ВЛИЯНИЕ ОБРАБОТКИ РАСТЕНИЙ МОНОФОСФАТОМ КАЛИЯ И ФУНГИЦИДОМ НА ЗЕРНОВУЮ ПРОДУКТИВНОСТЬ ИНТЕНСИВНЫХ СОРТОВ ОЗИМОЙ ПШЕНИЦЫ. Хлорофилл а и б в растениях озимой пшеницы
Гуляева А.Б., Гуляев Б.И. Влияние обработки фунгицидом при разных уровнях фосфорного питания на фотосинтез и продуктивность озимой пшеницы
Гуляева А.Б.1, Гуляев Б.И.21Институт физиологии растений и генетики Национальной академии наук Украины2Институт физиологии растений и генетики Национальной академии наук Украины, д.б.н., профессор
Gulyaeva A.B.1, Gulyaev B.I.21Institute of Plant Physiology and Genetics of National Academy of Sciences of Ukraine2Institute of Plant Physiology and Genetics of National Academy of Sciences of Ukraine, Ph.D. (doctor of sciences) in biology, professor
Библиографическая ссылка на статью:Гуляева А.Б., Гуляев Б.И. Влияние обработки фунгицидом при разных уровнях фосфорного питания на фотосинтез и продуктивность озимой пшеницы // Современные научные исследования и инновации. 2012. № 11 [Электронный ресурс]. URL: http://web.snauka.ru/issues/2012/11/18378 (дата обращения: 23.09.2018).
Проблема фосфорного питания озимой пшеницы стоит особенно остро в связи с тем, что растения этой культуры, в отличие от других сельскохозяйственных культур, имеют низкую способность к усвоению фосфора из почвы, но в то же время высокую чувствительность к фосфорным удобрениям. Так, для получения урожая зерна пшеницы до 100 ц / га оптимальная доза фосфора составляет 120-170 кг / га [2, 6].
Учитывая вышесказанное, актуальным является поиск альтернативных способов улучшения фосфорного питания растений пшеницы. По результатам наших предыдущих данных, к одному из таких способов можно отнести обработку растений фунгицидом амистар экстра 280 SC, в состав которого входят азоксисторбин, 200 г / л и ципроканозол 80 г / л – производные триазолов и стробилуринов [5].
Этот фунгицид применяется в качестве средства защиты растений пшеницы от болезней, таких как: септориоз колоса, мучнистая роса, бурая ржавчина листьев, альтернариоз. По регламенту применения за вегетацию, но не позднее, чем за 30 дней до уборки урожая, проводится до двух обработок дозой 0,5 – 0,75 л / га. Вместе с тем установлено, что этот фунгицид, кроме защиты растений от болезней, оказывает также позитивное воздействие на физиологические процессы в растениях: повышает эффективность использования ими воды, улучшает фотосинтез, оптимизирует азотный обмен, замедляет старение растений посредством подавления образования в растениях этилена [4].
В связи с этим, нами в 2011 году выполнено вегетационный опыт по изучению влияния обработки растений озимой пшеницы фунгицидом амистар экстра 280 SС при двух дозах фосфорного питания на интенсивность фотосинтеза, транспирации, дыхания растений и зерновую продуктивность, а так же структурные показатели урожая, а также содержание протеина в зерне.
Методика.
Место проведения опыта – вегетационный домик ИФРГ НАН Украины. Растения выращивали в сосудах Вагнера на 8 кг серого оподзоленные почвы на вариантах без обработки и с обработкой растений фунгицидом амистар экстра 280 SС в фазы кущения, выхода в трубку и цветения-колошения [1]. Схема опыта: 1 – NPK (90) – контроль, без обработки, 2 – NK (90) P (45), без обработки, 3 – NPK (90) + обработка растений амистар экстра 280 SC 4 – NK (90) P (45) + обработка растений амистар экстра 280 SC.
Варианты:
1) NK (90) P (90) – с обработкой и без обработки
2) NK (90) P (45) – с обработкой и без обработки
Определяли интенсивность фотосинтеза, темнового дыхания и фотодыхания, транспирации флагового листа в фазе колошения-цветения (30.06.2011) по стандартной методике при интенсивности ФАР 400 Вт / м и температуре 250C [3]. Содержание хлорофилла определяли с помощью хлорофиломира SPAD-502 (в условных единицах SPAD) («Konica Minolta») [7], а содержание протеина в зерне – по модифицированному микрометоду Кьельдаля [5]. Результаты обрабатывали с программой Microsoft Excel, статистическую обработку делали по Доспехову [1].
Результаты и обсуждение
Данные в табл. 1 свидетельствуют о существенном влиянии обработки фунгицидом амистар экстра 280 SC на интенсивность CO2 газообмена флагового листа. Так, в варианте с полной дозой фосфора – NPK (90) – у растений, обработанных фунгицидом, интенсивность фотосинтеза была выше, чем в контроле на 11,3%, а в варианте с двукратным снижением доли фосфора NPK (45) – на 40%, при одновременном снижении интенсивностей фотодыхания и темнового дыхания и повышение интенсивности транспирации на 48%. Последнее свидетельствует о снижении устьичного сопротивления листьев.
ТАБЛИЦА 1. Интенсивность фотосинтеза, фотодыхания, темнового дыхания в мг СО2 / дм2 час, интенсивность транспирации г Н2О / дм2 час в флаговом листке растений озимой пшеницы при двух уровнях фосфорного питания и обработки растений фунгицидом амистар экстра.
| Варианты | Фотосинтез | Фотодыхание | Темновое дыхание | Транспирация |
| Контроль (NPK(90)) | 28,4±0,4 | 5,6±0,2 | 2,0±0,1 | 1,93±0,1 |
| NK (90) P(45) | 20,0±0,9 | 4,4±0,2 | 1,8±0,1 | 1,77±0,1 |
| NPK(90) + амистар экстра | 31,6±1,4 | 5,0±0,3 | 1,8±0,1 | 2,85±0,1 |
| NK (90) P(45) + амистар экстра | 28,0±0,4 | 4,0±0,2 | 1,6±0,08 | 2,62±0,1 |
Установлено повышение хлорофилла в листьях на 4% в варианте NК (90) P (45) под действием обработки растений фунгицидом (Табл.2).
ТАБЛИЦА 2. Содержание хлорофилла в флаговом листке озимой пшеницы (в у.ед.) при двух уровнях фосфорного питания и обработки растений фунгицидом амистар экстра.
| Вариант | Содержание хлорофилла, (в у.ед. SPAD) |
| Контроль (NPK(90)) | 46,5±2,1 |
| NK (90) P(45) | 47,1±2,0 |
| NPK(90) + амистар экстра | 48,3±2,4 |
| NK (90) P(45) + амистар экстра | 46,7±2,3 |
Положительное влияние обработки фунгицидом наблюдался также на структурные показатели урожая и зерновую продуктивность (табл.3). Так, при дозе фосфора 90 мг на 1 кг почвы масса зерна растений в расчете на сосуд составила 22,7 г, а у растений, обработанных препаратом амистар экстра – 25,5 г – больше, на 12,3%, чем в варианте без обработки растений. В этих условиях наблюдалось также увеличение высоты растений и числа стеблей на растение – на 20%, количества колосков на побег и массы 1000 зерен главного колоса на 11%, колосков главного побега и колосьев боковых побегов соответственно на 9 и 27%. По дозы фосфора 45 мг на 1 кг почвы масса зерна у необработанных фунгицидом растений в расчете на сосуд составила 19,8 г, а у растений, обработанных фунгицидом амистар экстра – 24,3 г, т.е. больше на 22,7%, чем в варианте без обработки. Это позволяет сделать вывод о возможном участии обработки растений фунгицидом в регуляции фосфорного питания растений озимой пшеницы.
ТАБЛИЦА 3. Структура зерновой продуктивности растений озимой пшеницы сорта Смуглянка при двух уровнях фосфорного питания и обработки растений фунгицидом амистар экстра.
Биометрические показатели | Контроль (NPK(90)) | NK (90) P(45) | NPK(90) + амистар экстра | NK (90) P(45) + амистар экстра | |
Высота растений, см | 72,7±3,6 | 82,7±4,1 | 87,2±4,1 | 91±4,6 | |
| Количество побегов, шт. | 3±0,1 | 4±0,1 | 4±0,1 | 4±0,1 | |
| Масса 1000 зерен, г | 63,4±3,2 | 63,8±3,2 | 74,5±3,7 | 80,6±4,01 | |
| Число колосков в колосе главного побега | 17±0,9 | 18±0,8 | 19±0,9 | 20±1,0 | |
Масса зерна,г/колос | главного побега | 1,1±0,06 | 0,9±0,05 | 1,2±0,5 | 1,1±0,5 |
| боковых побегов | 1,1±0,5 | 1,1±0,5 | 1,4±0,6 | 1,5±0,6 | |
Масса снопа, г | 28,5±1,4 | 23,1±1,2 | 28,6±1,4 | 27,5±1,4 | |
| Масса зерна, г/ сосуд | 22,7±1,1 | 19,8±1,0 | 25,5±1,3 | 24,3±1,2 | |
Определение содержания азота в соломе и зерне показало снижение его содержания: в зерне на 0,5% при двукратном уменьшении внесении дозы фосфора в почву, а также тенденцию к его повышению на 0,3% в этих условиях при обработке растений фунгицидом, содержание которого увеличилось (табл. .4). Также был отмечен эффект обработки фунгицидом на содержание протеина в зерне на 1,5%. При этом влияние препарата на содержание азота в соломе не наблюдалось.
ТАБЛИЦА 4. Содержание азота и сырого протеина в органах озимой пшеницы при двух уровнях фосфорного питания и обработки растений фунгицидом амистар экстра.
| Варианты | содержание азота,% | содержание протеина,% | |||
| зерно | солома | почва | зерно | солома | |
| Контроль | 2,3±0,1 | 1,0±0,5 | 0,9±0,04 | 12,8±0,6 | 6,1±0,3 |
| NK90P45 | 1,8±0,6 | 0,9±0,4 | 0,9±0,04 | 10,5±0,5 | 5,8±0,3 |
| NPK90+ амистар экстра | 1,9±0,1 | 1,0±0,5 | 0,8±0,04 | 10,7±0,5 | 6,0±0,3 |
| NK90P45+ амистар экстра | 2,1±0,1 | 0,9±0,4 | 1,0±0,05 | 12,0±0,5 | 5,6±0,3 |
Таким образом, установлено повышение под действием обработки растений озимой пшеницы сорта Смуглянка фунгицидом амистар экстра 280 SС интенсивности фотосинтеза, зерновой продуктивности, а также содержания сырого протеина в зерне, при этом эффект был выше при недостатке фосфора в почве.
Положительный эффект от обработки растений пшеницы фунгицидом амистар экстра на интенсивность фотосинтеза листьев, зерновую продуктивность и качество зерна можно объяснить повышением активности корневой системы растений. Об этом свидетельствует значительное повышение под действием обработки растений фунгицидом интенсивности транспирации листьев (до 48%)
Таким образом, полученные данные позволяют сделать вывод о том, что трехкратная обработка растений озимой пшеницы фунгицидом амистар экстра в фазы кущения, выхода в трубку и колошения-цветения дает возможность значительно повышать зерновую продуктивность растений озимой пшеницы, а также качество зерна (массы 1000 зерен и содержание протеина в зерне) не только при в условиях недостатка фосфора в почве, а также при полной норме фосфора.
Увеличение под действием обработки растений в фазе цветения-колошения фунгицидом амистар экстра высоты растений, количества боковых побегов и их зерновой продуктивности дает основание сделать вывод о том, что под действием этого фунгицида в клетках растений озимой пшеницы в верхушках побегов корней, в молодых листьях, а также в растущих зародышах начинается синтез и накопление гиббереллина – фитогормона, который стимулирует рост клеток растяжением, вызывает увеличение высоты растений, улучшает развитие и семенную продуктивность боковых побегов.
Важно заметить, что улучшение эффективности усвоения фосфора растениями вызывает повышение эффективности усвоения ими азота [2, 6].
Таким образом, обработка посевов озимой пшеницы в указанные выше фазе вегетации фунгицидом амистар экстра может быть одним из эффективных способов повышения зерновой продуктивности озимой пшеницы.
Библиографический список- Доспехов Б.А. Методика полевого опыта (с основами статистической обработки результатов исследований). – М.: Агропромиздат, 1985. – 351 с.
- Моргун В.В., Швартау В.В., Киризий Д.А. Физиологические основы формирования высокой продуктивности зерновых злаков // Физиология и биохимия культ. растений.– 2010. – 42, № 5. – С. 371-393.
- Калинин Ф.Л. Биологически активные вещества в растениеводстве. 4.2.1. Гибереллины как один из компонентов комплекса факторов, активирующих формообразование у растений. Киев: Наук. думка, 1984.
- Клуб 100 центнерів. Сорти та технології вирощування високих урожаїв озимої пшениці. ІФРГ НАН України. Компанія Сингента, (Швейцарія). Київ. 2010.
- Перелік пестицидів і агрохімікатів, дозволених до використання в Україні Міністерство охорони навколишнього природнього средовища в Україні. Дніпропетровськ: Арт-Прес, 2006.
- Швартау В.В., Гуляев Б.И., Карлова А.Б. Особенности реакции растений на дефицит фосфора // Физиология и биохимия культурных растений. — 2009. — 41, №3.- С. 208- 220.
- Yildirim M., KilicH.,KendalE., KarahanT. Applicability of chlorophyll meter readings as yield predictor in durum wheat // J. of Plant Nutrition. – 2010. – 34. —Is 2. — P. 151-164.
Все статьи автора «Anna_Gulaeva»
web.snauka.ru
ВЛИЯНИЕ ИНОКУЛЯЦИИ НА СОДЕРЖАНИЕ ХЛОРОФИЛЛОВ В ЛИСТЬЯХ ПШЕНИЦЫ ОЗИМОЙ ПРИ ЗАГРЯЗНЕНИИ ПОЧВЫ ТЯЖЕЛЫМИ МЕТАЛЛАМИ
ВЛИЯНИЕ ИНОКУЛЯЦИИ НА СОДЕРЖАНИЕ ХЛОРОФИЛЛОВ В ЛИСТЬЯХПШЕНИЦЫ ОЗИМОЙ ПРИ ЗАГРЯЗНЕНИИ ПОЧВЫ ТЯЖЕЛЫМИ МЕТАЛЛАМИ
Людмила Александровна Чайковская
д-р с.-х. наук, гл. науч. сотр. лаб. растительно-микробного взаимодействия
Федерального государственного бюджетного
учреждения науки «НИИСХ Крыма»,
РФ, г. Симферополь
E-mail: [email protected]
Марина Ивановна Баранская
канд. с.-х.наук, зав. лаб. растительно-микробного взаимодействия
ФГБНУ «НИИСХ Крыма»,
РФ, г. Симферополь
E-mail: [email protected]
Ольга Леонидовна Овсиенко
научный сотрудник лаб. растительно-микробного взаимодействия
ФГБНУ «НИИСХ Крыма»,
РФ, г. Симферополь
E-mail: [email protected]
Нина Николаевна Клименко
научный сотрудник лаб. растительно-микробного взаимодействия
ФГБНУ «НИИСХ Крыма»,
РФ, г. Симферополь
E-mail: [email protected]
INOCULATION INFLUENSE ON CHLOROPHYLL’S CONTENT
IN WINTER WHEAT LEAVES WHEN SOIL POLLUTION’S BY HEAVY METALLS
Ludmila Chaikovska
doctor of Agriculture Science, Seniorstaff scientist, department of plant-microbeinteraction,
Federal state budgetary institution of science
“Scientific Research Institute of agriculture of Crimea”,
Russia, Simferopol
Marina Baranskaya
candidate of agricultural sciences,PhD,
Head of plant-microbeinteraction department, FSBIS “SRIAC of Crimea”,
Russia,
Olga Ovsienko
scientific researcher, Dep.ofplant-microbeinteraction, FSBIS “SRI AC of Crimea”,
Russia, Simferopol
Nina Кlymenko
scientific researcher, Dep. ofplant-microbeinteraction,FSBIS “SRI AC of Crimea”,
Russia, Simferopol
АННОТАЦИЯ
Исследовано влияние предпосевной инокуляции семян на количественное содержание хлорофиллов в листьях пшеницы озимой на ранних этапах онтогенеза растений и на повышение устойчивости пшеницы к стрессовому воздействию тяжелых металлов (ТМ) (Cr, Cu, Pb). Установлено положительное воздействие Диазофита на возрастание количества хлорофиллов (a+b) в листьях пшеницы озимой как в контроле, так и на фоне загрязнения почвы ТМ. Содержание пигментов в листьях бактеризованных растений в контроле и на различных уровнях ПДК ТМ превышало значения небактеризованных растений на 15 % и 7–15 % соответственно. Выявлено, что инокуляция снижает стрессовое воздействие ТМ: фитомасса бактеризованных растений при загрязнении почвы на уровне 1 ПДК возросла на 42 % по сравнению с растениями, выросшими из неинокулированных семян, а на уровне 2,5 и 5 ПДК – на 25 % и 90 % соответственно. Таким образом, полученные результаты свидетельствуют о позитивном влиянии Диазофита на количество хлорофиллов (a+b) в листьях пшеницы озимой в ранние этапы онтогенеза растений, что способствует повышению их продуктивности и роста благодаря смягчению стрессового воздействия ТМ.
ABSTRACT
It was studied the influence of presowingseeds inoculation on the quantitative content of chlorophylls in winter wheat leaves оn the early stages of plant ontogenesis and improvement of wheat resistance to stressful effect of HM (Cr, Cu, Pb). Was found the positive impact of Diazofit on increase the amount of chlorophyll (a+b) in leaves of winter wheat in the control and in the background of soil contaminated with the HM. The content of pigments in the leaves of inoculated plants in control and at different levels of HM MPC of none inoculated exceeds the value of the plant by 15 % and 7–15 %, respectively. It was revealed that inoculation reduces the stress effect of HM: phytomass of inoculated plants on contaminated soil at 1 MPC has increased by 42 % as compared to plants grown from uninoculated seeds, and at 2,5 and 5 MPC on 25 % and 90 %, respectively. Thus, the results indicate a positive influence of Diazofit on chlorophyll (a+b) in leaves of winter wheat in the early stages of plants ontogeny, thereby increasing productivity and growth through mitigation of HM stress.
Ключевые слова: инокуляция; Диазофит; озимая пшеница; хлорофиллы; тяжелые металлы.
Keywords: inoculation; Diazofit; winter wheat; chlorophyll; heavy metals.
Увеличение антропогенной нагрузки на окружающую среду оказывает все более негативное влияние на различные биологические объекты. Одним из распространенных загрязнителей, которые наносят значительный вред различным компонентам природной среды, являются тяжелые металлы (ТМ). Попадая в трофические цепи, ТМ могут значительно изменять интенсивность метаболических процессов растений (в частности фотосинтеза), что снижает их продуктивность и качество урожая. Так, выявлена позитивная корреляция между зерновой продуктивностью сельскохозяйственных растений и интенсивностью фотосинтеза [7, с. 60–66; 8, с. 411–415]. В почвах, загрязненных ТМ, нарушается интенсивность биологических процессов и снижается количество полезных микроорганизмов, что может негативно влиять на культурные растения [6, с. 60–66]. В то же время известно, что симбиотрофные микроорганизмы являются посредниками между эдафическими условиями (в том числе и токсичностью ТМ) и растениями, и способствуют значительному повышению устойчивости макросимбионта к стрессу [1, с. 17–22]. Именно поэтому поиск способов, которыеснижают негативное воздействие ТМ на продуктивность растений и качество продукции является актуальным и своевременным. Исследователями созданы эффективные микробные препараты, которые успешно применяют в технологиях выращивания сельскохозяйственных культур в различных агроклиматических зонах. Установлено, что биопрепараты улучшают питание растений, повышают их урожайность и способствуют получению экологически чистой продукции [5, с. 352–375; 2, с. 15–300]. В наших предыдущих исследованиях выявлено положительное влияние предпосевной бактеризации (на примере микробного препарата Фосфоэнтерин) на состав фотосинтезирующих пигментов и продуктивность растений при загрязнении почвы ТМ [9, с. 32–35].
Учитывая вышеизложенное,цель наших исследованийсостояла в изучении влияния предпосевной инокуляции семянна количественное содержание хлорофиллов в листьях на ранних этапах онтогенеза растений и повышение устойчивости пшеницы озимой к стрессовому воздействию ТМ (Cr, Cu, Pb).
Вегетационные опыты проведены в теплице, культура: пшеница озимая Triticum durumL. Для предпосевной инокуляции семян использован микробный препарат Диазофит (основа Agrobacterium radiobacter 204), контроль – без инокуляции. Растения выращивали в пластиковых сосудах (объем 0,5 л) в течение 6 недель, почва: чернозем южный. Повторность опытов пятикратная. При закладке опытов в сосуды вносили по 380 г воздушно-сухой почвы. В каждый сосуд были внесены и тщательно перемешаны с почвой растворы солей ТМ: Pb (Ch4COO)2, CuSO4, K2CrO4 из расчетов, которые соответствовали следующим уровням загрязнения: 1 ПДК, 2,5 ПДК, 5 ПДК. В контрольном варианте ТМ не вносили. Экстракцию фотосинтезирующихпигментов проводили при помощи этилового спирта, а их количественное содержание в листьях растений определяли колориметрическим методом [3, с. 127–134]. Вегетационные опыты и математическая обработка полученных результатов проведены согласно общепринятых методик [4].
Результаты исследований, полученные в условиях вегетационных опытов, показали, что загрязнение почвы ТМ снижает содержание фотосинтезирующих пигментов в листьях пшеницы озимой на ранних этапах онтогенеза (рис. 1). Так, содержание суммы хлорофиллов (a +b) в листьях растений, выросших на незагрязненной почве, достигало3,40 мг/г сырой фитомассы, а внесение в грунт ТМ снижало эти показатели до 3,24 мг/г.
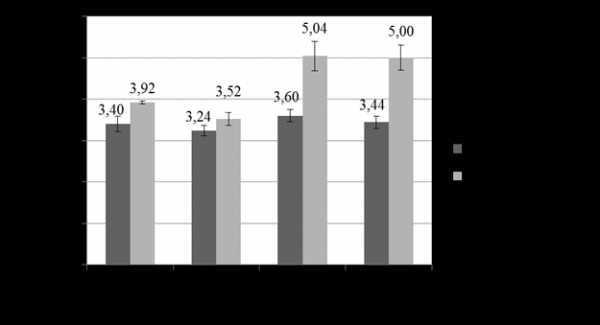
Рисунок 1. Влияние инокуляции на содержание хлорофиллов (a+b) в листьях пшеницы озимой при загрязнении почвы ТМ (Cr, Cu, Pb)
Анализ полученных результатов свидетельствуют о положительном воздействии предпосевной бактеризации семян на возрастание количества хлорофиллов в листьях пшеницы озимой как в контроле, так и на фоне загрязнения почвы ТМ. Содержание пигментов в листьях бактеризованных растений достигало 3,92 мг/г фитомассы (контроль) и 3,52–5,04 мг/г (различные уровни ПДК ТМ), что превышало значения небактеризованных растений на 15 % и 7–15 % соответственно.
Нарушение функционирования пигментного комплекса пшеницы озимой при загрязнении почвы ТМ снижает продуктивность растений: их фитомасса уменьшилась на уровне 1 и 2,5 ПДК на 7 %, а на уровне 5 ПДК – в 2,3 раза по сравнению с контролем (табл. 1). Использование Диазофита для предпосевной инокуляции семян способствовало увеличению фитомассы пшеницы как в контрольном варианте (на 19 %), так и при загрязнении почвы ТМ. Установлено, что бактеризация снижает стрессовое воздействие ТМ: фитомасса бактеризованных растений при загрязнении почвы на уровне 1 ПДК возросла на 42 % по сравнению с растениями, выросшими из неинокулированных семян, а на уровне 2,5 и 5 ПДК –на 25 % и 90 % соответственно.
Таблица 1.
Влияние инокуляции на продуктивность и высоту пшеницы озимой при загрязнении почвы ТМ (чернозем южный, вегетационный опыт)
| Вариант | Фитомасса | Высота | |||
| г/растение | % | см | % | ||
| Контроль (без внесения ТМ) | |||||
| Без инокуляции | 0,26 | 100 | 13,6 | 100 | |
| Диазофит | 0,31 | 119 | 13,6 | 100 | |
| НСР05 | 0,050 | 1,00 | |||
| ТМ (1 ПДК) | |||||
| Без инокуляции | 0,24 | 100 | 14,6 | 100 | |
| Диазофит | 0,34 | 142 | 17,4 | 119 | |
| НСР05 | 0,030 | 1,00 | |||
| ТМ (2,5 ПДК) | |||||
| Без инокуляции | 0,24 | 100 | 12,8 | 100 | |
| Диазофит | 0,30 | 125 | 15,2 | 119 | |
| НСР05 | 0,060 | 1,29 | |||
| ТМ (5 ПДК) | |||||
| Без инокуляции | 0,11 | 100 | 10,9 | 100 | |
| Диазофит | 0,21 | 190 | 15,6 | 143 | |
| НСР05 | 0,060 | 2,17 | |||
Необходимо отметить также положительное воздействие Диазофита на высоту пшеницы, выросшей на загрязненной почве: она превышала рост небактеризованных растений на 19–43 % в зависимости от уровня ПДК ТМ.
Таким образом, полученные результаты свидетельствуют о позитивном влиянии предпосевной инокуляции семян (Диазофит) на количественный состав суммы хлорофиллов (a+b) в листьях пшеницы озимой в ранние этапы онтогенеза растений, что способствует повышению их продуктивности и роста благодаря смягчению стрессового воздействия ТМ.
Список литературы:
- Белимов А.А, Тихонович И.А. Микробиологические аспекты устойчивости и аккумуляции тяжелых металлов у растений // Сельскохозяйственная биология. – 2011. – № 3. – С. 17–22.
- Волкогон В.В. Мікробніпрепарати у землеробстві. Теорія і практика / В.В. Волкогон, О.В. Надкернична, Т.М. Ковалевська та ін. – К.: Аграрна наука, 2006. – 312 с.
- Гавриленко В.Ф., Ладыгина М.Е., Хандобина Л.М. Большой практикум по физиологии растений.Фотосинтез. Дыхание: учебное пособие. – М.: Высшая школа, 1975. – C. 127–134.
- Доспехов Б.А. Методика полевого опыта. – М.: Агропромиздат, 1985. – 351 с.
- Иутинская Г.А. Биорегуляция микробно-растительных систем: Монография / Г.А. Иутинская, С.П. Пономаренко, Е.И. Андреюк и др. – К.: Ничлава, 2010. – С. 352–375.
- Иутинская Г.А., Колпа Ю.В., Степашко В.С. Моделирование динамики численности микроорганизмов в почве, загрязненной тяжелыми металлами // Мікробіологічний журнал. – 2002. – Т. 64, № 3. – С. 59–67.
- Прядкина Г.А., Шадчина Т.М. Связь между показателями мощности развития фотосинтетического аппарата и зерновой продуктивностью озимой пшеницы в разные по погодным условиям годы // Физиология и биохимия культурных растений. – 2009. – Т. 41, № 1. – С. 59–68.
- Шадчина Т.М., Прядкіна Г.О., Моргун В.В. Зв'язоки між характеристиками фотосинтетичногоапарату та зерновою продуктивністю у різних сортів озимої пшениці // Досягнення і проблеми генетики, селекції і біотехнології. Зб. наук. праць. – К.: Логос, 2007. – Т. 2. – С. 410–415.
- Чайковская Л.А., Cологуб Н.А., Баранская М.И. Влияние бактеризации на развитие Sorghum sudanense (Piper.) Stapf. и ризосферную микрофлору при воздействии тяжелых металлов // Промышленная ботаника. Сб. науч. трудов. – Донецк: Донецкий ботсад НАН Украины, 2011. – С. 32–35.
sibac.info
Гуляева А.Б., Гуляев Б.И. Влияние обработки растений монофосфатом калия и фунгицидом на зерновую продуктивность интенсивных сортов озимой пшеницы
Гуляева А.Б.1, Гуляев Б.И.21Институт физиологии растений и генетики Национальной академии наук Украины2Институт физиологии растений и генетики Национальной академии наук Украины, д.б.н., профессор
Gulyaeva A.B.1, Gulyaev B.I.21Institute of Plant Physiology and Genetics of National Academy of Sciences of Ukraine2Institute of Plant Physiology and Genetics of National Academy of Sciences of Ukraine, Ph.D. (doctor of sciences) in biology, professor
Библиографическая ссылка на статью:Гуляева А.Б., Гуляев Б.И. Влияние обработки растений монофосфатом калия и фунгицидом на зерновую продуктивность интенсивных сортов озимой пшеницы // Современные научные исследования и инновации. 2012. № 10 [Электронный ресурс]. URL: http://web.snauka.ru/issues/2012/10/17930 (дата обращения: 23.09.2018).
В полевых опытах установлено, что обработка растений озимой пшеницы фунгицидом амистар экстра, а также 3% раствором монофосфата калия (МКФ) и их смесью способствует повышению зерновой продуктивности, особенно при хорошей обеспеченности азотом на варианте – N100. В этих условиях зерновая продуктивность озимой пшеницы сорта Актер увеличилась на 21 – 25%. Обработка растений озимой пшеницы сортов Актёр и Смуглянка 3% раствором МКФ также способствовала повышению зерновой продуктивности, которая повысилась на 24% и 24,7% соответственно. В опытных вариантах обоих сортов наблюдался так же позитивный эффект обработки фунгицидом и раствором монофосфата калия на коэффициент выноса азота и эффективность его использования на растениях обоих сортов, хотя содержание белка в зерне опытных растений имело тенденцию к снижению, за исключением варианта с обработкой 3% МКФ растений озимой пшеницы сорта Смуглянка, где процент белка повышался на 1,8% : с 8,6% на контроле до 10% .
Фосфор относится к важным элементам минерального питания растений и его оптимизация является необходимым условием реализации потенциала продуктивности сельскохозяйственных культур. Содержание этого элемента в растениях составляет около 0,2% массы сухого вещества как компонент и ключевой элемент молекул нуклеиновых кислот, фосфолипидов и АТФ. Фосфор участвует в контроле и регуляции главных ферментативных реакций и метаболических путей в растительных клетках и организмах: процессах фотосинтеза, дыхание, роста и развития растений. Установлено, что существует взаимосвязь между содержанием азота и фосфора в органическом веществе вегетативных органов растений. Соотношение N / Р в растениях редко бывает ниже 0,04 г / г и выше 0,15 г / г, а в белках и нуклеиновых кислотах оно составляет около 0,055. По этой причине, эффективность процессов поглощения растениями азота и фосфора взаимосвязаны: при недостатке азота уменьшается поглощение фосфора, а при дефиците фосфора – усвоение азота это ухудшение связано с торможением синтеза органических кислот в растениях, что вызывает снижение усвоения аммиачного азота, поступающего через корни. При недостатке азота потребность растений в фосфоре снижается из-за ослабления процессов синтеза фосфорсодержащих органических веществ в клетках.
Учитывая это, актуальным является поиск нетрадиционных способов улучшения фосфорного питания озимой пшеницы. По нашим предварительным данным, в число одного из таких способов можно отнести обработку растений фунгицидом амистар экстра 280 SC. Этот фунгицид, в состав его входят азоксисторбин, 200 г / л и ципроканозол, 80 г / л предназначен для защиты растений пшеницы от таких болезней как: септориоз колоса, мучнистая роса, бурая ржавчина листьев, альтернариоз. Вместе с тем установлено [4], что этот фунгицид также оказывает положительное влияние на физиологические процессы в растениях: повышает эффективность использования воды, оптимизирует азотный обмен и тормозит синтез этилена в растениях, продлевая вегетацию растений [4]. В то же время, одним из возможных способов повышения зерновой продуктивности пшеницы может быть также обработка вегетирующих растений раствором монофосфата калия КН2РО4 в составе которого – Р2О5 и К2О, массовые части фосфора и калия равны соответственно 52 и 34%. Содержание ортофосфата в 3% растворе МКФ (моно фосфата калия): 1,1 г/10 м2 – КН2РО4.
Методика работы.
В 2011 – 2012г. нами были проведены исследования на полях опытного сельскохозяйственного производства ИФРГ НАНУ (пгт Глеваха Васильковского района, Киевской области) и на полях ООО «Агросервис» г. Жашков Черкасской области. Целью наших исследований было изучение воздействия на зерновую продуктивность интенсивных сортов озимой пшеницы обработки вегетирующих растений в фазу колошения-цветения 3% раствором монофосфата калия – КН2РО4 и фунгицидом амистар экстра 280 SC, а так же их смесью. На опытных полях в Киевской области почва серая лесная, фон минерального питания N45P45 K45. Площадь одного опытного участка составляла – 10 м2, повторность 4-х кратная. Объект изучения – растения сорта Смуглянка короткостебельной озимой мягкой пшеницы, интенсивного типа – селекции ИФРГ НАН Украины. В Черкасской области на опытных полях почва – чернозем, фон минерального питания N100P0K0. Площадь одного опытного участка – 10 м2, повторность 4-х кратная. Объект изучения сорт Актер немецкой селекции, устойчив к полеганию, мучнистой росе и бурой листовой ржавчины, который получил от Федеральной комиссии Германии по сортоиспытанию высокий балл.
Схемы опытов:
1. Контроль – без обработки растений; 2. Обработка растений 3% водным раствором МКФ; 3. Обработка растений раствором фунгицидом амистар экстра 280 SС; 4. Обработка растений смесью амистар экстра 280 SС и 3% раствора МКФ.
В фазу колошения-цветения во флаговом листе определяли содержание хлорофилла в условных единицах SPAD-502 с помощью хлорофилломера («Konica Minolta») (SPAD) [7], содержание азота в зерне определяли по модифицированному микрометоду Кьельдаля [6]. Содержание белка рассчитывали, используя показатели общего содержания азота (%) в зерне по следующей формуле:
Х=(m∙V∙V1 ∙100)∙5,7/(V2∙m1 ∙106),
где m – масса азота, найденная по калибровочному графику, мкг;
V – объем р-ра после озоления, мл;
V1 – объем р-ра взятого для фотометрирования, мл;
V2 - объем р-ра взятого для определения азота, мл;
m1 –масса навески, г;
5,7 – коэффициент для перерасчета азота на белок в зерне пшеницы.
Общее содержание азота в растении в период зрелости рассчитывали по следующей формуле [4]:
Nt = [масса зерна (г × м2) × концентрация элемента в зерне (%)] + [остальная биомасса (солома) при 0% влажности (г × м2 ) × концентрация элемента в остальной биомассе (соломы) (%)].
Кхоз рассчитывали по следующей формуле:
Кхоз = Gw/Tw,
Gw – масса зерна при 0% влажности, Tw - общая биомасса растений при 0% влажности (г × м2).
Коэф. выноса N считали по формуле: Nt / внесенное количество элемента. Эффективность продукции биомассы (ЕПБ) = Gw / Nt
Эффективность утилизации = индекс урожая × эффективность продукции биомассы.
Результаты обрабатывали Microsoft Excel, статистическую обработку делали по Доспехову [1].
Результаты и их обсуждение.
По данным табл.1 содержание хлорофилла флагового листа озимой пшеницы сорт Смуглянка (Актер) под действием обработки растений фунгицидом амистар экстра 280 SC имело тенденцию к увеличению на 3,9 (2,9%), на варианте с обработкой смесью амистар экстра 280 SС + МКФ – на 0,4 (2,2%), а при действием обработки раствором МКФ – на 1,3 (1,4%).
Таблица 1. Содержание хлорофилла в флаговом листе озимой пшеницы (в у.е. SPAD) под действием внекорневой обработки 3% раствором МКФ и фунгицидом амистар экстра.
| Варианты | Сорт Смуглянка | Сорт Актёр |
| Контроль (без обр.) | 46,5±2,1 | 48,9±2,3 |
| Обработка 3 % р-ром МКФ | 47,1±2,0 | 49,6±2,5 |
| Обработка фунгицидом | 48,3±2,4 | 50,3±2,5 |
| Обработка смесью: фунгицид + 3 % р-ром МКФ | 46,7±2,3 | 50,0±1,9 |
Обработка растений озимой пшеницы сортов Смуглянка и Актер 3% раствором монофосфата калия привела к повышению зерновой продуктивности на 6,5 ц / га: с 27 до 33,5 ц / га (24%) и на 9 ц / га, то есть с 38 до 47,4 ц / г (24,7%) соответственно. Под действием обработки растений фунгицидом наблюдалось повышение зерновой продуктивности озимой пшеницы сорта Актёр на 9,7 ц / га: – с 38,4 до 48,1 ц / га (25,3%), а обработка смесью фунгицида и 3% монофосфата калия – на 8 ц / га (с 38,4 до 46,4 ц / га (20,8%)) – эффект ниже (табл.2.). Так же нами был получен позитивный эффект от обработки фунгицидом и его смесью с 3% МКФ на биометрические показатели растений озимой пшеницы исследуемых сортов. По полученным данным влияние обработки фунгицидом амистар экстра и раствором МФК на показатель массы 1000 зерен в опыте с растениями озимой пшеницы сорта Смуглянка был незначителен: наблюдалась лишь тенденция к увеличению этого показателя на варианте с обработкой смесью фунгицид + 3% МКФ (на 2,3%), тогда как в опыте с растениями озимой пшеницы сорта Актёр влияние было существенным. Так, на варианте с обработкой фунгицидом масса 1000 зерен увеличилась на 7,3%, с обработкой 3% раствором МКФ – на 15,8%, а на варианте с обработкой смесью фунгицида + 3% МКФ – на 11,7%. Более значительное влияние обработки исследуемыми биологическими веществами на растения сорта Актер можно объяснить лучшей обеспеченностью растений азотом из почвы. Так, по нашим данным при обработке 3%-ным раствором МКФ масса зерна с колоса у растений озимой пшеницы сорта Актер повысилась на 64,3%, при обработке фунгицидом – на 85,7%, а на варианте с использованием смеси фунгицида + 3% МКФ – на 50%.
Обработка растений озимой пшеницы сорта Смуглянка 3% р – ром МКФ способствовала увеличению массы зерна с колоса на 31,6%.
Таблица 2. Влияние обработки растений фунгицидом амистар экстра и 3 % раствором МКФ в фазе колошения-цветения на показатели зерновой продуктивности сортов озимой пшеницы Смуглянка и Актер
| Варианты | Мз1* , ц/га | Кхоз2* | Мз10003*, г | Мз.кол.4*, г | белок, % |
| Сорт Смуглянка | |||||
| Контроль(без обработки) | 27,0 ± 1,4 | 0,77 | 46,4±1,3 | 1,6±0,02 | 8,6±0,2 |
| 3% МКФ | 33,3 ± 1,7 | 0,74 | 45,3±1,1 | 2,1 ± 0,02 | 10,4±0,2 |
| фунгицид | 32,8± 1,7 | 0,67 | 46,6±1,2 | 1,4 ± 0,02 | 8,0±0,2 |
| Смесь: фунгицид + 3% МКФ | 34,0± 1,7 | 0,71 | 47,5±1,2 | 1,6+0,02 | 8,6±0,2 |
| Сорт Актёр | |||||
| Контроль | 38,4± 1,9 | 0,46 | 41,1±1,2 | 1,4±0,02 | 14,5±0,3 |
| 3% МКФ | 48,1± 2,4 | 0,76 | 47,6±1,4 | 2,3±0,02 | 12,3±0,2 |
| фунгицид | 48,0± 2,3 | 0,98 | 44,1±1,3 | 2,6±0,02 | 14,1±0,3 |
| фунгицид + 3% МКФ | 46,5± 2,3 | 0,81 | 45,9±1,2 | 2,1±0,02 | 13,2±0,3 |
Мз 1*- урожай зерна, ц / га, Кхоз2* – показатель хозяйственной эффективности урожая; Мз 10003* – масса 1000 зерен, г Мз кол.4* = масса зерна с колоса (главного + боковых) на растение, г.
Известно, что воздействие определенных агрономических мероприятий на формирование урожая озимой пшеницы анализируется не только по зерновой продуктивности, а также по таким показателям, как эффективность использования отдельных элементов питания, индекс урожая (Кхозп), эффективность утилизации отдельных элементов и др. [4]. Проанализировав полученные данные мы пришли к выводу что, общее содержание азота в растениях озимой пшеницы (в расчете на урожай зерна с 1м2) пшеницы в период зрелости в опытных вариантах увеличивался, как при обработке 3% МФК, так и применении фунгицида (таблица 3). Применение 3 -% раствора МКФ привело к увеличению общего содержания азота в 1,8 раз в растениях исследуемого варианта (сорт Смуглянка) и в 1,5 раза (сорт Актер). Обработка фунгицидом привела к увеличению общего содержания азота в 2,4 раза (сорт Смуглянка) и 1,7 раз (сорт Актер) по отношению к контрольным растениям. Показатель индекса урожая Кхоз в этих условиях увеличился при обработке растений фунгицидом амистар экстра в 2,5 раза (сорт Смуглянка) и в 1,3 раза (сорт Актер). Отмечено так же, что применение обработки смесью фунгицида и 3% МКФ растений озимой пшеницы сорта Актер привело к двукратному увеличению индекса урожая.
Известно, что эффективность утилизации питательных элементов определяется как способность культуры превращать поглощённые элементы в урожай зерна [4]. По полученным данным обработка фунгицидом амистар экстра привела к повышению этого показателя в 1,1 г. (сорт Смуглянка) и в 1,45 г. (сорт Актер), а также в 3 раза при обработке смесью: амистар экстра +3% р-р МКФ на растениях озимой пшеницы сорта Актер. Эффективность использования азота растениями озимой пшеницы по отношению к урожаю зерна на единицу внесенного питательного элемента в опытных вариантах также выросла. При обработке растений озимой пшеницы сорта Смуглянка (сорта Актер) 3% р-ром МКФ эффективность использования азота выросла в 1,4 (1,8) раза, при обработке смесью амистар экстра и 3% МКФ – в 1,5 ( 1,6) раз, под действием лишь фунгицида – в 1,3 (1,7) раза. Отмечено, также увеличение коэффициента выноса азота растениями озимой пшеницы обоих сортов.
Таблица 3. Эффективность использования и утилизации азота озимой пшеницей под действием внекорневой обработки фунгицидом и 3% р-ром МКФ.
| Варианты | Nt , г∙%∙1м2 (Общее содержание азота в урожае зерна с м2 ), | Коэф. выноса N | Эффективность утилизации N | Эффективность использования N | |
| сорт Смуглянка | |||||
| Контроль (без обработки) | 202,7±10,3 | 2,0 | 0,23 | 11,2 | |
| 3% МКФ | 356,6±17,8 | 3,6 | 0,17 | 16,0 | |
| фунгицид | 485,4±24,3 | 4,9 | 0,26 | 14,6 | |
| Смесь: фунгіцид и 3% МКФ | 268,6±13,4 | 2,7 | 0,23 | 16,4 | |
| сорт Актёр | |||||
| Контроль (без обработки) | 340,0±17,0 | 1,5 | 0,11 | 10,0 | |
| 3% МКФ | 499,2±24,9 | 2,5 | 0,16 | 18,3 | |
| фунгид | 513,6±25,7 | 2,6 | 0,16 | 17,1 | |
| Смесь: фунгіцид и 3% МКФ | 395,7±19,8 | 2,0 | 0,33 | 16,2 | |
Выводы:
Таким образом, такие агрономические мероприятия, как обработка растений озимой пшеницы фунгицидом амистар экстра, а также 3%-ным раствором МКФ и их смесью приводит к повышению зерновой продуктивности, особенно при достаточном обеспечении почвы азотом (N100), наблюдавшееся в опытах с растениями озимой пшеницы сорта Актер, зерновая продуктивность которого выросла в пределах 21 – 25%. Обработка растений 3% раствором МКФ положительно повлияла на зерновую продуктивность обоих сортов, которая увеличилась: на 24 (Смуглянка) и на 24,7% ( Актёр). Увеличились также такие показатели, как коэффициент выноса и эффективность использования азота на всех опытных вариантах в обоих сортов, хотя содержание белка в зерне опытных растений имело тенденцию к снижению, за исключением варианта с обработкой 3% МКФ растений озимой пшеницы сорта Смуглянка, где процент белка повышался на 1,8% : с 8,6% на контроле до 10% .
Библиографический список- Доспехов Б.А. Методика полевого опыта (с основами статистической обработки результатов исследований). – М.: Агропромиздат, 1985. – 351 с.
- Моргун В.В., Швартау В.В., Киризий Д.А. Физиологические основы формирования высокой продуктивности зерновых злаков // Физиология и биохимия культ. растений.– 2010. – т. 42, № 5. – С. 371-393.
- Калинин Ф.Л. Биологически активные вещества в растениеводстве. 4.2.1. Гибереллины как один из компонентов комплекса факторов, активириющих формообразование у растений. Киев, Наукова думка, 1984.
- Клуб 100 центнерів. Сорти та технології вирощування високих урожаїв озимої пшениці. ІФРГ НАН України. Компанія Сингента, (Швейцарія). Київ. 2010.
- Перелік пестицидів і агрохімікаиів, дозволених до використання в Україні Міністерство охорони навколишнього природного средовища в Україні. Дніпропетровськ, Арт-Прес. 2006.
- Швартау В.В., Гуляев Б.И., Карлова А.Б. Особенности реакции растений на дефицит фосфора // Физиология и биохимия культурных растений. — 2009. — Т. 41, №3.- С. 208- 220.
Все статьи автора «Anna_Gulaeva»
web.snauka.ru
Потребность пшеницы в азотных удобрениях в зависимости от планируемой урожайности и показаний «n-тестера»
Таблицы и графики.
Опыт применения n-тестера в Германии
В Германии N-тестер широко применяется для определения доз азотных подкормок зерновых культурах в фазы ЕС30-32 и ЕС37-51, но без какой-либо привязки к планируемой урожайности. По приведенным ниже таблицам и графикам можно отметить, что кривые оптимальных доз азота близки к отечественным данным.
Озимая пшеница
| Фаза вегетации ЕС 30/32 | Фаза вегетации ЕС 37/51 | ||
| показания N-тестера | кг N (д.в.)/га | показания N-тестера | кг N (д.в.)/га |
| > 720 | 0 | > 720 | 0 |
| 720 – 690 | 20 | 720 – 706 | 10 |
| 689 – 675 | 30 | 705 – 691 | 20 |
| 674 – 660 | 40 | 690 – 676 | 30 |
| 659 – 640 | 50 | 675 – 661 | 40 |
| 639 – 620 | 60 | 660 – 641 | 50 |
| 619 – 595 | 70 | 640 – 621 | 60 |
| 594 – 570 | 80 | 620 – 601 | 70 |
| 569 – 545 | 90 | 600 – 576 | 80 |
| < 545 | 100 | < 576 | 90 |
Озимый ячмень
| Фаза вегетации ЕС 30/32 | Фаза вегетации ЕС 37/51 | ||
| показания N-тестера | кг N (д.в.)/га | показания N-тестера | кг N (д.в.)/га |
| > 705 | 0 | > 715 | 0 |
| 705 – 686 | 10 | 715 – 696 | 10 |
| 685 – 656 | 20 | 695 – 676 | 20 |
| 655 – 626 | 30 | 675 – 656 | 30 |
| 625 – 596 | 40 | 655 – 636 | 40 |
| 595 – 571 | 50 | 635 – 616 | 50 |
| 570 – 545 | 60 | 615 – 596 | 60 |
| < 545 | 70 | < 596 | 70 |
Озимая рожь
| Фаза вегетации ЕС 30/32 | Фаза вегетации ЕС 37/51 | ||
| показания N-тестера | кг N (д.в.)/га | показания N-тестера | кг N (д.в.)/га |
| > 715 | 0 | > 681 | 0 |
| 715 – 686 | 20 | 681 – 662 | 10 |
| 685 – 656 | 30 | 661 – 622 | 30 |
| 655 – 626 | 40 | 621 – 577 | 40 |
| 625 – 596 | 50 | < 577 | 50 |
| 595 – 571 | 60 | ||
| < 571 | 70 | ||
Тритикале
| Фаза вегетации ЕС 30/32 | Фаза вегетации ЕС 37/51 | ||
| показания N-тестера | кг N (д.в.)/га | показания N-тестера | кг N (д.в.)/га |
| > 715 | 0 | > 720 | 0 |
| 715 – 686 | 10 | 720 – 706 | 10 |
| 685 – 656 | 20 | 705 – 691 | 20 |
| 655 – 626 | 30 | 690 – 676 | 30 |
| 625 – 596 | 40 | 675 – 661 | 40 |
| 595 – 571 | 50 | 660 – 641 | 50 |
| 570 – 545 | 60 | 640 – 621 | 60 |
| 544 – 520 | 70 | 620 – 601 | 70 |
| < 520 | 80 | 600 – 576 | 80 |
| < 576 | 90 | ||
Способ быстрой калибровки n-тестера для различных культур, сортов и условий их выращивания.
Быстрый способ калибровки N-тестера для различных сортов и культур основан на том, что концентрация хлорофилла не может быть выше оптимальной величины для каждого сорта, культуры и условий выращивания даже при избыточном азотном питании. Поэтому на поле искусственно создаются условия избыточного внесения азота, таким образом, достигается максимальное содержание хлорофилла в растениях.
Данная методика более точно отражает потребность растений в азоте, так как применяется в год внесения удобрений и соответственно учитывает все изменения природного характера, по сравнению с применением расчетных коэффициентов.
Наибольшая взаимосвязь между показаниями прибора и потребностью растений в азоте наблюдается у зерновых культур. Наиболее изучены и приведены данные по 50 сортам озимой пшеницы, однако насыщенность хлорофиллом листьев ячменя, тритикале и ржи также как и у других сортов пшеницы имеет аналогичную зависимость между содержанием азота в листьях и показаниями прибора. Поэтому для определения оптимальной дозы азотной подкормки с помощью методики быстрой калибровки прибора, можно использовать графики потребности озимой пшеницы по фазам вегетации, но без учета урожайности.
studfiles.net
Диссертация на тему «Физиологические особенности формирования продуктивности озимой пшеницы при воздействии биологически активных веществ и минеральных удобрений» автореферат по специальности ВАК 03.00.12 - Физиология и биохимия растений
1. Абаимов В. Ф. Продуктивность посева и качество зерна озимой пшеницы при некорневых подкормках азотом и микроэлементами / В. Ф. Абаимов, В. Б. Щукин // Зерновые культуры. 1999. - № 2. - С. 17-18.
2. Авдонин Н. С. Научные основы применения удобрений / Н. С. Авдонин. -М.: Колос, 1972. 125 с.
3. Алейников И. Эффективная технология производства гуматов / И. Алейников, В. Кусайло, М. Овчаренко // Агрохимический вестник. 2002. -№3.-С. 34-35.
4. Алиев С. Парамагнитные свойства и физиологическая активность гумусовых веществ. / С. Алиев // Сб. науч. тр. Днепропетровский СХИ. 1983. - Т.VIII: Теория действия физиологически активных веществ. - С. 68-80.
5. Алиев Д. А. Использование энергии солнечной радиации посевами озимой пшеницы в зависимости от сортовых особенностей и условий минерального питания./ Д. А. Алиев, М. А. Юсифов. // Тр. Азербайджанского НИИ земледелия, -1981 Т.17. - С. 94-97.
6. Алиев Д. А. Фотосинтетическая деятельность, минеральное питание и продуктивность растений. / Д. А. Алиев,- Баку, 1974.-332 с.
7. Андрианова Ю. Е. Хлорофилл и продуктивность растений / Ю. Е. Андрианова, И. А. Тарчевский. М. : Наука, 2000. - 135 с.
8. Андреева Т.Ф. фотосинтез и азотный обмен растения / Т. Ф. Андреева // Физиология фотосинтеза. М., 1982. - С. 89 - 104.
9. Антыков А. Я. Почвы Ставрополья и их плодородие /А. Я. Алтыков, А. Я. Стоморев. Ставрополь. - 1970. - 414 с.
10. Апраксина С. М. Комплексообразующая способность веществ гуминовой природы в почве / С. М. Апраксина и др. // Химия в сельском хозяйстве. 1994. - № 5. - С. 8-10.
11. Бабенко В. И. Роль листьев различных ярусов в создании урожаев зерна у высоко и низкопродуктивных сортов пшеницы / В. И. Бабенко, Л. П. Колесник, С. В. Бирюков // Сельскохозяйственная биология. - 1977. Т. 12. -№2.-С. 198-203.
12. Бабенко В. И. Зависимость морозостойкости растений озимой пшеницы от степени их яровизации. / В. И. Бабенко, С. В. Бирюков // Методы и приемы повышения зимостойкости озимых зерновых культур./ М. : Колос, 1975.-С. 339-345.
13. Бадахова Г.Х. Ставропольский край: современные климатические условия / Г.Х. Бадахова, А.В. Кнутас. Ставрополь, ГУП СК Краевые сети связи, 2007.-272 с.
14. Баранина И. И. Содержание пигментов у озимой пшеницы при внекорневой подкормке мочевиной / И. И. Баранина, И. М. Унку ; под общ. ред. С. И. Тома; Макро и микроэлементы в регуляции обмена веществ растений. -Кишинев, «Штиинца», 1983. 99 с.
15. Баталкин Г. А. Гуминовые удобрения: Теория и практика их применения / Г. А. Баталкин и др. / Днепропетровск . 1980. - Т.7 - С. 67 - 73.
16. Батыгин Н. Ф. Онтогенез высших растений / Н. Ф. Батыгин. М. : Агропромиздат, 1986. - 100 с.
17. Бебякин В. М. Качество зерна пшеницы в зависимости от сорта и условий его выращивания / В. М. Бебякин, Н. И. Старичкова, А.А. Дорогобед // Зерновое хозяйство. 2003. - № 3. С. 22 - 24.
18. Бебякин В. М. Корреляционно-факторный анализ показателей качества зерна озимой пшеницы / В. М. Бебякин, Г. В. Пискунова, В. А. Матвеев // Зерновое хозяйство. -2003. 38.-С. 17-19.
19. Беденко В. П. Фотосинтез и продуктивность пшеницы на Юго Востоке Казахстана / В. П. Беденко. - Алма-Ата: Изд-во «Наука», 1980.224 с.
20. Безуглов В. Г. Эффективность удобрений, содержащих гумат натрия в баковых смесях с гербицидом на посевах озимой пшеницы / В. Г. Безуглов, Ф. М. Гафуров // Агрохимия. 2002. - № 9. - С. 41-46.
21. Безуглов В. Г. Микроэлементы с макроудобрениями под озимую пшеницу на почвах Северного Кавказа / В. Г. Безуглов, Ф. М. Гафуров // Аграрная наука. 2004. - № 1. - С. 23-24.
22. Бирюков С. В. Характер формирования зерновки у различных генотипов озимой пшеницы / С. В. Бирюков, В. П. Комарова // Репродуктивный процесс и урожайность полевых культур : Сб. науч. тр. / ВСГИ. 1981. -С. 19-26.
23. Бирюков С. В. Аттракция экзогенной сахарозы колосьями различных генотипов озимой пшеницы / С. В. Бирюков, В. П. Комарова // Вопросы селекции и генетики зерновых культур. М, 1983. С. 169 - 179.
24. Благовещенский А. В. Теоретические основы действия янтарной кислоты на растения / А. В. Благовещенский. М.: Изд-во «Наука», 1968. -117 с.У
25. Бобрышев Ф. И. Озимая пшеница в Ставропольском крае / Ф. И. Бобрышев, А. И. Войсковой, В. В. Дубина и др. / Ставрополь, Изд. СтГАУ «Агрус», 2003. 307 с.
26. Бобырь J1. Ф. О донорно акцепторных свойствах гумусовых кислот низинного торфа / JI. Ф. Бобырь, Л. А. Епимина // Физико - химия торфа : тез. докл. совещ. - Минск, 1976. - С. 70-71.
27. Булли В. А. Исследование биологической активности гуматов на сельскохозяйственных культурах / В. А. Булли, А. Л. Антонова, Н. А. Олей-ник // Химия в сельском хозяйстве. 1995. - №5. - С. 10-12.
28. Бутковский В. А. Требования к мукомольным и хлебопекарным качествам пшеницы / В. А. Бутковский // Зерновое хозяйство. 2003. - № 8. -С. 8-10.
29. Бутковский В. А. К вопросу о показателях мукомольных и хлебопекарных качеств пшеницы / В. А. Бутковский // Зерновое хозяйство. 2003. -№9.-С. 8-10.
30. Бухар Е. И. Пути повышения урожая и качества зерна озимой пшеницы в Молдавии / Е. И. Бухар. Кишинев. : Штиинца, 1983. - 109 с.
31. Вакар А. Б. Белковый комплекс клейковины. / А. Б. Вакар// : Сб. Растительные белки и их биосинтез. М., Наука. -1975.
32. Виноградова В. С. Влияние трофических и гормональных регуляторов на формирование урожая / В. С. Виноградова, Ю. В. Смирнова // Агрохимический вестник. 2002. - № 1. - С. 19-20.
33. Влияние янтарной кислоты на продуктивность сельскохозяйственных растений, урожай и его качество / Ю. Е. Андрианова, Н. И. Сафина, Н. Н. Максютова, М. Г. Кадошникова // Агрохимия. 1996. - №8-9. - С.18-123.
34. Волынкина О. В. Влияние густоты посева и средств химизации на урожайность и качество зерна яровой пшеницы / О. В. Волынкина, В. И. Во-лынкин // Агрохимия. 2003. - № 5. - С. 48-54.
35. Горовая А. И. Роль физиологически активных гумусовых препаратов в ругуляции процессов клеточного цикла / А. И. Горовая, А. Ф. Кулик, И. А. Огинава // Сб. науч. тр. / Киев. 1985. : Регуляция клеточного цикла. -С. 101-109.
36. Горышина Т. К. Фотосинтетический аппарат растений и условия среды / Т. К. Горышина. Ленинград : Изд-во Ленинградского университета, 1989.-202 с.
37. ГОСТ Р 52554 2006. / Пшеница. Технические условия/ М.: Стан-дартинформ, 2006. - 9 с.
38. Гуляев Б. И. Условия выращивания и показатели продуктивности сортов яровой пшеницы / Б. И. Гуляев, Ю. П. Шалин, А. Ю. Шалин // Физиология и биохимия культурных растений. 1989. - Т. 15. - Вып. 1. - С. 65- 72.
39. Деева В. П. Избирательное действие химических регуляторов роста на растения : Физиологические основы / В. П. Деева. Минск «Наука и техника». 1988. - 172-173 с.
40. Дёфлинг К. Гормоны растений. / К. Дёфлинг // Системный подход / под общ. ред. В. И. Кеорели. М., 1985. - С. 282-304.
41. Довнар В. С. Некоторые закономерности изменения продуктивности фотосинтеза и оптимальной площади листьев у кукурузы в Белоруссии/ В. С. Довнар // Важнейшие проблемы фотосинтеза в растениеводстве. М., «Колос». - 1970. - С. 298-316.
42. Долгушин Д. А. Стадия яровизации и некоторые биологические особенности современных сортов озимой пшеницы на Юге СССР / Д. А. Долгушин // Вестник сельскохозяйственной науки. 1980. - № 4. - С. 46-52.
43. Дорохов Б. JI. Зимний газообмен озимой пшеницы при различном минеральном питании / Б. JI. Дорохов, И. И. Баранина // Фотосинтез и пигменты основных сельскохозяйственных растений Молдавии. Кишинев, — 1976. С. 26-30.
44. Дорохов Б. JI. Оптические свойства, интенсивность и КПД фотосинтеза листьев фасоли при усилении минерального питания. / Б. JI. Дорохов, А. Г. Жакотэ // Биохимия и биофизика фотосинтеза :Сб. науч. тр./ Иркутск, - 1971. - С. 223-228.
45. Доспехов Б. А. Методика полевого опыта/ Б. А. Доспехов М.: Колос, 1979.-416 с.
46. Драганцев В. А. Экологический скрининг генофонда и методы конструирования сортов сельскохозяйственных растений по урожайности, устойчивости и качеству / В. А. Драганцев/ Методические рекомендации. СПб. : ВИР, 1997.
47. Евсеева Р. П. Технология возделывания озимых зерновых / Р. П. Евсеева // Суперагро. : Озимые зерновые. «МП». 2000. - 324 с.
48. Егоров Г. А. Технологическая характеристика зерна / Г. А. Егоров // Зерновое хозяйство. 2002. - № 7. - С. 28-31.
49. Ермаков А. А. Эффективность некорневых подкормок микроэлементами земляники, выращиваемой на разных агрохимических фонах / А. А. Ермаков // Агрохимический вестник. 2003. - № I. - С. 32-33.
50. Ерошенко Ф. В. Замедленная флуоресценция и фотосинтетическая продуктивность сортов озимой пшеницы / Ф. В. Ерошенко // Физиология озимой пшеницы при интенсивной технологии возделывания : Сб. науч. тр. / СНИИСХ. Ставрополь, 1992. - С. 62-71.
51. Ерошенко Ф. В. Особенности фотосинтетической деятельности сортов озимой пшеницы / Ф. В. Ерошенко. Ставрополь, 2006. - 198 с.
52. Животков Л. А. Пшеница / Л. А. Животков и др.. Киев : Урожай, 1989.-320 с.
53. Ивакин А. П. Фотосинтетическая деятельность и формирование урожая озимой пшеницы при различной густоте посева в условиях орошения : автореф. дис. . канд. биол. наук / Ивакин Алексей Петрович. Ленинград: 1970.-С. 20.
54. Исайчев В. А. Фотосинтетическая деятельность растений озимой пшеницы в зависимости от предпосевной обработки семян пектином и микроэлементами / В. А. Исайчев, Ф. А. Мударисов // Зерновое хозяйство. 2003. -№ 7. - С. 19-21.
55. Исайчев В. А. Влияние предпосевной обработки семян микроэлементами на фотосинтетическую деятельность посевов яровой пшеницы и сои / В. А. Исайчев А. В. Дозоров // Зерновые культуры. 1999. - № 6. - С. 12- 13.
56. Казарцева А. Т. Обесцвеченное зерно / А. Т. Казарцева, Р. А. Воробьева, М. И. Демченко // Зерновое хозяйство. 1989. - № 7. - С. 7-8.
57. Казарцева А. Т. Где теряем качество товарного зерна / А. Т. Казарцева, Н. В. Бородулина //Зерновое хозяйство. -1988. -№ 6. С. 35.
58. Калиненко И. Г. Новый сорт мягкой озимой пшеницы Донская полукарликовая. / И.Г. Калиненко, С.Н. Прищепов // Селекция и семеноводство зерновых и кормовых культур. :Сб. науч. тр./ -Зерноград, —1980. -С. 3—6.
59. Квасов Н. А. Применение гумата натрия и азотных удобрений с целью повышения урожая и качества зерна озимой пшеницы / Н. А. Квасов // Пути повышения качества зерна сельскохозяйственных культур : Сб. науч. тр. СНИИСХ. Ставрополь, 1999. - С. 111-116.
60. Керимханов С. У. Азотные удобрения под озимую пшеницу / С. У. Керимханов, К. М. Мамедгусейнов, В. В. Совоцкий // Химия в сельском хозяйстве. 1985. - № 10. - С. 26-27.
61. Климашевский Э. JI. Проблема генотипической специфики корневого питания / Э. Л. Климашевский // Сорт и удобрение. Сб. науч. тр. Иркутск, 1974.-С. 27-30.
62. Клочкова Н. М. СО2 газообмен растений гороха посевного различных морфотипов под действием янтарной кислоты и эпина при ранней корневой засухе / Н. М. Клочкова, Э. Н. Аканов, Н. Н. Третьяков // Сельскохозяйственная биология. - 2004. - №1. - С. 67-72.
63. Кириченко Ф. Т. Методы и результаты селекции прочностебельных и полукарликовых сортов озимой пшеницы для степных районов УССР / Ф. Т. Кириченко, С. Ф. Лыфенко// Селекция короткостебельных пшениц.: Сб. науч. тр./ М„ -1975. - С. 39-47.
64. Кириченко Ф. Т. Формирование урожая и накопление белка у сортов и форм озимой пшеницы. / Ф. Т. Кириченко, Н. А. Литвиненко // Вестник сельскохозяйственной науки. —1982. —№ 10. —С. 36—40.
65. Колмаков Ю. В. Обесцвеченное зерно / Ю. В. Колмаков, А. Н. Ти-мошкин, А. И. Бирюков // Зерновые культуры. 1996. - №1. - С. 11-13.
66. Комиссаров И. Д. Электронный парамагнитный резонанс гуминовых кислот / И. Д. Комиссаров, Л. Ф. Логинов// Гуминовые препараты : Сб. науч. тр. / ТСХИ. Тюмень, 1982. - Т.14. - С. 99-115.
67. Кондрашов А. Г. Гумат калия торфяной жидкий / А. Г. Кондрашов // Агрохимический вестник. - 2000. - 2.- С. 40.
68. Коробской Н. Ф. Отзывчивость сортов озимой пшеницы на минеральные удобрения / Н. Ф. Коробской, М. X. Ширинян, Н. Д. Кравцова // Агрохимия. 1997. - № 6. - С. 23-24.
69. Костин В.И. Динамика ростовых процессов озимой пшеницы в зависимости от обработки семян пектином и микроэлементами / В. И. Костин, В. А. Исайчев, Ф. А. Мударисов // Зерновое хозяйство. 2003. - № 4. - С. 1314.
70. Коф Э. М. Антистрессовое действие янтарной кислоты на проростки гороха / Э. М. Коф, Т. А. Борисова // Агрохимия. 1999. - № 8-9. - С. 118 -123.
71. Кошкин В. А. Влияние генов Ppd на хлорофилл-белковый комплекс сортов пшеницы с различной фотопериодической чувствительностью / В. А. Кошкин, И. А. Косарев, В. А. Драгавцев, И.И. Матвиенко // Доклады Россельхозакадемии. 1999. - № 4. - С. 6-7.
72. Крылов Е. А. Рациональное использование отходов / Е. А. Крылов и др. // Агрохимический вестник. 2003. - № 3. - С. 31-33.
73. Кузьмич М. А. Влияние гуминовых веществ на почву и растения / М.
74. A. Кузьмич // Агрохимия. 1990. №4. - С. 146-149.
75. Кумаков В. А. Влияние засухи на фотосинтетическую деятельность яровой пшеницы / В. А. Кумаков, К. М. Кузьмина, И. Ф. Алешин // Вопросы ботаники Юго-Востока. Саратов, 1975. Вып. 1. - С. 7-11.
76. Кумаков В. А. Коррелятивные отношения между органами растения в процессе формирования урожая / В. А. Кумаков // Физиол. Раст. 1980. -Т. 27. - Вып.5. - С.975-985.
77. Кумаков В. А. Фотосинтетическая деятельность в аспекте селекции /
78. B. А. Кумаков: в кн.: Физиология фотосинтеза. М., 1982. С.283-293.
79. Кумаков В. А. Анализ фотосинтетической деятельности растений и физиологическое обоснование модели сорта. В сб.: Фотосинтез и продукционный процесс /В. А. Кумаков // М.: Наука, 1988. С.247-251.
80. Кумаков В.А. Значение реутилизации в наливе зерна различных сортов яровой пшеницы. / В.А. Кумаков, Н.Ф. Матвеева, С.С. Павлова и др.// Доклады ВАСХНИЛ. -1979. -№ 8. С. 5-7.
81. Кумаков В. А. Оценка роли отдельных органов в наливе зерна пшеницы и ее селекционные аспекты./ В. А. Кумаков, А. П. Игошин, Б. В Бере-зин, Г. Д. Левина // Физиология и биохимия культурных растений. -1983, Т. 15. -№ 2. -С.163-169.
82. Кумаков В. А. Физиология яровой пшеницы. / В. А. Кумаков —М.: Наука, -1980. -103 с.
83. Кумаков В. А. Физиология формирования урожая яровой пшеницы и проблемы селекции/ Кумаков, В. А. // Сельскохозяйственная биология. -1995.-№ 5. С. 3-18.
84. Кумаков В. А. Эволюция показателей фотосинтетической деятельности в процессе селекции яровой пшеницы. / В. А. Кумаков // Теоретические основы фотосинтетической продуктивности. :Сб. науч. тр./ М., — 1972. -С. 500-503.
85. Кумаков В. А. Распределение сухого вещества между органами в связи с продуктивностью и засухоустойчивостью сортов пшеницы / В. А. Кумаков, О. А. Евдокимова, М. А. Буянова // Физиология растений. 2001. Т.48. -№ 3. - С. 421-426.
86. Князев Ю. В. Гумату «плодородие» — дорогу в производство / Ю. В. Князев // Агрохимический вестник. 2002. - № 5. - С. 27-28.
87. Куперман Ф. М. Морфофизиология растений / Ф. М. Куперман // М.: Высшая школа, 1984. 240 с.
88. Куприченков М. Т. Агротехника. Плодородие. Урожай / М. Т. Куприченков, В. И. Каргальцев. Ставрополь, 1988. - 111 с.
89. Куприченков М. Т. Плодородие почв и их пригодность для возделывания кормовых культур / М. Т. Куприченков, Л. Н. Петров // Система кормопроизводства Ставропольского края. Ставрополь, 1984. - С. 26-32.
90. Куприченков М. Т. Земельные ресурсы Ставрополья и их плодородие / М. Т. Куприченков, Т. Н. Антонова, Н. Ф. Симберев, А. С. Цыганов Ставрополь 2002 г., - 320 с.
91. Куприченков М. Т. Справочник по плодородию почв // М. Т. Ку-приченков., Ставрополь: Сервисшкола, 2007 г., 144 с.
92. Куркаев В. Т. Сельскохозяйственный анализ и основы биохимии / В. Т. Куркаев, С. М. Ерошкина, А. Н. Понамарев М.: Колос, 1977. - 240 с.
93. Курсанов A. JI. Транспорт ассимилятов в растении. // A. JI. Курса-нов -М.: Наука, -1976. 646 с.
94. Кухаренко Т. А. Окисление в пластах бурые угли. М., 1972. - 102с.
95. Кыдралиева К. А. Биотестирование продуктов анаэробной ферментации / К. А. Кыдралиева, Ш. Ж. Жеробекова, Н. В. Горбунова // Агрохимический вестник. 2003. - №2. - С. 36-38.
96. Литтл Т. М. Сельскохозяйственное опытное дело. Планирование и анализ / Т. М. Литтл, Ф. Дж. Хиллз; пер. с англ. Б. Д. Кирюшина; под ред. Д. В. Васильевой. -М.: Колос, 1981.-320 с.
97. Лукъяненко П. П. Селекция низкостебельных сортов озимой пшеницы, для условий орошения. / П. П. Лукъяненко // Вестник сельскохозяйственной науки. -1973. —№ 1. С. 8-15.
98. Лукьяненко П. П. О селекции низкостебельных сортов озимой пшеницы. / П. П. Лукьяненко // Селекция и сортовая агротехника озимой пшеницы. :Сб. науч. тр./-М.,-1971,-С. 13-24.
99. Лукьяненко П. П. Состояние и перспективы работ по селекции низкостебельных сортов озимой пшеницы для условий орошения. / П. П. Лукьяненко // Сб. научн. тр./ ВАСХНИЛ. -М., -1975, -С. 6-18.
100. Луценко Э. К. Возможные изменения качества зерна в зависимости от расположения в пространстве листового аппарата озимой пшеницы приинтенсивной технологии возделывания / Э. К. Луценко, О.И. Нешин // Сб. науч. тр. СНИИСХ. Ставрополь, 1992. - С. 159-164.
101. Лучник Н. А. Испытание гумата «Плодородие» в регионах / Н. А. Лучник // Агрохимический вестник. —2002. —№1. — С. 21-22.
102. Лучник Н. А. Испытание гумата «Плодородие» в Костромской области / Н. А. Лучник // Агрохимический вестник. 2002. —№1. - С. 6-13.
103. Лучник Н. А. Гумат натрия на посевах зерновых культур / Н. А. Лучник, А. Е. Иванов, А.И. Меркулов // Химия в сельском хозяйстве. -1997. -№2. -С. 28.
104. Макаров Р. Ф. Влияние удобрений на урожайность и качество мягкой пшеницы / Р.Ф. Макаров, В.В. Архипова // Зерновые культуры. 1999. -№2.-С. 25-26.
105. Максютова Н. Н. Белковый обмен растений при стрессе. Автореферат. дисс.докт. биол. наук-М., 1998. - С. 22 - 25.
106. Мартьянова А. И. Оценка технологических свойств товарных партий пшеницы / А. И. Мартьянова и др.. М. : Агропромиздат, 1986. 152 с.
107. Мартьянова А. И. Технологические свойства озимой и яровой мягкой пшеницы / А. И. Мартьянова, Т. А. Леонова, А. В. Лушина, Е. П. Меми-кина // Зерновое хозяйство. 2002. - № 8. - С. 9-10.
108. Мартьянова А. И. Пробная лабораторная выпечка хлеба прямой и надежный способ оценки качества зерна пшеницы / А. И. Мартьянова, Е. П. Пищугина // Зерновые культуры. - 1996. - № 1. - С. 11-13.
109. Медведев Н. П. Применение консерванта МиБАС К / Н. П. Медведев, М. П. Крысин, В. Р. Кочетов // Химия в сельском хозяйстве. - 1997. -№ 1. - С. 37-38.
110. МилаёваЯ. П. Сравнительное определение количества пигментов в листьях кукурузы и табака ускоренным методом / Я. П. Милаёва, И. П. Примак // Селекция и семеноводство. Киев, 1969. - Вып. 12. - С. 69-72.
111. Минеев В. Г. Агротехнические основы повышения качества зерна озимой пшеницы. / В. Г. Минеев, А. Н. Павлов — М.: Колос, 1981. — 342 с.
112. Мотовилова JI. В. Гуматы экологически чистые стимуляторы роста и развития растений / JI. В. Мотовилова, О. Н. Берман // Химия в сельском хозяйстве. - 1994. -№ 5. - С. 12-13.
113. Нальборчик Т.Э. Роль различных органов фотосинтеза в формировании урожая/ Т.Э. Нальборчик // Адаптивные реакции в формировании и активности фотосинтетического аппарата. Пущино, НЦБИ АН СССР, 1980. -С. 22-23.
114. Николаев Е. В. Технология выращивания сильной озимой пшеницы / Е. В. Николаев. Симферополь : Таврия, 1986. - 94 с.
115. Ничипорович А. А. (К 80-летию со дня рождения) Потенциальная продуктивность растений и принципы оптимального ее использования / А. А. Ничипорович //Сельскохозяйственная биология. —1979.—Т. 14. —№ 6, -С. 683-694.
116. Ничипорович А. А. Теория фотосинтетической продуктивности растений / А. А. Ничипорович // Итоги науки и техники. Серия Физиология растений. Теоретические основы повышения продуктивности растений. — М., -1977.-Т. 3.-С. 11-54.
117. Ничипорович А. А. Физиология фотосинтеза и продуктивность растений / А. А. Ничипорович // Физиология фотосинтеза.: Сб. науч. тр./ —М.: Наука, -1982. -С.7-34.
118. Ничипорович А. А. Фотосинтетическая деятельность растений и пути повышения их продуктивности./ А. А. Ничипорович //Теоретические основы фотосинтетической продуктивности. : Сб. науч. тр./ —М., -1972. — С. 511-527.
119. Нургалиева Р. В. Влияние гуми М на гормональный статус растений пшеницы при засолении / Р. В. Нургалиева и др. // Агрохимия. 2006. -№ 8 - С. 25-29.
120. Овчаренко М. М. Гуматы активаторы продуктивности сельскохозяйственных культур / М. М. Овчаренко // Агрохимический вестник. - 2002. -№2.-С. 13-14.
121. Орлов Д. С. Гуминовые кислоты почв / Д. С. Орлов М.: Изд-во МГУ, 1974. - 56 с.
122. Орлова И. Г. Оценка состояния озимых культур в процессе их вегетации и уход за посевом (с основами биологии развития озимой пшеницы) / И. Г. Орлова. Ставрополь : Изд-во СГУ, 2002. - 20 с.
123. Павлов А. Н. О причинах определяющих различный уровень накопления белка в зерне высоко- и низкобелковистых сортов пшеницы./ А. Н. Павлов, Т. И. Колесник// Физиология растений. —1974. -Т. 21 Вып. 2. -С. 329-335.
124. Павлов А. Н. Повышение белка в зерне. / А. П. Павлов М.: Наука, -1984.-119 с.
125. Павлов А. Н. Состояние и перспективы изучения действия удобрений на качество зерна. / А. Н. Павлов, В. Г. Минеев // Агрохимия-1982. -№ 1. — С. 134-141.
126. Петров Г. И. Влияние агроклиматических условий на формирование урожая озимой пшеницы в сухостепной полосе Ставрополья / Г. И. Петров. -Буденновск : Прикумье, 1996. - 334 с.
127. Петрова JI. Н. Производство сильного зерна пшеницы в Ставропольском крае и пути его увеличения / Л. Н. Петрова, А. Я. Чернов, Е. А. Прокудин// Пути повышения качества зерна сельскохозяйственных культур. : Сб. науч. тр./ — Ставрополь, -1999. -С. 5-10.
128. Петрова Л. Н. Урожай и качество зерна озимой пшеницы Безостая 1 при различных сроках внесения фосфорных внекорневых подкормок. / Л. Н. Петрова, В. Д. Костин // Тр. СНИИСХ. -Ставрополь -1999,- С. 107-111.
129. Петрова Л. Н. Физиолого-биологические основы питания и формирования урожая озимой пшеницы: Автореф. дис. . канд. биол. наук. / Л. Н. Петрова. Рига, -1971. -23 с.
130. Петрушин В. В. Эффективность гуминовых препаратов при выращивании сельскохозяйственных культур / В. В. Претрушин и др. // Агрохимический вестник. 2001. - №2. - С. 13-14.
131. Пикуш Г. Р. Некоторые особенности биологии кущения озимой пшеницы / Г. Р. Пикуш // Повышение продуктивности озимой пшеницы : Сб. науч. тр. Днепропетровск, 1980. - С. 22-29.
132. Пшцугина Н. С. Почвы экспериментального поля СНИИСХ / Н. С. Пищугина // Использование земельных ресурсов и пути повышения плодородия почв. : Сб. науч. тр./ Ставрополь, -1979. -Вып. 42. - С. 42-47.
133. Покинбара В. А. Испытание гуминовых препаратов / В. А. Покин-бара, С. В. Довыденко, Т. О. Скородумова // Агрохимический вестник. 2001. - № 2. - С. 9-11.
134. Попов Г. Н. Микроудобрения на орошаемых землях / Г. Н. Попов, Б. В. Егоров М.: Россельхозиздат, 1987. - 48 с.
135. Портуровская С. Н. Влияние минеральных удобрений на фотосинтетическую деятельность растений озимой пшеницы размещаемой после кукурузы на силос. / С. Н. Портуровская, Н. М. Шахзадов // Тр./ Ставропольский с.-х. ин-т. -1972. -Т. 1. -Вып. 35. -С. 20-23.
136. Прокудин Е. А. Изменчивость компонентов структуры урожая озимой пшеницы интенсивных сортов в зависимости от агрофона. / Е. А.Прокудин // Тр. СНИИСХ. -Ставрополь, -1977 -Вып. 36.-С. 15-18.
137. Прокудин Е. А. Условия повышения продуктивности озимой пшеницы. / Е. А. Прокудин, И. В. Нешин, О. П. Покатаева // Роль молодых ученных и специалистов, членов НТО в реализации продовольственной программы. :Сб. науч. тр./ Зерноград, -1982. - С. 28-30.
138. Прядкина Г. А. О связи между содержанием хлорофилла в отдельном листе и целом растении кукурузы в онтогенезе / Г. А. Прядкина // Сельскохозяйственная биология. 1999 - № 5. - С. 28-31.
139. Раскин М. С. Все о гуматах / ArpoXXI. 2000. - 11. - С. 16.
140. Ремесло В. Н. О селекции короткостебельных сортов для условий лесостепи Украины. / В. Н. Ремесло // Селекция короткостебельных пшениц : Сб. науч. тр./ М., -1975. - С. 19-27.
141. Рубин А. Б. Первичные процессы фотосинтеза и фотосинтетическая продуктивность / А. Б. Рубин, Т. Е. Кренделева, П. С. Венедиктов, Д. Н. Ма-торин // Сельхоз. биол. 1984. - №6. - С. 81 - 92.
142. Русеева 3. М. Агроклиматические ресурсы Ставропольского края (справочник). / 3. М. Русеева-Jl.: Гидрометеоиздат, —1974. -238 с.
143. Сандухадзе Б. И. Качество зерна озимой пшеницы / Б. И. Сандухад-зе, Е. Б. Егорова // Зерновое хозяйство. 2002. - №4. - С. 18-20.
144. Санькова Л. Н. Влияние погодных условий на развитие озимой пшеницы в Ставропольском крае./ Л. Н. Санькова // Озимая пшеница на Ставрополье. :Сб. науч. тр./- Ставрополь, -1981- С. 59-67.
145. Сахибгареев А. А. Обработка семян ярового ячменя микроэлементами / А. А. Сахибгареев, Т. А. Гаитов // Агрохимический вестник. 1999. -№5.-С. 24-25.
146. Середа Н. А. Эффективность макро- и микроудобрений на яровой пшенице сорта Башкирская 24 / Н. А. Середа, В. И. Никонов // Зерновые культуры. 2000. - №3. - С. 20-23.
147. Созинов А. А. Повышение качества зерна озимых пшениц / А. А. Созинов, В. Г. Козлов. -М.: Колос, 1970. 130 с.
148. Стрельникова М. М. Повышение качества зерна пшеницы/ М. М. Стрельникова. Киев, Урожай. - 1971.
149. Тарчевский И. А. К вопросу о передвижении ассимилятов у пшеницы и влиянии минерального питания на этот процесс. / И. А.Тарчевский, А. П. Иванова, У. А. Бектимиров // Тр. Биолого-почвенный ин-т. -Т. 20. —1973. -С. 174-178.
150. Тарчевский И. А. Основы фотосинтеза. / И. А. Тарчевский Казань, 1971.-246 с.
151. Тарчевский И. А. Основы фотосинтеза. / И.А. Тарчевский —М.: Высшая школа., 1977. -246 с.
152. Тарчевский И. А. Особенности фотосинтеза, оттока ассимилятов у различных сортов яровой пшеницы. / И. А. Тарчевский, А. Н. Иванова, В. И. Чиков, А. П. Сулейманова, Ю. Е. Андрианова //Тр. Татарский НИИСХ. Казань, -1974. -Вып 4. - С. 315-319.
153. Тарчевский И. А. Содержание пигментов, как показатель мощности развития фотосинтетического аппарата у пшеницы. / И. А. Тарчевский, Ю. Е. Андрианова// Физиология растений. -1980. -Т. 27. -№ 2. С. 341-347.
154. Тарчевский И. А. Янтарная кислота миметик салициловой кислоты / И. А. Тарчевский, Н. Н. Максютова, В. Г. Яковлева, А. Н. Гречкин // Физиология растений. - 1999. - Т.46. -№ 1. - С. 23-28.
155. Тооминг X. Г. Перспективы прогноза эффективности изменения параметров растений и оценки максимального урожая. / Х.Г.Тооминг // Программирование урожаев сельскохозяйственных культур. —М.: Колос, —1975. -С.403-414.
156. Удовенко Г. В. Методические рекомендации по проведению оценки атракции. / Г. В. Удовенко -М.: ВИР, 1992. 9 с.
157. Удовенко Г. В. Ростовая и аттрактивная активность пшеницы при разных терморежимах / Г. В.Удовенко, В. А. Драгавцев, А. А. Степанова, 3. А. Щедрина // Доклады Российской академии наук. 1998. - № 4. — С. 5-6.
158. Хотулев В. Я. Связь содержания хлорофилла с фотосинтетической продуктивностью короткостебельных и длинностебельных сортов озимой пшеницы. : Автореф. дис. . канд. биолог, наук. / В. Я. Хотулев. Казань, -1985.-19 с.
159. Христева JI. А. Действие физиологически активных гуминовых кислот на растение при неблагоприятных условиях / JT. А. Христева // Гумино-вые удобрения: теория и практика их применения. — Днепропетровск. 1973. - Т.4 - С. 5-24.
160. Христева JI. А. Гуминовые удобрения : теория и практика их применения / Л. А. Христева. Днепропетровск 1977. - Т.4 - С. 13-14.
161. Хьютсон А. Дисперсионный анализ / А. Хьютсон ; пер с англ. А. Г. Кругликова. -М.: Статистика, 1971. 88 с.
162. Церлинг В. В. Диагностика питания сельскохозяйственных культур. /В. В.Церлинг // Справочник М.: Агропромиздат, -1990 - 235 с.
163. Церлинг В. В. Методические указания по комплексной диагностике зерновых культур./ В. В. Церлинг М.: Колос, 1983.-326 с.
164. Чернышева Н. Ф. Факторы отзывчивости разных сортов растений в связи с их взаимодействием с удобрениями/ Н. Ф. Чернышева, Э. Л. Климашевский // Агрохимия. 1979. - № 6. - С. 57-65.
165. Чумаченко И. Н. Перспективы применения микроудобрений / И. Н. Чумаченко, В. А. Прошкин, Н. В. Войтович // Химия в сельском хозяйстве. 1995. - № 6. - С. 22-26.
166. Шафронов О. Д. Перспективный стимулятор роста растений «Гармония» / О. Д. Шафронов, В. Н. Полухин // Химия в сельском хозяйстве. -1994,-№5.-С. 13-14.
167. Шматысо И. Г. Устойчивость зеленых пигментов к водному дефициту и повышенным температурам / И. Г. Шматько, А. И. Шаповал,
168. Н. В. Шевчук // Методы оценки устойчивости растений к неблагоприятным условиям среды. М., - 1976. - 124 с.
169. Шпаар Д. Зерновые культуры/ Д. Шпаар, Ф. Эллмер, А. Постников, Н. Протасов и др.//Мн: ФУАинформ,. 2000. С. - 421.
170. Якубцинер М.М. Использование мировой коллекции в селекции озимой пшеницы / М.М. Якубцинер // Селекция и сортовая агротехника озимой пшеницы. -М., 1971. С. 60-74.
171. Arnon D. I. Regulatory electron transport pathways in cyclic photophos-phorylation. Reduction of c-550 and cytochrom Ьб by feredoxin in a dark / D. I. Arnon, R. IC.Chain // FEBS Lett. 1979. - Vol.- P. 133 - 138.
172. Anderson J.M. Consequence of higher plant chloroplast / J.M. Anderson //FEBS Lett. Vol. 124 - P. 1 - 10.
173. Aufhammer W. Das Sheichervermogen von Weizenahren in Abhangig-keit von Beziehungen zwischen den einzelnen Kornanlagen / W. Aufhammer, P. Zinsmaier // Ztsclir. Acker und Pflanztnbau. - 1982. - Bd/ 151 - H.4. - S. 249 -266.
174. Barber J. Membrane surface charges and potential in relation to photosynthesis / J. Barber // Biochem. et biophys. Acta. 1980. - Vol. 594. - P. 253 -308.
175. Burns R. G. Humus substances effect on soil and plants / R. G. Burns, G. DelFagnola, S. Miele, S. Nardis, G. Savoini, M. Sehnitzer, P. Segui, P. Vauch-gan, S. A. Visser/Reda edizioni perragricolturla. 1986. 170 p.
176. Galili D. R. Translational control of cellular and viral mRNAs / D. R. Galili // Plant Mol. Biol. 1996. V. 32. - P. 145 - 158.
177. Gerola P.D. Thilacoid membrane staking: structure and mechanism / P.D. Gerola // Physiol, veget. 1981. - Vol. 19. - P. 565 - 580.
178. Hall A. I. Assimilate source sink relation ships in Capsicum annum L. II Effects of fruiting and defloration on the photosynthetic capacity and senescence of the leaves/ A. I. Hall // Austral. J. Plant Physiol., 1977. - 4,- № 5. - P. 771-783.
179. Herold A. Regulation of photosynthesis by sink activity. The missing link/A. Herold// New Phytol., 1980. 86. - № 2. - P. 131-144.
180. Hole С. С. Effect of number and configuration of fruits photon flux and age on the growth and dry matter distribution of fruits of Pisum sativum L./ С. C. Hole, P. N. Skott // Plant, Cell and Environ., 1983. Vol. 6, № 6. - P. 31-38.
181. Jurcsilc I. Bot. koze /1. Jurcsik. 1984. Vol. 71. - № 3 - 4. P. 295-332.
182. Kazarda D. D. Structure and properties of a-gliadins / D. D. Kazarda // Ann. Technol/ Agr. 1980. V.29. N 2. P. 151 173.
183. Kozlowska-Ptaszynska, Glazewski, Z. Wplyw wielkosci powiorzchni asymilacyjnoj I intensywnosci fotosyntezy na process gromadzenia suchej masy w ziarnie hszenicy ozimej / Z. Kozlowska-Ptaszynska, Glazewski // Pamietnik Pulaw. 1975.-64. P. 149- 166.
184. Kruger J.E. Some experiences with monitoring alpha amylase levels in Canadian wheats / J.E. Kruger, K.H. Tippies // 7th. World cereal and bread congress. Abstracts, Praha 1982. - P. 200.
185. Loomis W.F. Cell Sci // W.F. Loomis, G. Shaulsry, N. Wang/ 1997. -V. llO.Pt.lO.P. 1141-1145.
186. Mae T. The remobilization of nitroden related to leaf growth and senescence in rice hlants (Oiyza sativa L.) / Т. Mae, K. Ohira // Plant and Cell Physiol. -1981. Vol. 22. -№ 6. - p. 1067 - 1074.
187. Osman A. M. Photosynthesis of wheat leaves in relation to age, illuminance and nutrient supply / A. M. Osman, F. L. Milthorpe // II Results, Photosyn-thetica, 1975. - V.5. -№ 1. - P. 61-70.
188. Savoini G. Responsabile recerca e sviluppo kometek S.A. Australian Journal of Agricultural Research, 1986. 50 p.
189. Walker D. A. // In: The intact chloroplast / Ed. J. Barber/ Amsterdam etc.: Elsevier, 1976. P. 235 278.
www.dissercat.com
Питание и урожайность пшеницы. Рост и развитие здорового растения
Продолжаем тему ограничивающих факторов урожая пшеницы. Рассмотрим взаимосвязь надземной части растения и корней, влияние денитрификации и ограничивающую роль в достижении урожая основных питательных веществ – макро- и микроэлементов(Продолжение, начало см. в №№7, 8, 9/2011)
Взаимосвязь надземной части растения и корней
Надземная часть растения и его корни очень зависят друг от друга. Любое ограничение одной из этих частей будет влиять на рост и развитие всего растения. Надземная часть обеспечивает корневую систему энергией, сконцентрированной в сахаре, произведенном в результате процесса фотосинтеза. В период засухи и высокотемпературного стресса, который требует значительного потребления воды и большего охлаждения за счет транспирации, растения способны выборочно направлять больше связанного углерода и энергии корням, чем надземной части, но здесь есть определенные ограничения.
Денитрификация
Плохая аэрация почвы может привести к денитрификации, то есть потере азота. Это может произойти вследствие активизации группы практически вездесущих бактерий почвы, которые при дыхании используют эфир азотной кислоты вместо кислорода. В этом случае эфир азотной кислоты переходит в газообразную закись азота, которая дифузирует (выходит) из почвы в атмосферу, и этот азот уже никогда не будет доступен растению. Такая почва становится средой обитания целого ряда строго аэробных и в большой степени не аэробных микроорганизмов. Необходимо помнить, что чем лучше дренаж и структура почвы, тем больше объем аэробной почвы. В почве, на поверхности которой были оставлены пожнивные остатки, в условиях теплой погоды при достаточном количестве осадков могут создаться анаэробные условия. Денитрификация здесь может наблюдаться временно, потому что проникновение кислорода в почву будет продолжаться вследствие потребления его микроорганизмами почвы.Исходя из изложенного, нужно не только уделять внимание внесению минерального азота, но и применять меры, сводящие к минимуму денитрификацию, – улучшение аэрации почвы, перевод азота в органическую форму, например с помощью сидерата, микроорганизмов и пожнивных остатков.
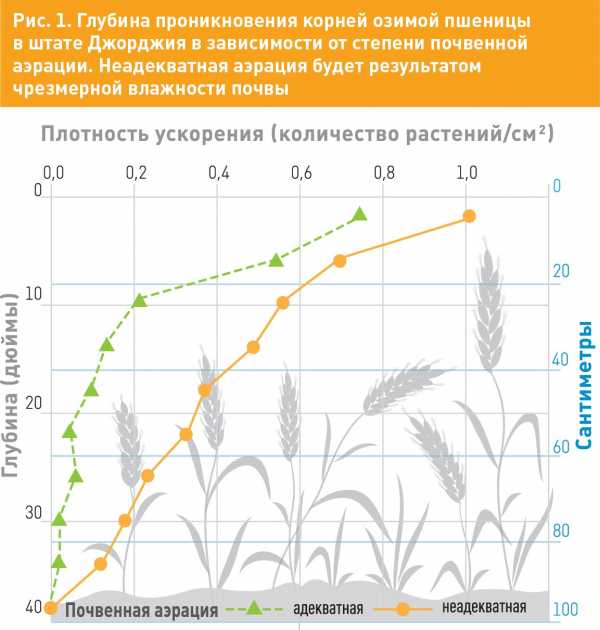
Питание растения и плодородность почвы
В предыдущей публикации мы говорили о взаимосвязи между обеспечением растения водой и его ростом. Чем больше снабжение водой, тем больше растение (при условии, что другие факторы не ограничивают его рост), но чем больше растение, тем больше воды ему необходимо для сохранения минимального тургора и непрерывного роста и развития.Если снабжение водой будет недостаточным для сохранения минимального тургора в клетках различных тканей, размер растения будет ограничен листьями, которые не разовьются до своей потенциальной ширины. Будет происходить сброс побегов (или они вовсе не будут формироваться), колосков или цветков (или отсутствие их формирования), не будет налива зерна. При лимите ресурсов растение генетически запрограммировано формировать по крайней мере один колос, одно или несколько семян в этом колосе. Вместе с тем, если нет ограничения в обеспечении растения водой, оно способно формировать много колосьев с максимальным количеством семян в каждом.По такому же принципу растение реагирует и на уровень обеспечения питательными веществами. Питательные вещества вместе с углекислым газом в атмосфере – строительный материал, из которого растение формирует себя как совокупность сложных органических веществ. Чем больше растение обеспечивается строительным материалом, тем больше оно развивается на любой стадии развития, тем ближе оно к своему максимальному (абсолютному) генетическому потенциалу. Следовательно, чем больше растение на любой стадии своего развития, тем в большей степени оно было обеспечено строительным материалом, необходимым для его роста и развития.При отсутствии заболеваний, засух или других неблагоприятных факторов пшеница в фазе выхода в трубку будет иметь половину (по массе) своего максимального зрелого размера. Далее, чтобы завершить вегетацию и сформировать минимальный от своего генетического потенциала урожай, пшеница нуждается в минимальном обеспечении как микро-, так и макроэлементами. Макроэлементы, необходимые для построения, сохранения и функционирования клеток и клетчатки, включают в себя: азот, фосфор, калий, кальций магний и серу. Микроэлементы очень важны для ключевых процессов, протекающих в клетке, но необходимы в незначительном количестве. К ним относятся цинк, медь, марганец, молибден, бор, железо и хлор.На фото 1-5 показаны примеры проявления дефицита питательных веществ на растениях пшеницы.




Азот
Растение пшеницы нуждается в азоте для выработки протеина. Так как протеин составляет внутреннее содержание клетки, он необходим в большем количестве, чем любой другой питательный элемент. Растение пшеницы без достаточного количества азота сначала становится светло-зеленым, а потом желтым (фото 1-2 и 5а). Этот симптом вначале проявляется на самых старых листьях, а затем и на самых молодых. Более старые листья в конечном счете становятся коричневыми и отмирают. Растение направляет ограниченное количество азота к самым молодым листьям.Пшеница может поглощать азот в виде аммония или нитрата. Аммоний не мобильный в почве. Имея положительный электрический заряд, этот ион удерживается в почве (в молекулярной решетке глины и органических веществ). Нитраты, будучи негативно заряженными, мобильны в почве и вымываются просачивающейся водой. Чтобы достичь аммония, корни должны широко разветвляться в почве. Диффузия аммония в корни, хотя и имеет место, но протекает очень медленно, чтобы удовлетворить потребность растения в азоте. Нитраты же, передвигающиеся в почвенном растворе, могут достигать даже базовых корней. Все полученные растением нитраты преобразовываются внутри растения сначала в аммоний, затем в аммонийную кислоту, а потом в протеин растения.Растение пшеницы реагирует на недостаток азота формированием маленьких листьев и меньшего количества побегов (или их отсутствием). Любой сбой в обеспечении растения азотом, совпадающий во времени с прорастанием спящей почки нового побега, выражается в том, что эта почка вовсе не прорастает.
Цель менеджмента выращивания качественной пшеницы – максимальное удержание азота в растении, сохранение этого элемента в пожнивных остатках для будущего урожая, а также минимизация потерь азота
Однако растение продолжает формировать новые побеги до тех пор, пока в него поступает азот, пока не начнут действовать какие-то другие ограничивающие факторы или не поступит сигнал (сумма активных температур или продолжительность дня) к удлинению стебля, тогда оно перейдет в фазу трубкования. После этого новые побеги перестанут формироваться. При достаточном количестве азота, а также фосфора и других питательных веществ озимая пшеница, высеянная в конце лета на выпас, может потенциально сформировать до 30 побегов на одном растении до перехода в фазу трубкования.Во время развития в распределении и перераспределении азота растение в первую очередь отдает предпочтение молодым побегам и их листьям. Так как почка побега формируется в период фазы третьего листа, то в это время особенно важно наличие азота для поддержания формирования побегов с самого начала (рис. 2).
Если для корректировки дефицита азота, выявленного во время формирования 4 и 5 листа на основном стебле, поверхностно внести удобрения после сформирования 6 листа, то это будет слишком поздно для формирования первых двух побегов. Третий побег может сформироваться, но первый и второй будут потеряны, а заодно и высокий урожайный потенциал. По сути, недостаток снабжения азотом после формирования первых двух или трех побегов ограничит размеры растения до одного главного стебля и трех побегов. Это неизбежно даже тогда, когда обеспечение водой и другие факторы позволяют иметь больше побегов и, следовательно, потенциально большее по размерурастение.После перехода в фазу удлинения стебля растение будет направлять азот на формирование не новых побегов, а основного стебля и новых листьев на существующих побегах (рис. 2).
Поверхностное внесение азотных удобрений после завершения фазы удлинения основного стебля может способствовать увеличению количества цветков в будущем колосе (азот должен попасть в растение) и, возможно, концентрации протеина в зерне. Но будет уже поздно влиять на количество колосьев и, вполне возможно, на количество колосков в них. Поверхностное внесение азотных удобрений после завершения фазы выхода в трубку уже не будет влиять на число колосков или цветков, но положительно скажется на содержании протеина в зерне (при условии, что азот будет абсорбирован растением, направлен в стебель, а потом в колос). Азот, внесенный на поверхность почвы после цветения, не повлияет на содержание протеина в зерне, тогда как листовые подкормки способны увеличить его содержание в зерне.
Сбой в обеспечении растения азотом, совпадающий во времени с прорастанием спящей почки нового побега, проявляется в непрорастании этой почки
При использовании азота очень важно четко представлять, какой вы хотите получить урожай и, соответственно, какие для этого необходимы параметры колосьев (их количество на единице площади, длина, количество колосков в них). Количество азота, которое нужно внести в той или иной фазе развития растения, можно рассчитать, определяя его содержание в растении в соответствующие фазы развития. При наличии избыточного количества питательных веществ во время формирования побегов и отсутствии других лимитирующих факторов растение поглощает азота все больше, в результате чего формирует больше побегов. В конце концов, оно потом может перенаправить азот от листьев к стеблю, а затем к колосу и зерну. В такой ситуации растение не будет консервировать азот в почве для дальнейшего его использования (если в ближайшее время оно не будет ограничено в его потреблении или не будет других лимитирующих факторов окружающей среды). Это как раз и объясняет тот факт, почему пшеница, будучи в подходящих для получения высокого урожая условиях окружающей среды, имеет низкое содержание протеина, а пшеница, выращиваемая в жестких условиях, сильно ограничивающих урожаи, имеет большее содержание протеина.
Общие требования
Здоровое культурное растение в Северной Америке обычно потребляет около 2,4-2,7 фунтов азота на 1 акр (2,7-2,9 кг/га) на каждый бушель пшеницы, полученный с акра (67 кг/га). Урожай 100 бушелей на акр (6,7 т/га) потребляет 240 фунтов азота на акр (270 кг/га). Это общее количество азота, содержащееся в корнях, листьях, стеблях и зерне пшеницы после созревания. Само зерно содержит около 0,8-1,0 фунтов азота в одном бушеле (1,3-1,7 кг азота в 100 кг) в мягкой (с низким содержанием протеина) пшенице и 1,5-1,6 фунтов в одном бушеле (2,5-2,7 кг на 100 кг) в твердой (с высоким уровнем содержания протеина) пшенице. Такое количество азота выносится с поля с зерном при сборе урожая. Остальной азот остается на поле в пожнивных остатках (солома, полова, корни) и становится доступным для следующей культуры после разложения остатков и минерализации азота.
Очень важно четко представлять, какой вы хотите получить урожай и, соответственно, какие для этого необходимы параметры колосьев (их количество на единице площади, длина, количество колосков в них)
Этот азот потом может теряться из-за выщелачивания, при котором он будет унесен с водой ниже корневой зоны. Из-за денитрификации, в процессе которой газообразные соединения азота улетучиваются в атмосферу, поверхностного стока и эрозии почвы, при которой с водой и почвой уносятся нитраты и аммоний. Удержание максимального количества азота в растении, сохранение почвенного азота в пожнивных остатках для следующей культуры сведет к минимуму его потери. Это должна быть одна из целей менеджмента выращивания высококачественной пшеницы, не пораженной вредителями и болезнями.
Фосфор
Фосфор входит в состав мембраны клетки и служит для передачи и сохранения энергии в растении. Как и азот, водород, кислород и углерод, он является частью структуры ключевых молекул в клетках, включая ДНК. Фосфор также имеет очень большое значение в некоторых вопросах содержания и интеграции существующих клеток растения. Без адекватного обеспечения этим элементом функции существующих клеток, а также генерация новых будут сильно ослаблены или прекращены (рис. 2).Растение пшеницы без адекватного количества фосфора, необходимого для нормального роста, задерживается в развитии и формировании колосьев, стебель будет тонким, корневая система плохой, листья – меньшего размера и по цвету темнее обычного, созревание растения будет затянуто (фото 3). Красноватые или пурпурные листья – один из симптомов дефицита фосфора в растении. Хорошее обеспечение растений фосфором будет стимулировать формирование мощной корневой системы и ускорит созревание пшеницы.
Пшеница, выращиваемая в монокультуре, из-за наличия в почве большой концентрации болезнетворных микроорганизмов, поражающих корни, может положительно реагировать на фосфорное удобрение даже тогда, когда анализ грунта указывает на среднюю или высокую концентрацию этого элемента
Доступность, полная потребность и отношение к здоровому состоянию корней
Фосфор в почве находится в виде фосфата, главным образом калия и железа, с относительно слабой растворимостью в воде, следовательно, он не очень мобильный, поэтому в почве корни должны дорасти до фосфора. Минерал легче поглощается корнями в почвах с рН около 6,0-7,5. Этот показатель будет иметь разное влияние на разных почвах и на разные растения. На кислотных почвах фосфор вступает в реакцию с железом и алюминием, образуя при этом менее растворимые соли, уменьшая доступность фосфора для растений пшеницы. В щелочных почвах он вступает в реакцию с кальцием, образовывая малорастворимые соли, которые также малодоступны растению пшеницы. Проблема закрепления фосфора в кислых и щелочных почвах может быть сведена к минимуму путем внесения фосфорных удобрений ленточным способом перед или во время сева. Ленточное внесение этого удобрения под семена во время сева предпочтительно внесению удобрений вместе с семенами, поскольку отсутствует непосредственный контакт нежных проростков с минеральными удобрениями (фосфорной кислотой). Внесение фосфора вместе с аммонийным азотом увеличивает его поступление в растения пшеницы. Подкормку пшеницы фосфором лучше всего проводить в соответствии с данными тестирования почвы. Фосфорные удобрения желательно применять, когда в почве мало питательных веществ. В зависимости от состояния корневой системы внесение фосфорных удобрений целесообразно осуществлять и при среднем содержании питательных веществ в почве, если же содержание фосфора оценивается как высокое, этого не нужно делать. Фактическая концентрация фосфора, которая считается низкой, средней или высокой, будет разной в разных почвах Северной Америки. Таким образом, нужно сверять свои данные с местными источниками, чтобы получить более точную рекомендацию.Урожайность пшеницы в зависимости от применения фосфора, особенно при ленточном его внесении, зависит от способности корневой системы достать (исследовать почву) и абсорбировать это питательное вещество. Растение пшеницы с развитой корневой системой и множеством корневых ворсинок идеально подходит для того, чтобы плотно заполнить почву в поисках фосфора. Однако если ворсинки были уничтожены болезнетворными микроорганизмами, растение может испытывать недостаток фосфора. Чем короче севооборот (чем чаще пшеница выращивается на поле), тем больше будет поражение корней болезнями и, соответственно, абсорбирующая способность корневой системы будет ниже.
Ленточное внесение фосфорных удобрений под семена во время сева предпочтительно внесению этих удобрений вместе с семенами, поскольку отсутствует непосредственный контакт нежных проростков пшеницы с минеральными удобрениями (фосфорной кислотой)
Пшеница, выращиваемая в трехпольном севообороте, будет расти с нормальными показателями даже при низком обеспечении ее фосфором. В то же время пшеница, выращиваемая в монокультуре, из-за наличия в почве большой концентрации болезнетворных микроорганизмов, поражающих корни, может положительно реагировать на фосфорное удобрение даже тогда, когда анализ грунта указывает на среднюю или высокую концентрацию этого элемента.Этим можно объяснить отсутствие реакции пшеницы на фосфор даже при его внесении ленточным способом непосредственно под семена на полях с постоянной безотвальной обработкой в случаях, когда тесты почвы указывают на то, что она нуждается в питательных веществах. Здоровое растение пшеницы в зрелом состоянии содержит 0,3-0,5 фунтов фосфора в каждом бушеле зерна (0,5-0,8 кг фосфора в 100 кг зерна). Таким образом, пшенице при урожае 100 бушелей на акр (6,7 т га) необходимо 33-50 фунтов фосфора (фосфата) на акр(35-55 кг/га), сосредоточенного вокруг корней, в листьях, стеблях (соломе) и зерне. Из этой суммы 20-25 фунтов (примерно фунта на один бушель) фосфора содержится в зерне, а остальное количество остается на поле с корневыми остатками и соломой.
Сера
Сера, как и азот, – составная часть протеина в растениях, поэтому является строительным материалом для формирования новых клеток и тканей растения. Симптомы дефицита серы в основном такие же, как и симптомы дефицита азота, но более ярко выраженные. Растение пшеницы при недостатке серы в фазе кущения становится низкорослым со слабой кустистостью, а также приобретает определенную окраску – от светло-зеленой до бриллиантовой или полностью желтой. Растение получает серу из почвы в виде сульфата, который содержит 35% серы. В зависимости от того, в какой форме сера находится в почве, она может быть достаточно мобильной (хотя и не такой мобильной, как азот), поэтому может поступать в корни вместе с почвенной водой, а также опускаться вниз по почвенному профилю. При интенсивной технологии выращивания пшеницы в монокультуре почва истощается. В этом случае необходимо вносить сульфатные удобрения.Будучи составной частью протеина, сера также является составной частью органических веществ, находящихся в почве. Она медленно высвобождается в процессе микробного разложения (минерализации органического вещества). Таким образом, почвы с малым содержанием органических веществ больше нуждаются в сульфатных удобрениях, чем высокогумусированные.Пшеница выносит 15-20 фунтов серы (40-60 фунтов сульфата) на каждые 100 бушелей произведенного зерна (2,5-5,5 кг/т). Около 2/3 ее количества содержится в зерне.
Калий
Калий необходим растению для сохранения баланса положительного и отрицательного заряда наружной и внутренней стороны клетки. Кроме того, он задействован в процессах транспортировки продуктов фотосинтеза и активизации определенных ферментных систем. Калий также выполняет определенные функции в регулировании напряжения тургора клетки. Ионы калия поглощаются или выталкиваются из клетки для сохранения электрохимического баланса. Ионы калия, будучи положительно заряженными, удерживаются в почвенном поглощающем комплексе, они относительно немобильны, особенно в некоторых глинистых почвах, если не заменяются другими положительно заряженными ионами. Вообще корни растения пшеницы должны дорастать до ионов калия. Калий довольно медленно проникает в корни.Взрослые растения пшеницы при дефиците калия уменьшаются в размерах и имеют слабый основной стебель (из-за коротких и тонких междоузлий). Листья становятся желтыми и со временем усыхают, начиная с верхней части стебля, затем начинают желтеть по краям и более старые листья (фото 5с). Корни дополнительных побегов либо вообще не развиваются, либо появляются, но не распространяются в почве с такой скоростью, как это происходит при отсутствии дефицита калия. Многие из этих симптомов типичны для растения, подверженного стрессу во время засухи.Большинство почв Великой Равнины и Тихоокеанского северо-востока располагают достаточным количеством калия для здорового роста пшеницы. Дефицит калия обычно проявляется на песчаных и в большой степени на выщелоченных почвах, где годовой уровень осадков превышает их испарение в атмосферу.Здоровые растения пшеницы выносят до 150-200 футов калия с каждыми 100 бушелями зерна (25-35 кг/т). В соломе содержится в три раза больше калия на акр, чем в зерне. Около 25 фунтов калия выносится с урожаем на каждые 100 бушелей зерна (4-4,5 кг/т зерна) и 80-100 фунтов возвращается в почву с соломой на каждые полученные 100 бушелей зерна (13-16 кг/т зерна).Поскольку калий, как и фосфор, в почве относительно немобильный, степень реакции пшеницы на применение калийных удобрений будет зависеть от абсорбирующей способности корневой системы. Заболевание корней, низкая температура почвы и другие факторы, которые могут повлиять на рост корней и их абсорбирующую способность, могут выразиться в дефиците калия, даже если данные тестирования почвы указывают на адекватное наличие питательных веществ в ней. Тестирование почвы является первым обязательным шагом в программе менеджмента применения удобрений. Но производители зерна должны иметь в виду, что севооборот, срок сева и другие моменты технологии выращивания пшеницы могут существенно влиять на необходимость применения калийных удобрений и их эффективность. Как и фосфорные, калийные удобрения вносятся в большинстве случаев ленточным способом. Отдача от них урожаем будет оптимальной только в том случае, когда калий для роста растения является лимитирующим фактором.
Кальций
Кальций в растении пшеницы необходим для мембраны клетки и ускорения развития корневых волосков (удлинения клеток на кончиках корневых волосков). Он также является скрепляющим материалом между клетками. Существует мнение, что кальций усиливает транспортировку и удержание некоторых питательных веществ (таких, как калий) в ткани корней. Так как кальций в растении немобильный, молодая ткань в первую очередь ощущает его дефицит. Характерными симптомами дефицита калия являются уменьшение размера корневой системы, а также снижение числа побегов.Вообще о симптомах дефицита кальция говорят очень редко. Одной из причин этого, как правило, является то, что его дефицит чаще всего наблюдается на кислых почвах. Он обычно маскируется негативным действием других факторов, таких как излишек алюминия и марганца. Кальций будет лимитирующим фактором для пшеницы на почвах с рН ниже 5,0. Пшеница поглощает около 20 фунтов кальция на каждые 100 бушелей (3-4 кг/т) полученного зерна.Когда применяется известь для раскисления почвы, то внесенный кальций будет, скорее всего, ограничивающим фактором, а не питательным элементом. Одним из положительных моментов применения извести является то, что она уменьшает концентрацию алюминия и марганца и в результате увеличивает доступность некоторых питательных элементов растению, которые были в ограниченном количестве в кислой почве. Кроме того, известь улучшает структуру почвы.
Магний
Магний играет структурную роль в хлорофилле, что является существенным фактором фотосинтеза в растении. Как и фосфор, магний необходим для нормального функционирования клеток растения. У нас нет сведений о почвах Северной Америки, где магний в дефиците для пшеницы и есть необходимость в применении специальных удобрений, содержащих магний. Пшеница содержит меньше 1,5-2 кг магния на 1 т зерна и выносит в целом меньше 20 фунтов во всей надземной массе растений на каждые 100 бушелей урожая (меньше 3,0-3,5 кг/т).Хотя были случаи заболевания крупного рогатого скота, который выпасали на юге Большой Равнины в местах, где выращивали озимую пшеницу. Причина заболевания заключалась в низком содержании магния в листьях пшеницы в этой местности. Концентрация магния в пшенице может быть повышена за счет внесения ленточным способом при посеве таких удобрений, как сульфат магния вместе с семенами пшеницы или по всходам пшеницы. Однако исследования не дали никаких убедительных доказательств того, что проблема заболевания животных может быть решена таким образом.
Микроэлементы
Железо, цинк, медь, марганец, бор, молибден и хлор участвуют во многих биохимических реакциях, протекающих в клетках растения. Одни из них только содействуют процессу образования протеина, а другие участвуют в построении больших молекул, и поэтому имеют критическое значение для функционирования клеток. Азот, фосфор и калий имеют огромное значение для основных функций клеток, а также для их восстановления, но общая потребность в них небольшая.В основном симптомы дефицита железа, марганца или меди проявляются в первую очередь на новых органах растения. Однако дефицит цинка проявляется сначала на старых листьях. Можно предположить, что растение само перенаправляет этот питательный элемент к новым листьям за счет старых. Растение, испытывающее дефицит одного из микроэлементов, отстает в росте, поэтому, как правило, имеет маленькие размеры и малое количество побегов. Самые очевидные симптомы проявляются на листьях: они вначале желтеют, обычно между жилками (междужильный хлороз), а потом полностью теряют свой цвет. Потребность пшеницы в микроэлементах мизерная – только одна сотая или одна тысячная ее потребностей в макроэлементах – кальции, магнии, сере или др.Микроэлементы широко не применяются в Северной Америке как обязательная часть программы использования удобрений. Однако их дефицит иногда становится ограничивающим фактором получения достижимого урожая.Дефицит в цинке, марганце, меди и железе больше будет создавать проблем на почвах с высоким рН. Известно, что с увеличением рН почв эти микроэлементы становятся менее доступными.
Р. Джеймс Кук, Министерство сельского хозяйства США, исследовательская сельскохозяйственная служба, Госуниверситет Вашингтона
Роджер Дж. Фесет,Служба по корпоративному распространению
www.zerno-ua.com
Лист озимой пшеницы
Листья растений озимой пшеницы появляются из верхнего слоя меристемы конуса нарастания. При делении клеток меристемы образуются выступы — листовые бугорки.
По мере разрастания верхнего бугорка возникает первый лист изумрудно-зеленого цвета, без опушения. Одновременно с ростом идет и дифференциация тканей листа. Листья, образовавшиеся осенью, сохраняются в зеленом виде до весны следующего года.
Листья озимой пшеницы бывают двух типов: прикорневые и стеблевые. Прикорневые образуются из подземных узлов, стеблевые — из надземной части стебля. Размер первого листа зависит от крупности зерна, величины зародыша и условий произрастания
Лист состоит из влагалища и листовой пластинки. Влагалище прикреплено к стеблю и своей нижней частью охватывает его в виде трубки. В нижней части листового влагалища имеется небольшое кольцеобразное утолщение, называемое листовым узлом, разрастающееся непосредственно над стеблевым узлом. Листовой узел по мере разрастания своей нижней частью давит на стебель, способствуя поднятию его после полегания.
Там, где влагалище переходит в листовую пластинку, у пшеницы имеется тонкая полупрозрачная пленка — продолжение внутренней части листового влагалища, которая называется лигулой, или язычком. Язычок довольно плотно прилегает к стеблю и предохраняет трубку влагалища от проникновения в нее воды и паразитов. По обоим бокам язычка образуются ушки, или рожки.
Листья выполняют очень важную функцию. В них протекает процесс фотосинтеза, то есть усвоение из внешней среды углекислоты за счет энергии солнечного света и преобразование ее в химическую энергию органических веществ. В процессе фотосинтеза создаются различные углеводистые соединения, превращающиеся затем в крахмал и сахар. При этом углерод и вода, содержащиеся в растении наряду с азотом и другими элементами, участвуют в образовании аминокислот и белков. Таким образом, сухая масса урожая зеленых растений состоит в основном из органических веществ, первично создаваемых в процессе фотосинтеза.
В последние годы профессор Горьковского университета И. И. Волский (1968) открыл, что растения, как и другие живые организмы (животные, микроорганизмы), ассимилируют свободный азот. Происходит это с помощью органических соединений, содержащихся в хлорофилле растений, при воздействии атмосферных ионов (заряженных частиц), под влиянием микроэлементов (меди, железа, молибдена, лития), а также с участием бактерий в процессе фотосинтеза. Новое открытие ассимиляции газообразного азота атмосферы повышает роль листьев растений в создании органических веществ (белков, углеводов и др.). Работы в этом направлении продолжаются.
Размеры и число листьев у озимой пшеницы довольно сильно колеблются, что связано с биологическими особенностями сорта. Лучшие сорта дают высокий урожай при листовой поверхности 30—50 тыс. м2 на 1 га (Корнилов, 1968). Площадь листьев значительно повышается на фоне удобрений. В опытах Ставропольского сельскохозяйственного института (Н. И. Перегудов), в 1966 г. листовая поверхность у пшеницы Безостая 1, посеянной после неудобренной кукурузы на силос, была равна 39,8 тыс. м2 на I га, а урожайность составила 47,1 ц с 1 га. На удобренном фоне (площадь листьев увеличилась до 46,8 тыс. м2, а урожайность — до 52 ц с 1 га.
По нашим наблюдениям, листьев на растениях удобренных фонов было значительно больше, чем на контрольных делянках. На всех вариантах опыта при раннем сроке посева (15 августа) число листьев увеличивалось.
Опыты ряда исследователей (Мединец, 1963) показывают, что высокий выход зерна у сортов озимой пшеницы обусловлен особенностями строения растений, в частности высотой растений и степенью их облиственности. Чем короче стебель и больше облиственность растений, тем больше выход зерна. Так, на Лохвицком и Козелыципском сортоучастках (Полтавская область) у сорта Безостая 1 растения были короче, облиственность их лучше, а выход зерна больше, чем у других сортов.
На увеличение листовой поверхности растений пшеницы влияют способы посева. По данным А. Г. Михайловского и В. П. Гудзь, в среднем за 10 лет получены следующие результаты. При внесении в занятом пару (горох и вико-овсяная смесь на сено) площадь листовом поверхности у сорта Мироновская 808 па 1 га в период выхода в трубку составляла при обычном рядовом способе посева 46,7 тыс. м2, прерывисто-рядовом (отрезками по 35 см обычного посева в рядке через каждые 15 см)—56,0 и пунктирном (размещение растений в рядке через каждые 4,5—5,0 см) —61,3 тыс. м2. При внесении навоза 20 т на 1 га и площадь листовой поверхности на 1 га была равна: при обычном рядовом посеве 51,3 тыс. м2, прерывисто-рядовом— 57,2 и пунктирном — 59,3 тыс. м2. Урожайность но минеральному фону при обычном рядовом севе составила 35,3 ц с 1 га, прерывисто-рядовом — 38,5 и пунктирном — 45,0; по органо-минеральному фону соответственно 35,3; 39,0 и 46,4 ц с 1 га.
Исследования показали, что наибольшее количество ФАР поглощаются растениями пшеницы при пунктирном посеве. Так, обычный посев на органическом фоне в период цветения поглощал солнечной радиации (ФАР) 38,8 ккал/га/мин, а некоторые растения — до 184,7 кал/мин, пунктирный же посев — 39,9 ккал/га/мин, а некоторые растения — 306,9 кал/мин.
Площадь листовой поверхности озимой пшеницы зависит также от предшественников и способов обработки почвы. Так, по данным Петровской селекционно-опытной станции (Пензенская область; Огарев, Разуваев, 1973), площадь листовой поверхности пшеницы на 1 га после вико-овсяной смеси на контроле в среднем за три года (1970—1972) составила (в тыс. м2): при лущении — 17,5, при вспашке— 13,5, после гороха на зерно соответственно 17,3 и 12,6, после кукурузы па зеленый корм — 15,4 и 14,3.
При внесении полного минерального удобрения площадь листовой поверхности была больше, чем па контроле, по лущению па 22%, а по вспашке — на 30%.
На увеличение площади листовой поверхности оказывают влияние нормы посева семян. И опытах А. М. Белоусова и Г. П. Устенко (1972) при увеличении нормы посева сорта Кавказ с 2,09 до 5,96 млн. всхожих зерен на 1 га максимальная площадь листьев возросла с 39 до 65 тыс. м2 на 1 га, а общая фотосинтезирукщая поверхность посева—с 58 до 84 тыс. м2 на 1 га. При увеличении нормы посева сорта Мироновская юбилейная с 1,23 до 5,05 млн. всхожих зерен на 1 га площадь листовой поверхности возросла с 35 до 62 тыс. м2 на 1 га, а общая фотосинтезирующая поверхность — с 16 до 81 тыс. м2 на 1 га.
Если вы нашли ошибку, пожалуйста, выделите фрагмент текста и нажмите Ctrl+Enter.
www.activestudy.info