
Содержание
Грибы. Бактерии. Лишайники. — огэ биология
Часть А (только один правильный ответ)
1. Клетка бактерии характеризуется наличием…
1) ядра
2) митохондрий
3) плазматической мембраны
4) аппарата Гольджи
2. Размножение бактерий происходит…
1) митозом
2) амитозом
3) мейозом
4) партеногенезом
3. Споры бактерий необходимы для…
1) размножения
2) запасания питательных веществ
3) пережидания неблагоприятных условий
4) питания
4. Тип питания бактерий:
1) автотрофный
2) гетеротрофный
3) автотрофный или гетеротрофный
4) миксотрофный
5. Оптимальным условием для жизни всех бактерий является…
1) кислород
2) солнечный свет
3) температура свыше 600 °С
4) щелочная или нейтральная среда
6. Для всех грибов характерно…
1) наличие плодового тела
2) автотрофное питание
3) неограниченный рост
4) мицелий
7. Какой признак является общим и для грибов, и для растений?
1) неподвижность
2) запасное вещество — гликоген
3) гетеротрофы
4) наличие тканей и органов
8. Основной признак грибов-сапрофитов:
Основной признак грибов-сапрофитов:
1) вызывают болезни
2) питаются мертвой органикой
3) способны к хемосинтезу
4) не имеют клеток
9. Назовите гриб, относящийся к паразитам:
1) дрожжи
2) опенок
3) фитофтора
4) пеницилл
10. Что такое микориза?
1) мицелий гриба
2) плодовое тело
3) симбиоз гиф гриба с корнями растений
4) оболочка клетки гриба
11. Какие грибы вызывают брожение?
1) плесневые грибы
2) дрожжи
3) шляпочные грибы
4) хлебная ржавчина
12. Назовите смертельно ядовитый гриб:
1) ложный опенок
2) сморчок
3) бледная поганка
4) масленок
13. Из предложенных организмов выберите лишайник:
1) кукушкин лен
2) пармелия
3) пеницилл
4) фитофтора
14. Какие организмы образуют лишайник?
1) мхи и водоросли
2) мхи и грибы
3) грибы и бактерии
4) грибы и цианобактерии
15. Что для лишайников нехарактерно?
1) медленный рост
2) требовательность к чистоте окружающей среды
3) гетеротрофное питание
4) размножение частями слоевища
Часть В
В заданиях В1 и В2 выберите три верных ответа из шести.
1. Бактерии, как и растения, …
1) способны к фотосинтезу
2) не имеют ядра
3) для размножения используют споры
4) в клетке содержат рибосомы и цитоплазму
5) относятся к прокариотам
6) имеют оболочку
2. Грибы-паразиты…
1) вызывают болезни растений и животных
2) способны к брожению
3) размножаются спорами
4) являются гетеротрофами
5) к ним относятся плесневые грибы
6) не имеют гиф
При выполнении задания В3 И В4 установите соответствие между содержанием первого и второго столбцов.
3. Установите соответствие между признаками и царствами живых организмов
ПРИЗНАК ЦАРСТВО
А) не имеют оформленного ядра 1) Грибы
Б) есть митохондрии 2) Бактерии
В) способны к хемосинтезу
Г) автотрофы и гетеротрофы
Д) размножаются спорами
Е) образуют микоризу
4. Установите соответствие между признаками и группами грибов.
Установите соответствие между признаками и группами грибов.
ПРИЗНАК ГРУППА ГРИБОВ
А) паразитический тип питания 1) Плесневые грибы
Б) развиваются на хлебе 2)Спорынья
В) развиваются на злаковых растениях
Г) синтезируют антибиотики
Д) есть плодовые тела
Часть С
1. Почему лишайники выделены в отдельную группу организмов?
2. Любая из бактерий способна размножаться делением каждые 15-20 минут. Потомки только одной бактерии за 1 сутки могли бы покрыть всю Землю слоем толщиной не менее 1 метра. Почему же этого в природе не происходит?
3. Почему при внесении на рисовые поля цианобактерий урожайность риса возрастает на 10-15 %?
4. Многие грибы-паразиты практически не встречаются на самоопыляемых растениях. Как вы можете объяснить это явление?
Биология 6 класс. Рабочая тетрадь.
 Растения. Бактерии. Грибы. Лишайники. К учебнику Трайтака Д.И., Трайтак Н.Д. «Биология. Растения. Бактерии. Грибы. Лишайники. 5-6 классы. Часть 2». ФГОС
Растения. Бактерии. Грибы. Лишайники. К учебнику Трайтака Д.И., Трайтак Н.Д. «Биология. Растения. Бактерии. Грибы. Лишайники. 5-6 классы. Часть 2». ФГОС
Биология 6 класс. Рабочая тетрадь. Растения. Бактерии. Грибы. Лишайники. К учебнику Трайтака Д.И., Трайтак Н.Д. «Биология. Растения. Бактерии. Грибы. Лишайники. 5-6 классы. Часть 2». ФГОС
- Описание
- Характеристики
- Отзывы (0)
ISBN: 978-5-00031-024-3
Автор: Бодрова Н.Ф.
Издательство: Учитель г.Воронеж
Год издания: 2014
Рабочая тетрадь разработана в соответствии с ФГОС и предназначена для организации самостоятельной работы учащихся в процессе обучения биологии в 6 классе по учебнику Трайтака Д.И., Трайтак Н.Д. «Биология. Бактерии. Грибы. Лишайники. 5-6 классы» в 2 частях (часть 2) (издательство «Мнемозина»).
Лишайники. 5-6 классы» в 2 частях (часть 2) (издательство «Мнемозина»).
Работа с таблицами, схемами и рисунками, представленными в тетради, способствует формированию и развитию универсальных учебных действий.
| Год Выпуска: | 2014 |
| Автор: | Бодрова Н.Ф. |
| Издательство: | Учитель г.Воронеж |
Пока нет отзывов
Оставить отзыв
Все поля обязательны к заполнению
Перед публикацией отзывы проходят модерацию
Формат
-
70х100 1/16 (4)
Год Выпуска
-
2018 (3)
-
2019 (1)
-
2011 (6)
-
2014 (6)
Автор
-
Бодрова Н. Ф. (16)
Ф. (16)
 Ф. (16)
Ф. (16)
Издательство
-
Учитель г.Воронеж (9)
-
МЕТОДА (3)
-
М-КНИГА (4)
Размер
-
235х165 мм (4)
Страниц
-
64 (1)
-
48 (3)
Тип обложки
-
мягкий (4)
Переплёт
-
мягкий переплет (крепление скрепкой или клеем) (4)
Класс
-
5 (3)
-
6 (1)
Экология грибов | OpenStax Biology 2e
Цели обучения
К концу этого раздела вы сможете делать следующее:
- Описывать роль грибов в различных экосистемах
- Описать мутуалистические взаимоотношения грибов с корнями растений и фотосинтезирующими организмами
- Опишите полезные отношения между некоторыми грибами и насекомыми
Грибы играют решающую роль в постоянно меняющемся «балансе» экосистем. Они колонизируют большинство мест обитания на Земле, предпочитая темные и влажные условия. Они могут процветать в, казалось бы, враждебной среде, такой как тундра, благодаря наиболее успешному симбиозу с фотосинтетическими организмами, такими как водоросли, для производства лишайников. В их сообществах грибы не так очевидны, как крупные животные или высокие деревья. Подобно бактериям, они действуют за кулисами как главные разлагатели. Благодаря своему универсальному метаболизму грибы расщепляют органические вещества, которые в противном случае не могли бы быть переработаны.
Они колонизируют большинство мест обитания на Земле, предпочитая темные и влажные условия. Они могут процветать в, казалось бы, враждебной среде, такой как тундра, благодаря наиболее успешному симбиозу с фотосинтетическими организмами, такими как водоросли, для производства лишайников. В их сообществах грибы не так очевидны, как крупные животные или высокие деревья. Подобно бактериям, они действуют за кулисами как главные разлагатели. Благодаря своему универсальному метаболизму грибы расщепляют органические вещества, которые в противном случае не могли бы быть переработаны.
Среда обитания
Хотя грибы в основном связаны с влажной и прохладной средой, обеспечивающей запас органических веществ, они колонизируют удивительно разнообразные среды обитания, от морской воды до человеческой кожи и слизистых оболочек. Хитриды встречаются в основном в водной среде. Другие грибы, такие как Coccidioides immitis , вызывающий пневмонию при вдыхании спор, процветают в сухой и песчаной почве на юго-западе США. В океане живут грибы, паразитирующие на коралловых рифах. Однако большинство представителей царства грибов растут на лесной подстилке, где темная и влажная среда богата разлагающимися остатками растений и животных. В этих средах грибы играют важную роль в качестве разлагателей и переработчиков, позволяя представителям других царств получать питательные вещества и жить.
В океане живут грибы, паразитирующие на коралловых рифах. Однако большинство представителей царства грибов растут на лесной подстилке, где темная и влажная среда богата разлагающимися остатками растений и животных. В этих средах грибы играют важную роль в качестве разлагателей и переработчиков, позволяя представителям других царств получать питательные вещества и жить.
Разлагатели и переработчики
Пищевая сеть была бы неполной без организмов, разлагающих органическое вещество ((Рисунок)). Некоторые элементы, такие как азот и фосфор, требуются биологическим системам в больших количествах, но в окружающей среде их мало. Действие грибов высвобождает эти элементы из разлагающихся веществ, делая их доступными для других живых организмов. Микроэлементы, присутствующие в небольших количествах во многих средах обитания, необходимы для роста и оставались бы связанными с гниющими органическими веществами, если бы грибки и бактерии не возвращали их в окружающую среду посредством своей метаболической активности.
Рисунок 1. Кротовидные грибы. Грибы являются важной частью круговорота питательных веществ в экосистеме. Эти трутовки, растущие на боку дерева, являются плодовыми структурами базидиомицета. Они получают питательные вещества через свои гифы, которые вторгаются в ствол дерева и разлагают его. (кредит: Кори Занкер)
Способность грибов разлагать многие большие и нерастворимые молекулы обусловлена их способом питания. Как было показано ранее, пищеварение предшествует приему пищи. Грибы производят различные экзоферменты для переваривания питательных веществ. Ферменты либо высвобождаются в субстрат, либо остаются связанными снаружи клеточной стенки грибов. Крупные молекулы расщепляются на мелкие, которые транспортируются в клетку с помощью системы белков-переносчиков, встроенных в клеточную мембрану. Поскольку движение малых молекул и ферментов зависит от присутствия воды, активный рост зависит от относительно высокого процента влажности окружающей среды.
Как сапробионты, грибы помогают поддерживать устойчивую экосистему для животных и растений, живущих в одной среде обитания. Помимо пополнения окружающей среды питательными веществами, грибы напрямую взаимодействуют с другими организмами полезными, а иногда и вредными способами ((рисунок)).
Рисунок 2. Полочные грибы. Шельфовые грибы, называемые так потому, что они растут на деревьях в штабелях, атакуют и переваривают ствол или ветви дерева. В то время как некоторые шельфовые грибы встречаются только на мертвых деревьях, другие могут паразитировать на живых деревьях и в конечном итоге вызывать смерть, поэтому они считаются серьезными патогенами деревьев. (кредит: Кори Занкер)
Взаимные отношения
Симбиоз — это экологическое взаимодействие между двумя организмами, которые живут вместе. Это определение не описывает тип или качество взаимодействия. Когда оба члена ассоциации получают выгоду, симбиотические отношения называются мутуалистическими. Грибы образуют мутуалистические ассоциации со многими типами организмов, включая цианобактерии, водоросли, растения и животных.
Грибы образуют мутуалистические ассоциации со многими типами организмов, включая цианобактерии, водоросли, растения и животных.
Мутуализм грибов и растений
Одной из самых замечательных ассоциаций между грибами и растениями является установление микориза . Микориза, происходящая от греческих слов myco , означающих гриб, и rhizo , означающих корень, относится к грибковому партнеру мутуалистической ассоциации между корнями сосудистых растений и их симбиотическими грибами. Почти 90 процентов всех видов сосудистых растений имеют микоризных партнеров. В микоризной ассоциации грибной мицелий использует свою разветвленную сеть гиф и большую площадь поверхности, контактирующую с почвой, для направления воды и минералов из почвы в растение. Взамен растение поставляет продукты фотосинтеза для подпитки метаболизма гриба.
Существует несколько основных типов микоризы. Эктомикоризы («внешние» микоризы) зависят от грибов, обволакивающих корни оболочкой (называемой оболочкой). Гифы прорастают из мантии в корень и покрывают внешние слои корневых клеток сетью гиф, называемой сетью Хартига ((Рисунок)). Гриб-партнер может принадлежать к Ascomycota, Basidiomycota или Zygomycota. Эндомикоризы («внутри» микоризы), также называемые арбускулярными микоризами , образуются, когда грибы растут внутри корня в виде разветвленной структуры, называемой arbuscule (от латинского «деревца»). Все грибковые партнеры эндомикоризных ассоциатов принадлежат к Glomeromycota. Грибковые арбускулы проникают в клетки корня между клеточной стенкой и плазматической мембраной и являются местом метаболического обмена между грибом и растением-хозяином ((Рисунок) b и (Рисунок) b ). Орхидеи полагаются на третий тип микоризы. Орхидеи — это эпифиты, которые обычно производят очень маленькие семена, переносимые по воздуху, без особого хранения для поддержания прорастания и роста. Их семена не прорастут без микоризного партнера (обычно базидиомицета).
Гифы прорастают из мантии в корень и покрывают внешние слои корневых клеток сетью гиф, называемой сетью Хартига ((Рисунок)). Гриб-партнер может принадлежать к Ascomycota, Basidiomycota или Zygomycota. Эндомикоризы («внутри» микоризы), также называемые арбускулярными микоризами , образуются, когда грибы растут внутри корня в виде разветвленной структуры, называемой arbuscule (от латинского «деревца»). Все грибковые партнеры эндомикоризных ассоциатов принадлежат к Glomeromycota. Грибковые арбускулы проникают в клетки корня между клеточной стенкой и плазматической мембраной и являются местом метаболического обмена между грибом и растением-хозяином ((Рисунок) b и (Рисунок) b ). Орхидеи полагаются на третий тип микоризы. Орхидеи — это эпифиты, которые обычно производят очень маленькие семена, переносимые по воздуху, без особого хранения для поддержания прорастания и роста. Их семена не прорастут без микоризного партнера (обычно базидиомицета). После того, как питательные вещества в семенах истощаются, грибковые симбионты поддерживают рост орхидеи, обеспечивая ее необходимыми углеводами и минералами. Некоторые орхидеи остаются микоризными на протяжении всего жизненного цикла.
После того, как питательные вещества в семенах истощаются, грибковые симбионты поддерживают рост орхидеи, обеспечивая ее необходимыми углеводами и минералами. Некоторые орхидеи остаются микоризными на протяжении всего жизненного цикла.
Art Connection
Рисунок 3. Два типа микоризы. (а) Эктомикоризы и (б) арбускулярные или эндомикоризы имеют разные механизмы взаимодействия с корнями растений. (кредит b: М.С. Турмел, Университет Манитобы, факультет растениеводства)
Если бы в почве отсутствовали симбиотические грибы, как вы думаете, какое влияние это оказало бы на рост растений?
Показать раствор
Рисунок 4. Микориза. (а) Инфицирование корней Pinus radiata (сосны Монтерей) гифами Amanita muscaria (мухоморы) приводит к тому, что сосна производит много маленьких разветвленных корешков. Гифы Amanita покрывают эти маленькие корни белой мантией. (b) Споры (круглые тела) и гифы (нитевидные структуры) очевидны на этой световой микрофотографии арбускулярной микоризы грибком на корне растения кукурузы. (кредит а: модификация работы Рэнди Молина, USDA; кредит b: модификация работы Сары Райт, USDA-ARS; данные масштабной линейки от Мэтта Рассела)
(кредит а: модификация работы Рэнди Молина, USDA; кредит b: модификация работы Сары Райт, USDA-ARS; данные масштабной линейки от Мэтта Рассела)
Другие примеры мутуализма между грибами и растениями включают эндофиты: грибы, которые живут внутри тканей, не повреждая растения-хозяина. Эндофиты выделяют токсины, отпугивающие травоядных, или придают устойчивость к стрессовым факторам окружающей среды, таким как заражение микроорганизмами, засуха или тяжелые металлы в почве.
Evolution Connection
Совместная эволюция наземных растений и микоризы
Как мы видели, микориза является грибковым партнером взаимовыгодной симбиотической ассоциации, которая возникла совместно между корнями сосудистых растений и грибами. Хорошо подтвержденная теория предполагает, что грибы сыграли важную роль в эволюции корневой системы растений и способствовали успеху покрытосеменных растений. Мохообразные (мхи и печеночники), которые считаются наиболее предковыми растениями и первыми, кто выжил и адаптировался на суше, имеют простые подземные ризоиды, а не настоящую корневую систему, и поэтому не могут выжить в засушливых районах. Однако у некоторых мохообразных есть арбускулярная микориза, а у некоторых нет.
Однако у некоторых мохообразных есть арбускулярная микориза, а у некоторых нет.
Настоящие корни впервые появились у предковых сосудистых растений: сосудистые растения, которые развили систему тонких отростков от своих корней, имели селективное преимущество перед несосудистыми растениями, потому что у них была большая площадь контакта с грибами-партнерами, чем у ризоидов. мхов и печеночников. Первые настоящие корни позволили бы сосудистым растениям получать больше воды и питательных веществ из земли.
Записи окаменелостей показывают, что грибы фактически предшествовали вторжению древних пресноводных растений на сушу. Первая ассоциация между грибами и фотосинтезирующими организмами на суше включала мохоподобные растения и эндофиты. Эти ранние ассоциации возникли еще до того, как у растений появились корни. Постепенно преимущества взаимодействия эндофитов и ризоидов для обоих партнеров привели к появлению современной микоризы: около 90 процентов современных сосудистых растений связаны с грибами в своей ризосфере.
Грибы, вовлеченные в микоризу, обладают многими характеристиками предковых грибов; они производят простые споры, мало разнообразны, не имеют полового репродуктивного цикла и не могут жить вне микоризной ассоциации. Растения выиграли от ассоциации, потому что микориза позволила им переместиться в новые места обитания и позволила увеличить поглощение питательных веществ, что дало им огромное селективное преимущество перед растениями, которые не устанавливали симбиотических отношений.
Лишайники
Лишайники имеют различные цвета и текстуры ((Рисунок)) и могут выживать в самых необычных и враждебных средах обитания. Они покрывают камни, надгробия, кору деревьев и землю в тундре, куда не могут проникнуть корни растений. Лишайники могут пережить длительные периоды засухи, когда они полностью высыхают, а затем быстро становятся активными, как только вода снова становится доступной.
Ссылка на обучение
Исследуйте мир лишайников, используя этот сайт Университета штата Орегон.
Рисунок 5. Лишайники. Лишайники имеют множество форм. Они могут быть (а) коркообразными, (б) волосовидными или (в) листовидными. (кредит a: модификация работы Джо Нейлор; кредит b: модификация работы «djpmapleferryman»/Flickr; кредит c: модификация работы Кори Занкера)
Важно отметить, что лишайники — это , а не единый организм , а скорее еще один замечательный пример мутуализма, при котором гриб (обычно представитель Ascomycota или Basidiomycota) живет в физических и физиологических отношениях с фотосинтетическим организмом (эукариотической водорослью или прокариотической цианобактерией) ((Рисунок)). Как правило, ни гриб, ни фотосинтезирующий организм не могут выжить в одиночку вне симбиотических отношений. Тело лишайника, называемое слоевищем, состоит из гиф, обернутых вокруг партнера по фотосинтезу. Фотосинтезирующий организм обеспечивает углерод и энергию в виде углеводов. Некоторые цианобактерии дополнительно фиксируют азот из атмосферы, внося в ассоциацию азотистые соединения. В свою очередь, гриб поставляет минералы и защищает от сухости и чрезмерного света, заключая водоросли в свой мицелий. Гриб также прикрепляет лишайник к своему субстрату.
В свою очередь, гриб поставляет минералы и защищает от сухости и чрезмерного света, заключая водоросли в свой мицелий. Гриб также прикрепляет лишайник к своему субстрату.
Рисунок 6. Строение лишайника. На этом поперечном срезе слоевища лишайника показана (а) верхняя кора гиф грибов, обеспечивающая защиту; (b) зона водорослей, где происходит фотосинтез, (c) сердцевина гиф грибов и (d) нижняя кора, которая также обеспечивает защиту и может иметь (e) ризины для прикрепления слоевища к субстрату.
Слоевище лишайников растет очень медленно, увеличивая свой диаметр на несколько миллиметров в год. И гриб, и водоросль участвуют в формировании дисперсионных единиц, называемых соредиями — скоплениями клеток водорослей, окруженными мицелием. Соредии разносятся ветром и водой и образуют новые лишайники.
Лишайники чрезвычайно чувствительны к загрязнению воздуха, особенно к аномальным уровням соединений азота и серы. Лесная служба США и Служба национальных парков могут контролировать качество воздуха, измеряя относительную численность и здоровье популяции лишайников в районе. Лишайники выполняют множество экологических ролей. Карибу и северные олени питаются лишайниками и служат укрытием для мелких беспозвоночных, которые прячутся в мицелии. В производстве текстиля ткачи использовали лишайники для окрашивания шерсти на протяжении многих веков, пока не появились синтетические красители. Пигменты, используемые в лакмусовой бумаге, также извлекаются из лишайников.
Лишайники выполняют множество экологических ролей. Карибу и северные олени питаются лишайниками и служат укрытием для мелких беспозвоночных, которые прячутся в мицелии. В производстве текстиля ткачи использовали лишайники для окрашивания шерсти на протяжении многих веков, пока не появились синтетические красители. Пигменты, используемые в лакмусовой бумаге, также извлекаются из лишайников.
Ссылка на обучение
Лишайники используются для контроля качества воздуха. Подробнее читайте на этом сайте Лесной службы США.
Мутуализм грибов и животных
Грибы развили мутуализм с многочисленными насекомыми типа членистоногих: суставноногие беспозвоночные с хитиновым экзоскелетом. Членистоногие зависят от грибка в защите от хищников и патогенов, в то время как гриб получает питательные вещества и способ распространения спор в новую среду. Одним из примеров является ассоциация между видами Basidiomycota и щитовками. Грибковый мицелий покрывает и защищает колонии насекомых. Чешуйчатые насекомые способствуют поступлению питательных веществ от зараженного растения к грибу.
Чешуйчатые насекомые способствуют поступлению питательных веществ от зараженного растения к грибу.
Во втором примере муравьи-листорезы из Центральной и Южной Америки буквально выращивают грибы. Они срезают с растений диски из листьев и складывают их в подземные сады ((Рисунок)). В этих дисковых садах выращивают грибы, которые переваривают целлюлозу в листьях, которую муравьи не могут расщепить. Как только меньшие молекулы сахара производятся и потребляются грибами, грибы, в свою очередь, становятся едой для муравьев. Насекомые также патрулируют свой сад, охотясь на конкурирующие грибы. И муравьи, и грибы выигрывают от этой мутуалистической ассоциации. Гриб получает постоянный запас листьев и свободу от конкуренции, в то время как муравьи питаются выращиваемыми ими грибами.
Рисунок 7. Муравей-листорез. Муравей-листорез переносит лист, который будет кормить выращиваемый гриб. (Фото: Scott Bauer, USDA-ARS)
Грибоядные
Для некоторых грибов важно распространение животных, поскольку они могут переносить грибковые споры на значительные расстояния от источника. Споры грибов редко полностью разлагаются в желудочно-кишечном тракте животного, и многие из них способны прорастать при попадании в фекалии. Некоторым «навозным грибам» действительно требуется прохождение через пищеварительную систему травоядных, чтобы завершить свой жизненный цикл. Черный трюфель — ценный деликатес для гурманов — является плодовым телом подземного аскомицета. Почти все трюфели являются эктомикоризными и обычно встречаются в тесной связи с деревьями. Животные едят трюфели и рассеивают споры. В Италии и Франции охотники за трюфелями используют самок свиней, чтобы вынюхивать трюфели (самок свиней привлекают трюфели, потому что грибок выделяет летучее соединение, тесно связанное с феромоном, вырабатываемым самцами свиней).0005
Споры грибов редко полностью разлагаются в желудочно-кишечном тракте животного, и многие из них способны прорастать при попадании в фекалии. Некоторым «навозным грибам» действительно требуется прохождение через пищеварительную систему травоядных, чтобы завершить свой жизненный цикл. Черный трюфель — ценный деликатес для гурманов — является плодовым телом подземного аскомицета. Почти все трюфели являются эктомикоризными и обычно встречаются в тесной связи с деревьями. Животные едят трюфели и рассеивают споры. В Италии и Франции охотники за трюфелями используют самок свиней, чтобы вынюхивать трюфели (самок свиней привлекают трюфели, потому что грибок выделяет летучее соединение, тесно связанное с феромоном, вырабатываемым самцами свиней).0005
Резюме раздела
Грибы колонизировали почти все среды на Земле, но часто встречаются в прохладных, темных, влажных местах с запасом разлагающегося материала. Грибы – это сапробы, разлагающие органические вещества. Во многих успешных мутуалистических отношениях участвуют грибок и другой организм. Многие грибы образуют сложные микоризные ассоциации с корнями растений. Некоторые муравьи выращивают грибы в качестве источника пищи. Лишайники представляют собой симбиотические отношения между грибком и фотосинтезирующим организмом, обычно водорослью или цианобактерией. Фотосинтезирующий организм обеспечивает энергию из накопленных углеводов, в то время как грибы поставляют минералы и защиту. Некоторые животные, потребляющие грибы, помогают распространять споры на большие расстояния.
Многие грибы образуют сложные микоризные ассоциации с корнями растений. Некоторые муравьи выращивают грибы в качестве источника пищи. Лишайники представляют собой симбиотические отношения между грибком и фотосинтезирующим организмом, обычно водорослью или цианобактерией. Фотосинтезирующий организм обеспечивает энергию из накопленных углеводов, в то время как грибы поставляют минералы и защиту. Некоторые животные, потребляющие грибы, помогают распространять споры на большие расстояния.
Art Connections
(Рисунок) Если в почве отсутствуют симбиотические грибы, как вы думаете, какое влияние это окажет на рост растений?
Показать раствор
Вопросы на повторение
Какой термин описывает тесную связь грибка с корнем дерева?
- ризоид
- лишайник
- а микориза
- эндофит
Показать раствор
Почему грибы являются важными редуцентами?
- Они производят много спор.
- Они могут расти в самых разных средах.
- Они производят мицелий.
- Они перерабатывают углерод и неорганические минералы в процессе разложения.

Показать раствор
Рассмотрим экосистему, в которой уничтожены все грибы, не связанные с микоризой. Как это повлияет на потребление азота растениями?
- Потребление азота увеличится.
- Потребление азота не изменится.
- Потребление азота уменьшится.
- Поступление азота прекратится.
Показать раствор
Free Response
Почему защита от света на самом деле приносит пользу фотосинтезирующему партнеру лишайников?
Показать раствор
Короеды-амброзии переносят споры гриба Ambrosiella на деревья, затем просверливают отверстия и откладывают вместе с грибом яйца. Когда новые личинки вылупляются, они поедают гриб, проросший в лунках.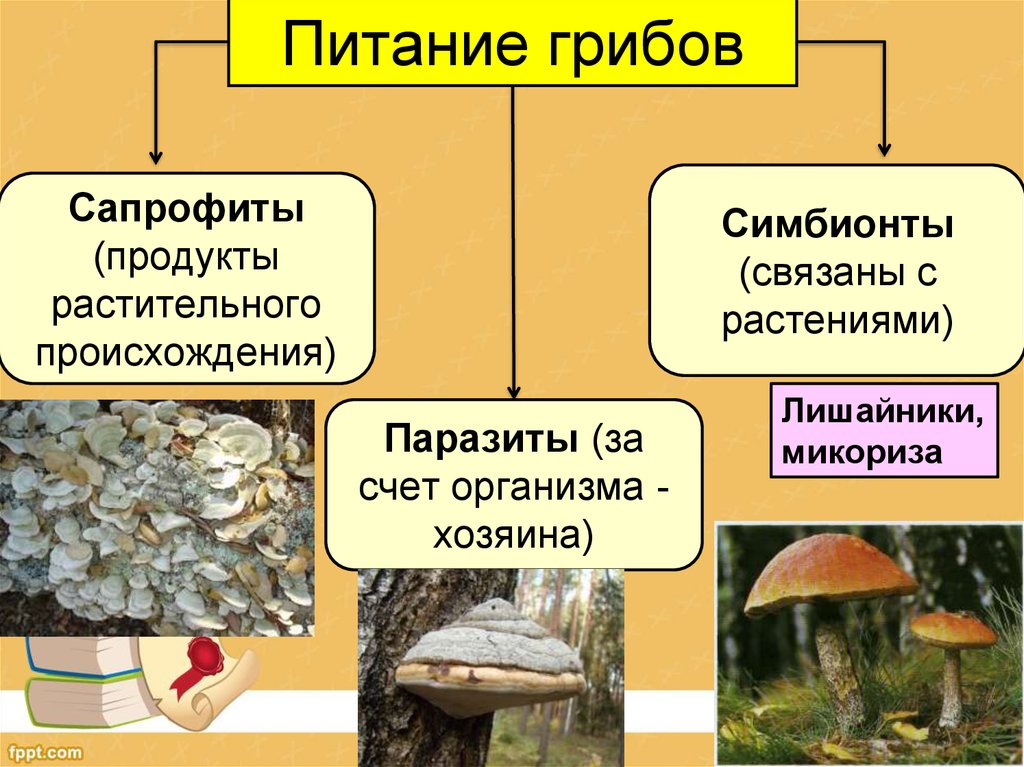 Опишите, как эти отношения можно классифицировать как мутуалистические.
Опишите, как эти отношения можно классифицировать как мутуалистические.
Показать решение
Экологи часто пытаются внедрить новые растения для восстановления деградировавших земель. В условиях засушливого климата ученые рекомендуют вводить растения с арбускулярной микоризой. Как микориза увеличит выживаемость растений по сравнению с растениями без микоризы?
Показать раствор
Глоссарий
- арбускулярная микориза
- микоризная ассоциация, при которой гифы грибов проникают в клетки корней и образуют обширные сети
- эктомикориза
- микоризные грибы, окружающие корни мантией и имеющие сеть Хартига, проникающую в корни между клетками
- лишайник
- тесная связь грибка с фотосинтезирующей водорослью или бактерией, приносящая пользу обоим партнерам
- микориза
- мутуалистическая ассоциация между грибами и корнями сосудистых растений
- соредия
- скопления клеток водорослей и мицелия, которые позволяют лишайникам размножаться
Грибы и лишайники | SpringerLink
Ахмаджян В.
, 1993. Симбиоз лишайников . Нью-Йорк: Уайли.Google ученый
Ариньо, X., и Саиз-Хименес, К., 1996. Лишайниковое повреждение консолидаторов, используемых при консервации каменных памятников. Лихенолог , 28 (4), 391–394.
Google ученый
Барсук М.Р., Пфанц Х., Бюдель Б., Хебер У. и Ланге О.Л., 1993. Доказательства функционирования фотосинтетических механизмов концентрации CO 2 в лишайниках, содержащих зеленые водоросли и цианобактериальные фотобионты. Планта , 191 , 57–70.
Перекрёстная ссылка
Google ученый
Белнап Дж., Бюдель Б. и Ланге О.Л., 2003. Биологические почвенные корки: характеристики и распространение. В Белнап, Дж., и Ланге, О.Л. (ред.), Экологические исследования 150: Биологические почвенные корки: структура, функции и управление .
Берлин: Springer, стр. 3–30.Перекрёстная ссылка
Google ученый
Benthelin, J., 1988. Микробные процессы выветривания в естественных условиях. У Лермана А. и Мейбека М. (ред.), Физическое и химическое выветривание в геохимических циклах . Берлин: Springer, стр. 33–60.
Перекрёстная ссылка
Google ученый
Bold, H.C., and Wynne, M.J., 1985. Введение в водоросли. Структура и воспроизведение , 2-е изд. Энглвудские скалы: Прентис-Холл.
Google ученый
Браун, Р. М., младший, Ларсон, Д. А., и Болд, Х. С., 1964. Водоросли в воздухе: их изобилие и неоднородность. Наука , 143 , 583–585.
Перекрёстная ссылка
Google ученый
Бюдель, Б., 2003. Биологические почвенные корки в умеренных и средиземноморских регионах Европы.
В Белнап, Дж., и Ланге, О.Л. (ред.), Экологические исследования 150: Биологические почвенные корки: структура, функции и управление . Берлин: Springer, стр. 75–86.Google ученый
Бюдель Б. и Ланге О.Л., 1991. Водный статус зеленых и сине-зеленых фикобионтов в слоевищах лишайников после гидратации за счет поглощения водяного пара: становятся ли они набухшими? Botanica Acta , 104 , 361–366.
Google ученый
Бюдель, Б., и Шайдеггер, К., 2008. Морфология и анатомия слоевища. В Нэше III Т.Х. (ред.), Биология лишайников . Кембридж, Издательство Кембриджского университета, стр. 40–68.
Перекрёстная ссылка
Google ученый
Бюдель Б., Беккер У., Фоллманн Г. и Стерфлингер К., 2000. Водоросли, грибы и лишайники на инзельбергах. В Porembski, S., и Barthlott, W. (ред.), Инзельбергс .
Берлин: Springer, стр. 69–90.Перекрёстная ссылка
Google ученый
Бюдель, Б., Вебер, Б., Кюль, М., Пфанц, Х., Зультемейер, Д. и Весселс, Д., 2004. Изменение формы поверхности песчаника криптоэндолитными цианоабктериями: биоалкализация вызывает химическое выветривание в засушливые пейзажи. Геобиология , 2 , 261–268.
Перекрёстная ссылка
Google ученый
Берфорд Э.П., Фомина М. и Гадд Г.М., 2003. Участие грибов в биовыветривании и биотрансформации горных пород и минералов. Минералогический журнал , 67 , 1127–1155.
Перекрёстная ссылка
Google ученый
Чен Дж., Блюм Х. П. и Бейер Л., 2000. Выветривание горных пород, вызванное заселением лишайниками – обзор. Катена , 39 , 121–146.
Перекрёстная ссылка
Google ученый
Деммиг-Адамс, Б.
, Магуас, К., Адамс, III, В.В., Мейер, А., Килиан, Э., и Ланге, О.Л., 1990. Влияние яркого света на эффективность фотохимического преобразования энергии у различных видов лишайников с зелеными и сине-зелеными фикобионтами. Планта , 180 , 400–409.Перекрёстная ссылка
Google ученый
Дьякумаку Э., Горбушина А. А., Крумбейн В. Э., Панина Л., Сухаревский С., 1995. Черные грибы в мраморе и известняках — эстетическая, химическая и физическая проблема консервации памятников. Наука об окружающей среде в целом , 167 , 295–304.
Перекрёстная ссылка
Google ученый
Дитц С., Бюдель Б., Ланге О.Л. и Билгер В., 2000. Пропускание света через кору лишайников из контрастных местообитаний. Bibliotheca Lichenologica , 75 , 171–182.
Google ученый
Дорниден Т.
, Горбушина А. А. и Крумбейн В. Е., 1997. Änderungen von physikalischen Eigenschaften von Marmor durch Pilzbewuchs. Международный журнал реставрации зданий и памятников , 3 , 441–456.Google ученый
Ertl, L., 1951. Über die Lichtverhältnisse in Laubflechten. Планта , 39 , 245–270.
Перекрёстная ссылка
Google ученый
Фридл Т. и Бюдель Б., 2008 г. Фотобионты. В Нэше III Т.Х. (ред.), Биология лишайников . Кембридж, Издательство Кембриджского университета, стр. 9–26.
Google ученый
Friedmann, E.I., 1982. Эндолитные микроорганизмы в холодной антарктической пустыне. Наука , 193 , 1247–1249.
Перекрёстная ссылка
Google ученый
Friedmann, E.I., and Weed, R., 1987. Микробное образование следов ископаемых, биогенное и абиотическое выветривание в холодной антарктической пустыне.
Наука , 236 , 703–705.Перекрёстная ссылка
Google ученый
Gadd, GM, 1999. Производство лимонной и щавелевой кислот грибами: важность в формировании металлов, физиологии и биогеохимических процессах. Достижения микробной физиологии , 41 , 47–92.
Перекрёстная ссылка
Google ученый
Галлоуэй, Д. Дж., 1996. Биогеография лишайников. В Нэше III Т.Х. (ред.), Биология лишайников . Кембридж, издательство Кембриджского университета, стр. 199–216.
Google ученый
Гейзер Д. М., Гейдан К., Миадликовска Дж., Лутцони Ф., Кауфф Ф., Хофштеттер В., Фракер Э., Шох С. Л., Тибелл Л., Унтеринер В. А. и Aptroot, A., 2006. Евротиомицеты: Eurotiomycetidae и Chaetothyriomycetidae. Микология , 98 , 1053–1064.
Перекрёстная ссылка
Google ученый
Голубич С.
, Фридман И. и Шнайдер Дж., 1981. Литобионтическая экологическая ниша с особым упором на микроорганизмы. Журнал осадочной петрологии , 51 (2), 475–478.Google ученый
Горбушина А.А., 2003. Микроколониальные грибы: потенциал выживания наземных растительных структур. Астробиология , 3 , 543–554.
Перекрёстная ссылка
Google ученый
Горбушина А.А., 2007. Жизнь на скалах. Экологическая микробиология , 9 , 1613–1631.
Перекрёстная ссылка
Google ученый
Грин, Т.Г.А., и Снелгар, В.П., 1982. Обмен углекислым газом в лишайниках: взаимосвязь между диффузионным сопротивлением углекислого газа и водяного пара. Лихенолог , 14 , 255–260.
Перекрёстная ссылка
Google ученый
Грин Т.
Г.А., Килиан Э. и Ланге О.Л., 1991. Pseudocyphellaria dissimilis: чувствительный к высыханию, хорошо адаптирующийся к тени лишайник из Новой Зеландии. Экология , 85 , 498–503.Перекрёстная ссылка
Google ученый
Гротцингер Дж., Джордан Т.Х., Пресс Ф. и Сивер Р., 2006. Понимание Земли. 5-е изд. Нью-Йорк: WH Фриман.
Google ученый
Хан, С., Шпеер, М., Мейер, А., и Ланге, О.Л., 1989. Фотосинтетическая первичная продукция эпигейских флехтенов в «Mainfränkischen Trockenrasen». I. Tagesläufe von Mikroklima, Wassergehalt und CO 2 -Gaswechsel zu den verschiedenen Jahreszeiten. Флора , 182 , 313–339.
Google ученый
Хенсен А., 1987. Lichenothelia, род микрогрибов на скалах. Bibliotheca Lichenologica , 25 , 257–293.
Google ученый
Онеггер Р.
, 1996. Микобионты. В Нэше III Т.Х. (ред.), Биология лишайников . Кембридж, Издательство Кембриджского университета, стр. 24–36.Google ученый
Каппен, Л., 1973. Реакция на экстремальные условия. В Ахмаджане, В., и Хейле, М. (ред.), Лишайники . Лондон: Академический, стр. 311–380.
Перекрёстная ссылка
Google ученый
Каппен Л., 1988. Экофизиологические взаимосвязи в различных климатических регионах. В Галун, М. (ред.), Справочник по лихенологии . Бока-Ратон: CRC, Vol. 2, стр. 39–99.
Google ученый
Каппен Л. и Фридман Э. И., 1983. Экофизиология лишайников в сухих долинах южной части Земли Виктории, Антарктида. II. СО 2 газообмен у криптоэндолитных лишайников. Полярная биология , 1 , 227–232.
Перекрёстная ссылка
Google ученый
Кершоу, К.
А., 1985. Физиологическая экология лишайников . Кембридж: Издательство Кембриджского университета.Google ученый
Крумбейн, У. Э., и Йенс, Дж., 1981. Биогенные скальные лаки пустыни Негев (Израиль): экологическое исследование трансформации железа и марганца цианобактериями и грибами. Экология , 50 , 25–38.
Перекрёстная ссылка
Google ученый
Lange, OL, 1953. Hitze- und Trockenresistenz der Flechten in Beziehung zu ihrer Verbreitung. Флора , 140 , 39–97.
Google ученый
Lange, OL, 1969. Experimentell-ökologische Untersuchungen an Flechten der Negev-Wüste. I. CO 2 -Gaswechsel von Ramalina maciformis (Del.) Bory unter kontrollierten Bedingungen im Laboratorium. Флора , 158 , 324–359.
Google ученый
Lange, O.
L., 2002. Фотосинтетическая продуктивность эпилитного лишайника Lecanora muralis : долгосрочный полевой мониторинг обмена CO 2 и его физиологическая интерпретация. Флора , 197 , 233–249.Перекрёстная ссылка
Google ученый
Lange, O.L., and Green, T.G.A., 1997. Высокое содержание воды в талломе может ограничивать продуктивность фотосинтеза накипных лишайников в полевых условиях. Bibliotheca Lichenologica , 68 , 81–99.
Google ученый
Lange, O.L., and Green, T.G.A., 2005. Лишайники показывают, что грибы могут адаптировать свое дыхание к сезонным изменениям температуры. Экология , 142 , 11–19.
Перекрёстная ссылка
Google ученый
Lange, O.L., and Wagenitz, G., 2003. Что такое «фиколишай»? различия и изменения в значении старого лихенологического термина.
Лихенолог , 35 (4), 341–345.Перекрёстная ссылка
Google ученый
Lange, O.L., Büdel, B., Meyer, A., and Kilian, E., 1993. Еще одно доказательство того, что активация чистого фотосинтеза сухими цианобактериальными лишайниками требует жидкой воды. Лихенолог , 25 (2), 175–189.
Google ученый
Lange, O.L., Green, T.G.A., Reichenberger, H., and Meyer, A., 1996. Фотосинтетическая депрессия при высоком содержании воды в лишайниках: одновременное использование методов газообмена и флуоресценции с цианобактериями и зелеными водорослями Пельтигера вида. Botanica Acta , 109 , 43–50.
Google ученый
Ланге О.Л., Белнап Дж., Рейхенбергер Х. и Мейер А., 1997. Фотосинтез зеленых водорослей лишайников почвенной корки засушливых земель на юге Юты, США: роль содержания воды в освещении и температуре.
ответы CO 2 обмен. Флора , 192 , 1–15.Google ученый
Lange, O.L., Green, T.G.A., and Reichenberger, H., 1999. Реакция фотосинтеза лишайников на внешний CO 2 концентрация и ее взаимодействие с водным статусом слоевища. Журнал физиологии растений , 154 , 157–166.
Перекрёстная ссылка
Google ученый
Lange, O.L., Büdel, B., Meyer, A., Zellner, H., and Zotz, G., 2000. Получение углерода лишайниками в тропических условиях: водные отношения и CO 2 обмен трех Leptogium вида нижних горных тропических лесов в Панаме. Флора , 195 , 172–190.
Google ученый
Ланге, О. Л., Грин, Т. Г. А., и Хебер, У., 2001. Зависимое от гидратации фотосинтетическое производство лишайников: что лабораторные исследования говорят нам о полевых условиях? Журнал экспериментальной ботаники , 52 (363), 2033–2042.
Перекрёстная ссылка
Google ученый
Lange, O.L., Büdel, B., Meyer, A., Zellner, H., and Zotz, G., 2004. Получение углерода лишайниками в тропических условиях: водные отношения и CO 2 обмен видами Lobariaceae из нижнего горного тропического леса в Панаме. Лихенолог , 36 (5), 329–342.
Перекрёстная ссылка
Google ученый
Мядликовска Дж., Кауфф Ф., Хофстеттер В., Фракер Э., Грубе М., Хафельнер Дж., Риб В., Ходкинсон Б. П., Куква М., Люкинг, Р., Хестмарк Г., Гарсия-Оталора М., Раухут А., Бюдель Б., Шайдеггер К., Тимдал Э., Стенроос С., Бродо И., Перлмуттер Г. Б., Эрц , Д., Дидерих П., Лендемер, Дж. К., Трипп, Э., Яр, Р., Мэй, П., Гейдан, К., Спатафора, Дж. В., Шох, К., Арнольд, А. Э., Робертсон, К. , and Lutzoni, F., 2006. Новый взгляд на классификацию и эволюцию Lecanoromycetes (Pezizomycotina, Ascomycota) на основе филогенетического анализа трех генов, кодирующих рибосомную РНК, и двух генов, кодирующих белок.
Микология , 98 , 1088–1103.Перекрёстная ссылка
Google ученый
Нэш, Т. Х., 1996а. Введение. В Nash III, TH (ed.), Lichen Biology . Кембридж, Издательство Кембриджского университета, стр. 1–7.
Google ученый
Нэш, Т. Х., 1996b. Фотосинтез, дыхание, продуктивность и рост. В Nash III, TH (ed.), Lichen Biology . Кембридж, Издательство Кембриджского университета, стр. 88–120.
Google ученый
Onofri, S., Barreca, D., Rabbow, E., de Vera, J.P., Selbmann, L., and Zucconi, L., 2007. Антарктические каменные грибы в условиях, смоделированных в космосе и на Марсе. Тезисы геофизических исследований , 9 , 9782–9783.
Google ученый
Палмер, Ф. Э., Эмери, Д. Р., Стеммлер, Дж., и Стейли, Дж. Т.
, 19 лет87. Выживание и рост микроколониальных каменных грибов при воздействии температуры и влажности. Новый фитолог , 107 , 155–162.Перекрёстная ссылка
Google ученый
Рундель, П. В., 1988. Водные отношения. In Galun, M. (ред.), Справочник по лихенологии , Бока-Ратон: CRC, Vol. 2, 17–36.
Google ученый
Schoch, C.L., Shoemaker, R.A., Seifert, K.A., Hambleton, S., Spatafora, J.W., and Crous, P.W., 2006. Мультигенная филогения Dothideomycetes с использованием четырех ядерных локусов. Микология , 98 , 1041–1052.
Перекрёстная ссылка
Google ученый
Schöller, H., 1997. Ökologie und Verbreitung. In Schöller, H. (ed.), Flechten: Geschichte, Biologie, Systematik, Ökologie, Naturschutz und kulturelle Bedeutung . Франкфурт-на-Майне: Senckenbergische Naturforschende Gesellschaft, стр.
83–109.Google ученый
Schöller, H., и Mollenhauer, D., 1997. Flechtensymbiose und Flechtenthallus. In Schöller, H. (ed.), Flechten: Geschichte, Biologie, Systematik, Ökologie, Naturschutz und kulturelle Bedeutung . Франкфурт-на-Майне: Senckenbergische Naturforschende Gesellschaft, стр. 15–47.
Google ученый
Зельбманн, Л., де Хоог, Г.С., Маццаглия, А., Фридманн, Э.И., и Онофри, С., 2005. Грибы на краю жизни: криптоэндолитические черные грибы из антарктической пустыни. Исследования по микологии , 51 , 1–32.
Google ученый
Sigler, L., Tsuneda, A., and Carmichael, J.W., 1981. Phaeotheca и Phaeosclera , два новых рода дематиевых гифомицетов и переописание Sarcinomyces 9444. Микотаксон , 12 , 449–467.
Google ученый
Sterflinger, K.
, De Baere, R., de Hoog, G.S., De Wachter, R., Krumbein, W.E., and Haase, G., 1997. Coniosporium Perforans и C. apollinis , два новых населяющие грибы, выделенные из мрамора в святилище Делос (Киклады, Греция). Антони ван Левенгук , 72 , 349–363.Перекрёстная ссылка
Google ученый
Тейлор-Джордж С., Палмер Ф., Стейли Дж. Т., Борнс Д. Дж., Кертисс Б. и Адамс Дж. Б., 19 лет83. Грибы и бактерии, участвующие в образовании пустынного нагара. Микробная экология , 9 , 227–245.
Перекрёстная ссылка
Google ученый
Tretiach, M., и Geletti, A., 1997. CO 2 обмен эндолитного лишайника Verrucaria baldensis из карстовых мест обитания в Северной Италии. Экология , 111 , 515–522.
Перекрёстная ссылка
Google ученый
Tretiach, M.
, and Pecchiari, M., 1995. Скорости газообмена и содержание хлорофилла в эпи- и эндолитных лишайниках из Триестского карста (СВ Италия). Новый Фотолог , 130 , 585–592.Перекрёстная ссылка
Google ученый
van den Hoek, C., Jahns, H.M., and Mann, D.G., 1993. Algen. 3. Ауфлаж . Штутгарт: Тиме.
Google ученый
Warscheid, T., Oelting, M., and Krumbein, W.E., 1991. Физико-химические аспекты процессов биодеградации горных пород с особым учетом органических загрязнителей. International Biodeterioration , 28 , 37–48.
Перекрёстная ссылка
Google ученый
Вебер, Б., и Бюдель, Б., 2001. Картирование и анализ моделей распространения лишайников в сельских средневековых церквях в Северо-Восточной Германии. Лихенолог , 33 (3), 231–248.
Перекрёстная ссылка
Google ученый
Вебер, Б.
, Шерр, К., Райхенбергер, Х., и Бюдель, Б., 2007. Быстрая реактивация при высокой влажности воздуха и фотосинтетических характеристиках альпийских лишайников, эндолитически растущих в известняке. Арктические, антарктические и альпийские исследования , 39 (2), 309–317.Перекрёстная ссылка
Google ученый
Вебер, Б., Олеховски, К., Кнерр, Т., Хилл, Дж., Дойчевич, К., Весселс, Д. К. Дж., Эйтель, Б. и Бюдель, Б., 2008. Новый подход к картографирование биологических почвенных корок в полупустынных районах гиперспектральной съемкой. Дистанционное зондирование окружающей среды , 112 , 2187–2201.
Перекрёстная ссылка
Google ученый
Wessels, D.C.J., and Schoeman, P., 1988. Механизмы и скорость выветривания песчаника Clarens эндолитным лишайником. Южноафриканский научный журнал , 84 , 274–277.
Google ученый
Винклер, Дж.
 , 1993. Симбиоз лишайников . Нью-Йорк: Уайли.
, 1993. Симбиоз лишайников . Нью-Йорк: Уайли. Берлин: Springer, стр. 3–30.
Берлин: Springer, стр. 3–30. В Белнап, Дж., и Ланге, О.Л. (ред.), Экологические исследования 150: Биологические почвенные корки: структура, функции и управление . Берлин: Springer, стр. 75–86.
В Белнап, Дж., и Ланге, О.Л. (ред.), Экологические исследования 150: Биологические почвенные корки: структура, функции и управление . Берлин: Springer, стр. 75–86. Берлин: Springer, стр. 69–90.
Берлин: Springer, стр. 69–90. , Магуас, К., Адамс, III, В.В., Мейер, А., Килиан, Э., и Ланге, О.Л., 1990. Влияние яркого света на эффективность фотохимического преобразования энергии у различных видов лишайников с зелеными и сине-зелеными фикобионтами. Планта , 180 , 400–409.
, Магуас, К., Адамс, III, В.В., Мейер, А., Килиан, Э., и Ланге, О.Л., 1990. Влияние яркого света на эффективность фотохимического преобразования энергии у различных видов лишайников с зелеными и сине-зелеными фикобионтами. Планта , 180 , 400–409. , Горбушина А. А. и Крумбейн В. Е., 1997. Änderungen von physikalischen Eigenschaften von Marmor durch Pilzbewuchs. Международный журнал реставрации зданий и памятников , 3 , 441–456.
, Горбушина А. А. и Крумбейн В. Е., 1997. Änderungen von physikalischen Eigenschaften von Marmor durch Pilzbewuchs. Международный журнал реставрации зданий и памятников , 3 , 441–456. Наука , 236 , 703–705.
Наука , 236 , 703–705. , Фридман И. и Шнайдер Дж., 1981. Литобионтическая экологическая ниша с особым упором на микроорганизмы. Журнал осадочной петрологии , 51 (2), 475–478.
, Фридман И. и Шнайдер Дж., 1981. Литобионтическая экологическая ниша с особым упором на микроорганизмы. Журнал осадочной петрологии , 51 (2), 475–478. Г.А., Килиан Э. и Ланге О.Л., 1991. Pseudocyphellaria dissimilis: чувствительный к высыханию, хорошо адаптирующийся к тени лишайник из Новой Зеландии. Экология , 85 , 498–503.
Г.А., Килиан Э. и Ланге О.Л., 1991. Pseudocyphellaria dissimilis: чувствительный к высыханию, хорошо адаптирующийся к тени лишайник из Новой Зеландии. Экология , 85 , 498–503. , 1996. Микобионты. В Нэше III Т.Х. (ред.), Биология лишайников . Кембридж, Издательство Кембриджского университета, стр. 24–36.
, 1996. Микобионты. В Нэше III Т.Х. (ред.), Биология лишайников . Кембридж, Издательство Кембриджского университета, стр. 24–36..jpg) А., 1985. Физиологическая экология лишайников . Кембридж: Издательство Кембриджского университета.
А., 1985. Физиологическая экология лишайников . Кембридж: Издательство Кембриджского университета. L., 2002. Фотосинтетическая продуктивность эпилитного лишайника Lecanora muralis : долгосрочный полевой мониторинг обмена CO 2 и его физиологическая интерпретация. Флора , 197 , 233–249.
L., 2002. Фотосинтетическая продуктивность эпилитного лишайника Lecanora muralis : долгосрочный полевой мониторинг обмена CO 2 и его физиологическая интерпретация. Флора , 197 , 233–249. Лихенолог , 35 (4), 341–345.
Лихенолог , 35 (4), 341–345. ответы CO 2 обмен. Флора , 192 , 1–15.
ответы CO 2 обмен. Флора , 192 , 1–15.
 Микология , 98 , 1088–1103.
Микология , 98 , 1088–1103. 83–109.
83–109. , De Baere, R., de Hoog, G.S., De Wachter, R., Krumbein, W.E., and Haase, G., 1997. Coniosporium Perforans и C. apollinis , два новых населяющие грибы, выделенные из мрамора в святилище Делос (Киклады, Греция). Антони ван Левенгук , 72 , 349–363.
, De Baere, R., de Hoog, G.S., De Wachter, R., Krumbein, W.E., and Haase, G., 1997. Coniosporium Perforans и C. apollinis , два новых населяющие грибы, выделенные из мрамора в святилище Делос (Киклады, Греция). Антони ван Левенгук , 72 , 349–363. , and Pecchiari, M., 1995. Скорости газообмена и содержание хлорофилла в эпи- и эндолитных лишайниках из Триестского карста (СВ Италия). Новый Фотолог , 130 , 585–592.
, and Pecchiari, M., 1995. Скорости газообмена и содержание хлорофилла в эпи- и эндолитных лишайниках из Триестского карста (СВ Италия). Новый Фотолог , 130 , 585–592. , Шерр, К., Райхенбергер, Х., и Бюдель, Б., 2007. Быстрая реактивация при высокой влажности воздуха и фотосинтетических характеристиках альпийских лишайников, эндолитически растущих в известняке. Арктические, антарктические и альпийские исследования , 39 (2), 309–317.
, Шерр, К., Райхенбергер, Х., и Бюдель, Б., 2007. Быстрая реактивация при высокой влажности воздуха и фотосинтетических характеристиках альпийских лишайников, эндолитически растущих в известняке. Арктические, антарктические и альпийские исследования , 39 (2), 309–317.