
Молекулярная формула фотосинтеза. Формула фотосинтеза у растений
Какая формула фотосинтеза?
Зелёные растения - основа жизни на планете. Это существа, которые одни из первых научились сохранять энергию солнца с помощью синтеза сложных молекул из имеющихся простых (вода, углекислота). Они превращают энергию, падающую на них в форме солнечного света, в энергию, запасенную в углеводах. "Суммарное уравнение фотосинтеза выглядит так:
вода + углекислый газ + свет → углеводы + кислород
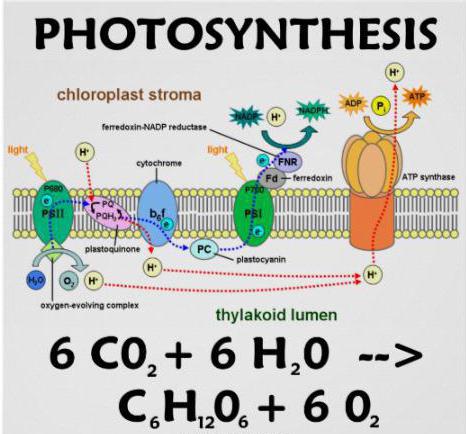
Растения поглощают углекислый газ, образовавшийся при дыхании, и выделяют кислород — продукт жизнедеятельности растений. К тому же, фотосинтез играет важнейшую роль в круговороте углерода в природе. ...Фотосинтез начинается с того, что излучаемые солнцем фотоны попадают в особые пигментные молекулы, находящиеся в листе, — молекулы хлорофилла. Хлорофилл содержится в клетках листа, в мембранах клеточных органелл хлоропластов (именно они придают листу зеленую окраску). Процесс улавливания энергии состоит из двух этапов и осуществляется в раздельных кластерах молекул — эти кластеры принято называть Фотосистемой I и Фотосистемой II. Номера кластеров отражают порядок, в котором эти процессы были открыты, и это одна из забавных научных странностей, поскольку в листе сначала происходят реакции в Фотосистеме II, и лишь затем — в Фотосистеме I.
Когда фотон сталкивается с 250-400 молекулами Фотосистемы II, энергия скачкообразно возрастает и передается на молекулу хлорофилла. В этот момент происходят две химические реакции: молекула хлорофилла теряет два электрона (которые принимает другая молекула, называемая акцептором электронов) и расщепляется молекула воды. Электроны двух атомов водорода, входивших в молекулу воды, возмещают два потерянных хлорофиллом электрона.
После этого высокоэнергетический («быстрый») электрон перекидывают друг другу, как горячую картофелину, собранные в цепочку молекулярные переносчики. При этом часть энергии идет на образование молекулы аденозинтрифосфата (АТФ), одного из основных переносчиков энергии в клетке (см. Биологические молекулы). Тем временем немного другая молекула хлорофилла Фотосистемы I поглощает энергию фотона и отдает электрон другой молекуле-акцептору. Этот электрон замещается в хлорофилле электроном, прибывшим по цепи переносчиков из Фотосистемы II. Энергия электрона из Фотосистемы I и ионы водорода, образовавшиеся ранее при расщеплении молекулы воды, идут на образование НАДФ-Н, другой молекулы-переносчика.
В результате процесса улавливания света энергия двух фотонов запасается в молекулах, используемых клеткой для осуществления реакций, и дополнительно образуется одна молекула кислорода. (Отмечу, что в результате еще одного, значительно менее эффективного процесса с участием одной лишь Фотосистемы I, также образуются молекулы АТФ.) После того как солнечная энергия поглощена и запасена, наступает очередь образования углеводов. Основной механизм синтеза углеводов в растениях был открыт Мелвином Калвином, проделавшим в 1940-е годы серию экспериментов, ставших уже классическими. Калвин и его сотрудники выращивали водоросль в присутствии углекислого газа, содержащего радиоактивный углерод-14. Им удалось установить химические реакции темновой фазы, прерывая фотосинтез на разных стадиях.
Цикл превращения солнечной энергии в углеводы — так называемый цикл Калвина — сходен с циклом Кребса (см. Гликолиз и дыхание): он тоже состоит из серии химических реакций, которые начинаются с соединения входящей молекулы с молекулой-«помощником» с последующей инициацией других химических реакций. Эти реакции приводят к образованию конечного продукта и одновременно воспроизводят молекулу-«помощника», и цикл начинается вновь. В цикле Калвина роль такой молекулы-«помощника» выполняет пятиуглеродный сахар рибулозодифосфат (РДФ). Цикл Калвина начинается с того, что молекулы углекислого газа соединяются с РДФ. За счет энергии солнечного света, запасенной в форме АТФ и НАДФ-H, сначала происходят химические реакции связывания углерода с образованием углеводов, а затем — реакции воссоздания рибулозодифосфата. На шести витках цикла шесть атомов углерода включаются в молекулы предшественников глюкозы и других углеводов. Этот цикл химических реакций будет продолжаться до тех пор, пока поступает энергия. Благодаря этому циклу энергия солнечного света становится доступной живым организмам."
vorum.ru
| Физиология растений Формула фотосинтеза
Две стадии фотосинтеза Процесс фотосинтеза состоит из двух последовательных и взаимосвязанных этапов: светового (фотохимического) и темнового (метаболического). На первой стадии происходит преобразование поглощенной фотосинтетическими пигментами энергии квантов света в энергию химических связей высокоэнергетического соединения АТФ и универсального восстановителя НАДФН — собственно первичных продуктов фотосинтеза, или так называемой «ассимиляционной силы». В темновых реакциях фотосинтеза происходит использование образовавшихся на свету АТФ и НАДФН в цикле фиксации углекислоты и ее последующего восстановления до углеводов. У всех фотосинтезирующих организмов фотохимические процессы световой стадии фотосинтеза происходят в особых энергопреобразующих мембранах, называемых тилакоидными, и организованы в так называемую электрон-транспортную цепь. Темновые реакции фотосинтеза осуществляются вне тилакоидных мембран (в цитоплазме у прокариот и в строме хлоропласта у растений). Таким образом, световая и темновая стадии фотосинтеза разделены в пространстве и во времени.НАДФН – Никотинамидадениндинуклеотидфосфа́т Синтез АТФ и образование НАДФН Одновременно с фотосинтетическим транспортом электронов происходит перенос протонов из стромы хлоропласта во внутритилакоидное пространство — возникает трансмембранный электрохимический градиент ионов водорода (pH-градиент), используемый затем комплексом фермента АТФ-синтазы для синтеза АТФ из АДФ и неорганического фосфата в процессе фотосинтетического фосфорилирования. При нециклическом токе электронов и сопряженном с ним фотофосфорилировании происходит образование восстановителя НАДФН и АТФ. При альтернативных путях переноса электронов — циклическом и псевдоциклическом — образуется только АТФ. Образующиеся в результате световых реакций первичные продукты фотосинтеза — НАДФН и АТФ — используются в ходе дальнейших ферментативных реакций для восстановления углекислоты до углеводов, жиров, белков. При неуглеводной направленности темнового метаболизма, когда преимущественно образуются аминокислоты, белки, органические кислоты, возрастает уровень потребления восстановителя НАДФН.Темновые реакции фотосинтеза (фотосинтетическая фиксация CO 2 ) Метаболические варианты фотосинтетической фиксации CO2 у растений принято классифицировать на С3-, С4- и САМ-фотосинтез. Образующиеся в темновых реакциях углеводы могут откладываться в виде крахмала в хлоропластах; выходить из хлоропластов и использоваться в образовании нового структурного материала клеток; служить источником энергии для различных метаболических процессов; транспортироваться в запасающие органы растения.С 3 -путь фотосинтеза Восстановительный пентозофосфатный цикл фиксации CO2 (С3-путь, или цикл Кальвина), открытый американскими учеными Э. Бенсоном и М. Калвином в 1950-е годы, универсален и обнаруживается практически у всех автотрофных организмов. В этом цикле (рис.5) фиксация СО2 осуществляется на пятиуглеродное соединение рибулезобисфосфат (РуБФ) при участии фермента рибулезобисфосфаткарбоксилазы (РуБФ-карбоксилазы). Первым стабильным продуктом являются две молекулы трехуглеродного соединения 3-фосфоглицериновой кислоты (3-ФГК), восстанавливаемая затем с использованием АТФ и НАДФН до трехуглеводных сахаров, из которых образуется конечный продукт фотосинтеза — шестиуглеродная глюкоза. Субстратом ключевого фермента фотосинтетической фиксации СО2 — РуБФ-карбоксилазы — наряду с СО2 может быть и О2. При взаимодействии РуБФ с кислородом реализуется гликолатный, или С2-путь, известный как фотодыхание. Большинство наземных растений осуществляют фотосинтез по С3-пути. Типичные представители этой группы — горох, фасоль, конские бобы, шпинат, салат, капуста, пшеница, овес, рожь, ячмень, свекла, подсолнечник, тыква, томаты и другие одно- и двудольные растения. С 4 -путь фотосинтеза У некоторых видов растений (в основном тропических и очень небольшого числа видов из умеренных широт) первыми стабильными соединениями при фиксации СО2 являются четырехуглеродные органические кислоты — яблочная и аспарагиновая. Такие растения отличаются видимым отсутствием фотодыхания (или очень низким уровнем), высокой скоростью фиксации СО2 в расчете на единицу поверхности листа, более высокой общей фотосинтетической продуктивностью, быстрой скоростью роста. Функционально и анатомически в ткани их листьев выделяют 2 типа фотосинтезирующих клеток — клетки паренхимной обкладки, окружающие проводящие пучки, и клетки мезофилла. Для всех растений этой группы характерна катализируемая ферментом фосфоенолпируваткарбоксилазой (ФЕП-карбоксилазой) фиксация СО2 на трехуглеродное соединение фосфоенолпируват (ФЕП) с образованием щавелевоуксусной кислоты, которая далее превращается в яблочную (малат) или аспарагиновую кислоту. Эти реакции протекают в цитоплазме клеток мезофилла листа. С4-кислоты затем поступают в клетки обкладки проводящих пучков, где подвергаются декарбоксилированию, а высвободившаяся СО2 фиксируется через цикл Кальвина. Следовательно, у С4-растений фотосинтетический метаболизм углерода пространственно разделен и осуществляется в клетках различного типа, т. е. по «кооперативному механизму», подробно описанному австралийскими исследователями М. Хетчем и К Слэком и советским биохимиком Ю. С. Карпиловым в конце 1960—1970 годов. В соответствии с первичным механизмом декарбоксилирования С4-кислот все С4-растения подразделяются на три группы. НАДФ-малатдегидрогеназные растения осуществляют декарбоксилирование малата с помощью фермента НАДФ-малатдегидрогеназы в хлоропластах клеток обкладки проводящих пучков. Типичные представители этой группы — кукуруза, сахарный тростник, сорго, росичка кроваво-красная и другие злаки. НАД-малатдегидрогеназные растения осуществляют декарбоксилирование малата с помощью митохондриальной НАД-малатдегидрогеназы. Первичным продуктом фиксации углекислоты у них является аспартат. К типичным представителям этой группы принадлежат различные виды амаранта, портулак огородный, просо обыкновенное, бизонья трава, растущая в прериях Северной Америки и др. Фосфоенолпируват-карбоксикиназные растения осуществляют декарбоксилирование аспартата в цитоплазме клеток обкладки проводящих пучков с образованием ФЕП. Типичные представители — некоторые виды проса, хлориса, бутелуа. У суккулентных растений, произрастающих в условиях водного дефицита, фиксация СО2 осуществляется с помощью так называемого САМ-пути (метаболизм кислот по типу растений семейства толстянковых). Первичный продукт фиксации углекислоты (яблочная кислота) образуется у них в темновой период и накапливается в вакуолях клеток листа. В дневное время при закрытых устьицах (которые закрываются для сохранения воды в тканях листа) осуществляется декарбоксилирование этой кислоты, а освобождающаяся СО2 поступает в цикл Кальвина. Возникновение С4- и САМ-путей фотоассимиляции СО2 связано с давлением на высшие наземные растения засушливого климата. С4-растения хорошо адаптированы к высокой интенсивности света, повышенным температурам и засухе. Оптимальная температура для осуществления фотосинтеза у них выше, чем у С3-растений. С4-растения наиболее многочисленны в зонах с высокими температурами. Они более экономно используют воду по сравнению с С3-растениями. В настоящее время известно, что все растения с С4-фотосинтезом — цветковые (из 19 семейств:16 — двудольных и 3 —однодольных). Не обнаружено ни одного семейства, которое бы состояло только из С4-растений.Дыхание растений Представляет процесс, соответствующий дыханию животных. Растение поглощает атмосферный кислород, а последний воздействует на органические соединения их тела таким образом, что в результате появляются вода и углекислота. Вода остается внутри растения, а углекислота выделяется в окружающую среду. При этом происходит уничтожение, трата органического вещества; следовательно, Д. прямо противоположно процессу ассимиляции углерода. До известной степени его можно уподобить окислению и горению вещества. Исходя из крахмала, схематическое уравнение Д. можно представить так: C6h20O5 (крахмал) +6O2 (кислород) = 6CO2 (углекислота) + 5h3O (вода) + 2875 кДж/моль Цикл Кребса. Механизмы регуляции цикла. Энергетическая эффективность процесса, значение В анаэробных условиях пировиноградная кислота (пируват) подвергается дальнейшим превращениям в ходе спиртового, молочнокислого и других видов брожений, при этом НАДH используется для восстановления конечных продуктов брожения, регенерируя в окисленную форму. Последнее обстоятельство поддерживает процесс гликолиза, для которого необходим окисленный НАД + . В присутствии достаточного количества кислорода пируват полностью окисляется до С02 и Н20 в дыхательном цикле, получившем названиецикла Кребса, цикла ди- или трикарбоновых кислот. Все участки этого процесса локализованы в мАТФиксе или во внутренней мембране митохондрий. Последовательность реакций в цикле Кребса. Участие органических кислот в дыхании давно привлекало внимание исследователей. Еще в 1910 г. шведский химик Т. Тунберг показал, что в животных тканях содержатся ферменты, способные отнимать водород от некоторых органических кислот (янтарной, яблочной, лимонной). В 1935 г. А. Сент-Дьердьи в Венгрии установил, что добавление к измельченной мышечной ткани небольших количеств янтарной, фумаровой, яблочной или щавелевоуксуснсй кислот резко активирует поглощение тканью кислорода. Учитывая данные Тунберга и Сент-Дьердьи и исходя из собственных экспериментов по изучению взаимопревращения различных органических кислот и их влияния на дыхание летательной мышцы голубя, английский биохимик Г. А. Кребс в 1937 г. предложил схему последовательности окисления ди- и трикарбоновых кислот до С02 через«цикл лимонной кислоты» да счет отнятия водорода. Этот цикл и был назван его именем. Непосредственно в цикле окисляется не сам пируват, а его производное -- ацетил-СоА. Таким образом, первым этапом на пути окислительного расщепления ПВК является процесс образования активного ацетила в ходе окислительного декарбоксилирования. Окислительное декарбоксилирование пирувата осуществляется при участии пируватдегидрогеназного мультиферментного комплекса. В состав его входят три фермента и пять коферментов. Коферментами служат тиаминпирофосфат (ТПФ) -- фосфорилированное производное витамина Вь липоевая кислота, коэнзим A, ФАД и НАД+. Пируват взаимодействует с ТПФ (декарбоксилазой), при этом отщепляется С02 и образуется гидроксиэтильное производное ТПФ (рис. 3). Последнее вступает в реакцию с окисленной формой липоевой кислоты. Дисульфидная связь липоевой кислоты разрывается и происходит окислительно-восстановительная реакция: гидроксиэтильная группа, присоединенная к одному атому серы, окисляется в ацетильную (при этом возникает высокоэнергетическая тиоэфирная связь), а другой атом серы липоевой кислоты восстанавливается. Образовавшаяся ацетиллипоевая кислота взаимодействует с коэнзимом А, возникают ацетил- СоА и восстановленная форма липоевой кислоты. Водород липоевой кислоты переносится затем на ФАД и далее на НАД + . В результате окислительного декарбоксилирования пирувата образуются ацетил-СоА, С02 и НАДH. Рис. 3. Окислительное декарбоксилирование ПВК Дальнейшее окисление ацетил-СоА осуществляется в ходе циклического процесса. Цикл Кребса начинается с взаимодействия ацетил-СоА с енольной формой щавелевоуксусной кислоты. В этой реакции под действием фермента цитратсинтазы образуется лимонная кислота (2). Следующий этап цикла включает две реакции и катализируется ферментом аконитазой, или аконитатгидратазой (3). В первой реакции в результате дегидратации лимонной кислоты образуется цис-аконитовая. Во второй реакции аконитат гидратируется и синтезируется изолимонная кислота. Изолимонная кислота под действием НАД- или НАДФ-зависимой изоцитратдегидрогеназы (4) окисляется в нестойкое соединение -- щавелевоянтарную кислоту, которая тут же декарбоксилируется с образованием б-кетоглутаровой кислоты (б-оксоглутаровой кислоты). б-Кетоглутарат, подобно пирувату, подвергается реакции окислительного декарбоксилирования. б-Кетоглутаратдегидрогеназный мультиэнзимный комплекс (5) сходен с рассмотренным выше пируватдегидрогеназным комплексом. В ходе реакции окислительного декарбоксилирования б-кетоглутарата выделяется С02, образуются НАДH и сукцинил-СоА. Подобно ацетил-СоА, сукцинил-СоА является высокоэнергетическим тиоэфиром. Однако если в случае с ацетил-СоА энергия тиоэфирной связи расходуется на синтез лимонной кислоты, энергия сукцинил-CoA может трансформироватся в образование фосфатной связи АТФ. При участии сукцинил- СоА-синтетазы (6) из сукцинил-СоА, АДФ и Н3Р04 образуются янтарная кислота (сукцинат), АТФ, регенерирует молекула СоА. АТФ образуется в результате субстратного фосфорилирования. На следующем этапе янтарная кислота окисляется до фумаровой. Реакция катализируется сукцинатдегидрогеназой (7), коферментом которой является ФАД. Фумаровая кислота под действием фумаразы или фумаратгидратазы (8), присоединяя Н20, превращается в яблочную кислоту (малат). И, наконец, на последнем этапе цикла яблочная кислота с помощью НАД- зависимой малатдегидрогеназы (9) окисляется в щавелевоуксусную. ЩУК, которая самопроизвольно переходит в енольную форму, реагирует с очередной молекулой ацетил-СоА и цикл повторяется снова. Следует отметить, что большинство реакций цикла обратимы, однако ход цикла в целом практически необратим. Причина этого в том, что в цикле есть две сильно экзергонические реакции -- цитратсинтазная и сукцинил-СоА-синтетазная. На протяжении одного оборота цикла при окислении пирувата происходит выделение трех молекул С02, включение трех молекул Н2О и удаление пяти пар атомов водорода. Роль Н2О в цикле Кребса подтверждает правильность уравнения Палладина, который постулировал, что дыхание идет с участием Н2О, кислород которой включается в окисляемый субстрат, а водород с помощью «дыхательных пигментов» (по современным представлениям -- коферментов дегидрогеназ) переносится на кислород . Энергетический выход цикла Кребса, его связь с азотным обменом. Цикл Кребса. играет чрезвычайно важную роль в обмене веществ растительного организма. Он служит конечным этапом окисления не только углеводов, но также белков, жиров и других соединений. В ходе реакций цикла освобождается основное количество энергии, содержащейся в окисляемом субстрате, причем большая часть этой энергии не теряется для организма, а утилизируется при образовании высокоэнергетических конечных фосфатных связей АТФ. Каков же энергетический выход цикла Кребса? В ходе окисления пирувата имеют место 5 дегидрирований, при этом получаются 3НАДH, НАДФH (в случае изоцитратдегидрогеназы) и ФАДh3. Окисление каждой молекулы НАДH (НАДФH) при участии компонентов электронтранспортной цепи митохондрий дает по 3 молекулы АТФ, а окисление ФАДh3 -- 2АТФ. Таким образом при полном окислении пирувата образуются 14 молекул АТФ. Кроме того, 1 молекула АТФ синтезируется ; в цикле Кребса в ходе субстратного фосфорилирования. Следовательно, при окислении одной молекулы пирувата может образоваться 15 молекул АТФ. А поскольку в процессе гликолиза из молекулы глюкозы возникают две молекулы пирувата, их окисление даст 30 молекул АТФ. Итак, при окислении глюкозы в процессе дыхания при функционировании гликолиза и цикла Кребса в общей сложности образуются 38 молекул АТФ (8 АТФ связаны с глико- лизом). Если принять, что энергия третьей сложноэфирнои фосфатной связи АТФ равняется 41,87 кДж/моль (10 ккал/моль), то энергетический выход гликолитического пути аэробного дыхания составляет 1591 кДж/моль (380 ккал/моль). Значение цикла Кребса не ограничивается его вкладом в энергетический обмен клетки. Не менее важную роль играет то обстоятельство, что многие промежуточные продукты цикла используются при синтезе различных соединений. Из кетокислот в ходе реакций переаминирования образуются аминокислоты. Для синтеза липидов, полиизопренов, углеводов и ряда других соединений используется ацетил-СоА. Регуляция цикла Кребса. Дальнейшее использование образующегося из пирувата ацетил-СоА зависит от энергетического состояния клетки. При малой энергетической потребности клетки дыхательным контролем тормозится работа дыхательной цепи, а следовательно, реакций ЦТК и образования интермедиатов цикла, в том числе оксалоацетата, вовлекающего ацетил-СоА в цикл Кребса. Это приводит к большему использованию ацетил-СоА в синтетических процессах, которые также потребляют энергию. Особенностью регуляции ЦТК является зависимость всех четырех дегидрогеназ цикла (изоцитратдегидрогеназы, б-кетоглутаратдегидрогеназы, сукцинатдегидрогеназы, малатдегидрогеназы) от отношения [НАДH]/[НАД+]. Активность цитратсинтазы тормозится высокой концентрацией АТФ и собственным продуктом -- цитратом. Изоцитратдегидрогеназа ингибируется НАДH и активируется цитратом. б-Кето- глутаратдегидрогеназа подавляется продуктом реакции -- сукцинил-СоА и активируется аденилатами. Окисление сукцината сукцинатдегидрогеназой тормозится оксалоацетатом и ускоряется АТФ, АДФ и восстановленным убихиноном (Qh3). Наконец, малатдегидрогеназа ингибируется оксалоацетатом и у ряда объектов -- высоким уровнем АТФ. Однако степень участия величины энергетического заряда, или уровня адениновых нуклеотидов, в регуляции активности цикла Кребса у растений до конца не выяснена. Регулирующую роль может играть также альтернативный путь транспорта электронов в растительных митохондриях. В условиях высокого содержания АТФ, когда активность основной дыхательной цепи снижена, окисление субстратов через альтернативную оксидазу (без образования АТФ) продолжается, что поддерживает на низком уровне отношение НАДH/НАД+ и снижает уровень АТФ. Все это позволяет циклу Кребса функционировать.
|

otnos.ru
Молекулярная формула фотосинтеза — Циклопедия
Молекулярная формула фотосинтеза — это один из способов графического изображения процесса фотосинтеза.
Это формула была разработана французским биохимиком Мишелем Гриньяром в 1881 году. Она используется для обозначения преобразования световой энергии Солнца в энергию химических связей.Молекулярная формула фотосинтеза:
I) Световая или Светозависимая фаза: 1) n(L)+[Ch(ē)]⇌ ē↑+[Ch](-)L(+)n + Q 2)HOH → H(+)+ OH(-) - реакция идёт под воздействием света(hv) 3)H(+)+ē+OH 4)OH(-)=ē+OH 5)4ОН → О2 + 2Н2О 6)ē↑+[Ch](-)+ē⇌n(L){ա}(+) + [Ch(ē)](-) II) Темновая фаза: 6СО2 + 24 Н2О --> C6h22O6 + 6 h3O - общее уравнение.Условные обозначения:
n(L) - кванты света [Ch(ē)] - подвижные электроны молекул хлорофилла ē↑ - возбуждённое состояние электронов Q - выделение тепла (hv) - вещества-переносчики электронов OH(-) - Гидроксид-ион H(+) - ион водорода (+) и (-) - обозначение зарядов: положительного и отрицательногоЭтот метод соответствует тем процессам, которые происходят в особых органоидах Зелёных растений — хлоропластах — пигмента хлорофилла они осуществляют фотосинтез. Процесс фотосинтеза по формуле Гриньяра:: Кванты света взаимодействуют с молекулами хлорофилла, в результате чего эти молекулы(точнее, их электроны) переходят в более богатое энергией «возбужденное состояние». Избыточная энергия части возбуждённых молекул преобразуется в теплоту или испускаеться в виде света(-ē↑). Другая её часть передаётся ионам водорода, всегда находящимся в водном растворе вследствие диссоциации воды . Образовавшиеся атомы водорода непрочно соединяются с молекулами — переносчиками водорода. Ионы гидроксила отдают свои электроны другим молекулам и превращаются в свободные радикалы OH. Радикалы OH взаимодействую друг с другом, в результате чего образуются вода и молекулярный кислород в соответствии с уравнением: 4ОН → О2 + 2Н2О Отсюда следует, что источником свободного кислорода служит вода. Этот процесс(разложение воды, под действием света) называется фотолизом воды. Кроме фотолиза воды энергия возбуждённых светом электронов хлорофилла используется для синтеза АТФ их АДФ и фосфата без участия кислорода — этот процесс называется нуклеодеоксигенезация и соответствует формуле: ē↑+[Ch](-)+ē⇌n(L){ա}(+) + [Ch(ē)](-) Накопленная в результате светозависимых реакций энергия и атомы водорода, образованные при фотолизе воды, используется для синтеза углеводов из CO2: 6СО2 + 24 Н2О --> C6h22O6 + 6 h3O — это процесс темновой фазы.
- M.J .Grignard . "Lumière dans la transformation de substances biochimiques
- Michel Grignard Jaurès. «Des méthodes pour déterminer les processus, en utilisant un photon»
- Boussingault, J.B. Mémoires de J.-B. Boussingault. París: Chamerot et Renouard, 1892—1903.
- McCosh, F.W.J. Boussingault: Chemist and Agriculturist. Dordrecht: D. Reidel, 1984 ISBN 90-277-1682-X.
- Лауреаты Нобелевской премии: Энциклопедия. Пер. с англ. — М.: Прогресс, 1992.
cyclowiki.org
что такое? Стадии фотосинтеза. Условия фотосинтеза
Вы когда-нибудь задумывались, сколько на планете живых организмов?! И ведь всем им нужно вдыхать кислород, чтобы выработать энергию и выдохнуть углекислый газ. Именно углекислый газ - основная причина такого явления, как духота в помещении. Она имеет место тогда, когда в нем находится много людей, а комната продолжительное время не проветривается. Кроме этого, ядовитыми веществами наполняют воздух производственные объекты, частный автомобильный и общественный транспорт.
С учетом вышесказанного возникает вполне логичный вопрос: как же мы тогда еще не задохнулись, если все живое является источником ядовитого углекислого газа? Спасителем всех живых существ в данной ситуации выступает фотосинтез. Что такое представляет собой этот процесс и в чем его необходимость?

Его результат – регулировка баланса углекислого газа и насыщение воздуха кислородом. Известен такой процесс только представителям мира флоры, то есть растениям, поскольку происходит только в их клетках.
Сам по себе фотосинтез — это чрезвычайно сложная процедура, зависящая от определенных условий и происходящий в несколько этапов.
Определение понятия
Согласно научному определению, органические вещества в процессе фотосинтеза преобразуются в органические на клеточном уровне у автотрофных организмов за счет воздействия света солнца.

Если сказать более понятным языком, фотосинтез представляет собой процесс, при котором происходит следующее:
- Растение насыщается влагой. Источником влаги может быть вода из грунта либо влажный тропический воздух.
- Происходит реакция хлорофилла (специального вещества, которое содержится в растении) на воздействие солнечной энергии.
- Образование необходимой представителям флоры пищи, которую самостоятельно добыть они не в состоянии гетеротрофным способом, а сами являются ее производителем. Иначе говоря, растения питаются тем, что сами производят. Это и есть результат фотосинтеза.
Этап первый
Практически каждое растение содержит зеленое вещество, благодаря которому оно может поглощать свет. Это вещество является не чем иным, как хлорофиллом. Его местонахождение – хлоропласты. А вот хлоропласты располагаются в стеблевой части растения и его плодах. Но особенно распространен в природе фотосинтез листа. Поскольку последний довольно прост по своей структуре и имеет относительно большую поверхность, а значит, объемы энергии, необходимой для протекания процесса-спасителя будут гораздо больше.

Когда свет поглощен хлорофиллом, последний пребывает в состоянии возбуждения и свои энергетические посылы передает другим органическим молекулам растения. Наибольшее количество такой энергии достается участникам процесса фотосинтеза.
Этап второй
Образование фотосинтеза на втором этапе не требует обязательного участия света. Он состоит в формировании химических связей с использованием ядовитого углекислого газа, образующегося из воздушных масс и воды. Также происходит синтез множества веществ, которые обеспечивают жизнедеятельность представителей флоры. Таковыми являются крахмал, глюкоза.
У растений такие органические элементы выступают источником питания для отдельных частей растения, одновременно обеспечивая нормальное протекание процессов жизнедеятельности. Такие вещества получают и представители фауны, которые употребляют растения в пищу. Человеческий же организм насыщается этими веществами через пищу, которая входит в ежедневный рацион.
Что? Где? Когда?
Чтобы органические вещества превратились в органические, нужно обеспечить соответствующие условия фотосинтеза. Для рассматриваемого процесса необходим в первую очередь свет. Речь идет и об искусственном, и о солнечном свете. На природе обычно деятельность растений характеризуется интенсивностью весной и летом, то есть тогда, когда существует необходимость в поступлении большого количества солнечной энергии. Чего не скажешь об осенней поре, когда света все меньше, день все короче. В результате листва желтеет, а потом и вовсе опадает. Но как только заблестят первые весенние лучики солнца, взойдет зеленая травка, тут же возобновят свою деятельность хлорофиллы, и начнется активная выработка кислорода и других питательных веществ, которые носят жизненно важный характер.
Условия фотосинтеза включают не только наличие освещенности. Влаги тоже должно быть достаточно. Ведь растение сперва поглощает влагу, а потом начинается реакция с участием солнечной энергии. Результатом такого процесса и являются продукты питания растений.
Только при наличии зеленого вещества происходит фотосинтез. Что такое хлорофиллы, мы уже рассказывали выше. Они выступают неким проводником между светом или солнечной энергией и самим растением, обеспечивая надлежащее протекание их жизни и деятельности. Зеленые вещества обладают способностью поглощения множества солнечных лучей.
Немалую роль играет и кислород. Чтобы процесс фотосинтеза прошел успешно, растениям нужно его много, поскольку в его составе содержится всего 0,03% углекислой кислоты. Значит, из 20 000 м3 воздуха можно получить 6 м3 кислоты. Именно последнее вещество - основной исходный материал для глюкозы, которая, в свою очередь, является веществом, необходимым для жизнедеятельности.

Существует две стадии фотосинтеза. Первая называется световая, вторая – темновая.
В чем механизм протекания световой стадии
Световая стадия фотосинтеза имеет еще одно название – фотохимическая. Основными участниками на этом этапе являются:
- энергия солнца;
- разнообразные пигменты.
С первой составляющей все понятно, это солнечный свет. А вот что представляют собой пигменты, знает не каждый. Они бывают зелеными, желтыми, красными или синими. К зеленым относятся хлорофиллы групп «А» и «Б», к желтым и красным/синим – фикобилины соответственно. Фотохимическую активность среди участников этой стадии процесса проявляют только хлорофиллы «А». Остальным принадлежит дополняющая роль, суть которой – сбор квантов света и их транспортировка к фотохимическому центру.
Поскольку хлорофилл наделен способностью эффективного поглощения солнечной энергии с определенной длиной волны, были идентифицированы следующие фотохимические системы:
- Фотохимический центр 1 (зеленые вещества группы «А») – в состав включен пигмент 700, поглощающий световые лучи, длина которых приблизительно 700 нм. Этому пигменту принадлежит основополагающая роль в создании продуктов световой стадии фотосинтеза.
- Фотохимический центр 2 (зеленые вещества группы «Б») – в состав включен пигмент 680, поглощающий световые лучи, длина которых 680 нм. Ему принадлежит роль второго плана, заключающаяся в функции восполнении электронов, утраченных фотохимическим центром 1. Достигается благодаря гидролизу жидкости.
На 350– 400 молекул пигментов, которые концентрируют в себе потоки света в фотосистеме 1 и 2 приходится только одна молекула пигмента, являющегося активным фотохимически — хлорофилла группы «А».
Что происходит?
1. Световая энергия, поглощаемая растением, оказывает воздействие на содержащийся в нем пигмент 700, который переходит из обычного состояния в состояние возбуждения. Пигмент теряет электрон, в результате чего образуется так называемая электронная дыра. Далее молекула пигмента, которая утратила электрон, может выступать в качестве его акцептора, то есть стороной, принимающей электрон, и возвращать свою форму.
2. Процесс разложения жидкости в фотохимическом центре светопоглощающего пигмента 680 фотосистемы 2. При разложении воды образуются электроны, которые изначально акцептируются таким веществом, как цитохром С550, и обозначаются буквой Q. Затем от цитохрома электроны попадают в цепь переносчиков и транспортируются в фотохимический центр 1 для восполнения электронной дыры, которая стала результатом проникновения квантов света и восстановительного процесса пигмента 700.
Бывают случаи, когда такая молекула получает обратно электрон, идентичный прежнему. Это приведет к выделению энергии света в виде тепла. Но практически всегда электрон, имеющий отрицательный заряд, соединяется со специальными железосерными белками и переносится по одной из цепей к пигменту 700 либо попадает в другую цепь переносчиков и воссоединяется с постоянным акцептором.
При первом варианте имеет место циклическая транспортировка электрона замкнутого типа, при втором – нециклическая.
Оба процесса попадают на первой стадии фотосинтеза под катализацию одной и той же цепью переносчиков электронов. Но стоит отметить, что при фотофосфорилировании циклического типа начальной и одновременно конечной точкой транспортировки является хролофилла, в то время когда нециклическая транспортировка подразумевает переход зеленого вещества группы «Б» к хлорофиллу «А».
Особенности циклической транспортировки
Фосфорилирование циклическое называется еще фотосинтетическим. В результате такого процесса образуются молекулы АТФ. В основе такой транспортировки лежит возвращение через несколько последовательных этапов электронов в возбужденном состоянии на пигмент 700, в результате чего высвобождается энергия, принимающая участие в работе фосфорилирующей ферментной системы с целью дальнейшей аккумуляции в фосфатных связях АТФ. То есть энергия не рассеивается.
Фосфорилирование циклическое представляет собой первичную реакцию фотосинтеза, в основе которой технология образования химической энергии на мембранных поверхностях тилактоидов хлоропластов благодаря использованию энергии солнечных лучей.
Без фотосинтетического фосфорилирования реакции ассимиляции в темновой фазе фотосинтеза невозможны.
Нюансы транспортировки нециклического типа
Процесс заключается в восстановлении НАДФ+ и образовании НАДФ*Н. Механизм основан на передаче электрона ферредоксину, его восстановительной реакцией и последующим переходом к НАДФ+ с дальнейшим восстановлением до НАДФ*Н.
В итоге электроны, которые потеряли пигмент 700, восполняются благодаря электронам воды, которая разлагается под световыми лучами в фотосистеме 2.
Нециклический путь электронов, протекание которого также подразумевает световой фотосинтез, осуществляется посредством взаимодействия обеих фотосистем между собой, связывает их электронно-транспортные цепи. Световая энергия направляет поток электронов обратно. При транспортировке от фотохимического центра 1 к центру 2 электроны теряют часть своей энергии в связи с аккумуляцией в качестве протонного потенциала на мембранной поверхности тилактоидов.
В темновой фазе фотосинтеза процесс создания потенциала протонного типа в транспортировочной цепи электрона и его эксплуатация для образования АТФ в хлоропластах практически полностью идентичен с таким же процессом в митохондриях. Но особенности все же присутствуют. Тилактоидами в данной ситуации выступают митохондрии вывернутые на изнаночную сторону. Это и является главной причиной того, что электроны и протоны движутся через мембрану в противоположном направлении относительно течения переноса в мембране митохондриальной. Электроны транспортируются к наружной стороне, а протоны накапливаются во внутренней части матрикса тилактоидного. Последний принимает только положительный заряд, а наружная мембрана тилактоида – отрицательный. Из этого следует, что путь градиента протонного типа противоположен его пути в митохондриях.
Следующей особенностью можно назвать большой уровень рН в потенциале протонов.
Третьей особенностью является наличие в тилактоидной цепи только двух участков сопряжения и как следствие соотношение молекулы АТФ к протонам равняется 1:3.
Вывод
На первой стадии фотосинтез является взаимодействием световой энергии (искусственной и неискусственной) с растением. Реагируют на лучи зеленые вещества – хлорофиллы, большая часть которых содержится в листьях.

Образование АТФ и НАДФ*Н - результат такой реакции. Эти продукты необходимы для протекания темновых реакций. Следовательно, световая стадия – обязательный процесс, без которого не состоится вторая стадия - темновая.
Темновая стадия: суть и особенности
Темновой фотосинтез и его реакции представляют собой процедуру углекислоты в вещества органического происхождения с получением углеводов. Осуществление таких реакций происходит в строме хлоропласта и активное участие в них принимают продукты первой стадии фотосинтеза – световой.
В основе механизма темновой стадии фотосинтеза положен процесс ассимиляции диоксида углерода (еще называется фотохимическим карбоксилированием, циклом Кальвина), который характеризуется цикличностью. Состоит из трех фаз:
- Карбоксилирование – присоединение СО2.
- Восстановительная фаза.
- Фаза регенерации рибулозодифосфата.
Рибулофосфат – сахар с пятью атомами углерода - поддается процедуре фосфорилирования за счет АТФ, в результате чего образуется рибулозодифосфат, который далее подвергается карбоксилированию благодаря соединению с СО2 продуктом с шестью углеродами, которые мгновенно разлагаются при взаимодействии с молекулой воды, создавая две молекулярные частицы кислоты фосфоглицериновой. Потом эта кислота проходит курс полного восстановления при осуществлении ферментативной реакции, для которой обязательно присутствие АТФ и НАДФ с образованием сахара с тремя углеродами – трехуглеродного сахара, триоза или альдегида фосфоглицеринового. Когда два таких триоза конденсируются, получается молекула гексозы, которая может стать составной частью молекулы крахмала и отлаживаться про запас.
Эта фаза завершается тем, что во время процесса фотосинтеза происходит поглощение одной молекулы СО2 и использование трех молекул АТФ и четырех атомов Н. Гексозофосфат поддается реакциям пентозофосфатного цикла, в результате чего происходит регенерация рибулозофосфата, который может вновь воссоединиться с другой молекулой углеродной кислоты.
Реакции карбоксилирования, восстановления, регенерации нельзя назвать специфическими исключительно для клетки, в которой протекает фотосинтез. Что такое «однородное» протекание процессов, тоже не скажешь, поскольку отличие все же существует – при восстановительном процессе используется НАДФ*Н, а не НАД*Н.
Присоединение СО2 рибулозодифосфатом подвергается катализации, которую обеспечивает рибулозодифосфаткарбоксилаза. Продуктом реакции является 3-фосфоглицерат, восстанавливающийся за счет НАДФ*Н2 и АТФ до глицеральдегид-3-фосфата. Процесс восстановления катализируется глицеральдегидом-3-фосфат-дегидрогеназом. Последний легко превращается в дигидроксиацетонфосфат. Происходит образование фруктозобисфосфата. Часть его молекул принимает участие в регенерирующем процессе рибулозодифосфата, замыкая цикл, а вторая часть эксплуатируется для создания запасов углеводов в клетках фотосинтеза, то есть имеет место фотосинтез углеводов.
Энергия света необходима для фосфорилирования и синтеза веществ органического происхождения, а энергия окисления органических веществ необходима для окислительного фосфорилирования. Именно поэтому растительность обеспечивает жизнь животным и иным организмам, которые относятся к гетеротрофным.

Фотосинтез в клетке растений происходит именно таким образом. Его продуктом являются углеводы, необходимые для создания углеродных скелетов множества веществ представителей мира флоры, которые имеют органическое происхождение.
Вещества азоторганического типа усваиваются в фотосинтезирующих организмах за счет восстановления нитратов неорганических, а сера – за счет восстановления сульфатов до сульфгидрильных групп аминокислот. Обеспечивает образование белков, нуклеиновых кислот, липидов, углеводов, кофакторов именно фотосинтез. Что такое «ассорти» веществ жизненно важно для растений, уже подчеркивалось, а вот о продуктах вторичного синтеза, которые являются ценными лекарственными веществами (флавоноиды, алкалоиды, терпены, полифенолы, стероиды, оргкислоты и другие), не было сказано ни слова. Следовательно, без преувеличения можно сказать, что фотосинтез – залог жизни растений, животных и людей.
fb.ru
Фотосинтез
Фотосинтез — синтез органических соединений из неорганических за счет энергии света (hv). Суммарное уравнение фотосинтеза:
6CO2 + 6h3O → C6h22O6 + 6O2
Фотосинтез протекает при участии фотосинтезирующих пигментов, обладающих уникальным свойством преобразования энергии солнечного света в энергию химической связи в виде АТФ. Фотосинтезирующие пигменты представляют собой белковоподобные вещества. Наиболее важный из них — пигмент хлорофилл. У эукариот фотосинтезирующие пигменты встроены во внутреннюю мембрану пластид, у прокариот — во впячивания цитоплазматической мембраны.
Строение хлоропласта очень похоже на строение митохондрии. Во внутренней мембране тилакоидов граны содержатся фотосинтетические пигменты, а также белки цепи переноса электронов и молекулы фермента АТФ-синтетазы.
Процесс фотосинтеза состоит из двух фаз: световой и темновой.
Световая фаза фотосинтеза протекает только на свету в мембране тилакоидов граны. В этой фазе происходит поглощение хлорофиллом квантов света, образование молекулы АТФ и фотолиз воды.
Под действием кванта света (hv) хлорофилл теряет электроны, переходя в возбужденное состояние:
Хл → Хл + e—
Эти электроны передаются переносчиками на наружную, т.е. обращенную к матриксу поверхность мембраны тилакоидов, где накапливаются.
Одновременно внутри тилакоидов происходит фотолиз воды, т.е. ее разложение под действием света
2h3O → O2 +4H+ + 4e—
Образующиеся электроны передаются переносчиками к молекулам хлорофилла и восстанавливают их: молекулы хлорофилла возвращаются в стабильное состояние.
Протоны водорода, образовавшиеся при фотолизе воды, накапливаются внутри тилакоида, создавая Н+-резервуар. В результате внутренняя поверхность мембраны тилакоида заряжается положительно (за счет Н+), а наружная — отрицательно (за счет e—). По мере накопления по обе стороны мембраны противоположно заряженных частиц нарастает разность потенциалов. При достижении критической величины разности потенциалов сила электрического поля начинает проталкивать протоны через канал АТФ-синтетазы. Выделяющаяся при этом энергия используется для фосфорилирования молекул АДФ:
АДФ + Ф → АТФ
Образование АТФ в процессе фотосинтеза под действием энергии света называются фотофосфорилированием.
Ионы водорода, оказавшись на наружной поверхности мембраны тилакоида, встречаются там с электронами и образуют атомарный водород, который связывается с молекулой-переносчиком водорода НАДФ (никотинамидадениндинуклеотидфосфат):
2H+ + 4e— + НАДФ+ → НАДФ • h3
Таким образом, во время световой фазы фотосинтеза происходят три процесса: образование кислорода вследствие разложения воды, синтез АТФ, образование атомов водорода в форме НАДФ • h3. Кислород диффундирует в атмосферу, АТФ и НАДФ • h3 участвуют в процессах темновой фазы.
Темновая фаза фотосинтеза протекает в матриксе хлоропласта как на свету, так и в темноте и представляет собой ряд последовательных преобразований CO2, поступающего из воздуха, в цикле Кальвина. Осуществляются реакции темновой фазы за счет энергии АТФ. В цикле Кальвина CO2 связывается с водородом из НАДФ • h3 с образованием глюкозы.
В процессе фотосинтеза кроме моносахаридов (глюкоза и др.) синтезируются мономеры других органических соединений — аминокислоты, глицерин и жирные кислоты. Таким образом, благодаря фотосинтезу растения обеспечивают себя и все живое на Земле необходимыми органическими веществами и кислородом.
Сравнительная характеристика фотосинтеза и дыхания эукариот приведена в таблице:
| Уравнение реакции | 6CO2 + 6h3O + Энергия света → C6h22O6 + 6O2 | C6h22O6 + 6O2 → 6h3O + Энергия (АТФ) |
| Исходные вещества | Углекислый газ, вода | Органические вещества, кислород |
| Продукты реакции | Органические вещества, кислород | Углекислый газ, вода |
| Значение в круговороте веществ | Синтез органических веществ из неорганических | Разложение органических веществ до неорганических |
| Превращение энергии | Превращение энергии света в энергию химических связей органических веществ | Превращение энергии химических связей органических веществ в энергию макроэргических связей АТФ |
| Важнейшие этапы | Световая и темновая фаза (включая цикл Кальвина) | Неполное окисление (гликолиз) и полное окисление (включая цикл Кребса) |
| Место протекания процесса | Хлоропласта | Гиалоплазма (неполное окисление) и митохондрии (полное окисление) |
jbio.ru
Темновая фаза фотосинтеза | Student Guru
 Posted in Биология
Posted in Биология  Tags: Фотосинтез
Tags: Фотосинтез За световой фазой следует темновая фаза фотосинтеза, во время которой происходит синтез моносахаридов (глюкозы) из углекислого газа с затратой энергии АТФ и восстановительных эквивалентов (НАДФН). Синтез глюкозы является результатом целого ряда последовательных ферментативных реакций, которые назвали циклом Кальвина. Как было сказано ранее в разделе «Кислородный этап энергетического обмена», в цикле Кребса в митохондриях от молекул органических кислот отрываются молекулы углекислого газа (CO2), промежуточные продукты цикла последовательно окисляются, отрываемые от них атомы водорода присоединяются к НАД+ (т.е. образуется НАДН). В цикле Кальвина происходит все наоборот, к молекулам субстрата присоединяется молекулы углекислого газа (СО2), и они восстанавливаются за счет НАДФН (т.е образуется НАДФ+).
Началом синтеза глюкозы является присоединение молекулы углекислого газа к молекуле пятиуглеродного сахара – рибулозо-1,5-бисфосфата. При этом образуется шестиуглеродная молекула, которая сразу же распадается на две молекулы трехуглеродной фосфоглицериновой кислоты, которая восстанавливается до трехуглеродных сахаров с затратой АТФ и НАДФН. В результате их дальнейших перестроек и конденсаций образуются рибулозомонофосфат и глюкоза — конечный продукт фотосинтеза. Рибулозомонофосфат фосфорилируется АТФ до рибулозобисфостата, который вновь вступает в цикл Кальвина. На образование одной молекулы глюкозы затрачивается 18 молекул АТФ и 12 молекул НАДФН, накопленных в процессе световой фазы фотосинтеза. Следовательно, для темновой фазы фотосинтеза можно представить следующее общее уравнение:
6СО2 + 12НАДФН + 12Н+ + 18АТФ —> С6Н12О6 + 6Н2О + 12НАДФ+ + 18АДФ + 18Фн
Даже если учесть частичные потери энергии на различных стадиях темновой фазы, общий КПД фотосинтеза остается очень высоким и составляет приблизительно 60%.
У некоторых растений (например, сахарного тростника или кукурузы) процесс фотосинтеза идет вначале не через трехуглеродные, а через четырехуглеродные соединения. Эти растения называются С4-растениями. В отличие от С3-растений им характерен быстрый рост и высокая эффективность фотосинтеза, который протекает даже при очень низких концентрациях углекислого газа. В этом случае углекислый газ присоединяется не к рибулозобисфосфату, а к одному из промежуточных продуктов гликолиза – фосфоенолпирувату. В результате образуются четырехуглеродные яблочная или аспарагиновая кислоты, которые диффундируют в клетки обкладки сосудистых пучков, где от них отщепляется СО2, вступая в цикл Кальвина. В этих клетках слабо выражено фотодыхание, связанное с окислением рибулозобисфосфата кислородом, поэтому энергозатраты на фотосинтез резко снижаются (на 50%). В последние годы благодаря необычайно высокой биологической продуктивности С4-растения привлекают внимание ученых как потенциальный источник органического сырья.
Перейти к оглавлению.
You can leave a response, or trackback from your own site.www.studentguru.ru
Удивительный химический процесс фотосинтеза
Фотосинтез – это химический процесс, о котором слышал каждый из нас. Однако мало кто из людей задумывались, насколько важен этот процесс для жизни на Земле и как загадочно он протекает.
Если перевести формулу фотосинтеза в слова, она будет звучать так: вода, углекислый газ и солнечный свет производят глюкозу и кислород.
Реакция фотосинтеза может показаться очень простой, но она чрезвычайно сложна и до сих пор наука не смогла разгадать ее механизм: фотосинтез протекает лишь в одном месте: в растениях, которые производят глюкозу для всех живых существ на Земле.
Травоядные животные поедают растения, а всеядные животные поедают растения и/или других животных. Человек не составляет исключение: мы получаем энергию из пищи, которую едим и которая образуется из того же источника. Каждое съеденное яблоко, картофелина, шоколад или кусок мяса снабжают нас энергией, идущей от Солнца.
Иными словами, растения постоянно очищают атмосферу, которая непрерывно загрязняется дышащими воздухом людьми и животными, получающими энергию в результате сгорания кислорода и выделения углекислого газа. Если бы растения не выделяли кислород, мы бы быстро использовали весь кислород атмосферы, и это бы означало конец всему. Но растения – эти удивительные лаборатории жизни – миллиарды лет беспрерывно пополняют запасы кислорода в атмосфере.
Без фотосинтеза невозможна жизнь растений, а без растений не существовало бы ни человека, ни животных. Эта удивительная химическая реакция, которая наука не может воспроизвести даже в суперсовременных лабораторных условиях, осуществляется повсюду в природе: в каждой травинке, на которую вы наступаете, в каждом дереве, которое вы, возможно, даже не замечаете. Эта реакция протекала и в овощах, находящихся сейчас на вашем обеденном столе. Фотосинтез представляет собой один из основополагающих процессов жизни.
Фотосинтез совершается в молекулах хлорофилла... Механизм фотосинтеза запускается при поглощении солнечного света молекулами хлорофилла. Но, чтобы это случилось, свет должен быть необходимого цвета. Если свет будет не того цвета, ничего не получится.
Хорошее сравнение – телевизор. Чтобы телевизор принимал нужный канал, он должен быть настроен на этот канал. Если вы не так настроите телевизор, изображения не будет. То же самое с фотосинтезом. Если следовать нашему примеру, Солнце действует как передатчик, а молекула хлорофилла как принимающий телевизор. Если эта молекула и Солнце не настроены правильно по отношению друг к другу в плане цвета, фотосинтеза не произойдет. Однако оказывается, что цвет солнечного света всегда выбран абсолютно правильно.
К примеру, наша планета по подсчетам возникла 4.6 миллиардов лет тому назад. Вокруг нее возникает некая атмосфера. Тогда как жизнь на земле существует всего 3.5 миллиардов лет. 3.5 миллиардов лет назад на земле появляются первые цианобактерии – одноклеточные существа, способные вырабатывать кислород и производить процесс фотосинтеза, жизненно необходимый для всего живого. Именно тогда, с выработки кислорода, и начинается жизнь. И заметьте, эти сине-зеленые водоросли-бактерии появляются на земле внезапно. Казалось бы примитивнейшие одноклеточные бактерии производят сложнейший процесс фотосинтеза, неподвластный синтезу человека даже на современном уровне.
Причем за 3.5 миллиардов лет в механизме фотосинтеза и уровне производимого ими кислорода не произошло ни малейшего изменения. То есть процентная доля кислорода, производимого этими бактериями на земле все 3.5 миллиардов лет - 20.94%. И это единственно возможный для жизни процентный состав! Но как, откуда одноклеточные существа знают, сколько им надо производить кислорода для того, чтобы мы могли жить? Кто назначил им эту программу действий? !!!
С появлением жизни на земле Творец создавал и новые формы жизни, с каждым годом число живых существ - животных и людей, потребляющих кислород и вырабатывающих углекислый газ, росло в геометрической прогрессии, но содержание кислорода не уменьшается, оно все время на одном уровне. Неужели выработка кислорода на земле может быть бесконтрольной? И кто рассчитывает, сколько бактерий должно быть на земле, чтобы кислорода хватало всем, чтобы его соотношение не уменьшалось, даже если число потребителей растет каждый день? Ведь потребление кислорода меняется и в течении всего дня, люди занимаются спортом, увеличивается потребность в кислороде, каждую секунду рождаются новые люди и животные. Население планеты увеличивае6тся с каждой секундой, им тоже нужен сразу кислород, но удивительным образом его хватает на всех. А это значит, что есть Тот, кто знает обо всех изменениях, кто рассчитывает ежесекундно то, в чем нуждаемся мы для жизни, и кто заставляет все созданное им обеспечивать нас, это и есть наш Творец!!!
И чем больше мы изучаем наше мироздание, законы жизни на планете, тем более мы укрепляем в точном знании о существовании Высшего Разума, Высшего Творца, контролирующего, гармонично регулирующего и создающего каждый миг жизни миллиардов живых существ. Мы видим, что на земле и в космосе нет ничего случайного и бессистемного.
Если бы атмосфера не была достаточно прозрачной и солнечный свет не мог проникать сквозь нее, от него было бы мало пользы. Однако наша атмосфера не только пропускает солнечный свет, но и служит препятствием для всех других видов излучения, которые вредны и опасны для жизни. Атмосфера пропускает видимый свет и инфракрасные лучи, необходимые для жизни, но, выступая в качестве фильтра, блокирует поступление других видов вредоносной радиации, поступающей из космоса.
Атмосферные газы поглощают практически весь спектр электромагнитного излучения по обе стороны видимого и инфракрасного света. Из всего диапазона излучения – от радиоволн до гамма-лучей – через атмосферу проходит очень узкий участок, включающий видимый и инфракрасный свет. Гамма-лучи, ультрафиолетовые лучи и микроволновое излучение не достигают поверхности Земли.
Невозможно не видеть удивительную мудрость этого Плана. Солнце посылает нам только 1 к 10 в ²³ степени полезного излучения, единственно нужного для нашей жизни, и лишь это излучение оказывается способно пройти сквозь атмосферу Земли.
Поразительно и то, что вода, также, как и воздух, обладает определенной степенью прозрачности, поэтому единственное излучение, способное проходить через воду, это участок спектра видимого света. Даже инфракрасное излучение, которое способно проникать сквозь атмосферу (и давать нам тепло), проходит в глубину воды всего лишь на несколько миллиметров. Вследствие этого только несколько миллиметров поверхности воды мировых океанов согреваются теплом солнечного света. Это тепло постепенно передается более глубоким слоям воды, и, благодаря этому, на определенной глубине температура морской воды подобна температуре, которую мы имеем на земле. Это создает вполне пригодную для жизни морских обитателей среду.
В отношении воды интересно и то, что различные цвета видимого света способны проникать в нее на разные расстояния. Например, красный свет не может проникать ниже восьми метров в глубину, в то время как желтый свет проникает на глубину до ста метров, а голубой и зеленый могут опускаться до двухсот сорока метров. Это обстоятельство - очень важная часть Господнего Замысла жизни, поскольку свет необходим для фотосинтеза, прежде всего, голубой и зеленый участки его спектра. Вода же, как выяснилось, пропускает эти два цвета на самую большую глубину, таким образом, растения, осуществляющие фотосинтез, способны жить только на глубине до двухсот сорока метров от поверхности.
Достижения современной науки, однако, показывают, что каждая деталь, каждый закон Вселенной содержит Высший смысл и цель, План, назначение которого – рождение и поддержание жизни человека. Все в этом Плане – даже такая его составляющая, как свет, о котором мы, возможно, никогда не задумывались, – столь очевидно «правильно», что этому нельзя не поразиться.
Пытаться объяснить этот рассчитанный во всех деталях План с бесчисленным множеством идеально работающих систем случайностью – в высшей степени неразумно.
Разве могут быть случайностью все те факты, о которых мы говорили выше: ограничение солнечного излучения, проникающего сквозь атмосферу до узкого участка 1 к 10 в ²³ степени всего электромагнитного спектра, т.е. того участка света, который необходим для жизни; отторжение атмосферой всех видов вредоносного излучения; способность воды не пропускать ни одну форму смертоносных лучей и пропускать только видимый свет. Эта удивительно правильно настроенная система есть результат Высшего Господнего Замысла, но никак не случайности.
Вселенная и все ее детали, свет Cолнца, согревающий нас, дающий нам возможность видеть, сотворены для нас, для нашей жизни.
2012-11-20 18:15:39www.harunyahya.ru










