Содержание
Общая характеристика водорослей
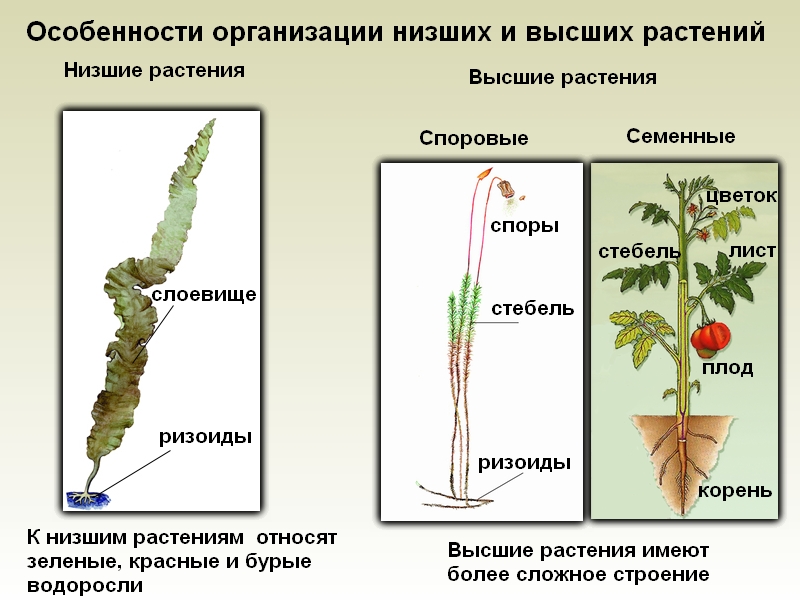
Особенности строения тела. Водоросли — сборная группа низших, преимущественно водных, растений. Характерной особенностью всех водорослей является то, что их тело не расчленено на вегетативные органы (корень, стебель, лист), а представлено талломом, или слоевищем. По этой причине их называют талломными, или слоевищными, растениями. В отличие от высших растений у них обычно отсутствуют ткани, а органы полового и бесполого размножения, как правило, одноклеточные. Общей для водорослей является также способность к фототрофному способу питания. Вместе с тем у некоторых водорослей наряду с автотрофным существует гетеротрофный тип питания (например, у эвгленовых).
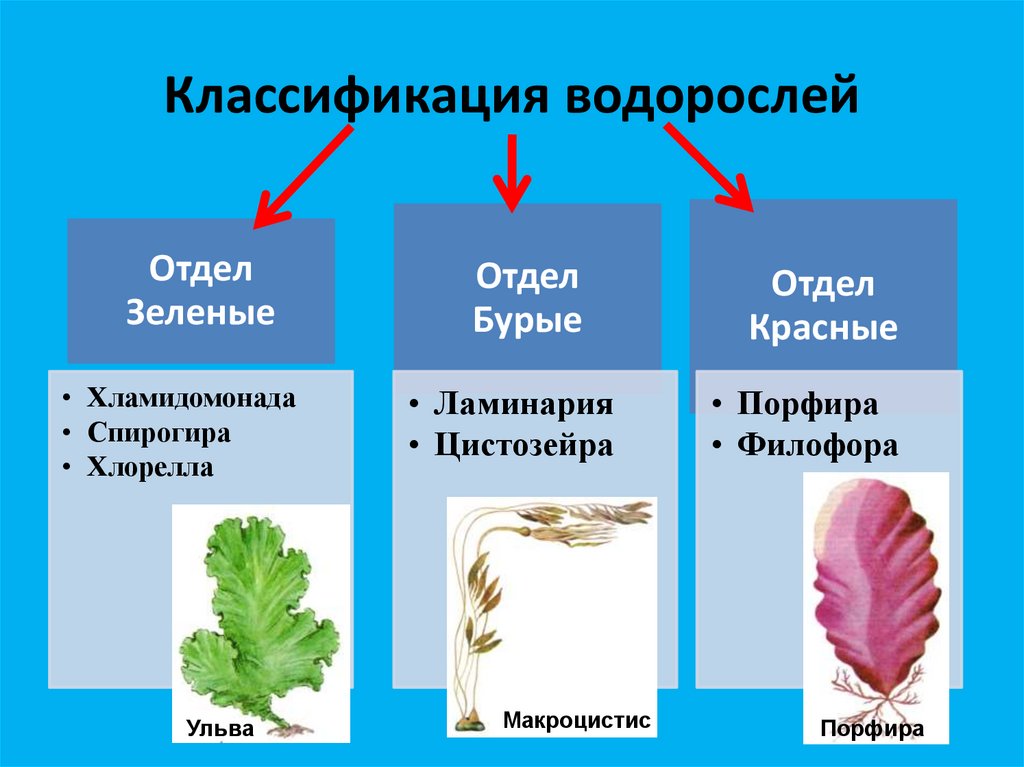
Известно более 40 тыс. видов водорослей, которые объединяются в два полцарства — Багрянки и Настоящие водоросли. Последние подразделяются на несколько обособленных отделов (Зеленые, Харовые, Диатомовые, Золотистые, Бурые и др.), которые отличаются друг от друга по ряду таких важнейших признаков, как структура таллома, набор фотосинтезирующих пигментов и запасных питательных веществ, особенности размножения и циклы развития, местообитание и т. п.
п.
| Строение многоклеточных водорослей. Слева клетка нитчатой спирогиры, справа – фукус пузырчатый |
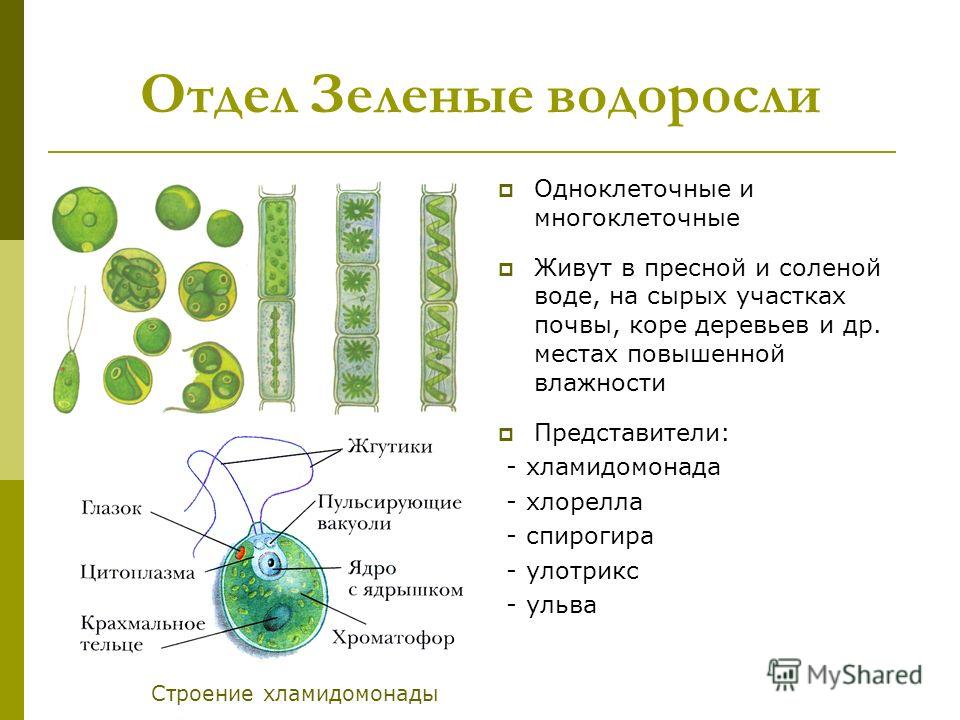
Водорослям присуще исключительное морфологическое разнообразие. Они бывают одноклеточные (хламидомонада, хлорелла), колониальные (вольвокс) и многоклеточные, среди которых встречаются нитчатые (спирогира, улотрикс) и пластинчатые (уль-ва, ламинария и др.). Каждая из этих форм имеет различные размеры — от I мкм до нескольких десятков метров (некоторые бурые и красные). Например, бурая водоросль макроцистис грушеносный вырастает за день на 45 см и достигает длины 160 м.
| Строение одноклеточных водорослей. Слева эвглена зелёная, справа – хламидомонада |
Строение клетки. Клетки большинства водорослей существенно не отличаются от типичных клеток высших растений, однако у них есть свои особенности.
Клетки большинства водорослей существенно не отличаются от типичных клеток высших растений, однако у них есть свои особенности.
Клетки водоросли имеют клеточную оболочку, состоящую из целлюлозы и пектиновых веществ. У многих из них в состав клеточной стенки входят добавочные компоненты: известь, железо, альгиновая кислота и др.
Цитоплазма у большинства водорослей расположена тонким слоем вдоль клеточной стенки и окружает большую центральную вакуоль. В цитоплазме хорошо различимы эндоплазмати-ческий ретикулум, митохондрии, аппарат Гольджи, рибосомы, одно или несколько ядер.
В клетках водорослей из органелл особенно заметны хромато-форы (хлоропласты), которые в отличие от хлоропластов высших растений более разнообразны по форме, размерам, числу, строению, местоположению и набору пигментов. Они могут быть чашевидными, лентовидными, пластинчатыми, звездчатыми, дис-ковидными и др.
В хроматофорах сосредоточены фотосинтезирующие пигменты: хлорофиллы а, b, с, d, каротиноиды (каротины и ксантофиллы), фикобилины (фикоцианин, фикоэритрин).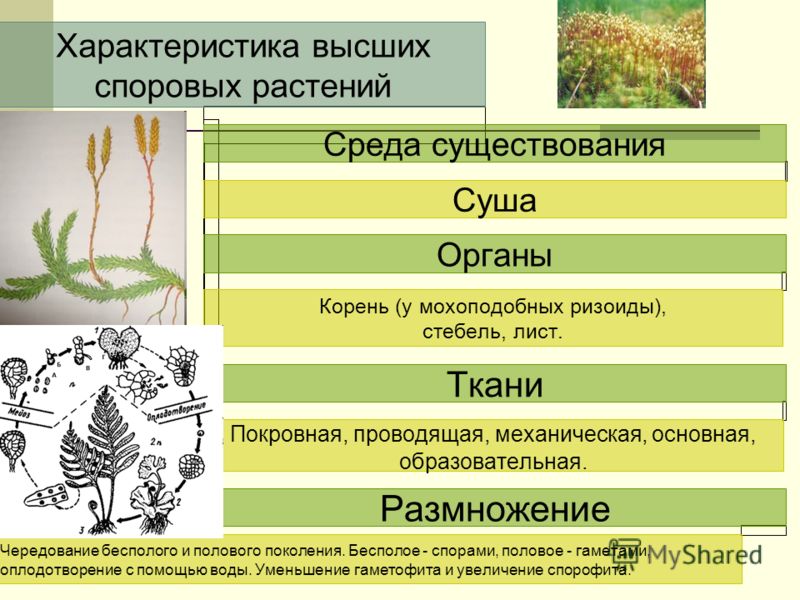 Кроме того, в матриксе хроматофора находятся рибосомы, ДНК, липидные гранулы и особые включения — пиреноиды. Пиреноиды присущи почти всем водорослям и небольшой группе мхов. Они являются не только местом скопления запасных питательных веществ, но и зоной их синтеза.
Кроме того, в матриксе хроматофора находятся рибосомы, ДНК, липидные гранулы и особые включения — пиреноиды. Пиреноиды присущи почти всем водорослям и небольшой группе мхов. Они являются не только местом скопления запасных питательных веществ, но и зоной их синтеза.
Веществами запаса у водорослей служат крахмал, масло, гликоген, волютин, водорастворимый полисахарид ламинарии и др.
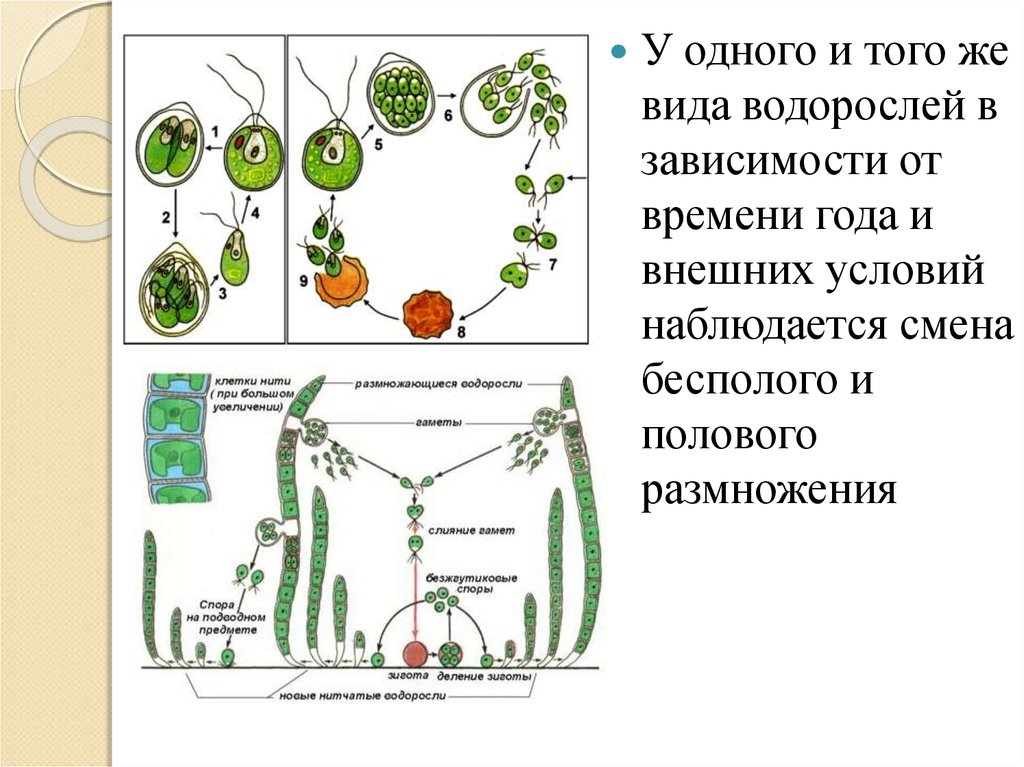
Размножение. Водоросли размножаются бесполым и половым путем. Бесполое размножение происходит фрагментами колонии или таллома, а также спорами.
У одних водорослей каждая особь способна формировать и споры, и гаметы в зависимости от времени года и условий среды, у других функции бесполого и полового размножения выполняют разные особи — спорофиты (образуют споры) и гаметофиты (формируют гаметы). У многих водорослей в цикле развития наблюдается строгое чередование поколений — спорофита и гаме-тофита (красные, бурые, Некоторые зеленые).
Экологические группировки водорослей.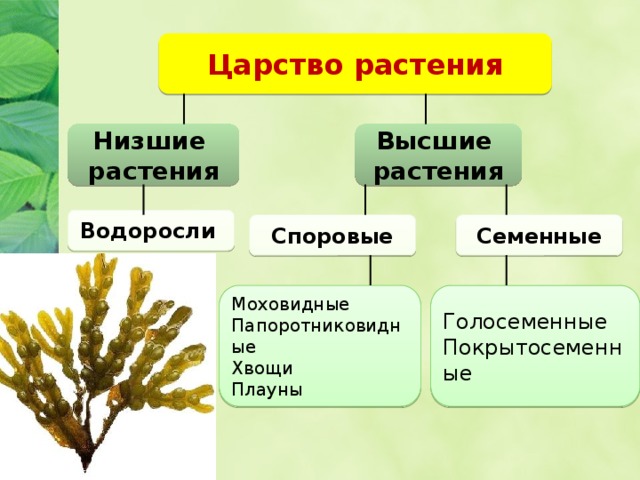 Большинство водорослей исторически приспособились к определенным комплексам экологических факторов и образовали своеобразные сообщества, или экологические группировки: планктонные, бентосные, наземные, почвенные и др.
Большинство водорослей исторически приспособились к определенным комплексам экологических факторов и образовали своеобразные сообщества, или экологические группировки: планктонные, бентосные, наземные, почвенные и др.
Планктоном называют совокупность преимущественно микроскопических свободно плавающих в толще воды организмов.
Среди них могут быть зеленые, диатомовые, золотистые, эвглено-вые водоросли. Для облегчения переноса водой планктонные водоросли имеют различные приспособления, уменьшающие плотность организмов (газовые вакуоли, включения липидов, насыщенность водой и студенистость) и увеличивающие их удельную поверхность (разветвленные выросты, приплюснутая или вытянутая форма тела и др.).
Бентосом называют водные организмы, живущие на дне водоемов или обрастающие различные водные предметы. Фито-бентос морей и океанов состоит преимущественно из бурых и красных водорослей, а в пресных водоемах представлены практически все отделы, кроме Бурых.
Обширную экологическую группировку составляют наземные, или воздушные, водоросли. Они образуют различно окрашенные налеты и пленки на коре деревьев, влажных камнях, заборах, крышах домов и т. д. Большинство из них относятся к отделам Зеленые и Диатомовые водоросли.
Почвенные водоросли живут в толще почвенного слоя. Это в основном желтозеленые, диатомовые и эвгленовые.
Кроме того, водоросли развиваются в горячих источниках, на поверхности снега и льда и т. п.
Источник : Н.А. Лемеза Л.В.Камлюк Н.Д. Лисов «Пособие по биологии для поступающих в ВУЗы»
Строение и жизнедеятельность водорослей — что это, определение и ответ
Характерной особенностью водорослей является то, что их тело НЕ расчленено на ткани и органы, а представлено талломом (или слоевищем). Именно поэтому водоросли относятся к низшим растениям. Так же для них характерно наличие хроматофоров – органоиды, подобные хлоропластам высших растений.
Вспомним! Наука о водорослях — альгология.
Строение клетки водоросли
Строение клетки водоросли в целом схоже со строением растительной клетки, однако она имеет свои особенности.
Одной из главных отличительных черт водорослевой клетки является организация фотосинтезирующего пигмента не в виде хлоропласта, а в виду специального образования – хроматофора, который может иметь самую замысловатую форму в зависимости от типа пигмента и вида самой водоросли.
Ниже представлено строение нитчатой водоросли спирогиры. Слева – схематичный рисунок, справа – фотография микропрепарата реальной спирогиры.
Хроматофор
Хоть тело водорослей и НЕ разделено на ткани и органы, однако они отличаются большим морфологическим многообразием: бывают одно- и многоклеточными, а также колониальными.
Колониальный организм отличается от многоклеточного тем, что в многоклеточном клетки дифференцируются по функциям (хотя бы частично), в колонии же каждая клетка в основном является практически самостоятельным организмом.
Колониальная водоросль вольвокс
Строение многоклеточных водорослей соответствует среде их обитания.
Особенности, характерные для растения, живущего в воде:
● Наличие целлюлозно-пектиновой оболочки позволяет водорослям сочетать прочность с возможностью интенсивного роста и высокой проницаемостью.
● У бентосных (донных) водорослей слоевище прикрепляется к дну с помощью ризоидов или базального диска. Это приспособление позволяет им не отрываться от субстрата во время прибоя и при ударах волн.
Ризоиды
● Таллом водоросли обычно не сплошной, а разделенный – это помогает водоросли снизить сопротивление толщи воды.
● Адаптацией к изменению спектра света, доходящего до глубин океана, стало формирование новых фотосинтезирующих пигментов, а также увеличение числа пигментов в клетке по мере роста глубины.
● Таллом водоросли достаточно часто бывает покрыт слизью, которая удерживает воду внутри водоросли, препятствует излишней ее потере.
● Внутри водоросли часто можно заметить воздушные пузырьки, которые помогают им удерживать вертикальное положение в воде.
● Размножение водорослей обычно происходит в период прилива. В это время споры и гаметы выходят из тела растения и дают начало новым водорослям. Молодые особи начинают свое развитие практически сразу же, чтобы не быть унесенными в море во время отлива.
Экология водорослей
Водоросли – самая древняя группа растений на Земле. Они обитают как в воде, так и на суше – во влажной почве, на коре деревьев. Также они способны вступать в симбиоз с грибами и образовывать лишайники. Бурые водоросли вместе с кораллами формируют океанические острова.
Водоросли являются началом цепей питания, так как способны улавливать энергию света и превращать ее в энергию химических связей в органических веществах, которые являются пищей для животных, начиная простейшими и заканчивая млекопитающими.

Не менее важной функцией водорослей является обогащение воды кислородом, который они выделяют в процессе фотосинтеза. Без кислорода в воде не смогли бы дышать ни животные, ни сами растения.
Водоросли поставляют многим животным среду для обитания: в толще их разветвленного таллома часто живут многие беспозвоночные, а также молодь рыб и земноводных.
Значение водорослей для человека
Человек также активно использует водоросли:
Их возделывают в крупных масштабах для получения ценных пищевых добавок в животноводстве
В промышленности из водорослей извлекают соли калия, йод, бром и другие вещества
Морская капуста (ламинария) имеет пищевое, диетическое и кормовое значение
Агар-агар, получаемый из некоторых бурых водорослей, используется в пищевой промышленности и в лабораториях.
Прочитайте темы: “Размножение водорослей”, “Классификация водорослей” и переходите к тестированию.
Что такое водоросли? | Live Science
Когда вы совершаете покупку по ссылкам на нашем сайте, мы можем получать партнерскую комиссию. Вот как это работает.
Водоросли могут стать пищей и топливом будущего.
(Изображение предоставлено: Богдан Ионеску, Шаттерсток)
Водоросли представляют собой разнообразную группу водных организмов, обладающих способностью к фотосинтезу. Некоторые водоросли знакомы большинству людей; например, морские водоросли (такие как ламинария или фитопланктон), прудовая пена или цветение водорослей в озерах. Однако существует огромный и разнообразный мир водорослей, которые не только полезны для нас, но и имеют решающее значение для нашего существования.
Определение
Термин «водоросли» охватывает множество различных организмов, способных производить кислород посредством фотосинтеза (процесс сбора световой энергии солнца для производства углеводов). Эти организмы не обязательно являются близкородственными. Однако некоторые черты объединяют их, отличая при этом от другой большой группы фотосинтезирующих организмов — наземных растений.
Эти организмы не обязательно являются близкородственными. Однако некоторые черты объединяют их, отличая при этом от другой большой группы фотосинтезирующих организмов — наземных растений.
Во-первых, водоросли не так высоко дифференцированы, как растения, согласно авторам «Водорослей: анатомия, биохимия и биотехнология, 2-е изд.». (CRC Press, 2014). Другими словами, у них отсутствуют настоящие корни, стебли и листья, а также сосудистая система для циркуляции воды и питательных веществ по телу. Во-вторых, согласно статье 2014 года, опубликованной в журнале Current Biology, многие водоросли являются одноклеточными. Они также встречаются в различных формах и размерах. Они могут существовать в виде одиночных микроскопических клеток; они могут быть макроскопическими и многоклеточными; живут колониями; или принять лиственный вид, как в случае водорослей, таких как гигантские водоросли. Диаметр пикопланктона составляет от 0,2 до 2 микрометров, а длина листьев гигантских водорослей достигает 60 метров. Наконец, водоросли встречаются в различных водных средах обитания, как пресноводных, так и соленых.
Наконец, водоросли встречаются в различных водных средах обитания, как пресноводных, так и соленых.
В силу этих характеристик общий термин «водоросли» включает прокариотические организмы — цианобактерии, также известные как сине-зеленые водоросли, — а также эукариотические организмы (все другие виды водорослей). «Поскольку «водоросли» не образуют естественную группу, которая произошла от общего предка, включение цианобактерий в неформальную группу «водоросли» является обычным явлением», — говорит Линда Грэм, профессор ботаники из Университета Висконсин-Мэдисон. «Термин «эукариотические водоросли» исключает цианобактерии». Интересно также отметить, что хлоропласты, являющиеся местом фотосинтеза у наземных растений, являются адаптированными формами цианобактерий. По данным Музея палеонтологии Калифорнийского университета, эти ранние цианобактерии были поглощены клетками примитивных растений где-то в конце протерозоя или в раннем кембрийском периоде.
(Прокариоты включают бактерии и археи. Это более простые организмы без организованной клеточной структуры, и их ДНК свободно плавает в цитоплазме в виде запутанной массы. С другой стороны, эукариоты — это все другие живые организмы: протисты, растения, грибы. Что такое Протисты и животные. Их клетки более организованы. У них есть структуры, называемые органеллами, для выполнения ряда клеточных функций, а их ДНК находится в центральном отделе, называемом ядром.)
Это более простые организмы без организованной клеточной структуры, и их ДНК свободно плавает в цитоплазме в виде запутанной массы. С другой стороны, эукариоты — это все другие живые организмы: протисты, растения, грибы. Что такое Протисты и животные. Их клетки более организованы. У них есть структуры, называемые органеллами, для выполнения ряда клеточных функций, а их ДНК находится в центральном отделе, называемом ядром.)
Общие характеристики
Среда обитания
Большинство водорослей живут в водной среде (Current Biology, 2014). Тем не менее, слово «водный» почти не может охватить разнообразие этих мест обитания. Эти организмы могут процветать в пресноводных озерах или в соленых океанах. Они также могут выдерживать различные температуры, концентрации кислорода или углекислого газа, кислотность и мутность. Например, гигантские водоросли встречаются на глубине более 200 метров под полярными ледяными щитами, согласно «Водорослям», в то время как одноклеточные виды зеленых водорослей 9Согласно обзорной статье 2005 года, опубликованной в журнале Saline Systems, 0023 Dunaliella salina встречается в очень соленой или гиперсоленой среде, такой как Мертвое море. Свободно плавающие, в основном одноклеточные водоросли, обитающие в освещенных областях воды, известны как планктонные. Те, которые прилипают к поверхности, известны как бентосные водоросли. Такие водоросли растут на иле, камнях, других водорослях и растениях или животных, согласно «Водорослям».
Свободно плавающие, в основном одноклеточные водоросли, обитающие в освещенных областях воды, известны как планктонные. Те, которые прилипают к поверхности, известны как бентосные водоросли. Такие водоросли растут на иле, камнях, других водорослях и растениях или животных, согласно «Водорослям».
Водоросли также способны выживать на суше. Некоторые неожиданные места, где они растут, — это стволы деревьев, мех животных, сугробы, горячие источники (согласно «Водорослям») и в почве, включая корки пустыни (Current Biology, 2014).
В основном водоросли живут независимо друг от друга в различных формах роста (отдельные клетки, колонии и т. д.), но они также могут вступать в симбиотические отношения с различными нефотосинтезирующими организмами, включая инфузорий, губок, моллюсков и грибов (таких как лишайники). Одним из преимуществ таких взаимоотношений является то, что они позволяют водорослям расширять горизонты своего обитания.
Питание
Как правило, водоросли способны к фотосинтезу и производят собственное питание, используя световую энергию солнца и углекислый газ для производства углеводов и кислорода.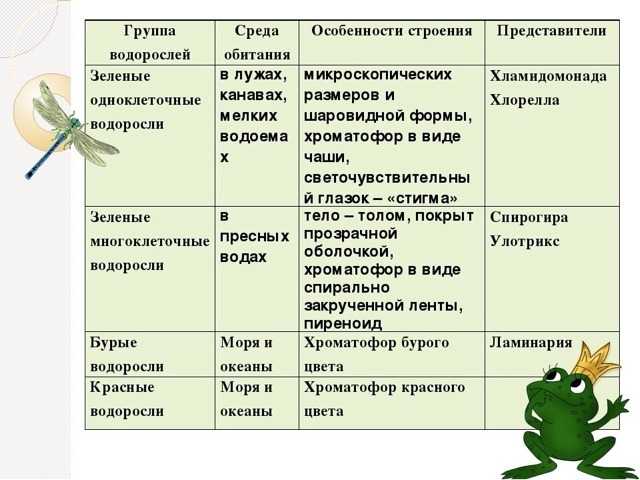 Другими словами, большинство водорослей являются автотрофами или, точнее, фотоавтотрофами (что отражает их использование энергии света для производства питательных веществ).
Другими словами, большинство водорослей являются автотрофами или, точнее, фотоавтотрофами (что отражает их использование энергии света для производства питательных веществ).
Однако существуют определенные виды водорослей, которым необходимо получать питание исключительно из внешних источников; то есть они гетеротрофны. Такие виды применяют различные гетеротрофные стратегии для получения питательных веществ из органических материалов (углеродсодержащих соединений, таких как углеводы, белки и жиры). Осмотрофия — это поглощение растворенных веществ, а фаготрофия включает поглощение бактерий или другой подобной добычи. Другим водорослям, известным как ауксотрофы, необходимо получать только основные витамины, такие как B 12 комплексные или жирные кислоты (согласно «Водоросли»).
Согласно авторам «Водорослей», широко признано, что пищевые стратегии водорослей существуют в спектре, сочетающем фотоавтотрофию и гетеротрофию. Эта способность известна как миксотрофия.
Размножение
Водоросли способны размножаться бесполым или вегетативным путем, а также половым путем.
По мнению авторов «Водорослей», бесполое размножение включает в себя производство подвижной споры, в то время как вегетативные способы включают простое деление клеток (митоз) для получения идентичного потомства и фрагментации колонии. Половое размножение включает объединение гамет (производящихся индивидуально у каждого родителя в результате мейоза).
Цветение водорослей в Северной Каролине, регионе страны, оборудованном для широкомасштабного роста водорослей. (Изображение предоставлено Ильдаром Сагдеевым через PNNL)
Классификация
Цианобактерии
Их также называют сине-зелеными водорослями. Хотя они способны проводить фотосинтез с выработкой кислорода и живут во многих из тех же сред, что и эукариотические водоросли, цианобактерии являются грамотрицательными бактериями и, следовательно, являются прокариотами.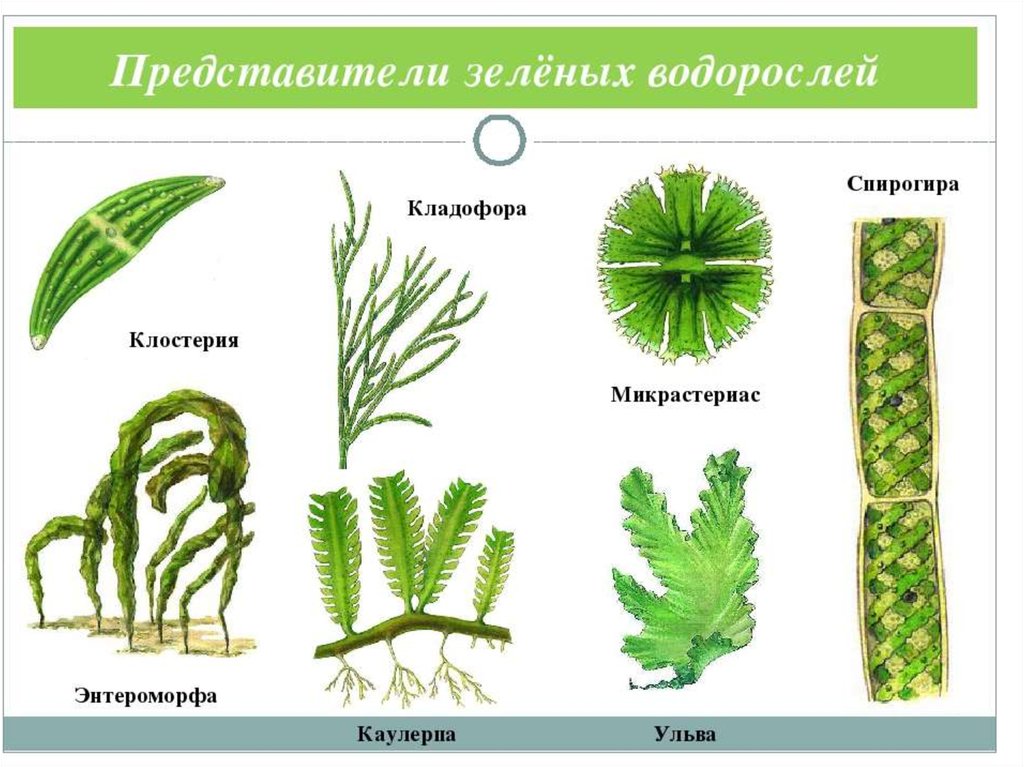 Они также способны самостоятельно проводить фиксацию азота, процесс преобразования атмосферного азота в пригодные для использования формы элемента, такие как аммиак.
Они также способны самостоятельно проводить фиксацию азота, процесс преобразования атмосферного азота в пригодные для использования формы элемента, такие как аммиак.
Приставка «циано» означает синий. Эти бактерии имеют пигменты, которые поглощают определенные длины волн света и придают им характерные цвета. Многие цианобактерии имеют синий пигмент фикоцианин, светособирающий пигмент (он поглощает красные длины волн света). Все цианобактерии имеют ту или иную форму зеленого пигмента хлорофилла, который отвечает за сбор энергии света в процессе фотосинтеза (Current Biology, 2014). Некоторые другие также имеют красный пигмент фикоэритрин, который поглощает свет зеленой областью и придает бактериям розовый или красный цвет.
Эукариотические водоросли
Эукариотические водоросли полифилетичны, что означает, что они не произошли от одного общего предка. Это ясно продемонстрировано в нашем нынешнем понимании древа жизни — генеалогического древа всех живых организмов, организованных их различными эволюционными отношениями.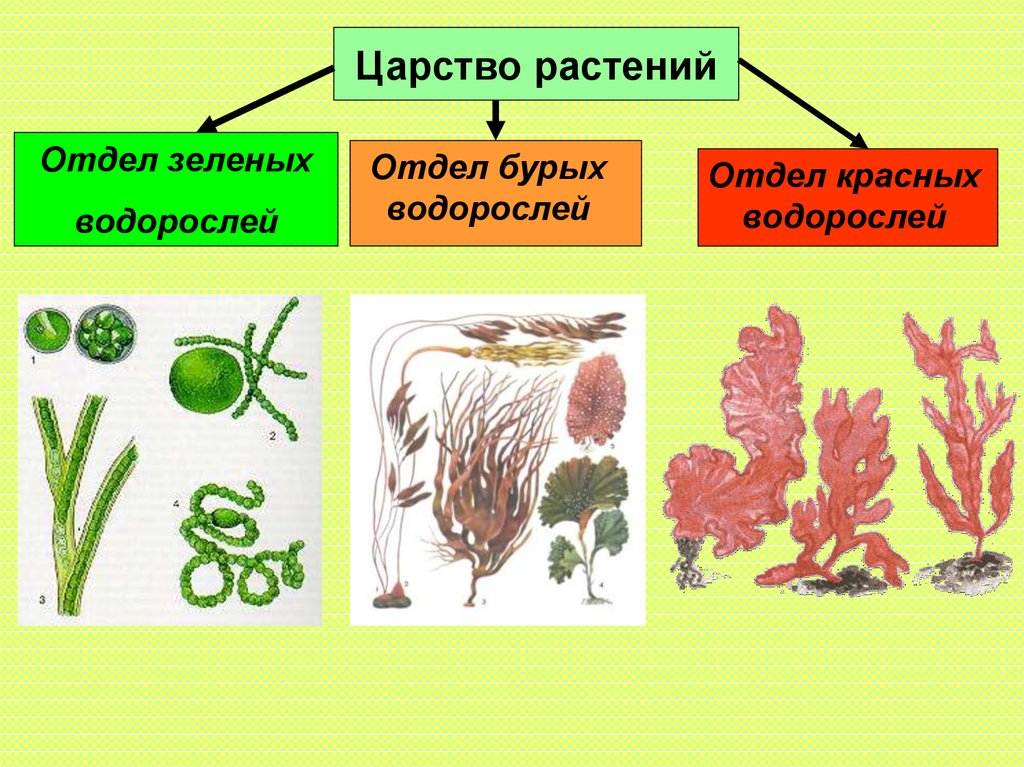 Эукариотические водоросли распространены среди многих различных групп или основных ветвей дерева.
Эукариотические водоросли распространены среди многих различных групп или основных ветвей дерева.
В обзорной статье 2014 года, опубликованной в журнале Cold Spring Harbour Perspectives in Biology, автор Фабьен Бурки перечисляет пять супергрупп эукариотических организмов: Ophiskontha, Amoebozoa, Excavata, Archaeplastida и SAR (в которую входят три группы: Stramenopiles, Alveolata и Rhizaria).
Archaeplastida включает растения и различные фотосинтезирующие виды водорослей, такие как хлорофиты (подгруппа зеленых водорослей), харофиты (главным образом пресноводные зеленые водоросли) и глаукоцистофиты (одноклеточные пресноводные водоросли). Хлорофиты — это зеленые водоросли, которые обычно образуют партнерские отношения лишайников с грибами.
Динофлагелляты встречаются в Альвеолатах. Это прежде всего одноклеточные морские и пресноводные организмы. Многие динофлагелляты утратили свои пластиды — место фотосинтеза — в ходе эволюции и стали фаготропными или живут как паразиты. Среди Alveolata, Excavata, Rhizaria и Chromista встречаются и другие виды водорослей (Current Biology, 2014).
Среди Alveolata, Excavata, Rhizaria и Chromista встречаются и другие виды водорослей (Current Biology, 2014).
Важность
Вероятно, наиболее важным вкладом водорослей в нашу окружающую среду и благополучие является выработка кислорода посредством фотосинтеза. «Водоросли незаменимы, потому что они производят примерно половину кислорода в атмосфере Земли», — сказал Грэм LiveScience.
Согласно обзорной статье 2010 года, опубликованной в журнале Biofuels, нефть частично получают из залежей древних водорослей. «Некоторые очень старые нефтяные месторождения приписываются цианобактериям, хотя личность производителей до сих пор неизвестна», — сказал Грэм. «Молодые залежи нефти, вероятно, возникли из эукариотических морских зеленых водорослей, кокколитофоридов и другого микроскопического морского фитопланктона». Эти залежи нефти являются ограниченным ресурсом и постепенно истощаются по мере использования человеком. В результате исследователи ищут возобновляемые альтернативы.
Биотопливо из водорослей является многообещающей заменой ископаемого топлива. Все водоросли обладают способностью производить масла, богатые энергией, а некоторые виды микроводорослей естественным образом накапливают высокие уровни масла в своей сухой массе. Более того, водоросли встречаются в разнообразных средах обитания и могут быстро размножаться. Они также эффективно используют углекислый газ. «Водоросли помогают поддерживать стабильный уровень углекислого газа в атмосфере, накапливая [газ] в органических материалах, включая месторождения нефти и неорганические карбонатные породы», — сказал Грэм. Зеленые водоросли, диатомовые водоросли и цианобактерии — это лишь некоторые из видов микроводорослей, которые считаются подходящими кандидатами для производства биотоплива (Biofuels, 2010).
Цветение водорослей
Водоросли в виде цветения водорослей имеют плохую репутацию за создание токсичных условий в океанах и озерах. По данным Агентства по охране окружающей среды (EPA), «цветение водорослей» относится к безудержному росту определенных микроводорослей, что, в свою очередь, приводит к выработке токсинов, нарушению естественных водных экосистем и увеличению затрат на очистку воды. Цветы приобретают цвет содержащихся в них водорослей. Грэм утверждает, что основными производителями токсинов в океанах являются некоторые динофлагелляты и диатомовые водоросли. В пресных водах цианобактерии являются основными производителями токсинов, хотя некоторые эукариотические водоросли также вызывают проблемы. Грэм отмечает, что в естественных условиях водоросли используют токсины, чтобы защитить себя от поедания мелкими животными, и им нужно лишь небольшое количество, чтобы защитить себя.
Цветы приобретают цвет содержащихся в них водорослей. Грэм утверждает, что основными производителями токсинов в океанах являются некоторые динофлагелляты и диатомовые водоросли. В пресных водах цианобактерии являются основными производителями токсинов, хотя некоторые эукариотические водоросли также вызывают проблемы. Грэм отмечает, что в естественных условиях водоросли используют токсины, чтобы защитить себя от поедания мелкими животными, и им нужно лишь небольшое количество, чтобы защитить себя.
Основной причиной цветения водорослей является явление, называемое загрязнением питательными веществами. При загрязнении питательными веществами возникает избыток азота и фосфора, что может подтолкнуть водоросли к безудержному росту. Это явление вызвано разнообразной деятельностью человека. По данным Агентства по охране окружающей среды, удобрения, которые мы используем в сельском хозяйстве, и навоз животных богаты азотом, в то время как неправильно очищенные сточные воды содержат много азота и фосфора.
«В обществе распространено мнение, что водоросли вредны и должны быть устранены при каждой возможности. Но это мнение ошибочно, потому что водоросли производят кислород, рыбу [они являются основным источником пищи для водных организмов], масло и многие другие полезные материалы», — сказал Грэм LiveScience. «Только несколько видов вызывают проблемы, и худший из них — 9.0023 Homo sapiens .»
Дополнительные ресурсы
Апарна Видьясагар — независимый научный журналист, специализирующийся на здравоохранении и науках о жизни. Апарна написал для ряда изданий, в том числе New Scientist, Science, PBS SoCal, Mental Floss и некоторых других. Апарна имеет докторскую степень в области клеточной и молекулярной патологии Университета Висконсин-Мэдисон, а также степень магистра и бакалавра в том же университете.
Водоросли как перспективные организмы для окружающей среды и здоровья
1. Эль-Бароти Г.С., Мусса М.Ю., Шаллан М.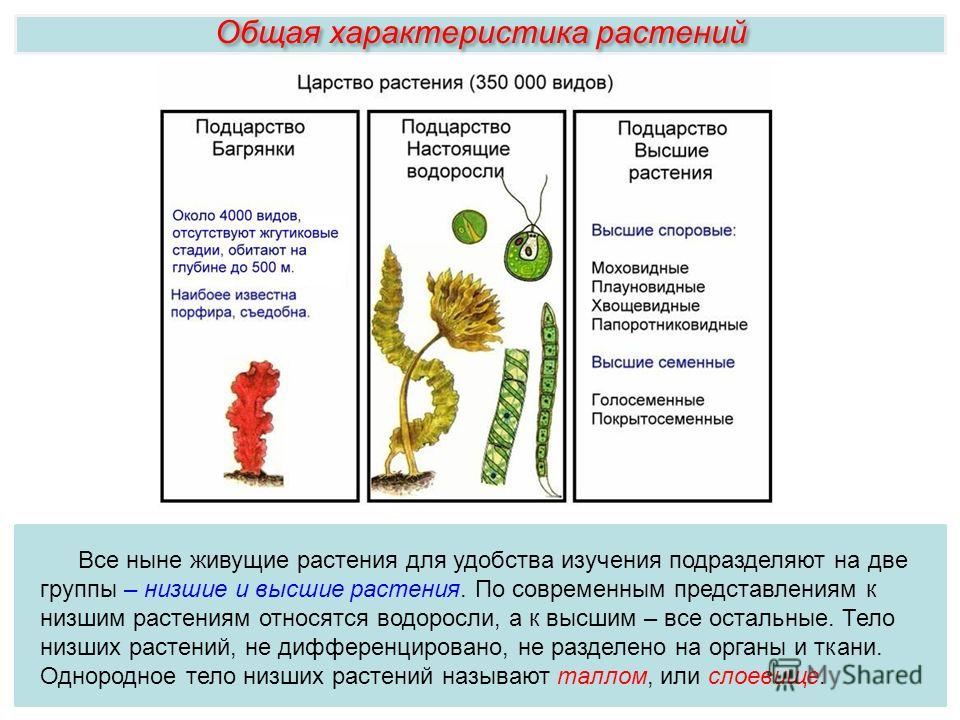 А., Али М.А., Сабх А.З., Шалаби Э.А. Вклад в содержание аромата, биологической активности, минералов, белков, пигментов и липидов красных водорослей: Asparagopsis taxformis (Delile) Trevisan. J App Sci Res. 2007; 3: 1825–1834. [Google Scholar]
А., Али М.А., Сабх А.З., Шалаби Э.А. Вклад в содержание аромата, биологической активности, минералов, белков, пигментов и липидов красных водорослей: Asparagopsis taxformis (Delile) Trevisan. J App Sci Res. 2007; 3: 1825–1834. [Google Scholar]
2. Lee-Saung H, Lee-Yeon S, Jung-Sang H, Kang-Sam S, Shin-Kuk H. Антиоксидантная активность фукостерола из морских водорослей Пельвеция кремнистая . Арч Фарм Рез. 2003; 26: 719–722. [PubMed] [Google Scholar]
3. Shalaby EA, Shanab SMM, Singh V. Повышение антиоксидантной и противовирусной эффективности Spirulina platensis при солевом стрессе. J Med Plants Res. 2010;4:2622–2632. [Google Scholar]
4. Шалабы ЕАА. Магистр Тезис. Кафедра биохимии сельскохозяйственного факультета Каирского университета; 2004. Химические и биологические исследования видов спирулины. [Google Scholar]
5. Шалабы ЕАА. Кандидат наук. Тезис. Кафедра биохимии сельскохозяйственного факультета Каирского университета; 2008.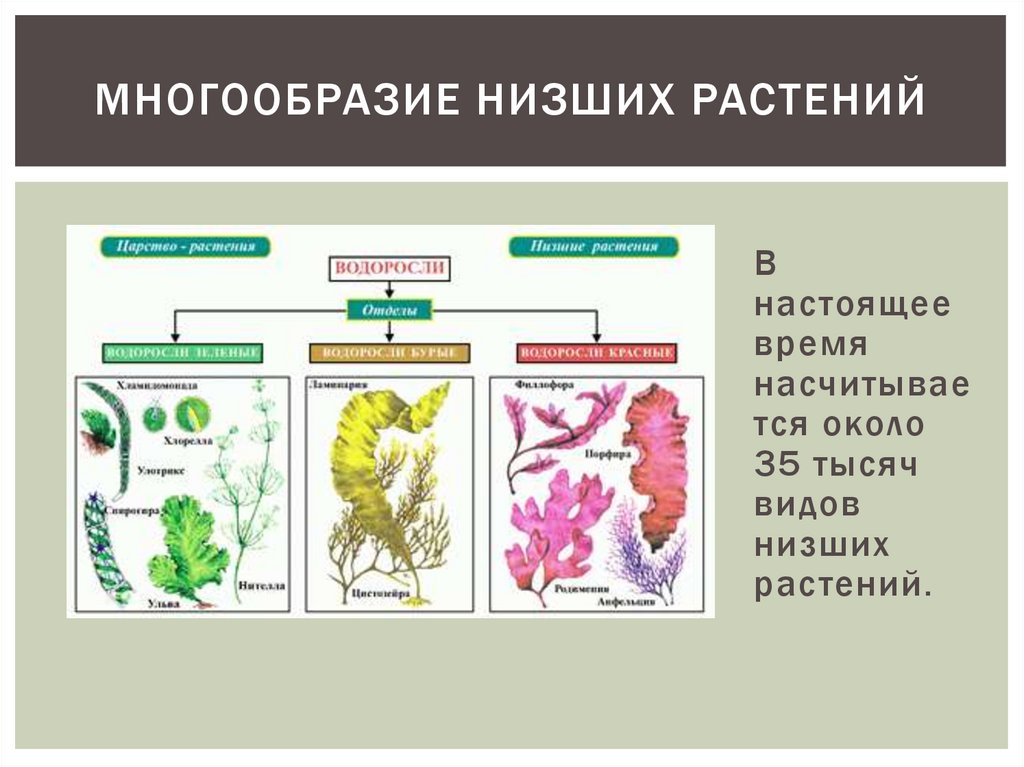 Биохимические и биотехнологические исследования некоторых морских водорослей. [Академия Google]
Биохимические и биотехнологические исследования некоторых морских водорослей. [Академия Google]
6. Абу Элалла Ф.М., Шалаби Э.А. Антиоксидантная активность экстракта и полуочищенных фракций морской красной макроводоросли, Gracilaria Verrucosa . Aust J Bas App Sci. 2009;3:3179–3185. [Google Scholar]
7. Shanab SM, Shalaby EA, El-Fayoumy E. Enteromorpha Compressa проявляет мощную антиоксидантную активность. Дж. Биомед Биотех. 2011:72640. [Бесплатная статья PMC] [PubMed] [Google Scholar]
8. Hehmann A, Kaya K, Watanabe MM. Селективный контроль Microcystis sp с помощью лабораторного анализа аминокислот. J Appl Phycol. 2002; 14:85–89. [Google Scholar]
9. Burja AM, Banaigs B, Abou-Mansour E, Burguess JG, Wright PC. Морские цианобактерии — ценный источник натуральных продуктов. Тетраэдр. 2001; 57: 9347–9377. [Google Scholar]
10. Бекер Э.В. Микроводоросли. Кембридж, Нью-Йорк: Издательство Кембриджского университета; 1994. [Google Scholar]
11. Blunt JW, Copp BR, Munro MHG, Northcote PT, Prinsep MR. Морские натуральные продукты. Nat Prod Rep. 2005; 22:15–61. [PubMed] [Google Scholar]
Blunt JW, Copp BR, Munro MHG, Northcote PT, Prinsep MR. Морские натуральные продукты. Nat Prod Rep. 2005; 22:15–61. [PubMed] [Google Scholar]
12. Puglisi MP, Tan LT, Jensen PR, Fenical W. Капистероны A и B из тропических зеленых водорослей Penicillus capitatus : неожиданная противогрибковая защита, нацеленная на морской патоген Lindra thallasiae . Тетраэдр. 2004; 60: 7035–7039. [Google Scholar]
13. Barros MP, Pinto E, Sigaud-Kutner TCS, Cardozo KHM, Colepicolo P. Ритмичность и окислительный/нитрозативный стресс у водорослей. Биол Ритм Рез. 2005; 36: 67–82. [Google Scholar]
14. Mayer AMS, Hamann MT. Морская фармакология в 2000 г.: морские соединения с антибактериальной, антикоагулянтной, противогрибковой, противовоспалительной, противомалярийной, антитромбоцитарной, противотуберкулезной и противовирусной активностью; влияя на сердечно-сосудистую, иммунную и нервную системы и другие разнообразные механизмы действия. Мар Биотехнолог. 2004; 6: 37–52. [Бесплатная статья PMC] [PubMed] [Google Scholar]
[Бесплатная статья PMC] [PubMed] [Google Scholar]
15. Дос Сантос, доктор медицинских наук, Гуаратини Т., Лопес JLC, Колепиколо П., Лопес Н.П. Культура растительных клеток и микроводорослей // Современная биотехнология в медицинской химии и промышленности. Керала, Индия: исследовательский указатель; 2005. [Google Scholar]
16. Макдермид К., Стюерке Б. Пищевой состав съедобных гавайских водорослей. J Appl Phycol. 2003; 15: 513–524. [Google Scholar]
17. Маршам С., Скотт Г.В., Тобин М.Л. Сравнение питательной химии ряда водорослей умеренного пояса. Пищевая хим. 2007; 100:1331–1336. [Академия Google]
18. Plaza M, Cifuentes A, Ibañez E. В поисках новых функциональных пищевых ингредиентов из водорослей. Технология пищевой науки. 2008; 19:31–39. [Google Scholar]
19. Гордон М.Х., Магос П. Влияние стеролов на окисление пищевых масел. Пищевая хим. 1983; 10:141. [Google Scholar]
20. Анггадиреджа Дж., Андьяни Р., Хаяти М. Антиоксидантная активность Sargassum polycystum (Phaeeophyta) и Laurencia obtus (Rhodophyta) с острова Серибу. J Appl Phycol. 1997;9:477–479. [Google Scholar]
J Appl Phycol. 1997;9:477–479. [Google Scholar]
21. Мацукава Р., Дубинаски З., Кисимото Э., Масаки К., Масуда Ю., Такеучи Т. и др. Сравнение методов скрининга антиоксидантной активности морских водорослей. J Appl Phycol. 1997; 9: 29–35. [Google Scholar]
22. Jia-Zhi S, Wu-Jian M, Zheng-Hai L, Jiang, Yin B, Zhu-Hong Y. Выделение и анализ состава фукозосодержащих сульфатированных полисахаридов из бурых водорослей Sargassum fusiformis Сетч Дж. Чжэцзянский сельскохозяйственный университет. 1998:670–673. [Google Scholar]
23. Weng XC, Wang W. Антиоксидантная активность соединений, выделенных из Sal через plebeian . Пищевая хим. 2000; 71: 489–493. [Google Scholar]
24. Новоа В., Мотидом М., Манчини-Фильо Дж., Линарес А.Ф., Танае М.М., Торрес Л.М. и соавт. Антиоксидантная активность, связанная с фенолокислотами, в водном экстракте морских водорослей Bryothamnion triquetrum (Gmelim S.G.) Howe. Revista-Brasilerra-deCiencias-Farmaceuticas.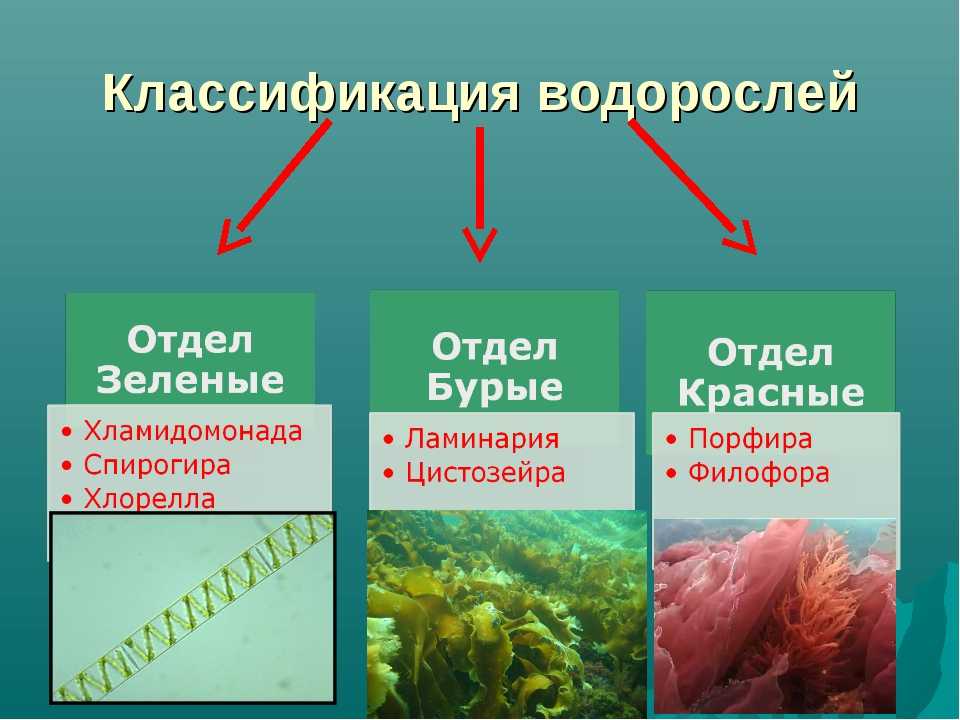 2001: 373–382. [Академия Google]
2001: 373–382. [Академия Google]
25. Ruberto G, Baratta MT, Biondi DM, Amico V. Антиоксидантная активность экстрактов морских водорослей рода Cystoseria в мицеллярной модельной системе. J Appl Phycol. 2001; 13: 403–407. [Google Scholar]
26. Хименес-Эскриг А., Хименес-Хименес И., Пулидо Р., Саура-Каликсто Ф. Антиоксидантная активность свежих и обработанных пищевых водорослей. J Наука о продуктах питания и сельском хозяйстве. 2001: 530–534. [Google Scholar]
27. Нагаи Т., Укимото Т. Приготовление и функциональные свойства напитков из морских водорослей. Пищевая хим. 2003; 81: 327–332. [Академия Google]
28. Cahyana AH, Shuto Y, Kinoshita Y. Пирофеофитин как антиоксидант из морских водорослей, Arame ( Eicenia bicyclis ) Biosci Biotechnol Agrochem. 1992; 56: 1533–1535. [Google Scholar]
29. Yan XJ, Chuda Y, Suzuki M, Nagata T. Фукоксантин как основной антиоксидант в Hijikia fusiformis . Биоски Биотехнолог Агрохим. 1999; 63: 605–607.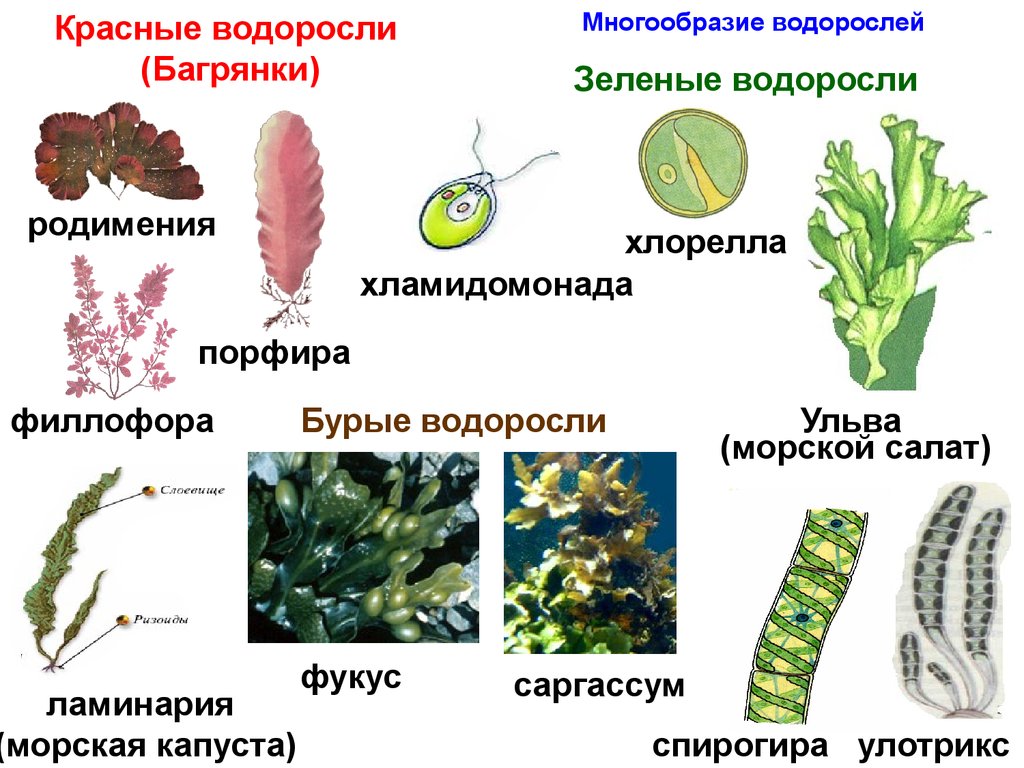 [PubMed] [Google Scholar]
[PubMed] [Google Scholar]
30. Mori J, Matsunaga T, Takahashi S, Hasegawa C, Saito H. Ингибирующая активность экстрактов морских бурых водорослей в отношении перекисного окисления липидов. Фитотер Рез. 2003;17:549–551. [PubMed] [Google Scholar]
31. Раймундо М.С., Орта П., Фетт Р. Антиоксидантная активность in vitro экстрактов некоторых зеленых водорослей (Chlorophyta) с южного побережья Бразилии. Revista-Brasileira-de-Ciencias-Farmaceuticas. 2004;40:495–503. [Google Scholar]
32. Maehira F, Motomura K, Nashiro T, Mekaru N, Sudo Y. Влияние красных водорослей, культивируемых в глубоководной воде, на окислительно-восстановительный статус метаболизма печени, легких, мозга и костей в SAMP1. и САМР1. Серия международных конгрессов. 2004; 1260:413–416. [Академия Google]
33. Heo S, Park E, Lee K, Jeon Y. Антиоксидантная активность ферментативных экстрактов бурых водорослей. Биоресурсная технология. 2005; 96: 1613–1623. [PubMed] [Google Scholar]
34. Jang-Kyoung H, Lee-Bong H, Choi-Byoung W, Lee-Hyi S, Shin-Jong H.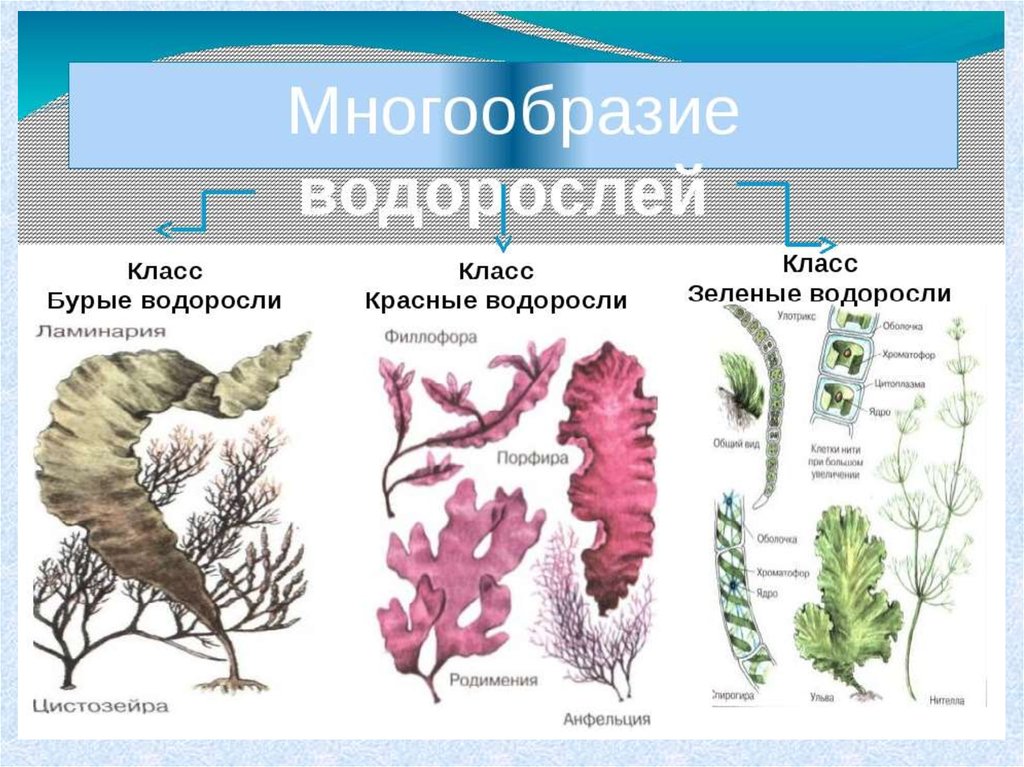 Chromenes из бурой водоросли Sargassum siliquastrum . Ж Натуральные продукты. 2005; 68: 719–723. [PubMed] [Google Scholar]
Chromenes из бурой водоросли Sargassum siliquastrum . Ж Натуральные продукты. 2005; 68: 719–723. [PubMed] [Google Scholar]
35. Нахас Р., Абатис Д., Анагностопулу М.А., Кефалас П., Вагиас С., Руссис В. Активность морских водорослей Эгейского моря по удалению радикалов. Пищевая хим. 2007; 102: 577–581. [Академия Google]
36. Чу Ю.Л., Лим Ю.Ю., Омар М., Ху К.С. Антиоксидантная активность трех съедобных водорослей из двух районов Юго-Восточной Азии. LWT. 2008;41:1067–1072. [Google Scholar]
37. Ганесан П., Кумар С.С., Бхаскар Н. Антиоксидантные свойства метанольного экстракта и его растворяющих фракций, полученных из выбранных индийских красных водорослей. Биоресурсная технология. 2008; 99: 2717–2723. [PubMed] [Google Scholar]
38. Ohigashi H, Sakai Y, Yamaguchi K, Umezaki I, Koshimizu K. Возможные противоопухолевые свойства морских водорослей и активность экстракта водорослей Wakama in vivo. Биоски Биотехнолог Биохим. 1992;56:994–995. [PubMed] [Google Scholar]
39. Рашид М.А., Густафсон К.Р., Карделлина Дж.Х.Х. Бромированный хамиграновый сесквитерпен дает новый профиль дифференцированной цитотоксичности в скрининге NCI in vitro. Нат Прод Летт. 1995; 6: 255–259. [Google Scholar]
Рашид М.А., Густафсон К.Р., Карделлина Дж.Х.Х. Бромированный хамиграновый сесквитерпен дает новый профиль дифференцированной цитотоксичности в скрининге NCI in vitro. Нат Прод Летт. 1995; 6: 255–259. [Google Scholar]
40. Harada H, Yamashita U, Kurihara H, Fukushi E, Kawabata J, Kamei Y. Противоопухолевое действие пальмитиновой кислоты, обнаруженной как селективное цитотоксическое вещество в морских красных водорослях. Антиканк Рез. 2002; 22: 2587–2590. [PubMed] [Академия Google]
41. Deslandes E, Pondaven P, Auperin T, Rossakis C, Guézennec J, Stiger V, et al. Предварительное исследование in vitro антипролиферативного действия гидроэтанольного экстракта субтропических водорослей Turbinaria ornata (Turner J. Agardh) на немелкоклеточную бронхолегочную карциному человека (NSCLC-N6) J Appl Phycol. 2000; 12: 257–262. [Google Scholar]
42. Сахара Х., Ханашима С., Ямадзаки Т., Такахаши С., Охтани С., Исикава М. и др. Противоопухолевый эффект химически синтезированных сульфолипидов на основе Sea Urchin’s натуральные сульфонохиновозилмоноацилглицеролы. Jpn J Рак Res. 2002; 93: 85–92. [Бесплатная статья PMC] [PubMed] [Google Scholar]
Jpn J Рак Res. 2002; 93: 85–92. [Бесплатная статья PMC] [PubMed] [Google Scholar]
43. Xu N, Fan X, Yan X, Tseng CK. Скрининг морских водорослей из Китая на их противоопухолевую активность. J Appl Phycol. 2004; 16: 451–456. [Google Scholar]
44. Юань Ю.В., Уолш Н.А. Антиоксидантная и антипролиферативная активность экстрактов различных съедобных водорослей. Пищевая химическая токсикол. 2006;44:1144–1150. [PubMed] [Академия Google]
45. Квон М.Дж., Нам Т.Дж. Порфиран индуцирует связанный с апоптозом сигнальный путь в клеточных линиях рака желудка AGS. Жизнь наук. 2006; 79: 1956–1962. [PubMed] [Google Scholar]
46. Kwon H, Bae S, Kim K, Han C, Cho S, Nam S, et al. Индукция апоптоза в клетках HeLa спиртовым экстрактом Corallina pilulifera . Пищевая хим. 2007; 1: 196–201. [Google Scholar]
47. Дар А., Байг Х.С., Сайфуллах С.М., Ахмад В.У., Ясмин С., Низамуддин М. Влияние сезонных колебаний на противовоспалительную активность Sargassum wightii , растущий на побережье Северного Аравийского моря в Пакистане J.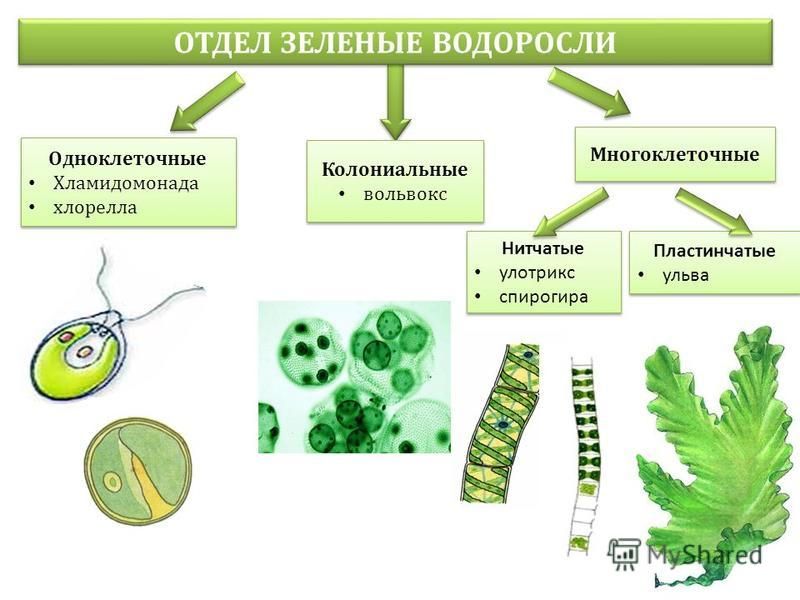 Exp Mar Biol Ecol. 2007; 351:1–9. [Google Scholar]
Exp Mar Biol Ecol. 2007; 351:1–9. [Google Scholar]
48. Kang JY, Khan MNA, Park NH, Cho JY, Lee MC, Fujii H и др. Жаропонижающее, обезболивающее и противовоспалительное действие морских водорослей Sargassum fulvellum и Sargassum thunbergii у мышей. J Этнофармакол. 2008; 116: 187–190. [PubMed] [Google Scholar]
49. Hehmann A, Kaya K, Watanabe MM. Селективный контроль Microcystis sp с помощью лабораторного анализа аминокислот. J Appl Phycol. 2002; 14:85–89. [Google Scholar]
50. Schrader KK, de Regt MQ, Tidwell PD, Tucker CS, Duke SO. Селективное ингибирование роста цианобактерий Oscillatoria cf., продуцирующих затхлый запах. chalybea природными соединениями. Булл Энвайрон. Контам Токсикол. 1998; 60: 651–658. [PubMed] [Google Scholar]
51. Чон С.Ю., Исида К., Ито Ю., Окада С., Мураками М. Бациламид, новый альгицид из морских бактерий, Bacillus sp. SY-1 против вредоносных динофлагеллят, Cochlodinium polykrikoides . Тетраэдр Летт.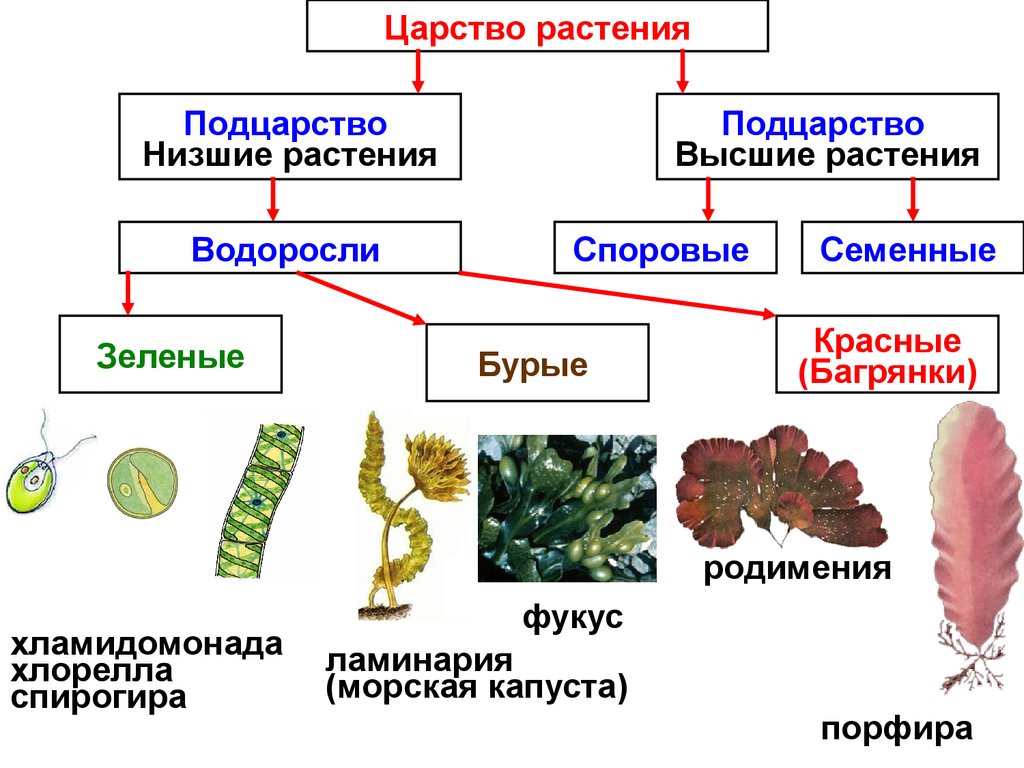 2003; 44:8005–8007. [Академия Google]
2003; 44:8005–8007. [Академия Google]
52. Папке У., Гросс Э.М., Франке В. Выделение, идентификация и определение абсолютной конфигурации Fischerellin B. Новый альгицид из пресноводных цианобактерий Fischerella muscicola (Thuret) Tetrahedron Lett. 1997; 38: 379–382. [Google Scholar]
53. Etchegaray A, Rabello E, Dieckmann R, Moon DH, Fiore MF, Dohren H, et al. Производство альгицидов нитчатыми цианобактериями Fischerella sp. CENA 19. J Appl Phycol. 2004; 16: 237–243. [Академия Google]
54. Duke SO, Dayan FE, Rimando AM, Schrader KK, Aliotta G, Oliva A, et al. Химические вещества от природы для борьбы с сорняками. Сорняк наук. 2002; 50: 138–151. [Google Scholar]
55. Мипагала К.М., Шрадер К.К., Ведж Д.Э., Дюк С.О. Альгицидные и противогрибковые соединения из корней Rutagraveolens и синтез их аналогов. Фитохимия. 2005; 66: 2689–2695. [PubMed] [Google Scholar]
56. Schrader KK, de Regt MQ, Tidwell PD, Tucker CS, Duke SO. Селективное ингибирование роста цианобактерий Oscillatoria cf. , продуцирующих затхлый запах. chalybea природными соединениями. Bull Environ Contam Toxicol. 1998;60:651–658. [PubMed] [Google Scholar]
, продуцирующих затхлый запах. chalybea природными соединениями. Bull Environ Contam Toxicol. 1998;60:651–658. [PubMed] [Google Scholar]
57. Kim JS, Hong KS, Lee BH, Kim JC, Cho KW. Гербицидная активность Polygonatum spp. и их экстракты против ряски, пресноводных водорослей и некоторых рисовых сорняков. Кор Дж. Сорняк Sci. 2005; 25:103–111. [Google Scholar]
58. Ким Дж. С., Ким Дж., Ли С., Ли Б., Чо К.Ю. Биологическая активность L-2-азетидинкарбоновой кислоты, выделенной из Polygonatum odoratum var. pluriflorum против некоторых водорослей. Аква Бот. 2006; 85: 1–6. [Академия Google]
59. Wang H, Liu Y, Gao X, Carter CL, Liu Z. Рекомбинантная b-субъединица C-фикоцианина ингибирует пролиферацию клеток и индуцирует апоптоз. Рак Летт. 2007; 247:150–158. [PubMed] [Google Scholar]
60. Nan C, Zhang H, Lin S, Zhao G, Liu X. Аллелопатические эффекты Ulva lactuca на отдельные виды вредных цветущих микроводорослей в лабораторных культурах.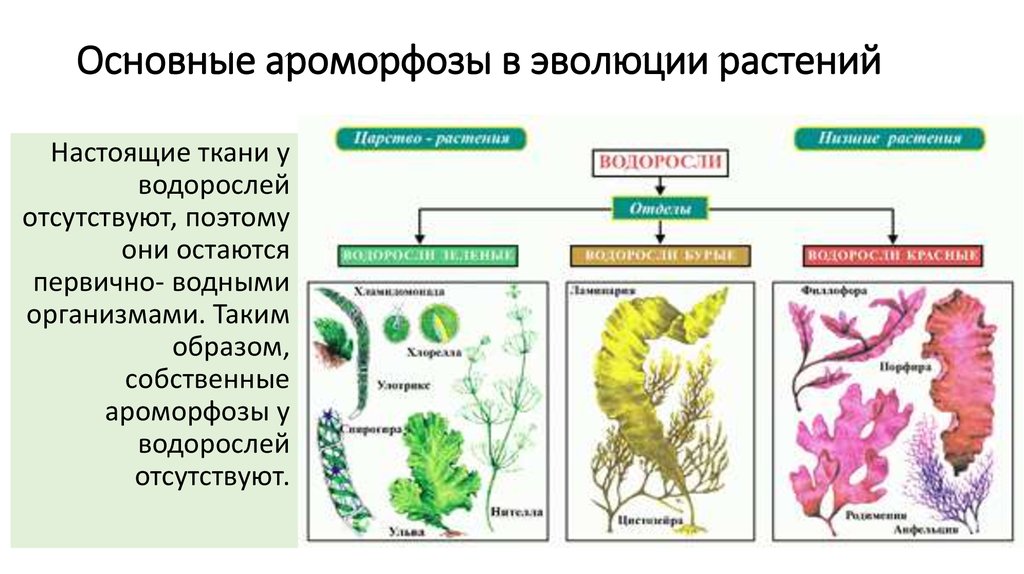 Аква Бот. 2008; 89: 9–15. [Google Scholar]
Аква Бот. 2008; 89: 9–15. [Google Scholar]
61. Гонсалес М., Аларкон Б., Карраско Л. Полисахариды как противовирусные агенты: противовирусный каррагенан. Антимикроб. Агенты Чемотер. 1987;31:1388–1393. [PMC free article] [PubMed] [Google Scholar]
62. Иванова В., Русева Р., Коларова М., Серкеджиева Дж., Рачев Р., Манолова Н. Выделение полисахарида с противовирусным действием из Ulva lactuca . Препарат Биохим. 1991; 24:83–97. [PubMed] [Google Scholar]
63. Witvrouw M, De Clercq E. Сульфатированные полисахариды, извлеченные из морских водорослей, как потенциальные противовирусные препараты. Генерал Фармак. 1997; 29: 497–511. [PubMed] [Google Scholar]
64. Carlucci MJ, Pujol CA, Ciancia M, Noseda MD, Matulewiez MC, Damonte EB, et al. Противогерпетические и антикоагулянтные свойства каррагинанов из красных водорослей Gigartina skottsbergii и их циклизированные производные: корреляция между структурой и биологической активностью. Int J Биол Макромоль. 1997; 20:97–105. [PubMed] [Google Scholar]
1997; 20:97–105. [PubMed] [Google Scholar]
65. Nowotny N, Bascunana CR, Ballagi-Pordany A, Gavier-Widen D, Uhlen M, Betak S. Филогенетический анализ геморрагической болезни кроликов и вирусов синдрома европейского бурого зайца путем сравнения последовательности гена капсидного белка. Арх Вирол. 1997; 142: 657–673. [PubMed] [Академия Google]
68. Бандара Б.М.Р., Гунатилака А.А.Л., Кумер Н.С., Вималасири В.Р. Антимикробная активность некоторых морских водорослей Шри-Ланки Дж. Национальный научный совет Шри-Ланки. 1988; 16: 209–221. [Google Scholar]
69. Perez GRM, Avila AJG, Perez GS, Martinez CA, Martinez CG. Антимикробная активность некоторых американских водорослей. J Этнофармакол. 1990; 29: 111–116. [PubMed] [Google Scholar]
70. Месмар М.Н., Абуссо М. Антибиотическая активность некоторых водных растений и экстрактов водорослей из Иордании. Qater University Science J. 1991;11:155–160. [Google Scholar]
71. Febles CI, Arias A, Hardisson A, Sierra-Lopez A, Gil-Rodriguez MC. Антимикробная активность экстрактов некоторых канареечных видов Phaeophyta и Chlorophyta. Фитотер Рез. 1995; 9: 385–387. [Google Scholar]
Антимикробная активность экстрактов некоторых канареечных видов Phaeophyta и Chlorophyta. Фитотер Рез. 1995; 9: 385–387. [Google Scholar]
72. Крауч И. Дж., Стаден В. Влияние концентратов морских водорослей на приживаемость и урожайность тепличных растений томатов. J Appl Phycol. 1992; 4: 291–296. [Google Scholar]
73. Zaccro MC, Salazer C, Caire Z, Cans S, Stella AM. Токсичность свинца в метаболизме порфиринов цианобактерий. Окружающая среда. Токсикольная вода Qual. 2001; 16:61–67. [PubMed] [Академия Google]
74. Iwamoto C, Minoura K, Oka T, Ohta T, Hagishita S, Numata A. Абсолютные стереоструктуры новых цитотоксических метаболитов, пеностатинов AE, из видов Penicillium, выделенных из водорослей Enteromorpha. Тетраэдр. 1999;55:14353–14368. [Google Scholar]
75. Киркпатрик Б., Флеминг Л.Э., Сквиччарини Д., Бакер Л.С., Кларк Р., Абрахам В. и др. Обзор литературы о красных приливах во Флориде: последствия для здоровья человека. Вредные водоросли. 2004; 3: 99–115.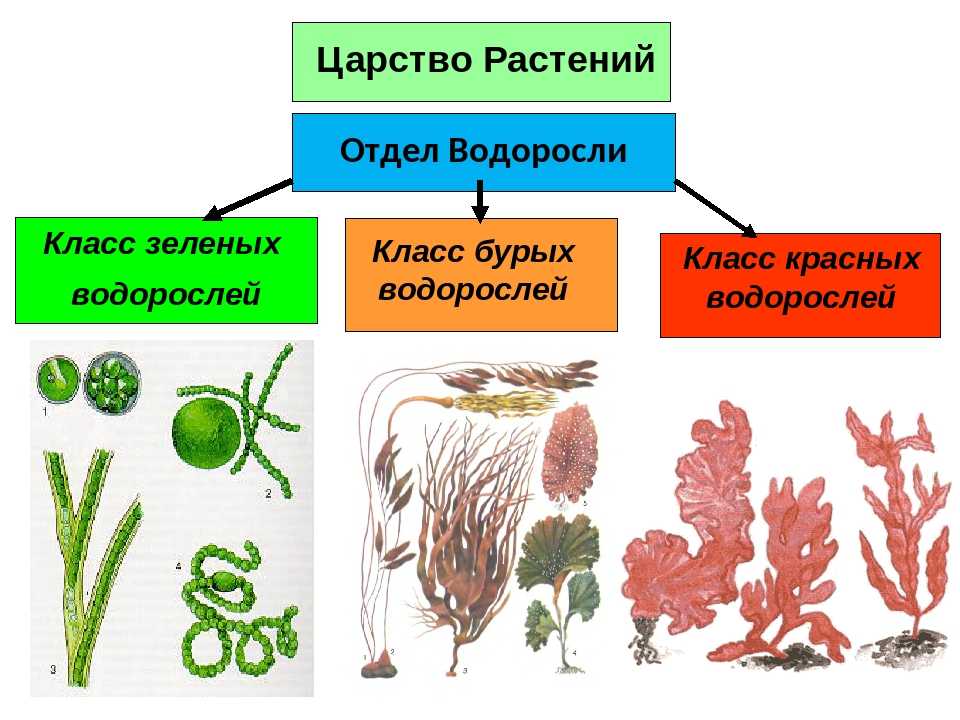 [Бесплатная статья PMC] [PubMed] [Google Scholar]
[Бесплатная статья PMC] [PubMed] [Google Scholar]
76. Кодд Г.А., Моррисон Л.Ф., Меткалф Дж.С. Цианобактериальные токсины: управление рисками для охраны здоровья. Toxicol Appl Pharmacol. 2005; 203: 264–272. [PubMed] [Google Scholar]
77. Bittencourt-Oliveira MC, Kujbida P, Cardozo KHM, Carvalho VM, Moura AN, Colepicolo P, et al. Новый ритм биосинтеза микроцистина описан у цианобактерии Microcystis panniformis . Biochem Biophys Res Commun. 2005; 326: 687–694. [PubMed] [Google Scholar]
78. Cazenave J, Wunderlin DA, Bistoni MDL, Ame MV, Krause E, Pflugmacher S, et al. Поглощение, распределение в тканях и накопление микроцистина-RR в Corydoras paeatus, Jenynsiamultidentata и Odontesthesbonariensis полевые и лабораторные исследования. Аква токсикол. 2005; 75: 178–190. [PubMed] [Google Scholar]
79. Kujbida P, Hatanaka E, Campa A, Colepicolo P, Pinto E. Влияние микроцистинов на полиморфноядерные лейкоциты человека. Biochem Biophys Res Commun.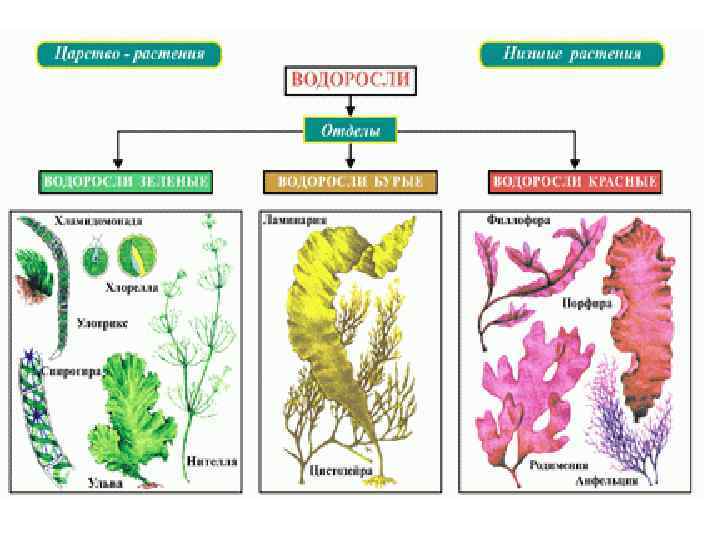 2006; 341: 273–277. [PubMed] [Google Scholar]
2006; 341: 273–277. [PubMed] [Google Scholar]
80. Denance M, Guyot M, Samadi M. Краткий синтез 16-гидрокси-5-холестан-3,-диона, нового цитотоксического морского оксистерола. Стероиды. 2006;71:599–602. [PubMed] [Google Scholar]
81. Awadalla FT, Pesic B. Биосорбция кобальта с помощью реагента для удаления металлов AMTTm. гидрометаллургия. 1992; 28: 65–80. [Google Scholar]
82. Холл Дж.Л. Клеточные механизмы детоксикации тяжелых металлов и толерантности. J Эксп. Бот. 2002; 53:1–11. [PubMed] [Google Scholar]
83. Ferrat L, Pergent-Martini C, Romeo M. Оценка использования биомаркеров в водных растениях для оценки качества окружающей среды: применение к морским водорослям. Аква токсикол. 2003; 65: 187–204. [PubMed] [Академия Google]
84. Moenne A. Эукариотические металлотионеины: структура белка, регуляция генов и гомеостаз меди. Cahiers Biologie Marine. 2001;42:125–135. [Google Scholar]
85. Клеменс С. Накопление токсичных металлов, реакция на воздействие и механизмы толерантности у растений. Биохимический. 2006; 88: 1707–1719. [PubMed] [Google Scholar]
Биохимический. 2006; 88: 1707–1719. [PubMed] [Google Scholar]
86. García-Ríos V, Freile-Pelegrin Y, Robledo D, Mendoza-Cózatl D, Moreno-Sánchez R, Gold-Bouchot G. Состав клеточной стенки влияет на Cd 2+ Накопление и внутриклеточные тиоловые пептиды у морских красных водорослей. Водная токсикология. 2007; 81: 166–170. [PubMed] [Google Scholar]
87. Руперес П. Содержание минералов в съедобных морских водорослях. Пищевая химия. 2002; 79: 23–26. [Google Scholar]
88. Cardozo KHM, Guaratini T, Barros MP, Falcao VR, Tonon AP, Lopes NP, et al. Метаболизм водорослей с экономическим эффектом. Сравнительная биохимия и физиология. 2006; 146: 60–78. [PubMed] [Google Scholar]
89. Росс А.Б., Джонс Дж.М., Кубаки М.Л., Бриджмен Л. Классификация макроводорослей как топлива и их термохимическое поведение. Биоресурсная технология. 2008;99:6494–6504. [PubMed] [Google Scholar]
90. Gerwick WH, Roberts MA, Proteau PJ, Chen J. Скрининг культивируемых морских микроводорослей на противораковую активность.