
Физиологическое действие ауксинов и распределение в растениях. Ауксин влияние на растения
Растительные гормоны. Ауксины. - Дачное Дело
Что такое ауксины?
Ауксины – это семейство растительных гормонов с различными ролями в морфогенезе растений, включая расширение клеток, образование корней и развитие почек, а также тропизм (реакция ориентирования клетки, то есть направление роста или движения клеток относительно раздражителя, химического, светового и др.).
- Пять природных (эндогенных) ауксинов в растениях включают индол-3-уксусную кислоту, 4-хлориндол-3-уксусную кислоту , фенилуксусную кислоту , индол-3-масляную кислоту и индол-3-пропионовую кислоту .
- Синтетические аналоги ауксина включают 1-нафталинуксусную кислоту , 2,4-дихлорфеноксиуксусную кислоту (2,4-D) и многие другие.
Ауксины, как правило, распределены в растении неравномерно. Особенно высоко их содержание в развивающихся тканях: в почках, молодых листьях и их зачатках, в проводящих пучках, в пыльце, в формирующихся семенах. В семенах некоторых растений с глубоким покоем концентрация ауксинов достигает ингибирующих (блокирующих рост) количеств.
Ауксины, эффекты воздействия

- Ауксины участвют в фототропизме, геотропизме, гидротропизме и других реакциях растения на влияние внешней среды.

| Фототропизм – реакция растения на источник света. Геотропизм – реакция растения на действие гравитации. Гидротропизм – реакция изгибания некоторых органов растений под влиянием неравномерного распределения влажности в окружающей их среде. |
Внешние причины, такие как боковая освещенность, сила тяжести, приводят к неравномерному росту ткани растения, и, как правило, именно ауксин определяет форму, направление и силу роста всех органов и их взаимодействие.
-
Ауксин стимулирует удлинение клеток. Присутствие гиббереллинов усиливает этот эффект .
-
Ауксин будет стимулировать деление клеток, если присутствуют цитокинины. Для увеличения скорости нарастания каллюса. Когда ауксин и цитокинин наносятся на каллюс , укоренение может быть получено, если концентрация ауксина выше, чем концентрация цитокинина. Ткани ксилемы могут быть получены, если концентрация ауксина равна концентрации цитокининов.
| Каллюс – ткань, образующаяся у растений на месте поранений и способствующая их заживлению или образованию корней на черенках Ксилема, или древесина — основная водопроводящая ткань наземных сосудистых растений; один из двух подтипов проводящей ткани растений, наряду с флоэмой — лубом. Флоэма, луб (φλοῦς — кора) — проводящая ткань сосудистых растений, по которой происходит транспорт продуктов фотосинтеза к частям растения, где происходит их использование. |
- Реакция на поранение. Ауксин вызывает образование и организацию флоэмы и ксилемы . Когда растение ранено, ауксин может индуцировать клеточную дифференциацию и регенерацию сосудистых тканей.

- Корневой рост и развитие. Ауксины способствуют инициированию корня. Ауксин индуцирует как рост ранее существовавших корней, так и случайное образование корней, т. е. ветвление корней. В садоводстве ауксины, особенно 1-Нафталинуксусная кислота и индол-3-масляная кислота, обычно применяются для стимулирования корнеобразования при укоренении черенков растений. Однако высокие концентрации ауксина подавляют корнеобразование.
- Ауксин индуцирует апикальное доминирование ; рост пазушных почек ингибируется ауксином. Высокая концентрация ауксина стимулирует синтез этилена в пазушных почках, вызывая ингибирование их роста. Если вершина растения, в которой концентрация ауксина максимальна, удаляется, то тормозящий эффект ауксина снижается, что стимулирует рост боковых почек.
| Апикальное доминирование — преимущественное развитие верхушечной почки, которая растёт быстрее и замедляет рост боковых почек. |
- Рост и развитие плодов. Ауксин необходим для роста и развития плодов. Экзогенный ауксин приводит к партенокарпии.
| Партенокарпия ( parthenos — девственница и karpos — плод) — частный случай партеногенеза, девственное оплодотворение без опыления у растений, обычно с образованием плодов без семян. |
- Цветение. Ауксин играет второстепенную роль в инициировании цветения и развития репродуктивных органов. В низких концентрациях он может замедлять старение.
- Биосинтез этилена. В низких концентрациях ауксин может ингибировать образование этилена в растениях. А высокие концентрации могут вызывать синтез этилена. Поэтому высокая концентрация может вызвать женский тип цветения у некоторых видов.
- Ауксин также вызывает накопление сахара и минералов в месте нанесения.
- Ауксин ингибирует абсцинирование до образования слоя абсцесса и, таким образом, препятствует старению листьев.
- Может действовать, как гербицид. Одним из наиболее известных синтетических ауксинов является 2,4-дихлорфеноксиуксусная кислота, более широко известная как 2,4-D. 2,4-D было одним из соединений (в сочетании с 2,4,5-T), использовавшийся США как дефолиант во время войны во Вьетнаме.
ИУК.
ИУК – индолил-3-уксусная кислота, природный фитогормон, стимулятор роста растений. Химическое вещество с высокой физиологической активностью.
Свойства ИУК:
- Белое кристаллическое вещество, быстро темнеющее на свету
- Хорошо растворяется в спиртах, в серном эфире и этилацетате.
- Плохо растворяется в воде. Растворимость в горячей воде увеличивается.
- Калиевая соль ИУК хорошо растворяется в воде.
- ИУК быстро разлагается в кислой среде.
- ИУК в щелочной среде более стабильна.
Читать по теме: Фитогормоны и их влияние на растения.
Применение ауксинов в садоводстве.Вконтакте
Google+
Поделиться через соцсеть
dachnoedelo.ru
| Наиболее ярким проявлением физиологического действия ауксина является его влияние на рост клеток в фазе растяжения. ИУК стимулирует выход протонов в клеточную стенку и увеличивает ее растяжимость. Под влиянием оптимальной концентрации ИУК рост в длину декапитированных отрезков стеблей гороха увеличивается более чем в два раза. Ауксины в некоторых случаях стимулируют деление клеток, например камбия. Под влиянием ауксинов может измениться направление дифференциации клеток. Поданным Торрея, ауксин вызывает дифференциацию ксилемы, индуцирует корнеобразование. В последнее время эти данные получили подтверждение. Так, введение в растения петунии гена синтеза ауксинов из агробактерии индуцировало дифференциацию проводящих пучков. В тоже время после внедрения в растения табака гена, снижающего содержание ИУК, уменьшилось и число сосудов. Стимуляция ауксином роста боковых корней показана на мутантах арабидопсиса, у которых образование массы корней происходило на фоне в 17 раз большего накопления ауксина. Таким образом, все проявления роста клеток находятся в определенной зависимости от содержания природных ауксинов. Большую роль играют ауксины при разрастании завязи и плодообразовании. Показано, что ауксины могут синтезироваться в пыльце, зародыше, эндосперме. На первых стадиях роста плодов ауксин поступает из эндосперма, а позднее — из зародыша. У некоторых растений ауксин стимулирует образование бессемянных (партенокарпических) плодов. Ауксины, так же как и другие фитогормоны, обусловливают взаимодействие отдельных органов растения (коррелятивный рост). Ауксин обуславливает явление апикального доминирования, проявляющегося в подавляющем влиянии верхушечной почки на рост пазушных. Участие ауксина в этом процессе установлено в1935 г. Прямые определения показали, что через 4 часа после декапитации содержания ауксина в узлах увеличивалось в 5 раз. С участием ауксина связана также регуляция двигательной активности растений, в частности тропизмы и настии. Действие ауксина находится в зависимости от его концентрации. Повышение концентрации ауксина выше оптимальной вызывает торможение роста. При этом для разных растений и для неодинаковых органов оптимальная концентрация ауксина резко различна. Одна и та же концентрация его может усилить рост одних органов и затормозить другие. Так, оптимальная концентрация для роста стебля составляет около 10 мг, тогда как для корня всего 0,01 мг ИУК на 1 кг массы растения. Концентрация ауксинов, усиливающая рост злаков, резко тормозит рост многих двудольных растений. Это может быть связано с тем, что ауксин стимулирует синтез другого фитогормона, ингибирующего рост, а именно этилена. При всех физиологических проявлениях ауксины усиливают поступление воды и питательных веществ (аттрагирующее влияние). Имеются многочисленные данные, что ауксины являются регуляторами притока воды и питательных веществ. Ауксины влияют на распределение питательных веществ в растении (Н.А. Максимов, Н.И. Якушкина). При внесении извне ауксинов или их синтетических аналогов они концентрируются в отдельных органах и клетках. Это вызывает приток к этим органам воды и питательных веществ и, как следствие, их усиленный рост. Одновременно рост других органов, содержащих меньше ауксинов, ослабляется, поскольку питательные вещества к ним поступают в меньшем количестве. Так, при обработке фитогормонами типа ауксина цветков томата происходит усиленное разрастание завязей, приток к ним питательных веществ значительно повышается, а рост боковых побегов тормозится. Общий вынос питательных веществ при этом не изменяется. Из этих опытов следует, что гормоны типа ауксина вызывают перераспределение питательных веществ в растении. В некоторых случаях ауксин может вызвать усиление притока питательных веществ и из внешней среды. Существуют разные гипотезы, объясняющие действие ауксина на передвижение веществ. Не вызывает сомнения, что транспорт веществ по растительному организму определенным образом связан с напряженностью энергетического обмена. В этой связи важно отметить, что еще в 1933 г. появились исследования, показывающие, что под влиянием ауксина интенсивность дыхания растет (Д. Боннер). В отсутствие кислорода действие ауксина или не проявляется, или значительно ослабевает. На различных объектах установлено, что влияние ауксина на повышение интенсивности дыхания опережает во времени его действие на ростовые процессы. Под влиянием ИУК возрастает сопряженность окисления и фосфорилирования (коэффициент Р/О) и содержание в клетках АТФ. Это дает основание считать, что ИУК увеличивает энергетическую эффективность дыхания растений (Н.И. Якушкина, И.А. Кулакова). Под влиянием ИУК возрастает и энергетический заряд клетки (отношение АТФ + АДФ к АМФ). Известно, что даже небольшие сдвиги в энергетическом потенциале клетки приводят к заметным изменениям в скорости различных ферментативных реакций. Положительные сдвиги в энергетическом обмене вызывают усиление передвижения питательных веществ и воды, что является одной из причин усиления роста растений. Решение вопроса о причинах усиления образования АТФ под влиянием ИУК связано с изучением первичных механизмов регуляторного влияния этого фитогормона. |
fizrast.ru
Применение ауксинов в садоводстве. - Дачное Дело

Применение ауксинов. Когда, где и как.
Ауксины – наиболее часто используемые в сельском хозяйстве стимуляторы.
- Применение ауксинов для вегетативного размножения.
Вследствие нестабильности самой ИУК чаще используют ее синтетические заменители, такие как 1-НУК (альфа-нафтилуксусная кислота – C12h20O2 ) и ИМК (Бета-Индолилмасляная кислота -C12h23NO2 ). Эти соединения наиболее стабильны и нефитотоксичны. Их применяют для замачивания зеленых и одревесневших черенков. Черенки замачивают на 8 — 24 часа, погружая в раствор на 1/3 или на ½ их длины. Раствор готовят из расчета 25 — 70 мг препарата на 1 л воды. Можно использовать и кратковременную (5 сек.) обработку нижней части черенков в водно-спиртовом (1:1) растворе ИМК. Концентрация ИМК в этом растворе 2,5 — 5 г на 1 литр водно-спиртовой смеси. Для зеленых черенков, а также для черенков травянистых декоративных растений требуется меньшая концентрация ростовых веществ.
- Нивелирование влияния стрессовых ситуаций, например при пересадке.
Снятие стрессовой ситуации возможно на фоне применения регуляторов роста, в частности, ауксинов. Полив раствором ИМК или 1-НУК из расчета 5 — 10 мг препарата на 1 л воды снижает влияние негативного внешнего воздействия. При пересадке срезы корней смазывают пастой из глины и торфа, приготовленной с добавлением растворов ИМК или 1-НУК или помещают корни растений на сутки в раствор стимулятора.
- Применение ауксинов для стимуляции плодообразования.
Ауксины используют для стимуляции плодообразования и получения бессемянных плодов. Чаще всего с этой целью применяют регуляторы роста при выращивании томатов, огурцов, баклажанов, перцев и некоторых других культур в теплицах, но возможно использование этого метода и на плантациях, особенно при неблагоприятных погодных условиях. Для этого в начале цветения растения опрыскивают растворами 2,4-Д (2,4-Дихлорфеноксиуксусная кислота – C8H6Cl2O3 ) или 2,4,5-Т (2,4,5-Трихлорфеноксиуксусная кислота C8H5Cl3O3 ), 2-НОУК (2-Нафтоксиуксусная кислота C12h20O3). Для обработки используют рабочий раствор 40 — 50 мг препарата на 1 л воды. 2-НОУК эффективен и для опрыскивания грядок земляники.
- Для обработки семян овощных культур.
Для этой цели чаще используется гетероауксин. Концентрация гетероауксина для обработки семян моркови — 600 мг на 1 литр раствора, столовой свеклы — 800 мг, томатов, огурцов — 500 мг/л. Усиливает действие гетероауксина совместное применение с витаминами. Стимуляторы смешивают с витаминами в соотношении 600 мг/л гетероауксина+ 100 мг витамина В1 или такое же количество никотиновой кислоты, или же все три компонента вместе. Время обработки 10 — 12 часов.
- Стимуляция цветения и плодоношения.
Опрыскивание растений картофеля 0,01% раствором янтарной кислоты ускоряет цветение, повышается устойчивость к фитофторозу и, как следствие, увеличение урожайности. Обработка томатов янтарной кислотой проводится в период бутонизации (40 — 60 мг/л) для повышения продуктивности и повторяют трижды. Интервал между обработками — 7 дней. Расход раствора 2 литра на сотку.
- Прореживания цветков и завязей плодовых растений.
Прореживание цветов или завязей применяется для уменьшения нагрузки и повышения качества урожая. Кроме того, регулируя цветение можно нивелировать такое неприятное явление, как годовая периодичность урожая. Годовая периодичность плодоношения некоторых плодовых деревьев, например у яблонь, связана с их “усталостью” в предыдущий урожайный год. Уменьшение нагрузки позволяет растению заложить цветочные почки и обеспечить урожай на следующий год. Для прореживания цветков и завязей у груш, яблонь, абрикосов, персиков обычно используют раствор 1-НУК в концентрации 15 — 50 мг/л. Кроны деревьев обрабатывают во второй половине периода цветения.
- Сокращение опадания плодов.
С этой целью используют обычно растворы 1-НУК или 2,4-Д в концентрации 0,0001 — 0,001% (1 — 10 мг препарата на 1 л воды). Действие препарата сохраняется в течение 2-х недель со дня обработки. Предуборочное опрыскивание лимонов и апельсинов растворами 2,4-Д (8 мг/л) или 1-НУК (20 мг/л) не только уменьшает падалицу, но и замедляет созревание плодов. Такие плоды лучше хранятся, в меньшей степени подвергаются заболеваниям.
- Для задержки цветения плодовых деревьев.
Цветение плодовых во время весенних возвратных заморозков может полностью лишить урожая. Осеннее опрыскивание ауксинами в период окончания роста и начала закладки цветочных почек задержит цветение весной на 5-10 дней и спасет урожай будущего года. Опрыскивание 1-НУК (25 — 50 мг/л) задерживает наступление периода цветения весной следующего года у яблонь и груш на 5 — 7 дней, абрикосов и персиков — на 10 дней. К тому же этот прием в год обработки ускоряет на несколько дней созревание плодов.
- Уменьшение отрицательного влияния заморозков на созревающий урожай.
С этой целью используют ауксины 30 — 40 мг на 1 л раствора. Такое опрыскивание ускоряет созревание плодов, что уменьшает вероятность попадания урожая под заморозок.
- Применение ауксинов при хранении корнеплодов и луковиц.
Обработка ауксинами позволяет продлить период покоя у корнеплодов и луковиц.Метиловый эфир а-нафтилуксусной кислоты, известный как препарат М-1, широко употребляется в виде порошка ( 3 – 4 % препарата и 96 % глины), как средство, задерживающее прорастание клубней картофеля при хранении. Слои картофеля при переборке обрабатывают смесью глины и М-1. На 1 тонну картофеля требуется 50 — 100 г препарата. М-1 резко тормозит прорастание глазков и потерю веса клубнями.Другой эффективный способ задержки прорастания клубней картофеля — опрыскивание ботвы за 2−3 недели до уборки 0,2% раствором ГМК (гидразида малеиновой кислоты). ГМК проникает в клубни и задерживает прорастание глазков в течение 8 месяцев при температуре +10 гр С. ГМК также способствует сохранению сахарозы в корнеплодах свеклы, ингибирует прорастание моркови, лука и других овощей при длительном хранении.
- Применение ауксинов для уничтожения сорняков (гербицидные свойства).
Читать по теме: Фитогормоны и их влияние на растения.
Растительные гормоны. Ауксины.
Вконтакте
Google+
Поделиться через соцсеть
dachnoedelo.ru
Физиологическое действие ауксинов и распределение в растениях
Под ауксинами большинство исследователей подразумевает определенную группу химических соединении (кислоты с ненасыщенным ядром или их производные), естественно встречающихся или не обнаруженных в растениях и характеризующихся способностью индуцировать рост клеток стебля в длину при определенных концентрациях.
Сверхоптимальные концентрации ауксинов вызывают торможение роста. В зависимости от вида растительной ткани границы оптимальных (стимулирующих) и сверхоптимальных (тормозящих) концентраций колеблются в определенных пределах.
Приведена рост-концентрационная кривая для типичного ауксина β-икдолилуксусной кислоты (ИУК) в тесте коротких 5-миллиметровых отрезков овсяных колеоптплей. Эта кривая больше характеризует ростовую реакцию молодых стеблей. Почти такой же является реакция тканей листа, незрелых плодов и других надземных органов растений. При этом, конечно, имеются в виду молодые, потенциально способные к росту без предварительной структурной перестройки ткани. Оптимальными для роста стеблей, листьев и плодов являются концентрации порядка 10-4— 10-5 М в пересчете на индолилуксусную кислоту. Такие же концентрации уже резко тормозят рост корней. Стимулирующими или по крайней мере нейтральными для них обычно являются концентрации порядка 10-11—10-9 М ИУК. Из этого вовсе не следует, что содержание ауксинов в корне гораздо ниже содержания их в стебле. Видимо, оно такое же, а иногда и выше, чем в стебле.
Исследуя распределение свободных ауксинов в растениях томатов на разных этапах их индивидуального развития, мы во всех случаях обнаруживали, что концентрация ауксинов в корнях равнялась или превышала концентрацию ауксинов в отличающихся интенсивным ростом частях надземных органов: верхушках, молодых листьях, верхних частях стебля.
Однако в силу какой-то специфики, обусловленной, очевидно, особенностями метаболизма, корни чувствительней к ауксинам, чем надземные органы. Различная чувствительность корней и стеблей к одинаковым концентрациям нативных ауксинов особенно хорошо проявляется, когда растения испытывают азотное голодание.
Давно известно, что недостаток азота в почве по-разному влияет на рост надземной части и корней, в результате чего отношение массы корней к массе надземных органов увеличивается. Рассматривая этот неравномерный рост корней и стеблей растений с точки зрения рострегулиругощего действия ауксинов, Зединг писал: «Если и в отношении корней также действует правило, согласно которому недостаток азота снижает образование ростовых веществ верхушками побегов, а избыток азота — усиливает, то этим можно объяснить влияние азотных удобрений на соотношение побегов и корней у растений. Избыточно удобренные растения вследствие интенсивного образования плазмы и ростовых веществ развивают мощные надземные органы; в то же время корни растут слабо, вероятно, как раз из-за слишком высокого содержания в них ростовых веществ, так как уже нормальное количество последних превышает оптимум для роста корней. У голодающих растений, наоборот, вследствие недостатка ростовых веществ, рост побегов уменьшается; для корней же это сниженное содержание ростовых веществ оказывается, вероятно, ближе к оптимальному, что и приводит к значительному усилению их роста. Следовательно, изменение соотношения надземных частей и корней при недостатке или избытке азота основывается на различной чувствительности к ростовым веществам побегов и корней. Совершенно ясно, что это объяснение является только гипотезой, пока оно экспериментально не проверено».
Определенные экспериментальные подтверждения высказанного Зедингом предположения дают наши исследования. Наблюдая изменения сырого веса и концентрации свободных ауксинов в корнях и надземной части молодых растений томатов, в фазе 2—3 листков перенесенных на среду без азота, мы установили, что более усиленный, по сравнению со стеблями, рост корней на начальном этапе азотного голодания вызывает постепенное увеличение соотношения веса корни/стебли и сопровождается падением концентрации свободных ауксинов и в надземной части, и в корнях почти до одинакового уровня. И, наоборот, первой реакцией длительно голодавших по азоту растений в ответ на азотную подкормку является резкое (в 3,5 раза) возрастание концентрации свободных ауксинов в стеблях и листьях и значительно меньшее (только в 1,5 раза) увеличение концентрации тех же ауксинов в корнях. Таким образом, различная интенсивность роста корней и надземных органов при отсутствии или недостатке азота в субстрате может быть объяснена разной чувствительностью этих частей растения к снижению концентрации свободных ауксинов в испытывающих недостаток азота растениях. Когда же такие растения попадают в благоприятные в отношении азотного питания условия, концентрация ауксинов в них сильно увеличивается как раз там, где испытывался их недостаток, т. е. в надземных органах.
Влияние ауксинов не ограничивается регуляцией роста клеток в фазе растяжения. Как правильно замечают Кеффорд и Голдакр, сосредоточение внимания исследователей именно на этом процессе и в значительно меньшей степени — на других аспектах действия ауксинов вызвано, очевидно, техническими удобствами применяемых для выявления ауксинов тестов, в основе которых лежит растяжение клеток. Сейчас доказано обязательное участие ауксинов в индуцируемом кининами делении клеток и образовании каллюсов, в дальнейшей дифференциации клеток каллюсов, связанной с возникновением зачатков корней или стеблей, в угнетении верхушечной почкой развития боковых побегов из латеральных почек, т. е. в явлениях корреляции роста целого растения.
Если вы нашли ошибку, пожалуйста, выделите фрагмент текста и нажмите Ctrl+Enter.
www.activestudy.info
Ауксин (гормон растений)
В 30-х гг. ХХ столетия Ф. Венту удалось подтвердить мысль Ч. Дарвина, что в верхушках колеоптилей злаков действительно образуется значительное количество способного к диффузии вещества, которое контролирует рост нижележащих зон. Это вещество назвали ауксином (от греческого слова auxein – расти). Большой вклад в изучение механизма действия ауксина внес Н. Г. Холодный.
Ауксин синтезируется растущими апикальными зонами стеблей, в том числе молодыми листьями. От апекса ауксин мигрирует в зону растяжения, где он специфически влияет на рост растяжением.
Было установлено, что природный ауксин представляет собой простое соединение – индолил-3-уксусную кислоту (ИУК):

которая синтезируется в растениях путем ферментативного превращения аминокислоты – триптофана. Триптофан образуется из шикимовой кислоты через ряд реакций:
Шикимовая кислота → индол → серин → L-триптофан
Сам синтез ИУК из триптофана довольно сложный процесс и имеет свои особенности в отдельных растениях (рис. 6.2). Образованная из триптофана ИУК, проходит дальнейшее превращение в растениях: обратимое связывание ИУК в неактивные комплексы с белками, аминокислотами, углеводами; необратимое окисление молекулы ИУК под воздействием ферментов типа ауксиноксидазы.

Рис. 6.2. Биосинтез индолил-3-уксусной кислоты
Транспорт ИУК происходит полярно со скоростью 10–15 мм/ч от вершины побегов к корням. Механизм полярного транспорта следующий: в апикальный конец клетки ИУК проникает пассивно совместно с Н+, а в базальном конце активно секретируется через клеточную мембрану.
Физиологическое действие ауксина сложное. Разные ткани отвечают на действие ауксина увеличением роста, которое обусловлено стимуляцией растяжения клеток (рис. 6.3).

Рис. 6.3. Схема основных функций ауксина в растении
Роль ауксина в стимуляции опадания листьев и цветов связана с заметным понижением его содержания в листьях. Это приводит к старению листьев. Стареющие ткани продуцируют этилен, который действует на отделительную зону (зона опадания).
Сам же ауксин задерживает опадание листа и цветов. Имеются доказательства, что ауксин, кроме участия в растяжении и опадании листьев, принимает также участие в процессах клеточного деления. Вероятно, ауксины стимулируют камбиальную активность. Считают, что ИУК участвует в дифференциации сосудистой ткани в период начала ростового процесса. Ауксин, вероятно, принимает участие в образовании боковых корней. Превращение завязи в плод – это еще один контролируемый ауксином процесс.
Первостепенную роль играет ауксин в ростовых движениях – тропизмах и настиях (теория гормональной регуляции тропизмов Холодного – Вента).
В основе действия ауксинов, как считают, существуют два механизма:
– быстрое влияние ауксинов на мембранную систему, где за счет энергии АТФ увеличивается транспорт водородных ионов из цитоплазмы в клеточную оболочку и ускоряется размягчение клеточной оболочки;
– относительно медленное влияние ауксинов через геномную систему на синтез белков, определяемых рост клеток.
biofile.ru
Ауксины. Физиологическая активность и механизмы действия. Явление апикального доминирования. Практическое использование ауксинов в растениеводстве и биотехнологии.
Наиболее ярким проявлением физиологического действия ауксина является его влияние на рост клеток в фазе растяжения. ИУК стимулирует выход протонов в клеточную стенку и увеличивает ее растяжимость. Под влиянием ауксинов может измениться направление дифференциации клеток. Ауксины, так же как и другие фитогормоны, обусловливают взаимодействие отдельных органов растения (коррелятивный рост). Ауксин обуславливает явление апикального доминирования, проявляющегося в подавляющем влиянии верхушечной почки на рост пазушных. При всех физиологических проявлениях ауксины усиливают поступление воды и питательных веществ (аттрагирующее влияние). Ауксины влияют на распределение питательных веществ в растении. В отсутствие кислорода действие ауксина или не проявляется, или значительно ослабевает. ИУК увеличивает энергетическую эффективность дыхания растений. Под влиянием ИУК возрастает энергетический заряд клетки.ИУК активирует протонную помпу в плазмалемме, что приводит к закислению и разрыхлению клеточной стенки и тем самым способствует росту клеток растяжением. Комплекс ИУК с рецептором транспортируется в ядро и активирует синтез РНК, что в свою очередь приводит к усилению синтеза белков.
С участием ауксина связана регуляция двигательной активности растений, в частности тропизмы и настии. Повышение концентрации ауксина выше оптимальной вызывает торможение роста. Ауксин стимулирует синтез другого фитогормона, ингибирующего рост, этилена. Гормоны типа ауксина вызывают перераспределение питательных веществ в растении.
Ауксин стимулирует развитие бессемянных плодов. Использование, как гербицида. Если ауксинов слишком много, то начинается синтез гормона-антагониста этилена. Этилен угнетает рост побегов и корней в длину и вызывает листопад. При обработке естественными ауксинами (ИУК, ИПВК и др.) начинается их окисление и/или образование конъюгатов.
4. Гиббереллины. Открытие, строение, содержание, транспорт и распределение в различных частях и органах растений. Физиологическая активность и механизмы действия. Практическое применение.
Открытие гормонов растений гиббереллинов связано с изучением болезни риса. Японские ученые показали, что эта болезнь вызывается выделением гриба Gibberellafujikuroi. Из выделений этого гриба было получено кристаллическое вещество — гиббереллин. Гиббереллины обладают высокой физиологической активностью и являютсяестественными фитогормонами. По химическойструктуре это производные дитерпенов — дитерпеноиды, состоящие из четырех изопреновых остатков. Растения на разных этапах онтогенеза могут различаться по набору гиббереллинов, активность которых может быть различной. Гиббереллины могут образовываться в разных, по преимуществу растущих частях растительного организма. Все же основное место синтеза гиббереллинов — это листья. Имеются данные, что гиббереллины образуются в пластидах. По-видимому, гиббереллины существуют в двух формах — свободной и связанной. Нередко наблюдаемое повышение содержания гиббереллинов связано с переходом их из связанной в свободную форму. В отличие от ауксинов гиббереллины передвигаются из листьев как вверх, так и вниз, как по ксилеме, так и по флоэме. Это пассивный процесс, не связанный с метаболизмом. Образование гиббереллина в хлоропластах идет путем превращения мевалоновой кислоты в геранилгераниол и далее через каурен в гибберелловую кислоту.Влияние факторов. Освещение увеличивает содержание гиббереллинов. При выращивании растений на красном свете в них содержится больше гиббереллинов по сравнению с выращиванием на синем свете. Улучшение питания растений азотом содержание гиббереллинов снижает.
Проявлением физиологического действия гиббереллина является способность резко усиливать рост стебля у карликовых форм растений. Генетическая карликовость вызвана изменениями на генном уровне (отсутствие гена который кодирует синтез гиббереллинов) и может быть связана с нарушениями в синтезе гиббереллинов. 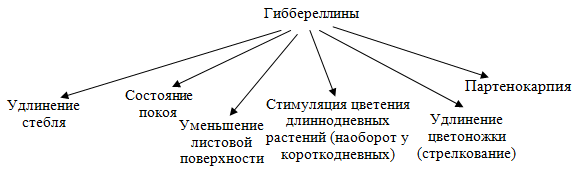
Партенокарпия-образование на растении плодов без оплодотворения
Гиббереллины заметно усиливают вытягивание стебля и у многих нормальных растений. Существует определенная зависимость между скоростью роста стебля растений и содержанием гиббереллинов. Увеличение роста стебля происходит за счет усиления деления клеток и их растяжения. Влияние гиббереллинов на растяжение связано с образованием белка клеточной стенки экстенсина и повышением активности ферментов. Гиббереллины накапливаются в почках при выходе из покоящегося состояния. При выходе семян из покоящегося состояния в них накапливаются гиббереллины. При действии гиббереллина возрастает общая масса растительного организма, он способствует не перераспределению питательных веществ, а общему их накоплению. Показано, что гиббереллин усиливает процесс фотосинтетического фосфорилирования, в первую очередь нециклического, и, как следствие, основных продуктов этого процесса — АТФ и НАДФН. Одновременно наблюдается снижение содержания хлорофилла. Следовательно, под влиянием гиббереллина повышается интенсивность использования единицы хлорофилла, возрастает ассимиляционное число. В темноте гиббереллин воздействует лишь на растяжение клеток, не вызывая возрастания интенсивности их деления.
5. Цитокинины. Природные и синтетические. Открытие, строение, содержание, места синтеза, транспорт и распределение в растениях. Физиологическая активность и механизмы действия. Взаимодействие с другими гормонами.
Учение о цитокининах начинается с открытия полуискусственного продукта – кинетина, который образуется из ДНК при кислотном гидролизе. Природный цитокинин был идентифицирован как зеатин и получен он из неспелых зерновок кукурузы.
Открытие цитокининов связано с обширными исследованиями по выращиванию каллуса, образовавшегося из изолированной ткани сердцевины стебля табака на питательной среде. Было показано, что клетки каллуса в стерильной культуре через определенный промежуток времени прекращают деление. Однако при добавлении к питательной среде производных ДНК, получающихся после ее автоклавирования, деление клеток возобновляется. Все известные цитокинины — это производные пуриновых азотистых оснований, а именно аденина, в котором аминогруппа в шестом положении замещена различными радикалами.
Соединения цитокининового типа обнаруживаются в растениях не только в свободном состоянии, но и в составе некоторых тРНК. Богаты цитокининами клетки апикальных побегов и меристем корня. Цитокинины образуются главным образом в корнях и пассивно в виде зеатинрибозида передвигаются в надземные органы по ксилеме. Вместе с тем имеются данные об образовании цитокининов в семенах (зрелые зародыши) и развивающихся плодах.
Ключевой фермент синтеза цитокининов — изопентенилтра
нсфераза и назван ipt-ген. Она катализирует синтез зеатина и рибозидзеатина из изопентенилпирофосфата. Изопентениловый остаток может образовываться из мевалоновой кислоты. Содержание цитокининов определяется скоростью их синтеза и разложения. Распад цитокининов регулируется ферментом цитокининоксидазой. Имеются сведения, что улучшение питания растений азотом усиливает образование цитокининов.
Основное место синтеза цитокининов – корни; однако в последнее время получены данные о том, что синтез цитокининов может происходить и в семенах (семена гороха).
Из корней цитокинины пассивно транспортируются в наземные органы по ксилеме.
infopedia.su
Фитогомоны ауксины
История изучения ауксинов
Ауксины – это вещества индольной природы. Основным фитогормоном типа ауксина является β-индолилуксусная кислота (ИУК). Открытие ауксинов связано с исследованиями Ч.Дарвина (1860). Дарвин установил, что, если осветить проросток злака с одной стороны, он изгибается к свету. Однако, если на верхушку проростка надеть непроницаемый для света колпачок и после этого поставить в условия одностороннего освещения, изгиба не происходит. Таким образом, органом, воспринимающим одностороннее освещение, является верхушка растения, тогда как сам изгиб происходит в нижней части проростка. Из этого Ч. Дарвин заключил, что в верхушке проростка под влиянием одностороннего освещения вырабатывается вещество, которое передвигается вниз и вызывает изгиб. Идеи Ч. Дарвина получили развитие лишь через 50 лет в работах датского исследователя П. Бойсен-Йенсена, который показал, что если срезанную верхушку вновь наложить на колеоптиль через слой желатины, то при одностороннем освещении наблюдается изгиб к свету. Было показано также, что удаление верхушки проростка (декапитация) резко замедляет рост нижележащих клеток, находящихся в фазе растяжения. При обратном накладывании верхушки проростка через слой желатина или агар-агара рост нижележащих клеток возобновляется. Далее исследования Вента показали, что если срезанную верхушку поместить на блок из агар-агара, а затем наложить этот блок на декапитированный колеоптиль, рост возобновляется. Если агаровый блок, на котором в течение некоторого времени была помещена верхушка колеоптиля, наложить на обезглавленный колеоптиль асимметрично, то происходит изгиб, причём более интенсивно растёт та сторона, на которую наложен блок. Все эти опыты привели к выводу, что в верхушке проростков вырабатывается особое вещество, которое, передвигаясь к нижележащим клеткам, регулирует их рост в фазе растяжения. Поскольку это вещество вырабатывается в одной части растения, а в другой вызывает физиологический эффект, оно было отнесено к гормонам роста растения.
Исследования, проведённые академиком Н.Г. Холодным, показали, что рост различных видов растений, а также различных органов одного и того же растения регулируется одним и тем же гормоном – ауксином. Оказалось, что фитогормоны типа ауксина – β-индолилуксусная кислота (ИУК) и некоторые близкие к ней соединения – широко распространены в растениях. Наиболее богаты ауксинами растущие части растительного организма: верхушки стебля, молодые растущие части листьев, почки, завязи, развивающие семена, а так же пыльца.
Биосинтез и деградация ауксинов
Ауксины синтезируются в растениях разными путями. Ранее считалось, что ИУК образуется единственным путём – из триптофана. Синтез ИУК идёт как минимум в три стадии: декарбоксилирование, дезаминирование, окисление.
Для исследования метаболических путей, ведущих от триптофана к ИУК, получены мутанты, не способные к синтезу триптофана, тем не менее у мутантов синтез ауксина не менялся. Очевидно, есть и другие пути синтеза ИУК, в которых триптофан не участвует. В некоторых растениях таких путей несколько: триптофановый и «нетриптофановые». После образования ИУК может связываться с сахарами, аминокислотами или небольшими белками, образуя неактивные (запасные) формы. При необходимости ИУК освобождается из конъюгатов.
В упомянутых опытах Ч. Дарвина с проростками злаков ауксин в апексе не синтезируется! ИУК образуется в период роста зародыша, а при созревании накапливаются ИУК-гликозиды. В проростках они транспортируются в апекс колеоптиля, где активный ауксин высвобождается и перераспредиляется по колеоптилю в зависимости от освещенности. Колеоптиль – временный орган прорстка злаков, необходимый для пробивания почвы. Внутри него заключена апикальная меристема побега, образующая стебель и листья. Именно в апикальной меристеме сосредоточен синтез ауксинов. ИУК может необратимо разрушаться. Это происходит либо специфически (с помощью ИУК-оксидазы), либо неспецифически (полифенолоксидазой).
Таким образом по мере удаления от точки синтеза концентрация ауксинов падает за счет необратимого окисления и связывания в неактивные формы.
Физиологические проявления действия ауксинов
Наиболее ярким проявлением физиологического действия ауксина является его влияние на рост клеток в фазе растяжения. ИУК стимулирует выход протонов в клеточную стенку и увеличивает её растяжимость. Под влиянием оптимальной концентрации ИУК рост в длину декапитированных отрезком стеблей гороха увеличивается более, чем в два раза. Ауксины в некоторых случаях стимулируют деление клеток, например камбия. Под влиянием ауксинов может измениться направление дифференциации клеток. Ауксин вызывает дифференциацию ксилемы, индуцирует корнеобразование.
Все проявления роста клеток находятся в определённой зависимости от содержания природных ауксинов. Большую роль играют ауксины при разрастании завязи и плодообразования. Показано, что ауксины могут синтезироваться в пыльце, зародыше, эндосперме. На первых стадиях роста плодов ауксин поступает из эндосперма, а позднее – из зародыша. У некоторых растений ауксин стимулирует образование бессемянных (партенокарпических) плодов. Ауксины, так же как и другие фитогормоны, обуславливает взаимодействие отдельных органов растения (коррелятивный рост). Ауксин обуславливающий явление апикального доминирования, проявляющегося в подавляющем влиянии верхушечной почки на рост пазушных. Участие ауксина в этом процессе установлено в 1935 г. Прямые определения показали, что через 4 часа после декапитации содержание ауксина в узлах увеличивалось в 5 раз. С участием ауксина связана также регуляция двигательной активности растений, в частности тропизмы и настии.
Действие ауксина находится в зависимости от его концентрации. Повышение концентрации ауксина выше оптимальной вызывает торможение роста. При этом для разных растений и для неодинаковых организмов оптимальная концентрация ауксина резко различна. Одна и та же концентрация может усилить рост одних органов и затормозить другие. Так, оптимальная концентрация для роста стебля составляет около 10мг, тогда как для корня всего 0,01мг ИУК на 1 кг массы растения. Концентрация ауксинов, усиливающая рост злаков, резко тормозит рост многих двудольных растений. Это может быть связано с тем, что ауксин стимулирует синтез другого фитогормона, ингибирующего рост, а именно этилена.
При всех физиологических проявлениях ауксины усиливают поступление воды и питательных веществ (аттрагирующее влияние). Имеются многочисленные данные, что ауксины являются регуляторами притока воды и питательных веществ. Ауксины влияют на распределение питательных веществ в растении. При внесении извне ауксинов или их синтетических аналогов они концентрируются в отдельных органах и клетках. Это вызывает приток к этим органам воды и питательных веществ и, как следствие, их усиленный рост. Одновременно рост других органов, содержащих меньше ауксинов, ослабляется, поскольку питательные вещества к ним поступают в меньшем количестве. Так, при обработке фитогормонами типа ауксина цветков томата происходит усиленное разрастание завязей, приток к ним питательных веществ значительно повышается, а рост боковых побегов тормозится. Общий вынос питательных веществ при этом не изменяется. Из этих опытов следует, что гормоны типа ауксина вызывают перераспределение питательных веществ в растении. В некоторых случаях ауксин может вызывать усиление притока питательных веществ из внешней среды.
Существуют разные гипотезы, объясняющие действие ауксина на передвижение веществ. Не вызывает сомнения, что транспорт веществ по растительному организму определенным образом связан с напряженностью энергетического обмена. В этой связи важно отметить, что ещё в 1933 г. появились исследования, показывающие, что под влиянием ауксина интенсивность дыхания растёт. В отсутствие кислорода действие ауксина или не проявляется, или значительно ослабевает. На различных объектах установлено, что влияние ауксина на повышение интенсивности дыхания опережает во времени его действие на ростовые процессы. Под влиянием ИУК возрастает сопряженность окисления и фосфорилирования (коэффициент Р/О) и содержание в клетках АТЫ. Это даёт основание считать, что ИУК увеличивает энергетическую эффективность дыхания растений. Под влиянием ИУК возрастает и энергетической заряд клетки (отношение АТФ + АДФ к АМФ). Известно, что даже небольшие сдвиги в энергетичекомпотенциале клетки приводят к заметным изменениям в скорости различных ферментативных реакций. Положительные сдвиги в энергетическом обмене вызывают усиление передвижения питательных веществ и воды, что является одной из причин усиления роста растений. Решение вопроса о причинах усиления образования АТФ под влиянием ИУК связано с изучением первичных механизмов регуляторного влияния этого фитогормона.
Ауксин и плоды
Ауксин стимулирует развитие бессемянных плодов. В норме плоды с многочисленными семенами лучше обеспечены питательными веществами, чем те, в которых семян нет. В силу аттрагирующего эффекта ауксинов чем больше семян образовалось в плоде (чем больше точек синтеза ауксинов), тем лучше плод обеспечен питательными веществами. При обработке бессемянных плодов ИУК у растения создаётся «иллюзия», что семена в них есть. Туда направляются питательные вещества, стимулируя развитие плодов.
Ауксин как гербицид
Если ауксинов слишком много, то начинается синтез гормона-антагониста этилена. Этилен угнетает рост побегов и корней в длину и вызывает листопад. При обработке естественными ауксинами (ИУК, ИПВК и др.) начинается их окисление и/или образование конъюгатов. Систем инактивации синтетических аналогов (НУК, 2,4-Д) у растений нет. При обработке растений 2,4-Д синтезируются антагонисты (в основном этилен), рост угнетается, листья желтеют и опадают и т.д. У двудольных под действием 2,4-Д отмирают кончики корней, и это позволяет использовать 2,4-Д как селективный гербицид. На
Гербицидные свойства ауксинов могут проявиться при укоренении черенков: превышенные дозы препарата или времени обработки ведёт к синтезу этилена, что неблагоприятно для растений. При подготовке статьи использовалась литература: Физиология растений: под. ред. И.П. Ермакова, Физиология растений под. ред. Якушкина, Н.И.
biofile.ru










