
Большая Энциклопедия Нефти и Газа. Запасные белки у растений накапливаются где
Запасные белок - Большая Энциклопедия Нефти и Газа, статья, страница 1
Запасные белок
Cтраница 1
Запасные белки, находящиеся в эндосперме или семядолях, малорастворимы ( или вовсе нерастворимы) в воде, не проходят через клеточные оболочки, не используются непосредственно развивающимся зародышем. При прорастании семян они претерпевают глубокий распад с образованием растворимых и подвижных аминокислот. Гидролизуются белки под действием протеолитических ферментов. В прорастающих зернах пшеницы активность протеиназы усиливается приблизительно в 40 раз и гидролиз белков протекает с большой скоростью. [1]
Запасные белки, которые служат источниками углерода и азота прорастающих семян, состоят из ограниченного повторяющегося набора аминокислот. Аминокислотный состав запасных белков семян можно немного изменить обычным скрещиванием, а недавно для этих целей были использованы генноинженер-ные методы. [2]
Запасные белки локализованы в определенных субклеточных структурах ( фото 51) - белковых телах, которые можно выделить из клеточных гомогенатов [77] и которые, вероятно, содержат только глобулины. [3]
Запасные белки, находящиеся в эндосперме или семядолях, малорастворимы ( или вовсе не растворимы) в воде, не проходят через клеточные оболочки, не используются непосредственно развивающимся зародышем. При прорастании семян они претерпевают глубокий распад с образованием растворимых и подвижных аминокислот. Гидролизуются белки под действием протеолитиче-ских ферментов. В прорастающих зернах пшеницы активность протеиназы усиливается приблизительно в 40 раз и гидролиз белков протекает с большой скоростью. Из получившихся аминокислот синтезируются новые белки, идущие на питание развивающегося зародыша и построение тканей растения. [4]
Запасные белки образуются в результате превращения конституционных белков. [5]
Запасные белки перевариваются ( гидролизуются) с высвобождением аминокислот - основных структурных единиц белка. Из этих аминокислот синтезируются ферменты ( все ферменты - белки), например а-амилаза; эти ферменты используются затем для переваривания питательных веществ эндосперма. [6]
Эти же запасные белки имеют дополнительную функцию связывания споровой ДНК, повышая таким образом ее устойчивость к облучению. Энергия для восполнения отсутствующих компонентов может быть получена из стабильного 3-фосфоглицерата путем его конвертации в источник энергии и фосфата - фосфоенолпируват. [8]
Местом отложения запасных белков в семенах являются алейроновые зерна, погруженные в цитоплазму. В алейроновых зернах белки находятся в виде кристаллоидов и глобоидов, содержащих различные по свойствам группы белков. [9]
Повышение интенсивности обновления запасных белков в растениях во время колошения, возможно, находится в связи с процессами интенсивного оттока белковых веществ из вегетативных частей растений в репродуктивные органы, когда происходит усиленное образование запасных белков для формирования колоса. Интенсивность же обновления конституционных белков в стареющих органах растений падает. Особенно резко это проявилось в опыте с тимофеевкой, где конституционные белки даже за 120 часов обновились только на 66 7 %, в то время как в молодых растениях овса и ржи конституционные белки полностью обновлялись в течение 72 часов. Интенсивность обновления хлорофилла при старении растений также падает. Если в молодых растениях о-вса и ржи ( 1) азот хлорофилла в течение 48 часов обновлялся примерно на 60 %, то в стадии колошения овса за тот же период обновилось только 25 % азота хлорофилла. [10]
В основном это касается запасных белков. [11]
В этом опыте обновление запасных белков происходило менее интенсивно, чем в предыдущем-с рожью, что, по-видимому, обусловлено особенностями метаболизма щавеля. [12]
В каких частях растения откладываются запасные белки. [13]
Хотя относительно протеаз, гидролизующих запасные белки семян, известно немногое, а об их регуляции по существу ничего не известно, было показано, что проростки способны переработать относительно большие количества аммиака, который образуется в результате окисления аминокислот. Глутаминсин-тетаза и аспарагинсинтетаза описаны в гл. [14]
Наконец, следует упомянуть группу запасных белков. [15]
Страницы: 1 2 3 4
www.ngpedia.ru
Запасные белок - Большая Энциклопедия Нефти и Газа, статья, страница 2
Запасные белок
Cтраница 2
Основная масса белков семян злаков - запасные белки, представляющие собой альбумины, глобулины, проламины и глюте-лины. [16]
Скорость обновления РНК в период накопления запасных белков с 8-го по 35 - й день неизвестна. [17]
Даниэльссон [25] показал, что основная масса запасных белков в развивающихся и созревающих семядолях гороха ( фиг. Эти два запасных белка не только различаются по растворимости, константам седиментации и аминокислотному составу, но и откладываются также с неодинаковой скоростью в период развития. Какое значение может иметь для растения наличие двух запасных глобулинов, не ясно. То, что оба эти белка играют запасную роль, подтверждается их исчезновением во время прорастания. Ферментативная активность у этих запасных глобулинов не обнаружена. Предполагают, что большую часть альбуминовой фракции составляют ферменты, не связанные с частицами. На эту фракцию приходится небольшая часть общего азота гороха, и в процессе развития и прорастания семян эта фракция незначительно изменяется. [18]
Кроме триглицеридов в созревающих масличных семенах накапливаются также запасные белки. Суммарное содержание масла ( запасных липидов) и запасных белков составляет до 80 % массы созревших масличных семян. [19]
При внесении меченой азотной подкормки содержание в корнях мобильных, запасных белков резко повышалось за счет их оттока из листьев растений. Высказано предположение, что переработка в корнях неорганического азота осуществляется при участии запасных белков, как носителей соответствующих ферментных систем, катализирующих Синтез аминокислот в растении. При повышенном притоке неорганического азота содержание этих белков в корнях становится недостаточным и растения восполняют этот недостаток оттоком запасных белков из листьев в корни. [20]
Выполненные в нашей лаборатории анализы В. М. Мака-ревич показали, что запасные белки всех исследованных растений совершенно не содержат пролина и что они значительно богаче триптофаном, аланином, валином и лейцином, чем конституционные белки. [21]
Если в ранних фазах роста и развития растений овса конституционные белки обновляются значительно быстрее и интенсивнее запасных белков ( см. табл. 3), то в фазе колошения конституционные и запасные белки обновляются примерно с одинаковой интенсивностью. В колосьях же запасные белки обновляются значительно быстрее конституционных. Резкое повышение интенсивности обновления запасных белков в колосьях в фазе колошения, невидимому, находится в связи с интенсивным оттоком белковых веществ из листьев и стеблей в репродуктивные органы. Наконец, в этой фазе развития растений значительно понижается интенсивность обновления азотистого состава хлорофилла. [22]
Сконструированы такие генноинженерные сорта сои, которые проявляют устойчивость к насекомым, гербицидам, вирусам и образуют больше запасных белков, обогащенных метионином. [23]
Характерной особенностью хлоропластов большинства водорослей являются так называемые пиреноиды - тела неправильной формы, содержащие, как предполагают, запасные белки и часто окруженные крахмальным сдоем ( см. фиг. По Бозе [67], масляные капельки, образующиеся при фотосинтезе у водорослей, сперва появляются в этих слоях. [24]
Из результатов этого опыта также следует, что скорость обновления двух групп выделенных нами из растений белковых веществ - конституционных и запасных белков - весьма различна. Конституционные белки обновляются со значительно большей интенсивностью, чем запасные. В других опытах, с более короткими экспозициями растений на N15, меченый азот мог быть обнаружен в конституционных белках в более ранние сроки, чем в запасных белках. Это дает известное основание предполагать, что вначале синтезируются конституционные белки, которые, претерпевая соответствующие изменения, превращаются в более подвижные запасные белки. [25]
При обсуждении результатов химического анализа растений мы указывали, что по этим данным невозможно было установить какие-либо закономерности в изменении содержания запасных белков в растениях при различных сроках их уборки. Результаты изотопного анализа, наоборот, указывают на сильное обновление азота этих ( белков через 48 и 96 часов после внесения подкормки с меченым азотом. Это заставляет нас признать, что в действительности зайасные белки, так же как и конституционные, подвергались непрерывным изменениям в организме растений. [26]
Кальций усиливает обмен веществ в растениях, играет важную роль в передвижении углеводов, оказывает влияние на превращение азотистых веществ, ускоряет распад запасных белков семени при прорастании. Кроме того, он имеет существенное значение для построения нормальных клеточных оболочек и для установления благоприятного кислотно-щелочного равновесия в растениях. [27]
Если в ранних фазах роста и развития растений овса конституционные белки обновляются значительно быстрее и интенсивнее запасных белков ( см. табл. 3), то в фазе колошения конституционные и запасные белки обновляются примерно с одинаковой интенсивностью. В колосьях же запасные белки обновляются значительно быстрее конституционных. Резкое повышение интенсивности обновления запасных белков в колосьях в фазе колошения, невидимому, находится в связи с интенсивным оттоком белковых веществ из листьев и стеблей в репродуктивные органы. Наконец, в этой фазе развития растений значительно понижается интенсивность обновления азотистого состава хлорофилла. [28]
Так, например, известно, что в целых колосьях пшеницы ( 20 дней после оплодотворения) включение аминокислот, меченных С14, в цитоплаз-матические белки ингибируется фторацетатом, тогда как включение в запасные белки оказывается нечувствительным к этому соединению. Включение меченых аминокислот в бесклеточной фракции белковых тел не зависит от добавления АТФ или системы, генерирующей энергию, а также от добавления рибосом или над-осадочной фракции. После разрушения изолированных белковых тел ультразвуком удается выделить рибосомы, активирующие ферменты и транспортную РНК. [29]
Повышение интенсивности обновления запасных белков в растениях во время колошения, возможно, находится в связи с процессами интенсивного оттока белковых веществ из вегетативных частей растений в репродуктивные органы, когда происходит усиленное образование запасных белков для формирования колоса. Интенсивность же обновления конституционных белков в стареющих органах растений падает. Особенно резко это проявилось в опыте с тимофеевкой, где конституционные белки даже за 120 часов обновились только на 66 7 %, в то время как в молодых растениях овса и ржи конституционные белки полностью обновлялись в течение 72 часов. Интенсивность обновления хлорофилла при старении растений также падает. Если в молодых растениях о-вса и ржи ( 1) азот хлорофилла в течение 48 часов обновлялся примерно на 60 %, то в стадии колошения овса за тот же период обновилось только 25 % азота хлорофилла. [30]
Страницы: 1 2 3 4
www.ngpedia.ru
Липидные (жировые) капли
Поиск ЛекцийСодержание
1.Введение…………………………………………………………...2
2.Запасные вещества………………………………………………3-6
3. Крахмальные зёрна……………………………………………..7-9
4.Липидные капли……………………………………………….10-11
5.Запасные белки………………………………………………..12-13
6. Характер включений…………………………………………14-15
7.Химический состав……………………………………………….16
8.Недостаток питательных веществ………………………………17
9.Вывод……………………………………………………………...18
10. Сприсок используемой литературы……………………………19
Введение
Растение, как и всякий живой организм, состоит из клеток, причем каждая клетка порождается тоже клеткой. Клетка — это простейшая и обязательная единица живого, это его элемент, основа строения, развития и всей жизнедеятельности организма.Существуют растения, построенные из одной единственной клетки. К ним относятся одноклеточные водоросли и одноклеточные грибы. Обычно это микроскопические организмы, но есть и довольно крупные одноклеточные (длина одноклеточной морской водоросли ацетабулярии достигает 7 см). Большинство растений, с которыми мы сталкиваемся в повседневной жизни,— это многоклеточные организмы, построенные из большого числа клеток.Запасные продукты — это вещества, временно выключенные из обмена веществ. Они расходуются на построение тела растения или на различные жизненные процессы как энергетический материал. Местом отложения их служат вакуоли и цитоплазма. В вакуолях запасные продукты накапливаются в виде растворов, в цитоплазме — в виде оформленных включений: алейроновых, крахмальных зерен, капель жирного масла и др. Алейроновые зерна — это гранулы запасного белка. Они обычно образуются в клетках запасающей ткани зрелых семян.
Запасные продукты
— это вещества, временно выключенные из обмена веществ. Они расходуются на построение тела растения или на различные жизненные процессы как энергетический материал. Местом отложения их служат вакуоли и цитоплазма. Вакуоли (рис.1) - полости в протопласте эукариотических клеток. У растений вакуоли - производные эндоплазматической сети , ограниченные мембраной – тонопластоми заполненные водянистым содержимым - клеточным соком. Протопласт - активное содержимое растительной клетки. Основной компонент протопласта - белок . У большинства зрелых растительных клеток центральную часть занимает крупная, заполненная клеточным соком вакуоль , главное содержимое которой - вода с растворенными в ней минеральными и органическими веществами. Клеточная оболочка и вакуоль представляют собой продукты жизнедеятельности протопласта . Большую часть протопласта растительной клетки занимает цитоплазма , меньшую по массе - ядро . От вакуоли протопласт отграничен мембраной, называемой тонопластом , от клеточной стенки - другой мембраной - плазмалеммой . В протопласте осуществляются все основные процессы клеточного метаболизма.По- видимому, существенную роль в образовании вакуолей имеет деятельность аппарата Гольджи .
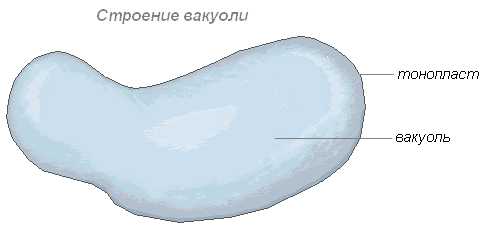 рис.1
рис.1
Функции вакуолей :
1)Они формируют внутреннюю водную среду клетки, и с их помощью осуществляется регуляция водно-солевого обмена.
2)Поддержание тургорного гидростатического давления внутриклеточной жидкости в клетке. 3) Обеспечение роста клетки. В молодых делящихся растительных клетках вакуоли представляют систему канальцев и пузырьков (провакуоли), по мере роста клеток они увеличиваются, а затем сливаются в одну большую центральную вакуоль. Она занимает от 70 до 90 % объёма клетки, в то время как протопласт располагается в виде тонкого постенного слоя.
3Накопление запасных веществ и "захоронение" отбросов, т.е. конечных продуктов метаболизма клетки. Иногда вакуоли разрушают токсичные или ненужные клетке вещества. Обычно это выполняется специальными небольшими вакуолями, содержащими соответствующие ферменты. Такие вакуоли получили название лизосомных .
В вакуолях запасные продукты накапливаются в виде растворов, в цитоплазме — в виде оформленных включений: алейроновых, крахмальных зерен, капель жирного масла и др.
Алейроновые зерна — это гранулы запасного белка. Они обычно образуются в клетках запасающей ткани зрелых семян.(рис.2)
При формировании семян в мелких вакуолях накапливается белок. В созревающих семенах вакуоли теряют воду и превращаются в алейроновые зерна. При прорастании семян, когда они обогащаются водой, алейроновые зерна вновь преобразуются в вакуоли.
Алейроновые зерна имеют округлую форму, диаметр их колеблется от 0,2 до 20 мкм. Снаружи они покрыты мембраной.
У простых алейроновых зерен белок находится в виде аморфной массы (бобовые, кукуруза, рис), у сложных — в аморфную массу включен один, реже 2—3 белковых кристалла и небольшое округлое тельце — глобоид, содержащее запасной фосфор. Белковые тела могут образовываться и в других частях клетки — в ядре, пластидах, митохондриях, эндоплазматическом ретикулуме. Алейроновые зёрна имеют округлую форму ,диаметр их колеблется от 0,2 до 20 мкм. Снаружи они покрыты мембраной. У простых алейроновых зёрнах белок находится в виде аморфной массы (бобовые, кукуруза, рис),у сложных – в аморфную массу вкючен один, реже 2-3 белковых кристалла и небольшое округлое тельце – глобоид, содержащее запасной фосфор.
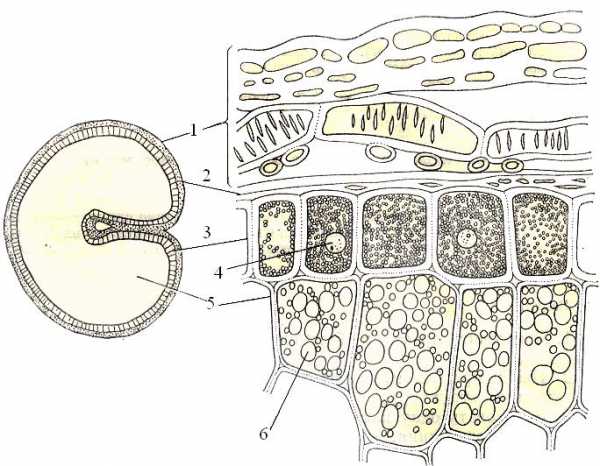
Рис.2Запасные вещества в зерновке пшеницы (Triticum aestivum) на поперечном срезе:
1 - околоплодник, 2 - кожура семени, 3 - алейроновый слой, 4 - ядро, 5 - клетки эндосперма с крахмальными зернами, 6 - крахмальные зерна.
Крахмальные зерна.
Зёрна крахмала в клетках картофельного клубня. Поперечный срез
Главнейшее и наиболее распространенное из них - полисахарид крахмал . Крахмал злаков , клубней картофеля, ряда тропических растений - важнейший источник углеводов в рационе человека.В хлоропластах откладывается откладываются зёрна первичного крахмала. Но здесь он не накапливается .При помощи ферментов первичный крахмал осахаривается и в виде глюкозы транспортируется из листа в другие органы.Вторичное превращение сахара в крахмал происходит в лейкопластах (амилопластах). Образование зерен вторичного крахмала начинается в определённых точках стромы амилопласта, называется образовательными центрами . Рост крахмальных зерен происходит путем наложения новых слоев крахмала на старые, поэтому они имеют слоистую структуру. Смежные слои могут иметь различные показатели преломления и поэтому видны под микроскопом.Слоистость бывает концентрической и эксцентрической.С ростом крахмального зерна объём стромы амилопласта уменьшается, и в определённый момент слой ее становится настолько тонким ,что неразлечим под световым микроскопом. Надо помнить, что двухмембранная оболочка и тонкий слой стромы всегда присутствует на поверхности зерна вторичного крахмала.
Если имеется один центр, вокруг которого откладываются слои крахмала, то возникает простое зерно, если два и более, то образуется сложное зерно, состоящее как бы из нескольких простых. Полусложное зерно формируется в тех случаях, когда крахмал сначала откладывается вокруг нескольких точек, а затем после соприкосновения простых зерен вокруг них возникают общие слои. Расположение слоев может быть концентрическим или эксцентрическим, что также определяет особенности строения крахмальных зерен.
В химическом отношении крахмал представляет собой полисахарид, сходный с целлюлозой, построенный из сотен глюкозных остатков.Первичный ассимиляционный крахмал образуется только в хлоропластах . Ночью, когда фотосинтез прекращается, ассимиляционный крахмал ферментативно гидролизуется до сахаров и транспортируется в другие части растения.
В запасающих тканях различных органов, особенно в клубнях, луковицах, корневищах и др., в особом типе лейкопластов - амилопластах часть сахаров откладывается в виде зерен вторичного крахмала .
Если в амилопласте имеется один образовательный центр ,то формируется простое зерно, если два и более – сложное зерно ,состоящее как бы из несколько простых. Полусложное зерно образуется в том случае, если крахмал сначала откладывается вокруг нескольких образовательных центров, а затем после соприкосновения простых зёрен вокруг них возникают общие слои.Размер крахмальных зёрен колеблется в больших пределах.Так, у картофеля диаметр их достигает 100мкм, у пшеницы и ржи бывают мелкие зёрна диаметром2-9 мкм и крупные диаметром 30-45 мкм, у кукурузы – диаметром 5-30 мкм.Форма, размер, структура крахмальных зёрен специфичны для каждого вида растения, а иногда и для сорта. Это обстоятельство используют при анализе состава муки.
Липидные (жировые) капли
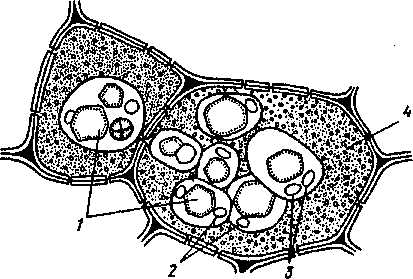
1-Алейроновое зерно;2- Оболочка алейронового зерна ; 3-Глобоиды; 4-Липидные капли
Обычно располагаются в гиалоплазме и встречаются практически во всех растительных клетках. Это основной тип запасных питательных веществ большинства растений. Липидные капли встречаются практически во всех растительных клетках. Жирные масла накапливаются у огромного количества растений и по своему значению являются второй после крахмала формой запасных питательных веществ. Особенно богаты ими семена и плоды. Семена некоторых растений (подсолнечник, хлопчатник, арахис) могут содержать до 40% масла от массы сухого вещества. Поэтому растительные жиры получают, главным образом, из семян.
Липидные капли накапливаются непосредственно в гиалоплазме. Они выглядят как мелкие сферические тела, каждая капля отделена от гиалоплазмы мембраной. Иногда липидные капли называют сферосомами.
Реактивом на жирное масло является краситель судан III, липидные капли окрашиваются им в оранжево-красный цвет.
Запасные белки
Запасные белки относятся к категории простых белков – протеинов, в отличие от сложных белков – протеидов, составляющих основу протопласта. В наибольшем количестве они откладываются в запасающей ткани сухих семян в виде алейроновых зерен, или белковых телец.
Относятся к категории простых белков - протеинов в отличие от сложных белков - протеидов , составляющих основу протопласта . Наиболее часто запасные белки откладываются в семенах. Очень богаты белками семена многих используемых в пищу и кормовых видов бобовых. Иногда протеины обнаруживаются в ядре и гиалоплазме в виде трудно различимых в световой микроскоп кристаллоподобных структур. Однако чаще запасные белки накапливаются в вакуолях и выпадают в осадок при потере влаги в процессе созревания семян.
Обычно осаждающиеся белки образуют зерна округлой или эллиптической формы, называемые алейроновыми зернами . Если алейроновые зерна не имеют заметной внутренней структуры, их называют простыми. Иногда же в алейроновых зернах среди аморфного белка заметны один или несколько кристаллоподобных структур (кристаллоидов), способных в отличие от настоящих кристаллов набухать в воде. Помимо кристаллоидов, в алейроновых зернах встречаются блестящие бесцветные тельца округлой формы - глобоиды. Алейроновые зерна, содержащие кристаллоиды и глобоиды, называют сложными. У каждого вида растений они, подобно зернам крахмала, имеют определенную структуру.
Растения в отличие от животных не имеют специальных выделительных органов и нередко накапливают конечные продукты жизнедеятельности протопласта в виде солей оксалата или карбоната кальция. Кристаллические включения в значительных количествах накапливаются в тканях и органах, которые растения периодически сбрасывают (листья, кора). Они откладываются исключительно в вакуолях. Форма этих включений достаточно разнообразна: одиночные многогранники - стилоиды (палочковидные кристаллы), игольчатые кристаллы - рафиды, скопления множества мелких кристаллов - кристаллический песок, сростки кристаллов - друзы. Форма кристаллов нередко специфична для определенных таксонов и иногда используется для их микродиагностики.
К кристаллическим включениям близки цистолиты. Они чаще всего состоят из карбоната кальция или кремнезема и представляют собой гроздевидные образования, возникающие на выступах клеточной оболочки, вдающейся внутрь клетки. Цистолиты характерны для растений семейств крапивных, тутовыхи др.Белковые включения можно окрасить реактивом Люголя в золотисто-желтый цвет.
По характеру все включения
- это продукты клеточного метаболизма. Они накапливаются главным образом в форме гранул, капель и кристаллов. Химический состав включений очень разнообразен.
Липоидыобычно откладываются в клетке в виде мелких капель. Большое количество жировых капель встречается в цитоплазме ряда простейших, например инфузорий. У млекопитающих жировые капли находятся в специализированных жировых клетках, в соединительной ткани. Часто значительное количество жировых включений откладывается в результате патологических процессов, например при жировом перерождении печени. Капли жира встречаются в клетках практически всех растительных тканей, очень много жира содержится в семенах некоторых растений.
Включения полисахаридов имеют чаще всего формулу гранул разнообразных размеров. У многоклеточных животных и простейших в цитоплазме клеток встречаются отложения гликогена. Гранулы гликогена хорошо видны в световом микроскопе. Особенно велики скопления гликогена в цитоплазме поперечнополосатых мышечных волокон и в клетках печени, в нейронах. В клетках растений из полисахаридов наиболее часто откладывается крахмал. Он имеет вид гранул различной формы и размеров, причем форма крахмальных гранул специфична для каждого вида растений и для определенных тканей. Отложениями крахмала богата цитоплазма клубней картофеля, зерен злаков; каждая крахмальная гранула состоит их отдельных слоев, а каждый слой, в свою очередь, включает радиально расположенные кристаллы, почти невидимые в световой микроскоп.
Белковые включения встречаются реже, чем жировые и углеводные. Белковыми гранулами богата цитоплазма яйцеклеток, где они имеют форму пластинок, шариков, дисков, палочек. Белковые включения встречаются в цитоплазме клеток печени, клеток простейших и многих других животных.
К клеточным включениям относятся некоторые пигменты, например распространенный в тканях желтый и коричневый пигмент липофусцин, круглые гранулы которого накапливаются в процессе жизнедеятельности клеток, особенно по мере их старения. Сюда же относятся пигменты желтого и красного цвета - липохромы. Они накапливаются в виде мелких капель в клетках коркового вещества надпочечников и в некоторых клетках яичников. Пигмент ретинин входит в состав зрительного пурпура сетчатки глаза. Присутствие некоторых пигментов связано с выполнением этими клетками особых функций. Примерами могут служить красный дыхательный пигмент гемоглобин в эритроцитах крови или пигмент меланин в клетках меланофорах покровных тканей животных.
В качестве включений во многих животных клетках присутствуют гранулы секрета, вырабатываемого в клетках разных типов, в первую очередь в железистых. Секреторные включения могут быть белками, сахаридами, липопротеидами и т. д
poisk-ru.ru








