Факторы, определяющие симбиотические взаимоотношения клубеньковых бактерий с бобовыми растениями. Клубеньковые бактерии и бобовые растения взаимоотношения
Урок№29. Лабораторная работа. Изучение взаимоотношений клубеньковых микроорганизмов и высших бобовых растений
Полная версия 46 (час) лабораторных работ с видеоматериалам
Тип урока – комплексное применение ЗУН учащихся
Метод обучения - основаны на самостоятельном проведении учащимися экспериментов, исследований и выдвижение гипотезы. Определение путей ее реализации, подбор необходимых приборов и материалов самими учащимися.
Формы организации деятельности учащихся
| Форма организации работы | Особенности, признаки | В каком случае выбираем ту или иную форму на теоретическом обучении | В каком случае выбираем ту или иную форму на практическому обучении |
| Парная | Когда преподаватель организует выполнение работы парами: сильный учащийся – слабый учащийся. Или два равных по успеваемости. | 1.В ходе актуализации опорных знаний, когда предстоящая работа требует серьезного предварительного осмысления. Пары учащихся обсуждают предстоящее задание. 2.В ходе лабораторно-практической работы возможна организация взаимоконтроля и взаимопомощи. Во время проведения итогов возможна организация взаимной оценки работ. | 1.В ходе вводного инструктажа, когда предстоящая практическая работа требует серьезного осмысления, пары учащихся обсуждают предстоящее задание. 2.В ходе практической работы возможна организация взаимоконтроля и взаимопомощи. 3.В ходе заключительного инструктажа возможна организация оценки работы |
ВЗАИМООТНОШЕНИЯ ОРГАНИЗМОВ В БИОЦЕНОЗАХ
Основу возникновения и существования любого биоценоза составляют отношения организмов, их связи, в которые они вступают друг с другом, населяя один и тот же биотоп.
Можно выделить следующие взаимоотношения
Конкуренция — взаимоотношения между организмами одного и того же вида (внутривидовая конкуренция) или разных видов (межвидовая конкуренция), соревнующимися за одни и те же ресурсы внешней среды, при недостатке последних.
Хищничество — способ добывания пищи и питания животных (редко растений), при котором они ловят, умертвляют и поедают других животных.
Паразитизм — форма взаимоотношений двух различных организмов, принадлежащих к разным видам, носящая антагонистический характер, когда один организм использует другой в качестве среды обитания или источника пищи.
Комменсализм — форма взаимосуществования, при которой один из партнеров системы питается остатками или продуктами выделения другого (хозяина), не причиняя последнему вреда.
Синойкия — форма симбиоза, разновидность комменсализма; совместное проживание двух организмов разных видов, полезное для одного и безразличное для другого
Мутуализм — форма симбиоза, взаимовыгодное сожительство разных видов.
Зоохория— форма межвидовых взаимоотношений, при которых животные содействуют растениям в распространении семян и плодов.
В.Н. Беклемишев классифицирует все межвидовые отношения живых организмов на следующие типы:
1.Трофические связи, которые возникают при питании одного вида другим (либо живыми организмами, либо мертвыми остатками).
2.Топические связи. Тип связи, при котором вид изменяет условия жизни другого.
3.Форические связи. Это участие одного вида в распространении другого. Перенос семян, спор, пыльцы растений называют зоохорией.
4.Фабрические связи. Это тип взаимоотношений, при котором вид использует для постройки своих сооружений другой вид, продукты его жизнедеятельности или мертвые остатки.
Цель работы: изучить тип симбиотических отношений между корневой системой высших бобовых растений и клубеньковыми бактериями.
Материалы и оборудование: фиксированные препараты корней различных видов бобовых; микроскопы; лупы; линейки; предметные и покровные стекла; метиловый голубой; препаровальные иглы; бритвы.
Ход работы.
Рассмотрите корневые системы трех видов бобовых, замерьте размеры, количество и форму клубеньков, данные занесите в таблицу. Сделайте фиксированные препараты микроорганизмов клубеньковых бактерий, зарисуйте.
Таблица 40
Особенности строения клубеньковых бактерий различных видов бобовых
| Вид бобового растения | Количество клубеньков на 1 дм2 корня | Форма клубеньков | Размеры клубеньков | Форма клеток микроорганизмов |
Контрольные вопросы
1.Какие типы биотических отношений в биоценозах вам известны?
2.Какое значение имеют биотические отношения в биоценозах?
3.Какой тип отношений вы изучили на примере бобовых? Что дает каждому из партнеров этот тип взаимоотношений?
Польза и вред бобовых
Семейство Бобовые
Кормовые бобовые растения
Семейство Мотыльковые Бобовые
Ресурсы:
ПРАКТИКУМ ПО ЭКОЛОГИИ Н.А. Иванова, Т. В. Сторчак, Э.Р. Юмагулова
(Учебно-методическое пособие)
Сайт YouTube: https://www.youtube.com /
Хостинг презентаций
- http://ppt4web.ru/nachalnaja-shkola/prezentacija-k-uroku-okruzhajushhego-mira-vo-klasse-chto-takoe-ehkonomika.html
xn--j1ahfl.xn--p1ai
Факторы, определяющие симбиотические взаимоотношения клубеньковых бактерий с бобовыми растениями
Страница 11 из 17
Факторы, определяющие симбиотические взаимоотношения клубеньковых бактерий с бобовыми растениями
Для симбиоза азотфиксаторов с растениями, обеспечивающего хорошее развитие последних, необходим определенный комплекс условий среды. Если условия окружающей среды будут неблагоприятными, то, даже, несмотря на высокую вирулентность, конкурентную способность и активность микросимбионта, эффективность симбиоза будет низкой. Для развития клубеньков оптимальна влажность 60-70% от полной влагоемкости почвы. Минимальная влажность почвы, при которой еще возможно развитие клубеньковых бактерий в почве, приблизительно равна 16% от полной влагоемкости. При влажности ниже этого предела клубеньковые бактерии обычно уже не размножаются, но, тем не менее, они не погибают и могут длительное время сохраняться в неактивном состоянии. Недостаток влаги приводит и к отмиранию уже сформировавшихся клубеньков. Нередко в районах с недостаточным увлажнением многие бобовые растения развиваются, не образуя клубеньков.
Большое влияние на жизнедеятельность клубеньковых бактерий и образование клубеньков оказывает реакция почвы. Для разных видов и даже штаммов клубеньковых бактерий значение рН среды обитания несколько различно. Так, например, клубеньковые бактерии клевера более устойчивы к низким значениям р.Н, чем клубеньковые бактерии люцерны. Очевидно, здесь также сказывается адаптация микроорганизмов к среде обитания. Клевер растет на более кислых почвах, чем люцерна. Реакция почвы как экологический фактор оказывает влияние на активность и вирулентность клубеньковых бактерий. Наиболее активные штаммы, как правило, легче выделить из почв с нейтральными значениями рН. В кислых почвах чаще встречаются неактивные и слабовирулентные штаммы. Кислая среда (рН 4,0 — 4,5) оказывает непосредственное влияние и на растения, в частности нарушая синтетические процессы обмена веществ растений и нормальное развитие корневых волосков. В кислой среде у инокулированных растений резко сокращается срок функционирования бактероидной ткани, что ведет к снижению степени азотфиксации.
Размеры симбиотической азотфиксации определяются в значительной степени условиями питания растения-хозяина, а не клубеньковых бактерий. Клубеньковые бактерии как эндотрофные симбионты растений зависят в основном от растения при получении углеродсодержащих веществ и минеральных элементов питания.
Для клубеньковых бактерий ткань хозяина представляет такую питательную среду, которая может удовлетворить даже самый требовательный штамм вследствие содержания в ткани всех типов питательных веществ. Тем не менее, после внедрения клубеньковых бактерий в ткань растения-хозяина их развитие определяется не только внутренними процессами, но и в значительной степени зависит от действия внешних факторов, оказывающих влияние на весь ход инфекционного процесса. Содержание или отсутствие того или иного питательного вещества в окружающей среде может быть определяющим моментом для проявления симбиотической азотфиксации.
Обеспеченность бобовых растений доступными формами минеральных соединений азота определяет эффективность симбиоза. На основании многочисленных лабораторных и вегетационных опытов известно, что с увеличением азотсодержащих соединений в окружающей среде, внедрение бактерий в корень растений затрудняется.
Большое значение в активации усвоения азота бобовыми растениями имеет фосфорное питание. При низком содержании фосфора в среде проникновение бактерий в корень происходит, но клубеньки при этом не образуются. Бобовым растениям присущи некоторые особенности в обмене фосфорсодержащих соединений. Семена бобовых отличаются повышенным содержанием фосфора. Запасной фосфор при прорастании семян используется не так, как у других культур, — сравнительно равномерно для формирования всех органов, а в большей степени сосредоточиваясь в корнях. Поэтому в ранние сроки развития бобовые растения, в отличие от злаковых, в большей степени удовлетворяют свои потребности в фосфоре за счет семядолей, а не запасов почвы. Чем крупнее семена, тем меньше бобовые растения зависят от фосфора почвы. Однако при симбиотическом способе существования потребность бобовых растений в фосфоре выше, чем при росте и развитии растений в среде без азотфиксаторов. Поэтому при недостатке фосфора в среде у инокулированных растений ухудшается снабжение растений азотом.
Также бобовые растения выносят с урожаем значительно больше калия, чем другие сельскохозяйственные культуры. Поэтому калийные и особенно фосфорно-калийные удобрения существенно повышают продуктивность азотфиксации бобовыми растениями. Положительное действие калия на образование клубеньков и интенсивность азотфиксации связано в значительной степени с физиологической ролью калия в углеводном обмене растения.
Кальций нужен не только для устранения излишней кислотности почвы. Он играет специфическую роль в развитии клубеньковых бактерий и обеспечении нормального симбиоза бактерий с растением-хозяином. Потребность клубеньковых бактерий в кальции частично может быть компенсирована стронцием. Интересно, что клубеньковые бактерии тропических культур, растущих на кислых латеритных почвах, не нуждаются в кальции. В этом опять проявляется экологическая адаптация клубеньковых бактерий, поскольку тропические почвы содержат очень небольшие количества кальция.
Для симбиотической азотфиксации необходимы также магний, сера и железо. При недостатке магния тормозится размножение клубеньковых бактерий, снижается их жизнедеятельность, подавляется симбиотическая азот-фиксация. Сера и железо оказывают также благоприятное влияние на образование клубеньков и процесс азотфиксации, в частности играя несомненную роль в синтезе леггемоглобина.
Из микроэлементов особо отмечена роль молибдена и бора. При недостатке молибдена клубеньки плохо образуются, в них нарушается синтез свободных аминокислот и подавляется синтез леггемоглобина. Молибден вместе с другими элементами с переменной валентностью (Fe, Co, Си) служит посредником при переносе электронов в окислительно-восстановительных ферментных реакциях. При дефиците бора в клубеньках не формируются сосудистые пучки, и вследствие этого нарушается развитие бактероидной ткани.
На формирование клубеньков у бобовых растений большое влияние оказывает углеводный обмен растений, определяемый рядом факторов: фотосинтезом, наличием в среде углекислого газа, физиологическими особенностями растений. Улучшение углеводного питания благоприятно сказывается на инокуляционном процессе и азотонакоплении. С практической точки зрения большой интерес представляет использование соломы и соломистого свежего навоза для удобрения бобовых растений как источника углеводов. Но в первый год после внесения соломы в почву при ее разложении накапливаются токсические вещества. Следует отметить, что не все виды бобовых растений чувствительны к токсическим продуктам распада соломы; горох, например, не реагирует на них.
Определенное значение в симбиозе клубеньковых бактерий и бобовых растений имеют биологические факторы. Большое внимание уделяется влиянию ризосферной микрофлоры на клубеньковые бактерии, которое может иметь как стимуляционный, так и антагонистический характер в зависимости от состава микроорганизмов ризосферы.
Много работ посвящено изучению фагов клубеньковых бактерий. Большинство фагов способны лизировать различные виды бактерий, некоторые специализированы лишь в отношении отдельных видов или даже штаммов клубеньковых бактерий. Фаги могут препятствовать внедрению бактерий в корень, вызывать лизис клеток в ткани клубенька, они наносят большой ущерб, лизируя препараты клубеньковых бактерий на заводах, вырабатывающих нитрагин.
Среди различных видов насекомых, наносящих вред клубеньковым бактериям, особенно выделяется полосатый клубеньковый долгоносик, личинки которого разрушают клубеньки на корнях многих видов бобовых растений (главным образом однолетних). Широко распространен и щетинистый клубеньковый долгоносик. Ранней весной самки клубеньковых долгоносиков откладывают от 10 до 100 яиц. Через 10-15 дней из яиц развиваются небольшие, до 5,5 мм, червеобразные, согнутые, белые, со светло-бурой головкой личинки, питающиеся преимущественно клубеньками и корневыми волосками. Только что вылупившиеся личинки проникают в клубенек и питаются его содержимым. Более взрослые личинки разрушают клубеньки снаружи. Одна личинка за 30-40 дней уничтожает от 2 до 6 клубеньков. Особенно большой вред они наносят в сухую и жаркую погоду, когда развитие растений замедляется.
Клубеньки люцерны и некоторых других видов бобовых растений повреждает также большой люцерновый долгоносик. Самки жука откладывают до 400 яиц, из которых развиваются безногие, дугообразные, желтовато-белые, с бурой головкой, покрытые бурыми щетинками личинки. Их длина 10 — 14 мм. Цикл развития большого люцернового долгоносика протекает в течение двух лет.
Наличие нематод в корневой зоне различных видов бобовых растений отмечают многие исследователи. В прикорневой зоне гороха, например, обнаружено 47 видов нематод, среди них 25 паразитических. На корнях молодых растений фасоли, люпина, клевера может паразитировать широко распространенная ростковая нематода. Самки этого вида, питающиеся корнями растений, откладывают яйца в ткани растения. Весь жизненный цикл развивающейся из яиц нематоды протекает обычно внутри тканей. В степных районах на корнях люцерны, клевера и сои обнаружена степная нематода. Самки перед откладкой яиц проникают в корень, куда откладывают от 12 до 20 яиц. В корнях личинки проходят три личиночные стадии развития, нарушая функции корня и клубеньков.
Распространение клубеньковых бактерий в природе
Являясь симбиотическими организмами, клубеньковые бактерии распространяются в почвах, сопутствуя определенным видам бобовых растений. После разрушения клубеньков клетки клубеньковых бактерий попадают в почву и переходят к существованию за счет различных органических веществ подобно другим почвенным микроорганизмам. Почти повсеместное распространение клубеньковых бактерий является доказательством высокой степени их приспособляемости к различным почвенно-климатическим условиям, способности вести симбиотический и сапрофитный способ жизни.
В целинных и окультуренных почвах присутствуют обычно в больших количествах клубеньковые бактерии тех видов бобовых растений, которые имеются в составе дикой флоры или культивируются длительное время в данной местности. Численность клубеньковых бактерий всегда наивысшая в ризосфере бобовых растений, несколько меньше их в ризосфере других видов и мало в почве вдали от корней.
В почвах встречаются как эффективные, так и неэффективные клубеньковые бактерии. Имеется много данных о том, что длительное сапрофитное существование клубеньковых бактерий, особенно в почвах с неблагоприятными свойствами (кислых, засоленных), ведет к снижению и даже утрате активности бактерий.
Перекрестная заражаемость разных видов бобовых растений нередко приводит в природе и сельскохозяйственной практике к появлению на корнях клубеньков, недостаточно активно фиксирующих молекулярный азот. Это, как правило, зависит от отсутствия в почве соответствующих видов клубеньковых бактерий. Особенно часто такое явление наблюдается при использовании новых видов бобовых растений, которые либо заражаются неэффективными видами бактерий перекрестных групп, либо развиваются без клубеньков.
selo-delo.ru
Факторы, определяющие симбиотические взаимоотношения клубеньковых бактерий с бобовыми растениями
Страница 11 из 17
Факторы, определяющие симбиотические взаимоотношения клубеньковых бактерий с бобовыми растениями
Для симбиоза азотфиксаторов с растениями, обеспечивающего хорошее развитие последних, необходим определенный комплекс условий среды. Если условия окружающей среды будут неблагоприятными, то, даже, несмотря на высокую вирулентность, конкурентную способность и активность микросимбионта, эффективность симбиоза будет низкой. Для развития клубеньков оптимальна влажность 60-70% от полной влагоемкости почвы. Минимальная влажность почвы, при которой еще возможно развитие клубеньковых бактерий в почве, приблизительно равна 16% от полной влагоемкости. При влажности ниже этого предела клубеньковые бактерии обычно уже не размножаются, но, тем не менее, они не погибают и могут длительное время сохраняться в неактивном состоянии. Недостаток влаги приводит и к отмиранию уже сформировавшихся клубеньков. Нередко в районах с недостаточным увлажнением многие бобовые растения развиваются, не образуя клубеньков.
Большое влияние на жизнедеятельность клубеньковых бактерий и образование клубеньков оказывает реакция почвы. Для разных видов и даже штаммов клубеньковых бактерий значение рН среды обитания несколько различно. Так, например, клубеньковые бактерии клевера более устойчивы к низким значениям р.Н, чем клубеньковые бактерии люцерны. Очевидно, здесь также сказывается адаптация микроорганизмов к среде обитания. Клевер растет на более кислых почвах, чем люцерна. Реакция почвы как экологический фактор оказывает влияние на активность и вирулентность клубеньковых бактерий. Наиболее активные штаммы, как правило, легче выделить из почв с нейтральными значениями рН. В кислых почвах чаще встречаются неактивные и слабовирулентные штаммы. Кислая среда (рН 4,0 — 4,5) оказывает непосредственное влияние и на растения, в частности нарушая синтетические процессы обмена веществ растений и нормальное развитие корневых волосков. В кислой среде у инокулированных растений резко сокращается срок функционирования бактероидной ткани, что ведет к снижению степени азотфиксации.
Размеры симбиотической азотфиксации определяются в значительной степени условиями питания растения-хозяина, а не клубеньковых бактерий. Клубеньковые бактерии как эндотрофные симбионты растений зависят в основном от растения при получении углеродсодержащих веществ и минеральных элементов питания.
Для клубеньковых бактерий ткань хозяина представляет такую питательную среду, которая может удовлетворить даже самый требовательный штамм вследствие содержания в ткани всех типов питательных веществ. Тем не менее, после внедрения клубеньковых бактерий в ткань растения-хозяина их развитие определяется не только внутренними процессами, но и в значительной степени зависит от действия внешних факторов, оказывающих влияние на весь ход инфекционного процесса. Содержание или отсутствие того или иного питательного вещества в окружающей среде может быть определяющим моментом для проявления симбиотической азотфиксации.
Обеспеченность бобовых растений доступными формами минеральных соединений азота определяет эффективность симбиоза. На основании многочисленных лабораторных и вегетационных опытов известно, что с увеличением азотсодержащих соединений в окружающей среде, внедрение бактерий в корень растений затрудняется.
Большое значение в активации усвоения азота бобовыми растениями имеет фосфорное питание. При низком содержании фосфора в среде проникновение бактерий в корень происходит, но клубеньки при этом не образуются. Бобовым растениям присущи некоторые особенности в обмене фосфорсодержащих соединений. Семена бобовых отличаются повышенным содержанием фосфора. Запасной фосфор при прорастании семян используется не так, как у других культур, — сравнительно равномерно для формирования всех органов, а в большей степени сосредоточиваясь в корнях. Поэтому в ранние сроки развития бобовые растения, в отличие от злаковых, в большей степени удовлетворяют свои потребности в фосфоре за счет семядолей, а не запасов почвы. Чем крупнее семена, тем меньше бобовые растения зависят от фосфора почвы. Однако при симбиотическом способе существования потребность бобовых растений в фосфоре выше, чем при росте и развитии растений в среде без азотфиксаторов. Поэтому при недостатке фосфора в среде у инокулированных растений ухудшается снабжение растений азотом.
Также бобовые растения выносят с урожаем значительно больше калия, чем другие сельскохозяйственные культуры. Поэтому калийные и особенно фосфорно-калийные удобрения существенно повышают продуктивность азотфиксации бобовыми растениями. Положительное действие калия на образование клубеньков и интенсивность азотфиксации связано в значительной степени с физиологической ролью калия в углеводном обмене растения.
Кальций нужен не только для устранения излишней кислотности почвы. Он играет специфическую роль в развитии клубеньковых бактерий и обеспечении нормального симбиоза бактерий с растением-хозяином. Потребность клубеньковых бактерий в кальции частично может быть компенсирована стронцием. Интересно, что клубеньковые бактерии тропических культур, растущих на кислых латеритных почвах, не нуждаются в кальции. В этом опять проявляется экологическая адаптация клубеньковых бактерий, поскольку тропические почвы содержат очень небольшие количества кальция.
Для симбиотической азотфиксации необходимы также магний, сера и железо. При недостатке магния тормозится размножение клубеньковых бактерий, снижается их жизнедеятельность, подавляется симбиотическая азот-фиксация. Сера и железо оказывают также благоприятное влияние на образование клубеньков и процесс азотфиксации, в частности играя несомненную роль в синтезе леггемоглобина.
Из микроэлементов особо отмечена роль молибдена и бора. При недостатке молибдена клубеньки плохо образуются, в них нарушается синтез свободных аминокислот и подавляется синтез леггемоглобина. Молибден вместе с другими элементами с переменной валентностью (Fe, Co, Си) служит посредником при переносе электронов в окислительно-восстановительных ферментных реакциях. При дефиците бора в клубеньках не формируются сосудистые пучки, и вследствие этого нарушается развитие бактероидной ткани.
На формирование клубеньков у бобовых растений большое влияние оказывает углеводный обмен растений, определяемый рядом факторов: фотосинтезом, наличием в среде углекислого газа, физиологическими особенностями растений. Улучшение углеводного питания благоприятно сказывается на инокуляционном процессе и азотонакоплении. С практической точки зрения большой интерес представляет использование соломы и соломистого свежего навоза для удобрения бобовых растений как источника углеводов. Но в первый год после внесения соломы в почву при ее разложении накапливаются токсические вещества. Следует отметить, что не все виды бобовых растений чувствительны к токсическим продуктам распада соломы; горох, например, не реагирует на них.
Определенное значение в симбиозе клубеньковых бактерий и бобовых растений имеют биологические факторы. Большое внимание уделяется влиянию ризосферной микрофлоры на клубеньковые бактерии, которое может иметь как стимуляционный, так и антагонистический характер в зависимости от состава микроорганизмов ризосферы.
Много работ посвящено изучению фагов клубеньковых бактерий. Большинство фагов способны лизировать различные виды бактерий, некоторые специализированы лишь в отношении отдельных видов или даже штаммов клубеньковых бактерий. Фаги могут препятствовать внедрению бактерий в корень, вызывать лизис клеток в ткани клубенька, они наносят большой ущерб, лизируя препараты клубеньковых бактерий на заводах, вырабатывающих нитрагин.
Среди различных видов насекомых, наносящих вред клубеньковым бактериям, особенно выделяется полосатый клубеньковый долгоносик, личинки которого разрушают клубеньки на корнях многих видов бобовых растений (главным образом однолетних). Широко распространен и щетинистый клубеньковый долгоносик. Ранней весной самки клубеньковых долгоносиков откладывают от 10 до 100 яиц. Через 10-15 дней из яиц развиваются небольшие, до 5,5 мм, червеобразные, согнутые, белые, со светло-бурой головкой личинки, питающиеся преимущественно клубеньками и корневыми волосками. Только что вылупившиеся личинки проникают в клубенек и питаются его содержимым. Более взрослые личинки разрушают клубеньки снаружи. Одна личинка за 30-40 дней уничтожает от 2 до 6 клубеньков. Особенно большой вред они наносят в сухую и жаркую погоду, когда развитие растений замедляется.
Клубеньки люцерны и некоторых других видов бобовых растений повреждает также большой люцерновый долгоносик. Самки жука откладывают до 400 яиц, из которых развиваются безногие, дугообразные, желтовато-белые, с бурой головкой, покрытые бурыми щетинками личинки. Их длина 10 — 14 мм. Цикл развития большого люцернового долгоносика протекает в течение двух лет.
Наличие нематод в корневой зоне различных видов бобовых растений отмечают многие исследователи. В прикорневой зоне гороха, например, обнаружено 47 видов нематод, среди них 25 паразитических. На корнях молодых растений фасоли, люпина, клевера может паразитировать широко распространенная ростковая нематода. Самки этого вида, питающиеся корнями растений, откладывают яйца в ткани растения. Весь жизненный цикл развивающейся из яиц нематоды протекает обычно внутри тканей. В степных районах на корнях люцерны, клевера и сои обнаружена степная нематода. Самки перед откладкой яиц проникают в корень, куда откладывают от 12 до 20 яиц. В корнях личинки проходят три личиночные стадии развития, нарушая функции корня и клубеньков.
Распространение клубеньковых бактерий в природе
Являясь симбиотическими организмами, клубеньковые бактерии распространяются в почвах, сопутствуя определенным видам бобовых растений. После разрушения клубеньков клетки клубеньковых бактерий попадают в почву и переходят к существованию за счет различных органических веществ подобно другим почвенным микроорганизмам. Почти повсеместное распространение клубеньковых бактерий является доказательством высокой степени их приспособляемости к различным почвенно-климатическим условиям, способности вести симбиотический и сапрофитный способ жизни.
В целинных и окультуренных почвах присутствуют обычно в больших количествах клубеньковые бактерии тех видов бобовых растений, которые имеются в составе дикой флоры или культивируются длительное время в данной местности. Численность клубеньковых бактерий всегда наивысшая в ризосфере бобовых растений, несколько меньше их в ризосфере других видов и мало в почве вдали от корней.
В почвах встречаются как эффективные, так и неэффективные клубеньковые бактерии. Имеется много данных о том, что длительное сапрофитное существование клубеньковых бактерий, особенно в почвах с неблагоприятными свойствами (кислых, засоленных), ведет к снижению и даже утрате активности бактерий.
Перекрестная заражаемость разных видов бобовых растений нередко приводит в природе и сельскохозяйственной практике к появлению на корнях клубеньков, недостаточно активно фиксирующих молекулярный азот. Это, как правило, зависит от отсутствия в почве соответствующих видов клубеньковых бактерий. Особенно часто такое явление наблюдается при использовании новых видов бобовых растений, которые либо заражаются неэффективными видами бактерий перекрестных групп, либо развиваются без клубеньков.
selo-delo.ru
Факторы, определяющие симбиотические взаимоотношения клубеньковых бактерий с бобовыми растениями
Для симбиоза, обеспечивающего хорошее развитие растений, необходим определенный комплекс условий среды. Если условия окружающей среды будут неблагоприятными, то, даже несмотря на высокую вирулентность, конкурентную способность и активность микросимбионта, эффективность симбиоза будет низкой.
Рис. 163. Начальный период формирования артроспор в бактероидах клевера. Увел. X 30 000.
Для развития клубеньков оптимальная влажность 60—70% от полной влагоемкости почвы. Минимальная влажность почвы, при которой еще возможно развитие клубеньковых бактерий в почве, приблизительно равна 16% от полной влагоемкости. При влажности ниже этого предела клубеньковые бактерии обычно уже не размножаются, но тем не менее они не погибают и могут длительное время сохраняться в неактивном состоянии. Недостаток влаги приводит и к отмиранию уже сформировавшихся клубеньков.
Нередко в районах с недостаточным увлажнением многие бобовые растения развиваются, не образуя клубеньков.
Поскольку размножение клубеньковых бактерий в отсутствие влаги не происходит, в случае засушливой весны инокулированные (искусственно зараженные) семена необходимо вносить глубже в почву. Например, в Австралии семена с нанесенными на них клубеньковыми бактериями глубоко заделывают в почву. Интересно, что клубеньковые бактерии почв засушливого климата более стойко переносят засуху, чем бактерии почв влажного климата. В этом проявляется их экологическая приспособленность.
Избыточная влажность, как и ее недостаток, также неблагоприятна для симбиоза — из-за снижения степени аэрации в зоне корней ухудшается снабжение корневой системы растения кислородом. Недостаточная аэрация отрицательно влияет и на живущие в почве клубеньковые бактерии, которые, как известно, лучше размножаются при доступе кислорода. Тем не менее высокая аэрация в зоне корней приводит к тому, что кислород начинают связывать восстановители молекулярного азота, снижая степень азотфиксации клубеньков.
Важную роль во взаимоотношениях клубеньковых бактерий и бобовых растений играет температурный фактор. Температурные характеристики разных видов бобовых растений различны. Также и разные штаммы клубеньковых бактерий имеют свои определенные температурные оптимумы развития и активной фиксации азота. Следует отметить, что оптимальные температуры развития бобовых растений, образования клубеньков и азотфиксации не совпадают. Так, в природных условиях образование клубеньков может наблюдаться при температурах несколько выше О °С, азот-фиксация при таких условиях практически не происходит. Возможно, лишь арктические симбиозирующие бобовые растения связывают азот при очень низких температурах. Обычно же этот процесс происходит лишь при 10 °С и выше. Максимальная азотфиксация ряда бобовых растений наблюдается при 20—25 °С. Температура выше 30 °С отрицательно влияет на процесс азотонакопления.
Экологическая адаптация к температурному фактору у клубеньковых бактерий значительно меньше, чем у многих типичных сапрофитных форм. По мнению Е.Н. Мишустина (1970), это объясняется тем, что естественной средой обитания клубеньковых бактерий являются ткани растений, где температурные условия регулируются растением-хозяином.
Большое влияние на жизнедеятельность клубеньковых бактерий и образование клубеньков оказывает реакция почвы. Для разных видов и даже штаммов клубеньковых бактерий значение рН среды обитания несколько различно. Так, например, клубеньковые бактерии клевера более устойчивы к низким значениям р.Н, чем клубеньковые бактерии люцерны. Очевидно, здесь также сказывается адаптация мпкро-ерганизмов к среде обитания. Клевер растет на более кислых почвах, чем люцерна. Реакция почвы как экологический фактор оказывает влияние на активность и вирулентность клубеньковых бактерий. Наиболее активные штаммы, как правило, легче выделить из почв с нейтральными значениями рН. В кислых почвах чаще встречаются неактивные и слабовирулентные штаммы. Кислая среда (рН 4,0 — 4,5) оказывает непосредственное влияние и на растения, в частности нарушая синтетические процессы обмена веществ растений и нормальное развитие корневых волосков. В кислой среде у инокулированных растений резко сокращается срок функционирования бакте-роидной ткани, что ведет к снижению степени азотфиксации.
В кислых почвах, как отмечает А. В. Петербургский, в почвенный раствор переходят соли алюминия и марганца, неблагоприятно действующие на развитие корневой системы растений и процесс азотоусвоения, а также снижается содержание усвояемых форм фосфора, кальция, молибдена и углекислоты. Неблагоприятную реакцию почвы лучше всего устраняет известкование.
Размеры симбиотической азотфиксации определяются в значительной степени условиями питания растения-хозяина, а не клубеньковых бактерий. Клубеньковые бактерии как эндотрофные симбионты растений зависят в основном от растения при получении углеродсодер-жащих веществ и минеральных элементов питания.
Рис. 164. Бактероид со сформировавшейся артроспорой клубеньковой бактерии клевера. Увел. X 35 000.
Для клубеньковых бактерий ткань хозяина представляет такую питательную среду, которая может удовлетворить даже самый требовательный штамм вследствие содержания в ткани всех типов питательных веществ. Тем не менее после внедрения клубеньковых бактерий в ткань растения-хозяина их развитие определяется не только внутренними процессами, но и в значительной степени зависит от действия внешних факторов, оказывающих влияние на весь ход инфекционного процесса. Содержание или отсутствие того или иного питательного вещества в окружающей среде может быть определяющим моментом для проявления симбиотической азотфиксации.
Степень обеспеченности бобовых растений доступными формами минеральных соединений азота определяет эффективность симбиоза. На основании многочисленных лабораторных и вегетационных опытов известно, что чем больше азотсодержащих соединений в окружающей среде, тем с большим трудом внедряются бактерии в корень.
Сельскохозяйственная практика требует однозначно решить задачу — целесообразнее удобрять бобовые культуры азотом или же правы те исследователи, которые утверждают, что минеральный азот подавляет симбиотическую азотфиксацию бобовых культур и поэтому экономически выгоднее такие растения азотом не удобрять. На кафедре агрономической и биологической химии Московской сельскохозяйственной академии им. К. А. Тимирязева были проведены опыты, результаты которых дали возможность получить картину поведения симбионтов в условиях вегетационных и полевых опытов при обеспеченности растений разными дозами азота в среде. Установлено, что повышение содержания растворимых азотсодержащих соединений в среде в полевых условиях при оптимальных условиях произрастания растений не препятствует их симбиозу с клубеньковыми бактериями. Снижение доли атмосферного азота, усваиваемого растениями при повышенной обеспеченности минеральным азотом, имеет только относительный характер. Абсолютное количество азота, усвоенного бактериями из атмосферы, практически не снижается, даже нередко увеличивается по сравнению с растениями, выращивающимися в присутствии клубеньковых бактерий, но без внесения в почву азота.
Рис. 165. Миграция артроспор в окружающую среду из клубеньков: 1 — клевера (увел. X 17 000), 2 — люцерны (увел. X 23 000).
Большое значение в активации усвоения азота бобовыми растениями имеет фосфорное питание. При низком содержании фосфора в среде проникновение бактерий в корень происходит, но клубеньки при этом не образуются. Бобовым растениям присущи некоторые особенности в обмене фосфорсодержащих соединений. Семена бобовых отличаются повышенным содержанием фосфора. Запасной фосфор при прорастании семян используется не так, как у других культур, — сравнительно равномерно для формирования всех органов, а в большей степени сосредоточиваясь в корнях. Поэтому в ранние сроки развития бобовые растения, в отличие от злаковых, в большей степени удовлетворяют свои потребности в фосфоре за счет семядолей, а не запасов почвы. Чем крупнее семена, тем меньше бобовые растения зависят от фосфора почвы. Однако при симбиотическом способе существования потребность бобовых растений в фосфоре выше, чем при автотроф-ном. Поэтому при недостатке фосфора в среде у инокулированных растений ухудшается снабжение растений азотом.
Бобовые растения, как известно, выносят с урожаем значительно больше калия, чем другие сельскохозяйственные культуры. Поэтому калийные и особенно фосфорно-калийные удобрения существенно повышают продуктивность азотфиксации бобовыми растениями.
Рис. 166. Прекратившие функционировать интерцеллюлярная (1) и интрацеллюлярная (2) инфекционные нити в клубеньках клевера. Увел. X 10 000.
Положительное действие калия на образование клубеньков и интенсивность азотфиксации связано в значительной степени с физиологической ролью калия в углеводном обмене растения.
Кальций нужен не только для устранения излишней кислотности почвы. Он играет специфическую роль в развитии клубеньковых бактерий и обеспечении нормального симбиоза бактерий с растением-хозяином. Потребность клубеньковых бактерий в кальции частично может быть компенсирована стронцием. Интересно, что клубеньковые бактерии тропических культур, растущих на кислых латеритных почвах, не нуждаются в кальции. В этом опять проявляется экологическая адаптация клубеньковых бактерий, поскольку тропические почвы содержат очень небольшие количества кальция.
Для симбиотической азотфиксации необходимы также магний, сера и железо. При недостатке магния тормозится размножение клубеньковых бактерий, снижается их жизнедеятельность, подавляется симбиотическая азот-фиксация. Сера и железо оказывают также благоприятное влияние на образование клубеньков и процесс азотфиксации, в частности играя несомненную роль в синтезе леггемоглобина.
Из микроэлементов особо отметим роль молибдена и бора. При недостатке молибдена клубеньки плохо образуются, в них нарушается синтез свободных аминокислот и подавляется синтез леггемоглобина. Молибден вместе с другими элементами с переменной валентностью (Fe, Co, Си) служит посредником при переносе электронов в окислительно-восстановительных ферментных реакциях. При дефиците бора в клубеньках не формируются сосудистые пучки, и вследствие этого нарушается развитие бактероидной ткани.
На формирование клубеньков у бобовых растений большое влияние оказывает углеводный обмен растений, определяемый рядом факторов: фотосинтезом, наличием в среде углекислого газа, физиологическими особенностями растений. Улучшение углеводного питания благоприятно сказывается на инокуляционном процессе и азотонакоплении. С практической точки зрения большой интерес представляет использование соломы и соломистого свежего навоза для удобрения бобовых растений как источника углеводов. Но в первый год после внесения соломы в почву при ее разложении накапливаются токсические вещества. Следует отметить, что не все виды бобовых растений чувствительны к токсическим продуктам распада соломы; горох, например, не реагирует на них.
Определенное значение в симбиозе клубеньковых бактерий и бобовых растений имеют биологические факторы.
Большое внимание уделяется влиянию ризо-сферной микрофлоры на клубеньковые бактерии, которое может иметь как стимуляционный, так и антагонистический характер в зависимости от состава микроорганизмов ризосферы.
Много работ посвящено изучению фагов клубеньковых бактерий. Большинство фагов способны лидировать различные виды бактерий, некоторые специализированы лишь в отношении отдельных видов или даже штаммов клубеньковых бактерий. Фаги могут препятствовать внедрению бактерий в корень, вызывать лизис клеток в ткани клубенька. Фаги наносят большой ущерб, лизируя препараты клубеньковых бактерий на заводах, вырабатывающих нитрагин.
Среди различных видов насекомых, наносящих вред клубеньковым бактериям, особенно выделяется полосатый клубеньковый долгоносик, личинки которого разрушают клубеньки на корнях многих видов бобовых растений (главным образом однолетних). Широко распространен и щетинистый клубеньковый долгоносик.
Ранней весной самки клубеньковых долгоносиков откладывают от 10 до 100 яиц. Через 10—15 дней из яиц развиваются небольшие (до 5,5 мм), червеобразные, согнутые, белые, со светло-бурой головкой личинки, питающиеся преимущественно клубеньками и корневыми волосками. Только что вылупившиеся личинки проникают в клубенек и питаются его содержимым. Более взрослые личинки разрушают клубеньки снаружи. Одна личинка за 30—40 дней уничтожает 2—6 клубеньков. Особенно большой вред они наносят в сухую и жаркую погоду, когда развитие растений замедляется.
Клубеньки люцерны и некоторых других видов бобовых растений повреждает также большой люцерновый долгоносик.
Самки жука откладывают до 400 яиц, из которых развиваются безногие, дугообразные, желтовато-белые, с бурой головкой, покрытые бурыми щетинками личинки. Их длина 10— 14 мм. Цикл развития большого люцернового долгоносика протекает в течение двух лет.
Наличие нематод в корневой зоне различных видов бобовых растений отмечают многие исследователи. В прикорневой зоне гороха, например, обнаружено 47 видов нематод, среди них 25 паразитических.
На корнях молодых растений фасоли, люпина, клевера может паразитировать широко распространенная ростковая нематода. Самки этого вида, питающиеся корнями растений, откладывают яйца в ткани растения. Весь жизненный цикл развивающейся из яиц нематоды протекает обычно внутри тканей.
В степных районах на корнях люцерны, клевера и сои обнаружена степная нематода. Самки перед откладкой яиц проникают в корень, куда откладывают от 12 до 20 яиц. В корнях личинки проходят три личиночные стадии развития, нарушая функции корня и клубеньков.
3-net.ru
Азотфиксирующие клубеньковые бактерии бобовых
Дополнение серии материалов о сидератах. В предыдущей статье разбирались самые интересные бобовые сидераты, секрет эффективности которых — азотфиксирующие клубеньковые бактерии.
Д.Н Прянишников: «Бобовые растения это миниатюрный завод по утилизации атмосферного азота работающий на дармовой солнечной энергии».
Зачем нужен азот?
Самым важным питательным веществом во всем растениеводстве является азот (N). Азот наиболее распространенный в атмосфере газ (78-79%), но его трудно найти в других местах нашей планеты. N частично растворен в дождевой воде, но его количество ничтожно — 15 кг на гектар в год, и при всем его изобилии в воздухе, атмосферный азот тяжело доступен. Чтобы азот сделать пригодным в биологических системах, необходимо разорвать тройную связь в его молекуле N2, а это чрезвычайно сложно проделать.

Азот трудно получить, но он является питательным веществом с высоким спросом. Без N не было бы никаких аминокислот и белков — строительных блоков жизни.
Кто такие ризобии?
К счастью, природа создала группу азотфиксирующих клубеньковых бактерий Ризобия (Rhizobium), которые решают эту проблему. Ризобии не единственные, кто ответственен за фиксацию азота, но они проделывают большую часть работы на сегодняшний день.
Бактерии Rhizobum имеют особые последовательности ДНК, кодирующие белок нитрогеназа. Нитрогеназа представляет собой фермент, разрывающий азотные связи и превращающий N в биологически доступную форму. Это занятие энергетически дорогое для бактерий. Но, благодаря партнерским отношениям с растением, ризобии получают углеводы, созданные растениями фотосинтезом из углекислого газа и солнечной энергии. Бактерии отдают азот взамен на сахара. Клубеньковые бактерии Ризобии проникают в клетки корневой системы растений из почвы и в процессе размножения создают огромные колонии, которые мы видим невооруженным взглядом в виде шариков на корнях бобовых.
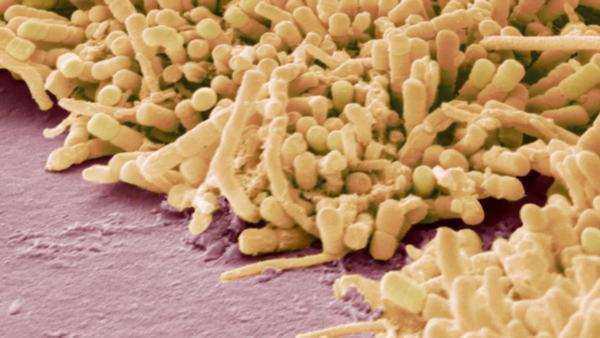
Ризобии
Только определенные растения имеют способность формировать отношения с азотфиксирующими клубеньковыми бактериями. Эта группа растений относится к ботанической семье бобовых. Хорошие примеры бобовых культур — клевер, люцерна, горох, фасоль, арахис.

Клубеньковые азотфиксирующие бактерии ризобии на корнях растений
Бобовые культуры полезно знать всем, кто хочет выращивать себе пищу. Без покупных удобрений не обойдешься, однако расходы можно существенно сократить, и это не единственное преимущество бобовых растений. В этой статье я не буду уделять внимание бобовым сидератам, с ними можно разобраться в этой статье. Упомяну лишь, что бобовые при заделке в почву оставляют в ней азот, легко доступный после разложения для других культур, посаженных на их месте.
Источник материала «Homestead and Gardens» , блог агронома из Айдахо, работающего на орехово-черносливной ферме.
Есть мнение, что для успешной фиксации азота необходимо искусственное подселение клубеньковых бактерий в почву с помощью специальных препаратов.
g.janecraft.net
Инфицирование бобового растения клубеньковыми бактериями
Страница 6 из 17
Инфицирование бобового растения клубеньковыми бактериями
Для обеспечения нормального процесса инфицирования корневой системы клубеньковыми бактериями нужно наличие довольно большого количества жизнеспособных клеток бактерий в прикорневой зоне. Мнения исследователей в отношении количества клеток, необходимых для обеспечения процесса инокуляции, различны. Так, по данным О. Аллена, для инокуляции мелкосеменных растений требуется 500-1000 клеток, для инокуляции крупносеменных — не менее 70 000 клеток на 1 семя. По мнению Дж. Винцента, в момент инокуляции на каждое семя должно приходиться по крайней мере несколько сотен жизнеспособных и активных клеток клубеньковых бактерий. Имеются данные, что в ткань корня могут внедряться и единичные клетки.
При развитии корневой системы бобового растения размножение клубеньковых бактерий на поверхности корня стимулируется выделениями корня. Продукты разрушения корневых чехликов и волосков играют также немаловажную роль в обеспечении клубеньковых бактерий подходящим субстратом. В ризосфере бобового растения резко стимулируется развитие клубеньковых бактерий, для злаковых растений такого явления не наблюдается.
На поверхности корня имеется слой слизистого вещества (матрица), образующийся независимо от наличия в ризосфере бактерий. Клубеньковые бактерии после инокуляции обычно устремляются к этому слою и скапливаются в нем благодаря стимулирующему действию корня, которое проявляется на расстоянии до 30 мм. В этот период, предшествующий внедрению клубеньковых бактерий в ткань корня, бактерии в ризосфере очень подвижны.
О механизме проникновения клубеньковых бактерий в корень растения существует ряд гипотез. Авторы одной из них утверждают, что клубеньковые бактерии проникают в корень через повреждения эпидермальной и коровой ткани (особенно в местах ответвления боковых корней). Эта гипотеза была выдвинута на основании исследований Бриля еще в 1888 году, вызвавшего образование клубеньков у бобовых растений путем прокалывания корней иглой, погруженной предварительно в суспензию клубеньковых бактерий. Некоторое подтверждение этому находят у арахиса, у которого клубеньки располагаются преимущественно в пазухах ответвлений корней, что наводит на мысль о проникновении клубеньковых бактерий в корень через разрывы при прорастании боковых корней.
Другая гипотеза обосновывает проникновение клубеньковых бактерий в ткань корня через корневые волоски рисунок 26. Путь прохождения клубеньковых бактерий через корневые волоски признает большинство исследователей.
Кажется убедительным предположение П. Дарта и Ф. Мерсера (1965) о том, что клубеньковые бактерии внедряются в корень в виде мелких (0,1-0,4 мкм) кокковидных клеток через промежутки (0,3-0,4 мкм) целлюлозной фибриллярной сети первичной оболочки корневых волосков. Электронно-микроскопические фотографии – рисунок 27 — поверхности корня, полученные методом реплик, и факт мельчания клеток клубеньковых бактерий в ризосфере бобовых растений подтверждают это положение.
Стрелкой показано движение клеток клубеньковых бактерий к вершине корневого волоска клевера.
Рисунок 26 — Скопления клубеньковых бактерий вокруг корневых волосков (по В. Шильниковой, 1974)
Рисунок 27 — Клетки клубеньковых бактерий на поверхности инфицированного корневого волоска бобового растения (по П. Дарту, Ф. Мерсеру).
Не исключено, что клубеньковые бактерии могут проникать в корень через эпидермальные клетки молодых верхушек корня. По мнению Пражмовского (1889), бактерии могут проникать в корень только через молодую клеточную оболочку (корневых волосков или эпидермальных клеток) и совершенно не способны преодолевать химически измененный или опробковевший слой коры. Этим можно объяснить, что клубеньки обычно развиваются на молодых участках главного корня и появляющихся боковых корнях.
В последнее время большую популярность получила ауксинная гипотеза. Авторы этой гипотезы считают, что клубеньковые бактерии проникают в корень благодаря стимуляции синтеза гетероауксина из триптофана, имеющегося всегда в корневых выделениях растений. С наличием гетероауксина связывается искривление корневых волосков, которое обычно наблюдается при инфицировании корневой системы клубеньковыми бактериями.
Источником бета-индолилуксусной кислоты в момент инфицирования растения, очевидно, служат не только растения, выделяющие через корневую систему триптофан, который многие виды бактерий, в том числе и клубеньковые, могут переводить в бета-индолилуксусную кислоту. Сами клубеньковые бактерии, а возможно, и другие виды почвенных микроорганизмов, живущие в зоне корня, также могут участвовать в синтезе гетероауксина. Однако действие гетероауксина неспецифично и вызывает искривление корневых волосков у разных видов растений, а не только бобовых. В то же время клубеньковые бактерии вызывают искривление корневых волосков лишь у бобовых растений, проявляя при этом довольно значительную избирательность. Если бы рассматриваемый эффект определялся только (З-индолилуксусной кислотой), то такой специфики не было бы. Кроме того, характер изменений корневых волосков под влиянием клубеньковых бактерий несколько иной, чем под влиянием гетероауксина.
Также отмечают, что в отдельных случаях инфицированию подвергаются не искривленные корневые волоски. Наблюдения показывают, что у люцерны и гороха искривляются и закручиваются 60-70% корневых волосков, а у клевера — около 50%. У некоторых видов клевера эта реакция отмечается не более чем у четверти заражаемых волосков. В реакции искривления, очевидно, имеет большое значение состояние корневого волоска. Растущие корневые волоски наиболее чувствительны к действию веществ, вырабатываемых бактериями.
Известно, что клубеньковые бактерии вызывают размягчение стенок корневых волосков. Однако ни целлюлазы, ни пектинолитических ферментов они не образуют. В связи с этим было высказано предположение, что клубеньковые бактерии проникают в корень благодаря выделению ими слизи полисахаридной природы, вызывающей синтез растениями фермента полигалактуроназы. Этот фермент, разрушая пектиновые вещества, влияет на оболочку корневых волосков, делая ее более пластичной и проницаемой. В небольших количествах полигалактуроназа всегда присутствует в корневых волосках и, очевидно, вызывая частичное растворение соответствующих компонентов оболочки, позволяет клетке растягиваться.
Некоторые исследователи полагают, что клубеньковые бактерии проникают в корень благодаря бактериям-спутникам, продуцирующим пектинолитические ферменты. Эта гипотеза была выдвинута на основании следующих фактов. При микроскопировании корневых волосков многие исследователи отмечали наличие светлого пятна, около которого скапливаются клубеньковые бактерии. Это пятно, возможно, является признаком начала мацерации (разрушения) ткани протопектиназой по аналогии с таким же признаком, наблюдающимся у растений при многих бактериальных заболеваниях. Кроме того, установлено, что авирулентные культуры клубеньковых бактерий в присутствии бактерий, продуцирующих пектинолитические ферменты, становятся способными проникать в корень.
Следует отметить еще одну гипотезу, по которой клубеньковые бактерии попадают в корень при образовании пальцевидного впячивания поверхности корневого волоска. Гипотеза инвагинации, по существу, не может быть отделена от ауксинной или ферментативной гипотезы, поскольку инвагинация происходит в результате воздействия либо ауксинного, либо ферментного фактора.
Процесс внедрения клубеньковых бактерий в ткань корня одинаков у всех видов бобовых растений и состоит из двух фаз. В первую фазу происходит инфицирование корневых волосков. Во вторую фазу интенсивно идет процесс образования клубеньков. Продолжительность фаз различна у разных видов растений: у Trifolium fragiferum первая фаза продолжается 6 дней, у Trifolium nigrescens — 3 дня. В некоторых случаях очень трудно обнаружить границы между фазами. Наиболее интенсивное внедрение клубеньковых бактерий в корневые волоски происходит на ранних этапах развития растения. Вторая фаза заканчивается в период массового образования клубеньков. Нередко внедрение клубеньковых бактерий в корневые волоски продолжается уже и после того, как клубеньки сформировались на корнях. Эта так называемая избыточная или дополнительная инфекция происходит потому, что инфицирование волосков не прекращается длительное время. Тип развития, структура и плотность корневых волосков не влияют на скорость внедрения клубеньковых бактерий. Места образования клубеньков не всегда связаны с местами расположения инфицированных волосков. Проникнув в корень (через корневой волосок, эпидермальную клетку, места повреждений корня),
клубеньковые бактерии далее перемещаются в ткани корня растения. Наиболее легко бактерии проходят через межклеточные пространства.
Внедриться в ткань корня может или одиночная клетка, или группа клеток бактерий. Если внедрилась отдельная клетка, она и в дальнейшем может перемещаться по ткани как одиночка. Путь инфицирования корня одиночными клетками свойствен растениям люпина. Однако в большинстве случаев внедрившаяся клетка, активно размножаясь, образует так называемые инфекционные нити и уже в виде таких нитей перемещается в ткани растения.
Термин «инфекционная нить» возник на основе изучения процесса инфицирования в световом микроскопе. Начиная с работ М. Бейеринка, инфекционная нить стала рассматриваться как слизистая гифообразная масса с заключенными в нее размножающимися бактериями. По существу, инфекционная нить — это колония размножившихся бактерий. Началом ее служит то место, куда проникла отдельная клетка или группа клеток. Не исключено, что колония бактерий, а следовательно, и будущая инфекционная нить, начинает формироваться еще на поверхности корня до момента внедрения бактерий в корень.
Количество инфицированных корневых волосков значительно различается у отдельных растений. Обычно инфекционные нити появляются в деформированных, искривленных корневых волосках. Однако есть указания, что и в прямых волосках иногда обнаруживаются подобные нити. Чаще в корневых волосках наблюдается одна разветвляющаяся нить, реже две. В некоторых случаях в одном корневом волоске имеется несколько нитей или же в нескольких имеются общие нити заражения, дающие начало одному клубеньку
Процент инфицированных корневых волосков в общем количестве деформированных очень низок. Он обычно колеблется от 0,6 до 3,2, изредка достигая 8,0. Доля удачных инфекций еще ниже, поскольку среди инфекционных нитей имеется много, до 80%, так называемых абортивных нитей, прекративших свое развитие. Скорость продвижения нормально развивающихся инфекционных нитей в растении составляет 5-8 мкм в час. При такой скорости путь через корневой волосок длиной 100-200 мкм инфекционная нить может пройти в течение одних суток.
selo-delo.ru
Инфицирование бобового растения клубеньковыми бактериями
Страница 6 из 17
Инфицирование бобового растения клубеньковыми бактериями
Для обеспечения нормального процесса инфицирования корневой системы клубеньковыми бактериями нужно наличие довольно большого количества жизнеспособных клеток бактерий в прикорневой зоне. Мнения исследователей в отношении количества клеток, необходимых для обеспечения процесса инокуляции, различны. Так, по данным О. Аллена, для инокуляции мелкосеменных растений требуется 500-1000 клеток, для инокуляции крупносеменных — не менее 70 000 клеток на 1 семя. По мнению Дж. Винцента, в момент инокуляции на каждое семя должно приходиться по крайней мере несколько сотен жизнеспособных и активных клеток клубеньковых бактерий. Имеются данные, что в ткань корня могут внедряться и единичные клетки.
При развитии корневой системы бобового растения размножение клубеньковых бактерий на поверхности корня стимулируется выделениями корня. Продукты разрушения корневых чехликов и волосков играют также немаловажную роль в обеспечении клубеньковых бактерий подходящим субстратом. В ризосфере бобового растения резко стимулируется развитие клубеньковых бактерий, для злаковых растений такого явления не наблюдается.
На поверхности корня имеется слой слизистого вещества (матрица), образующийся независимо от наличия в ризосфере бактерий. Клубеньковые бактерии после инокуляции обычно устремляются к этому слою и скапливаются в нем благодаря стимулирующему действию корня, которое проявляется на расстоянии до 30 мм. В этот период, предшествующий внедрению клубеньковых бактерий в ткань корня, бактерии в ризосфере очень подвижны.
О механизме проникновения клубеньковых бактерий в корень растения существует ряд гипотез. Авторы одной из них утверждают, что клубеньковые бактерии проникают в корень через повреждения эпидермальной и коровой ткани (особенно в местах ответвления боковых корней). Эта гипотеза была выдвинута на основании исследований Бриля еще в 1888 году, вызвавшего образование клубеньков у бобовых растений путем прокалывания корней иглой, погруженной предварительно в суспензию клубеньковых бактерий. Некоторое подтверждение этому находят у арахиса, у которого клубеньки располагаются преимущественно в пазухах ответвлений корней, что наводит на мысль о проникновении клубеньковых бактерий в корень через разрывы при прорастании боковых корней.
Другая гипотеза обосновывает проникновение клубеньковых бактерий в ткань корня через корневые волоски рисунок 26. Путь прохождения клубеньковых бактерий через корневые волоски признает большинство исследователей.
Кажется убедительным предположение П. Дарта и Ф. Мерсера (1965) о том, что клубеньковые бактерии внедряются в корень в виде мелких (0,1-0,4 мкм) кокковидных клеток через промежутки (0,3-0,4 мкм) целлюлозной фибриллярной сети первичной оболочки корневых волосков. Электронно-микроскопические фотографии – рисунок 27 — поверхности корня, полученные методом реплик, и факт мельчания клеток клубеньковых бактерий в ризосфере бобовых растений подтверждают это положение.
Стрелкой показано движение клеток клубеньковых бактерий к вершине корневого волоска клевера.
Рисунок 26 — Скопления клубеньковых бактерий вокруг корневых волосков (по В. Шильниковой, 1974)
Рисунок 27 — Клетки клубеньковых бактерий на поверхности инфицированного корневого волоска бобового растения (по П. Дарту, Ф. Мерсеру).
Не исключено, что клубеньковые бактерии могут проникать в корень через эпидермальные клетки молодых верхушек корня. По мнению Пражмовского (1889), бактерии могут проникать в корень только через молодую клеточную оболочку (корневых волосков или эпидермальных клеток) и совершенно не способны преодолевать химически измененный или опробковевший слой коры. Этим можно объяснить, что клубеньки обычно развиваются на молодых участках главного корня и появляющихся боковых корнях.
В последнее время большую популярность получила ауксинная гипотеза. Авторы этой гипотезы считают, что клубеньковые бактерии проникают в корень благодаря стимуляции синтеза гетероауксина из триптофана, имеющегося всегда в корневых выделениях растений. С наличием гетероауксина связывается искривление корневых волосков, которое обычно наблюдается при инфицировании корневой системы клубеньковыми бактериями.
Источником бета-индолилуксусной кислоты в момент инфицирования растения, очевидно, служат не только растения, выделяющие через корневую систему триптофан, который многие виды бактерий, в том числе и клубеньковые, могут переводить в бета-индолилуксусную кислоту. Сами клубеньковые бактерии, а возможно, и другие виды почвенных микроорганизмов, живущие в зоне корня, также могут участвовать в синтезе гетероауксина. Однако действие гетероауксина неспецифично и вызывает искривление корневых волосков у разных видов растений, а не только бобовых. В то же время клубеньковые бактерии вызывают искривление корневых волосков лишь у бобовых растений, проявляя при этом довольно значительную избирательность. Если бы рассматриваемый эффект определялся только (З-индолилуксусной кислотой), то такой специфики не было бы. Кроме того, характер изменений корневых волосков под влиянием клубеньковых бактерий несколько иной, чем под влиянием гетероауксина.
Также отмечают, что в отдельных случаях инфицированию подвергаются не искривленные корневые волоски. Наблюдения показывают, что у люцерны и гороха искривляются и закручиваются 60-70% корневых волосков, а у клевера — около 50%. У некоторых видов клевера эта реакция отмечается не более чем у четверти заражаемых волосков. В реакции искривления, очевидно, имеет большое значение состояние корневого волоска. Растущие корневые волоски наиболее чувствительны к действию веществ, вырабатываемых бактериями.
Известно, что клубеньковые бактерии вызывают размягчение стенок корневых волосков. Однако ни целлюлазы, ни пектинолитических ферментов они не образуют. В связи с этим было высказано предположение, что клубеньковые бактерии проникают в корень благодаря выделению ими слизи полисахаридной природы, вызывающей синтез растениями фермента полигалактуроназы. Этот фермент, разрушая пектиновые вещества, влияет на оболочку корневых волосков, делая ее более пластичной и проницаемой. В небольших количествах полигалактуроназа всегда присутствует в корневых волосках и, очевидно, вызывая частичное растворение соответствующих компонентов оболочки, позволяет клетке растягиваться.
Некоторые исследователи полагают, что клубеньковые бактерии проникают в корень благодаря бактериям-спутникам, продуцирующим пектинолитические ферменты. Эта гипотеза была выдвинута на основании следующих фактов. При микроскопировании корневых волосков многие исследователи отмечали наличие светлого пятна, около которого скапливаются клубеньковые бактерии. Это пятно, возможно, является признаком начала мацерации (разрушения) ткани протопектиназой по аналогии с таким же признаком, наблюдающимся у растений при многих бактериальных заболеваниях. Кроме того, установлено, что авирулентные культуры клубеньковых бактерий в присутствии бактерий, продуцирующих пектинолитические ферменты, становятся способными проникать в корень.
Следует отметить еще одну гипотезу, по которой клубеньковые бактерии попадают в корень при образовании пальцевидного впячивания поверхности корневого волоска. Гипотеза инвагинации, по существу, не может быть отделена от ауксинной или ферментативной гипотезы, поскольку инвагинация происходит в результате воздействия либо ауксинного, либо ферментного фактора.
Процесс внедрения клубеньковых бактерий в ткань корня одинаков у всех видов бобовых растений и состоит из двух фаз. В первую фазу происходит инфицирование корневых волосков. Во вторую фазу интенсивно идет процесс образования клубеньков. Продолжительность фаз различна у разных видов растений: у Trifolium fragiferum первая фаза продолжается 6 дней, у Trifolium nigrescens — 3 дня. В некоторых случаях очень трудно обнаружить границы между фазами. Наиболее интенсивное внедрение клубеньковых бактерий в корневые волоски происходит на ранних этапах развития растения. Вторая фаза заканчивается в период массового образования клубеньков. Нередко внедрение клубеньковых бактерий в корневые волоски продолжается уже и после того, как клубеньки сформировались на корнях. Эта так называемая избыточная или дополнительная инфекция происходит потому, что инфицирование волосков не прекращается длительное время. Тип развития, структура и плотность корневых волосков не влияют на скорость внедрения клубеньковых бактерий. Места образования клубеньков не всегда связаны с местами расположения инфицированных волосков. Проникнув в корень (через корневой волосок, эпидермальную клетку, места повреждений корня),
клубеньковые бактерии далее перемещаются в ткани корня растения. Наиболее легко бактерии проходят через межклеточные пространства.
Внедриться в ткань корня может или одиночная клетка, или группа клеток бактерий. Если внедрилась отдельная клетка, она и в дальнейшем может перемещаться по ткани как одиночка. Путь инфицирования корня одиночными клетками свойствен растениям люпина. Однако в большинстве случаев внедрившаяся клетка, активно размножаясь, образует так называемые инфекционные нити и уже в виде таких нитей перемещается в ткани растения.
Термин «инфекционная нить» возник на основе изучения процесса инфицирования в световом микроскопе. Начиная с работ М. Бейеринка, инфекционная нить стала рассматриваться как слизистая гифообразная масса с заключенными в нее размножающимися бактериями. По существу, инфекционная нить — это колония размножившихся бактерий. Началом ее служит то место, куда проникла отдельная клетка или группа клеток. Не исключено, что колония бактерий, а следовательно, и будущая инфекционная нить, начинает формироваться еще на поверхности корня до момента внедрения бактерий в корень.
Количество инфицированных корневых волосков значительно различается у отдельных растений. Обычно инфекционные нити появляются в деформированных, искривленных корневых волосках. Однако есть указания, что и в прямых волосках иногда обнаруживаются подобные нити. Чаще в корневых волосках наблюдается одна разветвляющаяся нить, реже две. В некоторых случаях в одном корневом волоске имеется несколько нитей или же в нескольких имеются общие нити заражения, дающие начало одному клубеньку
Процент инфицированных корневых волосков в общем количестве деформированных очень низок. Он обычно колеблется от 0,6 до 3,2, изредка достигая 8,0. Доля удачных инфекций еще ниже, поскольку среди инфекционных нитей имеется много, до 80%, так называемых абортивных нитей, прекративших свое развитие. Скорость продвижения нормально развивающихся инфекционных нитей в растении составляет 5-8 мкм в час. При такой скорости путь через корневой волосок длиной 100-200 мкм инфекционная нить может пройти в течение одних суток.
selo-delo.ru