: хлоропласты, лейкопласты и хромопласты. Пластиды водорослей именуются хроматофорами.
Хлоропласты – это зеленые пластиды высших растений, содержащие хлорофилл – фотосинтезирующий пигмент. Представляют собой тельца округлой формы размерами от 4 до 10 мкм. Химический состав хлоропласта: примерно 50% белка, 35% жиров, 7% пигментов, малое количество ДНК и РНК. У представителей разных групп растений комплекс пигментов, определяющих окраску и принимающих участие в фотосинтезе, отличается. Это подтипы хлорофилла и каротиноиды (ксантофилл и каротин). При рассматривании под световым микроскопом видна зернистая структура пластид – это граны. Под электронным микроскопом наблюдаются небольшие прозрачные уплощенные мешочки (цистерны, или граны), образованные белково-липидной мембраной и располагающиеся в непосредственно в строме. Причем некоторые из них сгруппированы в пачки, похожие на столбики монет (тилакоиды гран), другие, более крупные находятся между тилакоидами. Благодаря такому строению, увеличивается активная синтезирующая поверхность липидно-белково-пигментного комплекса гран, в котором на свету происходит фотосинтез.
Хромопласты – пластиды, окраска которых бывает желтого, оранжевого или красного цвета, что обусловлено накоплением в них каротиноидов. Благодаря наличию хромопластов, характерную окраску имеют осенние листья, лепестки цветов, созревшие плоды (помидоры, яблоки). Данные органоиды могут быть различной формы – округлой, многоугольной, иногда игольчатой.
Лейкопласты представляют собой бесцветные пластиды, основная функция которых обычно запасающая. Размеры этих органелл относительно небольшие. Они округлой либо слегка продолговатой формы, характерны для всех живых клеток растений. В лейкопластах осуществляется синтез из простых соединений более сложных – крахмала, жиров, белков, которые сохраняются про запас в клубнях, корнях, семенах, плодах. Под электронным микроскопом заметно, что каждый лейкопласт покрыт двухслойной мембраной, в строме есть только один или небольшое число выростов мембраны, основное пространство заполнено органическими веществами. В зависимости от того, какие вещества накапливаются в строме, лейкопласты делят на амилопласты, протеинопласты и элеопласты.
Все виды пластид имеют общее происхождение и способны переходить из одного вида в другой. Так, превращение лейкопластов в хлоропласты наблюдается при позеленении картофельных клубней на свету, а в осенний период в хлоропластах зеленых листьев разрушается хлорофилл, и они трансформируются в хромопласты, что проявляется пожелтением листьев. В каждой определенной клетке растения может быть только один вид пластид.
14. Транспирация, ее значение
В основе расходования воды растительным организмом лежит процесс испарения — переход воды из жидкого в парообразное состояние, происходящий при соприкосновении органов растения с ненасыщенной водой атмосферой. Однако этот процесс осложнен физиологическими и анатомическими особенностями растения, и его называют транспирацией.
Количество воды, испаряемой растением, во много раз превосходит объем содержащейся в нем воды. Экономный расход воды составляет одну из важнейших проблем сельскохозяйственной практики. К. А. Тимирязев назвал транспирацию, в том объеме, в каком она идет, необходимым физиологическим злом. Действительно, в обычно протекающих размерах транспирация не является необходимой. Так, если выращивать растения в условиях высокой и низкой влажности воздуха, то, естественно, в первом случае транспирация будет идти со значительно меньшей интенсивностью. Однако рост растений будет одинаков или даже лучше там, где влажность воздуха выше, а транспирация меньше. Вместе с тем транспирация в определенном объеме полезна растительному организму.
Рис.1. Транспирация
Транспирация спасает растение от перегрева, который ему грозит на прямом солнечном свете. Температура сильно транспирирующего листа может примерно на 7 °С быть ниже температуры листа завядающего, нетранспирирующего. Это особенно важно в связи с тем, что перегрев, разрушая хлоропласты, резко снижает процесс фотосинтеза (оптимальная температура для процесса фотосинтеза около 30—33 °С). Именно благодаря высокой транспирирующей способности многие растения хорошо переносят повышенную температуру.
Транспирация создает непрерывный ток воды из корневой системы к листьям, который связывает все органы растения в единое целое. С транспирационным током передвигаются растворимые минеральные и частично органические питательные вещества, при этом, чем интенсивнее транспирация, тем быстрее идет этот процесс. Как уже говорилось, механизм поступления питательных веществ и воды в клетку различен. Однако некоторое количество питательных веществ может поступать пассивно, и этот процесс может ускоряться с увеличением транспирации.
Различают два типа транспирации: устьичную — испарение воды через устьица и кутикулярную — испарение воды через всю поверхность листовой пластинки. Впервые разграничение на кутикулярную и устьичную транспирацию было введено в 1877 г. В том, что действительно испарение идет не только через устьица, но и через кутикулу, легко убедиться. Так, если взять листья, у которых устьица расположены только с нижней стороны (например, листья яблони), и замазать эту сторону вазелином, то испарение воды будет продолжаться, хотя и в значительно уменьшенном размере. Следовательно, определенное количество воды испаряется через кутикулу.
Кутикулярная транспирация
Снаружи листья имеют однослойный эпидермис, внешние стенки клеток которого покрыты кутикулой и воском, образующие эффективный барьер на пути движения воды. На поверхности листьев часто развиты волоски, которые также влияют на водный режим листа, так как снижают скорость движения воздуха над его поверхностью и рассеивают свет и тем самым уменьшают потери воды за счет транспирации.
Интенсивность кутикулярной транспирации варьирует у разных видов растений. У молодых листьев с тонкой кутикулой она может составлять около половины всей транспирации. У зрелых листьев с более мощной кутикулой кутикулярная транспирация равна 1/10 общей транспирации. В стареющих листьях из-за повреждения кутикулы она может возрастать. Таким образом, кутикулярная транспирация регулируется главным образом толщиной и целостностью кутикулы и других защитных покровных слоев на поверхности листьев. Кутикулярная транспирация обычно составляет около 10% от общей потери воды листом.
Однако в некоторых случаях у растений, листья которых характеризуются слабым развитием кутикулы, доля этого вида транспирации может повышаться до 30%. Имеет значение также возраст листа. Молодые листья, как правило, имеют слабо развитую кутикулу и, следовательно, более интенсивную кутикулярную транспирацию. У старых листьев доля кутикулярной транспирации снова возрастает, так как, хотя кутикула и сохраняет достаточную толщину, в ней появляются трещины, через которые легко проходят пары воды. Трещины в кутикуле могут появляться и после временного завядания листьев, благодаря чему транспирация усиливается. Имеются данные, что кутикулярная транспирация меньше зависит от условий внешней среды по сравнению с устьичной.
Устьичная транспирация
Основная часть воды испаряется через устьица. Устьица играют важную роль в газообмене между листом и атмосферой, так как являются основным путем для водяного пара, углекислого газа и кислорода. Устьица находятся на обеих сторонах листа. Есть виды растений, у которых устьица располагаются только на нижней стороне листа. В среднем число устьиц колеблется от 50 до 500 на 1 мм². Транспирация через устьица идет почти с такой же скоростью, как и с поверхности чистой воды. Это объясняется законом И. Стефана: через малые отверстия скорость диффузии газов пропорциональна не площади отверстия, а диаметру или длине окружности. Поэтому, хотя площадь устьичных отверстий мала по отношению к площади всего листа (0,5-2 %), испарение воды через устьица идет очень интенсивно.
Транспирация слагается из двух процессов:
1. передвижения воды в листе из сосудов ксилемы по симпласту и, преимущественно, по клеточным стенкам, так как в стенках транспорт воды встречает меньшее сопротивление
2. испарения воды из клеточных стенок в межклетники и подъустьичные полости с последующей диффузией в окружающую атмосферу через устьичные щели.
Чем меньше относительная влажность атмосферного воздуха, тем ниже его водный потенциал. Если водный потенциал воздуха меньше водного потенциала подъустьичных полостей, то молекулы воды испаряются наружу.
Основным фактором, влияющим на открывание и закрывание устьиц, является содержание воды в листе, в том числе и в замыкающих клетках устьиц. Клеточные стенки замыкающих клеток имеют неодинаковую толщину. Внутренняя часть стенки, примыкающая к устьичной щели, более толстая, а внешняя — более тонкая. По мере того как замыкающая клетка осмотически поглощает воду, более тонкая и эластичная часть ее клеточной стенки растягивается и оттягивает внутреннюю часть стенки. Замыкающие клетки принимают полукруглую форму и устьица раскрываются. При недостатке воды замыкающие клетки выпрямляются и устьичная щель закрывается. Кроме того, по мере увеличения водного дефицита в тканях растения повышается концентрация ингибитора роста абсцизовой кислоты. Она подавляет деятельность Н+ - насосов в плазмалемме замыкающих клеток, вследствие чего снижается их тургор и устьица закрываются. Абсцизовая кислота также ингибирует синтез фермента α-амилазы, что приводит к снижению гидролиза крахмала. По сравнению с низкомолекулярными углеводами крахмал не является осмотически активным веществом, поэтому сосущая сила замыкающих клеток уменьшается, и устьица закрываются.
В отличие от других клеток эпидермиса замыкающие клетки устьиц содержат хлоропласты. Синтез углеводов в процессе фотосинтеза в замыкающих клетках увеличивает их сосущую силу и вызывает поглощение воды, способствуя этим открыванию устьиц.
Состояние устьиц зависит от углекислого газа. Если концентрация СО2 в подъустьичной полости падает ниже 0,03%, тургор замыкающих клеток увеличивается и устьица открываются. Повышение концентрации СО2 в воздухе вызывает закрытие устьиц. Это происходит в межклетниках листа ночью, когда в результате отсутствия фотосинтеза и продолжающегося дыхания уровень углекислого газа в тканях повышается. Такое влияние углекислого газа объясняет, почему ночью устьица закрыты и открываются с восходом солнца. Сдвиг рН в щелочную сторону вследствие уменьшения концентрации СО2 увеличивает активность ферментов, участвующих в распаде крахмала, тогда как при кислом рН при повышении содержания СО2 в межклетниках повышается активность ферментов, катализирующих синтез крахмала.
На свету замыкающие клетки устьиц содержат значительно больше калия, чем в темноте. При открывании устьиц содержание калия в замыкающих клетках увеличивается в 4 раза при одновременном снижении его содержания в сопутствующих клетках. Установлено повышение содержания АТФ в замыкающих клетках устьиц в процессе их открывания. АТФ, образованная в процессе фотосинтетического фосфорилирования в замыкающих клетках, используется для усиления поступления калия. Усиленное поступление ионов калия повышает сосущую силу замыкающих клеток. В темноте ионы калия выделяются из замыкающих клеток и устьица закрываются.
Периодичность суточного хода транспирации наблюдается у многих растений, но у разных видов растений устьица функционируют неодинаково. У деревьев, теневыносливых растений, многих злаков и других гидростабильных видов с совершенной регуляцией устьичной транспирации испарение воды начинается на рассвете, достигает максимума в утренние часы. В полдень транспирация снижается и вновь увеличивается в предвечерние часы при снижении температуры воздуха. Такой ход транспирации приводит к незначительным суточным изменениям осмотического давления и содержания воды в листьях. У видов растений, способных переносить резкие изменения содержания воды в клетках в течение дня, то есть у гидролабильных видов, наблюдается одновершинный суточный ход транспирации с максимумом в полуденные часы. В обоих случаях ночью транспирация минимальна или полностью прекращается.
По способности регулировать свой водный обмен, растения делят на пойкилогидрические и гомойогидрические. Пойкилогидрическими (от греч. poikilos — различный, разнообразный и hydor — вода) называются растения, которые не могут сами регулировать свой водообмен. К этой группе относятся почвенные водоросли, лишайники, мхи, папоротники и некоторые покрытосеменные. Гомойогидрическими (от греч. homoios — сходный, одинаковый и hydor — вода) называются растения, которые регулируют свой водообмен. Гомойгидрическими являются покрытосеменные растения.
Различают два типа регуляции транспирации: устьичный и вне-устьичный. Устьичная регуляция осуществляется с помощью открывания и закрывания устьиц. Закрывание устьиц наполовину мало влияет на интенсивность транспирации, что вытекает из закона Стефана. Полное их закрывание сокращает транспирацию примерно на 90 %.
15. Физиологическая роль азота в растении. Круговорот азота в природе.
Азот - важный строительный материал для аминокислот, белков, нуклеиновых кислот и других соединений. Без него жизнь растительного организма была бы невозможна. При недостатке азота в почве уменьшается содержание зеленых пигментов, бледнеют листья, замедляется рост растения. А недостаток бывает часто ощутимый. Ведь в почве азота содержится всего от 0,02 до 0,5 процента и то лишь благодаря деятельности микроорганизмов некоторых растений и разложению органических веществ. И это в то время, когда миллионы тонн азота в атмосфере "давят" на поверхность Земли: над каждым гектаром почвы, образно говоря, висит 80 тысяч тонн относительно инертного газа! В чем же дело? А дело в том, что в воздухе азот находится в молекулярном состоянии, то есть в бездействии. И потому вполне оправдывает свое название: в переводе с греческого азот значит "безжизненный". Элементом жизни он становится только в химических соединениях - легкорастворимых азотнокислых и аммиачных солях. Однако связанного, даже в простые окислы, азота нет в воздухе. Бывает, что эти соединения образуются в небольшом количестве при вспышках молний. В почве растворимых солей азота тоже мало, потому что они легко вымываются. Получается парадоксальная картина: растения, буквально купающиеся в азоте, одновременно голодают из-за его нехватки. А без азота невозможен синтез белков и, следовательно, жизнь!
Каким же путем свободный азот попадает в почву в виде связанных соединений? Основные его поставщики - некоторые виды свободноживущих бактерий, среди которых, например, есть азотобактер, фиксирующая азот в кислородных условиях, или клебзилла, осуществляющая этот процесс только без кислорода. А бактерии рода ризобиум могут связывать безжизненный газ из воздуха лишь в симбиозе с определенными высшими растениями, в основном с бобовыми. Между прочим, бобовые растения не испытывают нехватки азота благодаря тем самым бактериям ризобиум. Их деятельность известна с древнейших времен. Уже тогда было описано благоприятное влияние бобовых растений - сои, земляных орехов, бобов, гороха, клевера - на плодородие почвы. Объясняется это способностью бактерий ризобиум образовывать клубеньки на корнях бобовых. Клубеньки представляют собой расширенные клетки, наполненные бактериями. Каждый вид бобовых связан с определенными бактериями. Так, бактерии, образующие клубеньки на корнях сои, не образуют их на люцерне. Это надо учитывать при искусственном размножении азотфиксирующих бактерии для повышения плодородия полей.
В Бразилии был зафиксирован интересный факт. Среди зерновых, выращиваемых на бедной азотом почве, некоторые растения были выше остальных. На их корнях обнаружили особые бактерии. Это натолкнуло на мысль о том, что не только бобовые могут обходиться без удобрений, но и другие растения, в частности зерновые.
В воде поставщиками связанного азота служат синезеленые водоросли (их теперь называют цианобактериями). Однако и на суше они выполняют эту функцию: вместе с азотфиксирующими бактериями повышают плодородие рисовых полей, заливаемых водой. Могут связывать азот и лишайники - симбиоз гриба и водоросли. Лишайники играют большую роль в экологическом равновесии, так как они первыми поселяются в местах, бедных питательными веществами, и накапливают азот прямо из воздуха.
Для растений азот – дефицитный элемент. Если некоторые микроорганизмы способны усваивать атмосферный азот, то растения могут использовать лишь азот минеральный. Растения никогда не выделяют азотистые соединения как продукты отброса и там, где это возможно, азотистые соединения заменены на безазотистые вещества. В почве азот находится в трех формах: Nh5+, NO3- и органический азот. Для растений наиболее доступной формой является NO3-, менее доступна форма Nh5+.
Для того чтобы сделать доступными для питания растений основные запасы азота в гумусе, необходимо разложить органическое вещество почвы. Этот процесс невозможен без участия различных микроорганизмов.
Процесс превращения органического азота почвы в NН4+ носит название аммонификации. Он осуществляется гетеротрофными микроорганизмами (питаются готовыми органическими веществами) и схематически может быть представлен следующим образом:
органический азот почв → RNН2 + СО2 + побочные продукты, RNН2 + Н2О = Nh4 + RОН, NН3 + Н2О = Nh5+ + ОH –.
Биологическое окисление NН3 или Nh5+ до NO3 – называется нитрификацией.
Еще в 1870 г. Шлезинг и Мюнц доказали опытным путем биологическую природу нитрификации. Для этого они добавляли к сточным водам хлороформ, в результате чего прекращалось окисление аммиака. Основная заслуга в раскрытии механизма нитрификации и выделении осуществляющих этот процесс микроорганизмов принадлежит выдающемуся российскому микробиологу С.Н.Виноградскому. В 1889 г. он открыл бактерии нитрификации и определил две фазы этого процесса.
Бактерии первой фазы окисляют аммиак до азотистой кислоты:
2Nh4 + 3O2 = 2HNO2 + 2h3O.
К ним относят бактерии следующих родов: Nitrosomonas, Nitrosocystis, Nitrosolobus, Nitrosospira. Из них наиболее изучен вид Nitrosomonas europaea.
2HNO2 + О2 = 2НNO3.
Среди бактерий, осуществляющих это превращение, различают три рода: Nitrobacter, Nitrospina, Nitrococcus.
Азотистая кислота в почве не накапливается, т. к. обе группы бактерий обычно функционируют последовательно. Ежегодно минерализуется 0,4–4 т/га органического вещества почвы, в основном гумуса. В результате образуется от 20 до 200 кг/га минерального азота. Таким образом, в почве накапливаются ионы Nh5+ и NO3 –, азот которых доступен для усваивания растениями.
Ионы Nh5+ не слишком подвижны, хорошо адсорбируются анионами, трудно вымываются осадками, и поэтому в почвенном растворе их концентрация значительно выше, чем NO3 –. В почвах, богатых глинистыми минералами, содержание азота в форме Nh5+ может достигать 2–3 т/га. В верхних слоях почвы фиксированный азот Nh5+ составляет 5–6% общего содержания азота в почве, в более глубоких слоях, где выше содержание глинистых частиц, – до 20% и более.
Анионы NO3 –, напротив, подвижны, плохо фиксируются в почве, легко вымываются почвенными водами в более глубокие слои и водоемы. Содержание нитратов в почве особенно возрастает весной, когда создаются условия, благоприятствующие деятельности нитрифицирующих бактерий. Количество азота NO3 – в почвенном растворе сильно варьируется в зависимости от скорости поглощения нитратов растениями, интенсивности микробиологических процессов и процессов вымывания.
В природе азотфиксирующие бактерии и водоросли по-разному приспособились защищать свой главный фермент от кислорода. У анаэробных бактерий этой проблемы не существует. Обитая в глубоких слоях почвы, они могут жить только в бескислородных условиях. Цианобактерии имеют специализированные толстые клетки, которые содержат особое вещество, препятствующее проникновению кислорода. Сходная защита возникла у аэробных (живущих в кислородных условиях) бактерий, у которых кислород при поступлении в клетку восстанавливается до воды.
Бобовые растения вместе с симбиотическими бактериями также выработали интересную защитную систему от кислорода, улавливаемого прежде, чем он достигнет клетки. Его "подстерегает" особый белок - леггемоглобин, синтезируемый в корневых клубеньках. Подобно гемоглобину животных, этот белок способен связывать кислород и отдавать его при необходимости.
Возможно, такая высокая чувствительность нитрогеназы к кислороду, а также потребность в большом количестве энергии для ее работы является причиной того, что в природе не так уж много организмов, способных осуществлять биологическую фиксацию азота.
При недостатке азота в среде обитания тормозится рост растений, ослабляется образование боковых побегов
cyberpedia.su
Микроэлементы. Медь
06.02.2017
Физиологическая роль микроэлемента. К наиболее важным микроэлементам, оказывающим ощутимое влияние на рост и развитие сельскохозяйственных культур, относится медь (Сu). Вместе с марганцем она участвует в окислительно-восстановительных процессах: повышает интенсивность фотосинтеза и образования хлорофилла, способствует активизации углеводного и азотного обмена. Достаточное количество меди в аграрных культурах улучшает сопротивляемость растений грибковым и бактериальным заболеваниям, увеличивает показатели засухо- и морозоустойчивости, а также стойкости к полеганию. Присутствие этого элемента в растительном питании способствует увеличению содержания белка в зерне, крахмала в клубнях картофеля, сахара в корнеплодах, жиров в зернах масличных культур, аскорбиновой кислоты и сахаров в ягодных и плодовых растениях.
Симптомы недостатка меди. При недостатке меди в питании растений происходит нарушение развития корневой системы, в связи с чем замедляется рост всей культуры. Внешние проявления можно определить по увяданию верхних листьев (возможно даже отмирание верхушек побегов), изменению их цвета (неоднородность окраски, появление более светлых пятен, пожелтение либо приобретение коричневатого оттенка), а иногда и формы (листья скручиваются и вянут).
Симптомы избытка меди. Высокие концентрации меди действуют на растения токсично. Переизбыток этого элемента приводит к замедлению развития растения, появлению бурых пятен на нижних листьях и их отмиранию. Кроме того, он может провоцировать дефицит железа в растениях. Поэтому важно правильно дозировать медьсодержащие удобрения, учитывая естественное количество меди, находящееся в плодородном слое почвы данного типа.
Содержание меди в почвах. Для этого элемента характерно сосредоточение в верхних слоях почвы. Количество меди в грунте, необходимое для оптимального обеспечения растений этим минералом, невелико и колеблется в пределах 5 – 20 мг/кг. При образовании биомассы сельскохозяйственные культуры поглощают из почвы от 10 до 300 г/га меди, но поступление этого элемента из атмосферы (метеоритная, космическая пыль, почвенно-грунтовые воды, вулканические газы, загрязнение биосферы в результате геотехнической деятельности человека), а также его количество в материнской породе почти полностью компенсируют эту потерю, а иногда и превышают ее. Происходит поглощение, накопление и распределение микроэлемента в почвенных слоях.

Содержание меди в различных типах грунтов может существенно отличаться. Большое количество ее характерно для кислых почв. Чтобы исправить эту ситуацию, применяют известкование. Бедные медью малогумусные песчаные, осушенные болотные, дерново-подзолистые с легким гранулометрическим составом, торфяные почвы, где медь находится в труднодоступной для растений форме органических соединений, требуют обогащения путем внесения медьсодержащих соединений.
Наиболее богаты медью желтоземы и красноземы. Чуть меньше ее в засоленных почвах и черноземах. Дерново-подзолистые, сероземы и каштановые грунты содержат более низкие концентрации этого микроэлемента. А верховые торфяники и дерново-карбонатные – самые бедные почвы по количеству находящейся в них меди.
Формы соединений меди. Помимо количественного показателя, который зависит от типа почвы, очень важна и форма медьсодержащих соединений, определяющая степень доступности этого элемента для растений. Растения могут питаться только водорастворимыми или поглощенными (обменно-сорбированными) формами меди.
Поглощение меди происходит различным образом: она может входить в кристаллическую решетку различных минералов, адсорбироваться коллоидными частицами почвы, входить в состав органических веществ почвы, а также образовывать водорастворимые соединения.
Катионы меди легко вступают в химическое взаимодействие с органическими и минеральными веществами, поэтому осаждаются различными анионами (сульфидом, карбонатом, гидроксидом), образуя малоподвижные формы. Водорастворимые соединения меди обычно составляют незначительную часть (до 1%) от общего ее количества в почве. К тому же они легко вымываются из грунта. Особенно это актуально для супесчаных и песчаных почв с небольшой поглощаемостью. Кроме водорастворимых хорошей доступностью для культур обладают обменно-сорбированные формы меди, когда она поглощается органическими или минеральными коллоидами почвы, или глинистыми минералами.

Применение удобрений, содержащих медь. Медные удобрения наиболее эффективны на торфяниках и заболоченных почвах легкого гранулометрического состава. Иногда торфяные почвы настолько бедны медью, что не позволяют получить урожай сельскохозяйственных культур. При их обогащении медьсодержащими удобрениями урожайность повышается на 2 – 5 ц/га.
В засушливые годы доступность подвижных форм меди, содержащихся в почве, снижается, поэтому применение удобрений становится более эффективным. Средние расчетные дозы меди при внесении в почву составляют 1,5 – 2,0 кг/га в год. Практика зарубежных исследований рекомендует одноразовое внесение (до 10 кг) на несколько лет. Такая технология имеет долговременный эффект, поэтому даже спустя 10 лет концентрация медьсодержащих соединений в грунте остается достаточно высокой. При многократных внесениях создается риск накопления меди в количествах, токсичных для растений.
Эффективность медьсодержащих удобрений зависит от вида растений и типа почв. Они увеличивают урожайность и повышают качество продукции зерновых, льна и кормовых культур, растущих на осушенных болотных и других почвах. Правильное применение медных удобрений позволяет повысить урожаи: на 2–5 ц/га для пшеницы, на 2–3 ц/га для ячменя, на 4–6 ц/га для овса, на 21% для зеленой массы кукурузы, на 9–13% для ее початков. Урожайность плодов сахарной свеклы, растущей на дерново-подзолистых почвах, удобренных медьсодержащими соединениями, повышается на 43 – 45%. При этом та же культура, произрастающая на дерново-карбонатных грунтах, богатых соединениями подвижной меди, не дает прибавки к урожаю в случае подкормки этим минералом.

Картофель, выращиваемый на дерново-подзолистых грунтах, благодаря внесению меди при определенных условиях не только повышает урожайность и качество корнеплодов, но и повышает устойчивость к таким заболеваниям как фитофтороз и черная ножка. У моркови медьсодержащие удобрения способствуют большему образованию в корнеплодах сахаров, каротина и азота, у томатов – повышению урожайности и витамина С в плодах. Кормовые травы более интенсивно наращивают зеленую массу, питательные качества которой также улучшаются.
Промышленное производство медьсодержащих удобрений базируется на переработке различных медных руд (окисленных, сульфидных или самородных), количество меди в которых составляет 0,7 – 3%. Еще один способ получения меди – переработка вторичных цветных металлов.
Среди медьсодержащих удобрений различают: сернокислую медь (медный купорос) и суперфосфат с медью, которые используют для внекорневой подкормки и предпосевной обработки семян; пиритные огарки (промышленные отходы медеплавильного производства), окисленные медные руды с низким процентом содержания меди и шлаки медеплавильного производства, которые применяют для внесения в почву во время зяблевой обработки; медьсодержащий порошок для опудривания семян.
На осваиваемых торфяниках целесообразно вносить 4 – 5 ц/га пиритных огарков (на 4 – 6 лет). Доза сульфата меди для почвенного внесения – 25 кг/га. Для равномерного распределения микроэлемента его предварительно тщательно смешивают с минеральным удобрением.
В настоящее время одним из наиболее перспективных и эффективных источников поступления меди для растений являются различные хелатные формы удобрений, содержащие этот микроэлемент. Так, например, микроудобрение для внекорневой подкормки "Оракул колофермин меди" украинского производства позволяет эффективно ликвидировать дефицит этого микроэлемента в растениях (при норме расхода 1 - 2 л/га). Благодаря отсутствию балластных добавок, этот хелатный комплекс не вызывает ожогов листьев и полностью усваивается листовой пластиной.

Взаимодействие меди с другими элементами. Содержание меди в почвенных растворах, преимущественно органических хелатах (до 80%), составляет 3 – 135 мкг/л. В почве медь сначала поглощается органическим веществом, а затем сорбируется минералами (эта форма легче подвергается десорбции). Недостаток меди часто проявляется на богатых гумусом почвах. При повышении рН снижается ее доступность. Также значительное конкурирующее влияние на поступление меди оказывают железо и цинк. Медь способна обострять дефицит молибдена в растениях. И наоборот: внесение молибдена может ухудшить доступность меди. Токсичные количества алюминия также способны затруднять поглощение меди растениями.
Из основных элементов питания наиболее сильно медь связана с азотом. Высокое содержание азотных удобрений вызывает интенсивный рост растений, что, в свою очередь, обуславливает их повышенную потребность в меди. Фосфатные соединения обладают высоким показателем адсорбции меди, но в результатате она снижает доступность фосфора для растений. Кальций в составе карбонатов приводит к осаждению меди и возникновению ее дефицита.
Для обработки семян и внекорневых подкормок широко используют сульфат меди (медный купорос), который содержит 23,4 – 24,9% меди. Этот кристаллический порошок хорошо растворяется в воде. Для предпосевной обработки семян его применяют в концентрации 0,1%. Целесообразно проводить такую обработку в комплексе с пестицидами (для протравливания посевного материала). Внекорневые подкормки требуют значительно меньшей концентрации раствора сульфата меди: 0,02 – 0,05%.
Наиболее отзывчивы на медные удобрения зерновые (пшеница, овес, ячмень), корнеплоды (свекла столовая, морковь), подсолнечник, овощи и плодово-ягодные культуры. Среди овощных особенно нуждаются в меди салат, шпинат, лук, огурец, фасоль, горох. Хороший эффект от применения медьсодержащих удобрений достигается у свеклы столовой и моркови. Более худший результат показывают капуста белокочаная, сельдерей, томат.
agrostory.com