Хромосомный механизм определения пола. Хромосомное определение пола у растений
Хромосомный механизм определения пола
Хромосомный механизм определения пола
Генетика пола. Наследование, сцепленное с полом.
Цель: сформировать у учащихся представление о генетике пола, наследовании признаков, сцепленных с полом.
Задачи:
1. Образовательные: сформировать понятия: аутосомы, гетерохромосомы, гомогаметный, гетерогаметный пол, сформировать представление о детерминации развития пола, признаках, сцепленных с полом, признаках наследуемых через Y-хромосому и Х-хромосому; познакомить учащихся с особенностями наследования половых хромосом, некоторыми патологическими состояниями человека, наследуемыми сцепленно с полом.
2. Развивающие: продолжить формирование умений и навыков решения генетических задач на сцепленное наследование генов, на наследование, сцепленное с полом, развивать мыслительные операции.
3. Воспитательные: формировать сознательное отношение к своему здоровью и здоровью потомков.
Оборудование: компьютер, мультимедиапроектор, экран (интерактивная доска), презентация в Power Point.
Тип урока:урок изучения новой темы.
Генетика пола
Генетика объяснила сущность удивительной и важной проблемы: равное распределение женских и мужских особей в поколениях животных и людей
· Для какого способа размножения характерно образование гамет? Половое
· Какой набор хромосом они имеют? n
· Как называется оплодотворенная яйцеклетка, и какой набор хромосом она имеет? Зигота, 2n
Для начала вспомним, что представляет собой хромосомный набор клеток человека.
В кариотипе человека состоит из скольких хромосом? из 46 хромосом
44 одинаковы у всех особей, независимо от пола (эти хромосомы называют аутосомами), а одной парой хромосом, называемых половыми, женщины отличаются от мужчин. Это общебиологическая закономерность для всех живых организмов, размножающихся половым путем.
Аутосомы – парные хромосомы, одинаковые и для мужских и женских организмов.
Половые хромосомы – хромосомы, набор которых отличает мужские и женские особи у животных и растений с хромосомным определением пола.
Диплоидная клетка организма человека: 46 хромосом =23 пары гомологичных хромосом, из которых 22 пары - аутосомы + 1 пара половые хромосомы:
· Как обозначаются половые хромосомы? у мужчины - ХY; у женщины - ХХ.
Пол можно рассматривать как один из признаков организма, как правило, определяется генами. Механизм же определения пола имеет иной характер - хромосомный.
Хромосомный механизм определения пола
Пол будущего потомка определяется сочетанием половых хромосом. Пол, имеющий одинаковые половые хромосомы, называют гомогаметным, так как он дает один тип гамет, а имеющий разные-гетерогаметным, так как он образует два типа гамет. У человека, млекопитающих, мухи дрозофилы гомогаметный пол женский, а гетерогаметный - мужской. Гетерогаметный женский у птиц, рептилий
· У мужского пола в процессе гаметогенеза формируется 2 типа гамет в равной пропорции, так как мужской пол - гетерогаметный: Х-сперматозоиды и Y-сперматозоиды.
· Поскольку у женского пола половые хромосомы одинаковы, так как женский пол - гомогаметный, то каждая яйцеклетка несет Х-хромосому.
Теоретически соотношение полов должно быть 1:1. Эта статистическая закономерность, обеспечиваемая условием равновероятной встречи гамет. Статистически так и получается.
·
Как думаете, какой гомогаметный или гетерогаметный пол будет определять пол?Пол будущего организма всегда определяет гетерогаметный пол(т.е. мужской), именно потому, что гаметы с Х- и Y- хромосомой образуются у мужского пола в равных количествах.
X- и Y-хромосомы отличаются по строению: Y-хромосома состоит как бы из двух участков - одного гомологичного Х-хромосоме, а другого негомологичного. А так же по набору генов, которые в них находятся.
studopedia.ru>
Хромосомный механизм определения пола
ПОЛ – это совокупность признаков и свойств организма, обеспечивающих его участие в воспроизводстве потомства и передача наследственной информации за счет образования гамет.
Самец и самка имеют закономерное различие, касающееся одной пары хромосом. Они называются ГЕТЕРОХРОМОСОМАМИ (половыми хромосомами). Остальные пары – АУТОСОМАМИ.
Пол, имеющий одинаковые половые хромосомы (ХХ) и образующий один тип гамет называется ГОМОГАМЕТНЫМ. Пол с разными половыми хромосомами, образующий два типа гамет, называется ГЕТЕРОГАМЕТНЫМ. Гетерогаметный пол бывает двух типов:
1. ХО (нет У хромосомы) – тип Protenor
2. ХУ – тип Lygaeus
Гетерогаметным может быть женский (птицы, пресмыкающиеся, бабочки) и мужской пол.
СИНГАМНОЕ определение пола происходит в момент слияния гамет в процессе оплодотворения, характерно для организмов с гетерогаметным мужским полом (человек, животные, большинство растений).
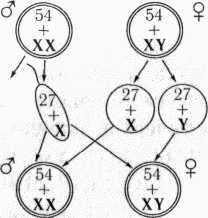
Пол потомка зависит от того, какой спермий оплодотворит яйцеклетку:
| Р: | ♀ ХХ | Х | ♂ ХУ |
| G: | Х Х | Х У | |
| яйцеклетки | спермии | ||
| F1: | 2ХХ | : | 2ХУ |
| Самки | : | Самцы | |
ПРОГАМНОЕ определение пола происходит в процессе созревания яйцеклеток при овогенезе, характерно для организмов с гетерогаметным женским полом (птицы, пресмыкающиеся, бабочки). Пол будущего потомка зависит от типа яйцеклетки: если яйцеклетка содержит Х-хромосому, то из нее после оплодотворения развивается самец, если яйцеклетка содержит У-хромосому, то из нее после оплодотворения развивается самка.
ЭПИГАМНОЕ определение пола является нехромосомным и происходит после оплодотворения в процессе индивидуального развития организма под влиянием условий внешней среды, характерно для организмов, у которых отсутствуют половые хромосомы и гены, отвечающие за половые признаки, распределены по всему генотипу (некоторые животные, морской червь Bonellia).
ЦИТОГЕНЕТИЧЕСКИЙ МЕТОД определения пола заключается в исследовании наличия полового хроматина (тельца Барра) в неделящихся соматических клетках слизистой оболочки щеки (буккальный соскоб) или на мазках крови в ядрах нейтрофиллоцитов ("барабанные палочки"). Он присутствует только у женщин (в норме).
studopedia.ru>
Хромосомный механизм определения пола
ПОЛ– это совокупность признаков и свойств организма, обеспечивающих его участие в воспроизводстве потомства и передача наследственной информации за счет образования гамет.
Самец и самка имеют закономерное различие, касающееся одной пары хромосом. Они называются ГЕТЕРОХРОМОСОМАМИ (половыми хромосомами). Остальные пары – АУТОСОМАМИ.
Пол, имеющий одинаковые половые хромосомы (ХХ) и образующий один тип гамет называется ГОМОГАМЕТНЫМ. Пол с разными половыми хромосомами, образующий два типа гамет, называется ГЕТЕРОГАМЕТНЫМ. Гетерогаметный пол бывает двух типов:
1. ХО (нет У хромосомы) – тип Protenor
2. ХУ – типLygaeus
Гетерогаметным может быть женский (птицы, пресмыкающиеся, бабочки) и мужской пол.
СИНГАМНОЕ определение пола происходит в момент слияния гамет в процессе оплодотворения, характерно для организмов с гетерогаметным мужским полом (человек, животные, большинство растений).
Пол потомка зависит от того, какой спермий оплодотворит яйцеклетку:
|
Р: |
♀ ХХ |
Х |
♂ ХУ |
|
G: |
Х Х |
Х У |
|
|
яйцеклетки |
спермии |
||
|
F1: |
2ХХ |
: |
2ХУ |
|
Самки |
: |
Самцы |
|
|
1 |
1 |
ПРОГАМНОЕ определение пола происходит в процессе созревания яйцеклеток при овогенезе, характерно для организмов с гетерогаметным женским полом (птицы, пресмыкающиеся, бабочки). Пол будущего потомка зависит от типа яйцеклетки: если яйцеклетка содержит Х-хромосому, то из нее после оплодотворения развивается самец, если яйцеклетка содержит У-хромосому, то из нее после оплодотворения развивается самка.
ЭПИГАМНОЕ определение пола является нехромосомными происходит после оплодотворения в процессе индивидуального развития организма под влиянием условий внешней среды, характерно для организмов, у которых отсутствуют половые хромосомы и гены, отвечающие за половые признаки, распределены по всему генотипу (некоторые животные, морской червьBonellia).
ЦИТОГЕНЕТИЧЕСКИЙ МЕТОД определения пола заключается в исследовании наличия полового хроматина (тельца Барра) в неделящихся соматических клетках слизистой оболочки щеки (буккальный соскоб) или на мазках крови в ядрах нейтрофиллоцитов ("барабанные палочки"). Он присутствует только у женщин (в норме).
Наследование, сцеплеНноЕ с полом
Признаки, определяемые генами, находящимися в половых хромосомах, называются ПРИЗНАКАМИ, СЦЕПЛЕННЫМИ С ПОЛОМ. Это явление было открыто Морганом у дрозофилы.
У человека с У-хромосомой связано несколько аномалий, которые передаются только по мужской линии: рыбья кожа (ихтиоз), синдактилия (перепончатые пальцы), гипертрихоз (оволоснение ушной раковины). В Х-хромосоме локализуются гены, обуславливающие развитие около 200 признаков.
ДОМИНАНТНЫЕ: гипофосфатемический рахит (аномалия костей, не лечащаяся витамином "D"), гипоплазия эмали (потемнение эмали зубов).
РЕЦЕССИВНЫЕ: дальтонизм, гемофилия, подагра, дистрофия Дюшена, отсутствие потовых желез и др.
Признаки, сцепленные с Х-хромосомой по рецессиву, передаются от матерей к сыновьям, а от отцов к дочерям. Такой тип передачи получил название крест-накрестиликрисс-кросс.
Признаки, сцепленные с У-хромосомой, передаются от отца к сыну и проявляются у самцов. Такой тип передачи называется ГОЛАНДРИЧЕСКОЕ НАСЛЕДОВАНИЕ.
StudFiles.ru>
Хромосомное определение пола
Большинство животных являются раздельнополыми организмами. Пол можно рассматривать как совокупность признаков и структур, обеспечивающих способ воспроизводства потомства и передачу наследственной информации. Пол чаще всего определяется в момент оплодотворения, то есть в определении пола главную роль играет кариотип зиготы. Кариотип каждого организма содержит хромосомы, одинаковые у обоих полов, — аутосомы, и хромосомы, по которым женский и мужской пол отличаются друг от друга, — половые хромосомы. У человека «женскими» половыми хромосомами являются две Х-хромосомы. При образовании гамет каждая яйцеклетка получает одну из Х-хромосом. Пол, у которого образуются гаметы одного типа, несущие Х-хромосому, называется гомогаметным. У человека женский пол является гомогаметным. «Мужские» половые хромосомы у человека — Х-хромосома и Y-хромосома. При образовании гамет половина сперматозоидов получает Х-хромосому, другая половина — Y-хромосому. Пол, у которого образуются гаметы разного типа, называется гетерогаметным. У человека мужской пол — гетерогаметный. Если образуется зигота, несущая две Х-хромосомы, то из нее будет формироваться женский организм, если Х-хромосому и Y-хромосому — мужской.
У животных можно выделить следующие четыре типа хромосомного определения пола.
1. Женский пол — гомогаметен (ХХ), мужской — гетерогаметен (ХY) (млекопитающие, в частности, человек, дрозофила).
Генетическая схема хромосомного определения пола у человека:
Генетическая схема хромосомного определения пола у дрозофилы:
2. Женский пол — гомогаметен (ХХ), мужской — гетерогаметен (Х0) (прямокрылые).
Генетическая схема хромосомного определения пола у пустынной саранчи:
3. Женский пол — гетерогаметен (ХY), мужской — гомогаметен (ХХ) (птицы, пресмыкающиеся).
Генетическая схема хромосомного определения пола у голубя:
Яндекс.ДиректВсе объявления
4. Женский пол — гетерогаметен (Х0), мужской — гомогаметен (ХХ) (некоторые виды насекомых).
Генетическая схема хромосомного определения пола у моли:
studopedia.ru>
Механизм определения пола у человека это:
Механизм определения пола у человекаМеханизм определения пола у человека
Определение пола у человека происходит по XY-механизму (см. также Определение пола). При этом гетерогаметным полом является мужской, гомогаметным - женский. Определение пола делится на три этапа: хромосомный, гонадный и фенотипический.
Содержание
|
Два основных правила определения пола у млекопитающих.
Классическими эмбриогенетическими исследованиями установлены два правила определения пола у млекопитающих. Первое из них сформулировано в 60-х годах Альфредом Жостом на основе экспериментов по удалению зачатка будущих гонад (гонадный валик) у ранних эмбрионов кроликов: удаление валиков до формирования гонады приводило к развитию всех эмбрионов как самок. Было высказано предположение о секретировании гонадами самцов (тестисами) эффектора (Тестостерон), ответственного за маскулинизацию плодов, и предсказано наличие второго эффектора антимюллеровского гормона (MIS), непосредственно контролирующего такие анатомические преобразования. Результаты наблюдений были сформулированы в виде правила: специализация развивающихся гонад в тестис или яичник определяет последующую половую дифференциацию эмбриона. Примерно до 1959 года предполагалось, что число Х-хромосом, которое, как известно, равно двум у самок и одному у самцов, является важнейшим фактором контроля пола у млекопитающих. Однако обнаружение индивидуумов с единственной X-хромосомой, развивающихся как самки, а особей с одной Y-хромосомой и множественными X-хромосомами как самцы заставило отказаться от таких представлений. Сформулировано второе правило определения пола у млекопитающих: Y-хромосома несет генетическую информацию, требуемую для детерминации пола у самцов. Комбинация приведенных выше двух правил иногда называется принципом роста: Хромосомный пол, связанный с присутствием или отсутствием Y-хромосомы, определяет дифференциацию эмбриональной гонады, которая, в свою очередь, контролирует фенотипический пол организма. Подобный механизм определения пола называют генетическим (GSD) и противопоставляют таковому, основанному на контролирующей роли факторов внешней среды (ESD) или соотношению половых хромосом и аутосом (CSD).
Физиологическая основа гонадного уровня детерминации пола.
Физиологической основой механизма определения пола является бисексуальность эмбриональных гонад млекопитающих. В таких прогонадах одновременно присутствуют Мюллеров проток и Вольфов канал- зачатки половых путей соответственно самок и самцов. Первичная детерминация пола начинается с появления в прогонадах специализированных клеточных линий - Клетка Сертоли. В последних синтезируется предсказанный Жостом MIS, ответственный за прямое или опосредованное ингибирование развития Мюллерова протока - зачатка будущих фаллопиевых труб и матки.
Генетический механизм определения пола.
Y-хромосома человека с указанием локализации SRY-генаВ 1987 году Дэвид Пэйдж и его коллеги, исследуя мужчину XX, унаследовавшего 280 т.п.н. фрагмент Y-хромосомспецифической ДНК, и женщину XY с делецией (нехваткой), захватывающей эту область в результате обмена участками между хромосомами, казалось, обнаружили ускользающий TDF. Им оказался присутствующий в Y-хромосоме всех настоящих зверей Eutheria и расположенный в области размером 140 т.п.н. в 100 т.п.н. от границы псевдоаутосомальной области ген ZFY. Гомолог ZFY – ZFX обнаружен в X-хромосоме, причем он избегает характерной для генов, в ней локализованных, инактивации. Оба эти фактора кодируют белок, образующий структуру так называемых цинковых пальцев, обладающий ДНК-связывающей активностью, который можно рассматривать как фактор транскрипции. Дальнейший детальный анализ Y-хромосом специфических последовательностей у особей с инверсией пола ограничил поиск районом размером 35 т.п.н. и привел к обнаружению гена, рассматриваемого как истинный эквивалент классического TDF. Такой ген получил название SRY (Sex determining Region Y gene). Приведем некоторые его характеристики, заставляющие считаться с этим предположением. SRY расположен в полопределяющей области и содержит консервативный домен (HMG-бокс), кодирующий белок размером 80 аминокислотных остатков. Его активность отмечена накануне периода дифференциации прогонады в тестис - 10-12-й день эмбрионального развития у мыши и по крайней мере на этой стадии не зависит от присутствия половых клеток. Специфические точковые мутации или делеции в HMG-боксе этого гена у женщин XY приводят к инверсии пола. Перенос 14 т.п.н. фрагмента ДНК, содержащего этот ген с фланкирующими участками, в оплодотворенную яйцеклетку гомогаметной особи с помощью микроинъекции (процедура трансгенеза - переноса генов) привел к появлению самца с XX-кариотипом. Правда, у этого животного отмечен дефектный сперматогенез.
Функции гена SRY.
Домен, кодируемый HMG-боксом SRY-гена, специфически связывается с ДНК, приводя к изгибанию ее молекулы. Такая деформация структуры ДНК, индуцируемая SRY-белком или родственными ему молекулами (известно более 100 белков с HMG-доменом), может механически передаваться на расстояние и играть важную роль в регуляции транскрипции, репликации и рекомбинации. Область ДНК, в которой локализуется SRY, ответственна за кодирование двух ключевых ферментов, участвующих в дифференцировке первичной гонады по мужскому типу: ароматазы Р450, контролирующей конверсию тестостерона в эстрадиол и фактора или гормона, ингибирующего развитие протоков Миллера, который вызывает обратное их развитие и способствует дифференцировке тестикул. Также SRY участвует в процессах половой дифференцировки в тесном взаимодействии с еще одним геном, названным K.McElreavey и соавт. (1993) геном Z, функция которого в норме заключается в угнетении специфических мужских генов. В случае нормального мужского генотипа 46XY ген SRY продуцирует белок, угнетающий ген Z, и специфические мужские гены активируются. В случае нормального женского генотипа 46ХХ, при котором отсутствует SRY, ген Z активируется и угнетает специфический мужской ген, что создает условия для развития по женскому типу.
См. также
- Пол
- Половые хромосомы
- Соотношение полов
- Определение пола
Wikimedia Foundation. 2010.
dic.academic.ru>
Читайте также
zna4enie.ru
25. Генетика пола, типы определения пола.
Генетика пола
Пол характеризуется комплексом признаков, определяемых генами, расположенными в хромосомах. В клетках организма человека хромосомы составляют парные диплоидные наборы. У видов с раздельнополыми особями хромосомный комплекс самцов и самок неодинаков и различается по одной паре хромосом (половые хромосомы). Одинаковые хромосомы этой пары назвали X (икс) -хромосомой, непарную, отсутствующую у другого пола — У (игрек) -хромосомой; остальные, по которым нет различий, — аутосомами (А).
Клетки женщины содержат две одинаковые половые хромосомы, которые обозначаются XX, у мужчин они представлены двумя непарными хромосомами X и Y. Таким образом, набор хромосом мужчины и женщины отличается только одной хромосомой: хромосомный набор женщины содержит 44 аутосомы + XX, мужчины — 44 аутосомы + XY.
Во время деления и созревания половых клеток у человека образуются гаметы с гаплоидным числом хромосом: яйцеклетки, как правило, содержат 22 + Х-хромосомы. Таким образом, у женщин образуется только один тип гамет (гаметы с Х-хромосомой). У мужчин гаметы содержат 22 + X или 22 + Y хромосом, и образуется два типа гамет (гамета с Х-хромосомой и гамета с Y-хромосомой). Если при оплодотворении в яйцеклетку попадает сперматозоид с Х-хромосомой, формируется зародыш женского пола, а с Y-хромосомой — мужского пола.
Следовательно, определение пола человека зависит от наличия в мужских половых клетках — сперматозоидах, оплодотворяющих яйцеклетку, X- или Y-хромосом.
Существует четыре основных типа хромосомного определения пола:
1. Мужской пол гетерогаметен; 50% гамет несут Х-, 50% -У – хромосому например, человек, млекопитающие, двукрылые, жуки, клопы (Слайд 4).
2. Мужской пол гетерогаметен; 50% гамет несут Х-, 50%– не имеют половой хромосомы, например, кузнечики, кенгуру (Слайд 7).
3. Женский пол гетерогаметен; 50% гамет несут Х- , 50% гамет– У– хромосому, например, птицы, пресмыкающиеся, хвостатые амфибии, шелкопряд (Слайд 7).
4. Женский пол гетерогаметен; 50% гамет несут Х- , 50% не имеют половой хромосомы, например, моль.
| Человек XY | Кузнечик XO |
| Шелкопряд XY | Моль XO |
|
|
|

Наследование признаков, гены которых локализованы в половых хромосомах, называют наследованием, сцепленным с полом.
26. Генотип как целостная система. Взаимодействие генов, множественное действие генов.
Генотип как целостная система
Свойства генов. На основании знакомства с примерами наследования признаков при моно- и дигибридном скрещивании может сложиться впечатление, что генотип организма слагается из суммы отдельных, независимо действующих генов, каждый из которых определяет развитие только своего признака или свойства. Такое представление о прямой и однозначной связи гена с признаком чаще всего не соответствует действительности. На самом деле существует огромное количество признаков и свойств живых организмов, которые определяются двумя и более парами генов, и наоборот, один ген часто контролирует многие признаки. Кроме того, действие гена может быть изменено соседством других генов и условиями внешней среды. Таким образом, в онтогенезе действуют не отдельные гены, а весь генотип как целостная система со сложными связями и взаимодействиями между ее компонентами. Эта система динамична: появление в результате мутаций новых аллелей или генов, формирование новых хромосом и даже новых геномов приводит к заметному изменению генотипа во времени.
Характер проявления действия гена в составе генотипа как системы может изменяться в различных ситуациях и под влиянием различных факторов. В этом можно легко убедится, если рассмотреть свойства генов и особенности их проявления в признаках:
Ген дискретен в своем действии, т. е. обособлен в своей активности от других генов.
Ген специфичен в своем проявлении, т. е. отвечает за строго определенный признак или свойство организма.
Ген может действовать градуально, т. е. усиливать степень проявления признака при увеличении числа доминантных аллелей (дозы гена).
Один ген может влиять на развитие разных признаков — это множественное, или плейотропное, действие гена.
Разные гены могут оказывать одинаковое действие на развитие одного и того же признака (часто количественных признаков) — это множественные гены, или полигены.
Ген может взаимодействовать с другими генами, что приводит к появлению новых признаков. Такое взаимодействие осуществляется опосредованно — через синтезированные под их контролем продукты своих реакций.
Действие гена может быть модифицировано изменением его местоположения в хромосоме (эффект положения) или воздействием различных факторов внешней среды.
Взаимодействия аллельных генов. Явление, когда за один признак отвечает несколько генов (аллелей), называется взаимодействием генов. Если это аллели одного и того же гена, то такие взаимодействия называются аллельными, а в случае аллелей разных генов —неаллельными.
Выделяют следующие основные типы аллельных взаимодействий: доминирование, неполное доминирование, сверхдоминирование и кодоминирование.
Доминирование —тип взаимодействия двух аллелей одного гена, когда один из них полностью исключает проявление действия другого. Такое явление возможно при следующих условиях: 1) доминантный аллель в гетерозиготном состоянии обеспечивает синтез продуктов, достаточный для проявления признака такого же качества, как и в состоянии доминантной гомозиготы у родительской формы; 2) рецессивный аллель совсем неактивен, либо продукты его активности не взаимодействуют с продуктами активности доминантного аллеля.
Примерами такого взаимодействия аллельных генов может служить доминирование пурпурной окраски цветков гороха над белой, гладкой формы семян над морщинистой, темных волос над светлыми, карих глаз над голубыми у человека и т. д.
Неполное доминирование, или промежуточный характер наследования, наблюдается в том случае, когда фенотип гибрида (гетерозиготы) отличается от фенотипа обеих родительских гомозигот, т. е. выражение признака оказывается промежуточным, с большим или меньшим уклонением в сторону одного или другого родителя. Механизм этого явления состоит в том, что рецессивный аллель неактивен, а степень активности доминантного аллеля недостаточна для того, чтобы обеспечить нужный уровень проявления доминантного признака.
Примером неполного доминирования является наследование окраски цветков у растений ночной красавицы (рис. 3.5). Как видно из схемы, гомозиготные растения имеют либо красные (АА), либо белые (аа) цветки, а гетерозиготные (Аа) — розовые. При скрещивании растения с красными цветками и растения с белыми цветками в F1, у всех растений цветки розовые, т. е. наблюдается промежуточный характер наследования. При скрещивании гибридов с розовой окраской цветков в F2 имеет место совпадение расщепления по фенотипу и генотипу, так как доминантная гомозигота (АА) отличается от гетерозиготы (Аа). Так, в рассматриваемом примере с растениями ночной красавицы расщепление в F2 по окраске цветков обычно следующее — 1 красная (АА): 2 розовые (Аа): 1 белая (аа).

Рис. 3.5. Наследование окраски цветков при неполном доминировании у ночной красавицы.
Неполное доминирование оказалось широко распространенным явлением. Оно наблюдается в наследовании курчавости волос у человека, масти крупного рогатого скота, окраски оперения у кур, многих других морфологических и физиологических признаков у растений, животных и человека.
Сверхдоминирование — более сильное проявление признака у гетерозиготной особи (Аа), чем у любой из гомозигот (АА и аа). Предполагается, что это явление лежит в основе гетерозиса (см. § 3.7).
Кодаминирвание— участие обоих аллелей в определении признака у гетерозиготной особи. Ярким и хорошо изученным примером кодоминирования может служить наследование IV группы крови у человека (группа АВ).
Эритроциты людей этой группы имеют два типа антигенов: антиген А (детерминируемый геном /\ имеющимся в одной из хромосом) и антиген В (детерминируемый геном /а, локализованным в другой гомологичной хромосоме). Только в этом случае проявляют свое действие оба аллеля — 1А (в гомозиготном состоянии контролирует II группу крови, группу А) и IB (в гомозиготном состоянии контролирует III группу крови, группу В). Аллели 1А и IB работают в гетерозиготе как бы независимо друг от друга.
Пример наследования групп крови иллюстрирует и прояв-ление множественного аллелизма: ген/может быть представлен тремя разными аллелями, а есть гены, имеющие десятки аллелей. Все аллели одного гена получили название серии мно-жественных аллелей, из которых каждый диплоидный организм может иметь два любых аллеля (и только). Между этими аллелями возможны все перечисленные варианты аллельных взаимодействий.
Явление множественного аллелизма распространено в природе. Известны обширные серии множественных аллелей, определяющих тип совместимости при оплодотворении у грибов, опылении у семенных растений, детерминирующих окраску шерсти животных и т. д.
Взаимодействия неаллельных генов.Неаллельные взаимодействия генов описаны у многих растений и животных. Они приводят к появлению в потомстве дигетерозиготы необычного расщепления по фенотипу: 9:3:4; 9:6:1; 13:3; 12:3:1; 15:1, т.е. модификации общей менделевской формулы 9:3:3:1. Известны случаи взаимодействия двух, трех и большего числа неаллельных генов. Среди них можно выделить следующие основные типы: комплементарность, эпистаз и полимерию.
Комплементарным, или дополнительным, называется такое взаимодействие неаллельных доминантных генов, в результате которого появляется признак, отсутствующий у обоих родителей. Например, при скрещивании двух сортов душистого горошка с белыми цветками появляется потомство с пурпурными цветками. Если обозначить генотип одного сорта ААbb, а другого — ааВВ, то
Гибрид первого поколения с двумя доминантными генами (А и В) получил биохимическую основу для выработки пурпурного пигмента антоциана, вто время как поодиночке ни ген А, ни ген B не обеспечивали синтез этого пигмента. Синтез антоциана представляет собой сложную цепь последовательных биохимических реакций, контролируемых несколькими неаллельными генами, и только при наличии как минимум двух доминантных генов (А-В-) развивается пурпурная окраска. В остальных случаях {ааВ- и A-bb) цветки у растения белые (знак «—» в формуле генотипа обозначает, что это место может занять как доминантный, так и рецессивный аллель).
При самоопылении растений душистого горошка из F1 в F2 наблюдалось расщепление на пурпурно- и белоцветковые формы в соотношении, близком к 9:7. Пурпурные цветки были обнаружены у 9/16 растений, белые — у 7/16. Решетка Пеннета наглядно показывает причину этого явления (рис. 3.6).
Эпистаз — это такой тип взаимодействия генов, при котором аллели одного гена подавляют проявление аллельной пары другого гена. Гены, подавляющие действие других генов, называются эпистатическими, ингибиторами или супрессорами. Подавляемый ген носит название гипостатический.
По изменению числа и соотношения фенотип и чес ких классов при дигибридном расщеплении в F2 рассматривают несколько типов эпистатических взаимодействий: доминантный эпистаз (А>В или В>А) с расщеплением 12:3:1; рецессивный эпистаз (а>В или b>А), который выражается в расщеплении 9:3:4, и т. д.
Полимерия проявляется в том, что один признак формируется под влиянием нескольких генов с одинаковым фенотипичес-ким выражением. Такие гены называются полимерными. В этом случае принят принцип однозначного действия генов на развитие признака. Например, при скрещивании растений пастушьей сумки с треугольными и овальными плодами (стручочками) в F1 образуются растения с плодами треугольной формы. При их самоопылении в F2 наблюдается расщепление на растения с треугольными и овальными стручочками в соотношении 15:1. Это объясняется тем, что существуют два гена, действующих однозначно. В этих случаях их обозначают одинаково— А1и A2 .

Рис. 3.6. Наследование окраски цветков у душистого горошка
Тогда все генотипы (А1 ,-А2,-, А1-а2а2, a1a1A2-) будут иметь одинаковый фенотип — треугольные стручочки, и только растения а1а1а2a2 будут отличаться —- образовывать овальные стручочки. Это случай некумулятивной полимерии.
Полимерные гены могут действовать и по типу кумулятивной полимерии. Чем больше подобных генов в генотипе организма, тем сильнее проявление данного признака, т. е. с увеличением дозы гена (А1 А2 А3 и т. д.) его действие суммируется, или кумулируется. Например, интенсивность окраски эндосперма зерен пшеницы пропорциональна числу доминантных аллелей разных генов в тригибридном скрещивании. Наиболее окрашенными были зерна А1А1А2А2А3,А 3 а зерна а1а1а2a2а3а 3 не имели пигмента.
По типу кумулятивной полимерии наследуются многие признаки: молочность, яйценоскость, масса и другие признаки сельскохозяйственных животных; многие важные параметры физической силы, здоровья и умственных способностей человека; длина колоса у злаков; содержание сахара в корнеплодах сахарной свеклы или липидов в семенах подсолнечника и т. д.
Таким образом, многочисленные наблюдения свидетельствуют о том, что проявление большей части признаков представляет собой результат влияния комплекса взаимодействующих генов и условий внешней среды на формирование каждого конкретного признака.
Взаимодействие генов
Отношение между генами и признаками достаточно сложное. В организме не всегда один ген определяет только один признак и, наоборот, один признак определяется только одним геном. Чаще один ген может способствовать проявлению сразу нескольких признаков, и наоборот. Генотип организма нельзя рассматривать как простую сумму независимых генов, каждый из которых функционирует вне связи с другими. Фенотипное проявления того или иного признака являются результатом взаимодействия многих генов.
Множественное действие генов (плейотропия) — процессы влияния одного гена на формирование нескольких признаков.
Например, у человека ген, определяющий рыжую окраску волос, обусловливает более светлую кожу и появление веснушек.
Иногда гены, определяющие морфологические признаки, влияют на физиологические функции, снижая жизнестойкость и плодовитость, или оказываются летальными. Так, ген, вызывающий голубую окраску у норки, снижает ее плодовитость. Доминантный ген серой окраски у каракулевых овец в гомозиготном состоянии детален, поскольку у таких ягнят недоразвит желудок и они погибают при переходе на питание травой.
Комплементарное взаимодействие генов. На развитие одного признака могут влиять несколько генов. Взаимодействие нескольких неаллельных генов, приводящее к развитию одного признака, называется комплементарным. Например, у кур имеются четыре формы гребня, проявление какой-либо из них связано со взаимодействием двух пар неаллельных генов. Розовидный гребень обусловлен действием доминантного гена одной аллели, гороховидный — доминантного гена другой аллели. У гибридов при наличии двух доминантных неаллельных генов образуется ореховидный гребень, а при отсутствии всех доминантных генов, т.е. у рецессивной гомозиготы по двум неаллельным генам, образуется простой гребень.
Результатом взаимодействия генов является окраска шерсти у собак, мышей, лошадей, форма тыквы, окраска цветков душистого горошка.
Полимерия — такое взаимодействие неаллельных генов, когда степень развития признака зависит от общего количества доминантных генов. По этому принципу наследуется окраска зерен овса, пшеницы, цвет кожи у человека. Например, у негров в двух парах неаллельных генов 4 доминантных, а у людей с белой кожей — ни одного, все гены рецессивные. Сочетания разного количества доминантных и рецессивных генов приводят к образованию мулатов с разной интенсивностью окраски кожи: от темной до светлой.
Различают две основных группы взаимодействия генов: взаимодействие между аллельными генами и взаимодействие между неаллельнимы генами. Однако следует понимать, что это не физическое взаимодействие самих генов, а взаимодействие первичных и вторичных продуктов, которые обусловят тот или иной признак. В цитоплазме происходит взаимодействие между белками - ферментами, синтез которых опрелятся генами, или между веществами, которые образовываются под влиянием этих ферментов.
Возможны следующие типы взаимодействия:
1) для образования определенного признака необходимо взаимодействие двух ферментов, синтез которых опрелятся двумя неаллельнимы генами;
2) фермент, что был синтезирован с участием одного гена, полностью подавляет или инактивирует действие фермента, что был образован другим неаллельным геном;
3) два ферменты, образование которых контролируется двумя неаллельми генами, влияющими на один признак или на один процесс так, что их совместное действие приводит к возникновению и усилению проявления признака.
Взаимодействие аллельных генов
Гены, которые занимают идентичные (гомологические) локусы в гомологичных хромосомах, называются аллельными. У каждого организма есть по два аллельных гена.
Известны такие формы взаимодействия между аллельными генами: полное доминирование, неполное доминирование, кодоминированием и сверхдоминирование.
Основная форма взаимодействия - полное доминирование, которое впервые описано Г. Менделем. Суть его заключается в том, что в гетерозиготном организме проявление одной из аллелей доминирует над проявлением другой. При полном доминировании расщепления по генотипу 1:2:1 не совпадает с расщеплением по фенотипу - 3:1. В медицинской практике с двух тысяч моногенных наследственных болезней почти в половины имеет место доминированое проявления патологических генов над нормальными. В гетерозигот патологический аллель проявляется в большинстве случаев признаками заболевания (доминантный фенотип).
Неполное доминирование - форма взаимодействия, при которой у гетерозиготного организма (Аа) доминантный ген (А) не полностью подавляет рецессивный ген (а), вследствие чего проявляется промежуточный между родительскими признак. Здесь расщепление по генотипу и фенотипу совпадает и составляет 1:2:1
При кодоминировании в гетерозиготных организмах каждый из аллельных генов вызывает формирование зависимого от него продукта, то есть оказываются продукты обеих аллелей. Классическим примером такого проявления является система групп крови, в частности система АBО, когда эритроциты человека несут на поверхности антигены, контролируемые обеими аллелями. Такая форма проявления носит название кодоминированием.
Сверхдоминирование - когда доминантный ген в гетерозиготном состоянии проявляется сильнее, чем в гомозиготном. Так, у дрозофилы при генотипе АА-нормальная продолжительность жизни; Аа - удлиненная триватисть жизни; аа - летальный исход.
Множественный алелизм
У каждого организма есть только по два аллельных гена. Вместе с тем нередко в природе количество аллелей может быть более двух, если какой то локус может находится в разных состояниях. В таких случаях говорят о множественные аллели или множественный аллеломорфизм.
Множественные аллели обозначаются одной буквой с разными индексами, например: А, А1, А3 ... Аллельные гена локализуются в одинаковых участках гомологичных хромосом. Поскольку в кариотипе всегда присутствуют по две гомологичных хромосомы, то и при множественных аллелях каждый организм может иметь одновременно лишь по два одинаковых или различных аллели. В половую клетку (вместе с различием гомологичних хромосом) попадает только по одному из них. Для множественных аллелей характерное влияние всех аллелей на один и тот же признак. Отличие между ними заключается лишь в степени развития признака.
Второй особенностью является то, что в соматических клетках или в клетках диплоидных организмов содержится максимум по две аллели из нескольких, поскольку они расположены в одном и том же локусе хромосомы.
Еще одна особенность присуща множественным аллелям. По характеру доминирования аллеломорфные признаки размещаются в последовательном ряду: чаще нормальный, неизмененный признак доминирует над другими, второй ген ряда рецессивный относительно первого, однако доминирует над следующими и т.д. Одним из примеров проявления множественных аллелей у человека есть группы крови системы АВО.
Множественный алелизм имеет важное биологическое и практическое значение, поскольку усиливает комбинативну изменчивость, особенно генотипического.
Взаимодействие неалельних генов
Известно много случаев, когда признак или свойства детерминируются двумя или более неалельнимы генами, которые взаимодействуют между собой. Хотя и здесь взаимодействие условно, потому что взаимодействуют не гены, а контролируемые ими продукты. При этом имеет место отклонение от менделивских закономерностей расщепления.
Различают четыре основных типа взаимодействия генов: комплементарность, эпистаз, полимерию и модифицирующее действие (плейотропия).
Комплементарность это такой тип взаимодействия неаллельних генов, когда один доминантный ген дополняет действие другого неаллельного доминантного гена, и они вместе определяют новый признак, который отсутствует у родителей. Причем соответственный признак развивается только в присутствии обоих неаллельних генов. Например, сера окраска шерсти у мышей контролируется двумя генами (А и В). Ген А детерминирует синтез пигмента, однако как гомозиготы (АА), так и гетерозиготы (Аа) - альбиносы. Другой ген В обеспечивает скопления пигмента преимущественно у основания и на кончиках волос. Скрещивания дигетерозигот (АаВЬ х АаВЬ) приводит к расщеплению гибридов в соотношении 9:3:4. Числовые соотношения при комплементарном взаимодействии могут быть как 9:7; 9:6:1 (видоизменение менделивского расщепления).
Примером комплементарного взаимодействия генов у человека может быть синтез защитного белка - интерферона. Его образование в организме связано с комплементарным взаимодействием двух неаллельних генов, расположенных в разных хромосомах.
Эпистаз -это такое взаимодействие неаллельных генов, при котором один ген подавляет действие другого неаллельного гена. Угнетение могут вызывать как доминантные, так и рецессивные гены (А> В, а> В, В> А, В> А), и в зависимости от этого розличают эпистаз доминантный и рецессивный. Подавляющий ген получил название ингибитора или супрессора. Гены-ингибиторы в основном не детерминируют развитие определенного признака, а лишь подавляют действие другого гена.
Ген, эффект которого подавляется, получил название гипостатичного. При епистатичном взаимодействияи генов расщепление по фенотипу в F2 составляет 13:3; 12:3:1 или 9:3:4 и др. Окрас плодов тыквы, масть лошадей определяются этим типом взаимодействия.
studfiles.net
Хромосомный механизм определения пола
ПОЛ– это совокупность признаков и свойств организма, обеспечивающих его участие в воспроизводстве потомства и передача наследственной информации за счет образования гамет.
Самец и самка имеют закономерное различие, касающееся одной пары хромосом. Они называются ГЕТЕРОХРОМОСОМАМИ (половыми хромосомами). Остальные пары – АУТОСОМАМИ.
Пол, имеющий одинаковые половые хромосомы (ХХ) и образующий один тип гамет называется ГОМОГАМЕТНЫМ. Пол с разными половыми хромосомами, образующий два типа гамет, называется ГЕТЕРОГАМЕТНЫМ. Гетерогаметный пол бывает двух типов:
1. ХО (нет У хромосомы) – тип Protenor
2. ХУ – тип Lygaeus
Гетерогаметным может быть женский (птицы, пресмыкающиеся, бабочки) и мужской пол.
СИНГАМНОЕ определение пола происходит в момент слияния гамет в процессе оплодотворения, характерно для организмов с гетерогаметным мужским полом (человек, животные, большинство растений).
Пол потомка зависит от того, какой спермий оплодотворит яйцеклетку:
| Р: | ♀ ХХ | Х | ♂ ХУ |
| G: | Х Х | Х У | |
| яйцеклетки | спермии | ||
| F1: | 2ХХ | : | 2ХУ |
| Самки | : | Самцы | |
| 1 | 1 |
ПРОГАМНОЕ определение пола происходит в процессе созревания яйцеклеток при овогенезе, характерно для организмов с гетерогаметным женским полом (птицы, пресмыкающиеся, бабочки). Пол будущего потомка зависит от типа яйцеклетки: если яйцеклетка содержит Х-хромосому, то из нее после оплодотворения развивается самец, если яйцеклетка содержит У-хромосому, то из нее после оплодотворения развивается самка.
ЭПИГАМНОЕ определение пола является нехромосомными происходит после оплодотворения в процессе индивидуального развития организма под влиянием условий внешней среды, характерно для организмов, у которых отсутствуют половые хромосомы и гены, отвечающие за половые признаки, распределены по всему генотипу (некоторые животные, морской червьBonellia).
ЦИТОГЕНЕТИЧЕСКИЙ МЕТОД определения пола заключается в исследовании наличия полового хроматина (тельца Барра) в неделящихся соматических клетках слизистой оболочки щеки (буккальный соскоб) или на мазках крови в ядрах нейтрофиллоцитов ("барабанные палочки"). Он присутствует только у женщин (в норме).
Наследование, сцеплеНноЕ с полом
Признаки, определяемые генами, находящимися в половых хромосомах, называются ПРИЗНАКАМИ, СЦЕПЛЕННЫМИ С ПОЛОМ. Это явление было открыто Морганом у дрозофилы.
У человека с У-хромосомой связано несколько аномалий, которые передаются только по мужской линии: рыбья кожа (ихтиоз), синдактилия (перепончатые пальцы), гипертрихоз (оволоснение ушной раковины). В Х-хромосоме локализуются гены, обуславливающие развитие около 200 признаков.
ДОМИНАНТНЫЕ: гипофосфатемический рахит (аномалия костей, не лечащаяся витамином "D"), гипоплазия эмали (потемнение эмали зубов).
РЕЦЕССИВНЫЕ: дальтонизм, гемофилия, подагра, дистрофия Дюшена, отсутствие потовых желез и др.
Признаки, сцепленные с Х-хромосомой по рецессиву, передаются от матерей к сыновьям, а от отцов к дочерям. Такой тип передачи получил название крест-накрестиликрисс-кросс.
Признаки, сцепленные с У-хромосомой, передаются от отца к сыну и проявляются у самцов. Такой тип передачи называется ГОЛАНДРИЧЕСКОЕ НАСЛЕДОВАНИЕ.
studfiles.net
Хромосомное определение пола
Одним из признаков у многих живых организмов является пол. Пол регулируется несколькими механизмами. Мужской или женский пол может определяться воздействием условий окружающей среды (эпигамный способ полоопределения), как, например, у морского кольчатого червя Bonnelia, или задаваться условиями развития яйцеклеток (прогамный способ), как у коловраток и кольчатых червей.
Тем не менее у большинства организмов пол определяется хромосомным составом. В генотипе имеются специальные половые хромосомы (X и У, или Z и W), остальные же представляют собой аутосомы.
Так, у человека и других млекопитающих женские особи имеют две X хромосомы, их кариотип может быть выражен как XX+2А (А — гаплоидный набор аутосом), а мужские — XY + 2А. Женские особи являются гомогаметными, поскольку формируют лишь один тип гамет, а мужские — гетерогаметными, так как производят два типа гамет в отношении половых хромосом. Мужские организмы являются также и гемизиготными: так как у них имеется всего одна их хромосома, то почти все гены, которые находятся в ней (не только доминантные, но и рецессивные), проявляются в фенотипе. Это происходит потому, что гены X хромосомы, как правило, не имеют гомологов в У хромосоме. У хромосома несет мало генов и поэтому является генетически инертной. Так же определяется генетический пол и у большинства высших растений. У птиц и бабочек гетерогаметным полом является женский пол, а гомогаметным — мужской. У них половые хромосомы часто обозначают как Z и W. У беспозвоночных регуляция пола более разнообразна. У некоторых из них (например, у некоторых клопов) самки имеют состав половых хромосом XX, самцы же несут всего одну из них (ХО). У плодовой мушки дрозофилы принадлежность к тому или иному полу определяется балансом числа X хромосом и аутосом. При соотношении Х/А = 1 развиваются обычные женские особи; Х/А = 0,5 — нормальные мужские; если оно имеет промежуточное значение 0,5 < Х/А < 1, развиваются интерсексы, имеющие проявления признаков обоих полов. У ряда перепончатокрылых (пчелы, муравьи) особи мужского пола развиваются из неоплодотворенных (гаплоидных) яиц, а женские — из оплодотворенных диплоидных. В этом случае пол потомства регулирует размножающаяся самка.
У цветковых растений встречаются случаи, когда принадлежность к тому или иному полу контролируется единичным числом генов. Если гены, определяющие формирование тех или иных фенотипических признаков, располагаются в половых хромосомах, то их наследование будет отклоняться от описанных выше закономерностей. Такие признаки называются признаками, сцепленными с полом. Если ген находится в У хромосоме, то он наследуется по однородительскому механизму (например, у млекопитающих, включая человека, — от отца к сыну), который носит название голандрического. При нахождении гена в X хромосоме в первом поколении в одном из направлений скрещиваний будет наблюдаться наследование типа крисс-кросс (или крест-накрест), при котором признак самца проявляется у его дочерей, а признак самки — у сыновей; во втором поколении проявляется соотношение 1:1. В другом же направлении скрещиваний будут наблюдаться обычные расщепления.
www.polnaja-jenciklopedija.ru
Хромосомное определение пола
У многих организмов соотношение между особями мужского и женского пола при изучении большого числа особей всегда примерно равное, т.е. расщепление по признаку пола проходит в отношении 1:1.
У плодовой мушки дрозофилы, на которой проведено множество генетических исследований, и у млекопитающих пол определяется следующим образом. В соматических клетках дрозофилы четыре пары хромосом. В число их входит три пары аутосом, т.е. хромосом, одинаковых у самца и самки, и одна пара хромосом, различных у особей мужского и женского пола. Это хромосомы, как было установлено, отвечают за наследование пола и поэтому были названы половыми хромосомами.
В клетках самок мух дрозофил имеются две одинаковые хромосомы, которые условно обозначают как Х-хромосомы. Следовательно, в диплоидных соматических клетках самки набор половых хромосом – ХХ. У самцов половые хромосомы отличаются от половых хромосом самок. В соматических клетках самца мухи дрозофилы имеется одна Х-хромосома и одна Y-хромосома. Поэтому набор половых хромосом самца обозначается XY. Следовательно, яйцеклетки женских организмов все одинаковы по хромосомному набору, т.к. в каждой из них имеются по одинаковому набору аутосом и одна половая хромосома. Все сперматозоиды также имеют по одному набору аутосом и одну половую хромосому, но половина сперматозоидов имеет Х-хромосому, а другая половина – Y-хромосому. Различаются они по набору генов, который в ней содержится. Поскольку гаметы с Х- и У-хромосом в результате мейоза образуются у самцов в равных количествах, то ожидаемое полов составляет 1:1, что совпадает с фактическими наблюдениями. Сходный способ определения полов присущим всем млекопитающим.
Наследование, сцепленное полом
В том случае, когда гены, ответственные за формирование признака, расположены в аутосомах, наследование осуществляется независимо от того, какой из родителей является носителем изучаемого признака. Однако ситуация резко изменяется, когда признаки определяются генами, лежащих в половых хромосомах. Черепаховая окраска, т.е. чередование черных и желтых пятен, встречается только у кошек. Котов с черепаховой окраской не бывает. Этот факт не могли объяснить, пока не стало известно, что наследование данного признака сцеплено с полом.
Рис. 32. Хромосомные комплексы самца и самки у дрозофилы
Рис. 33. Черепаховая окраска кошки
Черная окраска кошек определяется геном В, рыжая – геном b. Эти гены расположены в Х-хромосоме. В У-хромосоме они отсутствуют. Обозначим Х-хромосому, несущую аллель В, ХВ, а Х-хромосому с аллелем b – Хb. Поэтому возможны такие комбинации: ХBXb – черная кошка; ХBУ – черный кот Xb – рыжая кошка; ХbУ – рыжий кот ХBХb – черепаховая кошка
Так как мужская особь имеет только одну Х-хромосому, то кот может быть или черным, или рыжим, но не может иметь черепаховую окраску, потому что для ее развития необходимо одновременное присутствие генов B и b.
У человека также известны признаки, сцепленные с полом. К ним относится, например, очень тяжелое наследственное заболевание гемофилия, при котором кровь теряет способность свертываться. У гемофиликов даже небольшие царапины и ссадины вызывают тяжелые кровотечения. Это заболевание встречается, за редчайшими исключениями, только у мужчин. Было установлено, что гемофилия обусловлена рецессивным геном, расположенным в Х-хромосоме, поэтому гетерозиготные по данному гену женщины обладают обычной свертываемостью крови. Рассмотрим, какое потомство может появиться у гетерозиготной женщины, вступившей в брак с нормальным по этому признаку мужчиной.
Ген, обуславливающий нормальную свертываемость крови, обозначим Н, а ген, при котором кровь теряет способность свертываться – h. Учитывая, что в генотипе женщины присутствуют две Х-хромосомы, а у мужчины – одна Х-хромосома и одна У-хромосома, запишем схему наследования гемофилии:
| Р | ХНXh – носительница гена гемофилии | × | ХHУ – здоровый мужчина | |
| гаметы | ХН ; Xh | ХH ; У | ||
| F1 | ХНХН – здоровая женщина | ХНXh – здоровая женщина, носительница гена гемофилии | ХHУ – здоровый мужчина | ХhУ – мужчина-гемофилик |
Женщина передает половине своих сыновей Х-хромосому с геном нормальной свертываемости крови, а половине – Х-хромосому с геном гемофилии. Среди ее сыновей могут быть и здоровые и гемофилики.
В силу равновероятного расхождения хромосом при формировании гамет и их встречи в зиготе следует ожидать, что в потомстве большого числа браков, подобных только что рассмотренному, у половины сыновей разовьется гемофилия. В то же время все дочери в любом случае получают Х-хромосому от своего отца с геном ХH, поэтому у них всегда нормальная свертываемость крови, но половина дочерей будут гетерозиготными носительницами этого заболевания.
Ген, вызывающий дальтонизм (неспособность различать красный и зеленый цвет), также сцеплен с Х-хромосомой.
Взаимодействие генов
В примерах, которые мы до сих пор рассматривали, каждый ген определял только один признак, и развитие каждого признака зависело только от одного гена. Однако, как правило, отношения между генами и признаками гораздо сложнее.
Множественное действие гена. Один и тот же ген может влиять на формирование ряда признаков организма. Например, ген, вызывающий образование бурой семенной кожуры у гороха, способствует развитию пигмента и в других частях растения. Растения с семенами, покрытыми бурой семенной кожурой, имеют цветки фиолетовой окраски, а растения с белой кожурой семян – белые цветки. Влияние одного гена на развитие многих признаков называют множественным действием гена. Такое множественное действие характерно для большинства генов. Обычно судят о действии гена только по тем признакам, которые легко обнаруживаются. Например, у мухи дрозофилы глаза имеют красную окраску, развитие которой обусловлено присутствием гена W. При наличии аллелей w пигмент, влияющий на окраску глаз, отсутствует, и они становятся белыми. Оказалось, что аллель w влияет также на осветление окраски тела и изменение формы некоторых внутренних органов мухи.
В некоторых случаях проявление множественного признака легко наблюдать. Например, у человека есть ген, определяющий рыжую окраску волос. Этот же ген обусловливает более светлую окраску кожи, а также появление веснушек. У растений гороха ген, отвечающий за красную окраску цветов, определяет и красноватый оттенок стеблей.
Отношение ген – признак. При скрещивании в результате взаимодействия двух различных генов в потомстве могут возникать новые признаки, отсутствовавшие у родительских организмов. Это явление называется новообразованием при скрещиваниях. Его часто наблюдают при разведении домашних животных и культурных растений.
Приведем пример. У душистого горошка – садового растения – есть много сортов, которые отличаются по окраске цветков, в частности, например как в сорте с белыми цветками. При некоторых комбинациях скрещивания двух белых сортов между собой полученные семена дают растения с фиолетовыми цветками. Биохимический анализ показал, что фиолетовая окраска цветков душистого горошка – результат реакции между двумя веществами, каждое из этих веществ образуется под действием определенного гена. Например, есть два гена, каждый с двумя аллелями – С, с и Р, р. Окраска цветков образуется тогда, когда в генотипе растений присутствуют одновременно два доминантных гена С и Р. Скрещивание двух сортов душистого горошка с белыми цветками, когда гибриды первого поколения имеют фиолетовые цветки, записывают следующим образом:
|
Известно много примеров, когда гены влияют на характер проявления определенного неаллельного гена или на саму возможность проявления этого гена. Пример взаимодействия двух пар генов – наследование формы гребня у кур некоторых пород. В результате различных комбинаций этих генов возникают четыре варианта формы гребня.
Рис. 34. Форма гребня у петухов (А – простой (ааbb), Б – гороховидный (ааВВ или ааВb), В – ореховидный (ААВВ или АаВb), Г – розовидный (ААbb или Ааbb))
Таким образом, мы видим, что в генотипе любого организма гены взаимодействуют между собой; на фенотипическое проявление признака влияет целый комплекс генов. Сочетания генов в организме обусловливают индивидуальные различия особей одного вида.
infopedia.su
Хромосомное определение пола
Хромосомная теория наследственности позволила ответить на вопросы, которые давно интересовали человека: чем определяется пол у раздельнополых животных? Как достигается удивительное равенство числа особей обоих полов в каждом поколении? Оказалось, что пол будущего животного определяется в момент образования зиготы. В общем наборе хромосом всех раздельнополых животных существует два типа хромосом: аутосомы и половые хромосомы. В клетках тела организма обычно бывают две половые хромосомы. Пара половых хромосом может быть представлена двумя одинаковыми хромосомами в клетке; такие половые хромосомы называются X-хромосомами. А может быть, что в клетке есть пара неодинаковых половых хромосом — X-хромосома и Y-хромосома. Пол животного зависит от того, будут ли в его клетке половые хромосомы представлены XX или XY. У большей части животных, в том числе у человека, женскому полу соответствует набор половых хромосом XX, мужскому — XY. У других животных (птицы, бабочки), наоборот, XX означает мужской пол, а XY — женский. У некоторых видов хромосома Y может отсутствовать и тогда женский пол будут определять две X-хромосомы (XX), а мужской пол — только одна X-хромосома (X0).
Не только определение пола, но и числовое равенство полов в каждом поколении вытекает из механизма гаметогенеза. Когда образуются половые клетки, диплоидный набор хромосом распределяется на два гаплоидных. При этом распределяются и половые хромосомы. Если исходная клетка имела XX, то все гаметы будут иметь по одной X-хромосоме. При этом все образовавшиеся гаметы будут одинаковы. Если исходная клетка имела XY, то половина гамет будет иметь одну X-хромосому, а половина — одну Y-хромосому. В момент оплодотворения одинаково вероятно, что сольются в одну зиготу или гаметы, имеющие по X-хромосоме, или гаметы, одна из которых несет X-хромосому, а другая — Y-хромосому. В результате зиготы XX и XY в каждом поколении образуются примерно в одинаковом количестве. А это и означает, что число самцов и самок в каждом поколении примерно одно и то же.
Updated: 21.11.2014 at 2:26 пп
jbio.ru
Хромосомное определение пола — Энциклопедия по биологии
Хромосомное определение пола. Хромосомная теория наследственности позволила ответить на вопросы, которые давно интересовали человека: чем определяется пол у раздельнополых животных? Как достигается удивительное равенство числа особей обоих полов в каждом поколении? Оказалось, что пол будущего животного определяется в момент образования зиготы. В общем наборе хромосом всех раздельнополых животных существует два типа хромосом: аутосомы и половые хромосомы. В клетках тела организма обычно бывают две половые хромосомы. Пара половых хромосом может быть представлена двумя одинаковыми хромосомами в клетке; такие половые хромосомы называются Х-хромосомами. А может быть, что в клетке есть пара неодинаковых половых хромосом — Х-хромосома и У-хро-мосома. Пол животного зависит от того, будут ли в его клетке половые хромосомы представлены XX или ХУ. У большей части животных, в том числе у человека, женскому полу соответствует набор половых хромосом XX, мужскому — ХУ. У других животных (птицы, бабочки), наоборот, XX означает мужской пол, а ХУ — женский. У некоторых видов хромосома У может отсутствовать и тогда женский пол будут определять две Х-хромо-сомы (XX), а мужской пол — только одна Х-хромосома (ХО).
Не только определение пола, но и числовое равенство полов в каждом поколении вытекает из механизма гаметогенеза. Когда образуются половые клетки, диплоидный набор хромосом распределяется на два гаплоидных. При этом распределяются и половые хромосомы. Если исходная клетка имела XX, то все гаметы будут иметь по одной Х-хромосоме. При этом все образовавшиеся гаметы будут одинаковы. Если исходная клетка имела ХУ, то половина гамет будет иметь одну Х-хромосому, а половина — одну У-хромосому. В момент оплодотворения одинаково вероятно, что сольются в одну зиготу или гаметы, имеющие по Х-хромосоме, или гаметы, одна из которых несет Х-хромосому, а другая — У-хромосому. В результате зиготы XX и ХУ в каждом поколении образуются примерно в одинаковом количестве. А это и означает, что число самцов и самок в каждом поколении примерно одно и то же.
biologiya.net