
Что такое фотосинтез или почему трава зелёная? Фотосинтез растений это
Фотосинтез — WiKi
Классификация
У живых организмов обнаружено два типа пигментов, способных выполнять функцию фотосинтетических антенн. Эти пигменты поглощают кванты видимого света и обеспечивают дальнейшее запасание энергии излучения в виде энергии электрохимического градиента H+ на биологических мембранах. У подавляющего большинства организмов роль антенн играют хлорофиллы; менее распространен случай, при котором в качестве антенны служит производное витамина А, ретиналь. В соответствии с этим выделяют хлорофилльный и бесхлорофилльный фотосинтез.
Бесхлорофилльный фотосинтез
Система бесхлорофилльного фотосинтеза отличается значительной простотой организации, в связи с чем предполагается эволюционно первичным механизмом запасания энергии электромагнитного излучения. Эффективность бесхлорофилльного фотосинтеза как механизма преобразования энергии сравнительно низка (на один поглощённый квант переносится лишь один H+).
Открытие у галофильных архей
Dieter Oesterhelt и Walther Stoeckenius идентифицировали в «пурпурных мембранах» представителя галофильных архей Halobacterium salinarium (прежнее название Н. halobium) белок, который позже был назван бактериородопсином. В скором времени были накоплены факты, указывающие на то, что бактериородопсин является светозависимым генератором протонного градиента. В частности, было продемонстрировано фотофосфорилирование на искусственных везикулах, содержащих бактериородопсин и митохондриальную АТФ-синтазу, фотофосфорилирование в интактных клетках H. salinarium, светоиндуцируемое падение pH среды и подавление дыхания, причем все эти эффекты коррелировали со спектром поглощения бактериородопсина. Таким образом, были получены неопровержимые доказательства существования бесхлорофилльного фотосинтеза.
 Механизм бесхлорофилльного фотосинтеза галобактерий
Механизм бесхлорофилльного фотосинтеза галобактерий Механизм
Фотосинтетический аппарат экстремальных галобактерий является наиболее примитивным из ныне известных; в нём отсутствует электронтранспортная цепь. Цитоплазматическая мембрана галобактерий является сопрягающей мембраной, содержащей два основных компонента: светозависимую протонную помпу (бактериородопсин) и АТФ-синтазу. Работа такого фотосинтетического аппарата основана на следующих трансформациях энергии:
- Хромофор бактериородопсина ретиналь поглощает кванты света, что приводит к конформационным изменениям в структуре бактериородопсина и транспорту протона из цитоплазмы в периплазматическое пространство. Кроме того, дополнительный вклад в электрическую составляющую градиента вносит активный светозависимый импорт хлорид-аниона, который обеспечивает галородопсин[источник не указан 1349 дней]. Таким образом, в результате работы бактериородопсина энергия солнечного излучения трансформируется в энергию электрохимического градиента протонов на мембране.
- При работе АТФ-синтазы энергия трансмембранного градиента трансформируется в энергию химических связей АТФ. Таким образом, осуществляется хемиосмотическое сопряжение.
При бесхлорофилльном типе фотосинтеза (как и при реализации циклических потоков в электрон-транспортных цепях) не происходит образования восстановительных эквивалентов (восстановленного ферредоксина или НАД(Ф)Н), необходимых для ассимиляции углекислого газа. Поэтому при бесхлорофилльном фотосинтезе не происходит ассимиляции углекислого газа, а осуществляется исключительно запасание солнечной энергии в форме АТФ (фотофосфорилирование).
Значение
Основной путь получения энергии для галобактерий — аэробное окисление органических соединений (при культивировании используют углеводы и аминокислоты). При дефиците кислорода помимо бесхлорофилльного фотосинтеза источниками энергии для галобактерий может служить анаэробное нитратное дыхание или сбраживание аргинина и цитруллина. Однако в эксперименте было показано, что бесхлорофилльный фотосинтез может служить и единственным источником энергии в анаэробных условиях при подавлении анаэробного дыхания и брожения при обязательном условии, что в среду вносят ретиналь, для синтеза которого необходим кислород.
Хлорофилльный фотосинтез
Хлорофилльный фотосинтез отличается от бактериородопсинового значительно большей эффективностью запасания энергии. На каждый поглощённый квант излучения против градиента переносится не менее одного H+, и в некоторых случаях энергия запасается в форме восстановленных соединений (ферредоксин, НАДФ).
Аноксигенный
Аноксигенный (или бескислородный) фотосинтез протекает без выделения кислорода. К аноксигенному фотосинтезу способны пурпурные и зелёные бактерии, а также гелиобактерии.
При аноксигенном фотосинтезе возможно осуществление:
- Светозависимого циклического транспорта электронов, не сопровождающегося образованием восстановительных эквивалентов и приводящего исключительно к запасанию энергии света в форме АТФ. При циклическом светозависимом электронном транспорте необходимости в экзогенных донорах электронов не возникает. Потребность в восстановительных эквивалентах обеспечивается нефотохимическим путём, как правило, за счёт экзогенных органических соединений.
- Светозависимого нециклического транспорта электронов, сопровождающегося и образованием восстановительных эквивалентов, и синтезом АДФ. При этом возникает потребность в экзогенных донорах электронов, которые необходимы для заполнения электронной вакансии в реакционном центре. В качестве экзогенных доноров электронов могут использоваться как органические, так и неорганические восстановители. Среди неорганических соединений наиболее часто используются различные восстановленные формы серы (сероводород, молекулярная сера, сульфиты, тиосульфаты, тетратионаты, тиогликоляты), также возможно использование молекулярного водорода.
Оксигенный
Оксигенный (или кислородный) фотосинтез сопровождается выделением кислорода в качестве побочного продукта. При оксигенном фотосинтезе осуществляется нециклический электронный транспорт, хотя при определенных физиологических условиях осуществляется исключительно циклический электронный транспорт. В качестве донора электронов при нециклическом потоке используется крайне слабый донор электронов — вода.
Оксигенный фотосинтез распространён гораздо шире. Характерен для высших растений, водорослей, многих протистов и цианобактерий.
Этапы
Фотосинтез — процесс с крайне сложной пространственно-временной организацией.
Разброс характерных времен различных этапов фотосинтеза составляет 19 порядков: скорость процессов поглощения квантов света и миграции энергии измеряется в фемтосекундном интервале (10−15 с), скорость электронного транспорта имеет характерные времена 10−10−10−2 с, а процессы, связанные с ростом растений, измеряются днями (105−107 с).
Также большой разброс размеров характерен для структур, обеспечивающих протекание фотосинтеза: от молекулярного уровня (10−27 м3) до уровня фитоценозов (105 м3).
В фотосинтезе можно выделить отдельные этапы, различающиеся по природе и характерным скоростям процессов:
- Фотофизический;
- Фотохимический;
- Химический:
- Реакции транспорта электронов;
- «Темновые» реакции или циклы углерода при фотосинтезе.
На первом этапе происходит поглощение квантов света пигментами, их переход в возбуждённое состояние и передача энергии к другим молекулам фотосистемы (пластохинон).[1].
На втором этапе происходит разделение зарядов в реакционном центре. Молекула воды теряет электрон под воздействием катиона-радикала, образовавшегося из молекулы хлорофилла после потери ей своего электрона и передачи его пластохинону на первом этапе: h3O−e−⟶H++O˙H{\displaystyle {\ce {h3O-e^{-}->H^{+}{+}{\dot {O}}H}}} . Затем образовавшиеся гидроксильные радикалы под воздействием положительно заряженных ионов марганца преобразуются в кислород и воду: 4O˙H⟶O2+2h3O{\displaystyle {\ce {4{\dot {O}}H->O2{+}2h3O}}} [1]. Одновременно с этим процессом происходит перенос электронов по фотосинтетической электронотранспортной цепи, что заканчивается синтезом АТФ и НАДФН[1]. Первые два этапа вместе называют светозависимой стадией фотосинтеза.
 . Затем образовавшиеся гидроксильные радикалы под воздействием положительно заряженных ионов марганца преобразуются в кислород и воду: 4O˙H⟶O2+2h3O{\displaystyle {\ce {4{\dot {O}}H->O2{+}2h3O}}}
. Затем образовавшиеся гидроксильные радикалы под воздействием положительно заряженных ионов марганца преобразуются в кислород и воду: 4O˙H⟶O2+2h3O{\displaystyle {\ce {4{\dot {O}}H->O2{+}2h3O}}} [1]. Одновременно с этим процессом происходит перенос электронов по фотосинтетической электронотранспортной цепи, что заканчивается синтезом АТФ и НАДФН[1]. Первые два этапа вместе называют светозависимой стадией фотосинтеза.
[1]. Одновременно с этим процессом происходит перенос электронов по фотосинтетической электронотранспортной цепи, что заканчивается синтезом АТФ и НАДФН[1]. Первые два этапа вместе называют светозависимой стадией фотосинтеза. Третий этап заключается в поглощении второй молекулой хлорофилла кванта света и передаче ей электрона ферредоксину. Затем хлорофилл получает электрон после цепи его перемещений на первом и втором этапах. Ферредоксин восстанавливает универсальный восстановитель НАДФ.[1].
Четвёртый этап происходит уже без обязательного участия света и включает в себя биохимические реакции синтеза органических веществ с использованием энергии, накопленной на светозависимой стадии. Чаще всего в качестве таких реакций рассматривается цикл Кальвина и глюконеогенез, образование сахаров и крахмала из углекислого газа воздуха[1].
Пространственная локализация
Лист
Фотосинтез растений осуществляется в хлоропластах — полуавтономных двухмембранных органеллах, относящихся к классу пластид. Хлоропласты могут содержаться в клетках стеблей, плодов, чашелистиков, однако основным органом фотосинтеза является лист. Он анатомически приспособлен к поглощению энергии света и ассимиляции углекислоты. Плоская форма листа, обеспечивающая большое отношение поверхности к объёму, позволяет более полно использовать энергию солнечного света. Вода, необходимая для поддержания тургора и протекания фотосинтеза, доставляется к листьям из корневой системы по ксилеме — одной из проводящих тканей растения. Потеря воды в результате испарения через устьица и в меньшей степени через кутикулу (транспирация) служит движущей силой транспорта по сосудам. Однако избыточная транспирация является нежелательной, и у растений в ходе эволюции сформировались различные приспособления, направленные на снижение потерь воды. Отток ассимилятов, необходимый для функционирования цикла Кальвина, осуществляется по флоэме. При интенсивном фотосинтезе углеводы могут полимеризоваться, и при этом в хлоропластах формируются крахмальные зёрна. Газообмен (поступление углекислого газа и выделение кислорода) осуществляется путём диффузии через устьица (некоторая часть газов движется через кутикулу).
Поскольку дефицит углекислого газа значительно увеличивает потери ассимилятов при фотодыхании, необходимо поддерживать высокую концентрацию углекислоты в межклеточном пространстве, что возможно при открытых устьицах. Однако поддержание устьиц в открытом состоянии при высокой температуре приводит к усилению испарения воды, что приводит к водному дефициту и также снижает продуктивность фотосинтеза. Этот конфликт решается в соответствии с принципом адаптивного компромисса. Кроме того, первичное поглощения углекислого газа ночью, при низкой температуре, у растений с CAM-фотосинтезом позволяет избежать высоких транспирационных потерь воды.
У некоторых суккулентов с вырожденными листьями (например, у большинства видов кактусов) основная фотосинтетическая активность связана со стеблем.
Фотосинтез на тканевом уровне
На тканевом уровне фотосинтез у высших растений обеспечивается специализированной тканью — хлоренхимой. Она располагается близ поверхности тела растения, где получает достаточно световой энергии. Обычно хлоренхима находится непосредственно под эпидермой. У растений, растущих в условиях повышенной инсоляции, между эпидермой и хлоренхимой может располагаться один или два слоя прозрачных клеток (гиподерма), обеспечивающих рассеивание света. У некоторых тенелюбивых растений хлоропластами богата и эпидерма (например, кислица). Часто хлоренхима мезофилла листа дифференцирована на палисадную (столбчатую) и губчатую, но может состоять и из однородных клеток. В случае дифференцировки наиболее богата хлоропластами палисадная хлоренхима.
 Хлоропласты в клетках листа
Хлоропласты в клетках листа Хлоропласты
Внутреннее пространство хлоропласта заполнено бесцветным содержимым (стромой) и пронизано мембранами (ламеллами), которые, соединяясь друг с другом, образуют тилакоиды, которые, в свою очередь, группируются в стопки, называемые гранами. Внутритилакоидное пространство отделено и не сообщается с остальной стромой; предполагается также, что внутреннее пространство всех тилакоидов сообщается между собой. Световые стадии фотосинтеза приурочены к мембранам, автотрофная фиксация CO2 происходит в строме.
В хлоропластах имеются свои ДНК, РНК, рибосомы (типа 70s), идёт синтез белка (хотя этот процесс и контролируется из ядра). Они не синтезируются вновь, а образуются путём деления предшествующих. Всё это позволило считать их потомками свободных цианобактерий, вошедших в состав эукариотической клетки в процессе симбиогенеза.
Фотосинтетические мембраны прокариот
Цианобактерии, таким образом, как бы сами являются хлоропластом, и в их клетке фотосинтетический аппарат не вынесен в особую органеллу. Их тилакоиды не образуют стопок, а формируют различные складчатые структуры (у единственной цианобактерии Gloeobacter violaceus тилакоиды отсутствуют вовсе, а весь фотосинтетический аппарат находится в цитоплазматической мембране, не образующей впячиваний). У них и растений также есть различия в светособирающем комплексе (см. ниже) и пигментном составе.
Световая (светозависимая) стадия
В ходе световой стадии фотосинтеза образуются высокоэнергетические продукты: АТФ, служащий в клетке источником энергии, и НАДФ, использующийся как восстановитель. В качестве побочного продукта выделяется кислород.
Фотохимическая суть процесса
Хлорофилл имеет два уровня возбуждения (с этим связано наличие двух максимумов на спектре его поглощения): первый связан с переходом на более высокий энергетический уровень электрона системы сопряжённых двойных связей, второй — с возбуждением неспаренных электронов азота и магния порфиринового ядра. При неизменном спине электрона формируются синглетные первое и второе возбуждённые состояния, при изменённом — триплетное первое и второе.
Второе возбуждённое состояние наиболее высокоэнергетично, нестабильно, и хлорофилл за 10−12 сек переходит с него на первое с потерей 100 кДж/моль энергии только в виде теплоты. Из первого синглетного и триплетного состояний молекула может переходить в основное с выделением энергии в виде света (флуоресценция и фосфоресценция соответственно) или тепла с переносом энергии на другую молекулу, либо, поскольку электрон на высоком энергетическом уровне слабо связан с ядром, с переносом электрона на другое соединение.
Первая возможность реализуется в светособирающих комплексах, вторая — в реакционных центрах, где под воздействием кванта света переходящий в возбуждённое состояние хлорофилл становится донором электрона (восстановителем) и передаёт его на первичный акцептор. Чтобы предотвратить возвращение электрона на положительно заряженный хлорофилл, первичный акцептор передаёт его вторичному. Кроме того, время жизни полученных соединений выше, чем у возбуждённой молекулы хлорофилла. Происходит стабилизация энергии и разделение зарядов. Для дальнейшей стабилизации вторичный донор электронов восстанавливает положительно заряженный хлорофилл, первичным же донором в случае оксигенного фотосинтеза является вода.
Проблемой, с которой сталкиваются при этом проводящие оксигенный фотосинтез организмы, является различие окислительно-восстановительных потенциалов воды (для полуреакции h3O → O2 (E0=+0,82 В) и НАДФ+ (E0=-0,32 В). Хлорофилл при этом должен иметь в основном состоянии потенциал больше +0,82 В, чтобы окислять воду, но при этом иметь в возбуждённом состоянии потенциал меньше −0,32 В, чтобы восстанавливать НАДФ+. Одна молекула хлорофилла не может отвечать обоим требованиям. Поэтому сформировались две фотосистемы, и для полного проведения процесса необходимы два кванта света и два хлорофилла разных типов.
Светособирающие комплексы
Хлорофилл выполняет две функции: поглощения и передачи энергии. Более 90 % всего хлорофилла хлоропластов входит в состав светособирающих комплексов (ССК), выполняющих роль антенны, передающей энергию к реакционному центру фотосистем I или II. Помимо хлорофилла, в ССК имеются каротиноиды, а у некоторых водорослей и цианобактерий — фикобилины, роль которых заключается в поглощении света тех длин волн, которые хлорофилл поглощает сравнительно слабо.
Передача энергии идёт резонансным путём (механизм Фёрстера) и занимает для одной пары молекул 10−10—10−12 с, расстояние на которое осуществляется перенос составляет около 1 нм. Передача сопровождается некоторыми потерями энергии (10 % от хлорофилла a к хлорофиллу b, 60 % от каротиноидов к хлорофиллу), из-за чего возможна только от пигмента с максимумом поглощения при меньшей длине волны к пигменту с большей. Именно в таком порядке взаимно локализуются пигменты ССК, причём наиболее длинноволновые хлорофиллы находятся в реакционных центрах. Обратный переход энергии невозможен.
ССК растений расположен в мембранах тилакоидов, у цианобактерий основная его часть вынесена за пределы мембран в прикреплённые к ним фикобилисомы — палочковидные полипептидно-пигментные комплексы, в которых находятся различные фикобилины: на периферии фикоэритрины (с максимумом поглощения при 495—565 нм), за ними фикоцианины (550—615 нм) и аллофикоцианины (610—670 нм), последовательно передающие энергию на хлорофилл a (680—700 нм) реакционного центра.
Основные компоненты цепи переноса электронов
Фотосистема II
Фотосистема — совокупность ССК, фотохимического реакционного центра и переносчиков электрона. Светособирающий комплекс II содержит 200 молекул хлорофилла a, 100 молекул хлорофилла b, 50 молекул каротиноидов и 2 молекулы феофитина. Реакционный центр фотосистемы II представляет собой пигмент-белковый комплекс, расположенный в тилакоидных мембранах и окружённый ССК. В нём находится димер хлорофилла-a с максимумом поглощения при 680 нм (П680). На него в конечном счёте передаётся энергия кванта света из ССК, в результате чего один из электронов переходит на более высокое энергетическое состояние, связь его с ядром ослабляется и возбуждённая молекула П680 становится сильным восстановителем (химический потенциал E0=-0,7 В).
П680 восстанавливает феофитин, в дальнейшем электрон переносится на хиноны, входящие в состав ФС II и далее на пластохиноны, транспортируемые в восстановленной форме к комплексу b6f. Одна молекула пластохинона переносит 2 электрона и 2 протона, которые берутся из стромы.
Заполнение электронной вакансии в молекуле П680 происходит за счёт воды. В состав ФС II входит водоокисляющий комплекс, содержащий в активном центре ионы марганца в количестве 4 штук. Для образования одной молекулы кислорода требуется две молекулы воды, дающие 4 электрона. Поэтому процесс проводится в 4 такта и для его полного осуществления требуется 4 кванта света. Комплекс находится со стороны внутритилакоидного пространства, и полученные 4 протона выбрасываются в него.
Таким образом, суммарный результат работы ФС II — это окисление 2 молекул воды с помощью 4 квантов света с образованием 4 протонов во внутритилакоидном пространстве и 2 восстановленных пластохинонов в мембране.
Цитохром-b6/f-комплекс
b6f-комплекс — это насос, перекачивающий протоны из стромы во внутритилакоидное пространство и создающий градиент их концентрации за счёт выделяющейся в окислительно-восстановительных реакциях электронтранспортной цепи энергии. 2 пластохинона дают перекачку 4 протонов. В дальнейшем трансмембранный протонный градиент (pH стромы около 8, внутритилакоидного пространства — 5) используется для синтеза АТФ трансмембранным ферментом АТФ-синтазой.
Фотосистема I
Светособирающий комплекс I содержит примерно 200 молекул хлорофилла.
В реакционном центре первой фотосистемы находится димер хлорофилла a с максимумом поглощения при 700 нм (П700). После возбуждения квантом света он восстанавливает первичный акцептор — хлорофилл a, тот — вторичный (витамин K1 или филлохинон), после чего электрон передаётся на ферредоксин, который и восстанавливает НАДФ с помощью фермента ферредоксин-НАДФ-редуктазы.
Белок пластоцианин, восстановленный в b6f-комплексе, транспортируется к реакционному центру первой фотосистемы со стороны внутритилакоидного пространства и передаёт электрон на окисленный П700.
Циклический и псевдоциклический транспорт электрона
Помимо полного нециклического пути электрона, описанного выше, обнаружены циклический и псевдоциклический.
Суть циклического пути заключается в том, что ферредоксин вместо НАДФ восстанавливает пластохинон, который переносит его назад на b6f-комплекс. В результате образуется больший протонный градиент и больше АТФ, но не возникает НАДФН.
При псевдоциклическом пути ферредоксин восстанавливает кислород, который в дальнейшем превращается в воду и может быть использован в фотосистеме II. При этом также не образуется НАДФН.
Темновая фаза
В темновой стадии с участием АТФ и НАДФ происходит восстановление CO2 до глюкозы (C6h22O6). Хотя свет не требуется для осуществления данного процесса, он участвует в его регуляции.
С3-фотосинтез, цикл Кальвина
Цикл Кальвина или восстановительный пентозофосфатный цикл состоит из трёх стадий:
- карбоксилирования;
- восстановления;
- регенерация акцептора CO2.
На первой стадии к рибулозо-1,5-бисфосфату присоединяется CO2 под действием фермента рибулозобисфосфат-карбоксилаза/оксигеназа. Этот белок составляет основную фракцию белков хлоропласта и предположительно наиболее распространённый фермент в природе. В результате образуется промежуточное неустойчивое соединение, распадающееся на две молекулы 3-фосфоглицериновой кислоты (ФГК).
Во второй стадии ФГК в два этапа восстанавливается. Сначала она фосфорилируется АТФ под действием фосфороглицерокиназы с образованием 1,3-дифосфоглицериновой кислоты (ДФГК), затем при воздействии триозофосфатдегидрогеназы и НАДФН ацил-фосфатная группа ДФГК дефосфорилируется и восстанавливается до альдегидной и образуется глицеральдегид-3-фосфат — фосфорилированный углевод (ФГА).
В третьей стадии участвуют 5 молекул ФГА, которые через образование 4-, 5-, 6- и 7-углеродных соединений объединяются в 3 5-углеродных рибулозо-1,5-бифосфата, для чего необходимы 3АТФ.
Наконец, две ФГА необходимы для синтеза глюкозы. Для образования одной её молекулы требуется 6 оборотов цикла, 6 CO2, 12 НАДФН и 18 АТФ.
С4-фотосинтез
Отличие этого механизма фотосинтеза от обычного заключается в том, что фиксация углекислого газа и его использование разделены в пространстве, между различными клетками растения[2].
При низкой концентрации растворённого в строме CO2рибулозобифосфаткарбоксилаза катализирует реакцию окисления рибулозо-1,5-бифосфата и его распад на 3-фосфоглицериновую кислоту и фосфогликолевую кислоту, которая вынужденно используется в процессе фотодыхания.
Для увеличения концентрации CO2 растения С4 типа изменили анатомию листа. Цикл Кальвина у них локализуется в клетках обкладки проводящего пучка, в клетках мезофилла же под действием ФЕП-карбоксилазы фосфоенолпируват карбоксилируется с образованием щавелеуксусной кислоты, которая превращается в малат или аспартат и транспортируется в клетки обкладки, где декарбоксилируется с образованием пирувата, возвращаемого в клетки мезофилла.
С4-фотосинтез практически не сопровождается потерями рибулозо-1,5-бифосфата из цикла Кальвина, поэтому более эффективен. Однако он требует не 18, а 30 АТФ на синтез 1 молекулы глюкозы. Это оправдывает себя в тропиках, где жаркий климат требует держать устьица закрытыми, что препятствует поступлению CO2 в лист, а также при рудеральной жизненной стратегии.
Фотосинтез по пути С4 проводят около 7600 видов растений. Все они относятся к цветковым: многие Злаковые (61 % видов, в том числе культурные — кукуруза, сахарный тростник и сорго и др.[3][4]), Гвоздичноцветные (наибольшая доля в семействах Маревые — 40 % видов, Амарантовые — 25 %), некоторые Осоковые, Астровые, Капустные, Молочайные[5][6].
CAM-фотосинтез
При фотосинтезе типа CAM (англ. Crassulaceae acid metabolism — кислотный метаболизм толстянковых) происходит разделение ассимиляции CO2 и цикла Кальвина не в пространстве, как у С4, а во времени[2]. Ночью в вакуолях клеток по аналогичному вышеописанному механизму при открытых устьицах накапливается малат, днём при закрытых устьицах идёт цикл Кальвина. Этот механизм позволяет максимально экономить воду, однако уступает в эффективности и С4, и С3. Он оправдан при стресстолерантной жизненной стратегии.
Значение
Фотосинтез составляет энергетическую основу всего живого на планете, кроме хемосинтезирующих бактерий.
Возникновение на Земле более 3 млрд лет назад механизма расщепления молекулы воды квантами солнечного света с образованием O2 представляет собой важнейшее событие в биологической эволюции, сделавшее свет Солнца главным источником энергии биосферы.
Фототрофы обеспечивают конверсию и запасание энергии термоядерных реакций, протекающих на Солнце, в энергию органических молекул. Солнечная энергия при участии фототрофов конвертируется в энергию химических связей органических веществ. Существование гетеротрофных организмов возможно исключительно за счёт энергии, запасённой фототрофами в органических соединениях. При использовании энергии химических связей органических веществ гетеротрофы высвобождают её в процессах дыхания и брожения.
Фотосинтез является основой продуктивности как сельскохозяйственных растений, так и животной пищи.
Энергия, получаемая человечеством при сжигании биотоплива (дрова, биогаз, биодизель, этанол, метанол) и ископаемого топлива (уголь, нефть, природный газ, торф), также является запасённой в процессе фотосинтеза.
Фотосинтез служит главным входом неорганического углерода в биогеохимический цикл.
Большая часть свободного кислорода атмосферы — биогенного происхождения и является побочным продуктом фотосинтеза. Формирование окислительной атмосферы (кислородная катастрофа) полностью изменило состояние земной поверхности, сделало возможным появление дыхания, а в дальнейшем, после образования озонового слоя, позволило жизни существовать на суше.
История изучения
Первые опыты по изучению фотосинтеза были проведены Джозефом Пристли в 1770—1780-х годах, когда он обратил внимание на «порчу» воздуха в герметичном сосуде горящей свечой (воздух переставал поддерживать горение, а помещённые в него животные задыхались) и «исправление» его растениями. Пристли сделал вывод, что растения выделяют кислород, который необходим для дыхания и горения, однако не заметил, что для этого растениям нужен свет. Это показал вскоре Ян Ингенхауз.
Позже было установлено, что помимо выделения кислорода растения поглощают углекислый газ и при участии воды синтезируют на свету органическое вещество. В 1842 г. Роберт Майер на основании закона сохранения энергии постулировал, что растения преобразуют энергию солнечного света в энергию химических связей. В 1877 г. В. Пфеффер назвал этот процесс фотосинтезом.
Хлорофиллы были впервые выделены в 1818 г. П. Ж. Пельтье и Ж. Каванту. Разделить пигменты и изучить их по отдельности удалось М. С. Цвету с помощью созданного им метода хроматографии. Спектры поглощения хлорофилла были изучены К. А. Тимирязевым, он же, развивая положения Майера, показал, что именно поглощённые лучи позволяют повысить энергию системы, создав вместо слабых связей С-О и О-Н высокоэнергетические С-С (до этого считалось что в фотосинтезе используются жёлтые лучи, не поглощаемые пигментами листа). Сделано это было благодаря созданному им методу учёта фотосинтеза по поглощённому CO2: в ходе экспериментов по освещению растения светом разных длин волн (разного цвета) оказалось, что интенсивность фотосинтеза совпадает со спектром поглощения хлорофилла.
Окислительно-восстановительную сущность фотосинтеза (как оксигенного, так и аноксигенного) постулировал Корнелис ван Ниль, он же в 1931 году доказал, что пурпурные бактерии и зелёные серобактерии осуществляют аноксигенный фотосинтез[7][8]. Окислительно-восстановительный характер фотосинтеза означал, что кислород в оксигенном фотосинтезе образуется полностью из воды, что экспериментально подтвердил в 1941 г. А. П. Виноградов в опытах с изотопной меткой. В 1937 г. Роберт Хилл установил, что процесс окисления воды (и выделения кислорода), а также ассимиляции CO2 можно разобщить. В 1954—1958 гг. Д. Арнон установил механизм световых стадий фотосинтеза, а сущность процесса ассимиляции CO2 была раскрыта Мелвином Кальвином с использованием изотопов углерода в конце 1940-х гг., за эту работу в 1961 г. ему была присуждена Нобелевская премия.
В 1955 году был выделен и очищен фермент рибулозобисфосфат-карбоксилаза/оксигеназа. С4-фотосинтез был описан Ю. С. Карпиловым в 1960 г. и М. Д. Хэтчем и Ч. Р. Слэком в 1966 г.
Эволюция
Существующие данные указывают на то, что наиболее древними организмами, запасающими энергию света в форме химической энергии, были археи, осуществляющие бесхлорофилльный фотосинтез, при котором не происходит образования восстановителя (НАДФН) и фиксации углекислого газа, а энергия запасается лишь в форме АТФ.
Несколько позже (3,7—3,8 миллиарда лет назад) независимо от архей и многократно в ходе эволюции возникали организмы (зеленые, пурпурные бактерии и другие группы эубактерий) с одной из фотосистем, позволяющей осуществлять аноксигенный фотосинтез, при котором в качестве доноров электронов используются соединения с низким редокс-потенциалом (водород, сульфиды и сероводород, сера, соединения железа (II), нитриты) или осуществляется циклический поток электронов вокруг фотосистемы.
Система оксигенного фотосинтеза, при которой в электрон-транспортной цепи совместно функционируют две фотосистемы, характерная для цианобактерий и происшедших от них в ходе эндосимбиоза хлоропластов водорослей и высших растений, возникла в эволюции однократно по разным оценкам от 3,5 до 2,4 миллиардов лет назад. Появление фотосистемы II позволило использовать в качестве неограниченного донора электронов воду — соединение с высоким редокс-потенциалом, то есть не склонное выступать в окислительно-восстановительных процессах в роли восстановителя, однако крайне распространенное на Земле.
С момента появления оксигенных фотосинтезирующих организмов начинается увеличение содержания молекулярного кислорода (сильный окислитель) на Земле, что приводит к накоплению кислорода в водах мирового океана, к окислению горных пород, к формированию озонового экрана и в конечном счете к накоплению кислорода в атмосфере планеты. Таким образом возникновение системы оксигенного фотосинтеза рассматривается как причина кислородной катастрофы и в частности перестройки древней восстановительной атмосферы Земли и формированию современной атмосферы окислительного типа. Формирование озонового слоя, защищающего поверхность Земли от опасного для живых организмов высокоэнергетического ультрафиолетового излучения, сделало возможным выход жизни на сушу. Одновременно с описанными перестройками гидро-, лито- и атмосферы происходили значительные изменения в биосфере: накопление кислорода привело к смене доминирующих анаэробных сообществ аэробными.
Прочие факты
- Морской слизень Elysia chlorotica ассимилирует хлоропласты водоросли Vaucheria litorea в клетки пищеварительного тракта. Хлоропласты способны фотосинтезировать в организме слизня в течение нескольких месяцев, что позволяет слизню жить за счёт глюкозы, полученной в результате фотосинтеза. Геном слизня кодирует некоторые белки, необходимые хлоропластам для фотосинтеза.[9]
- Предполагается, что в естественной среде фотосинтезирующие бактерии могут использовать не только свет Солнца, но и другие источники света, а потому могут находиться в местах, не подвергающихся солнечному облучению.[10] В 2005 году Томас Битти из университета Британской Колумбии и Роберт Блейкеншип из университета Аризоны в глубоководных пробах, взятых в окрестностях глубоководного термального источника у побережья Коста Рики обнаружили серобактерию GSB1, сходную с серобактериями родов Chlorobium и Prosthecochloris, содержащую бактериохлорофилл. Они предположили, что вероятность контаминации образца невелика и, следовательно, GSB1 использует для фотосинтеза не солнечный свет (который не проникает сквозь 2,4-километровую толщу моря), а тусклый длинноволновый (~750 нм) свет, испускаемый гидротермальными источниками[10].
- По состоянию на конец 70-х годов XX века мощность солнечной энергии, перерабатываемой земной растительностью при фотосинтезе, всего лишь на порядок превосходила мощность всех электростанций в мире[11]. По другим данным, первичная мощность фотосинтеза составляет 1014{\displaystyle 10^{14}} Вт[12]. В 2015 г в мире произведено 24×1012{\displaystyle 24\times 10^{12}} кВт*час электроэнергии. Получаем среднюю мощность всех электростанций в мире в 2015 г.: P=24×1015×360024×365×3600=2,7×1012{\displaystyle P={\frac {24\times 10^{15}\times 3600}{24\times 365\times 3600}}=2,7\times 10^{12}} Вт. Это составляет около 3% от первичной мощности фотосинтеза.
- Для оценки эффективности трансформации энергии излучения в энергию химических связей органических соединений используются различные расчетные коэффициенты эффективности фотосинтеза.
 Вт[12]. В 2015 г в мире произведено 24×1012{\displaystyle 24\times 10^{12}}
Вт[12]. В 2015 г в мире произведено 24×1012{\displaystyle 24\times 10^{12}} кВт*час электроэнергии. Получаем среднюю мощность всех электростанций в мире в 2015 г.: P=24×1015×360024×365×3600=2,7×1012{\displaystyle P={\frac {24\times 10^{15}\times 3600}{24\times 365\times 3600}}=2,7\times 10^{12}}
кВт*час электроэнергии. Получаем среднюю мощность всех электростанций в мире в 2015 г.: P=24×1015×360024×365×3600=2,7×1012{\displaystyle P={\frac {24\times 10^{15}\times 3600}{24\times 365\times 3600}}=2,7\times 10^{12}} Вт. Это составляет около 3% от первичной мощности фотосинтеза.
Вт. Это составляет около 3% от первичной мощности фотосинтеза.См. также
Примечания
- ↑ 1 2 3 4 5 Полищук В. Р. Как разглядеть молекулу. - М., Химия, 1979. - Тираж 70000 экз. - С. 301-305
- ↑ 1 2 Стасевич К. Почему кактусы не высыхают // Наука и жизнь. — 2016. — № 12. — С. 70—73. — ISSN 0028-1263. — URL: http://www.nkj.ru/archive/articles/30088/
- ↑ Sage, Rowan. 16 // C4 Plant Biology. — 1999. — P. 551–580. — ISBN 0-12-614440-0.
- ↑ Ботаники объяснили появление энергоэффективных растений
- ↑ Sage, Rowan. 7 // C4 Plant Biology. — 1999. — P. 228–229. — ISBN 0-12-614440-0.
- ↑ Kadereit, G; Borsch, T; Weising, K; Freitag, H (2003). «Phylogeny of Amaranthaceae and Chenopodiaceae and the Evolution of C4 Photosynthesis». International Journal of Plant Sciences 164 (6): 959–86. DOI:10.1086/378649.
- ↑ Кондратьева Е. Н. Фототрофные бактерии // Жизнь растений : в 6 т. / гл. ред. Ал. А. Фёдоров. — М. : Просвещение, 1974. — Т. 1 : Введение. Бактерии и актиномицеты / под ред. Н. А. Красильникова и А. А. Уранова. — С. 323. — 487 с. — 300 000 экз.
- ↑ van Niel C.B (1932). «On the morphology and physiology of the purple and green sulphur bacteria». Archiv für Mikrobiologie 3 (1): 1-112. DOI:10.1007/BF00454965.
- ↑ Rumpho ME, Worful JM, Lee J, et al. (November 2008). «Horizontal gene transfer of the algal nuclear gene psbO to the photosynthetic sea slug Elysia chlorotica». Proc. Natl. Acad. Sci. U.S.A. 105 (46): 17867–17871. DOI:10.1073/pnas.0804968105. PMID 19004808. Проверено 2008-11-24.
- ↑ 1 2 Beatty, J. Thomas; Jörg Overmann, Michael T. Lince, Ann K. Manske, Andrew S. Lang, Robert E. Blankenship, Cindy L. Van Dover, Tracey A. Martinson, F. Gerald Plumley (2005). «An obligately photosynthetic bacterial anaerobe from a deep-sea hydrothermal vent». Proceedings of the National Academy of Sciences of the United States of America 102 (26): 9306 -9310. DOI:10.1073/pnas.0503674102. Проверено 2011-02-10.
- ↑ Углекислый газ в атмосфере земли, Б. М. Смирнов, Объединённый институт высоких температур РАН, 126 (11) (1978), Москва
- ↑ Пономарев Л. И. Под знаком кванта. - М., Наука, 1989. - Тираж 75 000 экз. - ISBN 5-02-014049-X. - C. 331-339
Литература
- Холл Д., Рао К. Фотосинтез: Пер. с англ. — М.: Мир, 1983.
- Физиология растений / под ред. проф. Ермакова И. П. — М.: Академия, 2007
- Молекулярная биология клетки / Альбертис Б., Брей Д. и др. В 3 тт. — М.: Мир, 1994
- Рубин А. Б. Биофизика. В 2 тт. — М.: Изд. Московского университета и Наука, 2004.
- Чернавская Н. М., Чернавский Д. С. Туннельный транспорт электронов в фотосинтезе. М., 1977.
- В. Любименко. Влияние света на усвоение органических веществ зелёными растениями // Известия Императорской Академии наук. VI серия. — 1907. — № 12. — С. 395—426, с 6 табл.
- Медведев С. С. Физиология растений — СПб,: СПбГУ, 2006
Ссылки
ru-wiki.org
Что такое фотосинтез? Описание, особенности, фазы и значение фотосинтеза
Со школьной скамьи понятие фотосинтез ассоциируется с зеленым цветом. Это цвет пигмента под названием хлорофилл. Без его скопления в листьях процесс фотосинтеза не возможен. Как же выживает белая секвойя?
Это растение-альбинос. Прочие травы, кусты и деревья без хлорофилла погибают. Секвойя же – паразит, подпитывающийся от корней других растений. Это избавляет дерево от необходимости преобразовывать энергию солнца в пищу для себя. Именно в этом заключается суть фотосинтеза. Ему и посвятим статью.
Процесс фотосинтеза
Фотосинтез растений зиждется на 0,4% световых лучей. Половина из них не доходит до поверхности планеты. Из оставшихся для фотосинтеза подходит только 1/8. Работают ограничения по длине световой волны. Из подходящих лучей растения забирают 0,4%.

Если переводить в энергию, это 1% от ее общего количества. Привычное течение фотосинтеза проходит под действием света солнца. Однако, искусственные лучи растения тоже научились использовать.
Световой фотосинтез сводится к получению глюкозы. Она идет на питание растений. Побочный продукт реакции — кислород. Он выбрасывается представителями флоры во внешнюю среду, пополняя атмосферу Земли.
Получаются кислород и глюкоза в ходе реакции меж углекислым газом и водой. Хлорофилл в этом взаимодействии – своеобразный катализатор. Без него реакция не возможна.
Интересно, что хлорофилл встречается только в растениях. Функции, возложенные на пигмент, напоминают работу крови в организме животных. Хлорофилл подобен молекуле гемоглобина, но с магнием в центре.

В клетках же человеческой крови задействовано железо. Тем не менее, на организмы людей хлорофилл оказывает близкое к гемоглобину действие, а именно, повышает уровень кислорода крови и ускоряет обмен азота.
Реакция фотосинтеза может протекать быстро, или медленно. Все зависит от условий среды. Важны: интенсивность светового потока, температура воздуха, его насыщение углекислым газом и кислородом. Идеалом считается достижение точки компенсации. Так называют совпадение скоростей дыхания растения и выделения им кислорода.
Если свет в клетки хлоропласты, в коих скапливается хлорофилл, поступает сверху, то воду для реакции растения выкачивают из почвы. Вот зачем нужен полив растений. Недостаток влаги тормозит реакции фотосинтеза. В итоге, растение желтеет, то есть теряет хлорофилл.
Полей представителя флоры в этот момент, листья не зазеленеют. Выкачивать воду из почвы тоже помогает хлорофилл. Получается замкнутый круг. Нет полива – нет хлорофилла, нет хлорофилла – нет доставки воды в растение.

Теперь, уделим внимание глюкозе. Раз зелень вырабатывает ее из воды и углекислого газа, значит, из неорганического получается органика. Присоединяя к сахару то фосфор, то серу, то азот, растения производят витамины, жиры, белки, крахмалы. Дополнения к глюкозе травы да деревья берут из почвы. Элементы поступают растворенными в воде.
Фазы фотосинтеза
Фазы фотосинтеза – это деление процесса на фотолиз и восстановительную реакцию. Первый протекает на свету и сводится к выделению водорода. Кислород служит побочным продуктом реакции, однако, тоже нужным растению. Оно использует газ в процессе дыхания.
Световая фаза фотосинтеза возбуждает хлорофилл. От переизбытка энергии, его электрон отрывается и начинает перемещение по цепи органических соединений. В ходе путешествия частица способствует синтезу аденозиндифосфорной кислоты из аденозинтрифосфорной.
На это уходит данная электрону энергия. АДФ нужна для образования растением нуклеотидов. Они входят в нуклеиновые кислоты, без которых не возможен метаболизм представителей флоры.

Растратив энергию, электрон возвращается к молекуле хлорофилла. Эта клетка фотосинтеза вновь захватывает квант света. Уставший от работы электрон подкрепляется ею, опять отправляясь на дело. Такова световая фаза процесса. Однако, он не останавливается и в темноте.
Темновой фотосинтез направлен на захват из внешней среды уже углекислого газа. Вместе с водородом он участвует в образовании 6-углеродного сахара. Это и есть глюкоза. Этот результат фотосинтеза сопровождается, так же, образованием веществ, помогающих захватывать новые порции углекислого газа.
Захватываются они опять же, хлоропластами. Те тратят энергию, накопленную за день. Ресурс идет на связывание углекислого газа с рибулозобисфосфатом. Это 5-углеродный сахар. Реакция дает две молекулы фосфоглицериновой кислоты.
В каждой из них по 3 атома углерода. Это один из этапов цикла Кальвина. Он протекает в строме, то есть подстилке хлоропластов. Состоит цикл из трех реакций. Сначала, углекислый газ присоединяется к рубулозо-1,5-дифосфату.

Для реакции обязательно присутствие рубулозобифосфата-карбоксилазы. Это фермент. В его присутствии рождается гексоза. Из нее и получаются молекулы фосфоглицериновой кислоты.
После получения фосфоглицеринового соединения растение восстанавливает его до глицеральдегида-3-фосфата. Его молекулы идут на два «направления». В первом образуется глюкоза, а во втором рубулозо-1,5-дифосфат. Он, как помним, подхватывает газ углекислый.
Фотосинтез на обеих стадиях протекает в растениях активно, поскольку те приспособились захватывать днем максимальное количество энергии солнца. Вспомним школьные классы. Фотосинтезу посвящены несколько уроков ботаники.

Учителя рассказывают, почему у большинства растений плоские и широкие листья. Так представители флоры увеличивают площадь для улавливания квантов света. Не зря и люди сделали солнечные батареи широкими, но плоскими.
Фотосинтез углекислого газа
Углекислый газ проникает в растения через устица. Это подобие пор в листьях, стволах. Процесс всасывания газа и выпуска после через те же устица кислорода напоминает дыхание у людей.
Разница лишь в чередовании стадий. Люди вдыхают кислород, а выдыхают углекислый газ. У растений все наоборот. Так на планете удерживается равновесие двух газов в атмосфере.
Продукты фотосинтеза несут в себе энергию солнца. Животные перерабатывать ее не умеют. Съесть растения – единственная возможность «зарядиться» от дневного светила.
Перерабатывая углекислое соединение, растения способны давать людям и животным в два раза больше. Представители флоры работают с 0,03% газа в атмосфере. Как видно, углекислый газ в ней не из преобладающих.
В искусственных условиях ученые доводили процент углекислого вещества в воздухе до 0,05%. Огурцы, при этом, давали в 2 раза больше плодов. Так же реагировали на изменения помидоры, картофель, капуста.

Уровень углекислого газа ученые повышали, сжигая в теплицах опилки и прочие отходы деревообрабатывающей промышленности. Интересно, что при концентрации газа в 0,1% растения уже не были рады.
Многие виды начинали болеть. У помидоров, к примеру, в атмосфере с переизбытком углекислого соединения начинали желтеть и скручиваться листья. Это еще одно подтверждение опасности перенасыщения атмосферы CO2. Продолжая вырубку лесов и развитие промышленности, человек рискует поставить оставшиеся растения в непригодные для них условия.
Повышать уровень углекислого газа до оптимального можно не только путем сжигания отходов древесины, но и внося в почву удобрения. Они провоцируют размножение бактерий.
Многие микроорганизмы вырабатывают углекислое соединение. Сосредотачиваясь у земли, оно тут же захватывается растениями, идя на благо представителей флоры и всего населения Земли.
Значение фотосинтеза
Если допустить повышение уровня углекислого газа в нижних слоях атмосферы повсеместно, а не только в экспериментальных теплицах, наступит парниковый эффект. Это то самое глобальное потепление, которое то ли уже приближается, то ли и не «светит».
Ученые не сходятся во мнениях. Если говорить о фактах, говорящих в пользу парникового эффекта, вспоминается таяние льдов Антарктики. Там обитают белые медведи. Уже несколько лет они включены в Красную книгу.

Частью жизни медведей исторически является преодоление водных широт на пути к новым ледникам. Устремляясь к ним, животные все чаще выбиваются из сил, так и не достигнув цели. Водные просторы увеличиваются.
Доплыть до клочков суши становится все сложнее. Порой, медведи гибнут в пути. Порой, краснокнижные хищники добираются до земли, но изможденными. Сил на охоту и переходов уже по твердой почве не остается.
Из вышесказанного делаем вывод: без фотосинтеза или с сокращением его доли, уровень углекислого газа в атмосфере спровоцирует парниковый эффект. Изменится не только климат планеты, но и состав ее обитателей, их облик, приспособления к окружающей среде.
Так будет до тех пор, пока доля углекислого соединения в воздухе не достигнет критического 1%. Далее, под вопрос встает сам фотосинтез. Воды мировых океанов могут остаться единственным его источником. Водоросли ведь тоже «дышат». Клетки, хранящие хлорофилл, у них другие.
Однако, суть процесса фотосинтеза у наземных и водных растений одна. Концентрация углекислого газа в атмосфере не обязательно передается водной среде. В ней баланс может сохраниться.

Некоторые ученые предполагают, что при постепенном увеличении доли углекислого газа в воздухе, представители флоры смогут приспособиться к новым условиям. Помидоры не станут сворачивать листья, капитулируя перед реалиями будущего.
Возможно, растения эволюционируют, научившись перерабатывать большее количество СО2. Догадка ученых относится к категории «лучше не проверять». Слишком рискованно.
Значение фотосинтеза связано не только с поддержанием жизни самих растений и насыщением атмосферы Земли кислородом. Ученые бьются над искусственным проведением реакций.
Расщепляемая под действием радиации солнца на водород и кислород вода – источник энергии. Энергия эта, в отличие от получаемой из нефтепродуктов и каменного угля, экологически чистая, безопасная.
Где происходит фотосинтез – не важно. Важна энергия, которую он несет с собой. Пока, человек получает ресурс, лишь поглощая растительную пищу. Возникает вопрос, как же выживают плотоядные? Они не зря охотятся на травоядных, а не себе подобных. В мясе животных, питающихся травами и листьями, сохраняется часть их энергии.
Кроме энергии фотосинтеза важны и его продукты. Кислород, к примеру, идет не только на дыхание животных, но и на образования озонового слоя. Он располагается в стратосфере Земли, на границе с космосом.

Озон – одна из модификаций кислорода, которую тот принимает, поднимаясь на тысячекилометровые высоты. Здесь элемент борется с радиацией Солнца. Не будь озонового слоя, излучение светила достигало бы поверхности планеты в опасных для всего живого дозах.
Интересно, что в деле поддержания баланса газов на планете могут помочь некоторые беспозвоночные. Слизень Elisia Chloroti, к примеру, научился ассимилировать хлоропласты водорослей.
Обитатель морей съедает их, «приручая» клетки с хлорофиллом в слизистой своего желудка. Геном слизня кодирует белки, необходимые зеленому пигменту для фотосинтеза.
Выработанные вещества поставляются хлоропластам и те «кормят» беспозвоночное сладенькой глюкозой. На ней и люди некоторое время способны выживать. Достаточно вспомнить больницы, где ослабленным вводят глюкозу внутривенно.

Сахар – основной источник энергии и, главное, быстрый. Цепочка преобразования глюкозы в чистую энергию короче, чем цепь преобразований жиров, белков. Конечно, сахар научились синтезировать искусственно.
Но, многие ученые склоняются к мнению, что полезнее для организма глюкоза растений, фруктов и овощей. Это подобно эффекту витаминов. У синтетических и природных один состав, но чуть разниться положение атомов. Опыты доказывают, что аптечный витамин С пользу дает сомнительную, а вот то же вещество из лимона или капусты – бесспорную.
Бесспорна и польза фотосинтеза. Он привычен и, одновременно, хранит еще много тайн. Познавайте их, дабы обеспечить счастливое будущее и себе, и планете в целом.
cadiogorod.ru
что такое, определение, фазы :: SYL.ru
Важнейшим органическим процессом, без которого существование всех живых существ нашей планеты было бы под вопросом, является фотосинтез. Что такое фотосинтез? известно всем со школы. Грубо говоря, это процесс образования органических веществ из углекислого газа и воды, который происходит на свету и сопровождается выделением кислорода. Более сложное определение звучит следующим образом: фотосинтез – процесс преобразования световой энергии в энергию химических связей веществ органического происхождения при участии фотосинтетических пигментов. В современной практике под фотосинтезом обычно понимают совокупность процессов поглощения, синтеза и использования света в ряде эндергонических реакций, одной из которых является превращение углекислого газа в органические вещества. А теперь давайте узнаем подробнее, как протекает фотосинтез и на какие фазы этот процесс делится!

Общая характеристика
Хлоропласты, которые есть у каждого растения, отвечают за фотосинтез. Что такое хлоропласты? Это овальные пластиды, в которых содержится такой пигмент, как хлорофилл. Именно хлорофилл определяет зеленую окраску растений. У водорослей данный пигмент представлен в составе хроматофор – пигментсодержащих светоотражающих клеток разной формы. Бурые и красные водоросли, которые обитают на значительных глубинах, куда плохо попадает солнечный свет, имеют иные пигменты.
Вещества фотосинтеза входят в состав автотроф – организмов, способных синтезировать из неорганических веществ органические. Они являются самой нижней ступенью пищевой пирамиды, поэтому входят в рацион всех живых организмов планеты Земля.
Польза фотосинтеза
Зачем же нужен фотосинтез? Кислород, который выделяется из растений во время фотосинтеза, поступает в атмосферу. Поднимаясь в ее верхние слои, он образует озон, который защищает земную поверхность от сильного солнечного излучения. Именно благодаря озоновому экрану живые организмы могут комфортно находиться на суше. Кроме того, как известно, кислород нужен для дыхания живых организмов.

Ход процесса
Все начинается с того, что в хлоропласты попадает свет. Под его влиянием органеллы вытягивают из почвы воду, а также делят ее на водород и кислород. Таким образом, имеют место два процесса. Фотосинтез растений начинается в момент, когда листья уже впитали воду и углекислый газ. Световая энергия аккумулируется в тилакоидах – специальных отсеках хлоропластов, и делит молекулу воды на две составляющие. Часть кислорода уходит на дыхание растения, а оставшаяся часть - в атмосферу.
Затем углекислый газ попадает в пиреноиды – белковые гранулы, окруженные крахмалом. Сюда же поступает водород. Смешавшись друг с другом, эти вещества образуют сахар. Эта реакция также проходит с выделением кислорода. Когда сахар (обобщающее название простых углеводов) смешивается с азотом, серой и фосфором, поступающими в растение из почвы, образуется крахмал (сложный углевод), белки, жиры, витамины и прочие вещества, необходимые для жизни растений. В абсолютном большинстве случаев фотосинтез происходит в условиях природного освещения. Однако искусственное освещение также может в нем поучаствовать.
Классы фотосинтеза
Вплоть до 60-х годов двадцатого века науке был известен один механизм восстановления углекислого газа – по С3-пентозофосфатному пути. Недавно австралийские ученые доказали, что у некоторых видов растений данный процесс может протекать по циклу С4-дикарбоновых кислот.
У растений, которые восстанавливают углекислый газ по С3 пути, фотосинтез лучше всего проходит при умеренной температуре и слабой освещенности, в лесах или темных местах. К таковым растениям можно отнести львиную долю культурных растений и почти все овощи, которые составляют основу нашего рациона.

У второго класса растений фотосинтез активнее всего протекает в условиях высокой температуры и сильной освещенности. В эту группу входят растения, которые произрастают в тропическом и теплом климате, к примеру кукуруза, сахарный тростник, сорго и так далее.
Метаболизм растений, кстати говоря, был обнаружен весьма недавно. Ученым удалось выяснить, что некоторые растения имеют специальные ткани для сохранения запасов воды. Углекислый газ у них скапливается в виде органических кислот и переходит в углеводы лишь через 24 часа. Этот механизм дает растениям возможность сэкономить воду.
Как проходит процесс?
Мы уже знаем в общих чертах, как протекает процесс фотосинтеза и какой фотосинтез бывает, теперь давайте познакомимся с ним глубже.
Начинается все с того, что растение поглощает свет. Ей в этом помогает хлорофилл, который в виде хлоропластов располагается в листьях, стеблях и плодах растения. Основное количество данного вещества сконцентрировано именно в листьях. Все дело в том, что благодаря своей плоской структуре, лист притягивает много света. А чем больше света, тем больше энергии для фотосинтеза. Таким образом, листья в растении выступают своеобразными локаторами, улавливающими свет.
Когда свет поглощен, хлорофилл пребывает в возбужденном состоянии. Он передает энергию другим органам растения, которые участвуют в следующей стадии фотосинтеза. Второй этап процесса протекает без участия света и состоит в химической реакции с участием воды, получаемой из почвы, и углекислого газа, получаемого из воздуха. На этой стадии синтезируются углеводы, которые крайне необходимы для жизни любого организма. В данном случае они не только питают само растение, но и передаются животным, которые его съедают. Люди также получают эти вещества, употребив продукты растительного или животного происхождения.

Фазы процесса
Будучи довольно сложным процессом, фотосинтез делится на две фазы: световую и темновую. Как можно понять из названия, для первой фазы обязательно наличие солнечного излучения, а для второй - нет. Во время световой фазы хлорофилл поглощает квант света, образуя молекулы АТФ и НАДН, без которых невозможен фотосинтез. Что такое АТФ и НАДН?
АТФ (аденозитрифосфат) – нуклеиновый кофермент, которые содержит высокоэнергетические связи и служит источником энергии в любом органическом превращении. Соединение часто называют энергетической волютой.
НАДН (никотинамидадениндинуклеотид) – источник водорода, который используется для синтеза углеводов с участием углекислого газа во второй фазе такого процесса, как фотосинтез.
Световая фаза
Хлоропласты содержат много молекул хлорофилла, каждая из которых поглощает свет. Его поглощают и другие пигменты, но они не способны к фотосинтезу. Процесс проходит лишь в части молекул хлорофилла. Остальные молекулы образуют антенные и светособирающие комплексы (ССК). Они накапливают кванты светового излучения и передают их в реакционные центры, которые также называют ловушками. Реакционные центры располагаются в фотосистемах, которых у фотосинтезирующего растения две. Первая содержит молекулу хлорофилла, способную поглощать свет с длиной волны 700 нм, а вторая – 680 нм.
Итак, два типа молекул хлорофилла поглощают свет и возбуждаются, что способствует переходу электронов на более высокий энергетический уровень. Возбужденные электроны, обладающие большим количеством энергии, отрываются и поступают в цепь переносчиков, расположенную в мембранах тилакоидов (внутренние структуры хлоропластов).

Переход электронов
Электрон из первой фотосистемы переходит от хлорофилла Р680 к пластохинону, а электрон из второй системы – к ферредоксину. При этом на месте отрыва электронов в молекуле хлорофилла образуется свободное место.
Для восполнения недостачи молекула хлорофилла Р680 принимает электроны из воды, образуя ионы водорода. А вторая молекула хлорофилла восполняет недостачу через систему переносчиков от первой фотосистемы.
Так протекает световая фаза фотосинтеза, суть которой состоит в переносе электронов. Параллельно электронному транспорту проходит перемещение ионов водорода через мембрану. Это приводит к их накоплению внутри тилакоида. Накапливаясь в большом количестве, они высвобождаются наружу с помощью сопрягающего фактора. Результатом транспорта электронов является образование соединения НАДН. А перенос иона водорода приводит к образованию энергетической валюты АТФ.
По окончании световой фазы в атмосферу поступает кислород, а внутри лепестка образуются АТФ и НАДН. Затем начинается темновая фаза фотосинтеза.

Темновая фаза
Для этой фазы фотосинтеза необходим углекислый газ. Растение постоянно поглощает его из воздуха. С этой целью на поверхности листка есть устьица – специальные структуры, которые при открывании всасывают углекислый газ. Поступая вовнутрь листка, он растворяется в воде и участвует в процессах световой фазы.
Во время световой фазы в большинстве растений углекислый газ связывается с органическим соединением, которое содержит 5 атомов углерода. В результате образуется пара молекул трехуглеродного соединения под названием 3-фосфоглицериновая кислота. Именно из-за того, что первичным результатом процесса является данное соединение, растения с таким типом фотосинтеза называют С3-растениями.
Дальнейшие процессы, проходящие в хлоропластах, весьма сложны для неискушенных обывателей. В конечном итоге получается шестиуглеродное соединение, синтезирующее простые или сложные углеводы. Именно в виде углеводов растение скапливает энергию. Небольшая часть веществ остается в листе и выполняет его нужды. Остальные углеводы циркулируют по всему растению и поступают в те места, где они больше всего нужны.
Фотосинтез зимой

Многие хотя бы раз в жизни задавались вопросом о том, откуда берется кислород в холодное время года. Во-первых, кислород вырабатывается не только лиственными растениями, но и хвойными, а также морскими растениями. И если лиственные растения зимой замирают, то хвойные продолжают дышать, хотя и менее интенсивно. Во-вторых, содержание кислорода в атмосфере не зависит от того, скинули ли деревья свои листья. Кислород занимает 21 % атмосферы, в любой точки нашей планеты в любое время года. Эта величина не меняется, так как воздушные массы перемещаются очень быстро, а зима наступает не одновременно во всех странах. Ну и, в-третьих, зимой в нижних слоях воздуха, которые мы вдыхаем, содержание кислорода даже больше, чем летом. Причина этого явления – низкая температура, из-за которой кислород становится плотнее.
Заключение
Сегодня мы вспомнили, что такое фотосинтез, что такое хлорофил, и как растения выделяют кислород, поглощая углекислый газ. Безусловно, фотосинтез является важнейшим процессом в нашей жизни. Он напоминает нам о необходимости бережного отношения к природе.
www.syl.ru
Фотосинтез - это... Что такое Фотосинтез?

 Лист растения
Лист растения 
Фотосинтез (от др.-греч. φῶς — свет и σύνθεσις — соединение, складывание, связывание, синтез) — процесс образования органических веществ из углекислого газа и воды на свету при участии фотосинтетических пигментов (хлорофилл у растений, бактериохлорофилл и бактериородопсин у бактерий).
В современной физиологии растений под фотосинтезом чаще понимается фотоавтотрофная функция — совокупность процессов поглощения, превращения и использования энергии квантов света в различных эндэргонических реакциях, в том числе превращения углекислого газа в органические вещества.Типы фотосинтеза
Бесхлорофилльный фотосинтез
Осуществляется археями рода Halobacterium, является наиболее примитивным типом фотосинтеза, кванты света поглощаются белком-бактериородопсином, имеющим сходство с родопсином в виде наличия ретиналя, этот тип фотосинтеза отличается отсутствием электрон-транспортной цепи, синтез АТФ осуществляется через создание электрохимического градиента протонов или ионов хлора при помощи бактериородопсиновой и галородопсиновой ионной помпы.
Хлорофилльный фотосинтез
Аноксигенный
Осуществляется пурпурными и зелёными бактериями, а также геликобактериями.
Оксигенный
Оксигенный фотосинтез распространён гораздо шире. Осуществляется растениями, цианобактериями и прохлорофитами.
Этапы фотосинтеза:
- фотофизический;
- фотохимический;
- химический.
На первом этапе происходит поглощение квантов света пигментами, их переход в возбуждённое состояние и передача энергии к другим молекулам фотосистемы. На втором этапе происходит разделение зарядов в реакционном центре, перенос электронов по фотосинтетической электронотранспортной цепи, что заканчивается синтезом АТФ и НАДФН. Первые два этапа вместе называют светозависимой стадией фотосинтеза. Третий этап происходит уже без обязательного участия света и включает в себя биохимические реакции синтеза органических веществ с использованием энергии, накопленной на светозависимой стадии. Чаще всего в качестве таких реакций рассматривается цикл Кальвина и глюкогенез, образование сахаров и крахмала из углекислого газа воздуха.
Первые процессы фотосинтеза у цианобактерий появились ещё в архейскую эру.
Пространственная локализация
 Хлоропласты в клетках листа
Хлоропласты в клетках листа Фотосинтез растений осуществляется в хлоропластах: обособленных двухмембранных органеллах клетки. Хлоропласты могут быть в клетках плодов, стеблей, однако основным органом фотосинтеза, анатомически приспособленным к его ведению, является лист. В листе наиболее богата хлоропластами ткань палисадной паренхимы. У некоторых суккулентов с вырожденными листьями (например, кактусы) основная фотосинтетическая активность связана со стеблем.
Свет для фотосинтеза захватывается более полно благодаря плоской форме листа, обеспечивающей большое отношение поверхности к объёму. Вода доставляется из корня по развитой сети сосудов (жилок листа). Углекислый газ поступает отчасти посредством диффузии через кутикулу и эпидермис, однако большая его часть диффундирует в лист через устьица и по листу по межклеточному пространству. Растения, осуществляющие CAM фотосинтез, сформировали особые механизмы для активной ассимиляции углекислого газа.
Внутреннее пространство хлоропласта заполнено бесцветным содержимым (стромой) и пронизано мембранами (ламеллами), которые соединяясь друг с другом образуют тилакоиды, которые в свою очередь группируются в стопки, называемые граны. Внутритилакоидное пространство отделено и не сообщается с остальной стромой, предполагается также что внутреннее пространство всех тилакоидов сообщается между собой. Световые стадии фотосинтеза приурочены к мембранам, автотрофная фиксация CO2 происходит в строме.
В хлоропластах имеются свои ДНК, РНК, рибосомы (70s типа), идёт синтез белка (хотя этот процесс и контролируется из ядра). Они не синтезируются вновь, а образуются путём деления предшествующих. Всё это позволило считать их потомками свободных цианобактерий, вошедших в состав эукариотической клетки в процессе симбиогенеза.
Цианобактерии, таким образом, как бы сами являются хлоропластом и в их клетке фотосинтетический аппарат не вынесен в особую органеллу. Их тилакоиды, однако, не образуют стопок, а формируют различные складчатые структуры (у единственной цианобактерии Gloeobacter violaceus тилакоиды отсутствуют вовсе, а весь фотосинтетический аппарат находится в цитоплазматической мембране, не образующей впячиваний). У них и растений также есть различия в светособирающем комплексе (см. ниже) и пигментном составе.
Световая (светозависимая) стадия
В ходе световой стадии фотосинтеза образуются высокоэнергетические продукты: АТФ, служащий в клетке источником энергии, и НАДФН, использующийся как восстановитель. В качестве побочного продукта выделяется кислород. В общем, роль световых реакций фотосинтеза заключается в том, что в световую фазу синтезируются молекула АТФ и молекулы-переносчики протонов, то есть НАДФ Н2.
Фотохимическая суть процесса
Хлорофилл имеет два уровня возбуждения (с этим связано наличие двух максимумов на спектре его поглощения): первый связан с переходом на более высокий энергетический уровень электрона системы сопряжённых двойных связей, второй — с возбуждением неспаренных электронов азота и магния порфиринового ядра. При неизменном спине электрона формируются синглетные первое и второе возбуждённое состояние, при изменённом — триплетное первое и второе.
Второе возбуждённое состояние наиболее высокоэнергетично, нестабильно и хлорофилл за 10-12 сек переходит с него на первое, с потерей 100 кДж/моль энергии только в виде теплоты. Из первого синглетного и триплетного состояний молекула может переходить в основное с выделением энергии в виде света (флуоресценция и фосфоресценция соответственно) или тепла, с переносом энергии на другую молекулу, либо, поскольку электрон на высоком энергетическом уровне слабо связан с ядром, с переносом электрона на другое соединение.
Первая возможность реализуется в светособирающих комплексах, вторая — в реакционных центрах, где переходящий в возбуждённое состояние под воздействием кванта света хлорофилл становится донором электрона (восстановителем) и передаёт его на первичный акцептор. Чтобы предотвратить возвращение электрона на положительно заряженный хлорофилл, первичный акцептор передаёт его вторичному. Кроме того, время жизни полученных соединений выше, чем у возбуждённой молекулы хлорофилла. Происходит стабилизация энергии и разделения зарядов. Для дальнейшей стабилизации вторичный донор электронов восстанавливает положительно заряженный хлорофилл, первичным донором же является в случае оксигенного фотосинтеза вода.
Проблемой, с которой сталкиваются при этом проводящие оксигенный фотосинтез организмы, является различие окислительно-восстановительных потенциалов воды (для полуреакции h3O → O2 (E0=+0,82 В) и НАДФ+ (E0=-0,32 В). Хлорофилл при этом должен иметь в основном состоянии потенциал больший +0,82 В чтобы окислять воду, но при этом иметь в возбуждённом состоянии потенциал меньший чем −0,32 В чтобы восстанавливать НАДФ+. Одна молекула хлорофилла не может отвечать обоим требованиям. Поэтому сформировались две фотосистемы, и для полного проведения процесса необходимы два кванта света и два хлорофилла разных типов.
Светособирающие комплексы
Хлорофилл выполняет две функции: поглощения и передачи энергии. Более 90 % всего хлорофилла хлоропластов входит в состав светособирающих комплексов (ССК), выполняющих роль антенны, передающей энергию к реакционному центру фотосистем I или II. Помимо хлорофилла в ССК имеются каротиноиды, а у некоторых водорослей и цианобактерий — фикобилины, роль которых заключается в поглощении света тех длин волн, которые хлорофилл поглощает сравнительно слабо.
Передача энергии идёт резонансным путём (механизм Фёрстера) и занимает для одной пары молекул 10−10—10−12 с, расстояние на которое осуществляется перенос составляет около 1 нм. Передача сопровождается некоторыми потерями энергии (10 % от хлорофилла a к хлорофиллу b, 60 % от каротиноидов к хлорофиллу), из-за чего возможна только от пигмента с максимумом поглощения при меньшей длине волны к пигменту с большей. Именно в таком порядке взаимно локализуются пигменты ССК, причём наиболее длинноволновые хлорофиллы находятся в реакционных центрах. Обратный переход энергии невозможен.
ССК растений расположен в мембранах тилакоидов, у цианобактерий основная его часть вынесена за пределы мембран в прикреплённые к ним фикобилисомы — палочковидные полипептидно-пигментные комплексы, в которых находятся различные фикобилины: на периферии фикоэритрины (с максимумом поглощения при 495—565 нм), за ними фикоцианины (550—615 нм) и аллофикоцианины (610—670 нм), последовательно передающие энергию на хлорофилл a (680—700 нм) реакционного центра.
Основные компоненты электронтранспортной цепи
Основная статья: Цепь переноса электронов фотосинтеза
Фотосистема II
Фотосистема — совокупность ССК, фотохимического реакционного центра и переносчиков электрона. Светособирающий комплекс II содержит 200 молекул хлорофилла a, 100 молекул хлорофилла b, 50 молекул каротиноидов и 2 молекулы феофитина. Реакционный центр фотосистемы II представляет собой пигмент-белковый комплекс, расположенный в тилакоидных мембранах и окружённый ССК. В нём находится димер хлорофилла a с максимумом поглощения при 680 нм (П680). На него в конечном счёте передаётся энергия кванта света из ССК, в результате чего один из электронов переходит на более высокое энергетическое состояние, связь его с ядром ослабляется и возбуждённая молекула П680 становится сильным восстановителем (E0=-0,7 В).
П680 восстанавливает феофитин, в дальнейшем электрон переносится на хиноны, входящие в состав ФС II и далее на пластохиноны, транспортируемые в восстановленной форме к b6f комплексу. Одна молекула пластохинона переносит 2 электрона и 2 протона, которые берутся из стромы.
Заполнение электронной вакансии в молекуле П680 происходит за счёт воды. В состав ФС II входит водоокисляющий комплекс, содержащий в активном центре ионы марганца в количестве 4 штук. Для образования одной молекулы кислорода требуется две молекулы воды, дающие 4 электрона. Поэтому процесс проводится в 4 такта и для его полного осуществления требуется 4 кванта света. Комплекс находится со стороны внутритилакоидного пространства и полученные 4 протона выбрасываются в него.
Таким образом, суммарный результат работы ФС II — это окисление 2 молекул воды с помощью 4 квантов света с образованием 4 протонов во внутритилакоидном пространстве и 2 восстановленных пластохинонов в мембране.
b6f или b/f-комплекс
b6f комплекс является насосом, перекачивающим протоны из стромы во внутритилакоидное пространство и создающий градиент их концентрации за счёт выделяющейся в окислительно-восстановительных реакциях электронтранспортной цепи энергии. 2 пластохинона дают перекачку 4 протонов. В дальнейшем трансмембранный протонный градиент (pH стромы около 8, внутритилакоидного пространства — 5) используется для синтеза АТФ трансмембранным ферментом АТФ-синтазой.
Фотосистема I
Светособирающий комплекс I содержит примерно 200 молекул хлорофилла.
В реакционном центре первой фотосистемы находится димер хлорофилла a с максимумом поглощения при 700 нм (П700). После возбуждения квантом света он восстанавливает первичный акцептор — хлорофилл a, тот — вторичный (витамин K1 или филлохинон), после чего электрон передаётся на ферредоксин, который и восстанавливает НАДФ с помощью фермента ферредоксин-НАДФ-редуктазы.
Белок пластоцианин, восстановленный в b6f комплексе, транспортируется к реакционному центру первой фотосистемы со стороны внутритилакоидного пространства и передаёт электрон на окисленный П700.
Циклический и псевдоциклический транспорт электрона
Помимо полного нециклического пути электрона, описанного выше, обнаружены циклический и псевдоциклический.
Суть циклического пути заключается в том, что ферредоксин вместо НАДФ восстанавливает пластохинон, который переносит его назад на b6f комплекс. В результате образуется больший протонный градиент и больше АТФ, но не возникает НАДФН.
При псевдоциклическом пути ферредоксин восстанавливает кислород, который в дальнейшем превращается в воду и может быть использован в фотосистеме II. При этом также не образуется НАДФН.
Темновая стадия
В темновой стадии с участием АТФ и НАДФН происходит восстановление CO2 до глюкозы (C6h22O6). Хотя свет не требуется для осуществления данного процесса, он участвует в его регуляции.
С3-фотосинтез, цикл Кальвина
Цикл Кальвина или восстановительный пентозофосфатный цикл состоит из трёх стадий:
- карбоксилирования;
- восстановления;
- регенерация акцептора CO2.
На первой стадии к рибулозо-1,5-бифосфату присоединяется CO2 под действием фермента рибулозобисфосфат-карбоксилаза/оксигеназа. Этот белок составляет основную фракцию белков хлоропласта и предположительно наиболее распространённый фермент в природе. В результате образуется промежуточное неустойчивое соединение, распадающееся на две молекулы 3-фосфоглицериновой кислоты (ФГК).
Во второй стадии ФГК в два этапа восстанавливается. Сначала она фосфорилируется АТФ под действием фосфороглицерокиназы с образованием 1,3-дифосфоглицериновой кислоты (ДФГК), затем при воздействии триозофосфатдегидрогеназы и НАДФН ацил-фосфатная группа ДФГК дефосфорилируется и восстанавливается до альдегидной и образуется глицеральдегид-3-фосфат — фосфорилированный углевод (ФГА).
В третьей стадии участвуют 5 молекул ФГА, которые через образование 4-, 5-, 6- и 7-углеродных соединений объединяются в 3 5-углеродных рибулозо-1,5-бифосфата, для чего необходимы 3АТФ.
Наконец, две ФГА необходимы для синтеза глюкозы. Для образования одной её молекулы требуется 6 оборотов цикла, 6 CO2, 12 НАДФН и 18 АТФ.
С4-фотосинтез
Основные статьи: Цикл Хетча-Слэка-Карпилова, С4-фотосинтез
При низкой концентрации растворённого в строме CO2 рибулозобифосфаткарбоксилаза катализирует реакцию окисления рибулозо-1,5-бифосфата и его распад на 3-фосфоглицериновую кислоту и фосфогликолевую кислоту, которая вынужденно используется в процессе фотодыхания.
Для увеличения концентрации CO2 растения С4 типа изменили анатомию листа. Цикл Кальвина у них локализуется в клетках обкладки проводящего пучка, в клетках мезофилла же под действием ФЕП-карбоксилазы фосфоенолпируват карбоксилируется с образованием щавелеуксусной кислоты, которая превращается в малат или аспартат и транспортируется в клетки обкладки, где декарбоксилируется с образованием пирувата, возвращаемого в клетки мезофилла.
С4 фотосинтез практически не сопровождается потерями рибулозо-1,5-бифосфата из цикла Кальвина, поэтому более эффективен. Однако он требует не 18, а 30 АТФ на синтез 1 молекулы глюкозы. Это оправдывает себя в тропиках, где жаркий климат требует держать устьица закрытыми, что препятствует поступлению CO2 в лист, а также при рудеральной жизненной стратегии.
САМ фотосинтез
При CAM (англ. Crassulaceae acid metabolism — кислотный метаболизм толстянковых) фотосинтезе происходит разделение ассимиляции CO2 и цикла Кальвина не в пространстве как у С4, а во времени. Ночью в вакуолях клеток по аналогичному вышеописанному механизму при открытых устьицах накапливается малат, днём при закрытых устьицах идёт цикл Кальвина. Этот механизм позволяет максимально экономить воду, однако уступает в эффективности и С4, и С3. Он оправдан при стресстолерантной жизненной стратегии.
Значение фотосинтеза
Фотосинтез является основным источником биологической энергии, фотосинтезирующие автотрофы используют её для синтеза органических веществ из неорганических, гетеротрофы существуют за счёт энергии, запасённой автотрофами в виде химических связей, высвобождая её в процессах дыхания и брожения. Энергия, получаемая человечеством при сжигании ископаемого топлива (уголь, нефть, природный газ, торф), также является запасённой в процессе фотосинтеза.
Фотосинтез является главным входом неорганического углерода в биологический цикл. Весь свободный кислород атмосферы — биогенного происхождения и является побочным продуктом фотосинтеза. Формирование окислительной атмосферы (кислородная катастрофа) полностью изменило состояние земной поверхности, сделало возможным появление дыхания, а в дальнейшем, после образования озонового слоя, позволило жизни выйти на сушу.
История изучения фотосинтеза
Первые опыты по фотосинтезу были проведены Джозефом Пристли в 1770—1780-х годах, когда он обратил внимание на «порчу» воздуха в герметичном сосуде горящей свечой (воздух переставал быть способен поддерживать горение, помещённые в него животные задыхались) и «исправление» его растениями. Пристли сделал вывод, что растения выделяют кислород, который необходим для дыхания и горения, однако не заметил, что для этого растениям нужен свет. Это показал вскоре Ян Ингенхауз.
Позже было установлено, что помимо выделения кислорода растения поглощают углекислый газ и при участии воды синтезируют на свету органическое вещество. В 1842 Роберт Майер на основании закона сохранения энергии постулировал, что растения преобразуют энергию солнечного света в энергию химических связей. В 1877 В. Пфеффер назвал этот процесс фотосинтезом.
Хлорофиллы были впервые выделены в 1818 П. Ж. Пельтье и Ж. Кавенту. Разделить пигменты и изучить их по отдельности удалось М. С. Цвету с помощью созданного им метода хроматографии. Спектры поглощения хлорофилла были изучены К. А. Тимирязевым, он же, развивая положения Майера, показал, что именно поглощенные лучи позволяют повысить энергию системы, создав вместо слабых связей С-О и О-Н высокоэнергетические С-С (до этого считалось что в фотосинтезе используются жёлтые лучи, не поглощаемые пигментами листа). Сделано это было благодаря созданному им методу учёта фотосинтеза по поглощённому CO2: в ходе экспериментов по освещению растения светом разных длин волн (разного цвета) оказалось, что интенсивность фотосинтеза совпадает со спектром поглощения хлорофилла.
Окислительно-восстановительную сущность фотосинтеза (как оксигенного, так и аноксигенного) постулировал Корнелис ван Ниль. Это означало, что кислород в фотосинтезе образуется полностью из воды, что экспериментально подтвердил в 1941 А. П. Виноградов в опытах с изотопной меткой. В 1937 г. Роберт Хилл установил, что процесс окисления воды (и выделения кислорода), а также ассимиляции CO2 можно разобщить. В 1954—1958 Д. Арнон установил механизм световых стадий фотосинтеза, а сущность процесса ассимиляции CO2 была раскрыта Мельвином Кальвином с использованием изотопов углерода в конце 1940-х, за эту работу в 1961 ему была присуждена Нобелевская премия.
В 1955 году был выделен и очищен фермент рибулозобисфосфат-карбоксилаза/оксигеназа. С4 фотосинтез был описан Ю. С. Карпиловым в 1960 и М. Д. Хэтчем и К. Р. Слэком в 1966.
Прочие факты
- Морской слизень Elysia chlorotica ассимилирует хлоропласты водоросли Vaucheria litorea в клетки пищеварительного тракта. Хлоропласты способны фотосинтезировать в организме слизня в течение нескольких месяцев, что позволяет слизню жить за счет глюкозы, полученной в результате фотосинтеза. Геном слизня кодирует некоторые белки, необходимые хлоропластам для фотосинтеза.[1]
- Предполагается, что в естественной среде фотосинтезирующие бактерии могут использовать не только свет Солнца, но и другие источники света, а потому могут находиться в местах, не подвергающихся облучению звезды[2] В 2005 году Томас Битти из университета Британской Колумбии и Роберт Блейкеншип из университета Аризоны в глубоководных пробах, взятых в окрестностях глубоководного термального источника у побережья Коста Рики обнаружили серобактерию GSB1, сходную с серобактериями родов Chlorobium и Prosthecochloris, содержащую бактериохлорофилл. Они предположили, что вероятность контаминации образца невелика и, следовательно, GSB1 использует для фотосинтеза не солнечный свет (который не проникает сквозь 2,4-километровую толщу моря), а тусклый длинноволновый (~750 нм) свет, испускаемый гидротермальными источниками[2].
- По состоянию на конец 70-х годов XX века, мощность солнечной энергии, перерабатываемой земной растительностью при фотосинтезе, всего лишь на порядок превосходила мощность всех электростанций в мире.[3]
См. также
Литература
- Холл Д., Рао К. Фотосинтез: Пер. с англ. — М.: Мир, 1983.
- Физиология растений / под ред. проф. Ермакова И. П. — М.: Академия, 2007
- Молекулярная биология клетки / Альбертис Б., Брей Д. и др. В 3 тт. — М.: Мир, 1994
- Рубин А. Б. Биофизика. В 2 тт. — М.: Изд. Московского университета и Наука, 2004.
- Чернавская Н. М., Чернавский Д. С. Туннельный транспорт электронов в фотосинтезе. М., 1977.
- В. Любименко. Влияние света на усвоение органических веществ зелёными растениями // Известия Императорской Академии наук. VI серия. — 1907. — № 12. — С. 395—426, с 6 табл.
- Медведев С. С. Физиология растений — СПб,: СПбГУ, 2004
Примечания
- ↑ Rumpho ME, Worful JM, Lee J, et al. (November 2008). «Horizontal gene transfer of the algal nuclear gene psbO to the photosynthetic sea slug Elysia chlorotica». Proc. Natl. Acad. Sci. U.S.A. 105 (46): 17867–17871. DOI:10.1073/pnas.0804968105. PMID 19004808. Проверено 2008-11-24.
- ↑ 1 2 Beatty, J. Thomas; Jörg Overmann, Michael T. Lince, Ann K. Manske, Andrew S. Lang, Robert E. Blankenship, Cindy L. Van Dover, Tracey A. Martinson, F. Gerald Plumley (2005). «An obligately photosynthetic bacterial anaerobe from a deep-sea hydrothermal vent». Proceedings of the National Academy of Sciences of the United States of America 102 (26): 9306 -9310. DOI:10.1073/pnas.0503674102. Проверено 2011-02-10.
- ↑ Углекислый газ в атмосфере земли, Б. М. Смирнов, Объединенный институт высоких температур РАН, 126 (11) (1978), Москва
Ссылки
dic.academic.ru
Фотосинтез растений и его особенности
Фотосинтез растений представляет собой сложный физико-биохимический процесс, благодаря которому растения преобразовывают электромагнитную энергию, находящуюся в солнечных лучах, в химическую энергию, используемую в органических соединениях. В основе данного процесса лежит цепочка окислительно-восстановительных химических реакций, в результате которых электроны переносятся от доноров-восстановителей, которыми являются водород и вода, к акцепторам, представляющим собой окислители. При этом образуются углеводы и выделяется O2 при окислении воды.
Фотосинтез растений имеет две последовательных стадии. Первая стадия называется световой (фотохимической). На этом этапе квантовая световая энергия преобразуется в химическую энергию для связей высокоэнергетических соединений, а также в универсальный восстановитель. На второй стадии, имеющей название темновой (метаболической), полученная химическая энергия и универсальный восстановитель проходят цикл для фиксации и восстановления углекислоты, в результате чего образовываются углеводы. Механизм фотосинтеза разделяет световую и темновую стадии не только во времени, но и в пространстве. Световой этап проходит в специальных тилакоидных энергопреобразующих мембранах, в то время как темновые реакции проходят либо в строме хлоропласта, либо в цитоплазме.
Фотосинтез и дыхание растений основывается на поглощении световых квантов, где основную роль играют хлорофиллы, спектр поглощения которых включает видимую область, а также ближние к ней части инфракрасных и ультрафиолетовых областей. Основным пигментом для всех растений, осуществляющих фотосинтез, является хлорофилл а. Зеленые водоросли, мхи и сосудистые растения имеют еще и хлорофилл b, который расширяет поглощаемый световой спектр. Некоторые виды водорослей содержат также хлорофиллы c и d. Помимо хлорофиллов, в процессе поглощения света принимают также участие каротиноиды и фикобилины.
После поглощения света наступает фотохимический этап, в котором принимают участие две фотосистемы типов I и II (ФС1 и ФС2). Каждая из фотосистем состоит из реакционного центра, где происходит разделение зарядов, электротранспортной цепи, где происходит окисление электронов, и набора компонентов, которые выполняют процессы по фотоокислению воды и регенерации реакционного центра. В реакционных центрах квантовая световая энергия преобразуется в химическую, а дальше электроны движутся согласно градиенту электрохимического потенциала, представляющего собой электрон-транспортную цепочку фотосинтеза.
Фотосистема типа II выполняет реакции по фотоокислению воды, в результате чего образуется кислород и протон Н+. Параллельно фотосинтетическому транспорту электронов происходит процесс переноса протонов из хлоропласта во внутритилакоидную область. В результате реакций образуются НАДФН и АТФ, которые являются первичными продуктами фотосинтеза. Далее фотосинтез растений образовывает ферментативные реакции, при которых из углекислот получаются белки, углеводы и жиры. Если же темновой метаболизм имеет неуглеводную направленность, то образуются аминокислоты, органические соединения и белки.
Метаболические процессы по типу фиксации CO2 подразделяются на С3-, С4- и САМ-фотосинтез. При этом углеводы, которые образуются на темновой стадии фотосинтеза, могут отложиться в хлоропластах в виде соединений крахмала, выйти из хлоропластов для образования новых клеток, выполнять функции источника энергии для метаболических реакций.
Фотосинтез растений использует лишь 1-2 процента поглощенной световой энергии. На интенсивность процесса фотосинтеза влияет спектральный состав и интенсивность света, температура, водный режим растения и его минеральное питание, концентрация СО2 и О2, а также другие факторы окружающей среды.
fb.ru
Фотосинтез растений
Фотосинтез - это уникальный физико-химический процесс, осуществляемый на Земле всеми зелеными растениями и некоторыми бактериями и обеспечивающий преобразование электромагнитной энергии солнечных лучей в энергию химических связей различных органических соединений. Основа фотосинтеза — последовательная цепь окислительно-восстановительных реакций, в ходе которых осуществляется перенос электронов от донора — восстановителя (вода, водород) к акцептору — окислителю (СО2, ацетат) с образованием восстановленных соединений (углеводов) и выделением O2, если окисляется вода
Фотосинтез играет ведущую роль в биосферных процессах, приводя в глобальных масштабах к образованию органического вещества из неорганического.
Фотосинтезирующие организмы, используя солнечную энергию в реакциях фотосинтеза, осуществляют связь жизни на Земле со Вселенной и определяют в конечном итоге всю ее сложность и разнообразие. Гетеротрофные организмы — животные, грибы, большинство бактерий, а также бесхлорофилльные растения и водоросли — обязаны своим существованием автотрофным организмам — растениям-фотосинтетикам, создающим на Земле органическое вещество и восполняющим убыль кислорода в атмосфере. Человечество все более осознает очевидную истину, впервые научно обоснованную К.А. Тимирязевым и В.И. Вернадским: экологическое благополучие биосферы и существование самого человечества зависит от состояния растительного покрова нашей планеты.
Процессы, происходящие в листе
Лист осуществляет три важных процесса – фотосинтез, испарение воды и газообмен. В процессе фотосинтеза в листьях из воды и двуокиси углерода под действием солнечных лучей синтезируются органические вещества. Днем, в результате фотосинтеза и дыхания, растение выделяет кислород и двуокись углерода, а ночью – только двуокись углерода, образующуюся при дыхании.
Большинство растений способно синтезировать хлорофилл при слабом освещении. При прямом солнечном освещении хлорофилл синтезируется быстрее. Необходимая для фотосинтеза световая энергия в известных пределах поглощается тем больше, чем меньше затемнен лист. Потому у растений в процессе эволюции выработалась способность поворачивать пластину листа к свету так, чтобы на нее падало больше солнечных лучей. Листья на растении располагаются так, чтобы не притеснять друг друга. Тимирязев доказал, что источником энергии для фотосинтеза служат преимущественно красные лучи спектра. На это указывает спектр поглощения хлорофилла, где наиболее интенсивная полоса поглощения наблюдается в красной, и менее интенсивное – в сине-фиолетовой части.
 Фото: Nat Tarbox
Фото: Nat Tarbox
Интенсивность фотосинтеза также зависит от фазы развития растения. Максимальная интенсивность фотосинтеза наблюдается в фазе цветения. Обычное содержание углекислоты в воздухе составляет 0,03% по объему. Уменьшение содержания углекислоты в воздухе снижает интенсивность фотосинтеза. Повышение содержания углекислоты до 0,5% увеличивает интенсивность фотосинтеза почти пропорционально. Однако при дальнейшем повышении содержания углекислоты, интенсивность фотосинтеза не возрастает, а при 1% - растение страдает.
Растения испаряют или трансперируют очень большое количество воды. Испарение воды является одной из причин восходящего тока. Вследствие испарения воды растением в нем накапливаются минеральные вещества, и происходит полезное для растения понижение температуры во время солнечного нагрева. Растение регулирует процесс испарения воды посредством работы устьиц. Отложение кутикулы или воскового налета на эпидерме, образование его волосков и другие приспособления направлены к сокращению нерегулируемой трансперации.
Процесс фотосинтеза и постоянное протекающее дыхание живых клеток листа требуют газообмена между внутренними тканями листа и атмосферой. В процессе фотосинтеза из атмосферы поглощается ассимилируемый углекислый газ и возвращается в атмосферу кислородом. Применение изотопного метода анализа показало, что кислород, возвращаемый в атмосферу 16O принадлежит воде, а не углекислому газу воздуха, в котором приобладает другой его изотоп - 15О. При дыхании живых клеток (окисление свободным кислородом органических веществ внутри клетки до углекислого газа и воды) необходимо поступление из атмосферы кислорода и возвращение углекислоты. Этот газообмен также в основном осуществляется через устьичный аппарат.
Процесс фотосинтеза состоит из двух последовательных и взаимосвязанных этапов: светового (фотохимического) и темнового (метаболического). На первой стадии происходит преобразование поглощенной фотосинтетическими пигментами энергии квантов света в энергию химических связей высокоэнергетического соединения АТФ и универсального восстановителя НАДФН — собственно первичных продуктов фотосинтеза, или так называемой «ассимиляционной силы». В темновых реакциях фотосинтеза происходит использование образовавшихся на свету АТФ и НАДФН в цикле фиксации углекислоты и ее последующего восстановления до углеводов. У всех фотосинтезирующих организмов фотохимические процессы световой стадии фотосинтеза происходят в особых энергопреобразующих мембранах, называемых тилакоидными, и организованы в так называемую электрон-транспортную цепь. Темновые реакции фотосинтеза осуществляются вне тилакоидных мембран (в цитоплазме у прокариот и в строме хлоропласта у растений). Таким образом, световая и темновая стадии фотосинтеза разделены в пространстве и во времени.
Интенсивность фотосинтеза древесных растений широко варьирует в зависимости от взаимодействия многих внешних и внутренних факторов, причем эти взаимодействия изменяются во времени и различны у разных видов.
Фотосинтетическую способность иногда оценивают по чистому приросту сухой массы. Такие данные имеют особое значение, потому что прирост представляет собой среднее истинное увеличение массы за большой промежуток времени в условиях внешней среды, включающих обычные периодически наступающие стрессы. Некоторые виды покрытосеменных эффективно осуществляют фотосинтез как при низкой, так и при высокой интенсивности света. Многие голосеменные гораздо более продуктивны при высокой освещенности. Сравнение этих двух групп при низкой и высокой интенсивности света часто дает различное представление о фотосинтетической способности с точки зрения накопления питательных веществ. Кроме того, голосеменные часто накапливают некоторое количество сухой массы в период покоя, тогда как листопадные покрытосеменные теряют ее вследствие дыхания. Поэтому голосеменное растение с несколько более низкой интенсивностью фотосинтеза, чем листопадное покрытосеменное во время периода роста, может накапливать в течение года столько же или даже больше общей сухой массы благодаря гораздо большей продолжительности периода фотосинтетической активности.
Первые опыты по фотосинтезу были проведены Джозефом Пристли в 1770-1780-х годах, когда он обратил внимание на "порчу" воздуха в герметичном сосуде горящей свечой (воздух переставал быть способен поддерживать горение, помещённые в него животные задыхались) и "исправление" его растениями. Пристли сделал вывод что растения выделяют кислород, который необходим для дыхания и горения, однако не заметил что для этого растениям нужен свет. Это показал вскоре Ян Ингенхауз. Позже было установлено что помимо выделения кислорода растения поглощают углекислый газ и при участии воды синтезируют на свету органическое вещество. В 1842 Роберт Майер на основании закона сохранения энергии постулировал что растения преобразуют энергию солнечного света в энергию химических связей. В 1877 В. Пфеффер назвал этот процесс фотосинтезом.
biofile.ru
Что такое фотосинтез или почему трава зелёная?
Жизнь человека, как и всего живого на Земле невозможна без дыхания. Мы вдыхаем из воздуха кислород, а выдыхаем углекислый газ. Но почему же кислород не кончается? Оказывается, воздух в атмосфере непрерывно подпитывается кислородом. И происходит это насыщение именно благодаря фотосинтезу.
Фотосинтез - просто и понятно!
Каждый человек обязан понимать, что такое фотосинтез. Для этого совсем не нужно писать сложные формулы, достаточно понять всю важность и волшебство этого процесса.
Главную роль в процессе фотосинтеза играют растения – трава, деревья, кустарники. Именно в листьях растений на протяжении миллионов лет происходит удивительное превращение углекислого газа в кислород, так необходимый для жизни любителям дышать. Попробуем разобрать весь процесс фотосинтеза по порядку.
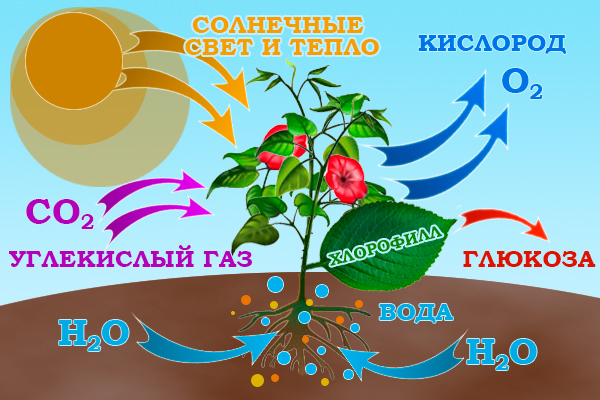
1. Растения берут из почвы воду с растворенными в ней минеральными веществами – азот, фосфор, марганец, калий, различные соли – всего больше 50 различных химических элементов. Это необходимо растениям для питания. Но из земли растения получают лишь 1/5 часть необходимых веществ. Остальные 4/5 они получают из воздуха!
2. Из воздуха растения поглощают углекислый газ. Тот самый углекислый газ, который мы выдыхаем каждую секунду. Углекислым газом растения дышат, как мы с вами дышим кислородом. Но и этого мало.
3. Незаменимый компонент в природной лаборатории - солнечный свет. Солнечные лучи в листьях растений пробуждают необычайную химическую реакцию. Как же это происходит?
4. В листьях растений есть удивительное вещество – хлорофилл. Хлорофилл способен улавливать потоки солнечного света и неутомимо перерабатывать полученные воду, микроэлементы, углекислый газ в органические вещества, необходимые каждому живому существу нашей планеты. В этот момент растения выделяют в атмосферу кислород! Именно эту работу хлорофилла ученые называют сложным словом – фотосинтез.
Презентацию по теме Фотосинтез можно скачать на образовательном портале Учителя.com
Так почему трава зелёная?
Теперь, когда мы знаем, что в клетках растений, содержится хлорофилл, на этот вопрос ответить очень легко. Недаром с древнегреческого языка хлорофилл переводится как «зелёный лист». Для фотосинтеза хлорофилл использует все лучи солнечного света, кроме зеленого. Мы видим траву, листья растений зелеными именно потому, что хлорофилл получается зеленым.

Значение фотосинтеза.
Значение фотосинтеза невозможно переоценить - без фотосинтеза в атмосфере нашей планеты накопилось бы слишком много углекислого газа, большинство живых организмов просто не смогли бы дышать и погибли. Наша Земля превратилась бы в безжизненную планету. Для того чтобы этого не допустить каждому человеку планеты Земля нужно помнить, что мы очень обязаны растениям.
Именно поэтому так важно в городах делать как можно больше парков и зелёных насаждений. Беречь от уничтожения тайгу и джунгли. Или просто посадить дерево рядом с домом. Или не ломать ветки. Только участие каждого человека планеты Земля поможет сохранить жизнь на родной планете.
Но важность фотосинтеза не ограничивается переработкой углекислого газа в кислород. Именно в результате фотосинтеза сформировался озоновый слой в атмосфере, защищающий планету от губительных лучей ультрафиолета. Растения это пища для большинства живых существ на Земле. Пища необходимая и полезная. Питательность растений это тоже заслуга фотосинтеза.
С недавнего времени хлорофилл стали активно использовать в медицине. Люди издавна знали, что больные животные инстинктивно едят зеленые листья, чтобы вылечиться. Ученые выяснили, что хлорофилл сходен с веществом в клетках крови человека и способен творить настоящие чудеса.
ya-uznayu.ru








