
«Кровь» растений: ксилема, флоэма и сок в физиологии растений. Ксилема развита у растений
ксилема, флоэма и сок в физиологии растений — Сириус агро плант
Флоэма и ксилема являются сосудистыми тканями трахеофитных растений, то есть в них расположены сосуды растений, которые осуществляют транспортировку сока в растительных организмах: папоротникообразных, голосеменных и покрытосеменных, сообщая корневую систему с лиственными структурами посредством стебля. Обе являются переносчиками сока.

Ксилема транспортирует необработанный «сырой» сок (сырье или неорганический сок, поскольку он состоит из питательных веществ, поглощенных из почвы растением, воды и минеральных солей) и расположена в самом внутреннем слое стебля. Она транспортирует сок от корней к листьям, где будет проходить фотосинтез. Ксилема образована мертвыми клетками, которые имеют функцию предотвращения деформации, вызванной давлением сока.
Флоэма транспортирует переработанный сок (или органический сок, продукт фотосинтеза, где питательные вещества превращаются в глюкозу) и находится в самом внешнем слое стебля. Она транспортирует переработанный сок из хлорофильных частей (мест, где происходит фотосинтез) к живыми частями растения — областям, где глюкоза будет превращаться в энергию. Флоэма образована живыми клетками, вытянутыми и без ядра.

Транспортировка необработанного сока, образованного водой и минеральными солями, осуществляемая ксилемой, происходит из способности поглощения корней и распределения в основной пункт назначения к листьям. Ее основной состав включает сосуды, трахеи, трахеиды, волокна и паренхимные клетки.
Транспортировка переработанного сока, образованного органическими веществами, полученными при фотосинтезе, осуществляется через флоэму, начиная от листьев в направлении остальных органов, главным образом, энергетического резерва (корней и стеблей). Она состоит из ситовидных элементов, сопровождающих клеток, волокон и паренхимных клеток.
Растения как автотрофные организмы
У растений автотрофное питание, другими словами, они могут производить органические молекулы (главным образом глюкозу) из простых неорганических веществ с помощью внешнего источника энергии, энергии света.
Итак, питательные вещества, которые требуются растению, очень просты: вода, минеральные соли и углекислый газ (мы не называем солнечную энергию, потому что это не вещество, а тип энергии). Растения в процессе фотосинтеза, происходящего в хлоропластах клеток, продуцируют глюкозу, которая является органической молекулой, служащей источником химической энергии (а также для создания других малых и больших структурных молекул, например, целлюлозы).
В связи с этим растение испытывает следующие трудности:
- Фотосинтез происходит главным образом в клетках листьев на определенной высоте над землей (иногда несколько метров), но воду и минеральные соли следует добывать из почвы, корнями. Поэтому растение должно транспортировать эти простые вещества вверх, преодолевая силу тяжести.
- Не все клетки растения выполняют фотосинтез (например, клетки корней нет). Поэтому, как только глюкоза вырабатывается в клетках листьев, часть ее должна транспортироваться в другие места для снабжения всех клеток.
Растение должно поглощать воду и минеральные соли через корни. Для этого клетки самого поверхностного слоя корня имеют очень тонкие расширения в виде нитей, которые выступают наружу, называемые абсорбирующими или корневыми волосками. Таким образом, контактная поверхность корня с внешней средой значительно увеличивается.
Клетки, которые составляют абсорбирующие волоски, имеют важную способность: они способны переносить минеральные соли из внешней среды во внутреннюю часть клетки, даже если концентрация солей внутри клетки больше, чем снаружи (затратив некоторую энергию, конечно). Таким образом, концентрация солей становится больше внутри клеток по отношению к внешней стороне.
Существуют две альтернативы для проникновения воды в корень: вода может проходить через клетки, переходя из одной в другую через небольшие отверстия, называемые плазмодесмами (внутриклеточный или упрощенный транспорт) или проходить через пространства клеточных стенок, которые отделяют одну клетку от другой (внеклеточный или апопластический транспорт).
Как только вода была поглощена, достигнута первая цель: вода и минеральные соли находятся внутри корневых клеток… но все еще далеки от их цели, которая является листьями.
Транспортировка необработанного сока через ксилему

Для доставки воды, минеральных солей и других веществ, производимых корневыми клетками, к листьям у растений есть гениальная система проводимости, образованная очень специализированными клетками, которые в совокупности называются ксилемой.
Ксилема — очень своеобразная ткань, потому что многие из ее клеток «жертвуют» собой, чтобы выполнить свою функцию! Хотя это звучит немного противоречиво, клетки ксилемы более эффективны мертвыми, чем живыми. По мере их роста они утолщают клеточные стенки, образуя спиральные или кольцевые структуры. Кроме того, они соединяются друг с другом для образования различных типов трубок. Наконец, они умирают, клетка исчезает, но ее клеточная стенка остается, что будет способствовать образованию удлиненных каналов, наподобие трубок, которые проходят по всему стеблю от корней до листьев.
Существует два основных типа проводящих трубок. Первый тип — трахеиды, представляющие собой очень узкие и удлиненные клетки, в которых прохождение веществ из одной клетки в другую осуществляется через окаймленные поры в клеточных стенках, расположенные на большой части этих стенок. Другим типом являются сосуды или трахеи с несколько большим диаметром и с законченными отверстиями (перфорациями) в зонах стенок, отделяющих от других клеток.
Вы, вероятно, задаетесь вопросом: «Если клетки трубок ксилемы мертвы, как им удается транспортировать сок?»
Хитрость заключается в использовании физических механизмов, которые не требуют вмешательства биологических процессов. Вода поднимается через трубки ксилемы благодаря нескольким физическим процессам, которые вместе образуют механизм, часто называемый натяжение-адгезия(прилипание)-когезия(сцепление). Основные идеи этого механизма были предложены более века тому назад Генри Диксоном. Механизм натяжение-адгезия-когезия основан на следующих явлениях или процессах.
Капиллярность
Капилярность — это любопытное явление, которое заставляет некоторые жидкости подниматься против гравитации спонтанно, внутри очень тонких протоков. Чем меньше диаметр канала, тем выше высота восходящего столба жидкости. Это движение, вызванное силой, возникшей в результате конфликта между двумя другими: поверхностное натяжение воды и адгезия воды к твердым поверхностям некоторых веществ:

- Когезия и поверхностное натяжение:
Молекулы воды, благодаря своей полярности, устанавливают между собой союзы, которые отвечают за сильное сцепление (объединение), существующее между ними.
Внутри массы воды каждая молекула одинаково привлекается в равной мере своими соседями, но только на поверхности жидкости, контактирующей с воздухом, все меняется: сплоченность молекул воды «вытягивает» молекулы воды внутрь жидкости, а также в направлении, параллельном поверхности жидкости.
Таким образом, сцепление заставляет слой молекул, контактирующих с воздухом, иметь натяжение, которое заставляет их вести себя как эластичная пленка.
Поверхность воды оказывает сопротивление деформации, поскольку ситуация максимального баланса будет такова, в которой поверхность контакта вода-воздух минимальна. Сила поверхностного натяжения отвечает за форму, которую принимают капли воды, а также за то, что легкие насекомые способны ходить по поверхности воды без погружения.
- Поверхностное натяжение и адгезия:
Между молекулами воды и твердыми поверхностями многих веществ также может быть установлена притягивающая сила, которая заставляет воду прилипать к этим поверхностям. Если адгезия к твердой поверхности сильнее когезии между молекулами воды, на краю жидкости, контактирующей с твердым материалом будет образовываться вогнутая кривизна поверхности.
Однако, поверхностное натяжение воды создает силу, которая тянет молекулы, образующие нижнюю часть кривой, чтобы попытаться уменьшить контактную поверхность воздух-вода, создавая плоскую пленку.
В результате создается возрастающая результирующая сила, которая приводит к небольшому подъему водного столба. Конечным результатом является то, что вода может буквально подняться по каналу, преодолевая силу тяжести.
Эвапотранспирация и натяжение
Капиллярность очень хорошо дает понять, как сырой сок может спонтанно подниматься через очень тонкие трубки ксилемы, но как только столб воды или сырой сок достигает конца трубки, капиллярности недостаточно, чтобы объяснить, как этот сок может продолжать подниматься из корня.
Механизм, который нам не ясен, связан с испарением воды, которая вырабатывается в листьях, и расходами этой воды при фотосинтезе.
Когда молекулы воды удаляются с конца трубки ксилемы из-за транспирации листьев, отверстие, как правило, немедленно заполняется другими молекулами, которые ниже. Это создает натяжение или отрицательное давление, которое буквально тянет столб воды вверх.
Сильное сцепление между молекулами воды предотвращает разрыв столбика сока внутри трубки, так что весь сок внутри трубки поднимается как единое целое. Естественно, из корней должна быть постоянная подача воды. В противном случае, как и в ситуациях засухи, столбик сока может быть прерван, что вызовет проблемы для растения.
Следующий рисунок объясняет явление натяжения, вызванного эвапотранспирацией:

Транспортировка переработанного сока через флоэму
К настоящему моменту мы не решили и более половины проблемы, потому что, как только фотосинтетические клетки образуют углеводы, необходимо распределить эти молекулы и их производные во многие другие отдаленные клетки, которые не способны их произвести.
Для этого распределения растения имеют другую ткань, называемую флоэмой, которая также образует каналы, проходящие через растение, но с некоторыми важными отличиями по сравнению с ксилемой.

Во-первых, проводящие клетки флоэмы живые, хотя они теряют свое ядро и довольно много органелл. В целом, они имеют меньший диаметр, чем проводники ксилемы.
Как и в ксилеме, клетки флоэмы выстраиваются рядами и соединяют свои стенки. В зонах соединения образуются структуры, называемые ситовидными пластинками. В этих областях имеются небольшие отверстия диаметром до 15 мкм (не такие большие, как перфорации ксилемы), которые позволяют передавать переработанный сок из одной ячейки в другую. Каналы, образованные объединением множества этих клеток, называются ситовидными трубками.
В течение некоторого времени считалось, что переработанные вещества могут транспортироваться путем диффузии через трубки флоэмы, но когда были сделаны расчеты, чтобы узнать скорость транспортировки через флоэму, стало ясно, что необходим другой механизм.
В настоящее время считается, что механизмом движения переработанного сока через флоэму является механизм, который предложил немецкий ученый Эрнст Мюнх в 1926 году. Этот механизм называется потоком под давлением и состоит в следующем.
Как только фотосинтетические клетки образуют органические молекулы, благодаря фотосинтезу, особенно глюкозу и фруктозу, большая часть этих молекул связывается с образованием сахарозы, которая является предпочтительной органической молекулой для переноса углеводов через растение.
Сахароза транспортируется во флоэму, из-за этого переработанный сок содержит высокую концентрацию этого вещества (сахароза составляет 90% веществ, присутствующих в переработанном соке, конечно, без учета воды). Поскольку концентрация сахарозы внутри флоэмы намного выше, чем снаружи, создается осмотическое давление, которое заставляет воду поступать во флоэму, тем самым увеличивая ее давление. Это давление подталкивает переработанный сок вдоль протоков флоэмы.
Как только они достигают места назначения, которое является конечным пунктом протоков флоэмы, большое количество сахарозы извлекается из флоэмы, «вытаскивая» с ней определенное количество воды. Это способствует циркуляции переработанного сока, вызывая падение давления в конечном пункте. Сахароза, поступив к месту назначения, может использоваться для разных целей.
Резюме
Подводя итоги, можно отметить, что корни поглощают минеральные соли посредством активного транспорта, что генерирует осмотическое давление, вызывающее поглощение воды.
Эти вещества достигают проводников ксилемы, которые отвечают за транспортировку сырого сока от корней к фотосинтетическим структурам за счет механизма когезия-адгезия-натяжение.
После того как вырабатываются органические вещества, проводники флоэмы отвечают за транспортировку переработанного сока, богатого органическими молекулами, из листьев в другие области растения посредством механизма потока под давлением, основанного на механизмах осмотического давления.
Инга Костенко, Mivena Украина
Анна Устименко, Клуб Sirius Agro Plant
siriusap.com
Ксилема - «Энциклопедия»
КСИЛЕМА (от греческого ξύλον – дерево, древесина), проводящая ткань плауновидных, папоротниковидных, хвощевидных и семенных растений, обеспечивающая передвижение поглощённой корнями воды с растворёнными в ней элементами минерального питания и фитогормонами, синтезируемыми верхушками корней. Вместе с другой проводящей тканью – флоэмой образует непрерывную разветвлённую систему, пронизывающую всё тело растения – от тончайших корешков до самых молодых побегов. По происхождению различают первичную и вторичную ксилему. Первичная ксилема дифференцируется из прокамбия; первые проводящие её элементы, развивающиеся во время роста органа, называются протоксилемой, проводящие элементы, формирующиеся позднее, – метаксилемой. Вторичная ксилема формируется камбием; у деревьев и кустарников её обычно называют древесиной.
 Ксилема – сложная ткань, состоящая из клеток разных типов. Самые важные из них – проводящие, или трахеальные, элементы (ТЭ). Они возникают из живых клеток, которые имеют тонкие растяжимые первичные клеточные стенки. После достижения ТЭ окончательных размеров происходит формирование вторичной стенки и автолиз протопласта клетки; клеточная стенка остаётся, а полость заполняется жидкостью. У подавляющего большинства цветковых растений ТЭ представлены сосудами (могут достигать нескольких метров в длину), которые состоят из одного ряда клеток-члеников, сообщающихся между собой одним или несколькими сквозными отверстиями (перфорациями) в смежных стенках, образующимися при автолизе протопласта; такие смежные стенки члеников называют перфорационными пластинками. У остальных растений, обладающих ксилемами, ТЭ представлены трахеидами – гомологами члеников сосудов, не превышающими в длину 1,5-2 мм. Трахеиды появились у растений в связи с выходом их на сушу, сосуды – значительно позднее, путём преобразования трахеид.
Ксилема – сложная ткань, состоящая из клеток разных типов. Самые важные из них – проводящие, или трахеальные, элементы (ТЭ). Они возникают из живых клеток, которые имеют тонкие растяжимые первичные клеточные стенки. После достижения ТЭ окончательных размеров происходит формирование вторичной стенки и автолиз протопласта клетки; клеточная стенка остаётся, а полость заполняется жидкостью. У подавляющего большинства цветковых растений ТЭ представлены сосудами (могут достигать нескольких метров в длину), которые состоят из одного ряда клеток-члеников, сообщающихся между собой одним или несколькими сквозными отверстиями (перфорациями) в смежных стенках, образующимися при автолизе протопласта; такие смежные стенки члеников называют перфорационными пластинками. У остальных растений, обладающих ксилемами, ТЭ представлены трахеидами – гомологами члеников сосудов, не превышающими в длину 1,5-2 мм. Трахеиды появились у растений в связи с выходом их на сушу, сосуды – значительно позднее, путём преобразования трахеид.
Реклама
В зависимости от строения боковых стенок ТЭ называются кольчатыми, спиральными, пористыми. В кольчатых ТЭ вторичная клеточная стенка в виде нескольких отдельных колец армирует тонкую первичную стенку, в спиральных – имеет вид одной (иногда двух) узкой спиральной ленты, в пористых – пронизана мелкими отверстиями-порами. Вытянутые поперёк трахеальных элементов поры называют лестничными, а округлые – точечными. Кольчатые и спиральные ТЭ, развивающиеся в протоксилеме, способны эффективно проводить воду вдоль тяжа ксилемы, а также загружать воду в ксилему или выгружать её; пористые ТЭ, свойственные метаксилеме и вторичной ксилеме, проводят воду вдоль тяжа ксилемы.
Движение воды по ксилеме обусловлено корневым давлением и сосущей силой тканей побега. Тонкую регуляцию транспорта по ксилеме осуществляют живые клетки ксилемной паренхимы, располагающиеся продольными (тяжевая паренхима) или радиальными (лучевая паренхима) тяжами. Кроме того, они могут запасать крахмал или содержать кристаллический оксалат кальция. Вторичная ксилема многих древесных растений, включающая плохо проводящие воду толстостенные волокнистые трахеиды и/или не проводящие воду волокна либриформа, выполняет опорную роль. Обычно вторичная ксилема древесных растений содержит кольца годовых приростов (смотри Годичные кольца). Как правило, воду проводят только 2-3 наружных, самых молодых кольца ксилемы. Более старая ксилема внутренних колец осушается, пропитывается разными полиароматическими соединениями и становится спелой или ядровой древесиной, выполняющей только механическую функцию. Паренхимные клетки в ней отмирают.
Лит.: Лотова Л. И. Морфология и анатомия высших растений. М., 2001.
А. К. Тимонин.
knowledge.su
ксилема, флоэма и сок в физиологии растений — Сириус агро плант
Флоэма и ксилема являются сосудистыми тканями трахеофитных растений, то есть в них расположены сосуды растений, которые осуществляют транспортировку сока в растительных организмах: папоротникообразных, голосеменных и покрытосеменных, сообщая корневую систему с лиственными структурами посредством стебля. Обе являются переносчиками сока.
Ксилема транспортирует необработанный «сырой» сок (сырье или неорганический сок, поскольку он состоит из питательных веществ, поглощенных из почвы растением, воды и минеральных солей) и расположена в самом внутреннем слое стебля. Она транспортирует сок от корней к листьям, где будет проходить фотосинтез. Ксилема образована мертвыми клетками, которые имеют функцию предотвращения деформации, вызванной давлением сока.
Флоэма транспортирует переработанный сок (или органический сок, продукт фотосинтеза, где питательные вещества превращаются в глюкозу) и находится в самом внешнем слое стебля. Она транспортирует переработанный сок из хлорофильных частей (мест, где происходит фотосинтез) к живыми частями растения — областям, где глюкоза будет превращаться в энергию. Флоэма образована живыми клетками, вытянутыми и без ядра.
Транспортировка необработанного сока, образованного водой и минеральными солями, осуществляемая ксилемой, происходит из способности поглощения корней и распределения в основной пункт назначения к листьям. Ее основной состав включает сосуды, трахеи, трахеиды, волокна и паренхимные клетки.
Транспортировка переработанного сока, образованного органическими веществами, полученными при фотосинтезе, осуществляется через флоэму, начиная от листьев в направлении остальных органов, главным образом, энергетического резерва (корней и стеблей). Она состоит из ситовидных элементов, сопровождающих клеток, волокон и паренхимных клеток.
Растения как автотрофные организмы
У растений автотрофное питание, другими словами, они могут производить органические молекулы (главным образом глюкозу) из простых неорганических веществ с помощью внешнего источника энергии, энергии света.
Итак, питательные вещества, которые требуются растению, очень просты: вода, минеральные соли и углекислый газ (мы не называем солнечную энергию, потому что это не вещество, а тип энергии). Растения в процессе фотосинтеза, происходящего в хлоропластах клеток, продуцируют глюкозу, которая является органической молекулой, служащей источником химической энергии (а также для создания других малых и больших структурных молекул, например, целлюлозы).
В связи с этим растение испытывает следующие трудности:
- Фотосинтез происходит главным образом в клетках листьев на определенной высоте над землей (иногда несколько метров), но воду и минеральные соли следует добывать из почвы, корнями. Поэтому растение должно транспортировать эти простые вещества вверх, преодолевая силу тяжести.
- Не все клетки растения выполняют фотосинтез (например, клетки корней нет). Поэтому, как только глюкоза вырабатывается в клетках листьев, часть ее должна транспортироваться в другие места для снабжения всех клеток.
Растение должно поглощать воду и минеральные соли через корни. Для этого клетки самого поверхностного слоя корня имеют очень тонкие расширения в виде нитей, которые выступают наружу, называемые абсорбирующими или корневыми волосками. Таким образом, контактная поверхность корня с внешней средой значительно увеличивается.
Клетки, которые составляют абсорбирующие волоски, имеют важную способность: они способны переносить минеральные соли из внешней среды во внутреннюю часть клетки, даже если концентрация солей внутри клетки больше, чем снаружи (затратив некоторую энергию, конечно). Таким образом, концентрация солей становится больше внутри клеток по отношению к внешней стороне.
Существуют две альтернативы для проникновения воды в корень: вода может проходить через клетки, переходя из одной в другую через небольшие отверстия, называемые плазмодесмами (внутриклеточный или упрощенный транспорт) или проходить через пространства клеточных стенок, которые отделяют одну клетку от другой (внеклеточный или апопластический транспорт).
Как только вода была поглощена, достигнута первая цель: вода и минеральные соли находятся внутри корневых клеток… но все еще далеки от их цели, которая является листьями.
Транспортировка необработанного сока через ксилему
Для доставки воды, минеральных солей и других веществ, производимых корневыми клетками, к листьям у растений есть гениальная система проводимости, образованная очень специализированными клетками, которые в совокупности называются ксилемой.
Ксилема — очень своеобразная ткань, потому что многие из ее клеток «жертвуют» собой, чтобы выполнить свою функцию! Хотя это звучит немного противоречиво, клетки ксилемы более эффективны мертвыми, чем живыми. По мере их роста они утолщают клеточные стенки, образуя спиральные или кольцевые структуры. Кроме того, они соединяются друг с другом для образования различных типов трубок. Наконец, они умирают, клетка исчезает, но ее клеточная стенка остается, что будет способствовать образованию удлиненных каналов, наподобие трубок, которые проходят по всему стеблю от корней до листьев.
Существует два основных типа проводящих трубок. Первый тип — трахеиды, представляющие собой очень узкие и удлиненные клетки, в которых прохождение веществ из одной клетки в другую осуществляется через окаймленные поры в клеточных стенках, расположенные на большой части этих стенок. Другим типом являются сосуды или трахеи с несколько большим диаметром и с законченными отверстиями (перфорациями) в зонах стенок, отделяющих от других клеток.
Вы, вероятно, задаетесь вопросом: «Если клетки трубок ксилемы мертвы, как им удается транспортировать сок?»
Хитрость заключается в использовании физических механизмов, которые не требуют вмешательства биологических процессов. Вода поднимается через трубки ксилемы благодаря нескольким физическим процессам, которые вместе образуют механизм, часто называемый натяжение-адгезия(прилипание)-когезия(сцепление). Основные идеи этого механизма были предложены более века тому назад Генри Диксоном. Механизм натяжение-адгезия-когезия основан на следующих явлениях или процессах.
Капиллярность
Капилярность — это любопытное явление, которое заставляет некоторые жидкости подниматься против гравитации спонтанно, внутри очень тонких протоков. Чем меньше диаметр канала, тем выше высота восходящего столба жидкости. Это движение, вызванное силой, возникшей в результате конфликта между двумя другими: поверхностное натяжение воды и адгезия воды к твердым поверхностям некоторых веществ:
- Когезия и поверхностное натяжение:
Молекулы воды, благодаря своей полярности, устанавливают между собой союзы, которые отвечают за сильное сцепление (объединение), существующее между ними.
Внутри массы воды каждая молекула одинаково привлекается в равной мере своими соседями, но только на поверхности жидкости, контактирующей с воздухом, все меняется: сплоченность молекул воды «вытягивает» молекулы воды внутрь жидкости, а также в направлении, параллельном поверхности жидкости.
Таким образом, сцепление заставляет слой молекул, контактирующих с воздухом, иметь натяжение, которое заставляет их вести себя как эластичная пленка.
Поверхность воды оказывает сопротивление деформации, поскольку ситуация максимального баланса будет такова, в которой поверхность контакта вода-воздух минимальна. Сила поверхностного натяжения отвечает за форму, которую принимают капли воды, а также за то, что легкие насекомые способны ходить по поверхности воды без погружения.
- Поверхностное натяжение и адгезия:
Между молекулами воды и твердыми поверхностями многих веществ также может быть установлена притягивающая сила, которая заставляет воду прилипать к этим поверхностям. Если адгезия к твердой поверхности сильнее когезии между молекулами воды, на краю жидкости, контактирующей с твердым материалом будет образовываться вогнутая кривизна поверхности.
Однако, поверхностное натяжение воды создает силу, которая тянет молекулы, образующие нижнюю часть кривой, чтобы попытаться уменьшить контактную поверхность воздух-вода, создавая плоскую пленку.
В результате создается возрастающая результирующая сила, которая приводит к небольшому подъему водного столба. Конечным результатом является то, что вода может буквально подняться по каналу, преодолевая силу тяжести.
Эвапотранспирация и натяжение
Капиллярность очень хорошо дает понять, как сырой сок может спонтанно подниматься через очень тонкие трубки ксилемы, но как только столб воды или сырой сок достигает конца трубки, капиллярности недостаточно, чтобы объяснить, как этот сок может продолжать подниматься из корня.
Механизм, который нам не ясен, связан с испарением воды, которая вырабатывается в листьях, и расходами этой воды при фотосинтезе.
Когда молекулы воды удаляются с конца трубки ксилемы из-за транспирации листьев, отверстие, как правило, немедленно заполняется другими молекулами, которые ниже. Это создает натяжение или отрицательное давление, которое буквально тянет столб воды вверх.
Сильное сцепление между молекулами воды предотвращает разрыв столбика сока внутри трубки, так что весь сок внутри трубки поднимается как единое целое. Естественно, из корней должна быть постоянная подача воды. В противном случае, как и в ситуациях засухи, столбик сока может быть прерван, что вызовет проблемы для растения.
Следующий рисунок объясняет явление натяжения, вызванного эвапотранспирацией:
Транспортировка переработанного сока через флоэму
К настоящему моменту мы не решили и более половины проблемы, потому что, как только фотосинтетические клетки образуют углеводы, необходимо распределить эти молекулы и их производные во многие другие отдаленные клетки, которые не способны их произвести.
Для этого распределения растения имеют другую ткань, называемую флоэмой, которая также образует каналы, проходящие через растение, но с некоторыми важными отличиями по сравнению с ксилемой.
Во-первых, проводящие клетки флоэмы живые, хотя они теряют свое ядро и довольно много органелл. В целом, они имеют меньший диаметр, чем проводники ксилемы.
Как и в ксилеме, клетки флоэмы выстраиваются рядами и соединяют свои стенки. В зонах соединения образуются структуры, называемые ситовидными пластинками. В этих областях имеются небольшие отверстия диаметром до 15 мкм (не такие большие, как перфорации ксилемы), которые позволяют передавать переработанный сок из одной ячейки в другую. Каналы, образованные объединением множества этих клеток, называются ситовидными трубками.
В течение некоторого времени считалось, что переработанные вещества могут транспортироваться путем диффузии через трубки флоэмы, но когда были сделаны расчеты, чтобы узнать скорость транспортировки через флоэму, стало ясно, что необходим другой механизм.
В настоящее время считается, что механизмом движения переработанного сока через флоэму является механизм, который предложил немецкий ученый Эрнст Мюнх в 1926 году. Этот механизм называется потоком под давлением и состоит в следующем.
Как только фотосинтетические клетки образуют органические молекулы, благодаря фотосинтезу, особенно глюкозу и фруктозу, большая часть этих молекул связывается с образованием сахарозы, которая является предпочтительной органической молекулой для переноса углеводов через растение.
Сахароза транспортируется во флоэму, из-за этого переработанный сок содержит высокую концентрацию этого вещества (сахароза составляет 90% веществ, присутствующих в переработанном соке, конечно, без учета воды). Поскольку концентрация сахарозы внутри флоэмы намного выше, чем снаружи, создается осмотическое давление, которое заставляет воду поступать во флоэму, тем самым увеличивая ее давление. Это давление подталкивает переработанный сок вдоль протоков флоэмы.
Как только они достигают места назначения, которое является конечным пунктом протоков флоэмы, большое количество сахарозы извлекается из флоэмы, «вытаскивая» с ней определенное количество воды. Это способствует циркуляции переработанного сока, вызывая падение давления в конечном пункте. Сахароза, поступив к месту назначения, может использоваться для разных целей.
Резюме
Подводя итоги, можно отметить, что корни поглощают минеральные соли посредством активного транспорта, что генерирует осмотическое давление, вызывающее поглощение воды.
Эти вещества достигают проводников ксилемы, которые отвечают за транспортировку сырого сока от корней к фотосинтетическим структурам за счет механизма когезия-адгезия-натяжение.
После того как вырабатываются органические вещества, проводники флоэмы отвечают за транспортировку переработанного сока, богатого органическими молекулами, из листьев в другие области растения посредством механизма потока под давлением, основанного на механизмах осмотического давления.
Инга Костенко, Mivena Украина
Анна Устименко, Клуб Sirius Agro Plant
siriusap.com
Ксилема (древесина) растений
Ксилема (древесина) растений
По ксилеме от корня к листьям передвигаются вода и растворенные в ней минеральные вещества. Первичная и вторичная ксилемы содержат клетки одних и тех же типов. Однако первичная ксилема не имеет сердцевинных лучей, отличаясь этим от вторичной.
В состав ксилемы входят морфологически различные элементы, осуществляющие функции как проведения, так и хранения запасных веществ, а также чисто опорные функции. Дальний транспорт осуществляется по трахеальным элементам ксилемы: трахеидам и сосудам , ближний - по паренхимным элементам. Опорные, а иногда и запасающие функции выполняют часть трахеид и волокна механической ткани либриформа , также входящие в состав ксилемы.
Трахеиды в зрелом состоянии - это мертвые прозенхимные клетки, суженные на концах и лишенные протопласта . Длина трахеид в среднем составляет 1-4 мм, поперечник же не превышает десятых и даже сотых долей миллиметра. Стенки трахеид одревесневают, утолщаются и несут простые или окаймленные поры , через которые происходит фильтрация растворов. Большая часть окаймленных пор находится около окончаний клеток, т.е. там, где растворы просачиваются из одной трахеиды в другую. Трахеиды есть у спорофитов всех высших растений , а у большинства хвощевидных , плауновидных , папоротниковидных и голосеменных они являются единственными проводящими элементами ксилемы.
Сосуды - это полые трубки, состоящие из отдельных члеников, располагающихся друг над другом. Между расположенными один над другим члениками одного и того же сосуда имеются разного типа сквозные отверстия - перфорации. Благодаря перфорациям вдоль всего сосуда свободно осуществляется ток жидкости. Эволюционно сосуды, по-видимому, произошли из трахеид путем разрушения замыкающих пленок пор и последующего их слияния в одну или несколько перфораций. Концы трахеид, первоначально сильно скошенные, заняли горизонтальное положение, а сами трахеиды стали короче и превратились в членики сосудов ( рис. 45 ).
Сосуды появились независимо в разных линиях эволюции наземных растений. Однако наибольшего развития они достигают у покрытосеменных , где являются главнейшими водопроводящими элементами ксилемы.
Возникновение сосудов - важное свидетельство эволюционного прогресса этого таксона, поскольку они существенно облегчают транспирационный ток вдоль тела растения.
Помимо первичной оболочки, сосуды и трахеиды в большинстве случаев имеют вторичные утолщения. В самых молодых трахеальных элементах вторичная оболочка может иметь форму колец, не связанных друг с другом (кольчатые трахеиды и сосуды). Позднее появляются трахеальные элементы со спиральными утолщениями. Затем следуют сосуды и трахеиды с утолщениями, которые могут быть охарактеризованы как спирали, витки которых связаны между собой (лестничные утолщения). В конечном итоге вторичная оболочка сливается в более или менее сплошной цилиндр, формирующийся внутрь от первичной оболочки. Этот цилиндр прерывается в отдельных участках порами .
Сосуды и трахеиды с относительно небольшими округлыми участками первичной клеточной оболочки, не прикрытыми изнутри вторичной оболочкой, нередко называют пористыми. В тех случаях, когда поры во вторичной оболочке образуют подобие сетки или лестницы, говорят о сетчатых или лестничных трахеальных элементах (лестничные сосуды и трахеиды).
Вторичная, а иногда и первичная оболочка, как правило, лигнифицируются, т.е. пропитываются лигнином , это придает дополнительную прочность, но ограничивает возможности дальнейшего их роста в длину.
Трахеальные элементы, т.е. трахеиды и сосуды, распределяются в ксилеме различным образом. Иногда на поперечном срезе они образуют хорошо выраженные кольца (кольцесосудистая древесина). В других случаях сосуды рассеяны более или менее равномерно по всей массе ксилемы (рассеяннососудистая древесина). Особенности распределения трахеальных элементов в ксилеме используют при определении древесин различных пород деревьев.
Помимо трахеальных элементов, ксилема включает лучевые элементы, т.е. клетки, образующие сердцевинные лучи ( рис. 46 ), сформированные чаще всего тонкостенными паренхимными клетками (лучевая паренхима). Реже в лучах хвойных встречаются лучевые трахеиды. По сердцевинным лучам осуществляется ближний транспорт веществ в горизонтальном направлении. В ксилеме покрытосеменных помимо проводящих элементов содержатся также тонкостенные неодревесневшие живые паренхимные клетки, называемые древесинной паренхимой. По ним наряду с сердцевинными лучами отчасти осуществляется ближний транспорт. Кроме того, древесинная паренхима служит местом хранения запасных веществ. Элементы сердцевинных лучей и древесинной паренхимы, подобно трахеальным элементам, возникают из камбия .
Ссылки:
Все ссылкиmedbiol.ru
18.Ксилема. Флоэма.
Проводящие ткани обеспечивают восходящий и нисходящий ток растения. Восходящий ток - это ток минеральных солей, растворенных в воде, идущих от корней по стеблю к листьям. Восходящий ток осуществляется по сосудам и трахеидам ксилемы (древесины). Нисходящий ток - это ток органических веществ, направляющийся от листьев к корням по ситовидным элементам флоэмы(луба).
Ксилема и флоэма - это сложные ткани, состоящие из 3 основных элементов (табл. 2.2).
Проводящие элементы ксилемы. Наиболее древними проводящими элементами ксилемы являются трахеиды - это вытянутые клетки с заостренными концами. Трахеиды имеют одревесневшую клеточную стенку. По характеру утолщения оболочек, размерам и расположению в них участков первичных оболочек различают 4 типа трахеид: кольчатые, спиральные, пористые и лестничные (рис. 2.14). К наиболее древним пористым трахеидам относят лестничные трахеиды.
Основные элементы ксилемы и флоэмы

Сосуды (или трахеи) представляют собой однорядный продольный тяж клеток, называемых члениками. В филогенезе членики трахеи произошли из трахеид. Благодаря перфорациям между члениками вдоль всего сосуда свободно осуществляется ток жидкости. Утолщения клеточных оболочек у сосудов бывают кольчатыми, спиральными, лестничными, сетчатыми и пористыми (точечными).
Проводящие элементы флоэмы у архегониальных растений, кроме мхов, представлены ситовидными клетками. На их продольных стенках имеются сквозные отверстия, напоминающие сито, а потому называемые ситовидными полями. У покрытосеменных растений в процессе эволюции сформировался 2-й тип проводящих элементов - ситовидные трубки, представляющие собой продольный тяж клеток, называемых члениками. Конечные стенки члеников превращены в ситовидные пластинки: простые (имеют по одному ситовидному полю) и сложные (имеют несколько ситовидных полей). Ситовидные поля могут встречаться и на боковых стенках члеников (у березы). В ситовидном элементе разрушается тонопласт («оболочка» вакуоли), и вакуолярный сок смешивается с гиалоплазмой. Ядро, как правило, при этом разрушается. Каждому членику ситовидной трубки сопутствуют (одна или несколько) специализированных паренхимных клеток - (клетки-спутницы), выполняющие вспомогательную роль в транспорте органических веществ (рис. 2.15). Передвижение по ситовидным трубкам - физиологический процесс, идущий под действием ферментов выделяемых клетками-спутницами.
19.Проводящие пучки, их строение, классификация.
Сосудисто-волокнистые пучки. Флоэма и ксилема образуют сосудисто-волокнистые пучки, которые располагаются в центральном осевом цилиндре и бывают открытыми и закрытыми.
Закрытые пучки состоят из ксилемы и флоэмы, между которыми отсутствует камбий и, таким образом, не происходит образования новых элементов флоэмы и ксилемы. Закрытые сосудисто-волокнистые пучки встречаются в стеблях и корневищах однодольных растений.
Открытые пучки имеют камбий между флоэмой и ксилемой. В результате деятельности камбия пучок разрастается и происходит утолщение органа. Открытые сосудисто-волокнистые пучки встречаются во всех осевых органах двудольных и голосеменных растений.
В зависимости от взаимного расположения флоэмы и ксилемы различают 4 типа сосудисто-волокнистых пучков
Чаще всего флоэма лежит по одну сторону от ксилемы, т.е. она примыкает к ксилеме и обращена к периферии органа. Такие пучки называют коллатеральными. Они могут быть как закрытыми, так и от- крытыми. Закрытые коллатеральные пучки встречаются у стеблей и корневищ однодольных растений, а открытые - у стеблей, корневищ и корней двудольных растений.
В биколлатеральных пучках флоэма примыкает к ксилеме с 2 сторон. Мощный участок флоэмы обращен к периферии органа и отделен от ксилемы слоем камбия. Внутренняя флоэма развита слабо и примыкает к ксилеме без камбия. Биколлатеральные пучки встречаются у стеблей, корней и корневищ двудольных растений.
Радиальные пучки составлены ксилемой, которая располагается по радиусам. Между лучами ксилемы находятся участки флоэмы. Эти пучки не имеют камбия и встречаются только у корней в первичном строении.
Концентрические пучки складываются из ксилемы, окружающей флоэму, - центрофлоэмные пучки(у большинства корневищ однодольных), а центроксилемные - когда флоэма окружает ксилему (у корневищ папоротников).
studfiles.net
Ксилема и флоэма | Учеба-Легко.РФ
Ксилема и флоэма
Ксилема и флоэма – это проводящие ткани, состоящие из нескольких типов клеток. Они имеются только у папоротникообразных и семенных растений. В проводящей ткани имеются как мёртвые, так и живые клетки.
|
|
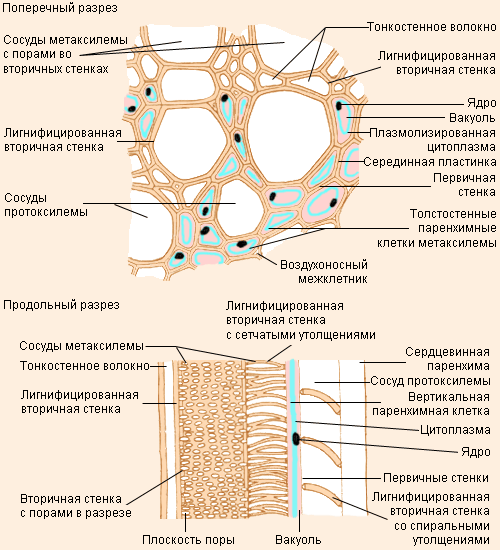
Строение ксилемы |
Ксилема выполняет в растении как опорную, так и проводящую функцию – по ней движутся вверх по растению вода и минеральные соли. В состав ксилемы входят элементы четырех типов: трахеиды, сосуды, паренхимные клетки и волокна. Трахеиды – мёртвые одиночные клетки веретеновидной формы. Их концы перекрываются, придавая растению необходимую прочность. Вода движется по пустым просветам трахеид, не встречая на своём пути помех в виде клеточного содержимого; от одной трахеиды к другой она передается через поры.

|
|
Рост сосудов протоксилемы |
|
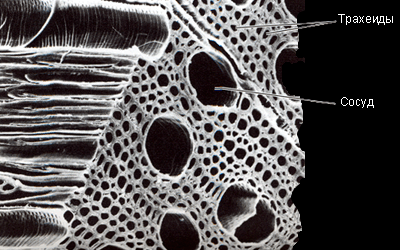
|
Сосуды и трахеиды в древесине клёна (увеличение в 350 раз) |
У покрытосеменных трахеиды развились в сосуды. Это очень длинные трубки, образовавшиеся в результате «состыковки» ряда клеток; остатки торцевых перегородок всё ещё сохраняются в сосудах в виде ободков. Размеры сосудов варьируют от нескольких сантиметров до нескольких метров. В первых по времени образования сосудах протоксилемы лигнин накапливается кольцами или по спирали. Это даёт возможность сосуду продолжать растягиваться во время роста. В сосудах метаксилемы лигнин сосредоточен более плотно – это идеальный «водопровод», действующий на большие расстояния.
Паренхимные клетки ксилемы образуют своеобразные лучи, соединяющие сердцевину с корой. Они проводят воду в радиальном направлении, запасают питательные вещества. Из других клеток паренхимы развиваются новые сосуды ксилемы. Наконец, древесинные волокна похожи на трахеиды. Они не проводят воду, но придают дополнительную прочность.
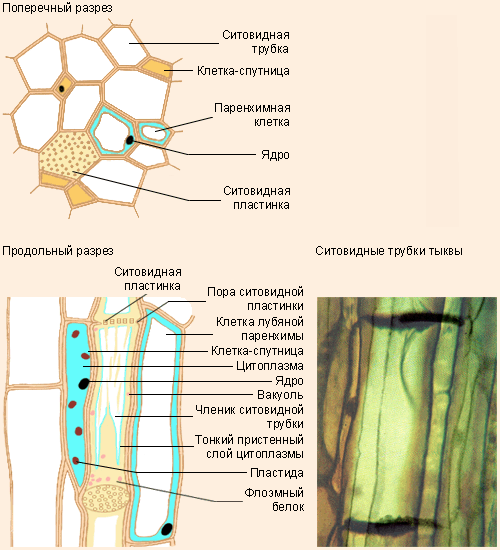
|
|
Строение флоэмы. Слева вверху – поперечный разрез, слева внизу – продольный, справа внизу – ситовидные трубки тыквы |
Во флоэме, как и в ксилеме, имеются трубчатые структуры, образованные, однако, живыми клетками. Основой этих структур являются ситовидные трубки, образующиеся в результате соединения ряда клеток. Ядра таких клеток после созревания отмирают, а цитоплазма прижимается к стенкам, освобождая путь для органических веществ. Торцевые стенки клеток ситовидных трубок постепенно покрываются порами и начинают напоминать сито – это ситовидные пластинки. Во флоэме имеются клетки и других видов: лубяные волокна, лубяная паренхима, склереиды.
Лекция добавлена 28.02.2013 в 21:59:45uclg.ru
Большая Энциклопедия Нефти и Газа, статья, страница 1
Ксилема
Cтраница 1
Ксилема ( основная водопроводящая ткань) представляет собой прочную ткань, состоящую из живых и отмерших клеток. Из ксилемы состоит большая часть древесины. В травянистых растениях также содержится ксилема, но в значительно меньших количествах, чем в древесных. Во-допроводящие функции ксилемы осуществляются через специализированные неживые клетки - трахеиды. Это удлиненные, суживающиеся к концам клетки с прочными стенками, обычно одревесневшими, хотя и не очень толстыми. Вода с растворенными в ней веществами ( клеточный сок) легко движется по полым трахеидам, диффундируя из клетки в клетку через многочисленные поры. Клеточные стенки у трахеид часто неравномерно утолщенные или скульптурные, благодаря чему они легко различимы на продольном срезе. [1]
Ксилема образуется также путем дифференциации верхушечных меристем корней и побегов. У многолетних древесных растений вторичная ксилема имеет вид хорошо известных годичных колец. Весенняя древесина состоит из более крупных и тонкостенных клеток и выглядит более светлой и менее плотной, чем летняя. [2]
Ксилема выполняет в растении две основные функции: по ней движется вода вместе с растворенными минеральными веществами и она служит опорой органам растения. Таким образом, ксилема играет в растении двоякую роль - физиологическую и структурную. В состав ксилемы входят гистологические элементы четырех типов: трахеиды, сосуды, паренхимные клетки и волокна. На рис. 6.9 эти гистологические элементы представлены и поперечном и продольном разрезах. [4]
Ксилема состоит из длинных трубок, образованных в результате слияния соседних клеток и разрушения поперечных стенок между ними. [5]
Ксилема плауна первичная, она возникает исключительно из прокамбия. [7]
Ксилема цветковых растений содержит два типа проводящих воду структур - трахеиды и сосуды. Строение вторичной ксилемы ( древесины) рассматривается в гл. Ксилема и флоэма образуют проводящую ткань высших, или сосудистых, растений. [8]
Ксилему, образующуюся из прокамбия, называют первичной ксилемой. Она отчетливо подразделяется на два структурных типа - протоксилему, образующуюся на самых первых этапах дифференциации элементов ксилемы, и возникающую несколько позднее мета ксилем у. Отличия в строении этих двух типов мы опишем несколько позднее, а сейчас рассмотрим порядок образования элементов первичной ксилемы. Начнем с того, что прокамбий у различных растений возникает в верхушках осевых органов различными путями. Он может представлять собой на поперечном срезе небольшой кружок ( а иногда и более сложную фигуру), лежащий в центре осевого органа, и, напротив, может развиться в виде небольшого пояска, к периферии и к центру от которого лежат неспециализированные клетки апикальной меристемы. [9]
Обесцвечивание ксилемы на первом узле оценивалось от 0 до 5, где О - отсутствие, а 5 - полное обесцвечивание. [10]
Слои ксилемы лигнифицируются ( древеснеют) и накапливаются в дереве, образуя основную часть ствола. В флоэме лигнификация либо вовсе отсутствует, либо происходит только в некоторых клетках; флоэма образуется из года в год снаружи слоя камбия. По мере образования новой флоэмы слой ее отодвигается все дальше и дальше к наружной стороне ствола и в конце концов образует кору. [11]
Слой ксилемы, образующийся в течение какого-то определенного года, обычно в шесть или восемь раз толще соответствующего слоя флоэмы, поэтому нарастание древесины в стволе дерева происходит гораздо быстрее, чем нарастание коры. [12]
В ксилеме ток жидкости направлен в одну сторону-к местам испарения влаги. Здесь транспортируются главным образом минеральные соли и азотсодержащие соединения, которые, вероятно, поступают сюда в результате активной секреции их специализированными клетками корневой паренхимы. [14]
ДРЕВЕСИНА ( ксилема), ткань древесных и кустарниковых растений, придающая им мех. [15]
Страницы: 1 2 3 4 5
www.ngpedia.ru









