15.Особенности полового размножения красных водорослей. Циклы воспроизведения. Циклы воспроизведения растений
24.Половое размножение растений, цикл воспроизведения » Шпоры для студентов
Вопросы по ботанике
Это наиболее совершенный способ размножения растений. Сущность Полового размножения Заключается в Слиянии, Или Копуляции, Женской и мужской гаплоидных половых клеток — Гамет С образованием диплоидной Зиготы, Дающей начало новому организму. Происходит соединение наследственных особенностей материнской и отцовской особей, вследствие чего потомство сочетает в себе признаки и свойства двух родительских форм. Такое потомство получается более разнообразным, жизнеспособным и лучше приспосабливается к различным условиям. У растений и грибов существует несколько Типов полового процесса Изогамия, гетерогамия, оогамия, гаметангиогамия, зигогамия, соматогамия. При Изогамии (водорослей и грибов) происходит соединение двух подвижных одинаковых гамет. При Гетерогамии (зеленым и бурым водорослям) сливаются две подвижные, но различные гаметы: женская — крупная, мужская — более мелкая. Оогамия — (низшие и высшие).Половые клетки образуются и развиваются в специальных органах — Гаметангиях Женская гамега — Яйцеклетка — крупная, неподвижная, которая развивается в Оогониях Или Архегониях. Мужские гаметы многочисленные, очень мелкие, развиваются в Антеридиях, Подэазделяются на подвижные Сперматозоиды (у низших, высших споровых) и неподвижные Спермии (у цветковых). Сформировавшаяся после оплодотворения зигота может находиться определенное время в стадии покоя, может сразу прорастать в новую особь или образовывать зооспоры, прорастающие в новые особи. Цикл воспроизведения – это отрезок жизни всегда ограниченный 2мя этапами. У каждого растения Полный жизненный цикл развития Возможен только при наличии бесполого и полового способов размножения Особь, образующая органы полового размножения с половыми клетками — гаметами, называется Половым поколением, Или Гаметофитом. Особь, пи которой образуются органы бесполого размножения со спорами, - Бесполым поколением, Или Спорофитом. Одновременно с чередованием поколений в цикле развития происходит Смена ядерных фаз. Она заключается в следующем. Споры имеют гаплоидный набор хромосом, так как образуются в спорангиях спорофитов из спорогенной ткани в результате редукционного деления. Из споры развивается гаплоидный гаметофит, на котором формируются гаплоидные гаметы. При их слиянии на гаметофите образуется зигота с диплоидным набором хромосом. Гаплоидная фаза ядра сменилась диплоидной фазой. Из зиготы развивается диплоидный спорофит. Затем при образовании спор число хромосом снова уменьшается вдвое и т. д. Имея различное число хромосом, гаметофит и спорофит различаются внешне.
shporiforall.ru
Цикл воспроизведения цветковых растений.
Реферат на тему
Цикл воспроизведения цветковых растений.
План:
Введение
-
Общая характеристика цикла воспроизведения цветковых растений
-
Характеристика гаплоидной фазы
-
Характеристика диплоидной фазы
-
Заключение
-
Схема цикла воспроизведения
Использованная литература
Введение
Высшие растения в ходе исторического развития приспособились к жизни в наземных условиях. У этих растений отмечается чередование полового (гаметофит) и бесполого (спорофит) поколений. Гаметофит продуцирует гаметы и защищает зародыш, тогда как спорофит продуцирует споры, которые обеспечивают следующую генерацию гаметофита. У растений этой группы доминирует диплоидный спорофит, который и определяет внешний вид растения.
В ходе эволюции на нашей планете среди царства растений наиболее большего развития получили отдел Покрытосемянные или Цветковые (Magnoliophyta). На сегодняшний день это самая процветающая группа растений, насчитывающая примерно 250 000 – 300 000 видов, что составляет примерно две трети видов царства растений. Типичными представителями являются рожь, осина, роза, лен, берёза, кактус, лотос, яблоня и др. Таким образом, представители этого отдела являются как травянистыми, так и кустарниковыми и деревьями, а также однодольными и двудольными, для которых характерно чередование поколений с характерной уменьшенной фазой гаметофита. Главным же отличием их от других отделов послужило наличие цветка, представляющий собой видоизменённый побег и являющийся производным спорофита, что в свою очередь ещё раз доказывает их высшее положение в современной систематике растений.
Цветковые (покрытосеменные) растения - самый обширный отдел растительного мира, к которому относится больше половины всех видов растений на Земле. Первые покрытосеменные растения появились на Земле 125 - 150 миллионов лет назад, что соответствует юрскому и меловому периодам мезозойской эры. Очевидно, предками первых покрытосеменных были какие-то вымершие голосеменные растения. Быстрое распространение покрытосеменных растений повлияло на животный мир Земли, обеспечив эволюцию насекомых, птиц и млекопитающих. Общепринятой системы классификации покрытосеменных нет. Однако их принято делить на 2 класса: двудольные и однодольные. Из общего числа видов цветковых растений двудольные составляют около 80%. Они насчитывают 292 семейства. Класс однодольных объединяет около 20% цветковых растений и насчитывает около 80 семейств.
Покрытосеменные характеризуются рядом четких признаков, отличающих только эту группу растений. Важнейшей отличительной особенностью является:
- наличие у них пестика и развивающегося из него плода;
-наличие рыльца - органа, улавливающего пыльцу и обеспечивающего прорастание пыльцевых зерен. После оплодотворения завязь разрастается и постепенно превращается в плод, а семяпочки развиваются в семена, скрытые внутри плода, почему растения и получили название – покрытосеменные;
- наличие у них цветка, снабженного околоцветником, в связи с чем их называют цветковыми. Цветок представляет собой своеобразный видоизмененный спороносный побег. Для покрытосеменных характерно огромное разнообразие в строении цветков.
infourok.ru
15.Особенности полового размножения красных водорослей. Циклы воспроизведения.
Размножение красных водорослей чрезвычайно сложный и многообразный процесс, отличающий их от других групп водорослей. Различные формы бесполого размножения, сложное строение половой системы, особенно женского репродуктивного органа, особенности развития зиготы, а также многообразие циклов развития — все эти признаки не только придают своеобразие , но являются основой, на которой строится классификация багрянок, их разделение на порядки, семейства, роды. Поэтому, чтобы составить целостное представление о красных водорослях, нельзя не остановиться на особенностях их размножения. Собственно говоря, сложным размножением характеризуются представители класса флоридеевых; у бангиевых строение репродуктивной системы намного проще. Вегетативное размножение флоридеевых происходит посредством образования дополнительных побегов, которые берут начало от подошвы или от стелющихся ветвей. Вертикальное слоевище в определенный период отмирает и остается только базальная часть растения, которая через какое-то время прорастает, образуя новые побеги. Самые примитивные багрянки из класса бангиевых размножаются только вегетативно. У одноклеточных и колониальных форм вегетативное размножение состоит в делении клетки на две или более дочерние клетки. Бесполое размножение. Для бесполого размножения служат различного рода споры. У низкоорганизованных багрянок бесполое размножение осуществляется моноспорами. Они характерны для всех представителей класса бангиевых, тогда как среди флоридеевых известны за очень редкими исключениями только в порядке немалиевых. Моноспорами называют споры, которые образуются по одной в клетке в результате превращения всего протопласта. Моноспоры не имеют жгутика и оболочки, после выхода из материнской клетки они способны к амебоидному движению. У бангиевых моноспоры образуются в любой клетке слоевища и до своего выхода не отличаются от вегетативных клеток. В классе флоридеевых (там, где они встречаются) моноспорангии образуются на веточках ограниченного роста. У водорослей, построенных по типу свободных нитей, в моноспорангии превращаются конечные клетки коротких боковых веточек. Иногда они образуются группами по два-три на одной и той же клетке-ножке (рис. 151, 1). У водорослей, имеющих более или менее плотное строение и дифференцированный коровой слой, в моноспорангии превращаются внешние клетки ассимиляционных нитей. Моноспоры отличаются от вегетативных клеток яйцевидной или шаровидной формой, богатым содержимым и интенсивной окраской. Созревшие моноспорангии открываются на вершине; голые моноспоры выходят в окружающую среду, через некоторое время одеваются оболочкой и начинают прорастать.
16. Подцарство миксомицеты. Строение. Образ жизни. Капустная кила. Цикл воспроизведения. Меры борьбы.
Миксомице́ты — тип слизевиков (грибоподобных организмов). Вегетативная фаза состоит из плазмодия (многоядерной подвижной протоплазматической массы, лишённой клеточных стенок) или псевдоплазмодия (агрегата голых одноядерных амёбоидных клеток, сохраняющих свою индивидуальность). Питание как голозойное, так и абсорбтивное. Жгутиконосные клетки, когда они имеются, обычно несут два неодинаковых жгутика. Споры и спорангии (вместилища спор) обычно многочисленные. Включает один отдел (тип) слизистые грибы, или миксомицеты ( Мухомусоta).
На определённой стадии жизненного цикла все миксомицеты имеют вид плазмодия или псевдоплазмодия. Классы Myxomycetes и Protosteliomycetes, имеющие плазмодий, относят к плазмодиальным (неклеточным) слизевикам, а Dictyosteliomycetes, которые имеют псевдоплазмодий — к клеточным. У большинства видов он виден невооружённым глазом и способен двигаться. Из плазмодия или псевдоплазмодия формируются спороношения, которые часто напоминают внешним видом плодовые тела грибов. Споры прорастают подвижными клетками — зооспорами или миксамёбами, из которых различными путями образуется плазмодий или псевдоплазмодий.
Слизевики — гетеротрофы. Зооспоры, миксамёбы и плазмодии способны питаться осмотрофно(всасыванием питательных веществ через клеточную мембрану) и (или) путём эндоцитоза (захватывать внутрь клетки пузырьки с частицами пищи).
Все миксомицеты свободноживущие и живут в наземных местообитаниях.
Капустная кила - это такой миксомицет, который проводит почти всю свою жизнь в тканях других растений.
studfiles.net
Цикл воспроизведения цветковых растений. страница 2
В жизненном цикле высших растений выделяют смену двух поколений: гаметофита и спорофита. Преобладающим поколением является спорофит обычно представляющий собой крупное листостебельное растение, существующее достаточно длительный срок. Изменения, произошедшие в цикле развития у покрытосеменных, дал им широкое распространение во всех климатических зонах.
1.Общая характеристика цикла воспроизведения цветковых растений
В цикле развития покрытосемянных присутствуют альтернативные процессы - оплодотворение и мейоз, обусловливающие смену ядерных фаз. В результате первого в типичных случаях происходит смена гаплоидной фазы фазой диплоидной, а второго - смена диплоидной фазы фазой гаплоидной. Диплоидную фазу в циклах развития называют спорофитом, гаплоидную - гаметофитом. У этой группы растений гаплоидное поколение сильно редуцировано и представлено пыльцевыми зернами и зародышевым мешком, само же растение диплоидно.
У высших растений гаметангии, т.е. особые органы в которых формируются специализированные гаметы, как и спорангии, многоклеточны. Гаметы всегда голые (не имеют твердой оболочки) и часто бывают снабжены жгутиками. Однако, у большинства семенных растений мужские гаметы (спермии) в ходе эволюции утратили жгутики. Половой процесс – оогамный.
Главное отличие семенных растений от высших споровых заключается в том, что гаметофиты, особенно женский, утратили способность к самостоятельному существованию и живут исключительно за счет спорофита. При этом осуществлено важное приспособление к наземному образу жизни, давшее
Огромное преимущество семенным растениям перед споровыми: отрыв от водной среды при оплодотворении.
2.Характеристика гаплоидной фазы
ГАМЕТОФИТ (от гаметы и греч. phyton - растение) - половое поколение, этап жизненного цикла растений; чередуется со спорофитом. Образуется из споры, содержит гаплоидный набор хромосом; продуцирует гаметы в гаметангиях. У покрытосеменных гаметофитов редуцирован до пылинки (мужской гаметофит) и зародышевого мешка (женский гаметофит).
Следует отметить, что на одном гаметофите одновременно могут развиваться как мужские половые органы, так и женские. Такой гаметофит называют однодомным (обоеполым). В других случаях гаметофиты растений, относящихся к одному и тому же виду, формируют либо только мужские органы, либо только женские. Такие гаметофиты называют двудомными (раздельнополыми — мужскими и женскими). Раздельнополые гаметофиты свойственны всем семенным растениям, у которых мужские гаметофиты развиваются из микроспор, образующихся в пыльниках, а женские — из мегаспор, образующихся в семяпочках.
Мужской гаметофит
Все развитие мужского гаметофита, включая образование мужских гамет, сводится лишь к двум митотическим делениям. Первое из этих делений происходит всегда под защитой оболочки микроспоры, которая превращается в новое образование — пыльцевое зерно. Второе деление совершается в одних случаях в пыльцевом зерне, а в других — лишь позднее, в пыльцевой трубке. В результате зрелые пыльцевые зерна бывают двухклеточными или трехклоточными, причем двухклеточные встречаются значительно чаще, чем трехклеточные, и известны приблизительно у 70% цветковых растений. Еще в 1926 г. известный немецкий эмбриолог растений П. Шюргоф пришел к выводу, что в процессе эволюции трехклеточная пыльца возникла из двухклеточной. Он справедливо считал, что прохождение обоих делений внутри пыльцевого зерна, в то время как последнее еще не отделилось от материнского растения, является прогрессивным, так как в этом случае развитие мужского гамотофита происходит под надежной защитой спородермы и более быстро. Действительно, двухклеточная пыльца характерна для многих относительно примитивных групп, в том число для всех представителей порядков магнолиевых, лавровых, нимфейных, лютиковых, а трехклеточная пыльца встречается чаще у более подвинутых порядков, в том числе у всех сложноцветных и злаков.
infourok.ru
Цикл воспроизведения цветковых растений. страница 4
В первом случае образование клеточных перегородок подавляется только при втором делении мейоза. Поэтому оба ядра, вошедшие в состав жизнеспособной клетки диады, принимают участие в образовании женского гаметофита. Двухъядерная диада — это ценоцит, продукт слияния (вернее, разделения) двух потенциальных мегаспор. Поскольку каждый такой ценоцит соответствует по происхождению паре обособившихся мегаспор, его рассматривают как двухспоровый ценоцит, а развивающийся из него женский гаметофит — как биспорический (двуспоровый). Биспорический женский гаметофит наблюдается у лука, пролески (Scilia), ландыша, амариллисовых, частуховых и некоторых других растений. Развитие такого женского гаметофита происходит более сокращенным путем и для образования восьмиядерной стадии требуется всего лишь два митотических деления.
Наконец, у некоторых растений, таких, как, поперомия, ильм, майник, рябчик, тюльпан, лилия, гусиный лук, плюмбаго, адокса и другие, образование клеточных перегородок подавляется при обоих мейотических делениях. Вся материнская клетка мегаспор превращается здесь в один четырехспоровый ценоцит, или тетрацепоспору. Четырехспоровый ценоцит соответствует четырем перазделившимся мегаспорам, почему женские гаметофиты этого типа называются тетраспорическими (четырехспоровыми). Для образования восьмиядерной стадии женского гаметофита требуется лишь одно митотическое деление ядер четырехспорового ценоцита.
Согласно И. Д. Романову, в отличие от моноспорического женского гаметофита, би - и тетраспорические называются ценоспорическими.
3.Характеристика диплоидной фазы
СПОРОФИТ (от споры и греч. phyton - растение) - бесполое поколение или этап жизненного цикла растений от зиготы до образования споры. В цикле развития чередуется с гаметофитом; характерен двойной (диплоидный) набор хромосом. Образуется С. после оплодотворения — слияния муж. и жен. гаплоидных гамет в диплоидную зиготу, из которой развивается многоклеточный зародыш, дающий начало взрослому растению. Каждая клетка. С, как и зигота, обычно содержит двойной набор хромосом. На С. формируются органы бесполого размножения — спорангии, в которых после редукционного деления (мейоза) образуются мейоспоры с гаплоидным набором хромосом, дающие начало половому поколению и гаметофиту, все клетки которого гаплоидны. В жизненных циклах разных групп растений С. выражен различно. У всех высших растений цикл развития гетероморфный. У папоротниковидных, голосеменных и цветковых С. доминирует (все высшие зелёные растения, за исключением моховидных представлены С). У моховидных С. сведён к органу (спорогонию), развивающемуся на гаметофите и не способному существовать автономно.
Заключение
Таким образом, с повышением уровня организации живых существ наблюдается увеличение диплоидной фазы, которая создает целый ряд биологических преимуществ для жизнедеятельности. В частности, диплоидность позволяет нейтрализовать многие вредные мутации, поскольку большинство из них рецессивны. В соматических клетках действие таких рецессивных аллелей подавляется доминантными нормальными аллелями.
infourok.ru
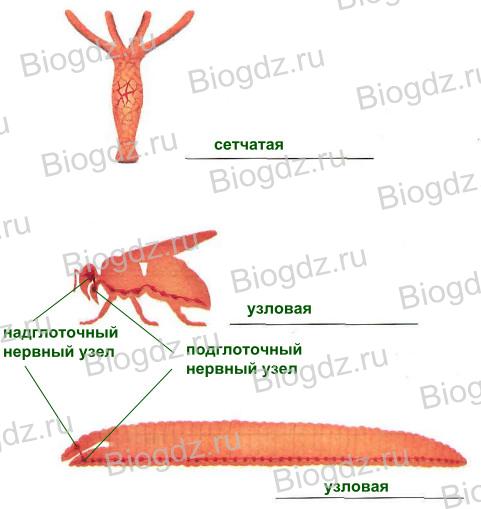
Понятие о цикле воспроизведения растений и смене ядерных фаз. Два типа без правильного чередования и с чередованием поколений. Направление циклов воспроизведения
Половое размножение осуществляется слиянием (копуляцией) двух половых клеток (гамет) и их ядер. Зигота, получающаяся в результате копуляции, у многих водорослей и низших грибов одевается толстой оболочкой и превращается в так называемую покоящуюся спору, которая лишь после некоторого периода покоя дает, так или иначе прорастая, новое растение. У большинства же зигота немедленно начинает делиться и образует новое растение или зародыш его; последний у семенных растений временно задерживается в дальнейшем развитии. Это новое растение, развившееся из зиготы, у одних вполне похоже на материнское, у других более или менее резко отличается от него. У большинства оно способно размножаться вегетативно; кроме того, у очень многих растений на нем или в нем образуются бесполым путем специальные клетки, служащие для размножения, носящие нередко различные названия и объединяемые под общим наименованием спор бесполого размножения. Они отделяются от материнского растения, рассеиваются и или непосредственно, или после некоторого периода покоя прорастают в новые растения, тождественные с материнским или непохожие на него. В дальнейшем опять наступает половое размножение. При этом у одних имеется правильное чередование полового и бесполого размножения; у других это в очень большой степени зависит от внешних условий, и половое размножение происходит после некоторого накопления запасных веществ, часто при наступлении неблагоприятных условий существования.
При слиянии половых ядер хромосомы их не сливаются, получающееся в результате слияния так называемое копуляционное ядро будет иметь двойной набор (2n) хромосом и называться диплоидным, ядра гамет, имеющие вдвое меньшее число хромосом (п), называют гаплоидными3. Каждый вид растений характеризуется определенным диплоидным и вдвое меньшим гаплоидным числом хромосом. Переход от гаплоидного к диплоидному совершается при оплодотворении, обратный - от диплоидного к гаплоидному - при редукционном делении ядра. Последнее происходит у разных групп растений в различной, но всегда строго определенной стадии их развития. Таким образом, в жизненном цикле каждого растения, имеющего половое размножение, мы находим так называемую смену ядерных фаз - гаплоидной и диплоидной. Растение с гаплоидным числом хромосом называют гаплонтом, с диплоидным - диплонтом. У многих водорослей и грибов в цикле развития доминирует гаплонт; например, у большинства зеленых водорослей диплоидна только зигота; первое же деление ее ядра в начале прорастания есть редукционное, и вся вновь развивающаяся водоросль будет гаплоидна. Прямой противоположностью этому является полное доминирование диплонта; например, у бурых фукусовых водорослей вся водоросль диплоидна, редукционное деление происходит при образовании гамет, а при слиянии их сейчас же восстанавливается диплоидная фаза. Противоположностью указанным случаям полного доминирования одной фазы является цикл развития многих красных, некоторых бурых, и немногих зеленых водорослей. У них водоросль в гаплоидной фазе размножается половым путем - дает гаметы, из слияния которых получается зигота. Последняя при прорастании образует новую водоросль с диплоидным числом хромосом, вполне похожую на гаплонта или более или менее, у некоторых даже очень резко, отличающуюся от него; эта диплоидная форма размножается бесполым путем - спорами, при образовании которых происходит редукционное деление. Из этих спор развивается гаплоидная водоросль, образующая гаметы, с которой мы начали знакомство с циклом развития их; зигота вновь дает диплоидную водоросль, размножающуюся бесполым путем, и т. п. Таким образом, здесь мы видим не только смену гаплоидной и диплоидной фаз, но и соответствующее им чередование поколений - полового и бесполого, ведущих каждое самостоятельный образ жизни и размножающихся половым или бесполым способом. Половое поколение называют гаметофитом, бесполое - спорофитом.
У высших растений мы имеем совпадающее со сменой ядерных фаз подобное же чередование поколений (смену генераций), причем у них одно какое-либо поколение доминирует в цикле развития, а другое и морфологически и по продолжительности его жизни развито слабее (это же бывает и у некоторых водорослей) и у многих не способно к самостоятельной жизни. У мхов доминирует гаметофит, а спорофит в виде коробочки со спорами живет на гаметофите. У папоротников, наоборот, доминирует спорофит - сам папоротник, размножающийся спорами, а гаметофит представлен очень маленькой, но живущей самостоятельно зеленой пластинкой, так называемым заростком. У семенных растений доминирует тоже спорофит - все растение, а гаметофит очень редуцирован, живет паразитически на спорофите и может быть узнан только после тщательного сравнительного морфологического сопоставления.Термин "чередование поколений" несколько неудачен, так как в обыденной жизни под "поколением" обычно понимают группу особей, проходящих все свое развитие (онтогенез) от рождения до смерти самостоятельно и сходно с родителями. Спорофит же и гаметофит у растений представляют собой различные фазы (или этапы) онтогенеза одной особи. Но понятие о чередовании поколений и сам этот термин имеют столетнюю давность, прочно вошли во всеобщее употребление у ботаников, и нет оснований заменять их чем-либо другим.
Необходимо еще отметить, что было бы ошибочно различия в организации и величине гаметофита и спорофита, имеющие место у большинства растений, ставить только в зависимость от различия в числе их хромосом.
У красных водорослей, многих бурых, некоторых зеленых (ульвовых) водорослей спорофит и гаметофит одинаковы по величине и форме, хотя и резко различны по числу хромосом. У большинства высших растений спорофит значительно крупнее и долговечнее гаметофита, но у мхов эти отношения обратные, хотя у тех и других число хромосом у спорофита всегда вдвое больше, чем у гаметофита.
megaobuchalka.ru
Цикл воспроизведения цветковых растений. страница 3
Женский гаметофит цветковых растений обычно называется зародышевым мешком. Возникновение этого названия восходит к тем временам, когда еще не была ясна морфологическая природа чрезвычайно своеобразного женского гаметофита цветковых растений. В настоящее время оба термина употребляют как синонимы.
У большинства цветковых растений (вероятно, не менее чем у 80%) начало женскому гаметофиту дает одна из четырех потенциальных мегаспор тетрады путем трех последовательных митотических делений. Этот тип образования женского гаметофита называется моноспорическим. Его развитие происходит следующим образом (рис. 28,Б).

В результате первого деления ядра функционирующей мегаспоры образуются два ядра, которые расходятся к полюсам (микропилярному и халазальному) сильно удлиняющейся мегаспоры, а между ними образуется крупная вакуоль. Далее каждое из этих двух ядер делится синхронно еще два раза и в результате у каждого полюса образуются по четыре ядра. Это восьмиядерная стадия развития женского гаметофита. От каждой из двух полярных четверок входит по одному ядру в центральную часть зародышевого мешка. Эти так называемые полярные ядра сближаются в центральной части и сливаются или сразу же, или только перед оплодотворением, а иногда и вовсе не сливаются. В результате слияния полярных ядер образуется ядро центральной клетки женского гаметофита, называемое часто вторичным ядром зародышевого мешка.
В дальнейшем (в процессе последнего митоза) возникают клеточные перегородки, и свободноядерная стадия развития женского гаметофита переходит в клеточную стадию. У микропилярного полюса образуются три клетки яйцевого аппарата, у противоположного (халазального) — три клетки, называемые антиподами, а между этими двумя группами клеток — центральная клетка (содержащая два свободных полярных ядра или одно вторичное ядро). Три клетки яйцевого аппарата дифференцируются на одну яйцеклетку и две клетки, называемые синергидами (от греч. synergos — вместе действующие) или вспомогательными клетками. Таково строение так называемого нормального моноспорического женского гаметофита.
В процессе эволюции этого основного для цветковых растений типа женского гамотофита возникли зародышевые мешки других типов, образуемые двумя или даже четырьмя мегаспорами. Произошло сокращение митотических делений до двух или одного, а также изменилось распределение ядер. Разные сочетания этих изменений обусловили возникновение нескольких типов женского гаметофита, которые отличаются как числом ядер (4, 8, 16), числом клеточных групп и полярных ядер (1, 2, 4, 7—14), так и числом клеток в группах (например, яйцевой аппарат может состоять из 1, 2, 3, 5 и 7 клеток), а также другими признаками.
Основной особенностью моноспорического типа образования женского гаметофита является последовательное заложение клеточных перегородок при мейотических делениях и образование обособленных мегаспор. Совершенно иной характер имеет мегаспорогенез, предшествующий образованию биспорического (двухспорового) и тетраспорического (четырехспорового) гаметофитов. Изменения в мегаспорогенезе заключаются в подавлении заложения клеточной перегородки при мейотическом делении, вследствие чего вместо тетрад мегаспор возникают неклеточные образования, называемые ценоцитами (от греч. koinos — общий и kylos — сосуд, клетка), которые и развиваются далее в женские гаметофиты. Здесь возможны два варианта.
infourok.ru