
Экофизиологическая роль фотопериодизма у растений. Какой процесс обусловлен фотопериодической чувствительностью растений
Фотопериод и фотопериодические реакции растений
Фотопериодическая реакция растений. Важнейшей характеристикой светового режима местообитания является фотопериод — продолжительность светлого времени суток, обусловленная вращением Земли вокруг своей оси. Он неодинаков в течение года и на разных широтах. Чем ближе к полюсу, тем длиннее там день в летнее время (рис. 6.33). В низких широтах фотопериод в течение года меняется мало, но при удалении от тропиков соотношение светлого и темного периодов суток все больше различается. В средних широтах зимой день вдвое короче, чем летом, а за Полярным кругом наступает долгая полярная ночь зимой после долгого полярного дня летом. Ближе к полюсам интенсивность света ослабляется, но освещенность становится продолжительнее и преобладает длинноволновый рассеянный свет. При движении к экватору день укорачивается, но интенсивность света становится выше, а он богаче коротковолновым излучением. Таким образом, в высоких широтах растения растут в условиях длинного дня, а в низких — короткого дня.
Растения приспособились к жизни при чередовании определенной продолжительности дня и ночи. Способность организмов реагировать на суточный ритм освещения (соотношение светлого
Рис. 6.33. Сезонное изменение длины дня на разных широтах Северного полушария (по А. С. Данилевскому, 1961)
и темного периодов) называется фотопериодической реакцией. Выражается она в изменении процессов роста и развития при изменении длины дня и обеспечивает адаптацию онтогенеза растений к сезонным особенностям климата. Длина дня используется растением как часы, показывающие лучшее время для перехода к цветению или для подготовки к неблагоприятному сезону.
Основным критерием фотопериодической реакции считается стимулирование некоторой продолжительностью дня перехода к генеративной фазе развития. Иногда достаточно всего нескольких суток с нужным соотношением света и темноты, чтобы растение зацвело (фотопериодическая индукция). Так, дурнишник (Xanthium strumarium) не цветет целое лето при неподходяще длинном дне, но зацветает уже после одного короткого дня. Оказалось, что растения способны воспринимать продолжительность дня с большой точностью. Так, для хризантемы (Chrysanthemum) критическая длина дня, обеспечивающая цветение, составляет 14 ч 40 мин, но уже при 13 ч 50 мин бутоны не образуются. А у дурнишника пенсильванского (X. pennsylvanicum) критическая длина дня лежит между 15 ч и 15 ч 30 мин (Т.К.Горышина, 1979).
Для растений важен также характер изменения дня (укорачивающийся или удлиняющийся). Так, у ряда многолетних злаков переход к цветению стимулируется удлинением дня весной, а сокращающийся день той же продолжительности в конце лета этого эффекта не вызывает. 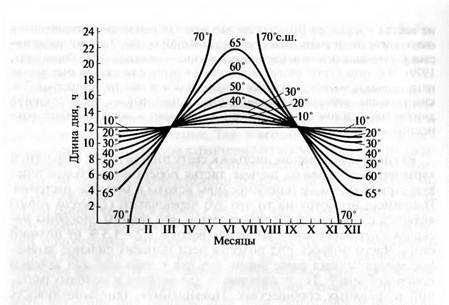
Фотопериодическая индукция обнаруживается при намного более слабой интенсивности света, чем фотосинтез. Например, дурнишник реагирует на длину дня при освещении 100-ваттной
лампой на расстоянии 150 м. При такой чувствительности растений к слабому свету при определении критической длины дня требуется учитывать также светлую часть сумерек.
eco-rasteniya.ru
Фотопериодизм | Info-Farm.RU
Фотопериодизм — физиологическая реакция организмов на суточный ритм освещения (соотношение длины дня и ночи). Встречается у растений и животных. Оказывается в колебаниях интенсивности физиологических процессов. В наибольшей степени фотопериодизм свойственный зеленым растениям, жизнедеятельность которых напрямую зависит от световой энергии Солнца.
Фотопериодизм открыто в 1920 г.. Американскими учеными В. Гарнером и Г. Аллард на примере табака.
У растений
Фотопериодизм у растений проявляется в изменении процессов роста и развития. Один из основных проявлений этой реакции — фотопериодических индукция цветения.
По типам ФПР различают следующие основные группы растений (Т. К. Горишина, 1980):
- Растения короткого дня, которым для перехода к цветению нужно света 12 и менее часов в сутки (конопля, табак)
- Растения длинного дня, для фазы цветения им нужна длина дня более 12:00 в сутки (картофель, пшеница, шпинат)
- Растения промежуточного типа, цветение в которых наступает при определенном диапазоне фотопериодизма, например гваюла;
- Растения фотопериодически нейтральные, цветение в которых происходит при любой длине дня (помидор, одуванчик и т.д.).
Особенно большую роль фотопериодизм играет в географическом распространении растений и в регуляции их сезонного ритма.
Фотопериодизм изучали американские физиологи растений В. Гарнер, Г. Аллард в 1920-1925 гг. В зависимости от реакции растений на длину дня ускоряет зацветания, они подразделяются на:
- длиннодневные (ДДР — растения длинного дня — злаки, крестоцветные, укроп; распространены в основном в умеренных и приполярных широтах)
- короткодневные (КДР — рис, соя, конопля; субтропики)
- довгокороткоденни (ДКДР) и
- короткодовгоденни (КДДР) — требуют чередование различных фотопериодов;
- нейтральные (ГДР — гречка, горох и др.).
Разделение растений на данные группы не связан с конкретной оптимальной длиной дня, а только дает представление о том, ускоряется цветения при увеличении или уменьшении продолжительности освещения в каждом фотопериоде.
Есть растения с качественными и количественными типами фотопериодических реакций. Интенсивность освещения, температура и другие факторы могут изменять характер фотопериодических проявлений растений.
Для КДР с качественной реакцией основным фактором служит продолжительность темнового периода. Его прерывания (хотя на 1 мин.) Препятствует переходу к цветению. Прерывание светового периода темнотой не влияет на сроки цветения. ДДР не требуют периода темноты и зацветают при непрерывном освещении.
Фотопериодических действие воспринимается главным образом листьями, а не апекс побегов. Максимальную чувствительность к фотопериоду имеют листья, которые только что закончили рост. Осуществляется это главным образом благодаря фитохрома. Считают, что его Темнов преобразования может служить способом измерения времени по типу песочных часов. Вспышка красного света устанавливает фазу эндогенного ритма.
Необходимость большой поверхности листьев и достаточной интенсивности света для перехода к цветению у многих растений объясняется потребностью растущих меристем в ассимиляты. В ГДР переход к цветению обеспечивается возрастными изменениями. Здесь срабатывает эндогенная регуляция. Данные факторы (температура и фотопериод) могут действовать последовательно, как, например, у озимых злаков.
Температурная и фотопериодических регуляция служат приспособлением растений к условиям существования, потому что обусловливают благоприятные сроки для перехода к цветению. В ходе фотопериодических индукции в листьях образуется стимулятор цветения, который транспортируется в вегетативные почки побегов, где включает вторую фазу инициации — эвокация.
У животных
У животных и человека фотопериодизм выражается главным образом в колебаниях интенсивности обмена веществ и энергии. У животных с фотопериодизмом связаны также сроки наступления и прекращения брачного периода, плодовитость, осенние и весенние лень, переход к зимовке, миграции и др. По отношению к свету животные делятся на группы в зависимости от того, какой образ жизни они ведут: дневные, ночные станков.
Видео по теме
info-farm.ru
Роль света в процессах роста и развития растений. Фотоморфогенез. Явление фотопериодизма. Регуляция фотопериодических реакций.
⇐ ПредыдущаяСтр 42 из 52Следующая ⇒На разные стороны ростовых процессов влияние света проявляется неоднозначно. Так, свет необходим для протекания процесса фотосинтеза, и поэтому накопление массы растения без света не идет. Вместе с тем рост клеток растяжением может идти в темноте, а на свету этот процесс тормозится. Свет оказывает большое влияние и на формообразовательные процессы.
В отсутствие света происходит упрощение анатомической структуры стебля. Слабо развиваются ткани центрального цилиндра, механические ткани. Вместе с тем растяжение клеток в темноте идет очень интенсивно. В результате образуются длинные, вытянутые стебли. Листья редуцированы, у двудольных растений вместо листовой пластинки образуются лишь небольшие чешуйки. Этиолированные проростки имеют слегка желтоватый оттенок.
Возможно, что вытягивание стебля в отсутствие света является следствием отсутствия ингибиторов роста. В темноте образуется много ауксинов. Нарушение соотношения ауксинов и ингибиторов вызывает несбалансированный рост. При выходе проростков на поверхность почвы происходят их внутренние и внешние изменения ( изгиб гипокотиля выпрямляется ).
На свету рост стебля тормозится, рост листьев усиливается, и они принимают обычную форму. Под влиянием света происходят анатомические изменения стебля, дифференцируется эпидермис, появляются волоски, изменяется окраска — синтезируется хлорофилл. Эти изменения получили название фотоморфогенеза.
Особое значение имеют пигментыфитохромы, криптохромыи фототропины, потому как они способны поглощать красный и синий свет, и у них нет прямой связи с фотосинтезом.
Растения, выросшие в полной темноте, называются этиолированными. Они бледно-жёлтые, сильно вытянутые, у них недоразвиты механические ткани, листовые пластинки, устьица и хлоропласты. Освещение этиолированных растений приводит к снижению скорости роста стебля, инициации формирования хлоропластов и синтеза фотосинтетических ферментов.
Наиболее эффективное регуляторное действие на рост и морфогенез имеет красный свет с длиной волны 650-680 нм. Свет действует как сигнал, который индуцирует переключение программы этиоляции на программу роста клеток над почвой на свету. Помимо этого, красный свет влияет на прорастание семян, «избегание тени», закладку листовых примордиев и развитие листа, закладку цветочных почек и цветение.
В основе многих реакций морфогенеза лежат обратимые превращения фитохрома; он состоит из двух субъединиц, а каждая субъединица из двух компонентов: светопоглощающей хромофорной группы (тетрапирольная структура; фитохромобилин) и полипептида (апопротеин), соединённые м/у собой тиоэфирной связью.
Фитохром существует в двух формах:
ФК (Ф660) – неактивная форма, голубого цвета, более устойчив. Поглощая красный свет (650-680 нм), переходит в Фдк.
ФДК (Ф730) – активная форма, зелено-желтого цвета, восстановленная форма Фк. При облучении дальним красным светом (710-740 нм), переходит в Фк. Активирует гены, ферменты, увеличивает проницаемость мембран, участвует в регуляции: дифференцировки тканей и органов, разных ростковых реакций, движений хлоропластов, в фотопериодич.реакциях.
Основная роль фотопревращенийфитохрома заключается в том, что таким образом растение получает информацию об изменении световых условий в окружающей среде и приспосабливается к ним. Фитохром является индикатором затененности и продвижения проростка на свет.
В регуляции роста синим светом участвуют криптохромы и фототропины.
Криптохромы состоят из апопротеинаи двух хромофоров (птерин и флавин). Спектр поглощения: сине-фиолетовая (390-500 нм) и ультрафиолетовая часть спектра (320-390 нм), а также захватывает и зелёную часть. Широта области поглощения зависит от птерина.
Важные функции синего света: регуляция ширины устьичной щели, распрямление верхушки побега, раскрытие семядолей, рост листьев, дифференцировка хлоропластов.
Криптохромы регулируют экспрессию генов синтеза антоцианов, процесс деэтиоляции, фотопериодическую реакцию. Возможно, участвуют в передаче фитохромного сигнала.
Фототропины отвечают за рецепцию синего света в фототропич. реакции. Имеют 2 максимума поглощения при 390 и 450 нм. В процессе рецепции синего света происходит автофосфорилированиефототропина, что вызывает активацию Ca-каналов в плазмалемме, в цитоплазму поступают иона кальция, что включает сигнальную систему в растительной клетке. Также фототропины вовлечены в регуляцию движения хлоропластов и открывания устьиц, в регуляции роста путём растяжения клеток.
Фотопериодизм– это реакция растения на соотношение продолжительности дня и ночи, связанная с приспособлением онтогенеза к сезонным изменениям внешних условий.
Фотопериодическое воздействие вызывает свет малой интенсивности (на порядок меньше того, при котором идёт фотосинтез). Рецептором выступает листовая пластинка – она воспринимает длину дня и ночи (с помощью фитохрома), а изменения, в результате которых начинается заложение цветков, происходит в меристеме. На фотопериодич.воздействие отвечает ближайшая к листу точка роста. Чувствительность листовой пластинки зависит от возраста растения: наибольшая чувствительность у листа, достигшего максимального размера.
Механизм фотопериодического действия: фитохром поглощает свет, активизируется и вызывает синтез гормонов в листе; в меристеме под влиянием притекающих туда гормонов происходят изменения, приводящие к заложению цветка. Фитохром может действовать как рецептор (воспринимать сигнал, который или индуцирует, или ингибирует цветение), или как синхронизатор, участвующий в измерении времени.
Растения, в зависимости от того, какая продолжительность дня им требуется для зацветания, подразделяютсяна:
Короткодневные – растения, кот.зацветают при длине дня 8-12 ч в сутки. Находясь в более длинном дне, будет вызван только интенсивный рост, а не цветение. Этим растениям требуется тёмный период определенной продолжительности, потому что подготовка к заложению цветков происходит в темноте, а на свету этот процесс тормозится. (рис, кукуруза, просо, соя, сах. тростник, хлопчатник, сорго)
Длиннодневные – растения, кот.зацветают при продолжительности дня не менее 16-18 ч/сут, в коротком дне приобретают розеточную форму. Непрерывное освещение вызывает ускорение цветения. Необходим длинный светлый период. Подготовка к зацветанию у них происходит на свету. (пшеница, ячмень, овес, горчица, свекла, лен, шпинат, клевер)
Нейтральные – растения, зацветающие при любой продолжительности дня. (фасоль, томат)
Длинно-короткодневные растения – для перехода из вегетативного в генеративное состояние сначала должны развиваться при длинном, а затем – при коротком свете. Цветут осенью (бриофиллум)
Коротко-длиннодневные – наоборот: сначала при коротком, потом при длинном. (колокольчик средний, пеларгония крупноцветковая)
Существуют промежуточные формы – им нужен и не короткий, и не длинный день (от 12 до 16 часов). Если условия не выполняются – растение только вегетирует. (фасоль многоколосая).
В каждой из этих групп есть растения, которые требуют определенного фотопериода для зацветания (это облигатные растения) и растения, которые только ускоряют зацветание при нужной длине дня (необлигатные).
Длинно- или короткодневность у растения зависит от географического происхождения вида или сорта.
Короткий или длинный день нужен не всю жизнь, а только в определенное время, после чего растение зацветет при любой длине дня. При этом фотопериод выступает индуктором. Фотопериодическая индукция – явление благоприятных фотопериодов на развитие растений, приводящее к последующему их зацветанию независимо от длины дня. Складывается из двух фаз:
Листовая– когда в листьях под влиянием определенного фотопериода идёт образование гормона цветения.
Стеблевая – когда в стеблевых почках под влиянием гормонов начинаются изменения в метаболизме и закладываются цветки.
mykonspekts.ru
Экофизиологическая роль фотопериодизма у растений
Работа добавлена на сайт samzan.ru: 2016-03-13Приднестровский государственный университет им. Т.Г. Шевченко
Кафедра ботаники и экологии
Курсовая работа по физиологии растений
На тему: Экофизиологичсекая роль фотопериодизма у растений
Тирасполь, 2009
Оглавление
Введение
1. Понятие фотопериодизма
2. Локализация фотопериодической реакции
3. Практическое использование явления фотопериодизма
Заключение
Литература
Введение
Огромное влияние на жизнедеятельность растений и животных оказывает соотношение светлого (длина дня) и темного (длина ночи) периодов суток в течение года. Реакция организмов на суточный ритм освещения, выражающаяся в изменении процессов их роста и развития, называется фотопериодизмом. Регулярность и неизменная повторяемость из года в год данного явления позволила организмам в ходе эволюции согласовывать свои важнейшие жизненные процессы с ритмом этих временных интервалов. Под фотопериодическим контролем находятся практически все метаболические процессы, связанные с ростом, развитием, жизнедеятельностью и размножением растений.
Центральным моментом развития организма является переход от вегетативного роста к репродуктивному развитию (у цветковых растений – к цветению). Условия среды могут влиять на реализацию генетической информации и тем самым ускорять или замедлять наступление определенных этапов развития, в первую очередь переход растения к цветению. Таким образом, развитие растений совершается при взаимодействии генетических потенций и факторов внешней среды. Основными факторами среды, оказывающими влияние на переход растения к репродукции – к цветению, является температура и продолжительность дневного освещения.
1. Понятие фотопериодизма
Понятия фотопериод и фотопериодизм были введены в науку американскими физиологами растений У. Гарнером и Г.А. Аллардом (1920-1923).
Фотопериодизм – ритмические изменения морфологических, биохимических и физиологических свойств и функций под влиянием чередования и длительности световых и темновых интервалов.
ФОТОПЕРИОДИЧЕСКИЕ ГРУППЫ - группы растений с различной фотопериодической реакцией, названные по длине дня, ускоряющей их зацветание.
В зависимости от этого растения делятся на:
1. Нейтральные растения (НР) - длина дня не оказывает заметного влияния, растения цветут по достижении определенного возраста или размера. Обычно нейтральные растения происходят из экваториальных областей.
2. Длиннодневные растения (ДДР) - зацветают только в том случае, если длина дня больше некоторой критической величины. ДДР происходят из умеренных областей с равномерным увлажнением по сезонам.
3. Короткодневные растения (КДР) - зацветают только тогда, когда длина дня меньше, чем некоторая критическая величина. Они происходят из субтропических и тропических областей с зимним максимумом увлажнения. (Есть и растения с количественной реакцией на короткий день: цветение ускоряется на коротком дне, хотя длина дня не играет принципиальной роли).
4. Длиннокороткодневные растения (ДКДР) - для цветения необходима определенная последовательность: сначала длинные дни, а затем короткие. Эти растения настроены на благоприятный осенний период.
5. Короткодлиннодневные растения (КДДР) - для цветения необходима смена коротких дней на длинные (но не наоборот). Благоприятный период у этой группы ассоциируется с весенним сезоном.
6. Среднедневные растения (СДР) - для цветения необходим определенный интервал длины дня: ни при увеличенной, ни при уменьшенной длине дня эти растения не цветут. Это - сравнительно редкий тип регуляции цветения.
7. Амфифотопериодичные растения (АФПР) - для цветения неблагоприятен узкий интервал, а при большей или меньшей длине дня цветение наступает. Этот тип физиологических ответов также достаточно редок.
Отнесение растений к той или иной фотопериодической группе не связано с конкретной оптимальной длиной дня, а дает представление о том, ускоряется ли переход к цветению при увеличении или уменьшении длительности освещения в каждом фотопериоде. Среди растений есть виды и разновидности, как с качественным, так и с количественным типам фотопериодических ответов растений. Для короткодневных с качественной реакцией решающим фактором служит длина темнового периода. Кратковременное освещение этих растений в середине темнового периода препятствует их переходу к цветению. Прерывание светового периода темнотой не влияет на сроки цветения. Длиннодневные растения не нуждаются в периодах темноты и зацветают на непрерывном свету.
Свет и его роль в жизни растений.
Живая природа не может существовать без света, так как солнечная радиация, достигающая поверхности Земли, является практически единственным источником энергии для поддержания теплового баланса планеты, создания органических веществ фототрофными организмами биосферы, что в итоге обеспечивает формирование среды, способной удовлетворить жизненные потребности всех живых существ.
Биологическое действие солнечного света зависит от его спектрального состава, продолжительности, интенсивности, суточной и сезонной периодичности.
Солнечная радиация представляет собой электромагнитное излучение в широком диапазоне волн, составляющих непрерывный спектр от 290 до 3 000 нм. Ультрафиолетовые лучи (УФЛ) короче 290 им, губительные для живых организмов, поглощаются слоем озона и до Земли не доходят. Земли достигают главным образом инфракрасные (около 50% суммарной радиации) и видимые (45%) лучи спектра. На долю УФЛ, имеющих длину волны 290-380 нм, приходится 5% лучистой энергии. Длинноволновые УФЛ, обладающие большой энергией фотонов, отличаются высокой химической активностью. В небольших дозах они оказывают мощное бактерицидное действие, способствуют синтезу у растений некоторых витаминов, пигментов, а у животных и человека - витамина D; кроме того, у человека они вызывают загар, который является защитной реакцией кожи. Инфракрасные лучи длиной волны более 710 нм оказывают тепловое действие.
В экологическом отношении наибольшую значимость представляет видимая область спектра (390-710 нм), или фотосинтетически активная радиация (ФАР), которая поглощается пигментами хлоропластов и тем самым имеет решающее значение в жизни растений. Видимый свет нужен зеленым растениям для образования хлорофилла, формирования структуры хлоропластов; он регулирует работу устьичного аппарата, влияет на газообмен и транспирацию, стимулирует биосинтез белков и нуклеиновых кислот, повышает активность ряда светочувствительных ферментов. Свет влияет также на деление и растяжение клеток, ростовые процессы и на развитие растений, определяет сроки цветения и плодоношения, оказывает формообразующее воздействие.
Световой режим любого местообитания зависит от его географической широты, высоты над уровнем моря, состояния атмосферы, растительности, сезона и времени суток, солнечной активности и т. д. Поэтому разнообразие световых условий на нашей планете чрезвычайно велико: от таких сильно освещенных территорий, как высокогорья, пустыни, степи, до сумеречного освещения в водных глубинах и пещерах. В разных местообитаниях различаются не только интенсивность света, но и его спектральный состав, продолжительность освещения, пространственное и временное распределение света разной интенсивности и т.д. Соответственно, разнообразны и приспособления растений к жизни при том или ином световом режиме.
Экологические группы растений по отношению к свету.
По отношению к количеству света, необходимого для нормального развития, растения подразделяют натри экологические группы.
Светолюбивые, или гелиофиты, с оптимумом развития при полном освещении; сильное затенение действует на них угнетающе. Это растения открытых, хорошо освещенных местообитаний: степные и луговые травы, прибрежные и водные растения (с плавающими листьями), большинство культурных растений открытого грунта, сорняки и др.
Тенелюбивые, или теневые, с оптимальным развитием в пределах 1/10-1/3 от полного освещения, т.е. для них приемлемы области слабой освещенности. К тенелюбам относятся растения нижних затененных ярусов сложных растительных сообществ - темнохвойных и широколиственных лесов, а также водных глубин, расщелин скал, пещер и т.д. Тенелюбами являются и многие комнатные и оранжерейные растения. В лесах Беларуси и России типичными теневыми растениями являются копытень европейский, ветреница дубравная, сныть обыкновенная, чистотел большой, кислица обыкновенная, майник двулистный и др.
Теневыносливые растения имеют широкую экологическую амплитуду выносливости по отношению к свету. Они лучше растут и развиваются при полной освещенности, но хорошо адаптируются и к слабому свету. К ним относится большинство видов зоны смешанных лесов - ель, пихта, граб, бук, лещина, бузина, брусника, ландыш майский и др.
Адаптация растений к световому режиму.
Под влиянием различных условий светового режима у растений выработались соответствующие приспособительные качества. Прежде всего это касается величины листовых пластинок: у гелиофитов по сравнению с теплолюбивыми они обычно более мелкие. Ориентация листьев у светолюбов вертикальная или имеет разный угол по отношению к солнечным лучам, чтобы избежать избыточного света и перегрева. Листья теневыносливых растений, напротив, ориентированы к свету всей поверхностью листовой пластинки и расположены так, чтобы не затенять соседние листья (листовая мозаика).
У многих гелиофитов поверхность листовой пластинки блестящая, покрыта светлым восковым налетом, густо опушена, что способствует отражению палящих солнечных лучей или ослаблению их действия.
Световые и теневые растения имеют четкие различия и по анатомическому строению. Так, у гелиофитов хорошо развиты осевые органы с оптимальным соотношением ксилемы и механических тканей, менее сложные по форме листья с характерной дифференцировкой мезофилла на столбчатый и губчатый, высокой степенью жилкования, большим числом устьиц на единицу поверхности листа. У светолюбивых растений количество хлоропластов, приходящихся на единицу площади листовой пластинки, в несколько раз больше, чем у тенелюбивых. Сами хлоропласты у гелиофитов более мелкие и светлые (с малым содержанием хлорофилла), способные к изменению ориентировки и перемещениям в клетке: на сильном свету они занимают постенное положение и становятся «ребром» к направлению лучей, что защищает хлорофилловое разрушения.
Теневыносливые растения встречаются в местообитаниях с различным световым режимом благодаря увеличению ассимилирующей поверхности, снижению интенсивности дыхания и уменьшению относительной массы нефотосинтезирующих тканей, увеличению размеров хлоропластов и концентрации хлорофилла. Кроме того, в листьях наблюдается слабая дифференцировка на столбчатый и губчатый мезофилл или таковая совсем отсутствует, отмечается сравнительно малое количество устьиц и т. д.
Таким образом, способность воспринимать длину дня и реагировать на нее широко распространена в мире живых существ. Это означает, что живые организмы способны ориентироваться во времени, т. е. они обладают биологическими часами. Другими словами, для многих организмов характерна способность ощущать суточные, приливные, лунные и годичные циклы, что позволяет им заранее готовиться к предстоящим изменениям среды.
Правильно подобрав режимы освещения, температуры и другие факторы, наиболее соответствующие биоритмам, можно заметно повысить жизнедеятельность и продуктивность разводимых животных и растений, причем без каких-либо дополнительных затрат. Например, благодаря увеличению в теплицах, оранжереях и парниках светового дня до 12-15 ч зимой выращивают овощные культуры и декоративные растения, ускоряют рост и развитие рассады. Продлив за счет искусственного освещения световой период зимой, можно увеличить яйценоскость кур, уток, гусей, регулировать размножение пушных зверей на зверофермах.
Влияние качества света на развитие.
Много внимания было уделено исследователями качеству света, необходимого для цветения растений. Красный свет воспринимается растениями как свет в процессе развития, а сине-фиолетовая часть спектра – как темнота. Однако при увеличении интенсивности синего света он тоже начинает восприниматься растениями как свет. Опыты показывают, что если выровнять интенсивность света по числу квантов, то красный и синий свет становиться одинаково эффективным.
Важно выяснить в каких условиях проходят соответствующие изменения у растений короткого и длинного дня. Как показали опыты, во время пребывания растения в темноте на небольшой отрезок времени, в несколько минут, прервать темноту, то цветения не наступает. Того же самого можно достигнуть, давая этим растениям мигающий свет, где интервалы между светом равны секундами или минутами. У развития и важным является лишь то, чтобы в сумме число освещения соответствовало числу часов, характерному для длинного дня [4].
фотопериодизм растение свет
2. Локализация фотопериодических реакций
Фотопериодические воздействия воспринимаются главным образом листьями, а не апексами побега. У большинства растений максимальной чувствительность к фотопериоду обладают листья, только что прекратившие рост. Для дурнишника, зацветание которого индуцируется одним короткодневным фотопериодом, для фотоиндукции зацветания достаточно нескольких квадратных сантиметров листа. Основную роль в восприятии фотопериода листом играет фитохром. Предполагается, что его темновое превращение может служить способом измерения времени по типу песочных часов. Вспышка красного света устанавливает фазу эндогенного ритма.
Необходимость большой поверхности листьев и достаточной интенсивности света для перехода к цветению у многих растений объясняется потребностью растущих меристем в ассимилятах. Устранение СО2 из воздуха во время светового периода снижает инициацию цветения у некоторых короткодневных растений и длиннодневных растения.
У фотопериодически нейтральных растений переход к зацветанию обеспечивается возрастным изменениям.
Таким образом, сущность фотопериодической реакции заключается в том, что циклическое чередование света и темноты переводит растение из вегетативного в репродуктивное состояние.
Фотопериодизм, представляет собой приспособительную реакцию, позволяющую растениям зацветать в определённое, наиболее благоприятное время года. Как правило, длиннодневные растения северные, а короткодневные – южные. Для короткодневных растений более благоприятны повышенные ночные температуры, тогда как для длиннодневных – пониженные. Фотопериодическая реакция затрагивает не только процесс развития растений, но и вызывает некоторые изменения ростовых процессов.
Растения сильно различаются по числу фотопериодических циклов (числу суток с определённой длинной дня), которые вызывают затем переход к цветению одним растениям достаточно одного цикла, иначе говоря, воздействия определенной длинной дня в течение одних суток. Другим растениям необходимо получить определенную длину дня в течении 25 суток (25 циклов). Таким образом, фотопериодическое воздействие необходимо растительному организму лишь на протяжении определенного периода, после чего растение зацветает уже при любом соотношении дня и ночи. Это явление называют фотопериодической индукцией.
Исследования, проведенные на растениях короткого дня, показали, что для их перехода к образованию репродуктивных органов (цветению) важна длительность не дня, а ночи. Иначе говоря, для короткодневных растений необходим теневой период определенной длины. Если в середине темнового периода дать хотя бы вспышку света, то растение короткого дня к цветению уже не переходит. Так, если 14-часовая ночь прерывается освещением на 10 мин, то короткодневные растения не зацветают. Для длиннодневных растений нужен именно длинный световой период.
Фотопериодическое воздействие вызывается светом малой интенсивности. Так, достаточно прервать темновой период суток вспышкой света интенсивностью в 3-5 люкс, чтобы короткодневные растения не зацвели. Даже лунный свет препятствует переходу к цветению короткодневных растений. Фотосинтез при таких интенсивностях освещения еще не идет. Эти данные показывают относительную независимость явлений фотопериодизма от фотосинтеза. Вместе с тем, по мнения М.Х. Чайлахяна, переходу растений к цветению должен предшествовать определенный период нормального протекания фотосинтеза.
Фотопериодическая реакция растений наиболее успешно проходит лишь в лучах определенной длины волны, что доказано опытами американских ученых Хендрикса и Борствика. Наиболее активными в смысле задержки цветения сои оказались красные лучи (длина 600Нм). Сине-фиолетовые лучи (используемые при фотосинтезе) оказались не активными. Таким образом спектр действия фотопериодической реакции оказался совпадающим со спектром действия фитохрома. Это подтверждается тем, что действие вспышки красного света (Фк) на задержку цветения короткодневных растений снимается действием дальнего красного света (Фдк). В ночной период Фдк под влиянием дальних красных лучей превращается в Фк, и это способствует началу реакций, приводящих короткодневные растения к цветению. При вспышке красного света Фк превращается в Фдк, и это ингибирует реакции, приводящие к цветению. Следовательно, для перехода к цветению короткодневных растений нужно меньшее содержание активного фитохрома, поглощающего дальние красные лучи, тогда как для перехода к цветению длиннодневных необходимо его более высокое содержание.
Для фотопериодического действия рецепторным(воспринимающим) органом является лист. В опытах, проведенных с короткодневными растениями, в частности хризантемой, воздействию короткого дня подвергали либо листья, либо обезлиственные побеги. С этой целью указанные части растения закрывали на определенное время пакетами из непроницаемой для света бумаги. Оказалось, что растение зацветало в том случае, если воздействию укороченного дня подвергали только листья или листья вместе с точкой роста. Те растения, у которых воздействию короткого дня подвергали только точку роста или обезлиственные побеги, не зацветали.
Таким образом, именно листья являются органами, воспринимающими фотопериодическое воздействие, под влиянием которого происходят ответные изменения в точке роста. Результатом этих изменений является превращение вегетативной почки в цветочную. Это дало основание М.Х. Чайланяну предположить, что в листе вырабатываются вещества (гормоны), которые вызывают переход к цветению. Оказалось далее, что возникающие в листе вещества по преимуществу локализуются в том побеге, где они образуются, так как передвижение идет от листьев к точке роста, расположенной на том же побеге.
Интересные результаты дали опыты с прививками. Оказалось, что, если побеги с листьями короткодневного растения ( например хризантемы), выдержанного на коротком дне, привит на растение этого же типа, выдержанного на длинном дне, то последнее зацветает. Это подтвердило положение, что в листьях первого растения образуется гормональные вещества, которые при прививке вызывают изменения – развитие цветочной почки - у второго растения. Важные результаты были получены при прививках длиннодневных растений на короткодневные и обратно. Оказалось, что при прививке растений топинамбура (растение короткого дня) на цветущее растение подсолнечника (растение короткого дня) топинамбур зацветал при длинном дне. Сходные результаты были получены в опытах в которых длиннодневные растения прививались на цветущее короткодневное. Таким образом, исследователи показали, что листья, выдержанные на соответствующем фотопериоде, содержат вещества, вызывающие цветение в независимости от условий, в которых содержаться остальные растения. При этом вещества, вызывающие цветение, идентичны как для короткодневных, так и для длиннодневных растений[3].
3. Практическое использование явления фотопериодизма
Размножение клубнями.
Способность к клубнеобразованию формируется в процессе онтогенеза постепенно и проявляется после завершения ювенильного этапа в процессе клубнеобразования вначале развиваются столоны – стебли с измененной геотропической реакцией(1-я фаза), а затем на них формируются клубни (2-я фаза). Наиболее существенное влияние на клубнеобразование оказывают температура и длина дня. При этом у одной группы растений клубни образуются только в условиях короткого дня, а у другой – как при коротком, так и при длинном дне. В опятах с прививками растений, не образующих клубни, на клубненосы (например, томатов на картофель, подсолнечника на топинамбур) было выяснено, что стимул клубнеобразования у фотопериодически чувствительных клубненосов вырабатывается в листьях, что он не обладает видоспецифичностью и имеет гормональную природу. Оказалось, что в этом процессе взаимодействуют основные группы фитогормонов: ауксин подавляет клубнеобразование; гиббереллины, индуцирующие рост стеблей, усиливают рост столонов и тем самым в определенном отношении способствуют росту клубней. Наиболее значительное влияние на формирование клубней оказывают цитокинины. В условиях короткого дня в корнях и столонах наблюдается высокое содержание цитокининов, а в листьях – абсцизинов. При длинном дне в листьях выше содержание гиббереллинов, а в стеблевых почках – ауксинов.
Согласно М.Х. Чайлахяну, механизм гормональной индукции клубнеобразования у видов, образующих клубни на коротком дне, включает в себя две фазы. В начале из листьев в нижние стеблевые почки поступают гиббереллины и абсцизины. Поскольку при коротком дне соотношение их сдвинуто в сторону преобладания абсцизинов (АБК/ГА), ускоренный рост столонов, вызываемый гиббереллином, тормозиться. Во время второй фазы образуются клубни, рост которых регулируется более высокой концентрацией цитокинина в корнях и столонах по сравнению с ауксином (ЦК/ИУК).
В условиях длинного дня при неблагоприятном для клубнеобразования соотношении гормонов ( в листьях ГА/АБК, в корнях – ИУК/Цк) происходит образование и рост корней и столонов. Таким образом, стимулом клубнеобразования, вырабатываемым в листьях, служит определенное соотношение гиббереллинов и абсцизинов, влияющее на первую фазу клубнеобразования.
Развитие и рост клубней поддерживается притоком к ним продуктов фотосинтеза, образуемых в листьях. При этом интенсивность транспорта ассимилятов в клубни коррелирует с содержанием в них ауксинов и гиббереллинов, создающих оттрагирующую силу клубней. Именно поэтому при интенсивном клубнеобразвании замедляется рост надземных частей растения.
У сортов картофеля, выращиваемых в условиях короткого дня, вначале идет рост надземных частей, а затем – интенсивное столоно-и клубнеобразование. Для сортов, образующих клубни и при длинном дне, характерен более протяженный период вегетации, когда развивается надземная масса, а формирование клубней начинается с ускорением дня во второй половине лета [1].
Размножение луковицами.
Если большинство клубненосов образует клубни на коротком дне, то для луковичных растений, напротив, характерно формирование луковиц на длинном дне. Возможно, это связано с эколого-географическим происхождением разных видов. Регуляция образования луковиц изучено гораздо слабее, чем регуляция клубнеобразования. Органами, рецептирующими действие фотопериода, в данном случае также являются листья. В опытах Ф.Э.Реймерса выдерживание даже части листьев на коротком дне замедляло формирование луковицы. Фотопериодический стимул может передаваться от одного побега лука к другому только при условии, что второй побег обезлиствен. Если же побеги с листьями подвергаются воздействию разных фотопериодов, то луковицу формирует, лишь побег, находившийся на длинном дне. При образовании луковиц фотпериодический стимул, как при клубнеобразовании и цветении, по-видимому, является комплексом фитогормонов. Так, формирование луковицы сопровождается уменьшением активности ауксинов, а повышение из концентрации ведет к росту листьев и луковичных чешуй в длину. Однако вегетативное размножение луковичных растений связано не столько с формирование луковицы, сколько с образованием на ней пазушных побегов – будущих дочерних луковичек, «деток». Они закладываются в период зимнего покоя луковиц. Обнаружено, что под действием кинетина, ГА3 и ИУК, нанесенных на донце луковицы, значительно усиливается образование почек, формирующих дочерние луковички. Тот же эффект наблюдается при повышении температуры зимнего хранения луковиц или при удалении цветоносного побега. Прорастание клубней и луковиц происходит по истечении периода покоя. У луковичных растений в отличии от семян и почек, ростовые процессы полностью не прекращаются. В период покоя, хотя и очень медленно, формируются новые органы. Выход из состояния окоя ускоряется после обработки луковиц низкой температурой, т.е. после яровизации. Состояние покоя луковиц поддерживается высоким содержанием в них АБК, а при прорастании снижается уровень ингибиторов и повышается содержание ауксинов, цитокининов и гиббериллинов.
Знание принципов регуляции состояния покоя клубней и луковиц позволяет предотвращать их прорастание в период хранения, например лука и картофеля, с помощью многих ингибиторов роста, таких, как гидразид малеиновой кислоты [1].
Регуляция цветения у покрытосеменных растений.
В любом онтогенезе происходит постепенная реализация генетической информации, заключенной в ДНК. Молекулярные основы наследственной информации, связанные с редупликацией ДНК (хранением), транскрипцией (передачей) и трансляцией (реализацией) генетической информации, очень универсальны и обнаруживают большую общность во всем мире растительных организмов. В то же время выявляется широкое разнообразие видовой и фазовой специфичности регуляции пути от ДНК до белка, которая, в частности, зависит от условий среды.
Соответственно, кроме фотопериодической реакции, регуляторными механизмами развития являются температурные и возрастные факторы. Все эти условия не исключают, а дополняют друг друга.
В результате световых и темновых реакций фотопериодизма синтезируются гормональные регуляторы цветения. В результате этих преобразований в точках роста происходят изменения вегетативного морфогенеза на репродуктивный. В случае, если происходит блокировка любого из звеньев ДНК – РНК - белок, цветение не наступает.
Рецепторная роль листа в фотопериодической реакции заставляет предполагать, что у всех растений это свойство связано с каким-то фундаментальным процессом. Эту общую основу ищут в нескольких направлениях, а именно в роли фитохрома или эндогенного циркадного (суточного) ритма или отводят основную роль окислительно- восстановительным процессам листа, а именно взаимодействию процессов фотосинтеза и дыхания.
По этому воззрению, на свету получают большое значение процессы, связанные с восстановлением СО2 и фотофосфолирированием, а в темноте – с окислительным фосфорилированием.
Таким образом, в листе происходит функциональное взаимодействие митохондрий и хлоропластов, регулируемое светом и темнотой, сопряженное с биосинтезом гормональных веществ.
Таково дальнейшее течение гормональной концепции развития растений, многие звенья которой еще пока гипотетичны. Все опыты по влиянию фитогормонов и других физиологически активных веществ представляет большой интерес с точки зрения того, что развитие растений обусловлено исторически в эволюционном процессе, однако проявляются различно, в зависимости от внешних условий, в связи с приспособлением растений к окружающим условиям, т.е. к длине дня, температуре и т.д.[4]
С. С. Шаин экспериментально доказал, что при посеве растений длинного дня радами с севера на юг, а короткого дня- с востока на запад они лучше развиваются и дают больший урожай. Растения, высеянные рядами с севера на юг, лучше освещаются, потому что лучи падают перпендикулярно к ним в утренние и вечерние часы, когда сравнительно больше длинноволновой фракции спектра. В полдень, когда в солнечном свете сравнительно много коротковолновых лучей, растения в рядах затеняют друг друга. Следовательно, посевы с рядами, расположенными с востока на запад, получают больше коротковолновых лучей в течении дня, чем при посеве с севера на юг.
В опятах лаборатории фотосинтеза Украинской сельскохозяйственной академии при пунктирных посевах кукурузы с ориентацией рядов с севера-востока на юго-запад КПД фотосинтеза за вегетационный период повышается на 0,15-0,21% по сравнению с ориентацией рядов с севера на юг. Урожайность зерна кукурузы при этом возросла на 10-13%. Современные методы измерения фотосинтетически активной радиации показали, что при высоте солнца над горизонтом 20о и в течении дня спектральный состав ФАР почти не изменяется. Кроме того, большая продуктивность посевов с определенной ориентацией рядов обуславливается не только условиями освещения. При разной ориентации рядов растений условия корневого питания и водного режима также будут неодинаковыми, что, в свою очередь, влияет на поглощение и использование ФАР.
Повышенную продуктивность посевов с определенной ориентацией рядов канадский ученый У. Питтман объясняет реакцией корневой системы на направленность магнитных силовых линий Земли. В результате этого создаются лучшие условия растений. Направленность рядов имеет большое значение при посевах и посадках лесных пород, а также при уборках, уходе за молодняком. В южных широтах и в условиях резко выраженного климата широтное распоряжение рядов деревьев имеет положительное значение, так как лучше обеспечивается их взаимная защита от избыточной радиации и перегрева. В северных широтах более эффективно меридиальное расположение рядов, что способствует лучшему прогреванию почвы в междурядьях[5].
Заключение
Под контролем фотопериода находятся самые разнообразные процессы. Так, у многих растений от длины дня зависит перестройка работы меристемы побега и образование цветков. При определенной длине дня возникают и почки у многих деревьев. Земляника образует расселительные побеги - усы - при наступлении длинных дней. Сбрасывание листьев у репчатого лука и тюльпанов происходит на длинном дне, а у березы и осины - на коротком. Образование запасающих органов (кочанов у капусты, клубней у диких видов картофеля и топинамбура) также зависит от длины дня. Растения пустынь могут изменять листья С-3 типа на С-4 и наоборот, а водные растения - сменяют подводные листья на надводные, получая из внешней среды фотопериодический сигнал. Из всего выше сказанного понятно что, фотопериодизм играет очень важную роль в жизни растений.
Литература
1. Полевой В.В. «Физиология растений». М.: Высшая школа. 1989.
2. Э.Ф. Шабельская. «Физиология растений». Минск: Высшая школа, 1987.
3. Н.И. Якушкина. «Физиология растений». М.: Просвещение, 1980.
4. П.А. Генкель. «Физиология растений». М.: Просвещение, 1975.
5. С.И. Лебедев «Физиология растений». М.: ВО «Агропромиздат» 1988.
6. Р. Ван Дер Вин, Г. Мейер «Свет и рост растений». М., 1962 г.
7. Рубин Б.А. «Курс физиологии растений». М.: Москва, 1982 г.
8. Чайлахян М.Х. «Фотопериодическая и гормональная регуляция клубнеобразования у растений». М.: 1984 г.
samzan.ru
Экофизиологическая роль фотопериодизма у растений
Приднестровский государственный университет им. Т.Г. Шевченко
Кафедра ботаники и экологии
Курсовая работа по физиологии растений
На тему: Экофизиологичсекая роль фотопериодизма у растений
Тирасполь, 2009
Оглавление
Введение
1. Понятие фотопериодизма
2. Локализация фотопериодической реакции
3. Практическое использование явления фотопериодизма
Заключение
Литература
Введение
Огромное влияние на жизнедеятельность растений и животных оказывает соотношение светлого (длина дня) и темного (длина ночи) периодов суток в течение года. Реакция организмов на суточный ритм освещения, выражающаяся в изменении процессов их роста и развития, называется фотопериодизмом. Регулярность и неизменная повторяемость из года в год данного явления позволила организмам в ходе эволюции согласовывать свои важнейшие жизненные процессы с ритмом этих временных интервалов. Под фотопериодическим контролем находятся практически все метаболические процессы, связанные с ростом, развитием, жизнедеятельностью и размножением растений.
Центральным моментом развития организма является переход от вегетативного роста к репродуктивному развитию (у цветковых растений – к цветению). Условия среды могут влиять на реализацию генетической информации и тем самым ускорять или замедлять наступление определенных этапов развития, в первую очередь переход растения к цветению. Таким образом, развитие растений совершается при взаимодействии генетических потенций и факторов внешней среды. Основными факторами среды, оказывающими влияние на переход растения к репродукции – к цветению, является температура и продолжительность дневного освещения.
1. Понятие фотопериодизма
Понятия фотопериод и фотопериодизм были введены в науку американскими физиологами растений У. Гарнером и Г.А. Аллардом (1920-1923).
Фотопериодизм – ритмические изменения морфологических, биохимических и физиологических свойств и функций под влиянием чередования и длительности световых и темновых интервалов.
ФОТОПЕРИОДИЧЕСКИЕ ГРУППЫ - группы растений с различной фотопериодической реакцией, названные по длине дня, ускоряющей их зацветание.
В зависимости от этого растения делятся на:
Нейтральные растения (НР) - длина дня не оказывает заметного влияния, растения цветут по достижении определенного возраста или размера. Обычно нейтральные растения происходят из экваториальных областей.
Длиннодневные растения (ДДР) - зацветают только в том случае, если длина дня больше некоторой критической величины. ДДР происходят из умеренных областей с равномерным увлажнением по сезонам.
Короткодневные растения (КДР) - зацветают только тогда, когда длина дня меньше, чем некоторая критическая величина. Они происходят из субтропических и тропических областей с зимним максимумом увлажнения. (Есть и растения с количественной реакцией на короткий день: цветение ускоряется на коротком дне, хотя длина дня не играет принципиальной роли).
Длиннокороткодневные растения (ДКДР) - для цветения необходима определенная последовательность: сначала длинные дни, а затем короткие. Эти растения настроены на благоприятный осенний период.
Короткодлиннодневные растения (КДДР) - для цветения необходима смена коротких дней на длинные (но не наоборот). Благоприятный период у этой группы ассоциируется с весенним сезоном.
Среднедневные растения (СДР) - для цветения необходим определенный интервал длины дня: ни при увеличенной, ни при уменьшенной длине дня эти растения не цветут. Это - сравнительно редкий тип регуляции цветения.
Амфифотопериодичные растения (АФПР) - для цветения неблагоприятен узкий интервал, а при большей или меньшей длине дня цветение наступает. Этот тип физиологических ответов также достаточно редок.
Отнесение растений к той или иной фотопериодической группе не связано с конкретной оптимальной длиной дня, а дает представление о том, ускоряется ли переход к цветению при увеличении или уменьшении длительности освещения в каждом фотопериоде. Среди растений есть виды и разновидности, как с качественным, так и с количественным типам фотопериодических ответов растений. Для короткодневных с качественной реакцией решающим фактором служит длина темнового периода. Кратковременное освещение этих растений в середине темнового периода препятствует их переходу к цветению. Прерывание светового периода темнотой не влияет на сроки цветения. Длиннодневные растения не нуждаются в периодах темноты и зацветают на непрерывном свету.
Свет и его роль в жизни растений.
Живая природа не может существовать без света, так как солнечная радиация, достигающая поверхности Земли, является практически единственным источником энергии для поддержания теплового баланса планеты, создания органических веществ фототрофными организмами биосферы, что в итоге обеспечивает формирование среды, способной удовлетворить жизненные потребности всех живых существ.
Биологическое действие солнечного света зависит от его спектрального состава, продолжительности, интенсивности, суточной и сезонной периодичности.
Солнечная радиация представляет собой электромагнитное излучение в широком диапазоне волн, составляющих непрерывный спектр от 290 до 3 000 нм. Ультрафиолетовые лучи (УФЛ) короче 290 им, губительные для живых организмов, поглощаются слоем озона и до Земли не доходят. Земли достигают главным образом инфракрасные (около 50% суммарной радиации) и видимые (45%) лучи спектра. На долю УФЛ, имеющих длину волны 290-380 нм, приходится 5% лучистой энергии. Длинноволновые УФЛ, обладающие большой энергией фотонов, отличаются высокой химической активностью. В небольших дозах они оказывают мощное бактерицидное действие, способствуют синтезу у растений некоторых витаминов, пигментов, а у животных и человека - витамина D; кроме того, у человека они вызывают загар, который является защитной реакцией кожи. Инфракрасные лучи длиной волны более 710 нм оказывают тепловое действие.
В экологическом отношении наибольшую значимость представляет видимая область спектра (390-710 нм), или фотосинтетически активная радиация (ФАР), которая поглощается пигментами хлоропластов и тем самым имеет решающее значение в жизни растений. Видимый свет нужен зеленым растениям для образования хлорофилла, формирования структуры хлоропластов; он регулирует работу устьичного аппарата, влияет на газообмен и транспирацию, стимулирует биосинтез белков и нуклеиновых кислот, повышает активность ряда светочувствительных ферментов. Свет влияет также на деление и растяжение клеток, ростовые процессы и на развитие растений, определяет сроки цветения и плодоношения, оказывает формообразующее воздействие.
Световой режим любого местообитания зависит от его географической широты, высоты над уровнем моря, состояния атмосферы, растительности, сезона и времени суток, солнечной активности и т. д. Поэтому разнообразие световых условий на нашей планете чрезвычайно велико: от таких сильно освещенных территорий, как высокогорья, пустыни, степи, до сумеречного освещения в водных глубинах и пещерах. В разных местообитаниях различаются не только интенсивность света, но и его спектральный состав, продолжительность освещения, пространственное и временное распределение света разной интенсивности и т.д. Соответственно, разнообразны и приспособления растений к жизни при том или ином световом режиме.
Экологические группы растений по отношению к свету.
По отношению к количеству света, необходимого для нормального развития, растения подразделяют натри экологические группы.
Светолюбивые, или гелиофиты, с оптимумом развития при полном освещении; сильное затенение действует на них угнетающе. Это растения открытых, хорошо освещенных местообитаний: степные и луговые травы, прибрежные и водные растения (с плавающими листьями), большинство культурных растений открытого грунта, сорняки и др.
Тенелюбивые, или теневые, с оптимальным развитием в пределах 1/10-1/3 от полного освещения, т.е. для них приемлемы области слабой освещенности. К тенелюбам относятся растения нижних затененных ярусов сложных растительных сообществ - темнохвойных и широколиственных лесов, а также водных глубин, расщелин скал, пещер и т.д. Тенелюбами являются и многие комнатные и оранжерейные растения. В лесах Беларуси и России типичными теневыми растениями являются копытень европейский, ветреница дубравная, сныть обыкновенная, чистотел большой, кислица обыкновенная, майник двулистный и др.
Теневыносливые растения имеют широкую экологическую амплитуду выносливости по отношению к свету. Они лучше растут и развиваются при полной освещенности, но хорошо адаптируются и к слабому свету. К ним относится большинство видов зоны смешанных лесов - ель, пихта, граб, бук, лещина, бузина, брусника, ландыш майский и др.
Адаптация растений к световому режиму.
Под влиянием различных условий светового режима у растений выработались соответствующие приспособительные качества. Прежде всего это касается величины листовых пластинок: у гелиофитов по сравнению с теплолюбивыми они обычно более мелкие. Ориентация листьев у светолюбов вертикальная или имеет разный угол по отношению к солнечным лучам, чтобы избежать избыточного света и перегрева. Листья теневыносливых растений, напротив, ориентированы к свету всей поверхностью листовой пластинки и расположены так, чтобы не затенять соседние листья (листовая мозаика).
У многих гелиофитов поверхность листовой пластинки блестящая, покрыта светлым восковым налетом, густо опушена, что способствует отражению палящих солнечных лучей или ослаблению их действия.
Световые и теневые растения имеют четкие различия и по анатомическому строению. Так, у гелиофитов хорошо развиты осевые органы с оптимальным соотношением ксилемы и механических тканей, менее сложные по форме листья с характерной дифференцировкой мезофилла на столбчатый и губчатый, высокой степенью жилкования, большим числом устьиц на единицу поверхности листа. У светолюбивых растений количество хлоропластов, приходящихся на единицу площади листовой пластинки, в несколько раз больше, чем у тенелюбивых. Сами хлоропласты у гелиофитов более мелкие и светлые (с малым содержанием хлорофилла), способные к изменению ориентировки и перемещениям в клетке: на сильном свету они занимают постенное положение и становятся «ребром» к направлению лучей, что защищает хлорофилловое разрушения.
Теневыносливые растения встречаются в местообитаниях с различным световым режимом благодаря увеличению ассимилирующей поверхности, снижению интенсивности дыхания и уменьшению относительной массы нефотосинтезирующих тканей, увеличению размеров хлоропластов и концентрации хлорофилла. Кроме того, в листьях наблюдается слабая дифференцировка на столбчатый и губчатый мезофилл или таковая совсем отсутствует, отмечается сравнительно малое количество устьиц и т. д.
Таким образом, способность воспринимать длину дня и реагировать на нее широко распространена в мире живых существ. Это означает, что живые организмы способны ориентироваться во времени, т. е. они обладают биологическими часами. Другими словами, для многих организмов характерна способность ощущать суточные, приливные, лунные и годичные циклы, что позволяет им заранее готовиться к предстоящим изменениям среды.
Правильно подобрав режимы освещения, температуры и другие факторы, наиболее соответствующие биоритмам, можно заметно повысить жизнедеятельность и продуктивность разводимых животных и растений, причем без каких-либо дополнительных затрат. Например, благодаря увеличению в теплицах, оранжереях и парниках светового дня до 12-15 ч зимой выращивают овощные культуры и декоративные растения, ускоряют рост и развитие рассады. Продлив за счет искусственного освещения световой период зимой, можно увеличить яйценоскость кур, уток, гусей, регулировать размножение пушных зверей на зверофермах.
Влияние качества света на развитие.
Много внимания было уделено исследователями качеству света, необходимого для цветения растений. Красный свет воспринимается растениями как свет в процессе развития, а сине-фиолетовая часть спектра – как темнота. Однако при увеличении интенсивности синего света он тоже начинает восприниматься растениями как свет. Опыты показывают, что если выровнять интенсивность света по числу квантов, то красный и синий свет становиться одинаково эффективным.
Важно выяснить в каких условиях проходят соответствующие изменения у растений короткого и длинного дня. Как показали опыты, во время пребывания растения в темноте на небольшой отрезок времени, в несколько минут, прервать темноту, то цветения не наступает. Того же самого можно достигнуть, давая этим растениям мигающий свет, где интервалы между светом равны секундами или минутами. У развития и важным является лишь то, чтобы в сумме число освещения соответствовало числу часов, характерному для длинного дня [4].
фотопериодизм растение свет
2. Локализация фотопериодических реакций
Фотопериодические воздействия воспринимаются главным образом листьями, а не апексами побега. У большинства растений максимальной чувствительность к фотопериоду обладают листья, только что прекратившие рост. Для дурнишника, зацветание которого индуцируется одним короткодневным фотопериодом, для фотоиндукции зацветания достаточно нескольких квадратных сантиметров листа. Основную роль в восприятии фотопериода листом играет фитохром. Предполагается, что его темновое превращение может служить способом измерения времени по типу песочных часов. Вспышка красного света устанавливает фазу эндогенного ритма.
Необходимость большой поверхности листьев и достаточной интенсивности света для перехода к цветению у многих растений объясняется потребностью растущих меристем в ассимилятах. Устранение СО2 из воздуха во время светового периода снижает инициацию цветения у некоторых короткодневных растений и длиннодневных растения.
У фотопериодически нейтральных растений переход к зацветанию обеспечивается возрастным изменениям.
Таким образом, сущность фотопериодической реакции заключается в том, что циклическое чередование света и темноты переводит растение из вегетативного в репродуктивное состояние.
Фотопериодизм, представляет собой приспособительную реакцию, позволяющую растениям зацветать в определённое, наиболее благоприятное время года. Как правило, длиннодневные растения северные, а короткодневные – южные. Для короткодневных растений более благоприятны повышенные ночные температуры, тогда как для длиннодневных – пониженные. Фотопериодическая реакция затрагивает не только процесс развития растений, но и вызывает некоторые изменения ростовых процессов.
Растения сильно различаются по числу фотопериодических циклов (числу суток с определённой длинной дня), которые вызывают затем переход к цветению одним растениям достаточно одного цикла, иначе говоря, воздействия определенной длинной дня в течение одних суток. Другим растениям необходимо получить определенную длину дня в течении 25 суток (25 циклов). Таким образом, фотопериодическое воздействие необходимо растительному организму лишь на протяжении определенного периода, после чего растение зацветает уже при любом соотношении дня и ночи. Это явление называют фотопериодической индукцией.
Исследования, проведенные на растениях короткого дня, показали, что для их перехода к образованию репродуктивных органов (цветению) важна длительность не дня, а ночи. Иначе говоря, для короткодневных растений необходим теневой период определенной длины. Если в середине темнового периода дать хотя бы вспышку света, то растение короткого дня к цветению уже не переходит. Так, если 14-часовая ночь прерывается освещением на 10 мин, то короткодневные растения не зацветают. Для длиннодневных растений нужен именно длинный световой период.
Фотопериодическое воздействие вызывается светом малой интенсивности. Так, достаточно прервать темновой период суток вспышкой света интенсивностью в 3-5 люкс, чтобы короткодневные растения не зацвели. Даже лунный свет препятствует переходу к цветению короткодневных растений. Фотосинтез при таких интенсивностях освещения еще не идет. Эти данные показывают относительную независимость явлений фотопериодизма от фотосинтеза. Вместе с тем, по мнения М.Х. Чайлахяна, переходу растений к цветению должен предшествовать определенный период нормального протекания фотосинтеза.
Фотопериодическая реакция растений наиболее успешно проходит лишь в лучах определенной длины волны, что доказано опытами американских ученых Хендрикса и Борствика. Наиболее активными в смысле задержки цветения сои оказались красные лучи (длина 600Нм). Сине-фиолетовые лучи (используемые при фотосинтезе) оказались не активными. Таким образом спектр действия фотопериодической реакции оказался совпадающим со спектром действия фитохрома. Это подтверждается тем, что действие вспышки красного света (Фк) на задержку цветения короткодневных растений снимается действием дальнего красного света (Фдк). В ночной период Фдк под влиянием дальних красных лучей превращается в Фк, и это способствует началу реакций, приводящих короткодневные растения к цветению. При вспышке красного света Фк превращается в Фдк, и это ингибирует реакции, приводящие к цветению. Следовательно, для перехода к цветению короткодневных растений нужно меньшее содержание активного фитохрома, поглощающего дальние красные лучи, тогда как для перехода к цветению длиннодневных необходимо его более высокое содержание.
Для фотопериодического действия рецепторным(воспринимающим) органом является лист. В опытах, проведенных с короткодневными растениями, в частности хризантемой, воздействию короткого дня подвергали либо листья, либо обезлиственные побеги. С этой целью указанные части растения закрывали на определенное время пакетами из непроницаемой для света бумаги. Оказалось, что растение зацветало в том случае, если воздействию укороченного дня подвергали только листья или листья вместе с точкой роста. Те растения, у которых воздействию короткого дня подвергали только точку роста или обезлиственные побеги, не зацветали.
Таким образом, именно листья являются органами, воспринимающими фотопериодическое воздействие, под влиянием которого происходят ответные изменения в точке роста. Результатом этих изменений является превращение вегетативной почки в цветочную. Это дало основание М.Х. Чайланяну предположить, что в листе вырабатываются вещества (гормоны), которые вызывают переход к цветению. Оказалось далее, что возникающие в листе вещества по преимуществу локализуются в том побеге, где они образуются, так как передвижение идет от листьев к точке роста, расположенной на том же побеге.
Интересные результаты дали опыты с прививками. Оказалось, что, если побеги с листьями короткодневного растения ( например хризантемы), выдержанного на коротком дне, привит на растение этого же типа, выдержанного на длинном дне, то последнее зацветает. Это подтвердило положение, что в листьях первого растения образуется гормональные вещества, которые при прививке вызывают изменения – развитие цветочной почки - у второго растения. Важные результаты были получены при прививках длиннодневных растений на короткодневные и обратно. Оказалось, что при прививке растений топинамбура (растение короткого дня) на цветущее растение подсолнечника (растение короткого дня) топинамбур зацветал при длинном дне. Сходные результаты были получены в опытах в которых длиннодневные растения прививались на цветущее короткодневное. Таким образом, исследователи показали, что листья, выдержанные на соответствующем фотопериоде, содержат вещества, вызывающие цветение в независимости от условий, в которых содержаться остальные растения. При этом вещества, вызывающие цветение, идентичны как для короткодневных, так и для длиннодневных растений[3].
3. Практическое использование явления фотопериодизма
Размножение клубнями.
Способность к клубнеобразованию формируется в процессе онтогенеза постепенно и проявляется после завершения ювенильного этапа в процессе клубнеобразования вначале развиваются столоны – стебли с измененной геотропической реакцией(1-я фаза), а затем на них формируются клубни (2-я фаза). Наиболее существенное влияние на клубнеобразование оказывают температура и длина дня. При этом у одной группы растений клубни образуются только в условиях короткого дня, а у другой – как при коротком, так и при длинном дне. В опятах с прививками растений, не образующих клубни, на клубненосы (например, томатов на картофель, подсолнечника на топинамбур) было выяснено, что стимул клубнеобразования у фотопериодически чувствительных клубненосов вырабатывается в листьях, что он не обладает видоспецифичностью и имеет гормональную природу. Оказалось, что в этом процессе взаимодействуют основные группы фитогормонов: ауксин подавляет клубнеобразование; гиббереллины, индуцирующие рост стеблей, усиливают рост столонов и тем самым в определенном отношении способствуют росту клубней. Наиболее значительное влияние на формирование клубней оказывают цитокинины. В условиях короткого дня в корнях и столонах наблюдается высокое содержание цитокининов, а в листьях – абсцизинов. При длинном дне в листьях выше содержание гиббереллинов, а в стеблевых почках – ауксинов.
Согласно М.Х. Чайлахяну, механизм гормональной индукции клубнеобразования у видов, образующих клубни на коротком дне, включает в себя две фазы. В начале из листьев в нижние стеблевые почки поступают гиббереллины и абсцизины. Поскольку при коротком дне соотношение их сдвинуто в сторону преобладания абсцизинов (АБК/ГА), ускоренный рост столонов, вызываемый гиббереллином, тормозиться. Во время второй фазы образуются клубни, рост которых регулируется более высокой концентрацией цитокинина в корнях и столонах по сравнению с ауксином (ЦК/ИУК).
В условиях длинного дня при неблагоприятном для клубнеобразования соотношении гормонов ( в листьях ГА/АБК, в корнях – ИУК/Цк) происходит образование и рост корней и столонов. Таким образом, стимулом клубнеобразования, вырабатываемым в листьях, служит определенное соотношение гиббереллинов и абсцизинов, влияющее на первую фазу клубнеобразования.
Развитие и рост клубней поддерживается притоком к ним продуктов фотосинтеза, образуемых в листьях. При этом интенсивность транспорта ассимилятов в клубни коррелирует с содержанием в них ауксинов и гиббереллинов, создающих оттрагирующую силу клубней. Именно поэтому при интенсивном клубнеобразвании замедляется рост надземных частей растения.
У сортов картофеля, выращиваемых в условиях короткого дня, вначале идет рост надземных частей, а затем – интенсивное столоно-и клубнеобразование. Для сортов, образующих клубни и при длинном дне, характерен более протяженный период вегетации, когда развивается надземная масса, а формирование клубней начинается с ускорением дня во второй половине лета [1].
Размножение луковицами.
Если большинство клубненосов образует клубни на коротком дне, то для луковичных растений, напротив, характерно формирование луковиц на длинном дне. Возможно, это связано с эколого-географическим происхождением разных видов. Регуляция образования луковиц изучено гораздо слабее, чем регуляция клубнеобразования. Органами, рецептирующими действие фотопериода, в данном случае также являются листья. В опытах Ф.Э.Реймерса выдерживание даже части листьев на коротком дне замедляло формирование луковицы. Фотопериодический стимул может передаваться от одного побега лука к другому только при условии, что второй побег обезлиствен. Если же побеги с листьями подвергаются воздействию разных фотопериодов, то луковицу формирует, лишь побег, находившийся на длинном дне. При образовании луковиц фотпериодический стимул, как при клубнеобразовании и цветении, по-видимому, является комплексом фитогормонов. Так, формирование луковицы сопровождается уменьшением активности ауксинов, а повышение из концентрации ведет к росту листьев и луковичных чешуй в длину. Однако вегетативное размножение луковичных растений связано не столько с формирование луковицы, сколько с образованием на ней пазушных побегов – будущих дочерних луковичек, «деток». Они закладываются в период зимнего покоя луковиц. Обнаружено, что под действием кинетина, ГА3 и ИУК, нанесенных на донце луковицы, значительно усиливается образование почек, формирующих дочерние луковички. Тот же эффект наблюдается при повышении температуры зимнего хранения луковиц или при удалении цветоносного побега. Прорастание клубней и луковиц происходит по истечении периода покоя. У луковичных растений в отличии от семян и почек, ростовые процессы полностью не прекращаются. В период покоя, хотя и очень медленно, формируются новые органы. Выход из состояния окоя ускоряется после обработки луковиц низкой температурой, т.е. после яровизации. Состояние покоя луковиц поддерживается высоким содержанием в них АБК, а при прорастании снижается уровень ингибиторов и повышается содержание ауксинов, цитокининов и гиббериллинов.
Знание принципов регуляции состояния покоя клубней и луковиц позволяет предотвращать их прорастание в период хранения, например лука и картофеля, с помощью многих ингибиторов роста, таких, как гидразид малеиновой кислоты [1].
Регуляция цветения у покрытосеменных растений.
В любом онтогенезе происходит постепенная реализация генетической информации, заключенной в ДНК. Молекулярные основы наследственной информации, связанные с редупликацией ДНК (хранением), транскрипцией (передачей) и трансляцией (реализацией) генетической информации, очень универсальны и обнаруживают большую общность во всем мире растительных организмов. В то же время выявляется широкое разнообразие видовой и фазовой специфичности регуляции пути от ДНК до белка, которая, в частности, зависит от условий среды.
Соответственно, кроме фотопериодической реакции, регуляторными механизмами развития являются температурные и возрастные факторы. Все эти условия не исключают, а дополняют друг друга.
В результате световых и темновых реакций фотопериодизма синтезируются гормональные регуляторы цветения. В результате этих преобразований в точках роста происходят изменения вегетативного морфогенеза на репродуктивный. В случае, если происходит блокировка любого из звеньев ДНК – РНК - белок, цветение не наступает.
Рецепторная роль листа в фотопериодической реакции заставляет предполагать, что у всех растений это свойство связано с каким-то фундаментальным процессом. Эту общую основу ищут в нескольких направлениях, а именно в роли фитохрома или эндогенного циркадного (суточного) ритма или отводят основную роль окислительно- восстановительным процессам листа, а именно взаимодействию процессов фотосинтеза и дыхания.
По этому воззрению, на свету получают большое значение процессы, связанные с восстановлением СО2 и фотофосфолирированием, а в темноте – с окислительным фосфорилированием.
Таким образом, в листе происходит функциональное взаимодействие митохондрий и хлоропластов, регулируемое светом и темнотой, сопряженное с биосинтезом гормональных веществ.
Таково дальнейшее течение гормональной концепции развития растений, многие звенья которой еще пока гипотетичны. Все опыты по влиянию фитогормонов и других физиологически активных веществ представляет большой интерес с точки зрения того, что развитие растений обусловлено исторически в эволюционном процессе, однако проявляются различно, в зависимости от внешних условий, в связи с приспособлением растений к окружающим условиям, т.е. к длине дня, температуре и т.д.[4]
С. С. Шаин экспериментально доказал, что при посеве растений длинного дня радами с севера на юг, а короткого дня- с востока на запад они лучше развиваются и дают больший урожай. Растения, высеянные рядами с севера на юг, лучше освещаются, потому что лучи падают перпендикулярно к ним в утренние и вечерние часы, когда сравнительно больше длинноволновой фракции спектра. В полдень, когда в солнечном свете сравнительно много коротковолновых лучей, растения в рядах затеняют друг друга. Следовательно, посевы с рядами, расположенными с востока на запад, получают больше коротковолновых лучей в течении дня, чем при посеве с севера на юг.
В опятах лаборатории фотосинтеза Украинской сельскохозяйственной академии при пунктирных посевах кукурузы с ориентацией рядов с севера-востока на юго-запад КПД фотосинтеза за вегетационный период повышается на 0,15-0,21% по сравнению с ориентацией рядов с севера на юг. Урожайность зерна кукурузы при этом возросла на 10-13%. Современные методы измерения фотосинтетически активной радиации показали, что при высоте солнца над горизонтом 20о и в течении дня спектральный состав ФАР почти не изменяется. Кроме того, большая продуктивность посевов с определенной ориентацией рядов обуславливается не только условиями освещения. При разной ориентации рядов растений условия корневого питания и водного режима также будут неодинаковыми, что, в свою очередь, влияет на поглощение и использование ФАР.
Повышенную продуктивность посевов с определенной ориентацией рядов канадский ученый У. Питтман объясняет реакцией корневой системы на направленность магнитных силовых линий Земли. В результате этого создаются лучшие условия растений. Направленность рядов имеет большое значение при посевах и посадках лесных пород, а также при уборках, уходе за молодняком. В южных широтах и в условиях резко выраженного климата широтное распоряжение рядов деревьев имеет положительное значение, так как лучше обеспечивается их взаимная защита от избыточной радиации и перегрева. В северных широтах более эффективно меридиальное расположение рядов, что способствует лучшему прогреванию почвы в междурядьях[5].
Заключение
Под контролем фотопериода находятся самые разнообразные процессы. Так, у многих растений от длины дня зависит перестройка работы меристемы побега и образование цветков. При определенной длине дня возникают и почки у многих деревьев. Земляника образует расселительные побеги - усы - при наступлении длинных дней. Сбрасывание листьев у репчатого лука и тюльпанов происходит на длинном дне, а у березы и осины - на коротком. Образование запасающих органов (кочанов у капусты, клубней у диких видов картофеля и топинамбура) также зависит от длины дня. Растения пустынь могут изменять листья С-3 типа на С-4 и наоборот, а водные растения - сменяют подводные листья на надводные, получая из внешней среды фотопериодический сигнал. Из всего выше сказанного понятно что, фотопериодизм играет очень важную роль в жизни растений.
Литература
Полевой В.В. «Физиология растений». М.: Высшая школа. 1989.
Э.Ф. Шабельская. «Физиология растений». Минск: Высшая школа, 1987.
Н.И. Якушкина. «Физиология растений». М.: Просвещение, 1980.
П.А. Генкель. «Физиология растений». М.: Просвещение, 1975.
С.И. Лебедев «Физиология растений». М.: ВО «Агропромиздат» 1988.
Р. Ван Дер Вин, Г. Мейер «Свет и рост растений». М., 1962 г.
Рубин Б.А. «Курс физиологии растений». М.: Москва, 1982 г.
Чайлахян М.Х. «Фотопериодическая и гормональная регуляция клубнеобразования у растений». М.: 1984 г.
doc4web.ru
Экофизиологическая роль фотопериодизма у растений
жению интенсивности дыхания и уменьшению относительной массы нефотосинтезирующих тканей, увеличению размеров хлоропластов и концентрации хлорофилла. Кроме того, в листьях наблюдается слабая дифференцировка на столбчатый и губчатый мезофилл или таковая совсем отсутствует, отмечается сравнительно малое количество устьиц и т. д.
Таким образом, способность воспринимать длину дня и реагировать на нее широко распространена в мире живых существ. Это означает, что живые организмы способны ориентироваться во времени, т. е. они обладают биологическими часами. Другими словами, для многих организмов характерна способность ощущать суточные, приливные, лунные и годичные циклы, что позволяет им заранее готовиться к предстоящим изменениям среды.
Правильно подобрав режимы освещения, температуры и другие факторы, наиболее соответствующие биоритмам, можно заметно повысить жизнедеятельность и продуктивность разводимых животных и растений, причем без каких-либо дополнительных затрат. Например, благодаря увеличению в теплицах, оранжереях и парниках светового дня до 12-15 ч зимой выращивают овощные культуры и декоративные растения, ускоряют рост и развитие рассады. Продлив за счет искусственного освещения световой период зимой, можно увеличить яйценоскость кур, уток, гусей, регулировать размножение пушных зверей на зверофермах.
Влияние качества света на развитие.
Много внимания было уделено исследователями качеству света, необходимого для цветения растений. Красный свет воспринимается растениями как свет в процессе развития, а сине-фиолетовая часть спектра - как темнота. Однако при увеличении интенсивности синего света он тоже начинает восприниматься растениями как свет. Опыты показывают, что если выровнять интенсивность света по числу квантов, то красный и синий свет становиться одинаково эффективным.
Важно выяснить в каких условиях проходят соответствующие изменения у растений короткого и длинного дня. Как показали опыты, во время пребывания растения в темноте на небольшой отрезок времени, в несколько минут, прервать темноту, то цветения не наступает. Того же самого можно достигнуть, давая этим растениям мигающий свет, где интервалы между светом равны секундами или минутами. У развития и важным является лишь то, чтобы в сумме число освещения соответствовало числу часов, характерному для длинного дня [4].
фотопериодизм растение свет
2. Локализация фотопериодических реакций
Фотопериодические воздействия воспринимаются главным образом листьями, а не апексами побега. У большинства растений максимальной чувствительность к фотопериоду обладают листья, только что прекратившие рост. Для дурнишника, зацветание которого индуцируется одним короткодневным фотопериодом, для фотоиндукции зацветания достаточно нескольких квадратных сантиметров листа. Основную роль в восприятии фотопериода листом играет фитохром. Предполагается, что его темновое превращение может служить способом измерения времени по типу песочных часов. Вспышка красного света устанавливает фазу эндогенного ритма.
Необходимость большой поверхности листьев и достаточной интенсивности света для перехода к цветению у многих растений объясняется потребностью растущих меристем в ассимилятах. Устранение СО2 из воздуха во время светового периода снижает инициацию цветения у некоторых короткодневных растений и длиннодневных растения.
У фотопериодически нейтральных растений переход к зацветанию обеспечивается возрастным изменениям.
Таким образом, сущность фотопериодической реакции заключается в том, что циклическое чередование света и темноты переводит растение из вегетативного в репродуктивное состояние.
Фотопериодизм, представляет собой приспособительную реакцию, позволяющую растениям зацветать в определённое, наиболее благоприятное время года. Как правило, длиннодневные растения северные, а короткодневные - южные. Для короткодневных растений более благоприятны повышенные ночные температуры, тогда как для длиннодневных - пониженные. Фотопериодическая реакция затрагивает не только процесс развития растений, но и вызывает некоторые изменения ростовых процессов.
Растения сильно различаются по числу фотопериодических циклов (числу суток с определённой длинной дня), которые вызывают затем переход к цветению одним растениям достаточно одного цикла, иначе говоря, воздействия определенной длинной дня в течение одних суток. Другим растениям необходимо получить определенную длину дня в течении 25 суток (25 циклов). Таким образом, фотопериодическое воздействие необходимо растительному организму лишь на протяжении определенного периода, после чего растение зацветает уже при любом соотношении дня и ночи. Это явление называют фотопериодической индукцией.
Исследования, проведенные на растениях короткого дня, показали, что для их перехода к образованию репродуктивных органов (цветению) важна длительность не дня, а ночи. Иначе говоря, для короткодневных растений необходим теневой период определенной длины. Если в середине темнового периода дать хотя бы вспышку света, то растение короткого дня к цветению уже не переходит. Так, если 14-часовая ночь прерывается освещением на 10 мин, то короткодневные растения не зацветают. Для длиннодневных растений нужен именно длинный световой период.
Фотопериодическое воздействие вызывается светом малой интенсивности. Так, достаточно прервать темновой период суток в
www.studsell.com









