Прямые (контактные) взаимодействия между растениями. Взаимоотношения между растениями и растениями
Формы взаимоотношений между растениями
Никогда не бывает так, что какое-нибудь растение имеет в качестве среды для своего существования и развития только среду неорганическую.
Растения живут большими массами, и для каждого индивидуума совокупность всех остальных индивидуумов играет не меньшую роль, чем совокупность экологических факторов неорганического происхождения. Корневые системы разных или одинаковых растений конкурируют в извлечении из почвы влаги и питательных веществ, надземные органы ведут борьбу за свет; в густом насаждении возникают иные условия температуры, влажности, движения воздуха, нежели в насаждении разреженном. Поэтому растения и «борются» друг с другом (чаще всего через среду) и в то же время взаимно приспособляются. Взаимоотношения между растениями могут быть сведены к немногим основным формам.
Сапрофитами называются растения, которые добывают себе средства к жизни из мёртвого органического вещества, так как, будучи лишёнными хлорофилла, они сами не в состоянии ассимилировать CO2. К сапрофитам относятся слизевики (Myxophyta), обитающие на гниющем дереве, коре, экскрементах животных; многие бактерии, вызывающие минерализацию органических веществ; миксобактерии, живущие на помёте животных; многие грибы, например плесени, дрожжи, огромное большинство шляпочных грибов и т. д.
В мире микроскопических организмов большую роль играют явления антагонизма, выражающиеся в том, что многие актиномицеты (лучистые грибки), плесени и бактерии вырабатывают химические вещества (антибиотики), способные подавлять жизнедеятельность других микробов либо убивать их. На этом, в частности, основывается сейчас лечебное применение антибиотиков, которые, будучи введены в организм больного, поражают там возбудителей болезни, не принося в то же время вреда клеткам человеческого тела. Для излечения заражения крови, менингита, ангин, возвратного тифа и др. широко применяется пенициллин (антибиотик некоторых плесневых грибков), для лечения разных форм туберкулёза — стрептомицин (антибиотик актиномицетов), для лечения гнойных ран, дизентерии, дифтерии — грамицидин (выделяемый некоторыми почвенными бактериями) и т. д.
Широко распространённой формой сожительства растений является паразитизм, т. е. такая физиологическая связь, когда одно растение, называемое паразитом, пользуется готовыми питательными веществами другого организма, называемого хозяином, нанося последнему ущерб и нередко доводя хозяина до гибели. В этом отношении особенно крупная роль принадлежит болезнетворным микробам, вызывающим болезни растений, животных и людей. Заболевания растений чаще всего вызываются грибами, но также бактериями и вирусами. Внутрь растений бактерии проникают через повреждения в тканях или через естественные отверстия {поры, устьица). Следствием их деятельности в растениях является отмирание клеток, ненормальное их разрастание, «гниль» (моркови, томатов), увядание растений, разрастание тканей («рак»), наросты на стволах и корнях, истечение камеди и пр.
Из грибов к паразитам относятся некоторые сумчатые грибы, вызывающие на листьях многих растений болезнь «мучную росу»; головнёвые грибы; паразитирующие на деревьях трутовики; гриб спорынья, поражающий завязи ржи и некоторых других злаков. Из более высоко организованных растений к паразитам относятся омела, многие виды семейства раффлезиевых, семейства заразиховых, петров крест, паразитирующий на корнях ясеня, ольхи, вяза, тополя и некоторых других лиственных пород.
Сожительство двух организмов, извлекающих из этого сожительства взаимную пользу, называют симбиозом. Исторически симбиоз должен был развиться из паразитизма: гибель хозяина означала вместе с тем и гибель паразита, и поэтому умеренный паразитизм имел в борьбе за существование известные преимущества. Он в конце концов и превратился в симбиоз. В большом количестве случаев, когда речь идёт о симбиозе, можно в нём. обнаружить именно признаки либо одностороннего ограниченного паразитизма, либо чередующегося взаимного паразитизма.
Классическим примером симбиоза в растительном мире долго считались лишайники, представляющие сожительство гриба и водоросли. Впоследствии оказалось, что в лишайнике налицо сложное сочетание симбиоза и паразитизма. Грибная ткань в лишайнике представлена чаще всего различными сумчатыми грибами, а водоросли — одноклеточными зелёными или синезелёными водорослями. Хозяином здесь является водоросль, обладающая способностью в фотосинтезу, паразитом же гриб; вместе с тем гриб тоже доставляет водоросли часть необходимой ей пищи.
От паразитов необходимо отличать эпифиты, т. е. такие растения, которые совершенно потеряли связь с почвой, живут на других растениях, но физиологически с ними не связаны. Прикрепляются они обычно с помощью присосков-ризоидов, которые проникают немного вглубь мёртвой коры хозяина, либо с помощью присосков-корней, плотно прижимающихся к субстрату. Зольные вещества эпифит добывает из частиц перегноя и пыли, осевших на хозяине, а воду впитывает либо всей поверхностью своего тела (как это имеет место у водорослей, мхов, лишайников), либо с помощью особых воздушных корней. В наших широтах к эпифитам относятся некоторые лишайники, живущие на коре деревьев, бородатый лишай (Usnea barbata), обитающий на ветвях, и т. п. Особенно огромное количество эпифитов свойственно флорам влажных тропиков, причём, помимо низших растений, здесь в качестве эпифитов встречаются папоротники, а также цветковые — как травянистые, так и деревянистые.
Наконец, отметим ещё группу лиан — лазающих и вьющихся растений, которые нуждаются в других растениях только как в опоре. Некоторые авторы относят к лианам только древесные растения со слабыми главными осями, нуждающиеся в подпорках, подставках и пр., за которые они цепляются усиками (виноград), корнями (плющ) и т. п. Другие же авторы относят к лианам вообще всякие лазающие и вьющиеся растения. Примерами лиан могут служить лазающие розы, плющ, ваниль, хмель, вьюнок, жимолость, виноград, многие виды бобовых и тыквенных и т. п.
Как уже было отмечено, растения живут не изолированно друг от друга, а целыми группами. Эти группы отнюдь не случайны, а представляют собой характерную комбинацию различных растений. Такого рода характерные комбинации получили название фитоценозов, или растительных сообществ. Фитоценоз — это вполне закономерное сочетание различных видов растений, сложившееся в результате весьма длительного исторического процесса под давлением конкурентных отношений между слагающими растениями и находящееся в связи с внешними условиями существования.
Лес, луг, степь — это фитоценозы. Лес лиственный и лес хвойный — тоже фитоценозы. Иначе говоря, фитоценоз — это общее понятие для различных закономерных растительных объединений.
В фитоценозе растения влияют друг на друга не только прямо (паразиты, эпифиты, симбионты), но и косвенно — через изменение среды (конкуренция за воду, минеральные соли, воздушное питание, затенение, движение воздуха и т. и.).
Если вы нашли ошибку, пожалуйста, выделите фрагмент текста и нажмите Ctrl+Enter.
www.activestudy.info
Взаимоотношения между растениями
Взаимоотношения между растениями довольно разнообразны, существенно влияя на жизнь растительных организмов и фитоценозов. Наиболее часто используется классификация взаимоотношений растений, предложенная В. Н. Сукачевым (1956). Он различал три основные формы влияния растений друг на друга при их совместном произрастании: прямые и косвенные — трансабиотические и трансбиотические (см. разд. 11.1). В прямых (контактных) отношениях растения действуют друг на друга непосредственно, а в косвенных — через посредство другого агента. При прямых отношениях растения могут механически давить, разрушать друг друга, поселяться друг на друге, питаться друг другом. При косвенных посредником могут быть неживая среда (трансабиотические отношения) и другие организмы (трансбиотические отношения). Среди трансабиотических отношений можно выделить конкуренцию за ресурсы, создание фитосреды и влияние через выделения (прижизненные и от разлагающихся остатков). Все формы отношений растений проявляются одновременно, действуют друг на друга, и поэтому в природе их влияние обычно трудно разграничить.
Формы прямых (контактных) взаимоотношений растений. В. Н. Сукачев (1956) различал две формы контактных взаимоотношений растений: со срастанием и без него. Обе довольно широко распространены в отношениях паразит-хозяин, лиана-опора, эпи- фит-форофит. Особый вариант контактных взаимоотношений — срастание корней, распространенное у древесных растений. Наиболее часто срастаются корни одной особи, реже — особей одного вида и еще реже — разных видов. С возрастом древостоя срастание корней существенно увеличивается. Экспериментально доказана возможность переноса веществ (включая органику) через сросшиеся корни.
При совместном произрастании нередко одни растения создают механические препятствия для нормальной жизнедеятельности других. Например, перехват пыльцы снижает вероятность опыления цветков. Плохо влияет на семенное размножение задерживание плодов и семян мхами, лишайниками и надземными органами других растений. Сплошной мохово-лишайниковый покров снижает также всхожесть семян и иногда приводит к гибели всхо
дов, что связано с истощением запасов семени до достижения корешком проростка почвы, с колебаниями влажности в слое мхов и лишайников, с «выдергиванием» всходов при набухании увлажненных лишайников.
Плоды и пыльца, оседая на поверхности листьев, могут сокращать их фотосинтезирующую поверхность. Такое воздействие плоских плодов вяза (Ulmus), обильной пыльцы сосны (Pinus) и ели (Picea) недолговременно. Однако растения могут надолго создавать препятствия для перемещения подземных органов других растений, которые вынуждены обходить препятствие или пронизывать его. Так, Т. А. Работнов (1993) отмечает прободение корневищами пырея ползучего (Elytrigia repens) и свинороя (Cynodon), корневищ щавеля конского (Rumex confertus), корней клевера лугового (Trifolium repens) и клубней картофеля (Solanum tuberosum). В этих взаимоотношениях один вид наносит другому вред, однако тратит на это много энергии.
Угнетение одних растений другими происходит и при передвижении особей в результате разрастания. Например, у белоуса (Nardus stricta) молодые растения испытывают давление разрастающихся мощных особей. И хотя всходы обычно гибнут от затенения и перехвата взрослыми особями воды и минеральных элементов раньше их физического уничтожения передвигающимися растениями, судьба молодых экземпляров может зависеть и от того, где по отношению к более мощным соседям они возникли. Быстрорастущие водоросли часто механически вытесняют медленнорастущие. А в некоторых случаях, например при гнездовом посеве моркови (Daucus carota), может происходить «выпирание одних особей другими».
eco-rasteniya.ru
|
Студопедия
АвтоАвтоматизацияАрхитектураАстрономияАудитБиологияБухгалтерияВоенное делоГенетикаГеографияГеологияГосударствоДомЖурналистика и СМИИзобретательствоИностранные языкиИнформатикаИскусствоИсторияКомпьютерыКулинарияКультураЛексикологияЛитератураЛогикаМаркетингМатематикаМашиностроениеМедицинаМенеджментМеталлы и СваркаМеханикаМузыкаНаселениеОбразованиеОхрана безопасности жизниОхрана ТрудаПедагогикаПолитикаПравоПриборостроениеПрограммированиеПроизводствоПромышленностьПсихологияРадиоРегилияСвязьСоциологияСпортСтандартизацияСтроительствоТехнологииТорговляТуризмФизикаФизиологияФилософияФинансыХимияХозяйствоЦеннообразованиеЧерчениеЭкологияЭконометрикаЭкономикаЭлектроникаЮриспунденкция |
В лесном сообществе взаимодействия между растениями, имея в виду цветковые и высшие споровые растения, особенно многогранны и сложны. Они осуществляются либо через непосредственное влияние одних на другие (паразитизм, например, омелы на сосне, липе и др.), через срастание корней, которое позволяет использовать поглощаемые вещества корневой системы одного растения другим, через охлестывание и повреждение ветвями одного дерева другим, например, ели березой, непосредственным давлением корневых систем и т. п., либо через другие организмы (микроорганизмы, животные), либо через выделение веществ жидких, газообразных и твердых надземными и подземными частями, действующих положительно или отрицательно на другие растения, через конкуренцию корневых систем из-за воды и минеральных веществ, через изменение светового и водного режима. Другие «влияния» растений: ослабление действия ветра, защита от ветровала и бурелома; накапливание из отмирающих и опадающих остатков растений, листьев, ветвей, плодов, семян и т.п. лесной подстилки, которая не только косвенно влияет на растения через изменение почвенных процессов, но и создает особые условия для прорастания семян и развития всходов и т.п. Изучение биоморф как моделей наиболее существенных экологических черт видов перспективно при выяснении общих ценогеографических закономерностей. д) Взаимосвязь растительности с животным миром Не менее тесна взаимосвязь растительности с животным миром, населяющим данный биогеоценоз. Животные в процессе своей жизнедеятельности многосторонне влияют на растительность как непосредственно, питаясь ею, вытаптывая ее, строя в ней или при помощи ее свои жилища, убежища, содействуя опылению цветков и распространению семян или плодов, так и косвенно, изменяя почву, удобряя ее, разрыхляя, вообще меняя ее химические и физические свойства, а в некоторой степени влияя и на атмосферу. ПРИМЕР. Изучение отдельных комплексов вредителей определенных видов или других подсистем ценотического порядка (паразито-, микробоценозов и др.) в совокупности с растениями-"хозяевами", т.е. консорций, перспективно в прикладном отношении – для использования их в области защиты растений. Взаимоотношения между разными трофическими уровнями относятся к трофико-энергетическому направлению (Odum, 1963) и являются объектом многих исследований, получившим широкое развитие в последние десятилетия. Это дает возможность выявить общий характер и количественные показатели обмена веществ и энергии, раскрывая тем самым биогеофизическую и биогеохимическую роль живого покрова. е) Взаимодействия между неживыми (абиотическими) компонентами Не только живые организмы взаимодействует с другими компонентами биогеоценоза, но и эти последние взаимодействуют друг с другом. Климатические условия (атмосфера) влияют на почвообразовательный процесс, а почвенные процессы, определяя выделение углекислоты и других газов (дыхание почвы), изменяют атмосферу. Почва влияет на животный мир, не только ее населяющий, но косвенно и на весь остальной животный мир. Животный же мир воздействует на почву.
Факторы, влияющие на взаимодействия компонентов биогеоценоза Рельеф и биогеоценоз. Всякий биогеоценоз, занимая известное место в природе, связан с тем или иным рельефом. Но рельеф сам по себе не входит в число компонентов биогеоценоза. Рельеф является лишь условием, влияющим на процесс взаимодействия названных выше компонентов, и, в соответствии с этим, на их свойства и структуру, определяя направление и интенсивность процессов взаимодействия. В то же время взаимодействие компонентов биогеоценоза нередко может приводить к изменению рельефа и к созданию особых форм микрорельефа, а в известных случаях и мезо- и макрорельефа. Влияние человека на биогеоценоз. Человек не входит в число компонентов биогеоценозов. Он, однако, является в высшей степени мощным фактором, могущим не только в той или иной мере изменять, но и создавать путем культуры новые биогеоценозы. Ныне почти нет лесных биогеоценозов, не испытавших влияние хозяйственной, а часто и бесхозяйственной деятельности человека. Взаимовлияния между биогеоценозами. Вместе с тем каждый биогеоценоз, так или иначе, влияет на другие биогеоценозы и вообще явления природы, соседние с ним или в той или иной мере удаленные от него, т. е. обмен веществом и энергией имеет место не только между компонентами данного биогеоценоза, но и между самими фитоценозами. Часто ведущим фактором являются конкурентные отношения между фитоценозами. Фитоценоз более мощный вытесняет фитоценоз менее устойчивый, например, приопределенных условиях происходит вытеснение соснового фитоценоза еловым, а вместе с этим изменяется и весь биогеоценоз. Таким образом, взаимодействие всех компонентов биогеоценоза, особенно лесного биогеоценоза (включая воду в почвогрунтах и атмосфере), очень разнообразно и сложно: - растительность все время находятся в зависимости от почвы, атмосферы, животного мира и микроорганизмов. - химический состав почвы, ее влага и физические свойства влияют на рост и развитие растений, на их плодоношение и возобновляемость, на технические свойства их древесины древесных пород, на их, на рост и развитие всей другой растительности. - вся растительность же в свою очередь в сильной степени воздействует на почву, определяя, главным образом, качество и количество органического вещества в почве, влияя на физические и химические ее особенности.
Типы отношений между организмами в ценнозе. Организмы могут взаимодействовать друг с другом постоянно, в течение всей жизни или кратковременно. При этом они либо соприкасаются друг с другом, либо оказывают влияние на другой организм на расстоянии. Взаимные влияния растений могут иметь то благоприятный для их роста и развития характер, то неблагоприятный. В первом случае условно говорят о «взаимопомощи», во втором – о «борьбе за существование» между растениями в широком, дарвинском смысле, или о конкуренции. Само собой разумеется, что все эти взаимовлияния между организмами в биоценозе в то же время играют большую роль и в биогеоценозе в целом. Они могут проходить между особями как разных видов, так и одного и того же вида, т. е. могут быть как межвидовыми, так и внутривидовыми.
СИМБИОЗ ("ВЗАИМНЫЙ ПАРАЗИТИЗМ")
– отношения между организмами, обычно разных видов и находящихся в более или менее длительном контакте, при которых один или оба организма извлекают из этих отношений пользу и ни один не испытывает ущерба. Первый тип симбиотических отношений, когда пользу извлекают оба организма, называется мутуализмом, второй, когда пользу извлекает лишь один из организмов, – комменсализмом («нахлебничеством»). Мутуализм Симбиоз азотфиксирующих организмов с голосеменными и цветковыми растениями – отношения между высшим растением и бактериями. На корнях многих растений встречаются клубеньки, образованные бактериями или реже грибами. Клубеньковые бактерии фиксируют атмосферный азот и переводят его в доступную для высших растений форму. Микориза – симбиотические отношения между высшим растением и грибом. Микоризы широко распространены среди диких и культурных растений. В настоящее время микориза известна более чем для 2000 видов высших растений (Федоров, 1954), но, несомненно, действительное число видов, для которых характерна микориза, значительно больше. Для высших растений, на корнях которых поселяются грибы, характерен особый тип питания – микотрофный. При микотрофном питании с помощью симбиотических грибов высшее растение получает зольные элементы пищи, в том числе азот, из органического вещества почвы. Что касается грибов, образующих микоризу, то они в большинстве не могут существовать без корневых систем высших растений, всасывающих влагу из почвы и поставляющих из кроны органическое вещество. Деревья растут гораздо лучше с микоризой, чем без нее. Различают два основных вида микоризы: эктотрофную и эндотрофную. При эктотрофной микоризе корень высшего растения окутывается плотным грибным чехлом, от которого отходят много численные гифы гриба. При эндотрофной микоризе мицелий гриба проникает в клетки корневой паренхимы корня, которые сохраняют свою жизнедеятельность. Промежуточную форму микоризы, при которой имеет место и наружное обрастание корня гифами гриба, и проникновение гифов внутрь корня, называют перитрофной (эктоэндотрофной), микоризой. Эктотрофная микориза – однолетняя. Она развивается летом или осенью и погибает к следующей весне. Она свойственна многим деревьям из семейств сосновых, буковых, березовых и др., а также некоторым травянистым растениям, например подъельнику. Эктотрофную микоризу чаще всего образуют базидиальные грибы из семейства Polyporaceae и особенно часто из рода Boletus. Так, подберезовик (В. scaber) образует микоризу на корнях березы, масленок – на корнях лиственницы (В. elegans) или сосны и ели (В. luteus), подосиновик (В. versipellis) – на корнях осины, белый гриб (В. edulus) – на корнях ели, дуба, березы (различные подвиды) и т. д.
Эндотрофная микориза широко распространена у растений семейств орхидных, вересковых, брусничных, а также у многолетних трав из семейства сложноцветных и у некоторых деревьев, на пример у красного клена (Acer rubrum) и др. В качестве второго компонента эндотрофной микоризы часто выступает гриб Phoma из группы несовершенных грибов. Эндотрофная микориза может быть образована Oreomyces (живет на корнях орхидей, по-видимому, может фиксировать азот) и некоторыми другими видами грибов. Как предполагалось ранее, этот гриб может усваивать азот из атмосферы. Это обстоятельство связано с тем, что вереск (Calluna) и другие представители семейства вересковых, а также виды семейства орхидных могут развиваться на безазотистой среде только в присутствии этого гриба. В отсутствие Phoma betake у этих растений не прорастают семена или гибнут проростки вскоре после прорастания семян. Гибель проростков у орхидей, грушанок и других лесных растений можно объяснить тем, что в их семенах почти полностью отсутствуют запасные питательные вещества в клетках, и поэтому без гифов гриба, доставляющих необходимые питательные вещества всходам, их развитие быстро прекращается.В сосновых лесах Центрального Предуралья (Логинова, Селиванов, 1968) насчитывается следующее содержание микотрофных видов в лесной микофлоре: в бору-беломошнике – 81%, в бору-брусничнике – 85, в бору-черничнике – 90, в бору сфагново-багульниковом – 45, в бору остепненном травяном – 89%. В пустынях Тау-Кум процент видов с микоризой в разных ассоциациях составляет от 42 до 69%. Значение микоризы в связи с ее широким распространением огромно. Многие растения – орхидные и вероятно вересковые, а также некоторые деревья без микоризы развиваются плохо или даже вообще не развиваются либо из-за недостатка питательных веществ в их мелких семенах, либо из-за недостаточного развития сосущих частей корешков, а также на бедных минеральными питательными веществами почвах. Грибы, образующие эндотрофную микоризу на их корнях, могут существовать лишь в кислой среде. Именно благодаря им многие представители орхидных и вересковых потому и живут только на кислых почвах. Следовательно, наличие в фитоценозе грибов, образующих микоризу, в значительной степени определяет видовой состав высших растений, входящих в этот фитоценоз, и служит немаловажным фактором в их борьбе за существование между растениями, поскольку отсутствие микоризы у растений, склонных к микотрофномy питанию, замедляет темпы их развития и ухудшает их положение по отношению к более быстро развивающимся видам, использующим микоризу. Коменсализм Наиболее характерные растения, которые можно привести в качестве примеров комменсализма по способу размещения в ценозе и по типу питания: эпифиты, лианы, почвенные и наземные сапрофиты.
Эпифиты - растения, как высшие, так и низшие, растущие на других (хозяевах): деревьях, кустарниках, которые служат ему опорой. Отношения эпифитов к их хозяевам могут быть определены как комменсализм, при котором один из вступающих в эти отношения видов получает какое-либо пре имущество, а второй не терпит ущерба. Преимущество при этом получает эпифит. Чрезмерное развитие эпифитов на стволах и ветвях может угнетать и даже быть причиной поломки ствола растения-хозяина. Эпифиты могут затруднять рост и ассимиляцию, а также способствовать загниванию тканей хозяина вследствие повышения влажности. На дереве (рис.1) различают четыре местообитания эпифитов (Ochsner, 1928).В зависимости от условий существования эпифиты (Ричардс, 1961) делятся на три группы: теневые, солнечные и крайне ксерофильные.
Теневые эпифиты обитают в условиях сильной затененности, малого и мало меняющегося дефицита насыщения, т. е. в условиях, почти не отличающихся от условий жизни наземных трав. Они обитают главным образом в третьем (нижнем) ярусе леса. Многие из них имеют гигроморфную структуру тканей. Группа солнечных эпифитов, наиболее богатая по числу видов и особей, связана с кронами деревьев верхних ярусов. Эти эпифиты живут в микроклимате, промежуточном между микроклиматами напочвенного покрова и открытых мест, и получают гораздо больше света, чем теневые эпифиты. Многие солнечные эпифиты более или менее ксероморфны; осмотическое давление у них выше, чем у теневых эпифитов. Крайне ксерофильные эпифиты живут на верхушечных ветвях более высоких деревьев. Условия их местообитания подобны условиям открытых мест, условия питания здесь крайне суровы. Эпифиты, как правило, сапротрофы, т.е, питаются за счет отмирающих тканей растения-хозяина. Обычно для разложения этих отмирающих тканей эпифиты используют грибы, образующие микоризу с корнями эпифита. Большую роль в питании играют некоторые животные. Лианы. К лианам относят высшие растения со слабыми стеблями, нуждающимися в какой-нибудь опоре, чтобы подняться вверх. Лианы, относятся к комменсалам, но изредка могут причинить ущерб и даже вызвать гибель деревьев. Лианы делят на две группы: мелкие и крупные. Среди мелких лиан преобладают травянистые формы, хотя есть и деревянистые. Они развиваются в нижних ярусах лесов, а иногда (вьюнок – Convolvulus, подмаренник – Galium, марена – Rubia, княжик – Clematis, и др.) и среди травяного покрова. Крупные лианы обычно бывают деревянистыми. Они достигают вершин деревьев второго, иногда первого яруса. У этих лиан обычно очень длинные и иногда настолько крупные водоносные сосуды, что они видны на поперечном срезе простым глазом. Эта особенность связана с необходимостью поднимать огромные количества воды в крону лианы, иногда не уступающую по величине кроне дерева, по стволу, диаметр которого во много раз меньше диаметра обычного дерева. Стебли лиан часто имеют очень длинные междоузлия и быстро растут не ветвясь, пока не достигнут того яруса, в котором обычно развертывается листва этих растений. В "уссурийской тайге" наряду с мелкими лианами растут крупные (рис.3), придавая особый колорит приморским лесам. Длина взрослых лиан актинидии и винограда амурского достигает нескольких десятков метров, а диаметр – 10 и более сантиметров. Крупные лианы иногда так быстро растут и развиваются в таких массах, что губят поддерживающие их деревья. Вместе с деревом-опорой лиана падает на землю и здесь гибнет или поднимается на другое дерево. Часто расстояние между основаниями стволов лианы и дерева-опоры измеряется десятком или несколькими десятками метров, что убеждает в том, что несколько промежуточных деревьев, служивших лиане опорой, погибли ранее. Часто лианы фестонами перекинуты с одного дерева на другое, достигая длины 70, а в исключительных случаях (ротанговые пальмы) 240 м. В лесах умеренного пояса распространены исключительно или почти исключительно мелкие лианы, поэтому они не играют здесь большой роли. Почвенные и наземные сапрофиты. Сапрофиты – растительные организмы, живущие полностью (полные сапрофиты) или частично (частичные сапрофиты) за счет отмерших органов животных и растений. Кроме эпифитов, которые относятся по способу питания к сапрофитам, в эту группу входят многие наземные растения и обитатели почвы. ПРИМЕРЫ. К сапрофитам относится большинство грибов и бактерий, играющих огромную роль в круговороте веществ в почве, а также некоторые цветковые растения из семейств орхидных (гнездоцветка клобучковая) и вертляницевых (вертляница одноцветковая) в лесах умеренного пояса и из семейств лилейных, орхидных, горечавковых, истодовых и некоторых других в лесах тропического пояса.Большинство этих цветковых растений – полные сапрофиты, некоторые, по крайней мере, орхидеи, содержат какое-то количество хлорофилла и, вероятно, частично способны к фотосинтезу. Цвет надземных частей этих растений белый, светло-желтый, розовый, голубой или пурпурный. Обитают сапрофиты из цветковых растений в тропиках в тенистых местах на почве или на лежащих мертвых стволах. Обычно эти растения связаны с микоризными грибами, живущими на их корнях. Как правило, они невысоки, обычно не превышают 20 см, за исключением сапрофитной тропической орхидеи галеоны высочайшей (Gualala altissimo), представляющей собой лазающую (при помощи корней) лиану, достигающую высоты 40 м. АНТАГОНИЗМ - отношения, при которых один или оба организма испытывают ущерб. Душители. Душители – корнесобственные растения, но начинают развитие в качестве эпифитов. Различные животные переносят их семена с одного дерева на другое. Птицы – основные переносчики семян душителей. Душитель образует корни двух родов: одни из них плотно прилегают к коре дерева-хозяина, ветвятся, и образуют плотную сеть, одевающую ствол дерева-хозяина, другие свешиваются вертикально вниз и, достигнув почвы, ветвятся в ней, доставляя душителю воду и минеральное питание. В результате затенения и сдавливания дерево-хозяин погибает, а душитель, к тому времени развивший мощный корневой «ствол», остается стоять на «собственных ногах». Многочисленные лианы фестонами свисают с дерева.Душители характерны для влажных тропиков. Душители находятся с деревьями-хозяевами в антагонистических отношениях. Некоторые южноамериканские виды-душители имеют настолько слабые корни, что при падении дерево-хозяин увлекает их за собой.Паразиты и полупаразиты. Паразитизм – отношения между организмами обычно разных видов, при которых один из них (паразит) проводит более или менее длительное время на теле или в теле другого организма (хозяина), получая питание из его тканей.Паразитические растения играют огромную роль в жизни фитоценозов, ослабляя одни растения, вызывая гибель других и тем самым, способствуют лучшему развитию третьих.Паразитов делят на две большие группы: факультативные и облигатные. Первые могут вести и паразитический, и сапрофитный образ жизни, как, например, грибы трутовики из рода полипорус (Polyporus), вторые ведут только паразитический образ жизни. В связи с этим облигатные паразиты обычно не сразу губят своего хозяина, а постепенно истощают его, в то время как факультативные паразиты весьма быстро приводят своего хозяина к гибели и переходят на питание его мертвыми тканями. Поэтому факультативные паразиты оказывают более существенное воздействие на растение-хозяина, чем облигатные.К числу паразитов относятся многие грибы, некоторые слизевики, водоросли и лишайники, а также ряд видов цветковых растений. Так, повилика европейская (Cuscuta europaea) иногда сильно поражает крапиву двудомную (Urtica dioica), а повилика хмелевидная (С. lupuliformis) и однопестичная (С. Monogyna) – иву (Salix). Паразитами следует считать и бактерии, вызывающие заболевания растений, животных и человека. Кроме полных паразитов, существуют гемипаразиты (полупаразиты). Последние, имея хлорофилл, способны к ассимиляции и от растения-хозяина получают преимущественно минеральное питание в водном растворе.Часть полупаразитов представляют виды, обычно обитающие на лугах, иногда в лесах, с очень слабыми корневыми системами. Эти растения присасываются своими корнями к корням цветковых растений. Это многие виды семейства норичниковых (Scrophulariaceae) из родов мытник (Pedicularis), марьянник (Melampyrum), зубянка (Odontites), очанка (Euphrasia), погремок (Alectorolophus) и др. Массовое их развитие может резко снизить качество луга, так как ценные в хозяйственном отношении травы сильно истощаются и резко снижают урожай.В условиях умеренного климата наиболее широко распространена омела белая (Viscum album) на лиственных, реже на хвойных породах.Хищничество – отношения между организмами разных видов (если организмы относятся к одному виду, то это каннибализм), при которых один из организмов (хищник) питается за счет второго организма (жертвы). Антибиоз – отношения между организмами, обычно относящимися к разным видам, при которых один из организмов причиняет вред другому (например, выделением вредных для другого организма веществ), не извлекая для себя из этих отношений видимого преимущества.Воздействие выделений одних растений на другие. Взаимоотношения между растениями, в которых ведущую роль играют специфически действующие продукты обмена веществ, Молиш (Molisch, 1937) назвал аллелопатией. Вещества, выделяемые надземными и подземными органами живых растений, и органические соединения, получающиеся в процессе разложения мертвых растительных остатков и оказывающие влияние на другие растения, называются колинами. Колины образуются как при свободной жизни растений рядом друг с другом, так и при паразитизме и взаимном влиянии пыльцы разных видов. Выделения веществ могут оказывать положительное или отрицательное влияние на растения того же вида или других видов. В частности, антибиотические вещества, препятствующие росту различных плесеней и бактерий, выделяют плесневые грибы, бактерии, актиномицеты, а также некоторые водоросли и лишайники. Эти выделения защищают растения от нападающих на них паразитов и тем самым ограничивают число видов, входящих в состав фитоценоза, или затрудняют развитие некоторых из них. Среди колинов различают: - газообразные выделения надземных органов растений, - прочие выделения наземных органов растений, - корневые выделения, - продукты распада мертвых растительных остатков. Среди газообразных выделений большую роль играет этилен, в значительных количествах вырабатываемый некоторыми растениями, например, яблоками. (Этилен задерживает рост, вызывает преждевременный опад листвы, ускоряет распускание почек и созревание плодов, оказывает положительное или отрицательное влияние на рост корней). Газообразные колины могут оказать воздействие на ход сезонных явлений в ценозе, а также подавить развитие тех или иных видов. Однако более или менее значительной роль газообразных колинов может быть лишь в аридных регионах, где обильно число растений, выделяющих различные легко испаряющиеся эфирные масла. Эти эфирные масла служат приспособлением к понижению температуры вокруг испаряющей поверхности, но одновременно могут оказывать определенное воздействие на те или иные растения. Твердые и жидкие выделения надземных органов растений представляют собой минеральные и сложные органические соединения, которые вымываются из надземных частей растений осадками, иногда в весьма значительных количествах, и оказывают действие на другие растения, попадая на них непосредственно с дождем, росой или через почву, куда они вмываются. ПРИМЕРЫ. Выделения полыни горькой (Artemisia absinthium) задерживают рост многих растений, то же указывается для веществ, содержащихся в листьях ореха черного (Juglans nigra), а также в листьях и хвое многих древесных пород и некоторых кустарников и трав. Конкуренция – вслед за Ч. Дарвином в широком смысле – это борьба за существование: борьба за пищу, за место или за какие-либо другие условия. Даже при достаточно высоком сходстве экологических требований, растения одних видов оказываются более сильными, более конкурентомощными при одних определенных значениях факторов среды, другие – при других. Это и служит причиной победы того или другого вида в межвидовой борьбе.
ПРИМЕР. На Крайнем Севере Дальнего Востока береза каменная, ольховый и кедровый стланики формируют чистые сообщества и сообщества с доминированием одного из них на склонах южных экспозиций. Часто они произрастают совместно и доминанта трудно выделить. Все три вида характеризуются очень близкими экологическими свойствами. Все они – реликтовые, и отличаются высоким тепло-, влаго- и светолюбием. Но при этом ольховник несколько более теневынослив и более требователен к влажности почв, береза – более требовательна к теплу и трофности почв, а кедровый стланик – к свету и влажности воздуха. В результате при совместном произрастании кедровостланиковые ценоэлементы, или парцеллы, обычно приурочены к повышенным элементам микрорельефа, более сухим и хорошо дренированным, каменноберезовые – к выровненным участкам с высокой трофностью и тоже хорошо дренированных, ольховниковые – к микропонижениям с очень высоким, но проточным увлажнением и высокой трофностью почв. Каменноберезняки чаще приурочены к распадкам и в горах не поднимаются выше стлаников, кедровый стланик образует чистые заросли на верхней границе леса и на грядах, расположенных полосами вдоль по склону, а заросли ольховника предпочитают седловины и перегибы поверхностей склона в местах с вогнутой поверхностью. Конкуренция отмечается между особями одного вида (внутривидовая борьба) и между особями разных видов (межвидовая борьба) в неблагоприятных условиях среды. Оба типа борьбы обычно тесно связаны друг с другом. В борьбе за пищу, за влагу, с вредителями и паразитами растения конкурируют и с особями своего вида, и с особями других видов. |

stydopedya.ru
Прямые и косвенные взаимодействия между растениями. — КиберПедия
Взаимоотношения между растениями
1. Прямые (контактные)
· Механические(охлестывание ветвями, эпифитизм, давление и сцепление стволов и корней)
· Физиологические (симбиоз, паразитизм и полупаразитизм, срастание корней)
2. Косвенные трансбиотические(через животных и микроорганизмы)
3. Косвенные трансбиотические( средообразующие влияние, конкуренция)
Возникают при соприкосновении или проникновении организмов друг в друга. Подразделяются на физиологические (паразитизм и симбиоз), когда между организмами осуществляется активный обмен веществом и энергией, и механические (взаимоотношения эпифитов с форофитами и лиан с опорными растениями) — когда таковой отсутствует.
Примером механического взаимодействия является повреждение ели и сосны в смешанных лесах от охлестывающего действия березы. Раскачиваясь от ветра, тонкие ветви березы ранят хвою ели, сбивают легкие молодые иглы. Очень заметно это сказывается зимой, когда ветви березы безлиственны. Взаимное давление и сцепление стволов нередко оказывает отрицательное воздействие на растения. Однако чаще такие контакты встречаются в подземной сфере, где большие массы корней тесно переплетаются в небольших объемах почвы. Типы контактов могут быть различны – от простого сцепления до прочного срастания. К форме механических контактов относится и использование в качестве субстрата одним растением другого. Растения, живущие на других растениях (на ветвях, стволах деревьев), без связи с почвой, получили название эпифитов, а поселяющиеся на листьях – эпифиллов. Наиболее богаты эпифитами тропические леса. К ним относятся многие виды бромелиевых, орхидейных .Экологический смысл эпифитизма состоит в своеобразной адаптации к световому режиму в густых тропических лесах : возможность выбраться к свету в верхних ярусах леса без больших затрат веществ на рост.
Физиологические контакты между растениями включают паразитизм, симбиоз, сапрофитизм, срастание корней. Паразитизм – наиболее яркий пример прямых физиологических воздействий между растениями, т. е. переход одного из партнеров на гетеротрофный способ питания и существование за счет организма-хозяина. Характерным примером тесного симбиоза, или мутуализма между растениями, является сожительство водоросли и гриба, организм-лишайник. Другой пример симбиоза – сожительство высших растений с бактериями, так называемаябактериотрофия. Симбиоз с клубеньковыми бактериями-азотофиксаторами широко распространен среди бобовых и мимозовых. Так, бактерии из рода Rhizobium, живущие в клубеньках на корнях бобовых растений, обеспечиваются пищей и местообитанием, а растения получают от них взамен доступную форму азота. Встречается симбиоз мицелия гриба с корнем высшего растения, или микоризообразование. Такие растения называют микотрофными, или микотрофами. Отдельную группу растений с гетеротрофным питанием составляют сапрофиты – виды, которые используют в качестве источника углерода органические вещества отмерших организмов. Встречаются среди цветковых у представителей семейств грушанковых, орхидных и др. Срастание корней близко растущих деревьев (одного и того же вида или родственных видов) относится также к прямым физиологическим контактам между растениями. Установлено, что между сросшимися деревьями существует обмен через корни в виде переноса питательных веществ и воды.
Косвенные трансбиотические взаимоотношения между растениями (через животных и микроогранизмы). Важная экологическая роль животных в жизни растений состоит в участии в процессах опыления, распространения семян и плодов. Опыление растений насекомыми, получившее название энтомофилии, способствовало выработке ряда приспособлений, как у растений, так и насекомых. Разнообразное и сложное строение цветков (различная форма лепестков, симметричное или несимметричное их расположение, наличие определенных соцветий), называемое гетеростилией, - все это приспособления к строению тела и поведению строго конкретных насекомых. Например, цветки дикой моркови, тмина, опыляемые муравьями, цветки копытня, опыляемые муравьями соответственно не поднимающиеся из-под лесной подстилки.
В опылении растений принимают участие и птицы. Опыление растений с помощью птиц, или орнитофилия. Распространение семян, плодов, спор растений при помощи животных называют зоохорией. Среди растений, чьи семена, плоды разносятся животными. В свою очередь, различают эпизоохорные, эндозоохорные и синзоохорные. Съедобные плоды и семена дуба, сосны сибирской животные не поедают сразу, а растаскивают и складывают в запас. Значительная их часть при этом теряется и дает при благоприятных условиях начало новым растениям. Данное распространение семян и плодов получило название синзоохории.
В косвенных трансбиотических взаимоотношениях между растениями нередко выступают микроорганизмы. Ризосфера корней многих деревьев, к примеру, дуба, сильно изменяет почвенную среду, особенно ее состав, кислотность, и тем самым создает благоприятные условия для поселения там различных микроорганизмов, в первую очередь бактерий. Эти бактерии, сразу же сказывается положительно на состоянии растений, особенно молодых.
Косвенные трансбиотические взаимоотношения между растениями (средообразующие влияния, конкуренция, аллелопатия).Изменение растениями среды – это наиболее универсальный и широко распространенный тип взаимоотношений растений при их совместном существовании. Один из них – взаимовлияния через изменения факторов микроклимата. Другой путь взаимодействия растений в сообществах – через напочвенный слой мертвых растительных остатков, называемых на лугах и в степях ветошью, травянистым опадом или «степным войлоком», а в лесу – подстилкой. Этот слой вызывает затруднение для проникновения семян и спор в почву. Существенный путь взаимного влияния растений – это взаимодействие через химические выделения. Растения выделяют в окружающую среду (воздух, воду, почву) разнообразные химические вещества в процессе гуттации, секреции нектара, эфирных масел, смол и т.д.; Химические выделения растений могут служить одним из способов взаимодействия между растениями в сообществе, оказывая на организмы либо токсичное, либо стимулирующее действие. Такие химические взаимовлияния получили название аллелопатия. В качестве примера можно назвать выделения соплодий свеклы, тормозящие прорастание семян куколя. Как крайнюю форму аллелопатии или невозможность существования того или иного вида в присутствии другого в результате интоксикации среды, называют аменсализмом. Растения, наиболее активно и глубоко преобразующие среду и определяющие условия существования для других сообитателей, называют эдификаторами. Различают сильные и слабые эдификаторы. К сильнымэдификаторам относят ель (сильное затенение, обеднение почв питательными веществами и др.), сфагновые мхи (задержание влаги и создание избыточного увлажнения, увеличение кислотности, особый температурный режим и т.д.). Слабыми эдификаторами являются лиственные породы с ажурной кроной, растения травянистого покрова лесов. В качестве особой формы трансбиотических взаимоотношений растений выделяют конкуренцию. Это взаимные или односторонние отрицательные влияния, которые возникают на основе использования энергетических и пищевых ресурсов местообитания. Примером конкуренции могут служить взаимоотношения лисохвоста лугового и типчака.Межвидовая конкуренция проявляется у растений так же, как и внутривидовая (морфологические изменения, снижение плодовитости, численности и т.д.). Доминирующий вид постепенно вытесняет или сильно снижает его жизнеспособность
cyberpedia.su
6.3.Фитогенные факторы
В отечественной литературе наиболее распространена классификация форм взаимоотношений между растениями по В. Н. Сукачеву (табл. 6.2).
Таблица 6.2
Основные формы взаимоотношений между растениями (по в. Н. Сукачеву, н. В. Дылису и др., 1964)
Прямые (контактные) взаимодействия между растениями.
Примером механического взаимодействия является повреждение ели и сосны в смешанных лесах от схлестывающего действия березы. Раскачиваясь от ветра, тонкие ветви березы ранят хвою ели, сбивают легкие молодые иглы. Очень заметно это сказывается зимой, когда ветви березы безлиственны.
Взаимное давление и сцепление стволов нередко оказывает отрицательное воздействие на растения. Однако чаще такие контакты встречаются в подземной сфере, где большие массы корней тесно переплетаются в небольших объемах почвы. Типы контактов могут быть различны — от простого сцепления до прочного срастания. Так, губительным в жизни многих деревьев тропического леса оказывается разрастание лиан, зачастую приводящее к обламыванию ветвей под их тяжестью и усыханию стволов в результате сдавливающего действия вьющимися стеблями или корнями. Не случайно некоторые лианы называют «душителями» (рис. 6.8).
К форме механических контактов относится и использование в качестве субстрата одним растением другого. Растения, живущие на других растениях (на ветвях, стволах деревьев), без связи с почвой, получили название эпифитов, а поселяющиеся на листьях — эпифиллов. В отличие от паразитов они не вступают в прямой физиологический контакт с растением-субстратом, а самостоятельно существуют как автотрофные организмы.
Рис. 6.8. Растения-лианы:
1 — фикус-душитель; 2 — повилика;
3 — жимолость вьющаяся (по Н. М. Черновой и др., 1995)
По мнению ученых, около 10% всех видов растений веду эпифитный образ жизни. Наиболее богаты эпифитами тропические леса. К ним относятся многие виды бромелиевых, орхидейных (рис. 6.9).
Рис. 6.9. Эпифитная орхидея с воздушными корнями:
А — общий вид; Б — поперечный срез воздушного корня с наружным слоем всасывающей ткани (1) (по В. Л. Комарову, 1949)
Экологический смысл эпифитизма состоит в своеобразной адаптации к световому режиму в густых тропических лесах: возможность выбраться к свету в верхних ярусах леса без больших затрат веществ на рост. Само происхождение эпифитного образа жизни связывают с борьбой растений за свет. Эволюция многих эпифитов зашла так далеко, что они уже потеряли способность расти вне растительного субстрата, т. е. являются облигатными эпифитами. Вместе с тем есть виды, которые в оранжерейных условиях могут расти и в почве.
Физиологические контакты между растениями включают паразитизм, симбиоз, сапрофитизм, срастание корней. Паразитизм — наиболее яркий пример прямых физиологических воздействий между растениями, т.е. переход одного из партнеров на гетеротрофный способ питания и существование за счет организма-хозяина. Например, повилика, питающаяся соками клевера, угнетает его, не только подавляя развитие вегетативной массы, но и заметно задерживая развитие растения. Как правило, урожай семян пораженного клевера невысокий. Через несколько лет на лугу, где появилась повилика, клевер полностью выпадает из травостоя. Паразиты, как уже было отмечено ранее, многочисленны среди грибов и бактерий, значительно меньше распространены среди цветковых растений.
Характерным примером тесного симбиоза, или мутуализма между растениями является сожительство водоросли и гриба, которые образуют особый целостный организм-лишайник (рис. 6.10).
Рис. 6.10. Лишайник-кладония (по Н. М. Черновой и др., 1995)
Связи между партнерами-симбионтами довольно сложны, и некоторые авторы называют симбиоз «хорошо урегулированным взаимным паразитизмом». Фотосинтезирующие водоросли в лишайнике снабжают гриб углеводами и другими органическими веществами (нуклеиновые кислоты, протеины, активаторы роста и др.). Грибы поставляют водоросли влагу и минеральные вещества. При недостатке же света и других неблагоприятных условий, угнетающих фотосинтез, водоросли могут получать от грибов и органические вещества, которые они поглощают из субстрата. Лишайники интересны тем, что на их примере можно наблюдать постепенную эволюцию от паразитизма к мутуализму. У более примитивных лишайников гриб фактически проникает в клетки водоросли и по существу является паразитом (рис. 6.11А). У эволюционно более развитых видов водоросль и гриб живут в гармоничных отношениях, принося друг другу взаимную пользу (рис. 6.11Б и В).
Рис. 6.11. Направление эволюции от паразитизма к мутуализму
у лишайников (по Е. Одуму, 1963)
Другой пример симбиоза — это сожительство высших растений с бактериями, так называемая бактериотрофия. Симбиоз с клубеньковыми бактериями-азотофиксаторами широко распространен среди бобовых (93% изученных видов) и мимозовых (87%). Так, бактерии из рода Rhizobium, живущие в клубеньках на корнях бобовых растений, обеспечиваются пищей (сахара) и ме-стообиганием, а растения получают от них взамен доступную форму азота (рис. 6.12).
Рис. 6.12. Клубеньки на корнях бобовых растений:
А — клевера красного; Б — фасоли; В — сои; Г — люпина (по А. П. Шенникову, 1950)
Встречается симбиоз мицелия гриба с корнем высшего растения, или микоризообразование. Такие растения называют микотрофными, или микотрофами. Поселясь на корнях растений, гифы гриба обеспечивают высшее растение колоссальной всасывающей способностью. Поверхность соприкосновения клеток корня и гиф в эктотрофной микоризе в 10—14 раз больше, чем поверхность контакта с почвой клеток «голого» корня, тогда как всасывающая поверхность корня за счет корневых волосков увеличивает поверхность корня лишь в 2—5 раз. Из изученных в нашей стране 3425 видов сосудистых растений микориза обнаружена у 79%.
В качестве примера симбиоза грибов с насекомыми можно привести симбиоз грибка Septobasidium с насекомым-червецом из сем. Coccidae, дающее новое симбиотическое образование — лаки, которое как единый организм введено в культуру человеком.
Отдельную группу растений с гетеротрофным питанием составляют сапрофиты — виды, которые используют в качестве источника углерода органические вещества отмерших организмов. В биологическом круговороте это важное звено, осуществляющее разложение органических остатков и перевод сложных соединений в более простые, представлено большей частью грибами, актиномицетами, бактериями. Встречаются среди цветковых у представителей семейств грушанковых, орхидных и др. Примерами цветковых, полностью утративших хлорофилл и перешедших на питание готовыми органическими веществами, являются сапрофиты хвойных лесов — подъельник обыкновенный (Monotropa hypopitis), надбородник безлистный (Epipogon aphylluon). Среди мхов и папоротников сапрофиты редки.
Срастание корней близко растущих деревьев (одного и того же вида или родственных видов) относится также к прямым физиологическим контактам между растениями. Явление не столь , уж редкое в природе. В густых насаждениях ели Picea flies срастаются корнями около 30% всех деревьев. Установлено, что между сросшимися деревьями существует обмен через корни в виде переноса питательных веществ и воды. В зависимости от степени различия или сходства потребностей сросшихся партнеров между ними не исключены отношения как конкурентного характера в виде перехвата веществ более развитым и сильным деревом, так и симбиотические.
Определенное значение имеет форма связей в виде хищничества. Хищничество широко распространено не только между животными, но и между растениями и животными. Так, ряд насекомоядных растений (росянка, непентес) относят к хищникам (рис. 6.13).
Рис. 6.13. Хищное растение росянка (по Е. А. Криксунову и др., 1995)
Косвенные трансбиотические взаимоотношения между растениями (через животных и микроорганизмы). Важная экологическая роль животных в жизни растений состоит в участии в процессах опыления, распространения С9мян и плодов. Опыление растений насекомыми, получившее название энтомофилии, способствовало выработке ряда приспособлений как у растений, так и насекомых. Назовем здесь такие интересные адаптации энтомофильных цветков:] узоры, образующие «путевые нити» к нектарникам и тычинкам, нередко видимые только в ультрафиолетовых лучах, доступных для зрения насекомых; различие окраски цветков до и после опыления; синхронизация суточных ритмов раскрывания венчика и тычинок, обеспечивающие безошибочное попадание пь1пьцы на тело насекомого, а с него — на рыльце другого цветка и т. д. (рис. 6.14).
Рис. 6.14. Насекомое на
цветке (по Н. М. Черно-
вой и др., 1995)
Разнообразное и сложное строение цветков (различная форма лепестков, симметричное или несимметричное их расположение, наличие определенных соцветий), называемое гетеростилией, — все это приспособления к строению тела и поведению строго конкретных насекомых. Например, цветки дикой моркови (Daucus carota), тмина (Carum carvi), опыляемые муравьями, цветки копытня Asarum europaeum, опыляемые муравьями и соответственно не поднимающиеся из-под лесной подстилки.В опылении растений принимают участие и птицы. Опыление растений с помощью птиц, или орнитофилия, находит широкое распространение в тропических и субтропических областях южного полушария. Здесь известно около 2000 видов птиц, которые опыляют цветки при поисках нектара или ловле насекомых, прячущихся в их венчиках. Среди них наиболее известные опылите-ли-нектарницы (Африка, Австралия, Южная Азия) и колибри (Южная Америка). Цветки орнитофильных растений крупные, ярко окрашенные. Преобладает ярко-красная окраска, наиболее привлекательная для колибри и других птиц. В некоторых орнитофильных цветках существуют специальные защитные устройства, которые не дают нектару вылиться при движении цветка.
Реже встречается опыление растений млекопитающими, или зоо-гамия. Большей частью зоогамия отмечается в Австралии, в лесах Африки и Южной Америки. Например, австралийские кустарники из рода Driandra опыляются с помощью кенгуру, охотно пьющих их обильный нектар, переходя от цветка к цветку.
Распространение семян, плодов, спор растений при помощи животных называют зоохорией. Среди растений, чьи семена, плоды разносятся животными, в свою очередь, различают эпизоо-хорные, эндозоохорные и синзоохорные. Эпизоохорные растения большей частью открытых мест обитания имеют у семян, плодов всевозможные приспособления для закрепления и удерживания на поверхности тела животных (выросты, крючки, прицепки и др.), например лопухи большой и паутинистый, липучка обыкновенная и т. д.
В кустарниковом ярусе лесов, где обитает много птиц, преобладают эндозоохорные виды растений. Их плоды съедобны или привлекательны для птиц яркой окраской или сочным околоплодником. Следует отметить, что у семян многих эндозоохорных растений повышается всхожесть, а иногда и способность к прорастанию только после прохождения через пищевой тракт животного — многие аралиевные, яблоня Сиверса (Malus sieversu) и др.
Съедобные плоды исемена дуба, сосны сибирской животные не поедают сразу, а растаскивают и складывают в запас. Значительная их часть при этом теряется и дает при благоприятных условиях начало новым растениям. Данное распространение семян и плодов получило название синзоохории.
В косвенных трансбиотических взаимоотношениях эдежду растениями нередко выступают микроорганизмы. Ризосфера корней многих деревьев, к примеру, дуба, сильно изменяет почвенную среду, особенно ее состав, кислотность, и тем самым создает благоприятные условия для поселения там различных микроорганизмов, в первую очередь бактерий, таких, как Azotobacter chroocoteum, Tricholome legnorum, Pseudomonas sp. Эти бактерии, поселившись здесь, питаются выделениями корней дуба и органическими остатками, создаваемыми гифами микоризообразующих грибов. Бактерии, живя рядом с корнями дуба, служат своеобразной «оборонительной линией» от проникновения в корни патогенных грибов. Этот биологический барьер создается при помощи антибиотиков, выделяемых бактериями. Поселение бактерий в ризосфере дуба сразу же сказывается положительно на состоянии растений, особенно молодых.
Косвенные трансабиотические взаимоотношения между растениями (средообразующие влияния, конкуренция, аллелопатия). Изменение растениями среды — это наиболее универсальный и широко распространенный тип взаимоотношений растений при их совместном существовании. Когда тот или иной вид или группа видов растений в Ц результате своей жизнедеятельности сильно изменяет в количественном и качественном отношении основные экологические факторы таким образом, что другим видам сообщества приходится жить в условиях, которые значительно отличаются от зонального комплекса факто- lsров физической среды, то это говорит о средообразующей роли, средообразующем влиянии первого вида по отношению к остальным. Один из них — взаимовлияния через изменения факторов микроклимата (например, ослабление солнечной радиации внутри растительного покрова, обеднение ее фотосинтетически активными лучами, изменение сезонного ритма освещенности и др.). Одни растения влияют на другие и через изменение температурного режима воздуха, его влажности, скорости ветра, содержания углекислоты и т. д.
Другой путь взаимодействия растений в сообществах — через напочвенный слой мертвых растительных остатков, называемых на лугах и в степях ветошью, травянистым спадом или «степным войлоком», а в лесу — подстилкой. Этот слой (иногда толщиной в несколько сантиметров) вызывает затруднение для проникновения семян и спор в почву. Прорастающие в слое ветоши (или на нем) семена часто гибнут от высыхания раньше, чем корни проростков достигнут почвы. Для семян, попавших в почву и прорастающих, напочвенные остатки могут являться серьезным механическим препятствием на пути ростков к свету. Возможны и взаимоотношения растений через содержащиеся в подстилке продукты распада растительных остатков, тормозящих или, напротив, стимулирующих рост растений. Так, в свежем опаде ели или бука содержатся вещества, тормозящие прорастание ели и сосны, а в местах со скудными осадками и слабым промыванием подстилки могут угнетать естественное возобновление древесных пород. Водные вытяжки из лесных подстилок отрицательно действуют и на рост многих степных трав.
Существенный путь взаимного влияния растений — это взаимодействие через химические выделения. Растения выделяют в окружающую среду (воздух, воду, почву) разнообразные химические вещества в процессе гуттации, секреции нектара, эфирных масел, смол и т. д.; при вымывании минеральных солей дождевыми водами листья, например, деревьев, теряют калий, натрий, магний и другие ионы; в ходе метаболизма (корневые выделения) газообразные вещества, выделяемые надземными органами, — непредельные углеводороды, этилен, водород и др.; при нарушении целостности тканей и органов растения выделяют летучие вещества, так называемые фитонциды, и вещества из отмерших частей растений (рис. 6.15).
Выделяемые соединения необходимы растениям, но с развитием большой поверхности тела растений их потеря столь же неизбежна, как и транспирация.
Химические выделения растений могут служить одним из способов взаимодействия между растениями в сообществе, оказывая на организмы либо токсичное, либо стимулирующее действие.
Рис. 6.15. Влияние одного растения на другое
(по А. М. Гродзинскому, 1965):
1 — миазмины; 2 — фитонцидные вещества; 3 — фитогенные вещества; 4 — активные прижизненные выделения; 5 — пассивные прижизненные выделения; 6 — посмертные выделения; 7 — переработка гетеротрофными организмами
Такие химические взаимовлияния получили название аллелопатии. В качестве примера можно назвать выделения соплодий свеклы, тормозящие прорастание семян куколя (Agrostemma githago). Нут (Cicer arietinum) подавляюще действует на картофель, кукурузу, подсолнечник, томаты и другие культуры, фасоль — на рост яровой пшеницы; корневые выделения пырея (Agropyron repens) и костреца (Bromus inermis) — на растущие вблизи с ними другие травянистые растения и даже деревья. Как крайнюю форму аллелопатии или невозможность существования того или иного вида в присутствии другого в результате интоксикации среды, называют аменсализмом. Аменсализм соответствует прямой конкуренции, антибиозу и антагонизму. Так, благодаря выделению корнями токсических веществ ястребянка (Hieracium pilosella) из семейства сложноцветных вытесняет другие однолетние растения и нередко образует чистые заросли на довольно больших площадях. Многие грибы и бактерии синтезируют антибиотики, которые тормозят рост других бактерий. Аменсализм широко распространен в водной среде.
У разных видов растений степень воздействия на среду и таким образом на жизнь обитателей неодинакова в соответствии с особенностями их морфологии, биологии, сезонного развития и др. Растения, наиболее активно и глубоко преобразующие среду и определяющие условия существования для других сооби-тателей, называют эдификаторами. Различают сильные и слабые эдификаторы. К сильным эдификаторам относят ель (сильное затенение, обеднение почв питательными веществами и др.), сфагновые мхи (задержание влаги и создание избыточного увлажнения, увеличение кислотности, особый температурный режим и т. д.). Слабыми эдификаторами являются лиственные породы с ажурной кроной (береза, ясень), растения травянистого покрова лесов.
В качестве особой формы трансбиотических взаимоотношений растений выделяют конкуренцию. Это те взаимные или односторонние отрицательные влияния, которые возникают на основе использования энергетических и пищевых ресурсов мес-тообитания. Сильное влияние на жизнь растений оказывает конкуренция за почвенную влагу (особенно четко выражена в областях с недостаточным увлажнением) и конкуренция за питательные вещества почвы, более заметная на бедных почвах. Примером конкуренции могут служить взаимоотношения лисохвоста лугового (Alopecurus pratensis) и типчака (Festuca sulcata). Типчак может произрастать во влажной почве, однако в сообществе лисохвостного луга не растет из-за подавления теневыносливым и быстро растущим лисохвостом. В формировании типчакового или лисохвостного фитоценоза решающее значение имеет не влажность почвы, а конкурентные взаимоотношения типчака и лисохвоста. В более сухих местообитаниях типчак заглушает лисохвост, а на увлажненных лугах выходит победителем лисохвост.
Межвидовая конкуренция проявляется у растений так же, как и внутривидовая (морфологические изменения, снижение плодовитости, численности и т. д.). Доминирующий вид постепенно вытесняет или сильно снижает его жизнеспособность.
Самая жесткая конкуренция, нередко с непредвиденными последствиями, возникает при введении в сообщества новых видов растений без учета уже сложившихся отношений.
studfiles.net
|
ТОП 10: |
Примером механического взаимодействия является повреждение ели и сосны в смешанных лесах от схлестывающего действия березы. Раскачиваясь от ветра, тонкие ветви березы ранят хвоюели, сбивают легкие молодые иглы. Очень заметно это сказывается зимой, когда ветви березы безлиственны. Взаимное давление и сцепление стволов нередко оказывает отрицательное воздействие на растения. Однако чаще такие контакты встречаются в подземной сфере, где большие массы корней тесно переплетаются в небольших объемах почвы. Типы контактов могут быть различны — от простого сцепления до прочного срастания. Так, губительным в жизни многих деревьев тропического леса оказывается разрастание лиан, зачастую приводящее к обламыванию ветвей под их тяжестью и усыханию стволов в результате сдавливающего действия вьющимися стеблями или корнями. Не случайно некоторые лианы называют «душителями» (рис. 6.8). К форме механических контактов относится и использование в качестве субстрата одним растением другого. Растения, живущие на других растениях (на ветвях, стволах деревьев), без связи с почвой, получили название эпифитов, а поселяющиеся на листьях — эпифиллов. В отличие от паразитов они не вступают в прямой физиологический контакт с растением-субстратом, а самостоятельно существуют как автотрофные организмы.
Рис. 6.8. Растения-лианы: 1 — фикус-душитель; 2 — повилика; 3 — жимолость вьющаяся (по Н. М. Черновой и др., 1995)
По мнению ученых, около 10% всех видов растений веду эпифитный образ жизни. Наиболее богаты эпифитами тропические леса. К ним относятся многие виды бромелиевых, орхидейных (рис. 6.9).
Рис. 6.9. Эпифитная орхидея с воздушными корнями: А — общий вид; Б — поперечный срез воздушного корня с наружным слоем всасывающей ткани (1) (по В. Л. Комарову, 1949)
Экологический смысл эпифитизма состоит в своеобразной адаптации к световому режиму в густых тропических лесах: возможность выбраться к свету в верхних ярусах леса без больших затрат веществ на рост. Само происхождение эпифитного образа жизни связывают с борьбой растений за свет. Эволюция многих эпифитов зашла так далеко, что они уже потеряли способность расти вне растительного субстрата, т. е. являются облигатными эпифитами. Вместе с тем есть виды, которые в оранжерейных условиях могут расти и в почве. Физиологические контакты между растениями включают паразитизм, симбиоз, сапрофитизм, срастание корней. Паразитизм — наиболее яркий пример прямых физиологических воздействий между растениями, т.е. переход одного из партнеров на гетеротрофный способ питания и существование за счет организма-хозяина. Например, повилика, питающаяся соками клевера, угнетает его, не только подавляя развитие вегетативной массы, но и заметно задерживая развитие растения. Как правило, урожай семян пораженного клевера невысокий. Через несколько лет на лугу, где появилась повилика, клевер полностью выпадает из травостоя. Паразиты, как уже было отмечено ранее, многочисленны среди грибов и бактерий, значительно меньше распространены среди цветковых растений. Характерным примером тесного симбиоза, или мутуализма между растениями является сожительство водоросли и гриба, которые образуют особый целостный организм-лишайник (рис. 6.10).
Рис. 6.10. Лишайник-кладония (по Н. М. Черновой и др., 1995)
Связи между партнерами-симбионтами довольно сложны, и некоторые авторы называют симбиоз «хорошо урегулированным взаимным паразитизмом». Фотосинтезирующие водоросли в лишайнике снабжают гриб углеводами и другими органическими веществами (нуклеиновые кислоты, протеины, активаторы роста и др.). Грибы поставляют водоросли влагу и минеральные вещества. При недостатке же света и других неблагоприятных условий, угнетающих фотосинтез, водоросли могут получать от грибов и органические вещества, которые они поглощают из субстрата. Лишайники интересны тем, что на их примере можно наблюдать постепенную эволюцию от паразитизма к мутуализму. У более примитивных лишайников гриб фактически проникает в клетки водоросли и по существу является паразитом (рис. 6.11А). У эволюционно более развитых видов водоросль и гриб живут в гармоничных отношениях, принося друг другу взаимную пользу (рис. 6.11Б и В).
Рис. 6.11. Направление эволюции от паразитизма к мутуализму у лишайников (по Е. Одуму, 1963)
Другой пример симбиоза — это сожительство высших растений с бактериями, так называемая бактериотрофия. Симбиоз с клубеньковыми бактериями-азотофиксаторами широко распространен среди бобовых (93% изученных видов) и мимозовых (87%). Так, бактерии из рода Rhizobium, живущие в клубеньках на корнях бобовых растений, обеспечиваются пищей (сахара) и ме-стообиганием, а растения получают от них взамен доступную форму азота (рис. 6.12).
Рис. 6.12. Клубеньки на корнях бобовых растений: А — клевера красного; Б — фасоли; В — сои; Г — люпина (по А. П. Шенникову, 1950)
Встречается симбиоз мицелия гриба с корнем высшего растения, или микоризообразование. Такие растения называют микотрофными, или микотрофами. Поселясь на корнях растений, гифы гриба обеспечивают высшее растение колоссальной всасывающей способностью. Поверхность соприкосновения клеток корня и гиф в эктотрофной микоризе в 10—14 раз больше, чем поверхность контакта с почвой клеток «голого» корня, тогда как всасывающая поверхность корня за счет корневых волосков увеличивает поверхность корня лишь в 2—5 раз. Из изученных в нашей стране 3425 видов сосудистых растений микориза обнаружена у 79%. В качестве примера симбиоза грибов с насекомыми можно привести симбиоз грибка Septobasidium с насекомым-червецом из сем. Coccidae, дающее новое симбиотическое образование — лаки, которое как единый организм введено в культуру человеком. Отдельную группу растений с гетеротрофным питанием составляют сапрофиты — виды, которые используют в качестве источника углерода органические вещества отмерших организмов. В биологическом круговороте это важное звено, осуществляющее разложение органических остатков и перевод сложных соединений в более простые, представлено большей частью грибами, актиномицетами, бактериями. Встречаются среди цветковых у представителей семейств грушанковых, орхидных и др. Примерами цветковых, полностью утративших хлорофилл и перешедших на питание готовыми органическими веществами, являются сапрофиты хвойных лесов — подъельник обыкновенный (Monotropa hypopitis), надбородник безлистный (Epipogon aphylluon). Среди мхов и папоротников сапрофиты редки. Срастание корней близко растущих деревьев (одного и того же вида или родственных видов) относится также к прямым физиологическим контактам между растениями. Явление не столь , уж редкое в природе. В густых насаждениях ели Picea flies срастаются корнями около 30% всех деревьев. Установлено, что между сросшимися деревьями существует обмен через корни в виде переноса питательных веществ и воды. В зависимости от степени различия или сходства потребностей сросшихся партнеров между ними не исключены отношения как конкурентного характера в виде перехвата веществ более развитым и сильным деревом, так и симбиотические.
Определенное значение имеет форма связей в виде хищничества. Хищничество широко распространено не только между животными, но и между растениями и животными. Так, ряд насекомоядных растений (росянка, непентес) относят к хищникам (рис. 6.13).
Рис. 6.13. Хищное растение росянка (по Е. А. Криксунову и др., 1995) Косвенные трансбиотические взаимоотношения между растениями (через животных и микроорганизмы). Важная экологическая роль животных в жизни растений состоит в участии в процессах опыления, распространения С9мян и плодов. Опыление растений насекомыми, получившее название энтомофилии, способствовало выработке ряда приспособлений как у растений, так и насекомых. Назовем здесь такие интересные адаптации энтомофильных цветков:] узоры, образующие «путевые нити» к нектарникам и тычинкам,
Разнообразное и сложное строение цветков (различная форма лепестков, симметричное или несимметричное их расположение, наличие определенных соцветий), называемое гетеростилией, — все это приспособления к строению тела и поведению строго конкретных насекомых. Например, цветки дикой моркови (Daucus carota), тмина (Carum carvi), опыляемые муравьями, цветки копытня Asarum europaeum, опыляемые муравьями и соответственно не поднимающиеся из-под лесной подстилки. В опылении растений принимают участие и птицы. Опыление растений с помощью птиц, или орнитофилия, находит широкое распространение в тропических и субтропических областях южного полушария. Здесь известно около 2000 видов птиц, которые опыляют цветки при поисках нектара или ловле насекомых, прячущихся в их венчиках. Среди них наиболее известные опылите-ли-нектарницы (Африка, Австралия, Южная Азия) и колибри (Южная Америка). Цветки орнитофильных растений крупные, ярко окрашенные. Преобладает ярко-красная окраска, наиболее привлекательная для колибри и других птиц. В некоторых орнитофильных цветках существуют специальные защитные устройства, которые не дают нектару вылиться при движении цветка. Реже встречается опыление растений млекопитающими, или зоо-гамия. Большей частью зоогамия отмечается в Австралии, в лесах Африки и Южной Америки. Например, австралийские кустарники из рода Driandra опыляются с помощью кенгуру, охотно пьющих их обильный нектар, переходя от цветка к цветку. Распространение семян, плодов, спор растений при помощи животных называют зоохорией. Среди растений, чьи семена, плоды разносятся животными, в свою очередь, различают эпизоо-хорные, эндозоохорные и синзоохорные. Эпизоохорные растения большей частью открытых мест обитания имеют у семян, плодов всевозможные приспособления для закрепления и удерживания на поверхности тела животных (выросты, крючки, прицепки и др.), например лопухи большой и паутинистый, липучка обыкновенная и т. д. В кустарниковом ярусе лесов, где обитает много птиц, преобладают эндозоохорные виды растений. Их плоды съедобны или привлекательны для птиц яркой окраской или сочным околоплодником. Следует отметить, что у семян многих эндозоохорных растений повышается всхожесть, а иногда и способность к прорастанию только после прохождения через пищевой тракт животного — многие аралиевные, яблоня Сиверса (Malus sieversu) и др. Съедобные плоды исемена дуба, сосны сибирской животные не поедают сразу, а растаскивают и складывают в запас. Значительная их часть при этом теряется и дает при благоприятных условиях начало новым растениям. Данное распространение семян и плодов получило название синзоохории. В косвенных трансбиотических взаимоотношениях эдежду растениями нередко выступают микроорганизмы. Ризосфера корней многих деревьев, к примеру, дуба, сильно изменяет почвенную среду, особенно ее состав, кислотность, и тем самым создает благоприятные условия для поселения там различных микроорганизмов, в первую очередь бактерий, таких, как Azotobacter chroocoteum, Tricholome legnorum, Pseudomonas sp. Эти бактерии, поселившись здесь, питаются выделениями корней дуба и органическими остатками, создаваемыми гифами микоризообразующих грибов. Бактерии, живя рядом с корнями дуба, служат своеобразной «оборонительной линией» от проникновения в корни патогенных грибов. Этот биологический барьер создается при помощи антибиотиков, выделяемых бактериями. Поселение бактерий в ризосфере дуба сразу же сказывается положительно на состоянии растений, особенно молодых. Косвенные трансабиотические взаимоотношения между растениями (средообразующие влияния, конкуренция, аллелопатия). Изменение растениями среды — это наиболее универсальный и широко распространенный тип взаимоотношений растений при их совместном существовании. Когда тот или иной вид или группа видов растений в Ц результате своей жизнедеятельности сильно изменяет в количественном и качественном отношении основные экологические факторы таким образом, что другим видам сообщества приходится жить в условиях, которые значительно отличаются от зонального комплекса факто- lsров физической среды, то это говорит о средообразующей роли, средообразующем влиянии первого вида по отношению к остальным. Один из них — взаимовлияния через изменения факторов микроклимата (например, ослабление солнечной радиации внутри растительного покрова, обеднение ее фотосинтетически активными лучами, изменение сезонного ритма освещенности и др.). Одни растения влияют на другие и через изменение температурного режима воздуха, его влажности, скорости ветра, содержания углекислоты и т. д. Другой путь взаимодействия растений в сообществах — через напочвенный слой мертвых растительных остатков, называемых на лугах и в степях ветошью, травянистым спадом или «степным войлоком», а в лесу — подстилкой. Этот слой (иногда толщиной в несколько сантиметров) вызывает затруднение для проникновения семян и спор в почву. Прорастающие в слое ветоши (или на нем) семена часто гибнут от высыхания раньше, чем корни проростков достигнут почвы. Для семян, попавших в почву и прорастающих, напочвенные остатки могут являться серьезным механическим препятствием на пути ростков к свету. Возможны и взаимоотношения растений через содержащиеся в подстилке продукты распада растительных остатков, тормозящих или, напротив, стимулирующих рост растений. Так, в свежем опаде ели или бука содержатся вещества, тормозящие прорастание ели и сосны, а в местах со скудными осадками и слабым промыванием подстилки могут угнетать естественное возобновление древесных пород. Водные вытяжки из лесных подстилок отрицательно действуют и на рост многих степных трав. Существенный путь взаимного влияния растений — это взаимодействие через химические выделения. Растения выделяют в окружающую среду (воздух, воду, почву) разнообразные химические вещества в процессе гуттации, секреции нектара, эфирных масел, смол и т. д.; при вымывании минеральных солей дождевыми водами листья, например, деревьев, теряют калий, натрий, магний и другие ионы; в ходе метаболизма (корневые выделения) газообразные вещества, выделяемые надземными органами, — непредельные углеводороды, этилен, водород и др.; при нарушении целостности тканей и органов растения выделяют летучие вещества, так называемые фитонциды, и вещества из отмерших частей растений (рис. 6.15). Выделяемые соединения необходимы растениям, но с развитием большой поверхности тела растений их потеря столь же неизбежна, как и транспирация. Химические выделения растений могут служить одним из способов взаимодействия между растениями в сообществе, оказывая на организмы либо токсичное, либо стимулирующее действие.
Рис. 6.15. Влияние одного растения на другое (по А. М. Гродзинскому, 1965): 1 — миазмины; 2 — фитонцидные вещества; 3 — фитогенные вещества; 4 — активные прижизненные выделения; 5 — пассивные прижизненные выделения; 6 — посмертные выделения; 7 — переработка гетеротрофными организмами
Такие химические взаимовлияния получили название аллелопатии. В качестве примера можно назвать выделения соплодий свеклы, тормозящие прорастание семян куколя (Agrostemma githago). Нут (Cicer arietinum) подавляюще действует на картофель, кукурузу, подсолнечник, томаты и другие культуры, фасоль — на рост яровой пшеницы; корневые выделения пырея (Agropyron repens) и костреца (Bromus inermis) — на растущие вблизи с ними другие травянистые растения и даже деревья. Как крайнюю форму аллелопатии или невозможность существования того или иного вида в присутствии другого в результате интоксикации среды, называют аменсализмом. Аменсализм соответствует прямой конкуренции, антибиозу и антагонизму. Так, благодаря выделению корнями токсических веществ ястребянка (Hieracium pilosella) из семейства сложноцветных вытесняет другие однолетние растения и нередко образует чистые заросли на довольно больших площадях. Многие грибы и бактерии синтезируют антибиотики, которые тормозят рост других бактерий. Аменсализм широко распространен в водной среде. У разных видов растений степень воздействия на среду и таким образом на жизнь обитателей неодинакова в соответствии с особенностями их морфологии, биологии, сезонного развития и др. Растения, наиболее активно и глубоко преобразующие среду и определяющие условия существования для других сооби-тателей, называют эдификаторами. Различают сильные и слабые эдификаторы. К сильным эдификаторам относят ель (сильное затенение, обеднение почв питательными веществами и др.), сфагновые мхи (задержание влаги и создание избыточного увлажнения, увеличение кислотности, особый температурный режим и т. д.). Слабыми эдификаторами являются лиственные породы с ажурной кроной (береза, ясень), растения травянистого покрова лесов. В качестве особой формы трансбиотических взаимоотношений растений выделяют конкуренцию. Это те взаимные или односторонние отрицательные влияния, которые возникают на основе использования энергетических и пищевых ресурсов мес-тообитания. Сильное влияние на жизнь растений оказывает конкуренция за почвенную влагу (особенно четко выражена в областях с недостаточным увлажнением) и конкуренция за питательные вещества почвы, более заметная на бедных почвах. Примером конкуренции могут служить взаимоотношения лисохвоста лугового (Alopecurus pratensis) и типчака (Festuca sulcata). Типчак может произрастать во влажной почве, однако в сообществе лисохвостного луга не растет из-за подавления теневыносливым и быстро растущим лисохвостом. В формировании типчакового или лисохвостного фитоценоза решающее значение имеет не влажность почвы, а конкурентные взаимоотношения типчака и лисохвоста. В более сухих местообитаниях типчак заглушает лисохвост, а на увлажненных лугах выходит победителем лисохвост. Межвидовая конкуренция проявляется у растений так же, как и внутривидовая (морфологические изменения, снижение плодовитости, численности и т. д.). Доминирующий вид постепенно вытесняет или сильно снижает его жизнеспособность. Самая жесткая конкуренция, нередко с непредвиденными последствиями, возникает при введении в сообщества новых видов растений без учета уже сложившихся отношений. Антропогенные факторы
Действие человека как экологического фактора в природе огромно и чрезвычайно многообразно. В настоящее время ни один из экологических факторов не оказывает столь существенного и всеобщего, т. е. планетарного, влияния, как человек, хотя это наиболее молодой фактор из всех действующих на природу. Влияние антропогенного фактора постепенно усиливалось, начиная от эпохи собирательства (где оно мало чем отличалось от влияния животных) до наших дней, эпохи научно-технического прогресса и демографического взрыва. В процессе своей деятельности человек создал большое количество самых разнообразных видов животных и растений, существенным образом преобразовывал естественные природные комплексы. На значительных территориях создал особые, нередко практически оптимальные условия жизни многим видам. Создавая огромное разнообразие сортов и видов растений и животных, человек способствовал появлению у них новых свойств и качеств, обеспечивающих им выживание в неблагоприятных условиях, как в борьбе за существование с другими видами, так и невосприимчивости к воздействию патогенных организмов. Изменения, производимые человеком в природной среде, создают для одних видов благоприятные условия для размножения и развития, для других — неблагоприятные. И как результат, между видами создаются новые численные отношения, перестраиваются пищевые цепи, возникают приспособления, необходимые для существования организмов в измененной среде. Таким образом, действия человека обогащают или обедняют сообщества. Влияние ант-ропогенного фактора в природе может быть как сознательным, так и случайным, или неосознанным. Человек, распахивая целинные и залежные земли, создает сельскохозяйственные угодья (аг-роценозы), выводит высокопродуктивные и устойчивые к заболеваниям формы, расселяет одних и уничтожает других. Эти воздействия часто являются положительными, но нередко носят отрицательный характер, например, необдуманное расселение многих животных, растений, микроорганизмов, хищническое уничтожение целого ряда видов, загрязнение среды и др. К случайным относятся воздействия, происходящие в природе под влиянием человеческой деятельности, но не были заранее предусмотрены и запланированы им, и таких примеров немало: распространение различных вредителей, паразитов, случайный завоз различных организмов с грузом, непредвиденные последствия, вызванные сознательными действиями в природе, например нежелательные явления, вызванные осушением болот, постройкой плотин, распашкой целины и др. Человек может оказывать на животных и растительный покров Земли как прямое влияние, так и косвенное. Разнообразие современных форм воздействия человека на растительность представлено в табл. 6.3. Таблица 6.3 |

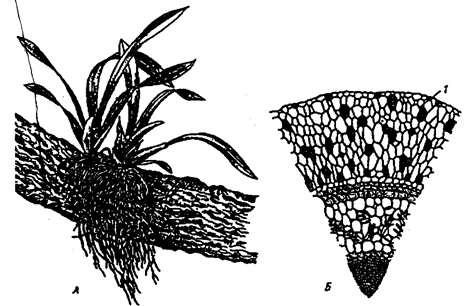

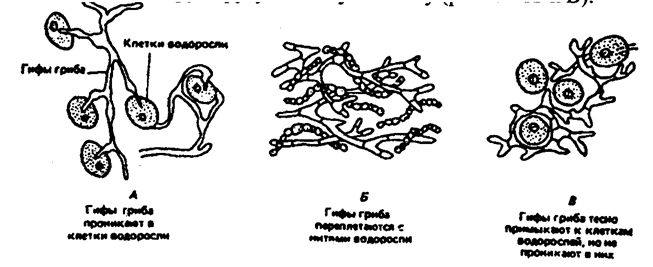
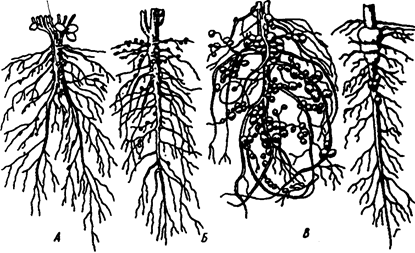

 нередко видимые только в ультрафиолетовых лучах, доступных для зрения насекомых; различие окраски цветков до и после опыления; синхронизация суточных ритмов раскрывания венчика и тычинок, обеспечивающие безошибочное попадание пь1пьцы на тело насекомого, а с него — на рыльце другого цветка и т. д. (рис. 6.14).
нередко видимые только в ультрафиолетовых лучах, доступных для зрения насекомых; различие окраски цветков до и после опыления; синхронизация суточных ритмов раскрывания венчика и тычинок, обеспечивающие безошибочное попадание пь1пьцы на тело насекомого, а с него — на рыльце другого цветка и т. д. (рис. 6.14).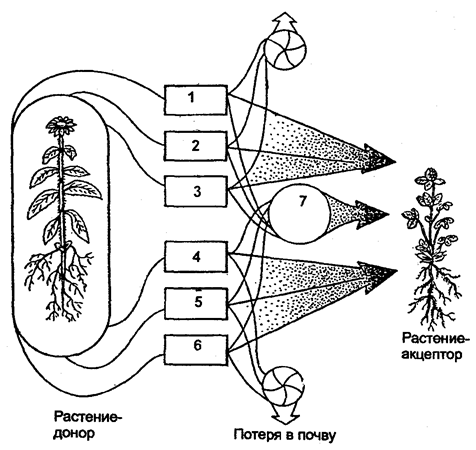
infopedia.su
Глава 2. Разноообразие взаимоотношений между муравьями и растениями. Разноообразие взаимоотношений между муравьями и растениями
Похожие главы из других работ:
Биосфера
7. Взаимоотношения между организмами
Среди огромного разнообразия взаимосвязей живых существ выделяют определенные типы отношений, имеющие много общего у организмов разных систематических групп. 1...
Биоэкологические особенности коллекции видов средиземноморья сочинского парка "Дендрарий"
1.2.2 Ионизация воздуха древесными растениями
Существуют аэроионы легкие, которые могут нести отрицательный или положительный заряды, и тяжелые - положительно заряженные. Наиболее благоприятное воздействие на окружающую среду оказывают легкие отрицательные ионы...
Взаимодействие микроорганизмов с высшими растениями
Глава I. Основы взаимодействия микроорганизмов с высшими растениями
1.1 Понятие и виды взаимодействия микроорганизмов с высшими растениями Растения вступают в тесные взаимоотношения с содержащейся в почве микрофлорой. Некоторые бактерии и грибы, обитающие в почве и на корнях...
Взаимодействие сил в природе
1. Зависимость сил взаимодействия между молекулами от расстояния между ними
Область пространства, в которой проявляется действие молекулярных сил, называют сферой молекулярного действия. Радиус этой сферы равен примерно 1*10-9 м. Силы молекулярного взаимодействия зависят от расстояния между молекулами...
Взаимосвязь и регуляция обмена углеводов, липидов, белков в организме человека
Взаимосвязь между обменами углеводов, липидов и белков
Обмен белков Основные функции: структурная (пластическая), каталитическая (ферменты), сократительная, защитная (антитела), регуляторная (пептидные гормоны), транспортная (мембранные белки-переносчики, сывороточные альбумины...
Дендритные потенциалы действия
Токи, протекающие между клетками
В большинстве случаев электрический ток не может течь напрямую с одной клетки на другую. Существуют, однако, электрически сопряженные клетки. Здесь речь пойдет об особых межклеточных структурах...
Живые организмы и окружающая среда
1.2. Факторы живой природы. Взаимосвязь между живыми организмами.
Каждое растение живет не обособленно, а взаимодействуя с другими растения-ми, бактериями, грибами, животными (рис. 199), Связи между организмами разных видов, обитаю-щих совместно...
Межклеточные контакты
3.3 Адгезионные соединения между клеткой и матриксом
Контакт образуется на небольшом по площади участке. В месте контакта в цитомембрану встроены трансмембранные белки б- и в-интегрины, которые соединяются с элементами межклеточного матрикса...
Особенности показателей вегетативной регуляции и функционального состояния у лиц с разным уровнем двигательной активности
1.1 Соотношения между парасимпатической и симпатической регуляциями сердца
Взаимодействие парасимпатических и симпатических регуляторных аппаратов сердца не подвергалось систематическому углубленному экспериментальному изучению...
Разноообразие взаимоотношений между муравьями и растениями
Глава 1. Типы и формы биотических взаимоотношений между растениями и обществеными перепончатокрылыми насекомыми
Основная черта, возникшая у общественных насекомых в процессе их эволюции и отсутствующая у других насекомых, -- это создание запасов пищи в гнездах, в которых находится и большая часть популяции данного вида. Обитатели гнезда защищают его...
Разноообразие взаимоотношений между муравьями и растениями
Глава 3. Морфологические, экологические, этологические и другие адаптивные приспособления к симбиотическим отношениям с растениями у муравьев
Биологическая адаптация (от лат. Adaptatio -- приспособление) -- приспособление организма к внешним условиям в процессе эволюции, включая морфофизиологическую и поведенческую составляющие...
Роль симметрии и асимметрии в научном познании
8. Асимметрия как разграничивающая линия между живой и неживой природой
Пастером было установлено, что все аминокислоты и белки, входящие в состав живых организмов, являются «левыми», т. е. отличаются оптическими свойствами. Объяснить происхождение «левизны» живой природы он пытался асимметрией...
Современное представление о механизмах и закономерностях эволюции
1.2 Противоречия между генетикой и дарвинизмом
Несмотря на то, что открытая Менделем дискретность наследственности устранила существенные затруднения, связанные с «кошмаром Дженкина»...
Теория эволюции
6. Соотношение между общей теорией эволюции и концепцией коэволюции.
Изучение проблем коэволюции открывает новое и, возможно, важнейшее направление фундаментальных исследований. Часто говорят, что в отличие от века пара, каким был век XIX, и века ХХ, который был веком электричества и атомной энергии...
Эволюция протозойных и грибковых заболеваний
1. Связь между протозойными заболеваниями человека и животных
Как видно из таблицы, большинство протозойных заболеваний до сих пор являются зоонозами. Так, балантиды являются мало патогенными или вовсе непатогенными паразитами свиней; у людей они вызывают либо бессимптомное носительство...
bio.bobrodobro.ru