
Плазматическая мембрана растительной клетки (плазмалемма). Её структура. Цитоплазматическая мембрана растений
функции, строение. Наружная цитоплазматическая мембрана :: SYL.ru
Наружная цитоплазматическая мембрана представляет собой тончайшую пленку. Ее толщина - порядка 7-10 нм. Просматривается пленка только в электронный микроскоп.  Далее рассмотрим, что собой представляет цитоплазматическая мембрана. Функции пленки также будут описаны в статье.
Далее рассмотрим, что собой представляет цитоплазматическая мембрана. Функции пленки также будут описаны в статье.
Структура
Какой состав имеет цитоплазматическая мембрана? Строение пленки достаточно разнообразно. В соответствии с химической организацией, она представляет собой комплекс белков и липидов. Цитоплазматическая мембрана клетки включает в себя бислой. Он выступает в качестве основы. Кроме этого, цитоплазматическая мембрана содержит холестерол и гликолипиды. Этим веществам свойственна амфипатричность. Другими словами, в них присутствуют гидрофобные ("боящиеся влаги") и гидрофильные ("любящие воду") концы. Последние (фосфатная группа) направлены наружу от мембраны, вторые (остатки от жирных кислот) ориентированы друг к другу. За счет этого и формируется липидный биполярный слой. Липидные молекулы обладают подвижностью. Они способны перемещаться в собственном монослое либо (что редко) из одного в другой. 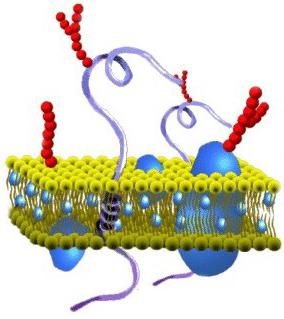 Липидный слой может иметь состояние твердого или жидкого кристалла. Монослои отличаются асимметричностью. Это значит, что в них различен состав липидов. За счет этого свойства цитоплазматические мембраны обладают специфичностью даже в рамках одной клетки. Ко второму обязательному компоненту пленки относят белки. Многие из этих соединений могут перемещаться в мембранной плоскости либо совершать вращения вокруг собственной оси. При этом они не способны переходить из одной части бислоя в другую. Защита внутренней среды – основная задача, которую выполняет цитоплазматическая мембрана. Строение пленки, кроме этого, обеспечивает течение различных процессов. За выполнение тех или иных задач отвечают белки. Благодаря липидам обеспечиваются структурные особенности пленки.
Липидный слой может иметь состояние твердого или жидкого кристалла. Монослои отличаются асимметричностью. Это значит, что в них различен состав липидов. За счет этого свойства цитоплазматические мембраны обладают специфичностью даже в рамках одной клетки. Ко второму обязательному компоненту пленки относят белки. Многие из этих соединений могут перемещаться в мембранной плоскости либо совершать вращения вокруг собственной оси. При этом они не способны переходить из одной части бислоя в другую. Защита внутренней среды – основная задача, которую выполняет цитоплазматическая мембрана. Строение пленки, кроме этого, обеспечивает течение различных процессов. За выполнение тех или иных задач отвечают белки. Благодаря липидам обеспечиваются структурные особенности пленки.
Цитоплазматическая мембрана: функции
Основными задачами являются:
- Барьерная. Защитная пленка обеспечивает активный, пассивный, избирательный, регулируемый обмен соединений с внешней средой. За счет избирательной проницаемости осуществляется отделение клетки и ее компартментов и снабжение их нужными веществами.
- Транспортная. Сквозь пленку осуществляется переход соединений от клетки к клетке. Благодаря этому доставляются питательные соединения, удаляются конечные продукты обмена, происходит секреция разных веществ. Кроме этого, формируются ионные градиенты, на оптимальном уровне поддерживаются ионная концентрация и рН. Они необходимы для активной деятельности ферментов клетки.
Вспомогательные задачи
- Матричная. Эта функция обеспечивает определенную ориентацию и взаиморасположение белков мембраны, а также оптимальное их взаимодействие.
- Механическая. За счет нее обеспечивается автономность клетки, внутренних структур. Также осуществляется соединение элемента с прочими аналогичными.

- Энергетическая. На фоне фотосинтеза в хлоропластах и при осуществлении клеточного дыхания в мембранах активны системы энергетического переноса. В них также участвуют и белковые соединения.
- Рецепторная. Ряд белков, которые присутствуют в мембране, обеспечивает восприятие различных сигналов. К примеру, циркулирующие в крови стероиды оказывают воздействие только на те клетки-мишени, которые обладают соответствующими гормонам рецепторами. Химические соединения, обеспечивающие проведение импульсов (нейромедиаторы), также связываются с помощью особых белков клеток-мишеней.
Особые свойства
К специфическим функциям мембраны относят:
- Ферментативную. Зачастую белки, которые содержит цитоплазматическая мембрана, выступают в качестве ферментов.
- Генерацию и проведение биопотенциалов.
- Маркировку. Цитоплазматическая мембрана включает в свой состав особые антигены. Они действуют как маркеры-"ярлыки". Благодаря им осуществляется распознание клеток. Маркеры представляют собой гликопротеины – белки, содержащие разветвленные олигосахаридные боковые цепи. Они выступают в качестве "антенн".
 Благодаря огромному количеству вариантов боковых цепей для того или другого типа клеток может быть сформирован особый маркер. При их помощи распознанные друг другом элементы начинают действовать согласованно. К примеру, так происходит при образовании тканей и органов. Маркировка также позволяет иммунитету определить чужеродные антигены.
Благодаря огромному количеству вариантов боковых цепей для того или другого типа клеток может быть сформирован особый маркер. При их помощи распознанные друг другом элементы начинают действовать согласованно. К примеру, так происходит при образовании тканей и органов. Маркировка также позволяет иммунитету определить чужеродные антигены.
Дополнительные сведения
Если какие-то частицы по тем или другим причинам не способны пройти сквозь фосфолипидный бислой (к примеру, вследствие гидрофильных свойств, поскольку внутри цитоплазматическая мембрана гидрофобна и такие соединения не пропускает, либо из-за больших размеров самих частиц), но они необходимы, то пройти они могут с помощью специальных белков-переносчиков (транспортеров) и белков-каналов. Либо проникновение их осуществляется посредством эндоцитоза.  В процессе пассивного транспорта пересечение веществами липидного слоя происходит путем диффузии. При этом энергия не затрачивается. В качестве одного из вариантов такого механизма может выступать облегченная диффузия. В ходе нее облегчает прохождение вещества какая-нибудь специфическая молекула. У нее может присутствовать канал, способный пропускать только однотипные частицы. При активном транспорте затрачивается энергия. Это связано с тем, что данный процесс осуществляется против концентрационного градиента. Цитоплазматическая мембрана содержит особые белки-насосы, АТФазу в том числе, которая способствует активному вхождению калиевых и выведению натриевых ионов.
В процессе пассивного транспорта пересечение веществами липидного слоя происходит путем диффузии. При этом энергия не затрачивается. В качестве одного из вариантов такого механизма может выступать облегченная диффузия. В ходе нее облегчает прохождение вещества какая-нибудь специфическая молекула. У нее может присутствовать канал, способный пропускать только однотипные частицы. При активном транспорте затрачивается энергия. Это связано с тем, что данный процесс осуществляется против концентрационного градиента. Цитоплазматическая мембрана содержит особые белки-насосы, АТФазу в том числе, которая способствует активному вхождению калиевых и выведению натриевых ионов.
Модели
Их существует несколько:
- "Бутербродная модель". Идею о трехслойном строении всех мембран высказали ученые Даусон и Даниэли в 1935 году. По их мнению, структура пленки была следующей: белки-липиды-белки. Такое представление существовало достаточно долго.
- "Жидкостно-мозаичная структура". Эта модель была описана Николсоном и Сингером в 1972 году. В соответствии с ней белковые молекулы не формируют сплошной слой, а погружаются в биполярный липидный в виде мозаики на различную глубину. Эта модель считается наиболее универсальной.
- "Белково-кристаллическая структура". В соответствии с этой моделью мембраны формируются за счет переплетения белковых и липидных молекул, которые объединены на базе гидрофильно-гидрофобных связей.
www.syl.ru
ЦИТОПЛАЗМАТИЧЕСКАЯ МЕМБРАНА. Функции. Структура
Оболочка клетки – это комплекс структур, отделяющий клетку от окружающей среды. Она состоит из наружного слоя – клеточной стенки и расположенной под ней плазматической мембраны.
Клетки животных и растений различаются по строению их наружного слоя. У растений и грибов на поверхности клеток расположена плотная оболочка - клеточная стенка. У большинства растений она состоит из целлюлозы, у грибов - из хитина. Клеточная стенка представляет собой защитную оболочку, обеспечивает форму растительных клеток, через клеточную стенку проходит вода, соли, молекулы многих органических веществ.
У животной клетки клеточной стенки нет. К цитоплазме примыкает плазматическая мембрана.
Под клеточной стенкой расположена плазматическая мембрана - плазмалемма (мембрана - кожица, пленка), граничащая непосредственно с цитоплазмой. Толщина плазматической мембраны около 10 нм.
Учитель: Сегодня на уроке мы познакомимся со строением и функциями плазматической мембраны.
Из истории открытия мембраны
Термин «мембрана» был предложен около ста лет назад для обозначения границ клетки, но с развитием электронной микроскопии стало ясно, что клеточная мембрана входит в состав структурных элементов клетки.
О наличии же пограничной мембраны между клеткой и окружающей средой было известно задолго до появления электронного микроскопа. Физико-химики отрицали существование плазматической мембраны и считали, что это просто граница раздела между живым коллоидным содержимым и окружающей средой, но Пфеффер (немецкий ботаник и физиолог растений) в 1890 году подтвердил существование ЦПМ.
В начале прошлого века Овертон (британский физиолог и биолог) обнаружил, что скорость проникновения многих веществ в эритроциты прямо пропорциональна их растворимости в липидах. В связи с этим ученый предположил, что мембрана содержит большое количество липидов и вещества, растворяясь в ней, проходят через нее и оказываются по ту сторону мембраны.
В 1925 году Гортер и Грендель (американские биологи) выделили липиды из клеточной мембраны эритроцитов. Полученные липиды они распределили по поверхности воды толщиной в одну молекулу. Оказалось, что площадь поверхности, занятой слоем липидов, в два раза больше площади самого эритроцита. Поэтому эти ученые сделали вывод, что клеточная мембрана состоит не из одного, а из двух слоев липидов.
Даусон и Даниэлли (английские биологи) в 1935 году высказали предположение, что в клеточных мембранах липидный бимолекулярный слой заключен между двумя слоями белковых молекул.

С появлением электронного микроскопа открылась возможность познакомиться со строением мембраны, и тогда обнаружилось, что мембраны животных и растительных клеток выглядят как трехслойная структура.

В 1959 году биолог Дж. Д. Робертсон, объединив имевшиеся в то время данные, выдвинул гипотезу о строении «элементарной мембраны», в которой он постулировал структуру, общую для всех биологических мембран.
Постулаты Робертсона о строении «элементарной мембраны»:
1. Все мембраны имеют толщину около 7,5 нм.
2. В электронном микроскопе все они представляются трехслойными.
3. Трехслойный вид мембраны есть результат именно того расположения белков и полярных липидов, которое предусматривала модель Даусона и Даниэлли – центральный липидный бислой заключен между двумя слоями белка.
Эта гипотеза о строении «элементарной мембраны» претерпела различные изменения и в 1972 году Сингером и Николсоном была предложена жидкостно-мозаичная модель мембраны, которая в настоящее время является общепризнанной.

Согласно этой модели основой любой мембраны является двойной слой фосфолипидов. У фосфолипидов (соединений, содержащих фосфатную группу) молекулы состоят из полярной головки и двух неполярных хвостов.

В фосфолипидном бислое гидрофобные остатки жирных кислот обращены внутрь, а гидрофильные головки, включающие остаток фосфорной кислоты, – наружу.

Фосфолипидный бислой представлен как динамическая структура, липиды могут перемещаться, меняя свое положение.
Двойной слой липидов обеспечивает барьерную функцию мембраны, не давая содержимому клетки растекаться, и препятствует попаданию в клетку токсических веществ.
Мембранные белки
В липидный бислой мембраны погружены молекулы белков, они образуют подвижную мозаику. По расположению в мембране и способу взаимодействия с липидным бислоем белки можно разделить на:
- поверхностные (или периферические) мембранные белки, связанные с гидрофильной поверхностью липидного бислоя;
- интегральные (мембранные) белки, погруженные в гидрофобную область бислоя.
Интегральные белки различаются по степени погруженности их в гидрофобную область бислоя. Они могут быть полностью погружены (интегральные) или частично погружены (полуинтегральные), а также могут пронизывать мембрану насквозь (трансмембранные).
Мембранные белки по своим функциям можно разделить на две группы:
- структурные белки. Они входят в состав клеточных мембран и участвуют в поддержании их структуры.
- динамические белки. Они находятся на мембранах и участвуют в происходящих на ней процессах.
Выделяют три класса динамических белков.
1. Рецепторные. С помощью этих белков клетка воспринимает различные воздействия на свою поверхность. То есть они специфически связывают такие соединения, как гормоны, нейромедиаторы, токсины на наружной стороне мембраны, что служит сигналом для изменения различных процессов внутри клетки или самой мембраны.
2. Транспортные. Эти белки транспортируют через мембрану те или иные вещества, также они образовывают каналы, через которые осуществляется транспорт различных ионов в клетку и из нее.
3. Ферментативные. Это белки-ферменты, которые находятся в мембране и участвуют в различных химических процессах.
На поверхности мембраны животных клеток образуется наружный рецепторный слой углеводов – гликокаликс. Образование гликокаликса, так же как и клеточных стенок растений, происходит благодаря жизнедеятельности самих клеток.
Основные функции клеточной мембраны
1. Структурная(клеточная мембрана отделяет клетку от окружающей среды).
2. Транспортная (через клеточную мембрану осуществляется транспорт веществ, причем клеточная мембрана является высокоизбирательным фильтром).
3. Рецепторная (находящиеся на поверхности мембраны рецепторы воспринимают внешние воздействия, передают эту информацию внутрь клетки, позволяя ей быстро реагировать на изменения окружающей среды).
Другие функции мембраны
Похожие статьи:
poznayka.org
| Общая характеристика. Плазматическая (цитоплазматическая) мембрана — обязательный компонент любой клетки. Она отграничивает клетку и обеспечивает сохранение существующих различий между клеточным содержимым и окружающей средой. Мембрана служит высокоизбирательным «входным» селективным фильтром и отвечает за активный транспорт веществ в клетку и из нее. Цитоплазматическую мембрану растительной клетки обычно называют плазмалеммой. Как и любая биологическая мембрана, она представляет собой липидный бислой с большим количеством белков. Основу липидного бислоя составляют фосфолипиды. Помимо них в состав липидного слоя входят гликолипиды и стерины. Липиды достаточно активно перемещаются в пределах своего монослоя, но возможны и их переходы из одного монослоя в другой. Такой переход, называемый «флип-флоп» (от англ. flip-flop), осуществляется ферментом флипазой. Кроме липидов и белков в плазмалемме присутствуют углеводы. Соотношение липидов, белков и углеводов в плазматической мембране растительной клетки составляет приблизительно 40:40: 20. Мембранные белки связаны с липидным бислоем различными способами. Первоначально белки мембран разделяли на два основных типа: периферийные и интегральные. Периферийные белки ассоциированы с мембраной за счет присоединения к интегральным белкам или липидному бислою слабыми связями: водородными, электростатическими, солевыми мостиками. Они в основном растворимы в воде и легко отделяются от мембраны без ее разрушения. Некоторые периферийные белки обеспечивают связь между мембранами и цитоскелетом. Интегральные белки мембран нерастворимы в воде.
Как минимум один из доменов интегрального белка встроен в гидрофобную часть бислоя мембраны, поэтому интегральный белок, как правило, не может быть удален из мембраны без ее разрушения. В последнее время показано существование третьей группы белков, так называемых «заякоренных» в мембране белков). Эти белки фиксируются в мембране за счет специальной молекулы, в качестве которой могут выступать жирная кислота (ЖК), стерин, изопреноид или фосфатидилинозитол. Белки, связанные с изопреноидами (пренилированные белки) или жирной кислотой, обратимо соединяются с эндоплазматической (внутренней) поверхностью мембраны. Из жирных кислот чаще используется миристиновая (С14) или пальмитиновая (C16). В первом случае образуется амидная связь с терминальной аминогруппой глицина. К остаткам пальмитиновой кислоты белки присоединяются за счет тиоэфирных связей с цистеинами в С-конце полипептидной цепи. Для пренилирования белков обычно используется фарнезил или геранилгеранил, которые также присоединяются к остаткам цистеина в карбоксильном конце полипептида. В отличие от этих двух групп белков фосфати-дилинозитолсвязанные белки находятся с внешней (экстрацеллюлярной, или люменальной) стороны мембраны. Подобным образом, по-видимому, связаны с плазматической мембраной большинство арабиногалактановых белков. Холестеринсвязанные белки недавно были обнаружены в плазматической мембране животных клеток, но в растительных клетках подобные белки пока не найдены. Особенностью липидного состава плазмалеммы по сравнению с другими мембранами растительной клетки является высокое содержание стеринов, но в отличие от плазматической мембраны животной клетки для плазмалеммы характерна высокая вариабельность их состава в зависимости от вида растения, органа и ткани. Например, у ячменя (Hordeum vulgaris) в клетках корня количество свободных стеринов превышает количество фосфолипидов более чем в два раза, тогда как в листьях фосфолипидов больше, чем стеринов почти в 1,5 раза. В листьях шпината (Spinacia oleracia) соотношение фосфолипиды: свободные стерины почти на порядок выше — 9:1.
Структурные особенности плазмалеммы. Помимо высокой степени вариабельности плазматическая мембрана растительной клетки имеет ряд структурных особенностей, отличающих ее от других эукариотических клеток. Жирнокислотный состав. Основными жирными кислотами плазмалеммы являются пальмитиновая (16:0), олеиновая (18:1; Δ9), линолевая (18 : 2; Δ9,12) и линоленовая (18 :3; Δ9,12,15). Практически отсутствует стеариновая кислота (18:0) и полностью — арахидоновая (20:4; Δ5'8,11,14), характерные для мембран клеток животных и грибов. Известно, что арахидоновая кислота в очень низких концентрациях является мощным стимулятором фитоиммунитета для некоторых видов растений (например, обеспечивает устойчивость картофеля к фитофторе). Набор стероидов. В плазмалемме, как и во всех мембранах растительной клетки, почти нет холестерина. Его заменяют фитостерины, которые являются С-24-замещенными стеринами. Основные фитостерины — ситостерин, стигмастерин и кампестерин. Помимо свободных фитостеринов присутствуют значительные количества эфиров, гликозидов и ацилгликозидов стеринов, что не характерно для клеток животных. Набор белков. Наличие специфических белков, прежде всего арабиногалактанов (AGPs), обеспечивающих структурное и функциональное взаимодействие плазмалеммы с клеточной стенкой. |


fizrast.ru
Цитоплазматическая мембрана
Цитоплазма бактериальной клетки ограничена от клеточной стенки тонкой полупроницаемой структурой толщиной 5-10 нм, называемой цитоплазматической мембраной (ЦПМ). ЦПМ состоит из двойного слоя фосфолипидов, пронизанных белковыми молекулами (рис. 6).

Рис. 6. Строение плазматической мембраны (по теории Давсона-Даниэлли). Два слоя фосфолипидных молекул, обращенных гидрофобными полюсами друг к другу и покрытых двумя слоями молекул глобулярного белка (А.Поликар, 1975).
С ЦПМ связаны многие ферменты и белки, участвующие в транслокации питательных веществ, а также ферменты и переносчики электронов конечных стадий биологического окисления (дегидрогеназы, цитохромная система, АТФ-аза). На ЦМП локализуются ферменты, катализирующие синтез пептидогликана, белков клеточной стенки, собственных структур. Мембрана является также местом превращения энергии при фотосинтезе, окислительном фосфорилировании.
Периплазматическое пространство
Периплазматическое пространство (периплазма) представляет собой зону между клеточной стенкой и ЦПМ. Толщина периплазмы составляет около 10 нм, объем зависит от условий среды и, прежде всего, от осмотических свойств раствора. Периплазма может включать до 20% всей находящейся в клетке воды, в ней локализуются некоторые ферменты (фосфатазы, пермеазы, нуклеазы и др.) и транспортные белки – переносчики соответствующих субстратов.
Мезосомы
Мезосомы представляют собой мембранные структуры, образуемые при закручивании ЦПМ. Морфологически мезосомы выглядят как ламеллярные стопки или спирально упакованные ламеллы, везикулярные или тубулярные структуры, а также смешанные мембранные системы, образованные трубочками, пузырьками и ламеллами (рис. 7). По расположению в клетке различают: мезосомы, образующиеся в зоне клеточного деления и формирования клеточной перегородки (септальные мезосомы) и мезосомы, сформированные в результате инвагинации периферических участков ЦПМ (латеральные мезосомы).

Рис. 7. Типы строения истинных мезосом. а - ламеллярный; б - г - тубулярные типы (Бирюзова, Поглазова, 1977).
Предполагается, что мезосомы полифункциональны, содержат различные ферментные системы и играют определенную роль в энергетическом метаболизме. Считают, что они являются сайтом для формирования клеточной стенки бактерий и прикрепления нуклеоида в процессе репликации ДНК. Септальные мезосомы участвуют в построении поперечной перегородки при делении бактерий.
Цитоплазма
Содержимое клетки, окруженное ЦПМ, составляет цитоплазму бактерий. Та часть цитоплазмы, которая имеет гомогенную коллоидную консистенцию и содержит растворимые РНК, ферменты, субстраты и продукты обмена веществ, обозначается как цитозоль. Другая часть цитоплазмы представлена различными структурными элементами: мезосомами, рибосомами, включениями, нуклеоидом, плазмидами.
Рибосомы - субмикроскопические рибонуклеопротеидные гранулы диаметром 15-20 нм. В рибосомах находится примерно 80-85% всей бактериальной РНК. Рибосомы прокариот имеют константу седиментации 70 S. Они построены из двух частиц: 30 S (малая субъединица) и 50 S (большая субъединица) (рис. 8). Рибосомы служат местом синтеза белка.

Рис. 8. Рибосома (а) и ее субчастицы -большая (б) и малая (в) (Блинов Н.П., 1989).
Некоторые бактерии способны накапливать фосфорную кислоту в виде гранул полифосфата (зерна волютина, метахроматические зерна, зерна Бабеша-Эрнста). Они играют роль фосфатных депо и регулярно выявляются у коринебактерий, микобактерий и спирилл в виде плотных, хорошо контурированных образований в форме шара или эллипса, располагающихся, в основном, у полюсов клетки. Обычно на полюсах бывает по одной грануле.
Наличие зерен волютина у бактерий определяют методом Нейссера.
studfiles.net
Цитоплазматическая мембрана (оболочка)
скачать ЦИТОПЛАЗМАЦитоплазма представляет собой водянистое вещество – цитозоль (90 % воды), в котором располагаются различные органеллы, а также питательные вещества (в виде истинных и коллоидных растворов) и нерастворимые отходы метаболических процессов. В цитозоле протекает гликолиз, синтез жирных кислот, нуклеотидов и других веществ. Цитоплазма является динамической структурой. Органеллы движутся, а иногда заметен и циклоз – активное движение, в которое вовлекается вся протоплазма.
ЦИТОПЛАЗМАТИЧЕСКАЯ МЕМБРАНАЦитоплазматическая мембрана (оболочка) - это тонкая структура, которая отделяет содержимое клетки от окружающей среды. Она состоит из двойного слоя липидов с белковыми молекулами. Клеточная мембрана сплошная, но у нее имеются многочисленные складки, извилины, и поры, что позволяет регулировать прохождение через нее веществ. ЭНДОПЛАЗМАТИЧЕСКАЯ СЕТЬ
Эндоплазматическая сеть – это сеть мембран, пронизывающих цитоплазму эукариотических клеток. Её можно наблюдать только при помощи электронного микроскопа. Эндоплазматическая сеть связывает органеллы между собой, по ней происходит транспорт питательных веществ. Гладкая ЭПС имеет вид трубочек, стенки которых представляют собой мембраны, сходные по своей структуре с плазматической мембраной. В ней осуществляется синтез липидов и углеводов. На мембранах каналов и полостей гранулярной ЭПС расположено множество рибосом; данный тип сети участвует в синтезе белка.
РИБОСОМЫ.
Рибосомы – мелкие (15–20 нм в диаметре) органеллы, состоящие из р-РНК и полипептидов. Важнейшая функция рибосом – синтез белка. Их количество в клетке весьма велико: тысячи и десятки тысяч. Рибосомы могут быть связаны с эндоплазматической сетью или находиться в свободном состоянии. В процессе синтеза обычно одновременно участвуют множество рибосом, объединённых в цепи, называемые полирибосомами.
КОМПЛЕКС ГОЛЬДЖИ.Аппарат Гольджи представляет собой стопку мембранных мешочков (цистерн) и связанную с ними систему пузырьков. На наружной, вогнутой стороне стопки из пузырьков (отпочковывающихся, по-видимому, от гладкой эндоплазматической сети) постоянно образуются новые цистерны, на внутренней стороне цистерны превращаются обратно в пузырьки. Основной функцией аппарата Гольджи является транспорт веществ в цитоплазму и внеклеточную среду, а также синтез жиров и углеводов, в частности, гликопротеина муцина, образующего слизь, а также воска, камеди и растительного клея. Аппарат Гольджи участвует в росте и обновлении плазматической мембраны и в формировании лизосом.
МИТОХОНДРИИ.Митохондрии иногда называют «клеточными электростанциями». Это спиральные, округлые, вытянутые или разветвлённые органеллы, длина которых изменяется в пределах 1,5–10 мкм, а ширина – 0,25–1 мкм. Митохондрии могут изменять свою форму и перемещаться в те области клетки, где потребность в них наиболее высока. В клетке содержится до тысячи митохондрий, причём это количество сильно зависит от активности клетки. Каждая митохондрия окружена двумя мембранами, внутри которых содержатся РНК, белки и митохондриальная ДНК, участвующая в синтезе митохондрий наряду с ядерной ДНК. Внутренняя мембрана сложена в складки, называемые кристами. Возможно, митохондрии некогда были свободнодвижущимися бактериями, которые, случайно проникнув в клетку, вступили с хозяином в симбиоз. Важнейшей функцией митохондрий является синтез АТФ, происходящий за счёт окисления органических веществ.
ЛИЗОСОМЫ.Лизосомы представляют собой мембранные мешочки, наполненные пищеварительными ферментами. Особенно много лизосом в животных клетках, здесь их размер составляет десятые доли микрометра. Лизосомы расщепляют питательные вещества, переваривают попавшие в клетку бактерии, выделяют ферменты, удаляют путём переваривания ненужные части клеток. Лизосомы также являются «средствами самоубийства» клетки: в некоторых случаях (например, при отмирании хвоста у головастика) содержимое лизосом выбрасывается в клетку, и она погибает.
КЛЕТОЧНЫЙ ЦЕНТР.Клеточный центр, митотический центр, постоянная структура почти всех животных и некоторых растительных клеток, определяет полюса делящейся клетки (см. Митоз). К. ц. обычно состоит из двух центриолей — плотных гранул размером 0,2—0,8 мкм, расположенных под прямым углом друг к другу. При образовании митотического аппарата центриоли расходятся к полюсам клетки, определяя ориентировку веретена деления клетки. Поэтому правильнее К. ц. называть митотическим центром, отражая этим его функциональное значение, тем более что лишь в некоторых клетках К. ц. расположен в ее центре. В ходе развития организма изменяются как положение К. ц. в клетках, так и форма его. При делении клетки каждая из дочерних клеток получает пару центриолей. Процесс их удвоения происходит чаще в конце предыдущего клеточного деления. Возникновение ряда патологических форм деления клетки связано с ненормальным делением К. ц.
ЦИТОСКЕЛЕТ.
Отличительная особенность эукарит. клетки является развитие в цитоплазме скелетных образований - микротрубочки и пучки белковых волокон. Элементы цитоскелета связаны с нар. цит. плазматической мембраной и ядерной оболочкой, образуют сложные переплетения в цитоплазме. Опорные элементы цитоплазмы определяют форму клетки, обеспечивают движение внутриклеточных структур и перемещение всей клетки. В некоторых типах клеток, простейших, хорошо выраженные элементы цитоскелета на протяжении всей жизни, в других случаях цитоскелет легко перестраивается, например у амеб, обеспечивая быстрое изменение формы клетки.
ЖГУТИКИ И РЕСНИЧКИ.Жгутики и реснички представляют собой подвижные цитоплазматические отростки, служащие либо для передвижения всего организма (у бактерий, водорослей, грибов, ресничных червей и др.), либо репродуктивных клеток (изогамет, спермиев, зооспор), либо для транспорта частиц и жидкостей (например, реснички у мерцательных клеток слизистой носа и трахеи, яйцеводов и т. д.).Жгутики эукариотических клеток имеют толщину до 200 нм и длину до 100 мкм и больше. Более короткие (обычно 10-20 мкм) жгутики, которых бывает много на одной клетке, называются ресничками. По всей длине жгутика или реснички проходят 20 микротрубочек: 9 периферических дублетов и 2 центральные одиночные. Дублет состоит из 23 протофиламентов - по 10 на каждую микротрубочку и 3 общих в области их соприкосновения. Дублеты имеют парные отростки (разделенные по длине трубочки расстояниями около 17 нм) из удлиненных молекул белка динеина. Эти отростки (толщиной 2-5 нм и длиной до 10-40 нм), подходят к соседним дублетам.
скачатьnenuda.ru
Цитоплазматическая мембрана
Цитоплазма бактериальной клетки ограничена от клеточной стенки тонкой полупроницаемой структурой толщиной 5-10 нм, называемой цитоплазматической мембраной (ЦПМ). ЦПМ состоит из двойного слоя фосфолипидов, пронизанных белковыми молекулами (рис. 6).

Рис. 6. Строение плазматической мембраны (по теории Давсона-Даниэлли). Два слоя фосфолипидных молекул, обращенных гидрофобными полюсами друг к другу и покрытых двумя слоями молекул глобулярного белка (А.Поликар, 1975).
С ЦПМ связаны многие ферменты и белки, участвующие в транслокации питательных веществ, а также ферменты и переносчики электронов конечных стадий биологического окисления (дегидрогеназы, цитохромная система, АТФ-аза). На ЦМП локализуются ферменты, катализирующие синтез пептидогликана, белков клеточной стенки, собственных структур. Мембрана является также местом превращения энергии при фотосинтезе, окислительном фосфорилировании.
Периплазматическое пространство
Периплазматическое пространство (периплазма) представляет собой зону между клеточной стенкой и ЦПМ. Толщина периплазмы составляет около 10 нм, объем зависит от условий среды и, прежде всего, от осмотических свойств раствора. Периплазма может включать до 20% всей находящейся в клетке воды, в ней локализуются некоторые ферменты (фосфатазы, пермеазы, нуклеазы и др.) и транспортные белки – переносчики соответствующих субстратов.
Мезосомы
Мезосомы представляют собой мембранные структуры, образуемые при закручивании ЦПМ. Морфологически мезосомы выглядят как ламеллярные стопки или спирально упакованные ламеллы, везикулярные или тубулярные структуры, а также смешанные мембранные системы, образованные трубочками, пузырьками и ламеллами (рис. 7). По расположению в клетке различают: мезосомы, образующиеся в зоне клеточного деления и формирования клеточной перегородки (септальные мезосомы) и мезосомы, сформированные в результате инвагинации периферических участков ЦПМ (латеральные мезосомы).

Рис. 7. Типы строения истинных мезосом. а - ламеллярный; б - г - тубулярные типы (Бирюзова, Поглазова, 1977).
Предполагается, что мезосомы полифункциональны, содержат различные ферментные системы и играют определенную роль в энергетическом метаболизме. Считают, что они являются сайтом для формирования клеточной стенки бактерий и прикрепления нуклеоида в процессе репликации ДНК. Септальные мезосомы участвуют в построении поперечной перегородки при делении бактерий.
Цитоплазма
Содержимое клетки, окруженное ЦПМ, составляет цитоплазму бактерий. Та часть цитоплазмы, которая имеет гомогенную коллоидную консистенцию и содержит растворимые РНК, ферменты, субстраты и продукты обмена веществ, обозначается как цитозоль. Другая часть цитоплазмы представлена различными структурными элементами: мезосомами, рибосомами, включениями, нуклеоидом, плазмидами.
Рибосомы - субмикроскопические рибонуклеопротеидные гранулы диаметром 15-20 нм. В рибосомах находится примерно 80-85% всей бактериальной РНК. Рибосомы прокариот имеют константу седиментации 70 S. Они построены из двух частиц: 30 S (малая субъединица) и 50 S (большая субъединица) (рис. 8). Рибосомы служат местом синтеза белка.

Рис. 8. Рибосома (а) и ее субчастицы -большая (б) и малая (в) (Блинов Н.П., 1989).
Некоторые бактерии способны накапливать фосфорную кислоту в виде гранул полифосфата (зерна волютина, метахроматические зерна, зерна Бабеша-Эрнста). Они играют роль фосфатных депо и регулярно выявляются у коринебактерий, микобактерий и спирилл в виде плотных, хорошо контурированных образований в форме шара или эллипса, располагающихся, в основном, у полюсов клетки. Обычно на полюсах бывает по одной грануле.
Наличие зерен волютина у бактерий определяют методом Нейссера.
studfiles.net
Цитоплазматическая мембрана - это... Что такое Цитоплазматическая мембрана?
У этого термина существуют и другие значения, см. Мембрана Изображение клеточной мембраны. Маленькие голубые и белые шарики соответствуют гидрофильным «головкам» липидов, а присоединённые к ним линии — гидрофобным «хвостам». На рисунке показаны только интегральные мембранные белки (красные глобулы и желтые спирали). Желтые овальные точки внутри мембраны — молекулы холестерола Желто-зеленые цепочки бусинок на наружной стороне мембраны — цепочки олигосахаридов, формирующие гликокаликс
Изображение клеточной мембраны. Маленькие голубые и белые шарики соответствуют гидрофильным «головкам» липидов, а присоединённые к ним линии — гидрофобным «хвостам». На рисунке показаны только интегральные мембранные белки (красные глобулы и желтые спирали). Желтые овальные точки внутри мембраны — молекулы холестерола Желто-зеленые цепочки бусинок на наружной стороне мембраны — цепочки олигосахаридов, формирующие гликокаликс Кле́точная мембра́на (или цитолемма, или плазмолемма, или плазматическая мембрана) отделяет содержимое любой клетки от внешней среды, обеспечивая ее целостность; регулируют обмен между клеткой и средой; внутриклеточные мембраны разделяют клетку на специализированные замкнутые отсеки — компартменты или органеллы, в которых поддерживаются определенные условия внутриклеточной среды.
Основные сведения
Клеточная стенка, если таковая у клетки имеется (обычно есть у растительных клеток), покрывает клеточную мембрану.
Клеточная мембрана представляет собой двойной слой (бислой) молекул класса липидов, большинство из которых представляет собой так называемые сложные липиды — фосфолипиды. Молекулы липидов имеют гидрофильную («головка») и гидрофобную («хвост») часть. При образовании мембран гидрофобные участки молекул оказываются обращены внутрь, а гидрофильные — наружу. Мембраны — структуры инвариабельные, весьма сходные у разных организмов. Некоторое исключение составляют, пожалуй, археи, у которых мембраны образованы глицерином и терпеноидными спиртами. Толщина мембраны составляет 7-8 нм.
Биологическая мембрана включает и различные белки: интегральные (пронизывающие мембрану насквозь), полуинтегральные (погруженные одним концом во внешний или внутренний липидный слой), поверхностные (расположенные на внешней или прилегающие к внутренней сторонам мембраны). Некоторые белки являются точками контакта клеточной мембраны с цитоскелетом внутри клетки, и клеточной стенкой (если она есть) снаружи. Некоторые из интегральных белков выполняют функцию ионных каналов, различных транспортеров и рецепторов.
Функции биомембран
- барьерная — обеспечивает регулируемый, избирательный, пассивный и активный обмен веществ с окружающей средой. Например, мембрана пероксисом защищает цитоплазму от опасных для клетки пероксидов. Избирательная проницаемость означает, что проницаемость мембраны для различных атомов или молекул зависит от их размеров, электрического заряда и химических свойств. Избирательная проницаемость обеспечивает отделение клетки и клеточных компартментов от окружающей среды и снабжение их необходимыми веществами.
- транспортная — через мембрану происходит транспорт веществ в клетку и из клетки. Транспорт через мембраны обеспечивает: доставку питательных веществ, удаление конечных продуктов обмена, секрецию различных веществ, создание ионных градиентов, поддержание в клетке соответствующего pH и ионной концентрации, которые нужны для работы клеточных ферментов.
Частицы, по какой-либо причине не способные пересечь фосфолипидный бислой (например, из-за гидрофильных свойств, так как мембрана внутри гидрофобна и не пропускает гидрофильные вещества, или из-за крупных размеров), но необходимые для клетки, могут проникнуть сквозь мембрану через специальные белки-переносчики (транспортеры) и белки-каналы или путем эндоцитоза.
При пассивном транспорте вещества пересекают липидный бислой без затрат энергии, путем диффузии. Вариантом этого механизма является облегчённая диффузия, при которой веществу помогает пройти через мембрану какая-либо специфическая молекула. У этой молекулы может быть канал, пропускающий вещества только одного типа.
Активный транспорт требует затрат энергии, так как происходит против градиента концентрации. На мембране существуют специальные белки-насосы, в том числе АТФаза, которая активно вкачивают в клетку ионы калия (K+) и выкачивают из неё ионы натрия (Na+).
- матричная — обеспечивает определенное взаиморасположение и ориентацию мембранных белков, их оптимальное взаимодействие;
- механическая — обеспечивает автономность клетки, ее внутриклеточных структур, также соединение с другими клетками (в тканях). Большую роль в обеспечение механической функции имеют клеточные стенки, а у животных — межклеточное вещество.
- энергетическая — при фотосинтезе в хлоропластах и клеточном дыхании в митохондриях в их мембранах действуют системы переноса энергии, в которых также участвуют белки;
- рецепторная — некоторые белки, сидящие в мембране, являются рецепторами (молекулами, при помощи которых клетке воспринимает те или иные сигналы).
Например, гормоны, циркулирующие в крови, действуют только на такие клетки-мишени, у которых есть соответствующие этим гормонам рецепторы. Нейромедиаторы (химические вещества, обеспечивающие проведение нервных импульсов) тоже связываются с особыми рецепторными белками клеток-мишеней.
- ферментативная — мембранные белки нередко являются ферментами. Например, плазматические мембраны эпителиальных клеток кишечника содержат пищеварительные ферменты.
- осуществление генерации и проведения биопотенциалов.
С помощью мембраны в клетке поддерживается постоянная концентрация ионов: концентрация иона К+ внутри клетки значительно выше, чем снаружи, а концентрация Na+ значительно ниже, что очень важно, так как это обеспечивает поддержание разности потенциалов на мембране и генерацию нервного импульса.
- маркировка клетки — на мембране есть антигены, действующие как маркеры — «ярлыки», позволяющие опознать клетку. Это гликопротеины (то есть белки с присоединенными к ним разветвленными олигосахаридными боковыми цепями), играющие роль «антенн». Из-за бесчисленного множества конфигурации боковых цепей возможно сделать для каждого типа клеток свой особый маркер. С помощью маркеров клетки могут распознавать другие клетки и действовать согласованно с ними, например, при формировании органов и тканей. Это же позволяет иммунной системе распознавать чужеродные антигены.
Структура и состав биомембран
Мембраны состоят из липидов трёх классов: фосфолипиды, гликолипиды и холестерол. Фосфолипиды и гликолипиды (липиды с присоединёнными к ним углеводами) состоят из двух длинных гидрофобных углеводородных «хвостов», которые связаны с заряженной гидрофильной «головой». Холестерол придаёт мембране жёсткость, занимая свободное пространство между гидрофобными хвостами липидов и не позволяя им изгибаться. Поэтому мембраны с малым содержанием холестерола более гибкие, а с большим — более жёсткие и хрупкие. Также холестерол служит «стопором», препятствующим перемещению полярных молекул из клетки и в клетку. Важную часть мембраны составляют белки, пронизывающие её и отвечающие за разнообразные свойства мембран. Их состав и ориентация в разных мембранах различаются.
Клеточные мембраны часто асимметричны, то есть слои отличаются по составу липидов, переход отдельной молекулы из одного слоя в другой (так называемый флип-флоп) затруднён.
Мембранные органеллы
Это замкнутые одиночные или связанные друг с другом участки цитоплазмы, отделённые от гиалоплазмы мембранами. К одномембранным органеллам относятся эндоплазматическая сеть, аппарат Гольджи, лизосомы, вакуоли, пероксисомы; к двумембранным — ядро, митохондрии, пластиды. Снаружи клетка ограничена так называемой плазматической мембраной. Строение мембран различных органелл отличается по составу липидов и мембранных белков.
Избирательная проницаемость
Клеточные мембраны обладают избирательной проницаемостью: через них медленно диффундируют глюкоза, аминокислоты, жирные кислоты, глицерол и ионы, причем сами мембраны в известной мере активно регулируют этот процесс-одни вещества пропускают, а другие нет. существует четыре основных механизма для поступления веществ в клетку или их из клеки наружу:диффузия, осмос, активный транспорт и экзо- или эндоцитоз. Два первых процесса носят пассивный характер, т.е. не требуют затрат энергии; два последних-активные процессы, связанные с потреблением энерги.
Избирательная проницаемость мембраны при пассивном транспорте обусловлена специальными каналами — интегральными белками. Они пронизывают мембрану насквозь, образовывая своего рода проход. Для элементов K, Na и Cl есть свои каналы. Относительно градиента концентрации молекулы этих элементов движутся в клетку и из неё. При раздражении каналы натриевых ионов раскрываются, и происходит резкое поступление в клетку ионов натрия. При этом происходит дисбаланс мембранного потенциала. После чего мембранный потенциал восстанавливается. Каналы калия всегда открыты, через них в клетку медленно попадают ионы калия.
Ссылки
- Bruce Alberts, et al. Molecular Biology Of The Cell. — 5th ed. — New York: Garland Science, 2007. — ISBN 0-8153-3218-1 — учебник по молекулярной биологии на англ. языке
- Рубин А.Б. Биофизика, учебник в 2 тт.. — 3-е издание, исправленное и дополненное. — Москва: издательство Московского университета, 2004. — ISBN 5-211-06109-8
- Геннис Р. Биомембраны. Молекулярная структура и функции: перевод с англ. = Biomembranes. Molecular structure and function (by Robert B. Gennis). — 1-е издание. — Москва: Мир, 1997. — ISBN 5-03-002419-0
- Иванов В.Г., Берестовский Т.Н. Липидный бислой биологических мембран. — Москва: Наука, 1982.
- Антонов В.Ф., Смирнова Е.Н., Шевченко Е.В. Липидные мембраны при фазовых переходах. — Москва: Наука, 1994.
См. также

Wikimedia Foundation. 2010.
dik.academic.ru








