
ГЕЛОФИТЫ (ГИДРОГИГРОФИТЫ) – ВОДНО-БОЛОТНЫЕ. Гелофиты примеры растений
Гелофиты
Гелофиты — разновидность гидрофитов — растения, обитающие на болотах и заболоченных лугах.[ ...]
Далее следует зона высокотравных гелофитов, представленная сообществами тростника обыкновенного и, реже - камыша озерного. На участках, особо подверженных действию волн, ветра, особенно на каменистых грунтах, разреженная растительность представлена различными видами прикрепленных погруженных гидрофитов, к которым могут примешиваться низкотравные гелофиты - стрелолист обыкновенный, сусак зонтичный.[ ...]
Зона 1 0—0.5 Торфяно- илистый Заросли гигрофитов, гелофитов, переходящие в юго-западном углу в злаково-осоковую сплавину с тростником, тростянкой овсяннице-видной, вехом, двукисточником тростниковидным, белокрыльником болотным, сабельником болотным.[ ...]
В растительности устьевых участков рек преобладают высокотравные гелофиты: Phragmites anstrulis, Scirpus lacustris, реже Typha angustifilia. Плавающая растительность представлена ценозами Nuphar lutea, Nymphaea Candida, Polygonum amphibia и Potamogeton natans. В растительном покрове юго-западного побережья Рыбинского водохранилища сочетаются черты растительности водоемов с постоянным и переменным уровнями. В водоемах с постоянным уровнем доминируют заросли тростника, камыша, манника, хвоща, с переменным уровнем — амфибийные группировки и прежде всего сообщества с господством жерушника земноводного и горца земноводного. Формирование растительных экотопов и характер сукцессий фитоценозов полностью зависит от изменений уровенного режима водохранилища.[ ...]
Зона 1 0-1 Илистый, илистопесчаный Разнообразные сообщества гигрофитов и гелофитов, чаще всего тростник, осоки (у береговой кромки). Глубже произрастает камыш и другие гелофиты с участием гидрофитов - рдестов стеблеобъемлющего и блестящего, урути, мелкой хары.[ ...]
| Скорость течения (а), общая степень зарастания (б), степень зарастания гидрофитами (в), степень зарастания гелофитами (г) по продольному профилю малых рек |  |
Основными растительными формациями в Ильменских озерах являются: Phragmiteta australis, Scirpeta lacustris, Typheta latifolii (группа формаций высокотравных гелофитов), Nuphareta lutei, Nymphaeeta candidae, Polygoneta amphibii (группа формаций прикрепленных гидрофитов с плавающими листьями), Stratioteta aloiditis, Myriophylleta spicati, Potamogetoneta lucentis, Potamogetoneta perfoliatis, Elodeeta canadensis, Ceratophylleta demersii (группа формаций прикрепленных погруженных гидрофитов), а также формации осок в прибрежной зоне.[ ...]
Горизонтальная и вертикальная структура сообществ макрофитов в озерах тесно связана с их экобиоморфологической структурой. Начиная с зоны гелофитов с увеличением глубины происходит смена жизненных форм доминантов в сторону увеличения их связи с водной средой и упрощение экобиоморфологической структуры за счет уменьшения доли гелофитов и гидрофитов с плавающими листьями. Соответственно, происходит упрощение ярусной структуры за счет выпадения надводного яруса гелофитов и наводного яруса прикрепленных гидрофитов с плавающими листьями.[ ...]
Показательно то, что вне зависимости от жизненной формы доминанта, во всех формациях по числу видов преобладают погруженные прикрепленные гидрофиты. Если гелофиты и прикрепленные гидрофиты с плавающими листьями в основном произрастают в строго ограниченных экологических условиях, где некоторые из них доминируют, представители других экобио-морфологических групп могут встречаться в различных местах обитания. Особенно это касается прикрепленных погруженных гидрофитов, которые, однако, могут доминировать также только в определенных биотопах.[ ...]
В оз. Б. Миассово, как по видовому богатству, так и по разнообразию жизненных форм выделяется формация тростника обыкновенного. В ее состав входят 25 видов 6-ти жизненных форм. Несмотря на то, что доминирующий вид является высокотравным гелофитом, по количеству видов здесь преобладают прибрежные гигрофиты (их присутствие характерно только для этой формации и только для этого озера) и прикрепленные погруженные гидрофиты. Высокотравные гелофиты литтть на третьем месте. Формация камыша озерного включает меньшее число видов — 13, принадлежащих к 5 эко биоморфо логическим группам, большая их часть — прикрепленные погруженные гидрофиты. В отличие от тростниковых сообществ отсутствуют прибрежные гигрофиты и плавающие на поверхности воды гидрофиты, и появляются приземные гелофиты.[ ...]
Зона 2 0.5—1 Мощный слой ила Преобладают погруженные гидрофиты. Распределение неравномерное. Доминируют телорез, элодея, рдест блестящий. Встречаются рдесты стеблеобъемлющий, сплюснутый, кубышка желтая. Изредка попадаются низкотравные гелофиты — стрелолист, ежеголовник простой.[ ...]
Зона 3 0.5-1 Илистый Разнообразные группировки гидрофитов погруженных и с плавающими листьями. Наиболее обильны гречиха земноводная, кувшинка чисто-белая, рдесты блестящий и стеблеобъемлющий, уруть колосовидная, мелкая хара. Из низкотравных гелофитов высокого обилия достигает хвощ топяной. Распределение неравномерное.[ ...]
При рассмотрении динамики флоры по структуре жизненных форм (по Раункиеру) можно сказать, что за 36 лет произошло увеличение групп фанерофитов (с 2 до 15 видов) и гемикриптофитов (с 25 до 54) и, в меньшей степени, увеличение групп геофитов (с 13 до 16) и терофитов (с 17 до 23), появилась группа хамефитов (6 видов). В группе гелофитов на первых этапах формирования происходило увеличение числа видов (от 1 до 5 за 1967-1972 гг.), а в последующие периоды - уменьшение до 1 вида, что связано с изменением эдафических условий.[ ...]
Наряду с анализом общего флористического списка, нами определялся состав жизненных форм различных растительных сообществ (рис. 54). Не учитывались прибрежные фитоценозы, так как состав жизненных форм здесь ограничивается преимущественно гигрофитами, кроме которых немалую роль играют три вида высокотравных гелофитов - тростник обыкновенный, рогозы широколистный и узколистный. Исходя из геоботанических описаний, определялось число видов макрофитов, принадлежащих к разным экобиоморфологическим группам, входящим в состав тех или иных растительных формаций.[ ...]
Большинство из растительных формаций в оз. Б. Ишкуль слагается меньшим числом видов и жизненных форм. Исключение составляют формации урути колосовидной, рдестов блестящего и стеблеобъемлющего, которые богаче и по видовому и по экобиоморфологическому составу, чем в оз. Б. Миассово. Отсутствуют представители приземных гелофитов и гидрофитов (это не относится, естественно, к фитоценозам прибрежной полосы и прибрежным сплавинам).[ ...]
В Шарканском районе (центральная часть УР) этот гибридогенный таксон в обилии произрастал по мелководьям Росовского пруда на окраине с. Шаркан вблизи ценозов Т. angustifolia. Характерными местообитаниями для Т. smimovíi были также придорожные лужи вблизи того же населенного пункта, где он произрастал совместно с другими видами гелофитов. Таким образом, изучаемые нами гибридогенные виды занимали свободные экологические ниши, освободившиеся в результате различного рода нарушений первичных биоценозов, и образовывали при этом обширные монодоминантные заросли.[ ...]
Анализ полученных данных показал, что в целом во флоре озерных макрофитов разнообразнее всех групп представлены гигрофиты — 32 вида, или 39.5 % от общего числа. Среди собственно водных растений выделяется группа погруженных гидрофитов, прикрепляющихся к субстрату (20 видов, или 25 %). Богат видовой состав низкотравных гелофитов (10 видов, или 12.3 %), несколько беднее — группы высокотравных гидрофитов и прикрепленных гидрофитов с плавающими листьями (6 видов, или 7.4 %). Остальные группы включают по 2—3 вида, что составляет 2.5—3.7 %.[ ...]
Выше (раздел 6.2.1) было показано, что количественное обилие зоопланктона зарослей макрофитов малых рек не имеет отличий на быстро- и медленнотекущих участках и не зависит от длины водотоков. При этом некоторые элементы видового состава зоопланктона определяются скоростью течения, общей степенью зарастания, а также зарастанием гидрофитами или гелофитами.[ ...]
Зона 1 0—1 Илисто- песчаный У самой кромки воды наряду с лесными растениями очень узкой полосой произрастают некоторые виды гигрофитов (осоки, дербенник иволистный, вербейник обыкновенный, шлемник обыкновенный), а также тростник. В воде — сообщества урути колосистой, рдеста стеблеобъемлющего с участием лютика водяного, рдеста курчавого, элодеи, иногда — рдеста блестящего. Встречаются также гелофиты — стрелолист обыкновенный, сусак зонтичный.[ ...]
Таким образом, в эвтрофных озерах макрофитная растительность занимает гораздо больший процент площади, чем в мезотрофных. В оз. Б. Таткуль практически все дно покрыто макрофитами, а в оз. Аргаяш от них свободен лишь небольшой участок в центральной, самой глубокой части. В эвтрофных озерах прибрежные сплавины занимают относительно большую часть площади, чем в мезотрофных. Соответственно, доля сообществ гидрофитов меньше. Сообщества гелофитов, характерные для песчаных и каменистых мелководий мезотрофных озер, в эвтрофных озерах сосредоточены, главным образом, на сплавинах, в воде же встречаются редко (в основном это касается тростниковой и рогозовой формаций).[ ...]
Наземные представители полушника обитают на очень влажных почвах с постоянным подтоком грунтовых вод и в условиях очень влажного воздуха. Например, полушник Дюрьё (I. durieui) в Испании произрастает на травянистых тенистых склонах, заболоченных почти в течение всего года, а в районах сильных туманов он может встречаться даже на замшелых скалах, прикрытых тонким слоем почвы. По сути дела, он является не чисто наземным, а болотным растением — гелофитом.[ ...]
В основу классификации экологических групп флоры города был положен критерий отношения растений к обеспечению водой. Экологический анализ показал преобладание мезофитов - растений, приуроченных к местообитаниям со средней степенью увлажненности (279 видов; 54,0 % СФ). Многочисленны виды сухих местообитаний - ксеромезофиты и ксерофиты (соответственно 120 и 10 видов, что в сумме составляет 25,1 % СФ). На долю видов переувлажненных местообитаний (гидрофиты, гелофиты, гигрогелофиты, гигрофиты, гигромезофиты) приходится 20,9 % СФ (108 видов). Соотношение экологических групп также свидетельствует о сходстве флоры города с флорами более южных территорий вследствие ее антропогенной трансформации.[ ...]
Что касается собственно водных макрофитов, то процент погруженных гидрофитов, прикрепленных к субстрату, самым высоким оказался в оз. Б. Миассово, несколько ниже — в оз. Б. Ишкуль и Аргаяш, и совсем небольшой — в оз. Процент гидрофитов, плавающих на поверхности воды, почти в два раза выше в эвтрофных озерах. Укореняющиеся гидрофиты с плавающими листьями составляют большую часть макрофитов в оз. Б. Ишкуль и Аргаяш чем в оз. Б. Миассово и Б. Таткуль. Высокотравных гелофитов несколько больше в эвтрофных озерах. Видовой состав низкотравных гелофитов в Б. Таткуле намного беднее, чем в остальных озерах.[ ...]
Увеличение степени зарастания на медленнотекущих участках влияло на численность и биомассу веслоногих ракообразных (г = 0.44 и г = 0.47, р [ ...]
Увеличение скорости течения способствует повышению обилия коловраток и снижению выравненное™ зоопланктона. Однако зоопланктон зарослей макрофитов быстро- и медленнотекущих участков не различается, хотя на быстротекущих участках глубже проявляется зависимость развития от различных гидрологических и морфометрических характеристик водотоков. В частности, при увеличении длины водотоков снижается число видов коловраток, но увеличивается общее число видов за счет повышения разнообразия кладоцер. При усилении скорости течения сокращается число видов, но возрастает обилие коловраток. Увеличение степени зарастания быстротекущих участков способствует повышению доли коловраток в биомассе зоопланктона. На медленнотекущих участках при увеличении степени зарастания гидрофитами наблюдается рост обилия веслоногих, а при увеличении зарастания гелофитами также возрастает доля веслоногих и выравненность зоопланктона по биомассе, но снижается численность ветвистоусых ракообразных.[ ...]
Степень развития растительности, видовой и экобиомор-фологический состав макрофитов и их распределение в водоеме подчиняются определенным закономерностям и зависят от многих экологических факторов (морфологические характеристики водоема, оптические свойства воды, подвижность водных масс, действие ветра, химические факторы, газовый режим, механические и химические свойства грунтов, температурный режим и другие). Особо нужно отметить антропогенный фактор, оказывающий существенное влияние на макрофитную растительность. Все эти факторы тесно связаны друг с другом и в комплексе создают все разнообразие мест обитания, которое отражается в распределении макрофитов в водоеме. При благоприятных для жиз ни растений условиях у берегов различают несколько зон, или поясов растительности, сменяющих друг друга с увеличением глубины (Катанская, 1981). Каждая из зон образуется видами, относящимися к определенным экобиоморфам. Как правило, они располагаются в следующем порядке: 1) зона прибрежных влаголюбивых растений; 2) зона воздушно-водных растений, или гелофитов; 3) зона гидрофитов с плавающими листьями; 4) зона высоких погруженных гидрофитов; 5) зона придонных гидрофитов.[ ...]
ru-ecology.info
Светолюбивые - гелиофиты.
ВЛИЯНИЕ УСЛОВИЙ ВНЕШНЕЙ СРЕДЫ НА МОРФОЛОГИЧЕСКОЕ И АНАТОМИЧЕСКОЕ СТРОЕНИЕ ОРГАНОВ РАСТЕНИЙ. ЭКОЛОГИЧЕСКИЕ ФАКТОРЫ И ЭКОЛОГИЧЕСКИЕ ГРУППЫ РАСТЕНИЙ.
Экологические группы выделяются по отношению к одному фактору среды, который играет большое значение в формировании и изменении органов высшего растения.
Наиболее важными экологическими факторами, влияющими на структуру растения являются:
- влажность воздуха, почвы;
- свет;
- температурные условия;
- особенности почв;
- конкуренция сообществ и т.д.
В связи с экологическими факторами выделяют соответствующие экологические группы. Экологическая группа растений - это совокупность видов или их популяций, произрастающих в определенных близких экологических условиях.
К одним и тем же условиям среды растения приспосабливаются по-разному, поэтому в пределах одной экологической группы можно найти растения, резко отличающиеся по внешнему облику друг от друга, и ещё по анатомическому строению, то есть растения имеют различнуюжизненную форму (биоморфу) – своеобразный внешний облик, возникающий в результате индивидуального развития (онтогенеза) в определённых экологических условиях и отражающий совокупность основных приспособительных черт. Жизненная форма растений зависит от особенностей роста подземных и надземных вегетативных органов, это результат длительной эволюции, она закреплена наследственно у особей одного и того же вида, но в зависимости от конкретных условий среды может варьировать. Экологические группы и формы растений между собой тесно связаны.
I. По отношению к влаге:
Гидрофиты.
Растения, нормально растущие в воде, встречающиеся часто по берегам водоёмов. В связи со спецификой местообитания у них наблюдаются следующие особенности – губчатость, рыхлость тканей, наличие крупных межклетников, то есть из тканей преобладает аэренхима. Система воздухоносных межклетников очень хорошо развита и является типичной чертой их строения. Полости частей растения, погружённых в воду, обычно сообщаются с устьицами непогружённых листьев, что является важным для газообмена. Воздушные полости есть во всех органах, даже в плодах (Nuphar lutea), что обеспечивает плавучесть и распространение семян. Губчатость тканей и наличие воздушных полостей в листовых пластинках и черешках также поддерживает плавучесть растения. У полностью погружённых растений (Elodea sp.) воздушные полости могут аккумулировать кислород в течение дня, который затем используется растением на дыхание ночью. У подводных растений в органах нет кутикулы и перидермы, устьица не функционируют. Отсутствие кутина и суберина способствует поглощению воды и питательных веществ непосредственно поверхностью тела, что является важным для питания растения. Корни обычно короткие и не разветвлённые, их окончания лишены корневых волосков. Отсутствуют механические и водопроводящие ткани. Недостаток арматурных тканей компенсируется плавучестью отдельных органов.
В зависимости от того, со сколькими средами контактирует растение, выделяются следующие подгруппы гидрофитов:
а) плавающие на поверхности, но не связанные с почвой:
Lemna minor, Salvinia sp.
б) полностью погруженные в воде, взвешенные в её толще,заселяют хорошо освещённые и аэрируемые слом воды, не связаны с почвой:
Batrachium eradicatum, Lemna trisulca, Sargassum sp., Utricularia intermedia.
в) полностью погруженные, плавающие, укореняющиеся:
Zostera marina, Isöetes lacustris, Potamogeton natans.
г) плавающие на поверхности, но укореняющиеся:
Batrachium peltatum, Nuphar lutea, Nymphaea candida, Sparganium sp.
Гелофиты.
Травянистые растения, вегетативные побеги которых располагаются в воздушной среде, а почки возобновления в неблагоприятный для вегетации период находятся под водой или в почве. Экологически разнообразная группа, занимающая переходное положение между световыми гигрофитами и гидрофитами. Нередко к ним относят только болотные или прибрежно-водные растения (амфифиты) – укореняющиеся растения мелководных местообитаний, стебли и листья которых высоко возвышаются, есть мощное корневище с толстыми придаточными корнями. Хорошо размножаются вегетативно.
Alisma plantago-aquatica, Calla palustris, Carex sp., Menyanthes trifoilata, Sagittaria sagittifolia, Scirpus lacustris, Typha latifolia.

Рис. 1. Поперечный разрез стебля белокрыльника болотного Calla palustris или частухи подорожниковой Alisma plantago-aquatica.
Гигрофиты.
Сухопутные растения, онтогенез которых проходит при хороших условиях – достаточной влажности воздуха, поэтому у них нет особых анатомо-морфологических приспособлений к засухе, слабо приспособлены к изменению влажного режима. Подземные органы, а часто и основания побегов таких растений находятся в воде. Корни и листья этих растений снабжены аэренхимой, в листьях слабо развита столбчатая паренхима и склеренхима, кутикула не выражена, клетки эпидермы имеют тонкие стенки. Это влаголюбивые растения - обитатели влажной и тёплой атмосферы, обитающие по берегам водоемов (Carex aquatilis), на болотах (Menyanthes trifoliata), на низинных лугах (Bistorta major), в травянистых лесах, тенистых лесах умеренной зоны, где воздух должен быть насыщен парами воды.
Гигрофиты умеренной зоны по отношению к свету делятся на следующиеподгруппы:
а) теневые гигрофиты тенистых лесов, имеют тонкие листья, нуждаются в высокой влажности приземного слоя воздуха:
Adoxa moschatellina;
б) световые (солнечные) гигрофиты, растущие на открытых местах, но на постоянно сырых почвах:
Caltha palustris, Cardamine sp., Carex sp.
Ксерофиты.
Растения этой группы имеют разнообразные приспособления к поддержанию водного баланса при недостатке почвенной и атмосферной влаги. Это самая разнообразная морфологически группа растений. Способность переносить неблагоприятные условия связана с морфологическими признаками, по которым среди них выделяют:
а) суккуленты – сочные, мясистые растения с сильно развитой водозапасающей паренхимой в листьях (Alöe, Agava), в стебле – кактусы, молочаи, в корне (Ceiba). Суккуленты – основные ландшафтные растения пустынь Центральной Америки (кактусы) и Африки (молочаи). Суккулентность растения связана с разрастанием клеток паренхимы при одновременном увеличении вакуолей и сильном сокращении межклетников. Основная отличительная черта суккулентов – способность запасать большое количество воды, сохранять и экономно её расходовать. Суккуленты имеют низкую интенсивность транспирации. Днём ни закрывают устьица. Потери воды могут быть уменьшены за счёт формы растения. Форма и замыкание устьиц днём затрудняет газообмен, поэтому у суккулентов особый цикл фотосинтеза. Многие кактусы фотосинтезируют в течение почти всего года, но образуют небольшое количество углеводов, что замедляет их рост. Корневая система у них небольшая, они используют воду из поверхностных слоёв почвы.
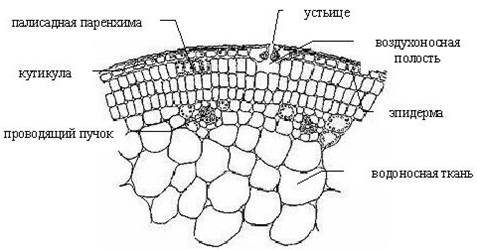
Рис. 2. Поперечный разрез листа алоэ Alöe sp.
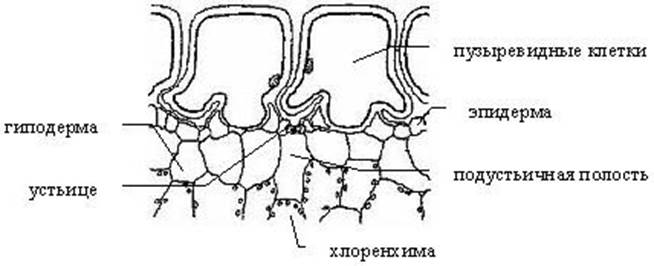
Рис. 3. Поперечный разрез толстянки Crassula falcata с пузыревидными клетками эпидермы.
В нашей флоре мало суккулентов: Mertensia maritima, Rhodiola arctica, Rh. rosea, Sedum acre.
б) склерофиты – по внешнему облику это суховатые и жёсткие растения с мало обводнёнными тканями. Они обладают высокой устойчивостью к завяданию, могут без заметного вреда терять до 25% воды. Имеют высокое осмотическое давление клеточного сока, что позволяет им увеличивать сосущую силу корней и получать воду из сухих почв. Корневая система развита сильнее надземной. Ксероморфные признаки проявляются, прежде всего, в особенностях строения эпидермы, клетки которой обладают утолщёнными наружными стенками. Мощная кутикула покрывает эпидерму и заходит глубоко в устьичные щели. На поверхности эпидермы образуются восковые выделения – зёрна, чешуйки, есть различные виды волосков (трихомов). Густой покров из волосков уменьшает транспирацию. Также характерно погружение устьиц в ямки (крипты) – и для суккулентов, их стенки могут иметь сложную конфигурацию.
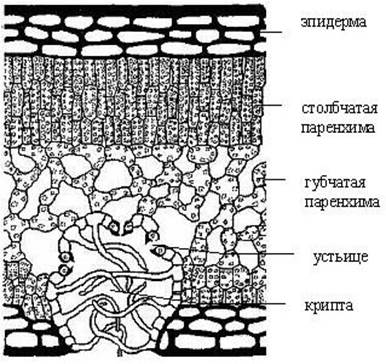
Рис. 4. Поперечный срез листа олеандра (Oleander sp.).
Внутреннюю ткань склерофитов отличает также склерификация – сокращение межклеточных пространств и внутренней испаряющей поверхности. Склерификация может захватывать многие органы. Она образует тяжи и слои, прилегающие к эпидерме. Образование твёрдых шипов по краю листа часто сопровождает склерификацию – преобразование побега в колючку. Листья многих злаков имеют различные приспособления к свёртыванию при недостатке влаги. У луговика дернистого - щучки (Deschampsia cespitosa) склеренхима залегает под листом, а устьица наверху на боковых сторонах гребней. В углублениях между гребнями находятся моторные клетки, которые представляют собой большие живые, с тонкими стенками клетки, способные изменять объём. Они раскрывают или закрывают лист в зависимости от влажности.
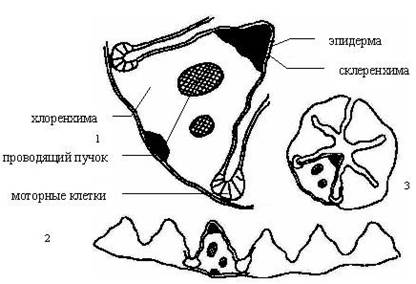
Рис. 5. Поперечный разрез листа луговика дернистого Deschampsia cespitosa:
1 – часть пластинки листа при большом увеличении;
2 – разрез всей пластинки листа;
3 – пластинка листа в свёрнутом состоянии.
Редукция листьев характерна для многих кустарников пустынь Средней Азии, Средиземноморья (саксаул, эфедра), их стебли фотосинтезируют, а листья либо недоразвиты, либо опадают рано весной. В стеблях над эпидермой хорошо развита столбчатая (палисадная) паренхима. Ксерофиты приспособлены к яркому освещению, поэтому не всегда можно разграничить признаки к недостатку воды и к яркому освещению. Корневая система очень обширна с глубоко идущими в почву корнями или распростёрта для использования кратковременных дождей.
Andromeda polifolia, Deschampsia cespitosa, Festuca ovina, Hieracium pilosella, Oxycoccus microcarpus, Vaccinium vitis-idaea.
Мезофиты.
Растения, живущие в условиях слабого увлажнения, находящиеся между ксерофитами и гидрофитами – имеют промежуточные черты гидроморфной и ксероморфной организации. К ним относятся лиственные древесные породы, сорняки, культурные растения. Для них характерно высокое содержание воды в тканях. Они слабо регулируют водный обмен.
Convallaria majalis, Geranium sylvaticum, Gymnocarpium dryopteris, Oxalis acetosella, Sorbus gorodkovii, Trifolium pratense.
Психрофиты.
Растения, приспосабливающиеся к влажным и холодным местообитаниям – высокогорья, средние широты.
Криофиты.
Растения, приспосабливающиеся к сухим и холодным местообитаниям. Это зимнезелёные растения с широкими кожистыми листьями. У них имеется толстая губчатая паренхима, крупные клетки и межклетники, слабо развитая механическая ткань, мало устьиц.
Нет резкой границы между психрофитами и криофитами. Их структура объясняется «физиологической» сухостью. Ксероморфная структура тундровых растений больше связана с холодным климатом и недостатком питания, чем с недостатком воды.
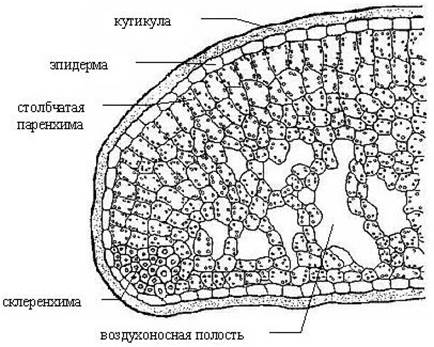
Рис. 6. Поперечный разрез листа брусники Vaccinium vitis-idaea.
II. По отношению к субстрату:
Эвтрофы.
Растения богатых минеральными солями почв, которые характерны для пойменных лугов, низинных болот и лиственных лесов. Среди эвтрофов выделяют особые группы растений, требующих большого содержания в почве определенных химических соединений. Так, нитрофилы требуют большого содержания в почве солей азота. Большинство из них относятся к сорным и рудеральным растениям (Chenopodium album, Urtica dioica). Кальцефилы предпочитают карбонатные, т. е. богатые солями кальция, почвы (Circaea alpina, Cypripedium calceolus, Epilobium alsinifolium, E. davuricum, Larix sibirica).
Мезотрофы.
Растения, которые по требовательности к минеральному богатству почвы занимают промежуточное положение между олиготрофами и эвтрофами (Achillea millefolium, Picea obovata).
Олиготрофы.
Растения, довольствующиеся небольшим содержанием минеральных солей в почве и произрастающие на скалах, болотах и подзолистых почвах (Andromeda polifolia, Arcostahylos uva-ursi, Chamaedaphne calyculata, Hieracium pilosa, Ledum palustre). Обладают чертами ксероморфизма. При избытке влаги даже они имеют мелкие жёсткие листья и другие признаки, говорящие об экономии воды. Среди них выделяют психрофиты - растения холодных, кислых и избыточно увлажнённых почв. Морфология и анатомия растений на недостаток почвенного питания сходна с типами строения растений на недостаток влаги. Особенности их структуры заключаются в сочетании ксероморфных признаков с наличием обширных межклетников, необходимых для аэрации, восполнении недостатка кислорода в болотной почве.
Рис. 7. Поперечный разрез листа багульника болотного Ledum palustre.
Галофиты.
Растения, живущие на сильно засолённых субстратах. Обладают чертами ксероморфизма. Так как поступление воды плохое, они обладают высоким осмотическим потенциалом клеточного сока, что является необходимым условием для всасывания воды и поддержания равновесия с внешней средой. Некоторые из них суккуленты (Mertensia maritima, Plantago maritima, Salicornia sp., Salsola sp.). Другие имеют жёсткие мелкие листья, выделяющие кристаллики соли.
Один и тот же вид может встречаться в разных экологических условиях и его трудно отнести к строго определенной экологической группе. В этом случае растение характеризуют, например, как мезогигрофит, олигомезотроф и т.д.
III. По отношению к свету:
Наиболее сильно сказывается влияние света на форме и анатомической структуре листьев. Многие признаки световых листьев совпадают с признаками ксероморфизма, так как прямому яркому освещению сопутствует всегда и нагревание и повышенная транспирация. Наоборот, гигроморфные признаки наблюдаются у теневых листьев.
Светолюбивые - гелиофиты.
Болотные, луговые, реже лесные растения верхнего яруса.
(Alopecurus pratensis, Betula nana).
Похожие статьи:
poznayka.org
Светолюбивые - гелиофиты.
ВЛИЯНИЕ УСЛОВИЙ ВНЕШНЕЙ СРЕДЫ НА МОРФОЛОГИЧЕСКОЕ И АНАТОМИЧЕСКОЕ СТРОЕНИЕ ОРГАНОВ РАСТЕНИЙ. ЭКОЛОГИЧЕСКИЕ ФАКТОРЫ И ЭКОЛОГИЧЕСКИЕ ГРУППЫ РАСТЕНИЙ.
Экологические группы выделяются по отношению к одному фактору среды, который играет большое значение в формировании и изменении органов высшего растения.
Наиболее важными экологическими факторами, влияющими на структуру растения являются:
- влажность воздуха, почвы;
- свет;
- температурные условия;
- особенности почв;
- конкуренция сообществ и т.д.
В связи с экологическими факторами выделяют соответствующие экологические группы. Экологическая группа растений - это совокупность видов или их популяций, произрастающих в определенных близких экологических условиях.
К одним и тем же условиям среды растения приспосабливаются по-разному, поэтому в пределах одной экологической группы можно найти растения, резко отличающиеся по внешнему облику друг от друга, и ещё по анатомическому строению, то есть растения имеют различнуюжизненную форму (биоморфу) – своеобразный внешний облик, возникающий в результате индивидуального развития (онтогенеза) в определённых экологических условиях и отражающий совокупность основных приспособительных черт. Жизненная форма растений зависит от особенностей роста подземных и надземных вегетативных органов, это результат длительной эволюции, она закреплена наследственно у особей одного и того же вида, но в зависимости от конкретных условий среды может варьировать. Экологические группы и формы растений между собой тесно связаны.
I. По отношению к влаге:
Гидрофиты.
Растения, нормально растущие в воде, встречающиеся часто по берегам водоёмов. В связи со спецификой местообитания у них наблюдаются следующие особенности – губчатость, рыхлость тканей, наличие крупных межклетников, то есть из тканей преобладает аэренхима. Система воздухоносных межклетников очень хорошо развита и является типичной чертой их строения. Полости частей растения, погружённых в воду, обычно сообщаются с устьицами непогружённых листьев, что является важным для газообмена. Воздушные полости есть во всех органах, даже в плодах (Nuphar lutea), что обеспечивает плавучесть и распространение семян. Губчатость тканей и наличие воздушных полостей в листовых пластинках и черешках также поддерживает плавучесть растения. У полностью погружённых растений (Elodea sp.) воздушные полости могут аккумулировать кислород в течение дня, который затем используется растением на дыхание ночью. У подводных растений в органах нет кутикулы и перидермы, устьица не функционируют. Отсутствие кутина и суберина способствует поглощению воды и питательных веществ непосредственно поверхностью тела, что является важным для питания растения. Корни обычно короткие и не разветвлённые, их окончания лишены корневых волосков. Отсутствуют механические и водопроводящие ткани. Недостаток арматурных тканей компенсируется плавучестью отдельных органов.
В зависимости от того, со сколькими средами контактирует растение, выделяются следующие подгруппы гидрофитов:
а) плавающие на поверхности, но не связанные с почвой:
Lemna minor, Salvinia sp.
б) полностью погруженные в воде, взвешенные в её толще,заселяют хорошо освещённые и аэрируемые слом воды, не связаны с почвой:
Batrachium eradicatum, Lemna trisulca, Sargassum sp., Utricularia intermedia.
в) полностью погруженные, плавающие, укореняющиеся:
Zostera marina, Isöetes lacustris, Potamogeton natans.
г) плавающие на поверхности, но укореняющиеся:
Batrachium peltatum, Nuphar lutea, Nymphaea candida, Sparganium sp.
Гелофиты.
Травянистые растения, вегетативные побеги которых располагаются в воздушной среде, а почки возобновления в неблагоприятный для вегетации период находятся под водой или в почве. Экологически разнообразная группа, занимающая переходное положение между световыми гигрофитами и гидрофитами. Нередко к ним относят только болотные или прибрежно-водные растения (амфифиты) – укореняющиеся растения мелководных местообитаний, стебли и листья которых высоко возвышаются, есть мощное корневище с толстыми придаточными корнями. Хорошо размножаются вегетативно.
Alisma plantago-aquatica, Calla palustris, Carex sp., Menyanthes trifoilata, Sagittaria sagittifolia, Scirpus lacustris, Typha latifolia.
Рис. 1. Поперечный разрез стебля белокрыльника болотного Calla palustris или частухи подорожниковой Alisma plantago-aquatica.
Гигрофиты.
Сухопутные растения, онтогенез которых проходит при хороших условиях – достаточной влажности воздуха, поэтому у них нет особых анатомо-морфологических приспособлений к засухе, слабо приспособлены к изменению влажного режима. Подземные органы, а часто и основания побегов таких растений находятся в воде. Корни и листья этих растений снабжены аэренхимой, в листьях слабо развита столбчатая паренхима и склеренхима, кутикула не выражена, клетки эпидермы имеют тонкие стенки. Это влаголюбивые растения - обитатели влажной и тёплой атмосферы, обитающие по берегам водоемов (Carex aquatilis), на болотах (Menyanthes trifoliata), на низинных лугах (Bistorta major), в травянистых лесах, тенистых лесах умеренной зоны, где воздух должен быть насыщен парами воды.
Гигрофиты умеренной зоны по отношению к свету делятся на следующиеподгруппы:
а) теневые гигрофиты тенистых лесов, имеют тонкие листья, нуждаются в высокой влажности приземного слоя воздуха:
Adoxa moschatellina;
б) световые (солнечные) гигрофиты, растущие на открытых местах, но на постоянно сырых почвах:
Caltha palustris, Cardamine sp., Carex sp.
Ксерофиты.
Растения этой группы имеют разнообразные приспособления к поддержанию водного баланса при недостатке почвенной и атмосферной влаги. Это самая разнообразная морфологически группа растений. Способность переносить неблагоприятные условия связана с морфологическими признаками, по которым среди них выделяют:
а) суккуленты – сочные, мясистые растения с сильно развитой водозапасающей паренхимой в листьях (Alöe, Agava), в стебле – кактусы, молочаи, в корне (Ceiba). Суккуленты – основные ландшафтные растения пустынь Центральной Америки (кактусы) и Африки (молочаи). Суккулентность растения связана с разрастанием клеток паренхимы при одновременном увеличении вакуолей и сильном сокращении межклетников. Основная отличительная черта суккулентов – способность запасать большое количество воды, сохранять и экономно её расходовать. Суккуленты имеют низкую интенсивность транспирации. Днём ни закрывают устьица. Потери воды могут быть уменьшены за счёт формы растения. Форма и замыкание устьиц днём затрудняет газообмен, поэтому у суккулентов особый цикл фотосинтеза. Многие кактусы фотосинтезируют в течение почти всего года, но образуют небольшое количество углеводов, что замедляет их рост. Корневая система у них небольшая, они используют воду из поверхностных слоёв почвы.
Рис. 2. Поперечный разрез листа алоэ Alöe sp.
Рис. 3. Поперечный разрез толстянки Crassula falcata с пузыревидными клетками эпидермы.
В нашей флоре мало суккулентов: Mertensia maritima, Rhodiola arctica, Rh. rosea, Sedum acre.
б) склерофиты – по внешнему облику это суховатые и жёсткие растения с мало обводнёнными тканями. Они обладают высокой устойчивостью к завяданию, могут без заметного вреда терять до 25% воды. Имеют высокое осмотическое давление клеточного сока, что позволяет им увеличивать сосущую силу корней и получать воду из сухих почв. Корневая система развита сильнее надземной. Ксероморфные признаки проявляются, прежде всего, в особенностях строения эпидермы, клетки которой обладают утолщёнными наружными стенками. Мощная кутикула покрывает эпидерму и заходит глубоко в устьичные щели. На поверхности эпидермы образуются восковые выделения – зёрна, чешуйки, есть различные виды волосков (трихомов). Густой покров из волосков уменьшает транспирацию. Также характерно погружение устьиц в ямки (крипты) – и для суккулентов, их стенки могут иметь сложную конфигурацию.
Рис. 4. Поперечный срез листа олеандра (Oleander sp.).
Внутреннюю ткань склерофитов отличает также склерификация – сокращение межклеточных пространств и внутренней испаряющей поверхности. Склерификация может захватывать многие органы. Она образует тяжи и слои, прилегающие к эпидерме. Образование твёрдых шипов по краю листа часто сопровождает склерификацию – преобразование побега в колючку. Листья многих злаков имеют различные приспособления к свёртыванию при недостатке влаги. У луговика дернистого - щучки (Deschampsia cespitosa) склеренхима залегает под листом, а устьица наверху на боковых сторонах гребней. В углублениях между гребнями находятся моторные клетки, которые представляют собой большие живые, с тонкими стенками клетки, способные изменять объём. Они раскрывают или закрывают лист в зависимости от влажности.
Рис. 5. Поперечный разрез листа луговика дернистого Deschampsia cespitosa:
1 – часть пластинки листа при большом увеличении;
2 – разрез всей пластинки листа;
3 – пластинка листа в свёрнутом состоянии.
Редукция листьев характерна для многих кустарников пустынь Средней Азии, Средиземноморья (саксаул, эфедра), их стебли фотосинтезируют, а листья либо недоразвиты, либо опадают рано весной. В стеблях над эпидермой хорошо развита столбчатая (палисадная) паренхима. Ксерофиты приспособлены к яркому освещению, поэтому не всегда можно разграничить признаки к недостатку воды и к яркому освещению. Корневая система очень обширна с глубоко идущими в почву корнями или распростёрта для использования кратковременных дождей.
Andromeda polifolia, Deschampsia cespitosa, Festuca ovina, Hieracium pilosella, Oxycoccus microcarpus, Vaccinium vitis-idaea.
Мезофиты.
Растения, живущие в условиях слабого увлажнения, находящиеся между ксерофитами и гидрофитами – имеют промежуточные черты гидроморфной и ксероморфной организации. К ним относятся лиственные древесные породы, сорняки, культурные растения. Для них характерно высокое содержание воды в тканях. Они слабо регулируют водный обмен.
Convallaria majalis, Geranium sylvaticum, Gymnocarpium dryopteris, Oxalis acetosella, Sorbus gorodkovii, Trifolium pratense.
Психрофиты.
Растения, приспосабливающиеся к влажным и холодным местообитаниям – высокогорья, средние широты.
Криофиты.
Растения, приспосабливающиеся к сухим и холодным местообитаниям. Это зимнезелёные растения с широкими кожистыми листьями. У них имеется толстая губчатая паренхима, крупные клетки и межклетники, слабо развитая механическая ткань, мало устьиц.
Нет резкой границы между психрофитами и криофитами. Их структура объясняется «физиологической» сухостью. Ксероморфная структура тундровых растений больше связана с холодным климатом и недостатком питания, чем с недостатком воды.
Рис. 6. Поперечный разрез листа брусники Vaccinium vitis-idaea.
II. По отношению к субстрату:
Эвтрофы.
Растения богатых минеральными солями почв, которые характерны для пойменных лугов, низинных болот и лиственных лесов. Среди эвтрофов выделяют особые группы растений, требующих большого содержания в почве определенных химических соединений. Так, нитрофилы требуют большого содержания в почве солей азота. Большинство из них относятся к сорным и рудеральным растениям (Chenopodium album, Urtica dioica). Кальцефилы предпочитают карбонатные, т. е. богатые солями кальция, почвы (Circaea alpina, Cypripedium calceolus, Epilobium alsinifolium, E. davuricum, Larix sibirica).
Мезотрофы.
Растения, которые по требовательности к минеральному богатству почвы занимают промежуточное положение между олиготрофами и эвтрофами (Achillea millefolium, Picea obovata).
Олиготрофы.
Растения, довольствующиеся небольшим содержанием минеральных солей в почве и произрастающие на скалах, болотах и подзолистых почвах (Andromeda polifolia, Arcostahylos uva-ursi, Chamaedaphne calyculata, Hieracium pilosa, Ledum palustre). Обладают чертами ксероморфизма. При избытке влаги даже они имеют мелкие жёсткие листья и другие признаки, говорящие об экономии воды. Среди них выделяют психрофиты - растения холодных, кислых и избыточно увлажнённых почв. Морфология и анатомия растений на недостаток почвенного питания сходна с типами строения растений на недостаток влаги. Особенности их структуры заключаются в сочетании ксероморфных признаков с наличием обширных межклетников, необходимых для аэрации, восполнении недостатка кислорода в болотной почве.
Рис. 7. Поперечный разрез листа багульника болотного Ledum palustre.
Галофиты.
Растения, живущие на сильно засолённых субстратах. Обладают чертами ксероморфизма. Так как поступление воды плохое, они обладают высоким осмотическим потенциалом клеточного сока, что является необходимым условием для всасывания воды и поддержания равновесия с внешней средой. Некоторые из них суккуленты (Mertensia maritima, Plantago maritima, Salicornia sp., Salsola sp.). Другие имеют жёсткие мелкие листья, выделяющие кристаллики соли.
Один и тот же вид может встречаться в разных экологических условиях и его трудно отнести к строго определенной экологической группе. В этом случае растение характеризуют, например, как мезогигрофит, олигомезотроф и т.д.
III. По отношению к свету:
Наиболее сильно сказывается влияние света на форме и анатомической структуре листьев. Многие признаки световых листьев совпадают с признаками ксероморфизма, так как прямому яркому освещению сопутствует всегда и нагревание и повышенная транспирация. Наоборот, гигроморфные признаки наблюдаются у теневых листьев.
Светолюбивые - гелиофиты.
Болотные, луговые, реже лесные растения верхнего яруса.
(Alopecurus pratensis, Betula nana).
Похожие статьи:
www.poznayka.org
Гелофиты Википедия

Гигрофи́ты (от — влажный + — растение) — растения, обитающие в местах с высокой влажностью воздуха и (или) почвы. Категория гигрофитов выделяется на основе особенностей физиологии и морфологии растений; наиболее характерный признак гигрофитов — отсутствие приспособлений, ограничивающих расходование влаги.
Гигрофиты, как правило, отличаются большими и тонкими листьями с незначительным поверхностным защитным слоем (кутикулой), что приводит к высокой кутикулярной транспирации; незначительным утолщением эпидермиса, слаборазвитой сеткой жилок; устьица у листьев крупные, но их удельное количество на единицу площади невелико. Часто листья на поверхности имеют волоски из живых клеток эпидермиса для увеличения площади листа. За счёт крупных устьиц растения достигают оптимального регулирования транспирации. Морфологической особенностью листьев гигрофитов является наличие специфических устьиц — гидатод, которые способны выделять влагу из листа в капельно-жидкой форме. Это необходимо растению для нормальной жизнедеятельности в условиях атмосферы, перенасыщенной водяными испарениями. Выделение капель воды из листа называется гуттацией.
Среди других морфологических особенностей гигрофитов — слаборазвитая корневая система, наличие длинных стеблей при неразвитости механических тканей, слабое одревеснение стенок сосудов. Подобные особенности приводят к увяданию даже при незначительной нехватке влаги.
Наиболее характерными гигрофитами являются травянистые растения влажных тропических лесов.
Болотные травянистые растения обычно также относят к гигрофитам, однако по ряду морфологических признаков они имеют отличия. В силу постоянного воздействия солнечного света и более сухой атмосферы у них наличествует более толстая кутикула, что приводит к уменьшению кутикулярной транспирации; а листья обычно толще и меньше. Иногда болотные растения выделяют в особую подгруппу — гелофиты ( — болото).
К гигрофитам близки гидрофиты (водные растения, погружённые в воду нижней частью) и гидатофиты (водные растения, полностью или бо́льшей частью находящиеся в воде). Иногда в силу схожести морфологии и условий жизнеобитания все эти группы рассматривают как одну.
См. также
Ссылки
wikiredia.ru
ГЕЛОФИТЫ (ГИДРОГИГРОФИТЫ) – ВОДНО-БОЛОТНЫЕ
Экология  ГЕЛОФИТЫ (ГИДРОГИГРОФИТЫ) – ВОДНО-БОЛОТНЫЕ
ГЕЛОФИТЫ (ГИДРОГИГРОФИТЫ) – ВОДНО-БОЛОТНЫЕ
просмотров - 81
РАСТЕНИЯ:
1. Надводные растения с поднимающимися над поверхностью воды стеблями и
листьями, укореняющиеся (тростник, рогоз, камыш, сусак, ежеголовник,
стрелолист, частуха и др.) Все они успешно развиваются и проходят полный
цикл развития, как в воде, так и на влажных берегах водоемов.
Глава I. Характеристика базовых групп растений по отношению к воде.
По приуроченности к местообитаниям с разными условиями увлажнения и выработке соответствующих приспособлений среди наземных растений различают три базовых экологических типа: гигрофиты, мезофиты и ксерофиты.
Растения, для которых вода не только необходимый экологический фактор, но непосредственная среда обитания, относятся к водным, называемым гидрофитами.
ГИГРОФИТЫ. Это растения избыточно увлажненных местообитаний с высокой влажностью воздуха и почвы. При довольно большом разнообразии местообитаний, особенностей водного режима и анатомо-морфологических черт всех гигрофитов объединяет отсутствие приспособлений, ограничивающих расход воды, и неспособность выносить даже незначительную ее потерю. Ярко выраженные гигрофиты - травянистые растения и эпифиты влажных тропических лесов, не выносящие сколько-нибудь заметного понижения влажности воздуха. Даже в разгар сезона дождей мелкие эпифитные папоротники на стволах деревьев теряют тургор и засыхают, если на них в течение 2-3 часов падают солнечные лучи. Черты гигрофитов имеют травянистые растения темнохвойных лесов (кислица, майник двулистный, двулепестник альпийский). К гигрофитам можно отнести и виды, растущие на открытых и хорошо освещенных местообитаниях, но в условиях избытка почвенной влаги - близ водоемов, в дельтах рек, в местах выхода грунтовых вод. В наших широтах примером могут служить прибрежные виды: калужница - Caltha palustris, плакун-трава - Lythrum salikaria, а в странах жаркого климата - папирус, болотные пальмы. Из культурных растений сюда можно отнести рис, культивируемый на полях, залитых водой.
КСЕРОФИТЫ. Это растения сухих местообитаний, способные переносить значительный недостаток влаги - почвенную и атмосферную засуху. Οʜᴎ распространены, обильны и разнообразны в областях с жарким и сухим климатом. К этой группе принадлежат виды пустынь, сухих степей, саванн, колючих редколесий, сухих субтропиков. В более гумидных районах ксерофиты принимают участие в растительном покрове лишь в наиболее прогреваемых и наименее увлажненных местообитаниях (к примеру, на склонах южной экспозиции).
Неблагоприятный водный режим растений в сухих местообитаниях обусловлен, во-первых, ограниченным поступлением воды при ее недостатке в почве и, во-вторых, увеличением расхода влаги на транспирацию при большой сухости воздуха и высоких температурах. Следовательно, для преодоления недостатка влаги возможны разные пути: увеличение ее поглощения и сокращение расхода, кроме того, способность переносить большие потери воды. Все это используется ксерофитами при адаптации к сухости, но у разных растений в неодинаковой степени, в связи с чем некоторые авторы различают два базовых способа преодоления ксерофитами засухи: возможность противостоять иссушению тканей, или активное регулирование водного баланса, и способность выносить сильное иссушение.
Учитывая зависимость отструктурных черт и способов регулирования водного режима различают несколько разновидностей ксерофитов (по Генкелю П.А.): эуксерофиты, гемиксерофиты, пойкилоксерофиты.
К группе ксерофитов относят и суккуленты - растения с сочными листьями или стеблями. Различают листовые суккуленты (агавы, алоэ) и стеблевые, у которых листья редуцированы, а наземные части представлены мясистыми стеблями(кактусы, некоторые молочаи).
Ксерофиты с наиболее ярко выраженными ксероморфными чертами строения листьев имеют своеобразный внешний облик, за что получили название склерофитов. Облик типичного склерофита легко представить на примере чертополоха - Carduus crispus и пустынных полыней, ковылей, саксаулов.
МЕЗОФИТЫ. Эта группа включает растения, произрастающие в средних условиях увлажнения. Сюда относятся растения лугов, травяного покрова лесов, лиственные древесные и кустарниковые породы из областей умеренно влажного климата͵ а также большинство культурных растений.
Мезофиты - группа весьма разнообразная не только по видовому составу, но и по различным экологическим оттенкам, обусловленным разным сочетанием факторов в природных местообитаниях. Οʜᴎ связаны переходами с другими экологическими типами растений по отношению к воде, так что четкую границу между ними провести очень трудно. Так, среди луговых мезофитов выделяются виды с повышенным влаголюбием, предпочитающие постоянно сырые или временно заливаемые участки (лисохвост луговой - Alopecurus pratensis, бекмания обыкновенная - Beckmannia eruciformis).
Их объединяют в переходную группу гигромезофитов наряду с некоторыми влаголюбивыми лесными травами, предпочитающими наиболее сырые леса, лесные овраги (недотрога - Impatiens nolitangere). С другой стороны в местообитаниях с переодическим или постоянным (небольшим) недостатком влаги много мезофитов с теми или иными ксероморфными признаками с повышенной физиологической устойчивостью к засухе. Эта группа переходная между мезофитами ксерофитами, - ксеромезофиты. Примером могут служить многие виды северных степей, сухих сосновых боров, песчаных местообитаний: клевер-белоголовка - Trifolium montanum, подмаренник желтый - Galium verum и другие.
Особое место среди мезофитов занимают степные и пустынные весенние эфемеры и эфемероиды. К этой группе принадлежат растения, ранней весной покрывающие степи и пустыни разноцветным цветущим ковром (многолетники - тюльпаны, гусиные луки; однолетники - маки, вероники). Это виды с чрезвычайно краткой вегетацией и длительным периодом покоя, который однолетние эфемеры переживают в виде семян, а многолетние эфемероиды - в виде покоящихся луковиц, клубней, корневищ. Кроме весенних существуют и осенние эфемероиды, произрастающие в районах с климатическим ритмом средиземноморского типа. Сюда относятся виды родов Crocus, Scilla и другие.
По многим особенностям структуры и физиологии близки к ксерофитам растения, которые по тем или иным причинам испытывают недостаток влаги, сопряженный с действием низких температур. Иногда такие виды в качестве особого подразделения включают в группу ксерофитов, иногда выделяют в самостоятельные экологические типы - психрофиты и криофиты.
Психрофиты - растения влажных и холодных почв в холодных местообитаниях высокогорий и северных широт. Несмотря на достаточное увлажнение почвы, они часто испытывают недостаток влаги (или из-за физиологической сухости, вызванной низкими температурами, или в связи с преобладанием в почве недоступной влаги, как, к примеру, на торфянистых почвах). Среди психрофитов есть травянистые растения (к примеру злаки северных лугов: белоус - Nardus strikta; высокогорные кавказские злаки: овсяница пестрая -Festuka varia), высокогорные, болотные и тундровые кустарники и кустарнички, как вечнозеленые (вереск - Calluna vulgaris), так и с опадающей листвой (карликовые ивы - Salix polaris, S. herbacea).К психрофитам относятся и хвойные древесные породы умеренных и северных широт.
Криофиты в экологическом отношении очень близки к психрофитам и связаны с ними переходными формами. Это растения сухих и холодных местообитаний - сухих участков тундр, скал, осыпей. Обычно они рассматриваются и характеризуются вместе с психрофитами, поскольку у них много сходных морфологических и физиологических черт. Но среди криофитов есть и весьма своеобразные формы - это растения-подушки высокогорных холодных пустынь.
ГИДРОФИТЫ. Это водные растения. По образу жизни и строению среди них можно выделить погруженные растения и растения с плавающими листьями. Погруженные растения подразделяют на укореняющиеся в донном грунте и взвешенные в толще воды. Из высших растений к первым принадлежат телорез - Stratiotes aloides, шильник водяной - Subularia aquatika. В эту же группу входят водоросли, прикрепленные к грунту. Из растений, взвешенных в толще воды, можно назвать роголистник погруженный--Ceratophyllum demersum, пузырчатку обыкновенную - Utrikularia vulgaris, а также многочисленные виды планктонных водорослей.
Растения с плавающими листьями используют частично водную, частично воздушную среду. Из них укореняются в грунте кувшинки из рода Nymphaea, кубышки из рода Nuphar, рдесты, орех водяной - Trapa natans.
Многие виды наряду с плавающими на поверхности воды листьями имеют и подводные. Плавают на поверхности воды, не укореняясь, ряски, водокрас.
К настоящим водным растениям очень близко примыкает и обычно вместе с ними рассматривается группа гелофитов или амфибий - земноводных растений. Это виды береговых и прибрежных местообитаний с избыточным или переменным увлажнением. Οʜᴎ могут расти как в воздушной среде, так и частично погруженными в воду, могут выносить и полное временное заливание. Как в природе нет резкой границы между водными и наземными местообитаниями для растений, так и группа гелофитов связана незаметными переходами, с одной стороны, с настоящими гидрофитами, с другой - с наземными гигрофитами и гигромезофитами. Примеры гелофитов - растений прибрежной полосы пресноводных водоемов и рек: стрелолист - Sagittaria sagittifolia, ежеголовка - Sparganium ramosum.
ГЛАВА II. Анатомо-морфологические приспособления растений к водному режиму.
Растения, приуроченные к местообитаниям с разными условиями увлажнения, выработали соответствующие приспособления к водному режиму.
Характерные структурные черты гигрофитов - тонкие нежные листовые пластинки с небольшим числом устьиц, не имеющие толстой кутикулы, рыхлое сложение тканей листа с крупными межклетниками, слабое развитие водопроводящей ткани, тонкие слаборазветвленные корни.
Для ксерофитов большое значение имеют разнообразные структурные приспособления к условиям недостатка влаги.
Корневые системы обычно сильно развиты, что помогает растениям увеличить поглощение почвенной влаги. По общей массе корневые системы ксерофитов нередко превышают надземные части, иногда весьма значительно. Так, у многих травянистых и кустарниковых видов среднеазиатских пустынь подземная масса больше надземной в 9-10 раз, а у кесрофитов памирских высокогорных холодных пустынь - в300-400 раз. Корневые системы ксерофитов часто бывают экстенсивного типа, то есть растения имеют длинные корни, распространяющиеся в большом объеме почвы, но сравнительно мало разветвленные. Проникновение таких корней на большую глубину позволяет ксерофитам использовать влагу глубоких почвенных горизонтов, а в отдельных случаях - и грунтовых вод.
У других видов корневые системы интенсивного типа: они охватывают относительно небольшой объем почвы, но благодаря очень густому ветвлению максимально используют почвенную влагу. Корни ряда ксерофильных видов имеют специальные приспособления для запасания влаги. Надземные органы ксерофитов также отличаются своеобразными (так называемыми ксероморфными чертами), которые носят отпечаток трудных условий водоснабжения. У них сильно развита водопроводящая система, что хорошо заметно по густоте сети жилок в листьях, подводящих воду к тканям. Эта черта облегчает ксерофитам пополнение запасов влаги, расходуемой на транспирацию.
Разнообразные структурные приспособления защитного характера, направленные на уменьшение расхода воды, в основном сводятся к следующему:
1.Общее сокращение транспирирующей поверхности. Многие ксерофиты имеют мелкие, узкие, сильно редуцированные листовые пластинки. В особо засушливых пустынных местообитаниях листья некоторых древесных и кустарниковых пород редуцированы до едва заметных чешуек. У таких видов фотосинтез осуществляют зеленые ветви.
2.Уменьшение листовой поверхности в наиболее жаркие и сухие периоды вегетационного сезона. Для многих кустарников среднеазиатских, североафриканских и других пустынь, а так же для некоторых видов сухих субтропиков средиземноморья характерен сезонный деформизм листьев: ранней весной при еще благоприятном водном режиме образуются относительно крупные листья, которые летом, при наступлении жары и сухости, сменяются мелкими листьями более ксероморфного строения с меньшей интенсивностью транспирации.
3.Защита листьев от больших потерь влаги на транспирацию. Она достигается благодаря развитию мощных покровных тканей - толстостенного, иногда многослойного эпидермиса, часто несущего различные выросты и волоски, которые образуют густое “войлочное” опушение поверхности листа. У других видов поверхность покрыта водонепроницаемым слоем толстой кутикулы или воскового налета. Развитие защитных покровов на листьях причина того, что степной травостой имеет тусклые, седоватые оттенки, резко отличающиеся от яркой зелени лугов.
Устьица у ксерофитов обычно защищены от чрезмерной потери влаги, к примеру, расположены в специальных углублениях в ткани листа͵ иногда снабженных волосками и прочими дополнительными защитными устройствами. У ковылей и других степных злаков существует интересный механизм защиты устьиц в самые жаркие и сухие часы дня: при больших потерях воды крупные тонкостенные водоносные клетки эпидермиса теряют тургор, и лист свертывается в трубку; так устьица оказываются изолированными от окружающего сухого воздуха внутри замкнутой полости, где благодаря транспирации создается повышенная влажность. Во влажную погоду клетки эпидермиса восстанавливают тургор, и листовая пластинка вновь развертывается.
4.Усиленное развитие механической ткани.
Клетки тканей листьев у ксерофитов отличаются мелкими размерами и весьма плотной упаковкой, то есть малым развитием межклетников, благодаря чему сильно сокращается внутренняя испаряющая поверхность листа. Поскольку ксерофиты обычно обитают на открытых, хорошо освещенных местообитаниях, многие черты ксероморфной структуры листа - это одновременно и черты световой структуры. Так у многих видов листья имеют мощную иногда многорядную палисадную паренхиму, часто расположенную с обеих сторон.
Основные морфолого-анотомические черты мезофитов - средние между чертами гигрофитов и ксерофитов. Мезофиты имеют умеренно развитые корневые системы как экстенсивного, так и интенсивного типа, со всеми переходами между ними. Для листа характерна дифференцировка тканей на более или менее плотную палисадную паренхиму и рыхлую губчатую паренхиму с системой межклетников. Сеть жилок сравнительно негустая. Покровные ткани могут иметь отдельные ксероморфные черты, но не столь ярко выраженные, как у ксерофитов.
Психрофиты имеют ярко выраженную ксероморфную структуру листа. Так, психрофильные злаки узколистны, имеют хорошо развитую проводящую и механическую ткани; некоторые из них способны к свертыванию листовой пластинки в трубку, напоминая степные ксерофиты. Вечнозеленые кустарнички имеют плотные кожистые листья, иногда с весьма мощной кутикулой, плотной палисадной паренхимой. Нижняя сторона, несущая устьица, часто опушена или покрыта восковым налетом. Вместе с тем в структуре листа ясно заметны и некоторые гигроморфные черты, главным образом, крупные размеры клеток и развитие больших межклетников в губчатой ткани.
Анатомо-морфологические черты гидрофитов существенно отличают их от наземных растений. Заметна редукция механических тканей. Столь же сильно редуцированна и проводящая система. В случае если у сухопутных мезофитов длина жилок на 1 кв.см. листа составляет около100 мм и более, а у ксерофитов доходит до300, то у водных и прибрежных растений она в несколько раз меньше. Вот несколько примеров (по Гесснеру Ф., 1959):
| Вид | Плавающие листья, мм | Подводные листья, мм | |
| Nelumbium nuciferum | -- | ||
| Victoria cruciana | -- | ||
| Victoria regia | -- | ||
| Potamogeton praelongus | -- | ||
У некоторых погруженных растений, не прикрепленных к грунту, корни полностью редуцированны, другие корни сохранили, но отдельно плавающие части растений могут обходиться и без них. Корни укрепляющихся гидрофитов слабоветвистые, без корневых волосков. Вместе с тем ряд видов имеет толстые и прочные корневища, которые играют роль якоря, хранилища запасных веществ и органа вегетативного размножения.
Листья погруженных гидрофитов очень тонки и нежны, имеют упрощенное строение мезофилла без заметной дифференциации на палисадную и губчатую паренхиму. Подводные листья без устьиц. В отдельных местах находятся группы клеток эпидермиса с утонченными стенками. Считается, что они играют большую роль в поглощении воды и растворенных минеральных солей.
У растений лишь частично погруженных в воду, хорошо выражена гетерофиллия - различие строения надводных и подводных листьев на одной и той же особи. Первые имеют черты, обычные для листьев наземных растений, вторые - очень тонкие или рассеченные листовые пластинки. Гетерофиллия отмечена у водного лютика - Ranunculus diversifolius, кувшинок и кубышек, стрелолиста и других видов. Интересный пример - поручейник, на стебле которого можно видеть несколько форм листьев, представляющих все переходы от типично наземных до водных.
Глава III.Физиологические адаптации растений, приуроченных к местообитаниям разной увлажненности.
Наряду с морфологическими особенностями у растений, приуроченных к местам с разными условиями увлажненности, выработались и физиологические.
Способность гигрофитов к регуляции водного режима ограничена: устьица большей частью широко открыты, так что транспирация мало отличается от физического испарения. Благодаря беспрепятственному потоку воды и отсутствию защитных приспособлений интенсивность транспирации очень высока: у световых гигрофитов в дневное время листья могут терять за час количество воды, в 4-5 раз превышающее массу листа. Высокая оводненность тканей гигрофитов поддерживается в основном за счет постоянного притока влаги из окружающей среды.
Другие характерные физиологические черты гигрофитов, обусловленные легкой доступностью влаги, - низкое осмотическое давление клеточного сока, незначительная водоудерживающая способность, приводящая к быстрой потере запасов воды.особенно показательны для гигрофитов небольшие величины сублетального водного дефицита: так, для кислицы и майника потеря 15%-20% запаса воды уже необратима и ведет к гибели. В некоторых случаях у растений сильно увлажненных местообитаний возникает крайне важность удаления избытка влаги. Обычно, это бывает, когда почва хорошо прогрета и корни активно всасывают воду, а транспирация отсутствует (к примеру, утром или при тумане, когда влажность воздуха 100%). Избыточная влага удаляется путем гуттации - выделение воды через специальные выделительные клетки, расположенные по краю или на острие листа.
Ксерофиты обладают рядом разнообразных физиологических адаптаций, позволяющих им успешно выдерживать недостаток влаги.
У ксерофитов обычно повышено осмотическое давление клеточного сока, позволяющее всасывать воду даже при больших водоотнимающих силах почвы, то есть использовать не только легкодоступную, но и труднодоступную почвенную влагу. Оно измеряется тысячами кПа, а у некоторых пустынных кустарников зарегистрированы цифры, достигающие 10000-30000 кПа.
С давних пор пристальное внимание привлекала проблема расхода воды ксерофитами на транспирацию. Казалось бы, многочисленные анатомические приспособления, достаточно надежно защищающие наземные части ксерофитов от сильного испарения, должны способствовать значительному снижению транспирации. При этом выяснилось, что в действительности это не так. При достаточном водоснабжении большинство ксерофитов имеют довольно высокую транспирацию, но при наступлении засушливых условий, они сильно сокращают ее. При этом играет роль и закрывание устьиц, и сильное обезвоживание листа при начинающемся подвядании. Несомненно, анатомо-морфологические приспособления имеют определенное значение, но основную роль в засухоустойчивости ксерофитов в настоящее время отводят физиологическим механизмам.
К числу этих механизмов принадлежит высокая водоудерживающая способность тканей и клеток, обусловленная рядом физиологических и биохимических особенностей.
Большое значение для выживания ксерофитов при резком недостатке влаги имеет их способность переносить глубокое обезвоживание тканей без потери жизнеспособности и способности восстановления нормального содержания воды в растении при возобновлении благоприятных условий. Ксерофиты способны потерять до 75% всего водного запаса и, тем не менее, остаться живыми. Ярким примером в этом отношении служат пустынные растения, которые летом высыхают до состояния, близкого к воздушно-сухому, и впадают в анабиоз, но после дождей возобновляют рост и развитие.
Еще одна система адаптаций, обеспечивающих выживание ксерофитов в аридных условиях, - выработка сезонных ритмов, дающих возможность растениям использовать для вегетации наиболее благоприятные периоды года и резко сократить жизнедеятельность во время засухи. Так, в областях со средиземноморским климатом с резко выраженным летним сухим периодом многие ксерофильные виды имеют “двухтактный” ритм сезонного развития: весенняя вегетация сменяется летним покоем, во время которого растения сбрасывают листву и снижают интенсивность физиологических процессов; в период осенних дождей вегетация возобновляется, и затем уже следует зимний покой. Сходное явление наблюдается и у растений сухих степей в середине и конце лета: потеря части листовой поверхности, приостановка развития, сильное обезвоживание тканей и т.д. Такое состояние, получившее название полупокоя, длится вплоть до осенних дождей, после которых у степных ксерофитов начинают отрастать листья.
Физиологические адаптации суккулентов столь своеобразны, что их крайне важно рассмотреть отдельно.
Основной способ преодоления засушливых условий у суккулентов - накопление больших запасов воды в тканях и крайне экономное ее расходование. В условиях жаркого и сухого климата весь водный запас мог бы быть быстро растрачен, но растения имеют защитные приспособления, направленные к сокращению транспирации. Одно из них - своеобразная форма надземных частей суккулентов. В дополнение к этому у многих суккулентов поверхность защищена восковым налетом опушением, хотя есть и суккуленты с тонким не защищенным эпидермисом. Устьица очень немногочисленны, часто погружены в ткань листа или стебля. Днем устьица обычно закрыты, и потеря воды идет в основном через покровные ткани.
Транспирация у суккулентов чрезвычайно мала. Ее трудно уловить за короткий период и приходится определять расход воды не за час, а за сутки или за неделю. Водоудерживающая способность тканей суккулентов значительно выше, чем у других растений экологических групп, благодаря содержанию в клетках гидрофильных веществ. По этой причине и без доступа влаги суккуленты расходуют водный запас очень медленно и долго сохраняют жизнеспособность даже в гербарии.
Ограничения, обусловленные особенностями водного режима суккулентов, создают и другие трудности для жизни этих растений в аридных условиях. Слабая транспирация сводит к минимуму возможность терморегуляции, с чем связано сильное нагревание массивных надземных органов суккулентов. Затруднения создаются и для фотосинтеза, поскольку днем устьица обычно закрыты, а открываются ночью, следовательно, доступ углекислоты и света не совпадают во времени. По этой причине у суккулентов выработался особый путь фотосинтеза, при котором в качестве источника углекислоты, частично используются продукты дыхания. Иными словами, в крайних условиях растения частично используют принцип замкнутой системы с реутилизацией отходов метаболизма.
В силу всех этих ограничений интенсивность фотосинтеза суккулентов невелика, рост и накопление массы идут очень медленно, вследствие чего они не отличаются высокой биологической продуктивностью и не образуют сомкнутых растительных сообществ.
Физиологические показатели водного режима мезофитов подтверждают их промежуточную позицию: для них характерны умеренные величины осмотического давления, содержания воды в листьях, предельного водного дефицита. Что касается транспирации, то ее величина в большей степени зависит от условий освещенности и других элементов микро климата.
Один и тот же мезофильный вид, попадая в разные по водоснабжению условия, обнаруживает известную пластичность, приобретая в сухих условиях более ксероморфные, а во влажных более гигроморфные черты.
Пластичность листьев проявляется не только в разных местообитаниях, но даже у одной и той же особи. К примеру, у деревьев на опушке леса листья на стороне, обращенной в сторону леса, имеют более мезофильный и теневой характер по сравнению с несколько ксероморфными листьями внешней стороны дерева (см. табл. ниже ). Листья разных высотных ярусов одних и тех же растений находятся в неодинаковых условиях водоснабжения, так как поступление воды в верхние части связано с преодолением большого сопротивления. К тому же у деревьев верхние листья обычно находятся в условиях иноВодная среда существенно отличается от воздушной, в связи с этим у водных растений существует ряд своеобразных физиологических адаптивных черт. Интенсивность света в воде сильно ослаблена, поскольку часть падающей радиации отражается от поверхности воды, другая - поглощается ее толщей. В связи с ослаблением света фотосинтез у погруженных растений сильно снижается с увеличением глубины. Считают, сто выживанию глубоководного фитопланктона в зонах, где освещенность ниже точки компенсации, способствуют его периодические вертикальные перемещения в верхние зоны, где идет интенсивный фотосинтез и пополнение запасов органических веществ.
В воде кроме недостатка света растения могут испытывать и другое затруднение, существенное для фотосинтеза, - недостаток доступной СО2 . Углекислота поступает в воду в результате растворения СО2 , содержащегося в воздухе, дыхания водных организмов, разложения органических остатков и высвобождения из карбонатов. При интенсивном фотосинтезе растений идет усиленное потребление СО2 , в связи с чем легко возникает ее дефицит. На увеличение содержания СО2 в воде гидрофиты реагируют заметным повышением фотосинтеза.
У погруженных растений транспирации нет, значит, нет и “верхнего двигателя”, поддерживающего ток воды в растении. При этом данный ток, доставляющий к тканям питательные вещества, существует, при чем с явной суточной периодичностью: днем больше, ночью отсутствует. Активная роль в его поддержании принадлежит корневому давлению и деятельности специальных клеток, выделяющих воду, - водяных устьиц.
Плавающие или торчащие над водой листья обычно имеют сильную транспирацию, хотя и расположены в слое воздуха, который непосредственно граничит с водой и имеет повышенную влажность. Устьица широко открыты и закрываются полностью только в ночное время.
Столь же велика транспирация у прибрежных растений, при чем у них значительное количество воды расходуется не только листьями, но и стеблями.
Осмотическое давление у водных и прибрежных растений очень низкое, так как им не приходится преодолевать водоудерживающую силу почвы при поглощении воды.
Примеры осмотического давления (в кПа) у водных растений и гелофитов (по Гесснеру Ф.,1959)
| Название растения | Давление в корне | Давление в листе | |
| Водные растения | |||
| Hippuris vulgaris | |||
| Nymphaea dentata | |||
| Victoria regia | |||
| Гелофиты | |||
| Polygonum amphibium | -- | ||
| Menyanthes trifoliata | |||
| Lythrum salicaria | -- | ||
| Typha angustifolia | -- | ||
Библиографический список.
1. Ботаника с основами экологии: Учеб. Пособие для студентов пед. ин-тов /Л.В. Кудряшов и др. - М.: Просвещение,1979
2. Горышина Т.К. Экология растений. - М.: Высшая школа,1979
3. Гусев Н.А. Некоторые закономерности водного режима растений. - М.: Изд-во АН СССР, 1959.
4. Двораковский М.С. Экология растений. -
М.: Просвещение 1964
5. Жданов В.С. Аквариумные растения. / Под. ред. Коровина. -
М.: Лесн. пром-ть , 1981
6. Крафтс А. Вода и ее значение в жизни растений. -
М.: Иност. лит., 1951
7. Культиасов И.М. Экология растений. - М.: Изд-во московского ун-та͵ 1982
8. Мак-Дуголл В.Б. Экология растений. - М.: Учпедгиз, 1935.
9. Пономарева И.Н. Экология растений с основами биогеоценологии. - М.: Просвещение, 1978
10. Рычин Ю.В. Флора гигрофитов. Определитель. - М.: Сов. наука, 1948
11. Шенников А.П. Экология растений. - М.: Сов. наука, 1950
го микроклимата.
Водная среда существенно отличается от воздушной, в связи с этим у водных растений существует ряд своеобразных физиологических адаптивных черт. Интенсивность света в воде сильно ослаблена, поскольку часть падающей радиации отражается от поверхности воды, другая – поглощается ее толщей. В связи с ослаблением света фотосинтез у погруженных растений сильно снижается с увеличением глубины. Считают, сто выживанию глубоководного фитопланктона в зонах, где освещенность ниже точки компенсации, способствуют его периодические вертикальные перемещения в верхние зоны, где идет интенсивный фотосинтез и пополнение запасов органических веществ.
В воде кроме недостатка света растения могут испытывать и другое затруднение, существенное для фотосинтеза, - недостаток доступной СО2 . Углекислота поступает в воду в результате растворения СО2 , содержащегося в воздухе, дыхания водных организмов, разложения органических остатков и высвобождения из карбонатов. При интенсивном фотосинтезе растений идет усиленное потребление СО2 , в связи с чем легко возникает ее дефицит. На увеличение содержания СО2 в воде гидрофиты реагируют заметным повышением фотосинтеза.
У погруженных растений транспирации нет, значит, нет и “верхнего двигателя”, поддерживающего ток воды в растении. При этом данный ток, доставляющий к тканям питательные вещества, существует, при чем с явной суточной периодичностью: днем больше, ночью отсутствует. Активная роль в его поддержании принадлежит корневому давлению и деятельности специальных клеток, выделяющих воду, - водяных устьиц.
Плавающие или торчащие над водой листья обычно имеют сильную транспирацию, хотя и расположены в слое воздуха, который непосредственно граничит с водой и имеет повышенную влажность. Устьица широко открыты и закрываются полностью только в ночное время.
Столь же велика транспирация у прибрежных растений, при чем у них значительное количество воды расходуется не только листьями, но и стеблями.
Осмотическое давление у водных и прибрежных растений очень низкое, так как им не приходится преодолевать водоудерживающую силу почвы при поглощении воды.
Примеры осмотического давления (в кПа) у водных растений и гелофитов (по Гесснеру Ф.,1959)
| Название растения | Давление в корне | Давление в листе |
| Водные растения | ||
| Hippuris vulgaris | ||
| Nymphaea dentata | ||
| Victoria regia | ||
| Гелофиты | ||
| Polygonum amphibium | — | |
| Menyanthes trifoliata | ||
| Lythrum salicaria | — | |
| Typha angustifolia | — |
Библиографический список
Ботаника с основами экологии: Учеб. Пособие для студентов пед. ин-тов /Л.В. Кудряшов и др. – М.: Просвещение,1979
Горышина Т.К. Экология растений. – М.: Высшая школа,1979
Гусев Н.А. Некоторые закономерности водного режима растений. – М.: Изд-во АН СССР, 1959.
Двораковский М.С. Экология растений. – М.: Просвещение 1964
Жданов В.С. Аквариумные растения. / Под. ред. Коровина. - М.: Лесн. пром-ть , 1981
Крафтс А. Вода и ее значение в жизни растений. – М.: Иност. лит., 1951
Культиасов И.М. Экология растений. - М.: Изд-во московского ун-та͵ 1982
Мак-Дуголл В.Б. Экология растений. – М.: Учпедгиз, 1935.
Пономарева И.Н. Экология растений с основами биогеоценологии. – М.: Просвещение, 1978
Рычин Ю.В. Флора гигрофитов. Определитель. – М.: Сов. наука, 1948
Шенников А.П. Экология растений. – М.: Сов. наука, 1950
Краткие сведения о классификации
oplib.ru
ГЕЛОФИТЫ (ГИДРОГИГРОФИТЫ) – ВОДНО-БОЛОТНЫЕ
РАСТЕНИЯ:
1. Надводные растения с поднимающимися над поверхностью воды стеблями и
листьями, укореняющиеся (тростник, рогоз, камыш, сусак, ежеголовник,
стрелолист, частуха и др.) Все они успешно развиваются и проходят полный
цикл развития, как в воде, так и на влажных берегах водоемов.
Глава I. Характеристика основных групп растений по отношению к воде.
По приуроченности к местообитаниям с разными условиями увлажнения и выработке соответствующих приспособлений среди наземных растений различают три основных экологических типа: гигрофиты, мезофиты и ксерофиты.
Растения, для которых вода не только необходимый экологический фактор, но непосредственная среда обитания, относятся к водным, называемым гидрофитами.
ГИГРОФИТЫ. Это растения избыточно увлажненных местообитаний с высокой влажностью воздуха и почвы. При довольно большом разнообразии местообитаний, особенностей водного режима и анатомо-морфологических черт всех гигрофитов объединяет отсутствие приспособлений, ограничивающих расход воды, и неспособность выносить даже незначительную ее потерю. Ярко выраженные гигрофиты - травянистые растения и эпифиты влажных тропических лесов, не выносящие сколько-нибудь заметного понижения влажности воздуха. Даже в разгар сезона дождей мелкие эпифитные папоротники на стволах деревьев теряют тургор и засыхают, если на них в течение 2-3 часов падают солнечные лучи. Черты гигрофитов имеют травянистые растения темнохвойных лесов (кислица, майник двулистный, двулепестник альпийский). К гигрофитам можно отнести и виды, растущие на открытых и хорошо освещенных местообитаниях, но в условиях избытка почвенной влаги - близ водоемов, в дельтах рек, в местах выхода грунтовых вод. В наших широтах примером могут служить прибрежные виды: калужница - Caltha palustris, плакун-трава - Lythrum salikaria, а в странах жаркого климата - папирус, болотные пальмы. Из культурных растений сюда можно отнести рис, культивируемый на полях, залитых водой.
КСЕРОФИТЫ. Это растения сухих местообитаний, способные переносить значительный недостаток влаги - почвенную и атмосферную засуху. Они распространены, обильны и разнообразны в областях с жарким и сухим климатом. К этой группе принадлежат виды пустынь, сухих степей, саванн, колючих редколесий, сухих субтропиков. В более гумидных районах ксерофиты участвуют в растительном покрове лишь в наиболее прогреваемых и наименее увлажненных местообитаниях (например, на склонах южной экспозиции).
Неблагоприятный водный режим растений в сухих местообитаниях обусловлен, во-первых, ограниченным поступлением воды при ее недостатке в почве и, во-вторых, увеличением расхода влаги на транспирацию при большой сухости воздуха и высоких температурах. Следовательно, для преодоления недостатка влаги возможны разные пути: увеличение ее поглощения и сокращение расхода, кроме того, способность переносить большие потери воды. Все это используется ксерофитами при адаптации к сухости, но у разных растений в неодинаковой степени, в связи с чем некоторые авторы различают два основных способа преодоления ксерофитами засухи: возможность противостоять иссушению тканей, или активное регулирование водного баланса, и способность выносить сильное иссушение.
В зависимости от структурных черт и способов регулирования водного режима различают несколько разновидностей ксерофитов (по Генкелю П.А.): эуксерофиты, гемиксерофиты, пойкилоксерофиты.
К группе ксерофитов относят и суккуленты - растения с сочными листьями или стеблями. Различают листовые суккуленты (агавы, алоэ) и стеблевые, у которых листья редуцированы, а наземные части представлены мясистыми стеблями(кактусы, некоторые молочаи).
Ксерофиты с наиболее ярко выраженными ксероморфными чертами строения листьев имеют своеобразный внешний облик, за что получили название склерофитов. Облик типичного склерофита легко представить на примере чертополоха - Carduus crispus и пустынных полыней, ковылей, саксаулов.
МЕЗОФИТЫ. Эта группа включает растения, произрастающие в средних условиях увлажнения. Сюда относятся растения лугов, травяного покрова лесов, лиственные древесные и кустарниковые породы из областей умеренно влажного климата, а также большинство культурных растений.
Мезофиты - группа весьма разнообразная не только по видовому составу, но и по различным экологическим оттенкам, обусловленным разным сочетанием факторов в природных местообитаниях. Они связаны переходами с другими экологическими типами растений по отношению к воде, так что четкую границу между ними провести очень трудно. Так, среди луговых мезофитов выделяются виды с повышенным влаголюбием, предпочитающие постоянно сырые или временно заливаемые участки (лисохвост луговой - Alopecurus pratensis, бекмания обыкновенная - Beckmannia eruciformis).
Их объединяют в переходную группу гигромезофитов наряду с некоторыми влаголюбивыми лесными травами, предпочитающими наиболее сырые леса, лесные овраги (недотрога - Impatiens nolitangere). С другой стороны в местообитаниях с переодическим или постоянным (небольшим) недостатком влаги много мезофитов с теми или иными ксероморфными признаками с повышенной физиологической устойчивостью к засухе. Эта группа переходная между мезофитами ксерофитами, - ксеромезофиты. Примером могут служить многие виды северных степей, сухих сосновых боров, песчаных местообитаний: клевер-белоголовка - Trifolium montanum, подмаренник желтый - Galium verum и другие.
Особое место среди мезофитов занимают степные и пустынные весенние эфемеры и эфемероиды. К этой группе принадлежат растения, ранней весной покрывающие степи и пустыни разноцветным цветущим ковром (многолетники - тюльпаны, гусиные луки; однолетники - маки, вероники). Это виды с чрезвычайно краткой вегетацией и длительным периодом покоя, который однолетние эфемеры переживают в виде семян, а многолетние эфемероиды - в виде покоящихся луковиц, клубней, корневищ. Кроме весенних существуют и осенние эфемероиды, произрастающие в районах с климатическим ритмом средиземноморского типа. Сюда относятся виды родов Crocus, Scilla и другие.
По многим особенностям структуры и физиологии близки к ксерофитам растения, которые по тем или иным причинам испытывают недостаток влаги, сопряженный с действием низких температур. Иногда такие виды в качестве особого подразделения включают в группу ксерофитов, иногда выделяют в самостоятельные экологические типы - психрофиты и криофиты.
Психрофиты - растения влажных и холодных почв в холодных местообитаниях высокогорий и северных широт. Несмотря на достаточное увлажнение почвы, они часто испытывают недостаток влаги (или из-за физиологической сухости, вызванной низкими температурами, или в связи с преобладанием в почве недоступной влаги, как, например, на торфянистых почвах). Среди психрофитов есть травянистые растения (например злаки северных лугов: белоус - Nardus strikta; высокогорные кавказские злаки: овсяница пестрая -Festuka varia), высокогорные, болотные и тундровые кустарники и кустарнички, как вечнозеленые (вереск - Calluna vulgaris), так и с опадающей листвой (карликовые ивы - Salix polaris, S. herbacea).К психрофитам относятся и хвойные древесные породы умеренных и северных широт.
Криофиты в экологическом отношении очень близки к психрофитам и связаны с ними переходными формами. Это растения сухих и холодных местообитаний - сухих участков тундр, скал, осыпей. Обычно они рассматриваются и характеризуются вместе с психрофитами, поскольку у них много сходных морфологических и физиологических черт. Но среди криофитов есть и весьма своеобразные формы - это растения-подушки высокогорных холодных пустынь.
ГИДРОФИТЫ. Это водные растения. По образу жизни и строению среди них можно выделить погруженные растения и растения с плавающими листьями. Погруженные растения подразделяют на укореняющиеся в донном грунте и взвешенные в толще воды. Из высших растений к первым принадлежат телорез - Stratiotes aloides, шильник водяной - Subularia aquatika. В эту же группу входят водоросли, прикрепленные к грунту. Из растений, взвешенных в толще воды, можно назвать роголистник погруженный--Ceratophyllum demersum, пузырчатку обыкновенную - Utrikularia vulgaris, а также многочисленные виды планктонных водорослей.
Растения с плавающими листьями используют частично водную, частично воздушную среду. Из них укореняются в грунте кувшинки из рода Nymphaea, кубышки из рода Nuphar, рдесты, орех водяной - Trapa natans.
Многие виды наряду с плавающими на поверхности воды листьями имеют и подводные. Плавают на поверхности воды, не укореняясь, ряски, водокрас.
К настоящим водным растениям очень близко примыкает и обычно вместе с ними рассматривается группа гелофитов или амфибий - земноводных растений. Это виды береговых и прибрежных местообитаний с избыточным или переменным увлажнением. Они могут расти как в воздушной среде, так и частично погруженными в воду, могут выносить и полное временное заливание. Как в природе нет резкой границы между водными и наземными местообитаниями для растений, так и группа гелофитов связана незаметными переходами, с одной стороны, с настоящими гидрофитами, с другой - с наземными гигрофитами и гигромезофитами. Примеры гелофитов - растений прибрежной полосы пресноводных водоемов и рек: стрелолист - Sagittaria sagittifolia, ежеголовка - Sparganium ramosum.
ГЛАВА II. Анатомо-морфологические приспособления растений к водному режиму.
Растения, приуроченные к местообитаниям с разными условиями увлажнения, выработали соответствующие приспособления к водному режиму.
Характерные структурные черты гигрофитов - тонкие нежные листовые пластинки с небольшим числом устьиц, не имеющие толстой кутикулы, рыхлое сложение тканей листа с крупными межклетниками, слабое развитие водопроводящей ткани, тонкие слаборазветвленные корни.
Для ксерофитов большое значение имеют разнообразные структурные приспособления к условиям недостатка влаги.
Корневые системы обычно сильно развиты, что помогает растениям увеличить поглощение почвенной влаги. По общей массе корневые системы ксерофитов нередко превышают надземные части, иногда весьма значительно. Так, у многих травянистых и кустарниковых видов среднеазиатских пустынь подземная масса больше надземной в 9-10 раз, а у кесрофитов памирских высокогорных холодных пустынь - в300-400 раз. Корневые системы ксерофитов часто бывают экстенсивного типа, то есть растения имеют длинные корни, распространяющиеся в большом объеме почвы, но сравнительно мало разветвленные. Проникновение таких корней на большую глубину позволяет ксерофитам использовать влагу глубоких почвенных горизонтов, а в отдельных случаях - и грунтовых вод.
У других видов корневые системы интенсивного типа: они охватывают относительно небольшой объем почвы, но благодаря очень густому ветвлению максимально используют почвенную влагу. Корни ряда ксерофильных видов имеют специальные приспособления для запасания влаги. Надземные органы ксерофитов также отличаются своеобразными (так называемыми ксероморфными чертами), которые носят отпечаток трудных условий водоснабжения. У них сильно развита водопроводящая система, что хорошо заметно по густоте сети жилок в листьях, подводящих воду к тканям. Эта черта облегчает ксерофитам пополнение запасов влаги, расходуемой на транспирацию.
Разнообразные структурные приспособления защитного характера, направленные на уменьшение расхода воды, в основном сводятся к следующему:
1.Общее сокращение транспирирующей поверхности. Многие ксерофиты имеют мелкие, узкие, сильно редуцированные листовые пластинки. В особо засушливых пустынных местообитаниях листья некоторых древесных и кустарниковых пород редуцированы до едва заметных чешуек. У таких видов фотосинтез осуществляют зеленые ветви.
2.Уменьшение листовой поверхности в наиболее жаркие и сухие периоды вегетационного сезона. Для многих кустарников среднеазиатских, североафриканских и других пустынь, а так же для некоторых видов сухих субтропиков средиземноморья характерен сезонный деформизм листьев: ранней весной при еще благоприятном водном режиме образуются относительно крупные листья, которые летом, при наступлении жары и сухости, сменяются мелкими листьями более ксероморфного строения с меньшей интенсивностью транспирации.
3.Защита листьев от больших потерь влаги на транспирацию. Она достигается благодаря развитию мощных покровных тканей - толстостенного, иногда многослойного эпидермиса, часто несущего различные выросты и волоски, которые образуют густое “войлочное” опушение поверхности листа. У других видов поверхность покрыта водонепроницаемым слоем толстой кутикулы или воскового налета. Развитие защитных покровов на листьях причина того, что степной травостой имеет тусклые, седоватые оттенки, резко отличающиеся от яркой зелени лугов.
Устьица у ксерофитов обычно защищены от чрезмерной потери влаги, например, расположены в специальных углублениях в ткани листа, иногда снабженных волосками и прочими дополнительными защитными устройствами. У ковылей и других степных злаков существует интересный механизм защиты устьиц в самые жаркие и сухие часы дня: при больших потерях воды крупные тонкостенные водоносные клетки эпидермиса теряют тургор, и лист свертывается в трубку; так устьица оказываются изолированными от окружающего сухого воздуха внутри замкнутой полости, где благодаря транспирации создается повышенная влажность. Во влажную погоду клетки эпидермиса восстанавливают тургор, и листовая пластинка вновь развертывается.
4.Усиленное развитие механической ткани.
Клетки тканей листьев у ксерофитов отличаются мелкими размерами и весьма плотной упаковкой, то есть малым развитием межклетников, благодаря чему сильно сокращается внутренняя испаряющая поверхность листа. Поскольку ксерофиты обычно обитают на открытых, хорошо освещенных местообитаниях, многие черты ксероморфной структуры листа - это одновременно и черты световой структуры. Так у многих видов листья имеют мощную иногда многорядную палисадную паренхиму, часто расположенную с обеих сторон.
Основные морфолого-анотомические черты мезофитов - средние между чертами гигрофитов и ксерофитов. Мезофиты имеют умеренно развитые корневые системы как экстенсивного, так и интенсивного типа, со всеми переходами между ними. Для листа характерна дифференцировка тканей на более или менее плотную палисадную паренхиму и рыхлую губчатую паренхиму с системой межклетников. Сеть жилок сравнительно негустая. Покровные ткани могут иметь отдельные ксероморфные черты, но не столь ярко выраженные, как у ксерофитов.
Психрофиты имеют ярко выраженную ксероморфную структуру листа. Так, психрофильные злаки узколистны, имеют хорошо развитую проводящую и механическую ткани; некоторые из них способны к свертыванию листовой пластинки в трубку, напоминая степные ксерофиты. Вечнозеленые кустарнички имеют плотные кожистые листья, иногда с весьма мощной кутикулой, плотной палисадной паренхимой. Нижняя сторона, несущая устьица, часто опушена или покрыта восковым налетом. Вместе с тем в структуре листа ясно заметны и некоторые гигроморфные черты, главным образом, крупные размеры клеток и развитие больших межклетников в губчатой ткани.
Анатомо-морфологические черты гидрофитов существенно отличают их от наземных растений. Заметна редукция механических тканей. Столь же сильно редуцированна и проводящая система. Если у сухопутных мезофитов длина жилок на 1 кв.см. листа составляет около100 мм и более, а у ксерофитов доходит до300, то у водных и прибрежных растений она в несколько раз меньше. Вот несколько примеров (по Гесснеру Ф., 1959):
| Вид | Плавающие листья, мм | Подводные листья, мм | |
| Nelumbium nuciferum | -- | ||
| Victoria cruciana | -- | ||
| Victoria regia | -- | ||
| Potamogeton praelongus | -- | ||
У некоторых погруженных растений, не прикрепленных к грунту, корни полностью редуцированны, другие корни сохранили, но отдельно плавающие части растений могут обходиться и без них. Корни укрепляющихся гидрофитов слабоветвистые, без корневых волосков. Вместе с тем ряд видов имеет толстые и прочные корневища, которые играют роль якоря, хранилища запасных веществ и органа вегетативного размножения.
Листья погруженных гидрофитов очень тонки и нежны, имеют упрощенное строение мезофилла без заметной дифференциации на палисадную и губчатую паренхиму. Подводные листья без устьиц. В отдельных местах находятся группы клеток эпидермиса с утонченными стенками. Считается, что они играют большую роль в поглощении воды и растворенных минеральных солей.
У растений лишь частично погруженных в воду, хорошо выражена гетерофиллия - различие строения надводных и подводных листьев на одной и той же особи. Первые имеют черты, обычные для листьев наземных растений, вторые - очень тонкие или рассеченные листовые пластинки. Гетерофиллия отмечена у водного лютика - Ranunculus diversifolius, кувшинок и кубышек, стрелолиста и других видов. Интересный пример - поручейник, на стебле которого можно видеть несколько форм листьев, представляющих все переходы от типично наземных до водных.
Глава III.Физиологические адаптации растений, приуроченных к местообитаниям разной увлажненности.
Наряду с морфологическими особенностями у растений, приуроченных к местам с разными условиями увлажненности, выработались и физиологические.
Способность гигрофитов к регуляции водного режима ограничена: устьица большей частью широко открыты, так что транспирация мало отличается от физического испарения. Благодаря беспрепятственному потоку воды и отсутствию защитных приспособлений интенсивность транспирации очень высока: у световых гигрофитов в дневное время листья могут терять за час количество воды, в 4-5 раз превышающее массу листа. Высокая оводненность тканей гигрофитов поддерживается в основном за счет постоянного притока влаги из окружающей среды.
Другие характерные физиологические черты гигрофитов, обусловленные легкой доступностью влаги, - низкое осмотическое давление клеточного сока, незначительная водоудерживающая способность, приводящая к быстрой потере запасов воды.особенно показательны для гигрофитов небольшие величины сублетального водного дефицита: так, для кислицы и майника потеря 15%-20% запаса воды уже необратима и ведет к гибели. В некоторых случаях у растений сильно увлажненных местообитаний возникает необходимость удаления избытка влаги. Обычно, это бывает, когда почва хорошо прогрета и корни активно всасывают воду, а транспирация отсутствует (например, утром или при тумане, когда влажность воздуха 100%). Избыточная влага удаляется путем гуттации - выделение воды через специальные выделительные клетки, расположенные по краю или на острие листа.
Ксерофиты обладают рядом разнообразных физиологических адаптаций, позволяющих им успешно выдерживать недостаток влаги.
У ксерофитов обычно повышено осмотическое давление клеточного сока, позволяющее всасывать воду даже при больших водоотнимающих силах почвы, то есть использовать не только легкодоступную, но и труднодоступную почвенную влагу. Оно измеряется тысячами кПа, а у некоторых пустынных кустарников зарегистрированы цифры, достигающие 10000-30000 кПа.
С давних пор пристальное внимание привлекала проблема расхода воды ксерофитами на транспирацию. Казалось бы, многочисленные анатомические приспособления, достаточно надежно защищающие наземные части ксерофитов от сильного испарения, должны способствовать значительному снижению транспирации. Однако выяснилось, что в действительности это не так. При достаточном водоснабжении большинство ксерофитов имеют довольно высокую транспирацию, но при наступлении засушливых условий, они сильно сокращают ее. При этом играет роль и закрывание устьиц, и сильное обезвоживание листа при начинающемся подвядании. Несомненно, анатомо-морфологические приспособления имеют определенное значение, но основную роль в засухоустойчивости ксерофитов в настоящее время отводят физиологическим механизмам.
К числу этих механизмов принадлежит высокая водоудерживающая способность тканей и клеток, обусловленная рядом физиологических и биохимических особенностей.
Большое значение для выживания ксерофитов при резком недостатке влаги имеет их способность переносить глубокое обезвоживание тканей без потери жизнеспособности и способности восстановления нормального содержания воды в растении при возобновлении благоприятных условий. Ксерофиты способны потерять до 75% всего водного запаса и, тем не менее, остаться живыми. Ярким примером в этом отношении служат пустынные растения, которые летом высыхают до состояния, близкого к воздушно-сухому, и впадают в анабиоз, но после дождей возобновляют рост и развитие.
Еще одна система адаптаций, обеспечивающих выживание ксерофитов в аридных условиях, - выработка сезонных ритмов, дающих возможность растениям использовать для вегетации наиболее благоприятные периоды года и резко сократить жизнедеятельность во время засухи. Так, в областях со средиземноморским климатом с резко выраженным летним сухим периодом многие ксерофильные виды имеют “двухтактный” ритм сезонного развития: весенняя вегетация сменяется летним покоем, во время которого растения сбрасывают листву и снижают интенсивность физиологических процессов; в период осенних дождей вегетация возобновляется, и затем уже следует зимний покой. Сходное явление наблюдается и у растений сухих степей в середине и конце лета: потеря части листовой поверхности, приостановка развития, сильное обезвоживание тканей и т.д. Такое состояние, получившее название полупокоя, длится вплоть до осенних дождей, после которых у степных ксерофитов начинают отрастать листья.
Физиологические адаптации суккулентов столь своеобразны, что их необходимо рассмотреть отдельно.
Основной способ преодоления засушливых условий у суккулентов - накопление больших запасов воды в тканях и крайне экономное ее расходование. В условиях жаркого и сухого климата весь водный запас мог бы быть быстро растрачен, но растения имеют защитные приспособления, направленные к сокращению транспирации. Одно из них - своеобразная форма надземных частей суккулентов. В дополнение к этому у многих суккулентов поверхность защищена восковым налетом опушением, хотя есть и суккуленты с тонким не защищенным эпидермисом. Устьица очень немногочисленны, часто погружены в ткань листа или стебля. Днем устьица обычно закрыты, и потеря воды идет в основном через покровные ткани.
Транспирация у суккулентов чрезвычайно мала. Ее трудно уловить за короткий период и приходится определять расход воды не за час, а за сутки или за неделю. Водоудерживающая способность тканей суккулентов значительно выше, чем у других растений экологических групп, благодаря содержанию в клетках гидрофильных веществ. Поэтому и без доступа влаги суккуленты расходуют водный запас очень медленно и долго сохраняют жизнеспособность даже в гербарии.
Ограничения, обусловленные особенностями водного режима суккулентов, создают и другие трудности для жизни этих растений в аридных условиях. Слабая транспирация сводит к минимуму возможность терморегуляции, с чем связано сильное нагревание массивных надземных органов суккулентов. Затруднения создаются и для фотосинтеза, поскольку днем устьица обычно закрыты, а открываются ночью, следовательно, доступ углекислоты и света не совпадают во времени. Поэтому у суккулентов выработался особый путь фотосинтеза, при котором в качестве источника углекислоты, частично используются продукты дыхания. Иными словами, в крайних условиях растения частично используют принцип замкнутой системы с реутилизацией отходов метаболизма.
В силу всех этих ограничений интенсивность фотосинтеза суккулентов невелика, рост и накопление массы идут очень медленно, вследствие чего они не отличаются высокой биологической продуктивностью и не образуют сомкнутых растительных сообществ.
Физиологические показатели водного режима мезофитов подтверждают их промежуточную позицию: для них характерны умеренные величины осмотического давления, содержания воды в листьях, предельного водного дефицита. Что касается транспирации, то ее величина в большей степени зависит от условий освещенности и других элементов микро климата.
Один и тот же мезофильный вид, попадая в разные по водоснабжению условия, обнаруживает известную пластичность, приобретая в сухих условиях более ксероморфные, а во влажных более гигроморфные черты.
Пластичность листьев проявляется не только в разных местообитаниях, но даже у одной и той же особи. Например, у деревьев на опушке леса листья на стороне, обращенной в сторону леса, имеют более мезофильный и теневой характер по сравнению с несколько ксероморфными листьями внешней стороны дерева (см. табл. ниже ). Листья разных высотных ярусов одних и тех же растений находятся в неодинаковых условиях водоснабжения, так как поступление воды в верхние части связано с преодолением большого сопротивления. К тому же у деревьев верхние листья обычно находятся в условиях иноВодная среда существенно отличается от воздушной, поэтому у водных растений существует ряд своеобразных физиологических адаптивных черт. Интенсивность света в воде сильно ослаблена, поскольку часть падающей радиации отражается от поверхности воды, другая - поглощается ее толщей. В связи с ослаблением света фотосинтез у погруженных растений сильно снижается с увеличением глубины. Считают, сто выживанию глубоководного фитопланктона в зонах, где освещенность ниже точки компенсации, способствуют его периодические вертикальные перемещения в верхние зоны, где идет интенсивный фотосинтез и пополнение запасов органических веществ.
В воде кроме недостатка света растения могут испытывать и другое затруднение, существенное для фотосинтеза, - недостаток доступной СО2 . Углекислота поступает в воду в результате растворения СО2 , содержащегося в воздухе, дыхания водных организмов, разложения органических остатков и высвобождения из карбонатов. При интенсивном фотосинтезе растений идет усиленное потребление СО2 , в связи с чем легко возникает ее дефицит.На увеличение содержания СО2 в воде гидрофиты реагируют заметным повышением фотосинтеза.
У погруженных растений транспирации нет, значит, нет и “верхнего двигателя”, поддерживающего ток воды в растении. Однако этот ток, доставляющий к тканям питательные вещества, существует, при чем с явной суточной периодичностью: днем больше, ночью отсутствует. Активная роль в его поддержании принадлежит корневому давлению и деятельности специальных клеток, выделяющих воду, - водяных устьиц.
Плавающие или торчащие над водой листья обычно имеют сильную транспирацию, хотя и расположены в слое воздуха, который непосредственно граничит с водой и имеет повышенную влажность. Устьица широко открыты и закрываются полностью только в ночное время.
Столь же велика транспирация у прибрежных растений, при чем у них значительное количество воды расходуется не только листьями, но и стеблями.
Осмотическое давление у водных и прибрежных растений очень низкое, так как им не приходится преодолевать водоудерживающую силу почвы при поглощении воды.
Примеры осмотического давления (в кПа) у водных растений и гелофитов (по Гесснеру Ф.,1959)
| Название растения | Давление в корне | Давление в листе | |
| Водные растения | |||
| Hippuris vulgaris | |||
| Nymphaea dentata | |||
| Victoria regia | |||
| Гелофиты | |||
| Polygonum amphibium | -- | ||
| Menyanthes trifoliata | |||
| Lythrum salicaria | -- | ||
| Typha angustifolia | -- | ||
Список литературы.
1. Ботаника с основами экологии: Учеб. Пособие для студентов пед. ин-тов /Л.В. Кудряшов и др. - М.: Просвещение,1979
2. Горышина Т.К. Экология растений. - М.: Высшая школа,1979
3. Гусев Н.А. Некоторые закономерности водного режима растений. - М.: Изд-во АН СССР, 1959.
4. Двораковский М.С. Экология растений. -
М.: Просвещение 1964
5. Жданов В.С. Аквариумные растения. / Под. ред. Коровина. -
М.: Лесн. пром-ть , 1981
6. Крафтс А. Вода и ее значение в жизни растений. -
М.: Иност. лит., 1951
7. Культиасов И.М. Экология растений. - М.: Изд-во московского ун-та, 1982
8. Мак-Дуголл В.Б. Экология растений. - М.: Учпедгиз, 1935.
9. Пономарева И.Н. Экология растений с основами биогеоценологии. - М.: Просвещение, 1978
10. Рычин Ю.В. Флора гигрофитов. Определитель. - М.: Сов. наука, 1948
11. Шенников А.П. Экология растений. - М.: Сов. наука, 1950
го микроклимата.
Водная среда существенно отличается от воздушной, поэтому у водных растений существует ряд своеобразных физиологических адаптивных черт. Интенсивность света в воде сильно ослаблена, поскольку часть падающей радиации отражается от поверхности воды, другая – поглощается ее толщей. В связи с ослаблением света фотосинтез у погруженных растений сильно снижается с увеличением глубины. Считают, сто выживанию глубоководного фитопланктона в зонах, где освещенность ниже точки компенсации, способствуют его периодические вертикальные перемещения в верхние зоны, где идет интенсивный фотосинтез и пополнение запасов органических веществ.
В воде кроме недостатка света растения могут испытывать и другое затруднение, существенное для фотосинтеза, - недостаток доступной СО2 . Углекислота поступает в воду в результате растворения СО2 , содержащегося в воздухе, дыхания водных организмов, разложения органических остатков и высвобождения из карбонатов. При интенсивном фотосинтезе растений идет усиленное потребление СО2 , в связи с чем легко возникает ее дефицит.На увеличение содержания СО2 в воде гидрофиты реагируют заметным повышением фотосинтеза.
У погруженных растений транспирации нет, значит, нет и “верхнего двигателя”, поддерживающего ток воды в растении. Однако этот ток, доставляющий к тканям питательные вещества, существует, при чем с явной суточной периодичностью: днем больше, ночью отсутствует. Активная роль в его поддержании принадлежит корневому давлению и деятельности специальных клеток, выделяющих воду, - водяных устьиц.
Плавающие или торчащие над водой листья обычно имеют сильную транспирацию, хотя и расположены в слое воздуха, который непосредственно граничит с водой и имеет повышенную влажность. Устьица широко открыты и закрываются полностью только в ночное время.
Столь же велика транспирация у прибрежных растений, при чем у них значительное количество воды расходуется не только листьями, но и стеблями.
Осмотическое давление у водных и прибрежных растений очень низкое, так как им не приходится преодолевать водоудерживающую силу почвы при поглощении воды.
Примеры осмотического давления (в кПа) у водных растений и гелофитов (по Гесснеру Ф.,1959)
| Название растения | Давление в корне | Давление в листе |
| Водные растения | ||
| Hippuris vulgaris | ||
| Nymphaea dentata | ||
| Victoria regia | ||
| Гелофиты | ||
| Polygonum amphibium | — | |
| Menyanthes trifoliata | ||
| Lythrum salicaria | — | |
| Typha angustifolia | — |
Список литературы
Ботаника с основами экологии: Учеб. Пособие для студентов пед. ин-тов /Л.В. Кудряшов и др. – М.: Просвещение,1979
Горышина Т.К. Экология растений. – М.: Высшая школа,1979
Гусев Н.А. Некоторые закономерности водного режима растений. – М.: Изд-во АН СССР, 1959.
Двораковский М.С. Экология растений. – М.: Просвещение 1964
Жданов В.С. Аквариумные растения. / Под. ред. Коровина. - М.: Лесн. пром-ть , 1981
Крафтс А. Вода и ее значение в жизни растений. – М.: Иност. лит., 1951
Культиасов И.М. Экология растений. - М.: Изд-во московского ун-та, 1982
Мак-Дуголл В.Б. Экология растений. – М.: Учпедгиз, 1935.
Пономарева И.Н. Экология растений с основами биогеоценологии. – М.: Просвещение, 1978
Рычин Ю.В. Флора гигрофитов. Определитель. – М.: Сов. наука, 1948
Шенников А.П. Экология растений. – М.: Сов. наука, 1950
Краткие сведения о классификации
5rik.ru
Гелофиты - Википедия
Материал из Википедии — свободной энциклопедии
относится к гигрофитам Гигрофи́ты (от др.-греч. ὑγρός — влажный + φυτόν — растение) — растения, обитающие в местах с высокой влажностью воздуха и (или) почвы. Категория гигрофитов выделяется на основе особенностей физиологии и морфологии растений; наиболее характерный признак гигрофитов — отсутствие приспособлений, ограничивающих расходование влаги.
Гигрофиты, как правило, отличаются большими и тонкими листьями с незначительным поверхностным защитным слоем (кутикулой), что приводит к высокой кутикулярной транспирации; незначительным утолщением эпидермиса, слаборазвитой сеткой жилок; устьица у листьев крупные, но их удельное количество на единицу площади невелико. Часто листья на поверхности имеют волоски из живых клеток эпидермиса для увеличения площади листа. За счёт крупных устьиц растения достигают оптимального регулирования транспирации. Морфологической особенностью листьев гигрофитов является наличие специфических устьиц — гидатод, которые способны выделять влагу из листа в капельно-жидкой форме. Это необходимо растению для нормальной жизнедеятельности в условиях атмосферы, перенасыщенной водяными испарениями. Выделение капель воды из листа называется гуттацией.
Среди других морфологических особенностей гигрофитов — слаборазвитая корневая система, наличие длинных стеблей при неразвитости механических тканей, слабое одревеснение стенок сосудов. Подобные особенности приводят к увяданию даже при незначительной нехватке влаги.
Наиболее характерными гигрофитами являются травянистые растения влажных тропических лесов.
Болотные травянистые растения обычно также относят к гигрофитам, однако по ряду морфологических признаков они имеют отличия. В силу постоянного воздействия солнечного света и более сухой атмосферы у них наличествует более толстая кутикула, что приводит к уменьшению кутикулярной транспирации; а листья обычно толще и меньше. Иногда болотные растения выделяют в особую подгруппу — гелофиты (др.-греч. χέλος — болото).
К гигрофитам близки гидрофиты (водные растения, погружённые в воду нижней частью) и гидатофиты (водные растения, полностью или бо́льшей частью находящиеся в воде). Иногда в силу схожести морфологии и условий жизнеобитания все эти группы рассматривают как одну.
См. также[ | ]
Ссылки[ | ]
encyclopaedia.bid









