
Содержание
а водоросли; б мхи; в плауны; г хвощи
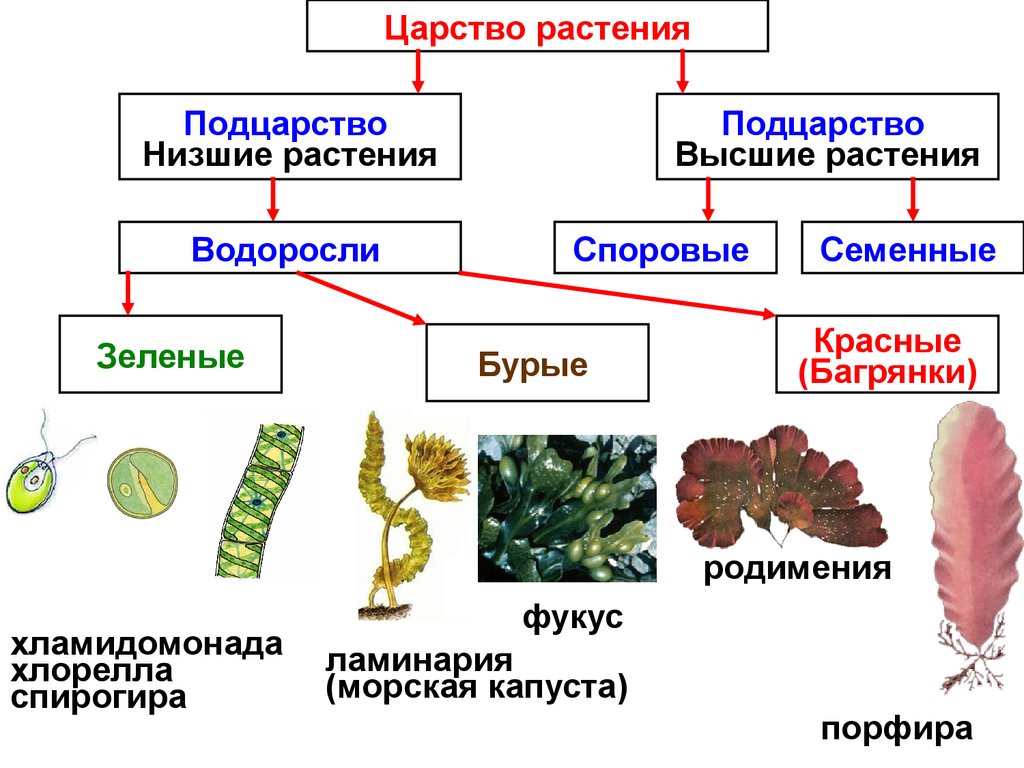
К низшим споровым растениям относятся: а водоросли; б мхи; в плауны; г хвощи
1 2 3 4 Связанные:
1 2 3 4 База данных защищена авторским правом ©psihdocs.ru 2022 | Главная страница Автореферат Анализ Анкета Бизнес-план Биография Бюллетень Викторина Выпускная работа Глава Диплом Дипломная работа |


Распределение мономеров лигнина и эволюция лигнификации среди низших растений
. 2011 Январь; 13 (1): 59-68.
doi: 10.1111/j.1438-8677.2010.00345.x.
Ж. М. Эспиньейра
М. Эспиньейра
1
, E Novo Uzal, LV Gómez Ros, J S Carrión, F Merino, A Ros Barceló, F Pomar
Филиалы
принадлежность
- 1 Кафедра биологии животных, биологии растений и экологии Университета Ла-Коруньи, Ла-Корунья, Испания.
PMID:
21143726
DOI:
10.1111/j.1438-8677.2010.00345.x
Дж. М. Эспиньера и соавт.
Растение Биол (Штутт).
2011 9 января0003
. 2011 Январь; 13 (1): 59-68.
doi: 10.1111/j.1438-8677. 2010.00345.x.
2010.00345.x.
Авторы
Ж. М. Эспиньейра
1
, Э. Ново Узал, Л. В. Гомес Рос, Х. С. Каррион, Ф. Мерино, А. Рос Барсело, Ф. Помар
принадлежность
- 1 Кафедра биологии животных, биологии растений и экологии, Университет Ла-Коруньи, Ла-Корунья, Испания.
PMID:
21143726
DOI:
10.1111/j.1438-8677.2010.00345.x
Абстрактный
Путем применения химического, биохимического и гистохимического анализов получены новые данные об отсутствии/наличии сирингиллигнинов у водорослей видов Mastocarpus stellatus, Cystoseira baccata и Ulva Rigida, мохообразных Physcomitrella patens и Marchantia polymorpha, плауновидных Selaginella martensii, Isoetes fluitans. и Isoetes histrix, сфенофит Equisetum telmateia, папоротники Ceratopteris thalictroides, Ceratopteris cornuta, Pteridium aquilinum, Phyllitis scolopendrium и Dryopteris affinis и покрытосеменное Posidonia oceanica. Лигнины, и особенно сирингиллигнины, распространяются от несосудистых базальных наземных растений, таких как печеночники, до плауновидных и папоротников. Это распределение, наряду с уже сообщенным присутствием сирингил-лигнинов в гинкгупсидах, предполагает, что сирингил-лигнин является примитивным признаком в эволюции наземных растений. Здесь мы обсуждаем, был ли путь рекрутирования синапилового спирта итеративным во время эволюции наземных растений или, альтернативно, он был включен в самые ранние наземные растения и впоследствии репрессирован у некоторых базальных печеночников, плаунов, эквизетопсид и папоротников. Эта последняя гипотеза, которая подтверждается недавними исследованиями транскрипционной регуляции биосинтеза лигнинов, подразумевает, что лигнификация возникла как активатор развития в периферических тканях протрахеофитов и только позже была использована для усиления трахеид у эвтрахеофитов.
и Isoetes histrix, сфенофит Equisetum telmateia, папоротники Ceratopteris thalictroides, Ceratopteris cornuta, Pteridium aquilinum, Phyllitis scolopendrium и Dryopteris affinis и покрытосеменное Posidonia oceanica. Лигнины, и особенно сирингиллигнины, распространяются от несосудистых базальных наземных растений, таких как печеночники, до плауновидных и папоротников. Это распределение, наряду с уже сообщенным присутствием сирингил-лигнинов в гинкгупсидах, предполагает, что сирингил-лигнин является примитивным признаком в эволюции наземных растений. Здесь мы обсуждаем, был ли путь рекрутирования синапилового спирта итеративным во время эволюции наземных растений или, альтернативно, он был включен в самые ранние наземные растения и впоследствии репрессирован у некоторых базальных печеночников, плаунов, эквизетопсид и папоротников. Эта последняя гипотеза, которая подтверждается недавними исследованиями транскрипционной регуляции биосинтеза лигнинов, подразумевает, что лигнификация возникла как активатор развития в периферических тканях протрахеофитов и только позже была использована для усиления трахеид у эвтрахеофитов.
© 2010 Немецкое ботаническое общество и Королевское ботаническое общество Нидерландов.
Похожие статьи
Присутствие синапиллигнина в культурах клеток гинкго двулопастного меняет наши взгляды на эволюцию биосинтеза лигнина.
Узал Э.Н., Гомес Рос Л.В., Помар Ф., Берналь М.А., Парадела А., Албар Х.П., Рос Барсело А.
Узал Э.Н. и соавт.
Завод Физиол. 2009 февраль;135(2):196-213. doi: 10.1111/j.1399-3054.2008.01185.x. Epub 2008 26 ноября.
Завод Физиол. 2009.PMID: 19055540
Структурные мотивы сирингилпероксидаз предшествуют не только дивергенции голосеменных и покрытосеменных, но и радиации трахеофитов.
Гомес Рос Л.В., Габальдон С., Помар Ф., Мерино Ф., Педреньо М.А., Барсело АР.

Гомес Рос Л.В. и соавт.
Новый Фитол. 2007;173(1):63-78. дои: 10.1111/j.1469-8137.2006.01898.х.
Новый Фитол. 2007.PMID: 17176394
Структурные мотивы сирингилпероксидаз сохраняются в ходе эволюции покрытосеменных.
Гомес Рос Л.В., Аснар-Асенсио Г.Дж., Эрнандес Х.А., Берналь М.А., Нуньес-Флорес М.Дж., Куэльо Х., Рос Барсело А.
Гомес Рос Л.В. и соавт.
J Agric Food Chem. 2007 г. 16 мая; 55 (10): 4131-8. doi: 10.1021/jf063164t. Epub 2007, 18 апреля.
J Agric Food Chem. 2007.PMID: 17439232
[Достижения в исследованиях биосинтеза лигнина и его генной инженерии].
Чжао Х.И., Вэй Д.Х., Сун Ю.Р.
Чжао ХИ и др.
Чжи Ву Шэн Ли Ю Фен Цзы Шэн Ву Сюэ Сюэ Бао. 2004 авг; 30 (4): 361-70.
Чжи Ву Шэн Ли Ю Фен Цзы Шэн Ву Сюэ Сюэ Бао. 2004.PMID: 15627683
Рассмотрение.
Китайский язык.Лигнификация клеточных стенок растений.
Рос Барсело А.
Рос Барсело А.
Int Rev Cytol. 1997; 176:87-132.
Int Rev Cytol. 1997.PMID: 9394918
Рассмотрение.

 2004.
2004.Посмотреть все похожие статьи
Цитируется
Глубинное происхождение и постепенная эволюция транспортных тканей: взгляд со всех сторон наземных растений.
Вуденберг С., Ренема Дж., Томеску А.М.Ф., Де Рибель Б., Вейерс Д.
Вауденберг С. и соавт.
Завод Физиол. 2022 авг 29;190(1):85-99. doi: 10.1093/plphys/kiac304.
Завод Физиол. 2022.PMID: 35904762
Бесплатная статья ЧВК.Методы флуоресцентной микроскопии для анализа и характеристики лигнина.
Маседа А, Террасас Т.
Маседа А. и др.
Полимеры (Базель). 2022 28 февраля; 14 (5): 961. doi: 10.3390/polym14050961.
Полимеры (Базель). 2022.PMID: 35267784
Бесплатная статья ЧВК.Рассмотрение.
Структурное выяснение загадочных альгицидов в морских водоросль-бактериальных симбиозах с помощью ЯМР-спектроскопии и MicroED.
Пак Д.Д., Ли И., Мун К., Хан Э.Дж., Ли С.Р., Сейедсаямдост М.Р.
Парк Джей Ди и др.
Angew Chem Int Ed Engl. 2022 21 января; 61 (4): e202114022. doi: 10.1002/anie.202114022. Epub 2021 10 декабря.
Angew Chem Int Ed Engl. 2022.PMID: 34852184
Бесплатная статья ЧВК.Новое понимание структур, образующих лигниноподобные фракции растений-предков.
Ренкорет Х.
, Гутьеррес А., Маркес Г., Дель Рио Х.С., Тобимацу Ю., Лам П.Ю., Перес-Боада М., Руис-Дуэньяс Ф.Дж., Барраса Х.М., Мартинес А.Т.
Ренкорет Дж. и соавт.
Фронт завод науч. 2021 7 окт;12:740923. doi: 10.3389/fpls.2021.740923. Электронная коллекция 2021.
Фронт завод науч. 2021.PMID: 34691117
Бесплатная статья ЧВК.Филогенетическое возникновение фенилпропаноидного пути и биосинтеза лигнина в растениях.
Яо Т., Фэн К., Се М., Баррос Дж., Чаплински Т.Дж., Тускан Г.А., Мучеро В., Чен Дж.Г.
Яо Т. и др.
Фронт завод науч. 2021 17 авг;12:704697. doi: 10.3389/fpls.2021.704697. Электронная коллекция 2021.
Фронт завод науч. 2021.PMID: 34484267
Бесплатная статья ЧВК.Рассмотрение.
 , Гутьеррес А., Маркес Г., Дель Рио Х.С., Тобимацу Ю., Лам П.Ю., Перес-Боада М., Руис-Дуэньяс Ф.Дж., Барраса Х.М., Мартинес А.Т.
, Гутьеррес А., Маркес Г., Дель Рио Х.С., Тобимацу Ю., Лам П.Ю., Перес-Боада М., Руис-Дуэньяс Ф.Дж., Барраса Х.М., Мартинес А.Т.Просмотреть все статьи «Цитируется по»
Типы публикаций
термины MeSH
вещества
Мохообразные
Коллекция мохообразных в гербарии Трейси состоит из трех основных источников.
 Во-первых, объединение гербария Департамента биологии (TAMU) добавило около 2800 образцов в результате слияния TAES-TAMU в 2012 году. внес значительный вклад. Баланс коллекции был построен за счет обмена, подарков и частого депонирования образцов отдельными лицами и группами, участвовавшими в бриофлористических исследованиях по всему штату.
Во-первых, объединение гербария Департамента биологии (TAMU) добавило около 2800 образцов в результате слияния TAES-TAMU в 2012 году. внес значительный вклад. Баланс коллекции был построен за счет обмена, подарков и частого депонирования образцов отдельными лицами и группами, участвовавшими в бриофлористических исследованиях по всему штату.
Бриология, как определено в словаре Вебстера, — это раздел ботаники, который занимается [изучением] мохообразных. Термин «мохообразный» произошел от греческого языка и относится к «растениям, которые набухают при увлажнении».
Эта способность набухать при намокании определенно намекает на способность этих организмов восстанавливать свой прежний рост после высыхания, что является ключевой особенностью физиологической экологии мохообразных. Часто рассматриваемые как «бедные родственники» более ярких покрытосеменных или как «низшие растения», эти характеристики, несомненно, намекают на их раннее эволюционное появление. Мохообразные часто объединяют в свободную группу организмов, называемых криптогамами. Криптогамы ( kryptos, означает «скрытый» и gameein, означает «жениться») представляют собой разрозненное скопление организмов, таких как водоросли, лишайники, грибы и папоротники, большинство из которых в лучшем случае отдаленно связаны с другими членами искусственной группы. Таким образом, этот термин в значительной степени устарел в современной систематике растений, хотя он действительно иллюстрирует игнорируемый статус этих организмов.
Криптогамы ( kryptos, означает «скрытый» и gameein, означает «жениться») представляют собой разрозненное скопление организмов, таких как водоросли, лишайники, грибы и папоротники, большинство из которых в лучшем случае отдаленно связаны с другими членами искусственной группы. Таким образом, этот термин в значительной степени устарел в современной систематике растений, хотя он действительно иллюстрирует игнорируемый статус этих организмов.
Мохообразные в традиционном понимании включают Bryophyta (мхи), Marchantiophyta (печеночники) и Anthocerotophyta (роголистники).
Все эти группы являются несосудистыми растениями со сходными привычками и часто занимают одни и те же местообитания. Хотя эти группы регулярно объединяются в единую группу из-за общего отсутствия сложности и схожих стратегий жизненного цикла, у этих групп очень мало общего и, вероятно, они отражают разные наследственные линии. Как несосудистые растения мохообразные обычно считаются первыми наземными растениями. Мишлер и др. (1994) пришли к выводу, что Charophyceae, одна из четырех основных линий водорослей, являются предками всех существующих наземных растений, включая мохообразные и трахеофиты. Однако размещение групп мохообразных в этой базовой линии еще не достигло окончательного консенсуса. Обобщая отношения между мохообразными по отношению к их предкам (харофитам) и их потомкам (трахеофитам), можно предположить, что сходство гипотез ставит роголистники в качестве наиболее базовой группы, а мхи и печеночники — в монофилетическую группу, сестринскую по отношению к трахеофитам.
Мишлер и др. (1994) пришли к выводу, что Charophyceae, одна из четырех основных линий водорослей, являются предками всех существующих наземных растений, включая мохообразные и трахеофиты. Однако размещение групп мохообразных в этой базовой линии еще не достигло окончательного консенсуса. Обобщая отношения между мохообразными по отношению к их предкам (харофитам) и их потомкам (трахеофитам), можно предположить, что сходство гипотез ставит роголистники в качестве наиболее базовой группы, а мхи и печеночники — в монофилетическую группу, сестринскую по отношению к трахеофитам.
Мохообразные как группа, насчитывающая от 15 000 до 25 000 видов во всем мире, являются второй по величине группой растений.
Они встречаются на всех континентах и в каждой среде обитания, где произрастают сосудистые растения. Фактически мохообразные являются доминирующими растениями по видовому разнообразию и биомассе в ряде регионов мира. Известно, что на антарктическом континенте сохранились только три вида сосудистых растений: Deschampsia antarctica Desvaux (антарктическая злаковая трава), Poa annua Linnaeus (мятлик однолетний) и Colobanthus quitnsis (Kunth) Bartling (антарктическая мшанка), остальная флора состоит из мхов и печеночников.