
Справочник химика 21. Крупная вакуоль у растений
Вакуоли растений - Справочник химика 21
Пигментная система фотосинтезирующих растений — сложная смесь, анализ которой представляет немало трудностей. Экстрагирование разрушает химические единицы, содержащие пигменты пластид в естественном состоянии, разбавляет их пигментами вакуолей и клеточных стенок, не имеющими отношения к фотосинтезу, и приводит их в соприкосновение с клеточными компонентами последние могут оказывать химическое действие на пигменты (например кислоты и энзимы). Разделение экстрагированной смеси на ее компоненты легко может повести к дальнейшей деструкции при соприкосновении с воздухом, растворителем или адсорбентом. Полное разделение затрудняется еще и тем, что смесь пигментов содержит изомеры или другие компоненты, мало отличающиеся друг от друга по растворимости и химическим свойствам. [c.401] В вакуолях растения могут накапливаться конечные и некоторые вторичные продукты метаболизма. Из конечных продук- [c.207]Получение препаратов НК из растений, особенно из дифференцированных тканей, осложняется низким содержанием НК. при наличии высокоактивных нуклеаз, обилием углеводных компонентов, полифосфатов, вакуолей и трудностью разрушения клеточных оболочек. При выделении нуклеиновых кислот из растений нео бходимо уделять особое внимание ингибиции нуклеаз, полноте гомогенизации и очистке препарата от сопутствующих примесей. [c.53]
Интересно, что многие растения накапливают значительные количества малата в цитоплазме и в вакуолях. Очевидно, он служит резервным материалом, использующимся при синтезе углеводов. [c.60]
Однако в модельных опытах невозможно учесть все многообразие условий, которые возникают в растительной клетке во время контакта с гифой гриба. В частности, неясно, как и когда гифа проникающего паразита вступает в контакт с фенольными соединениями клетки, находящимися в вакуоли, отделенной от оболочки слоем цитоплазмы. В некоторых, весьма немногочисленных исследованиях, например в работах Арциховской [2, 4], приводится анатомическая картина проникновения гриба и его распространения по тканям листа капусты и отмечается, что еще ни разу не удалось обнаружить проникновения гиф в живую клетку во всех случаях этому проникновению предшествует гибель клетки под влиянием токсина гриба. Факультативные паразиты образуют большое количество ферментов, расщепляющих пластические вещества растений и переводящих их в легко усвояемую для паразитов форму. В таких загнивших тканях содержание спирторастворимых фенолов значительно снижается (табл, 2).. [c.276]
Базисные ткани относятся к разряду мало специализированных, возникающих у растений из клеток апикальных меристем, у грибов имеются немногие соответствующие органоиды (не ткани), которые функционально сходны с базисными тканями — это преимущественно вакуоли с запасными питательными веществами [c.119]
Каллус можно механически разделить на одиночные клетки и небольшие комочки клеток и выращивать их затем в суспензионной культуре. В такой культуре клетки очень похожи друг на друга все они имеют тонкую первичную клеточную стенку и крупные вакуоли, пересеченные тонкими цитоплазматическими нитями (рис. 19-66). В ряде случаев из одиночных клеток, выделенных из суспензии, удавалось вырастить целое взрослое растение. Эта [c.205]
Широко распространена в растениях. Часто присутствует в большом количестве в вакуолях [c.153]Трудности криосохранения растений связаны со спецификой растительных клеток. Клетки растений имеют большие размеры (в культуре тканей они изменяются от 15 до 1000 мкм), прочную целлюлозную стенку и вакуоли. Причем именно степень вакуолизации играет основную роль в устойчивости клеток к действию низких температур. В зрелой клетке центральная вакуоль занимает [c.200]
Так называемым органом локального выделения (Резник, [801) у растений являются вакуоли и стенки клеток. В результате химических превращений токсичные вещества становятся нерастворимыми, нетоксичными или летучими. Этим, по-видимому, объясняется поразительное разнообразие вторичных веществ, найденных в высших растениях. Водорастворимые вещества, подобные антоцианам, проникают в вакуоли, в то время как другие фенольные соединения преобразуются в лигнины стенок клетки. Из работы, рассмотренной в этой главе, ясно, что обе группы возникают как метаболиты ароматических аминокислот, особенно фенилаланина. [c.278]
При токсическом действии металлов вырабатывается общая переносимость (толерантность) организма к повышенному количеству металлов или к чужеродным металлам, происходит неспецифическое связывание металлов или их накопление в коже, костях, волосах, в вакуолях растений и лизо-сомах до определенного порогового уровня. Часто выделяются слизистые жидкости (особенно у рыб), содержащие полисахариды и другие связывающие металлы вещества, богатые группами -ОН, -СООН, -SO3H все они вступают в реакцию с катионами. Вот почему в организме рыб обнаруживают высокие концентрации металлов свинца, цинка, марганца и др. [c.41]
Некоторые исследователи подчеркивали значение таннинов для движений мимозы. Эта гетерогенная группа конденсированных фенольных соединений может образовывать комплексы как с белками, так и неорганическими ионами. Таннины денатурируют белки и могли бы повреждать клетку, но не вызывают вредных эффектов, пока они изолированы в мембранных мешочках вакуолей. Видимо, их особенно много в вакуолях растений, способных к тигмонастическим движениям (рис. 13.4). Во время таких движений танниновые агрегаты в вакуолях диссоциируют, и некоторые таннины, видимо, выделяются через мембраны моторных клеток. Затем они диффундируют в гидатоды (поры) в эпидермисе листовой подушечки. Поскольку таннины обладают вяжущим вкусом и действуют как природные репелленты, выделение таннинов при движении листа может способствовать защите растения от насекомых. [c.395]
Другой источник Са + — внутриклеточные мембранные системы (как правило, сеть эндоплазматического ретикулума, митохондрии, вакуоли растений, ядра). Выход Са=+ из ретикулярных резервуаров осуществляется под действием электрического сигнала, передающегося с плазмалеммы (быстрые скелетные мышцы), самого Са=+, поступающие извне (сердце, медленные мышцы), инозиттрисфосфата (клетки гладких мышц, тромбоциты, большинство других клеток). Быстрый выброс Са + из внутреннего объема ретикулума обеспечивается открытием селективных каналов. [c.114]
Цитоплазма эвглены содержит ядро и многочисленные (более двадцати) зеленые овальные хлоропласты, придающие ей зеленый цвет. В хлоропластах содержится хлорофилл, с помощью которого этот организм фотосинтезирует клеточное вещество, как растения. Но хлорофилл исчезает, когда эвглена попадает в темноту. В новых условиях она усваивает растворенные органические вещества. Следовательно, этот организм на свету проявляет шризнаки растения, а в темноте — животного, Продукты обмена и избыточная влага выводятся из организма через сократительную вакуоль. Разм 10жается эвглена простым делением. Образует цисты. [c.277]
Биосинтез Ф. тесно связан с метаболизмом сахарозы, к-рая служит как донором, так и акцептором остатков -D-фруктофуранозы. В клетках растений Ф. накапливаются в вакуолях и выполняют роль резервного материала (источник фруктозы), осморегулятора и антифриза. Содержание Ф. доходит до 30% от сухой массы в листьях, а в специализир. запасающих органах (корневищах, клубнях и луковицах) может превышать 60%. [c.191]
Растительные клетки имеют возможность откладывать в запас белки в основных своих компартментах. Величина такого накопления очень изменчива, неодинакова у разных растений, органов, тканей. Бесспорно, семена содержат самые большие количества этих запасных белков и поэтому служат важным источником питания для человека и животных. Запасаемые и хранимые в вакуолях, эти белки находятся в форме белковых телец, или алейроновых зерен, число, распределение, структура и состав которых характерны для определенного ботанического вида. Скопления белков, которые можно наблюдать в растительных клетках за пределами вакуолей, почти всегда имеют кристаллическую или псевдокристаллическую структуру и, как правило, менее обильны. Поскольку с помош,ью генетических методов пытаются повысить содержание белковых телец вакуольного происхождения, вероятно, можно предполагать получение таких видов растений, которые будут накапливать большое количество белков и в других отделах клетки. [c.141]
Флавоноиды являются почти исключительно пигментами высших растений. Они могут синтезироваться и накапливаться во всех растительных тканях — в листьях, древесине, корнях, плодах, семенах и во всех частях цветков, главным образом в лепестках. Природные гликозиды растворимы в воде и обнарухклеточном соке или в вакуолях. В недавних сообщениях высказано предположение, что небольшие количества флавоноидов могут присутствовать в хло--ропластах высших растений. [c.137]
Ингибиторы амилаз у танидоносных растений (таннины) находятся в растворенном состоянии в клеточных вакуолях. С помощью гистохимических реакций было установлено, что большинство таниинов локализовано в листьях в обкладочных клетках, окружающих жилки. Это дало основание предположить, что таннины образуются в листьях, откуда проникают в другие части растений. [c.215]
Фруктаны. Фруктаны обычно присутствуют в клеточном соке вакуолей в растворенном состоянии. Полисахариды, построенные из остатков D-фруктозы, как и крахмал, выполняют роль энергетического резерва для организма, и, кроме того, их присутствие необходимо для поддержания так называемого "тургорного раствора" [4 . Фруктаны обнаружены в ряде высших растений, в первую очередь в представителях сложноцветных и злаковых. Известны два типа растительных фрукта-нов тип инулина и тип флеана [3]. [c.267]
В клетках растений резервные полисахариды могут находиться и в растворенном состоянии — в клеточном соке вакуолей. В этом соке обычно присутствуют фруктаны, а также сахароза и аналогичные олигосахариды, вьшолняюш,ие, по-видимому, близкие биологические функции. Полисахариды, содержащиеся в вакуолярном соке, важны не только как резервы пищи организма. Их присутствие необходимо для поддержания так называемого тургорного давления — давления изнутри содержимого клетки на клеточную стенку, что имеет первостепенное значение для всей жизнедеятельности растений. [c.600]
Тонопласты - крупные прозрачные вакуоли, которые окружены типичными мембранами, встречаются у грибов и растений. Вакуоли содержат различные гидролитические ферменты 2 типа протеаз, эс-теразу, РНКазу, аминопептидазу и липоамиддегидрогеназу (НАДН-зависимую). Высказываются предположения, что эти ферменты не связаны с тонопластом. Вероятно, их роль транспортная, хотя есть много вопросов. [c.43]
Клетки трибов и водорослей по своей организации похожи на клетки высших растений. Основными частями клетки являются оболочка, протоплазма (цитоплазма) и ядро (нуклеус). В состав оболочки входит целлюлоза. Протоплазма представляет собой сложное коллоидное образование с резко выраженным поверхностным натяжением. В этой коллоидной системе непрерывной фазой является вода, а дисперсной фазой — липопротеиновые соединения. В протоплазме одноклеточных грибных организмов — дрожжей— легко обнаруживаются вакуоли, представляющие собой пустоты, заполненные клеточным соком. При делении вакуоли дочерней клетки образуются путем отпочковы-вания от вакуоли материнской клетки. В протоплазме имеются также мельчайшие гранулы-—рибосомы (микросомы), размеры которых составляют 200 ммк, обнаружить их можно лишь методом электронной микроскопии. Б рибосомах, состоящих из рибонуклеиновой кислоты и белка, происходит белковый синтез- [c.113]В вакуолях могут накапливаться и храниться самые различные молекулы, в том числе те необходимые для клетки вещества, которые потенциально опасны, если поступают в цитоплазму в больших количествах. Например, у некоторых растений в вакуолях специализированных клеток накапливаются такие хорошо известные продукты, как каучук и опиум. Даже вездесущие ионы Na могут селективно накапливаться в этих органеллах, где их осмотическая активность способствует поддержанию тургорного давления. Исследования, проведенные на клетках ЫНеНа, показали, что натриевые насосы в то-нопласте поддерживают относительно низкую концентрацию N3 в цитозоле за счет создания 4-5-кратного избытка зтих ионов в вакуолях. А поскольку вакуоль занимает в клетке ИНеИа гораздо больший объем, чем цитоплазма, получается, что основной пул клеточного натрия сосредоточен в вакуолях. [c.186]
Среди продуктов, запасаемых в вакуолях, важное место занимают различные метаболиты. Например, растения-суккуленты ночью погло1Шют из воздуха двуокись углерода и хранят ее в вакуолях в форме малата, пока на следующий день он не будет с помощью солнечной энергии превращен в сахар. Органические молекулы могут храниться в вакуолях и в течение гораздо более длительного срока, как, например, запасвые белки в клетках многих семян (рис. 19-34). Часто в одной и той же клетке имеются вакуоли, выполняю- [c.186]
Клеточная оболочка — это мембрана, которая регулирует связь цитоплазмы с другими клетками и 1С внещней средой. Мембрана избирательно проницаема для различных веществ, ее проницаемость зависит от природы проникающих в клетку молекул и физиологических особенностей клетки. В цитоплазме находятся различные включения — капельки жира, зерна крахмала и т. д., вакуоли. В вакуолях содержится клеточный сок. В клеточный сок растений входят различные пигменты, определяющие окраску растений и их отдельных органов. Желтая окраска обусловлена флавонами, а красная и фиолетовая— антоцианинами. Окраска зависит также от кислотности сока. Главнейщими клеточными структурами, которые содержатся в цитоплазме, являются ядро, пластиды, митохондрии и микросомы. Пластиды—довольно крупные гранулы овальной формы, митохондрии — мелкие палочковидные частицы, а микросомы — мельчайшие округлые частицы. Митохондрии и микросомы хотя и значительно меньше ядра или пластид, но на их долю приходится до 50% массы протоплазмы. В протоплазме имеется сложная система мембран, образующих каналы, связанные с оболочкой ядра. Эта система представляет структурную основу клеточной цитоплазмы и называется эндоплаз-матической сетью. [c.28]
Рад веществ, накапливаемых в вакуолях, участвует во взаимодействии растений с животными или другими растениями. Антощ анины, например, придают окраску лепесткам цветков, что способствует привлечению насекомых-опылителей. Другие вещества выполн иот защитные функции. Растения не могут передвигаться и таким образом избегать уничтожения травоядными животными вместо этого они синтезируют бесчисленное множество ядовитых веществ, которые высвобождаются из вакуолей при повреждения клеток. Здесь можно найти и высокотоксичные алкалоиды, и просто неприятные на вкус вещества, отрицательно влияющие на пищеварение. На протяжешга всей своей истории растения, так же как и животные, постоянно разнообразили средства ведения скрытой химической войны. Равновесие смещалось в ту или иную сторону, когда, например, в растительном мире возникал новый мошщ>1Й репеллент для растительноядных видов или, наоборот, когда какое-нибудь насекомое в ходе эволюция научалось нейтрализовать или разрушать токсичный растительный метаболит и получало таким образом возможность поедать синтезирующее его растение. Сам токсин в этом случае мог становиться уже не репеллентом, а аттрактантом. [c.187]
Тонкие клегки, образующие волоски на поверхности растений, прозрачны, что позволяет без труда наблюдать движение цитоплазмы на живом объекте. Эти клетки содержат крупные вакуоли, через которые тянутся тонкие (толщиной около I мкм) жгуты цитоплазмы (рис. 19-46). Видно, как по этим цитоплазматическим тяжам быстро движутся различные частицы, например митохондрии. Создается впечатление, что эти тяжи расходятся из области, лежащей по соседству с клеточным ядром при этом они непрерывно меняют свою форму и расположение, сливаются друг с другом, ветвятся, сжимаются, исчезают и образуются заново. [c.194]
Имеется множество данных, что цикл Кребса распространен в растениях, но было предпринято очень мало попыток определить его количественную роль в дыхании. Ряд исследователей наблюдали, что при подкормке листьев меченными С органическими кислотами скорость включения метки в другие кислоты цикла Кребса относительно низка. На основании этих данных еще нельзя делать вывод о медленном протекании реакций цикла Кребса, так как меченые кислоты могут быть разбавлены большим количеством эндогенных немеченых кислот, присутствующих в вакуолях. Другой подход к этой проблеме заключается в решении вопроса, достаточно ли активны различные ферменты цикла для того, чтобы за счет их действия можно было отнести наблюдаемые скорости дыхания. Утверждали, что сукцинатоксидазная активность митохондрий достаточна, чтобы объяснить общую скорость дыхания [c.195]
У большинства видов этих водорослей клетки имеют целлюлозную оболочку, вакуоль с клеточным соком, как правило, одно дифференцированное ядро и хроматофо-ры, форма которых разнообразна пластинки, сеточки, звездочки, диски. Зеленые водоросли содержат те же пигменты, что и высшие растения, т. е. хлорофилл и каротин. Размножение половое и бесполое с образованием подвижных зооспор. [c.44]
Своим названием вся эта группа организмов обязана самым заметным ее представителям-шляпочным грибам (греч. mykes, лат. fungus). Грибы относятся к эукариотам. С растениями их сближает ряд общих признаков наличие клеточной стенки и вакуолей, заполненных клеточным соком хорошо видимое под микроскопом движение протоплазмы неспособность к активному перемещению. У грибов, однако, нет фотосинтетических пигментов это С-гетеротрофы (точнее, хемоорганогетеро--трофы). Грибы растут в аэробных условиях и получают энергию путем окисления органических веществ. По сравнению с растениями, имеющими стебель, корни и листья, грибы слабо дифференцированы морфологически, и у них почти нет разделения функций между разными частями организма. [c.155]
По-видимому, существуют различия между мхами и высшими растениями в поглощении свинца и в его распределении в тканях. У различных мхов были найдены электроноплотные отложения свинца в ядрах, пластидах, вакуолях, митохондриях и плазмодесмах. В отличие от этого у рдеста (Potamogeton) свинец изолирован в виде электроноплотного осадка в клеточной стенке и лишь в незначительном количестве поглощается путем пиноцитоза. Видимо, таким же образом живые деревья депонируют свинец в коре вне клеток в форме электроноплотного материала. Эти различия могли бы быть одной из причин чувствительности низших растений к загрязнению воздуха. [c.72]
В непосредственной близости к кончику корешка (на расстоянии примерно один миллиметр от (его) находится зона меристемы (делящихся клеток) выше ее расположен зона растяжения, в которой клетки увеличиваются в объеме, удерживают много воды, образуют вакуоли. Здесь начинается дифференцирование тканей и возникновение проводящей системы одни клетки дают начало флоэме, по которой передвигаются органические вещества, другие — ксиле11е, по ней движется вода. Проводящие системы связывают корень с надземцой частью растения. Дифференцирование тканей и завершение образования проводящей системы заканчивается в зоне возникновения корневых волосков. Все эти зоны имеют сравнительно проницаемую наружную целлюлозную оболочку, лишенную кутикулы. Схемати-ческл строение корня в зоце корневых волосков показано на рисунках 4—5 (поперечный и продольный разрезы при сильном увеличении). Еще выше расположена зона ветвления корней. [c.49]
Однако каким же образом передвигаются поглощенные плазмолеммой ионы через основную массу протоплазмы (мезоплазму) и тонопласт (внутренний поверхностный слой протоплазмы) в вакуоль с клеточным соком (рхте. 9) и в конечном счете — в сосудистую систему корня, чтобы затем, уже будучи подхваченными током транспирации, подняться в надземную часть растения [c.61]
Результаты Миллера показывают, что гликозиды, образовавшиеся из введенных агликонов, не влияют на нормальный рост и развитие растения. Вероятно, в клетках агликоны в виде гликозидов переносятся в места с низкой метаболической активностью, такие, как вакуоли, где и остаются до отмирания клетки. Миллер также установил, что эти гликозиды не переносятся из тканей, в которых они синтезировались. В побегах из клубней картофеля и луковиц гладиолуса, содержащих 2-хлорэтил-р-в-глюкозид, не было глюкозида. Луковицы гладиолуса, содержащие 2-хлорфенилгентиобиозид, выращивали до образования новых луковиц, однако гликозид не был обнаружен ни в полученных побегах, ни в дочерней луковице, хотя старые луковицы еще содержали измеримые количества гликозида [14]. Семена табака, который накапливает большие количества гликозида хлоральгидрата в листьях, побегах и корнях, не содержали этого соединения [13]. [c.201]
Чирх [9] выдвинул положение о существовании у хлоропластов мембраны, утверждая, что она предохраняет хлоропласты от срастайся и защищает хлорофилл от разрушения органическими кислотами, присутствующими в соке многих растений. Другие исследователи, например Шмитц [7], считали мембрану оптической ил.люзией. В более позднее время Вилер [34] подтвердил существование мембраны у хлоропластов Elodea anadensis, а Граник [50] установил, что полупроницаемая мембрана позволяет изолированным хлоропластам томатов и табака оставаться неразрушенными в течение нескольких часов в 0,5 Ж растворе глюкозы. Изолированные хлоропласты, помещенные в дестиллированную воду, набухают, вакуоли-зируются и разрушаются [57]. [c.361]
chem21.info
"Центральная вакуоль растительной клетки" страница 2
Для клеток растений характерна хорошо развитая система вакуолей, в значительной степени обеспечивающая их соматические свойства.
Почти во всех клетках, особенно в старых, хорошо заметны полости – вакуоли. Они заполнены клеточным соком.
Клеточный сок – это вода с растворенными в ней сахарами и другими органическими веществами. В клеточном соке могут содержаться красящие вещества, придающие синюю, фиолетовую, малиновую окраску лепесткам и другим органам растений. Осенняя окраска листьев также обусловлена окрашенным клеточным соком. Разрезая спелый плод или другую сочную часть растения, мы повреждаем клетки, и из их вакуолей вытекает сок. Цитоплазма в каждой из клеток оттеснена вакуолью к оболочке. Молодые клетки, в отличии от старых, неспособных делиться, содержат много вакуолей. В старой клетке обычно имеется одна большая вакуоль.
II. ОСНОВНОЕ СОДЕРЖАНИЕ.
Р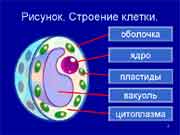 астительные клетки отличаются наличием толстой целлюлозной клеточной стенки, пластид, крупной центральной вакуоли, смещающей ядро к периферии. Клеточный центр высших растений не содержит центриоли. Запасным углеводом является крахмал.
астительные клетки отличаются наличием толстой целлюлозной клеточной стенки, пластид, крупной центральной вакуоли, смещающей ядро к периферии. Клеточный центр высших растений не содержит центриоли. Запасным углеводом является крахмал.
Вакуоль (франц. vacuole, от лат. vacuus — пустой) – наполненный жидкостью мембранный мешочек. В животных клетках могут наблюдаться небольшие вакуоли, выполняющие фагоцитарную, пищеварительную, сократительную и другие функции. Растительные клетки имеют одну большую центральную вакуоль. Жидкость, заполняющая её, называется клеточным соком. Это концентрированный раствор сахаров, минеральных солей, органических кислот, пигментов и других веществ. Вакуоли накапливают воду, могут содержать красящие пигменты, защитные вещества (например, таннины), гидролитические ферменты, вызывающие автолиз клетки, отходы жизнедеятельности, запасные питательные вещества. У растений вакуоли - производные эндоплазматической сети, окружены полупроницаемой мембраной - тонопластом. Вся система вакуолей растительной клетки называется вакуомом, который в молодой клетке представлен системой канальцев и пузырьков; по мере роста и дифференцировки клетки они увеличиваются и сливаются в одну большую центральную вакуолю, занимающую 70- 95% объема зрелой клетки.
Вакуоли растений, строение и функции.
В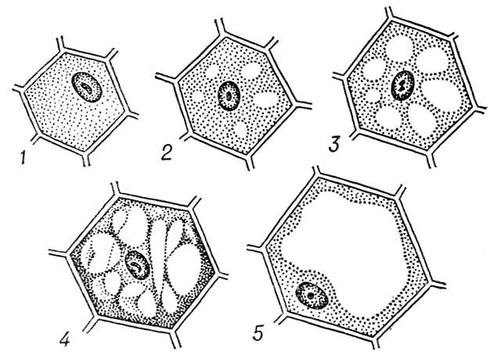 очень молодых клетках вакуолей нет или они почти незаметны по мере роста и дифференцировки клетки вакуоли появляются в разных её участках, а затем, постепенно увеличиваясь, сливаются друг с другом и образуют одну или несколько крупных вакуолей, занимающих до 80% объема всей клетки.
очень молодых клетках вакуолей нет или они почти незаметны по мере роста и дифференцировки клетки вакуоли появляются в разных её участках, а затем, постепенно увеличиваясь, сливаются друг с другом и образуют одну или несколько крупных вакуолей, занимающих до 80% объема всей клетки.
Центральные вакуоли отделены от цитоплазмы одинарной липоидно-белковой полупроницаемой мембраной, сходной по толщине с плазмалеммой. Мембрана, ограничивающая центральные вакуоли, носит название тонопласта. Возникают центральные вакуоли из мелких пузырьков, отщепившихся от эндоплазматической сети. Такие первичные провакуоли растут в объеме, сливаются друг с другом и, в конце концов, образуют одну или несколько крупных вакуолей, оттесняющих цитоплазму с ядром и органоидами к периферии клетки.
infourok.ru
Вакуоли растительных клеток - Справочник химика 21
Вакуоли. Растительные клетки отличаются от животных хорошо развитой системой вакуолей, заполненных клеточным со- [c.128] Однако в модельных опытах невозможно учесть все многообразие условий, которые возникают в растительной клетке во время контакта с гифой гриба. В частности, неясно, как и когда гифа проникающего паразита вступает в контакт с фенольными соединениями клетки, находящимися в вакуоли, отделенной от оболочки слоем цитоплазмы. В некоторых, весьма немногочисленных исследованиях, например в работах Арциховской [2, 4], приводится анатомическая картина проникновения гриба и его распространения по тканям листа капусты и отмечается, что еще ни разу не удалось обнаружить проникновения гиф в живую клетку во всех случаях этому проникновению предшествует гибель клетки под влиянием токсина гриба. Факультативные паразиты образуют большое количество ферментов, расщепляющих пластические вещества растений и переводящих их в легко усвояемую для паразитов форму. В таких загнивших тканях содержание спирторастворимых фенолов значительно снижается (табл, 2).. [c.276]На рис. 13.1. показаны полупроницаемые мембраны, имеющие важное значение для водного режима растительной клетки. Клеточная стенка обычно полностью проницаема для любых растворенных молекул, поэтому ее нельзя считать осмотическим барьером. В клетке часто имеется крупная центральная вакуоль, содер- [c.100]
Эта система участвует не только в синтезе ферментов, которые сек-ретируются клеткой, но и в образовании новых мембран. По-видимому, шероховатый ЭР поставляет мембранный материал гладкому ЭР и аппарату Гольджи, а компоненты мембран Гольджи включаются в состав наружной клеточной мембраны. В растительных клетках наружные мембраны митохондрий и мембраны, окружающие вакуоли, также образуются непосредственно из ЭР [19]. Компоненты наружных клеточных мембран, вероятно, могут использоваться повторно, включаясь в соответствующую структуру в ходе эндоцитоза [20]. [c.33]
Живая клетка имеет три части оболочку живое содержимое -протопласт вакуоли. У растительных клеток формирование оболочки (клеточной стенки) обусловлено деятельностью протопласта. Вакуоль -это полость с клеточным соком. Вакуоли в клетках образуются и увеличиваются в объеме по мере роста клеток. Протопласт (протоплазма) состоит из цитоплазмы и включенных в нее органоидов (органелл). [c.195]
В процессе роста растительной клетки происходит образование маленьких пузырьков, которые увеличиваются в объеме и, наконец, сливаются друг с другом, образуя вакуоли, так что в конце концов протоплазма оттесняется к клеточной оболочке, создавая здесь тонкий пристеночный слой. Вакуоль окружена типичной элементарной мембраной, известной под названием тонопласта, или вакуолярной мембраны. Вопрос о происхождении этой мембраны часто обсуждался в литературе по этому поводу существуют две распространенные точки зрения. Одни авторы утверждают, что вакуоль появляется в результате дальнейшего развития пузырьков эндоплазматического ретикулума. Такой механизм ставится, однако, под сомнение из-за очевидного отсутствия связи между мембраной вакуоли с одной стороны и эндоплазматическим ретикулумом или мембраной ядра — с другой. [c.53]
Типичная растительная клетка состоит из протопласта, окруженного полупроницаемой мембраной. Этот протопласт со своей мембраной напоминает соответствующие структуры клеток всех высших организмов, если пе считать того, что в растущей клетке он содержит большую вакуоль, наполненную водой с растворенными в ней веществами. Вакуоль представляет собой, [c.505]
При изменении нормального физико-химического состояния протоплазмы изменяется отношение ее,к красителям. Некоторые красители (нейтральный красный, метиленовый синий, тионин и др.), проникнув в нормальную клетку, оставляют цитоплазму и ядро неокрашенными. В животной клетке они откладываются в цитоплазме в виде мелких гранул или же связываются с некоторыми клеточными включениями. В растительной клетке эти красители гранул не образуют и обычно накапливаются в клеточной вакуоли. Цитоплазма и ядро и здесь остаются бесцветными. При повреждении любой клетки распределение в ней красителя нарушается. Прежде всего парализуется способность клетки откладывать краситель в цитоплазме в виде гранул. Одновременно цитоплазма и ядро приобретают сродство к красителю и начинают все более и более прокрашиваться. [c.17]
В растительных клетках роль лизосом могут играть крупные центральные вакуоли. Впрочем, в цитоплазме растительных клеток иногда видны тельца, напоминающие по своему виду лизосомы животных клеток. [c.199]
Вакуоль наполнена клеточным (вакуолярным) соком, в кото-ро.м содержатся различные метаболиты,— продукты жизнедеятельности протоплазмы, различные неорганические и органические соединения, в их числе органические кислоты, свободные аминокислоты, растворимые белки, алкалоиды и глюкозиды, воднорастворимые пигменты (антоцианы и др.), дубильные вещества и т. п. В последние годы установлено присутствие в клеточном соке ряда ферментов, что указывает на его физиологическую активность. С наличием вакуоли в большой степени связаны осмотические свойства растительной клетки, являющиеся одним из факторов, регулирующих процессы поступления и передвижения воды и минеральных солей. [c.34]
Как уже отмечалось, осмотические явления наблюдаются лишь в системах, содержащих так называемые полупроницаемые перепонки, свободно пропускающие через себя растворители и лишь частично проницаемые для растворенных в последних веществ. В растительной клетке роль такой полупроницаемой мембраны выполняет вся протоплазма и, в первую очередь, ее пограничные слои — мембраны. Считают, что различия в проницаемости клеточной оболочки и пограничных слоев протоплазмы, а также имеющаяся в клетке вакуоль с клеточным соком являются факторами, от которых в значительной степени зависит скорость проникновения в клетку и выделения клеткой воды. [c.68]Лизосомы также ограничены однослойной мембраной. Матрикс их оптически неоднороден и содержит ряд уплотнений. В лизосомах локализован набор гидролитических ферментов, участвующих в разрушении продуктов клеточного метаболизма, причем при помощи специального протонного насоса поддерживается низкое значение pH (не более 4,5), способствующее эффективному гидролизу. Внутриклеточные структуры, подлежащие разрушению, поступают в лизосомы, где и подвергаются гидролизу. Процесс селекции и поступления в лизосомы только отработанного материала обусловлен его специфическим мечением. Так, нативные белки в лизосомы не поступают. По истечении же времени функционирования происходит их инактивация цитоплазматическими протеиназами или присоединение убиквитина, что является сигналом для транспорта в лизосомы модифицирбванного белка. Кроме молекул, лизосомы могут разрушать органеллы или целые клетки (митохондрии, эритроциты). Процесс транспорта веществ в лизосомы является энергозависимым и требует затраты энергии. В растительных клетках гидролитические ферменты обычно локализованы в вакуолях — прообразе лизосом. [c.13]
Клетки высших растений содержат те же внутриклеточные компартменты, которые ранее были описаны для животных клеток, - это цитозоль, аппарат Гольджи, эндоплазматический ретикулум, ядро, митохондрии, пероксисомы и лизосомы. Кроме гого, растительные клетки обладают цитоскелетом, состоящим из актиновых филаментов, микротрубочек и промежуточных филаментов, сравнимых с теми, которые обнаруживаются в клетках животных. Тем не менее клетки растений легко отличить от клеток животных по присутствию в них двух особых типов компартментов, окруженных мембраной -вакуолей и пластид. Наличие данных [c.411]
Среда должна обладать достаточной буферной емкостью это позволяет избежать сдвига pH цитоплазмы, обусловленного высвобождением из вакуоли растительной клетки органических кислот. Чаще всего используют тр с-(оксиметил)аминометановый буфер] (или просто трис-буфер) с pH 7—8 (именно таков pH цитоплазмы). Такой буфер в концентрации 0,05 М достаточен для поддержания pH для всех (за исключением особенно кислых) растительных тканей. [c.10]
Рассмотрим теперь поступление воды в вакуоль и из вакуоли растительной клетки in situ (т. е. в растении). Вакуоль и окружающий ее протопласт заключены внутри клеточной стенки, через которую вода диффундирует свободно. (Клеточная стенка в этом смысле напоминает фильтровальную бумагу, которая тоже состоит из целлюлозы.) Клеточная стенка насыщена водой до тех пор, пока влажность почвы достаточна, а транспирация не слишком интенсивна. В этих условиях в об- [c.173]
Два типа органелл-пластиды и вакуоли-свойственны только растительным клеткам. Пластиды составляют неоднородную группу органелл, из которых наиболее известны фотосинтезирующие хлоропласты, имеющиеся во всех зеленых тканях. Вакуоль представляет собой крупную внутриклеточную полость, заполненную водным раствором того или иного состава и ограниченную мембраной, назмваемой тонопластом. Растительные клетки используют вакуоли в самых разных целях-например, для экономного заполнения внутриклеточного пространства при росте, для хранения запасов питательных веществ или для накопления вредных продуктов обмена. Хотя сами растительные клетки не обладают способностью двигаться, их цитоплазма, особенно в клетках с большими вакуолями, постоянно перемешиваетя в результате поддерживаемых в ней направленных потоков. Показано, что по крайней мере в некоторых случаях движение цитоплазмы связано с функцией цитоплазматических актиновых филаментов. [c.196]
Растительные клетки имеют возможность откладывать в запас белки в основных своих компартментах. Величина такого накопления очень изменчива, неодинакова у разных растений, органов, тканей. Бесспорно, семена содержат самые большие количества этих запасных белков и поэтому служат важным источником питания для человека и животных. Запасаемые и хранимые в вакуолях, эти белки находятся в форме белковых телец, или алейроновых зерен, число, распределение, структура и состав которых характерны для определенного ботанического вида. Скопления белков, которые можно наблюдать в растительных клетках за пределами вакуолей, почти всегда имеют кристаллическую или псевдокристаллическую структуру и, как правило, менее обильны. Поскольку с помош,ью генетических методов пытаются повысить содержание белковых телец вакуольного происхождения, вероятно, можно предполагать получение таких видов растений, которые будут накапливать большое количество белков и в других отделах клетки. [c.141]
Поскольку мы упоминали (см. стр. 239) о том, как должна выглядеть дифференцированная растительная клетка, поговорим более подробно именно о ней. Кстати, рассмотрим еще раз рис. 107. Это нам поможет. Прежде всего нужно отметить значительное увеличение размеров вакуоли, которое происходит главным образом за счет поглощения воды. Клеточное ядро не увеличивается совсем или увеличивается очень незначительно, объем основного вещества цитоплазмы увеличивается, но ненамного. Пропластиды, превращаясь в хлоропласты, увеличиваются в числе то же самое происходит и с митохондриями (предшественников которых аналогично называют промитохондриями). [c.259]
На основании изложенных выше литературных и экспериментальных данных можно предложить следующую гипотетическую схему биохимических процессов, возникающих в простейшем случае лод действием ауксина в растительной клетке, находящейся в фазе растяжения. Молекула ИУК, являющаяся донором электрона, образует лабильный комплекс с гипотетическим рибонуклеопротеидом-переносчиком поверхностной мембраны. Образование такого комплекса приводит к увеличению количества фосфатных групп, освобождающихся от связи с белком. Активированный таким образом переносчик связывает кальций пектатов клеточных стенок свободными фосфатными группами и транспортирует его на внутреннюю сторону мембран. Эта реакция идет с использованием энергии АТФ, в результате чего усиливается окислительное фосфорилирование и дыхание. В реакции переноса кальция принимают участие сократительные белки, содержащие сульфгидрильные группы. Перемещение кальция сопровождается изменением мембранного потенциала и активности ферментов, локализованных в мембранах и клеточных стенках (аскорбатоксидазы, метилпектинэстеразы). Изменяется также поглощение и выделение ряда катионов и анионов, в частности, увеличивается поглощение калия. В результате удаления части кальция клеточная стенка становится более пластичной, вследствие чего возрастают сосущая сила и поступление воды в вакуоль. Начинается растяжение клеточной оболочки. Переносчик под действием РНК-азы распадается на внутренней стороне мембраны и затем ресинтезируется для переноса новых ионов кальция. Растяжение клеточной стенки индуцирует системы синтеза пектинов, целлюлозы и других компонентов оболочки. Эти процессы также сопровождаются затратой энергии и усилением интенсивности дыхания. Растяжение и увеличение гидратации цитоплазмы приводит к уменьшению ее вязкости и активизации гидролитических ферментов. Вслед за поглощением воды в вакуоль поступают осмотически активные вещества, поддерживающие сосущую силу клетки. [c.42]Клеточный метаболизм требует того, чтобы субстраты, промежуточные продукты, кофакто1 л, сигнальные молекулы и ферменты могли перемещаться из -С одной части клетки в другую. В мелких клетках, таких как бактерии или даже большинство животных клеток, диффузия дает возможность низкомолекулярным растворенным веществам в доли секунды преодолевать расстояния, сравнимые с размерами самой клетки. Одиако растительные клетки благодаря наличию у них клеточной стенки, вакуолей и тургора могут достигать весьма крупных размеров обычно их длина превышает 100 мкм, а в отдельных случаях измеряется миллиметрами и даже сантиметрами. Диффузия здесь относительно неэффективна, так как время, необходимое для того, чтобы какая-то молекула достигла места своего назначения с помощью одной только диффузии, пропорционально квадрату расстояния до этого мес- [c.193]
Интересны результаты применения люминесцентной микроскопии к исследованию растительных тканей. В растительных клетках происходит более или менее значите гьное накопление флуорохромов в вакуолях оно протекает с разной интенсивностью в зависимости от pH и гН окружающей клетки водной среды, а также от состава веществ, растворенных в вакуолях. Флуорохромы образуют различного типа соедине- [c.315]
Растительные клетки содержат также пластиды других типов. В бесцветных лейкопластах запасаются крахмал и масла. Значительное место во многих растительных клетках занимают окруженные одиночной мембраной крупные пузьфь-ш-вакуоли (см. рис. 2-21). Они заполнены клеточным соком и различными продуктами, являющимися отходами метаболизма. Эти продукты часто агрегируют с образованием кристаллических отложений. В молодых клетках вакуоли имеют небольшую величину, но по мере старения клеток их размеры увеличиваются, и часто они заполняют весь объем клетки. Вакуоли встречаются также и в некоторых животных клетках, но здесь они, как правило, значительно мельче. У растительных клеток нет ни ресничек, ни жгутиков. [c.47]
В растительных клетках галогены накапливаются в вакуолях, главным образом в виде свободных ионов. Однако были идентифицированы и некоторые органические галогенсодержащие метаболиты. В основном это соединения микробного нроисхождения. Петти [122] в своей работе приводит список, насчитывающий 29 микробных галометаболитов. Галоген, входящий во все эти соединения, — хлор, причем во всех случаях, кроме одного, хлор присоединен к углеродному атому, входящему в пяти- или шестичленное кольцо. Упомянутое исключение — хлорамфеникол ( 11h22O5 I2), в молекуле которого присутствуют 2 атома хлора, соединенные с концевым атомом углерода. Ни один из этих хлорсодержащих метаболитов не имеет какой-либо существенной метаболической функции. У высших растений хлорметабо-литы не известны. [c.260]
Кристаллы оксалатов, разумеется, встречаются далеко не во всех растительных клетках, и эта микрофотография, собственно говоря, приведена здесь не ради них самих, а потому, что на ней превосходно видна граница между вакуолью и цитоплазмой.. Это опять-таки элементарная мембрана, точно такая же, как и плазмалемма. Первоначально вся вакуоль, т. е. пространство, заполненное клеточным соком, совместно с огра-ничиваюш,ей его мембраной, была названа тонопластом (Гуго де Фриз, 1885). Однако в результате известной небрежности большинство исследователей сейчас называют тонопластом только оболочку вакуоли. Тонопласт и плазмалемму можно считать братьями плазмалемма отграничивает цитоплазму от окружающей среды, а тонопласт — ее же от центральной вакуоли. [c.241]
Закончившая рост взрослая типичная живая растительная клетка имеет следующие части оболочку, протопласт и вакуоль с клеточным соком. Протопласт — живое содержимое клетки. Оболочка и клеточный сок — продукты жизнедеятельности протопласта. Протопласт состоит из протоплазмы (цитоплазмы) и включенных в нее органоидов ядра, пластид, митохондрий (хондриозом). В молодой клетке, образовавшейся в результате деления, цитоплазма заполняет ее сплошь или почти сплошь. Вакуолизация, т. е. появление полостей, заполненных клеточным соком, происходит постепенно, по мере роста клетки. Во взрослой клетке цитоплазма тонким слоем выстилает внутреннюю поверхность оболочки образовавшаяся внутри клетки полость наполнена клеточным соком. [c.13]
Экзоцитоз — процесс обратный эндоцитозу. Таким способом различные материалы выводятся из клеток из пищеварительных вакуолей удаляются оставшиеся непереваренными плотные частицы, а из секретарных клеток путем пиноцитоза наоборот выводится их секрет. Именно так секретируются в частности ферменты поджелудочной железы (рис. 5.29). В растительных клетках путем экзоцитоза экспортируются материалы, необходимые для построения клеточных стенок (рис. 5.30). [c.192]
Если растительная клетка контактирует с раствором, водный потенциал которого ниже, чем у ее содержимого (например, с концентрированным сахарным сиропом см. опыт 13.1), то вода будет выходить из нее за счет осмоса через плазмалемму (рис. 13.2). Сначала воды станет меньше в цитоплазме, а затем и в вакуоле, откуда она выйдет сквозь тонотшаст. Протопласт, т. е. живое содержимое растительной клетки, окруженное клеточной стенкой, спадется и отойдет от этой стенки, как бы сжавшись внутри нее. Этот процесс называется плазмолизом, а клетка в таком состоянии — плазмолизироваппой. Момент, когда протопласт еще прилегает к клеточной стенке, но уже перестал оказывать на нее давление, называется начальным плазмолизом. В этот момент клетка теряет тургор, т. е. становится вялой. Вода будет покидать протопласт до тех пор, пока его содержимое по водному потенциалу не сравняется с окружающим раствором. Тогда установится равновесное состояние, и спадение протопласта прекратится. [c.101]
Интересно, почему же чеснок не пахнет, пока его не начнут резать или толочь Оказывается, запах чеснока появляется в результате распада содержащегося в нем вещества аллиина под действием фермента аллиинлиазы. В целой дольке чеснока они находятся в разных местах растительной клетки аллиин — в цитоплазме (части протоплазмы, не входящей в ядро клетки), а фермент — в полостях клетки (вакуолях). Когда целостность дольки чеснока нарушена, эти два вещества вступают в химическое взаимодействие, и выделяется очень неустойчивая аминокислота аллицин, которая тут же распадается с выделением диаллилсульфида. Он и придает чесноку характерный заг1ах. [c.140]
Вакуоли. Протоплазма растительной клетки характеризуется высокой оводненностью, значительно превышающей таковую клеток животных. В связи с этим у зеленых растений сильно развит вакуолярный аппарат. Цитоплазма молодых клеток лишена единой вакуоли, в ней имеются лишь мельчайшие цистерны, расположенные в каналах эндоплазматической сети. В результате процессов, осуществляющихся в элементах эндоплаз-ыатической сети, в них накапливаются различные низкомолекулярные соединения (сахара, азотистые вещества, пектины и др.). В ходе дальнейшей гидратации протоплазмы размеры этих зачатков будущей вакуоли увеличиваются. Это ведет к их частичному слиянию и превращению в обособленную вакуоль, которая у взрослой клетки занимает большую часть ее объема (рис. 7). [c.34]
Клеточный метаболизм требует того, чтобы субстраты, промежуточные продукты, кофакторы, сигнальные молекулы и ферменты могли перемещаться из одной части клетки в другую. В мелких клетках, таких как бактерии или даже большинство животных клеток, диффузия дает возможность низкомолекулярным растворенным веществам в доли секунды преодолевать расстояния, сравнимые с размерами самой клетки. Однако растительные клетки благодаря наличию у них клеточной стенки, вакуолей и тургора могут достигать весьма крупных размеров обычно их длина превышает 100 мкм, а в отдельных случаях измеряется миллиметрами и даже сантиметрами. Диффузия здесь относительно неэффективна, так как время, необходимое какой-либо молекуле для достижения пункта своего назначения с помощью лишь диффузии, пропорционально квадрату расстояния до этого места (см. разд. 3.1.3). Между тем некоторые клетки взрослого растения могут быть значительно удалены от источника кислорода и питательных веществ. Поэтому неудивительно, что в крупных растительных клетках существуют интенсивные токи цитоилазмы, перемешивающие ее компоненты и обеспечивающие их быструю циркуляцию. [c.423]Если растительная клетка выращивается изолированно, то форма ее обычно приближается к сферической (рис. 2.2), но если она растет в окружении других клеток, то они сдавливают ее, и тогда она принимает форму многогранника. Клетка из зоны растяжения стебля или корня по форме напоминает коро бочку длиной около 50 мкм, шириной 20 мкм и высотой 10 мкм. Объем ее равен приблизительно 10 000 мкм . В одном кубическом сантиметре (1 см ) при плотной упаковке помещается до 100-10 таких клеток. Структура растительной клетки сложна и высокодифференцированна, но в первом приближении мы можем вычленить в ней три главные зоны 1) клеточную стенку — сравнительно жесткое образование, по всей вероятности неживое, представляющее собой высокоструктурированную и в химическом отношении сложную смесь веществ, выделяемых протопластом 2) протопласт — живую часть клетки, в которой заключены все клеточные органеллы, суспендированные здесь в сложном растворе, и 3) вакуоли — неживые образования, как бы мембранные мешки, служащие резервуарами или хранилищами клетки они заполнены водным раствором поглощенных клеткой неорганических солей и органических веществ, представляющих собой продукты метаболической активности клетки. Клеточные стенки у растения играют роль скелета, т. е. обеспе [c.24]
Благодаря своей жесткости клеточные стенки обеспечивают растительным клеткам определенную форму и некий минимальный размер, т. е. служат для растения своего рода скелетом. В клетках с сильно лигнифицированными вторичными клеточными стенками, например в древесине, этот скелет может обеспечить сохранение размера и формы даже при отсутствии всех других поддерживающих сил. В тонкостенных клетках, например в тканях листа, стенки слишком податливы и неспособны сохранять свою форму без поддержки со стороны клеточного содержимого. Поддержку эту создает главным образом давление, развиваемое вакуолью, о чем мы еще будем говорить в тл. 6. [c.72]
chem21.info









