
Стебли древесных двудольных и хвойных растений. Срез хвойного растения
Строение стебля древесных двудольных и хвойных растений.
Стебли двудольных древесных растений (яблони, липы, дуба и др.) имеют типичное непучковое строение. Если у трав все клетки камбия к осени превращаются в клетки постоянных тканей, то в древесных стеблях камбий функционирует в течение всей жизни.
Массовый транспорт веществ в стволе идет по молодым слоям луба и древесины, расположенным около камбия. Луб теряет эту способность к проведению веществ обычно через год (после перезимовки), древесина служит дольше — несколько лет. На смену стареющим тканям камбий откладывает молодые деятельные слои.
Основная масса ствола состоит из мертвых клеток и не принимает непосредственного участия в транспорте веществ: нефункционирующая древесина поддерживает тяжесть кроны, а мертвые ткани коры защищают внутренние живые ткани.
Рассмотрим строение ствола дерева на примере липы. Образующийся из почки в процессе весеннего роста побег липы покрыт эпидермой. На поперечном срезе его заметны сердцевина с примыкающей к ней первичной ксилемой и очень рано возникающие сплошные слои вторичной ксилемы, камбия, флоэмы и первичной коры. Однако уже летом под эпидермой закладывается пробковый камбий — феллоген и формируется перидерма. Под некоторыми устьицами образуются чечевички. С образованием перидермы, заканчивающимся к осени, клетки эпидермы отмирают. Остатки их сохраняются на поверхности стебля несколько лет. Под перидермой у дву - трехлетней ветви липы находятся кора (первичная и вторичная), камбий, древесина и сердцевина.
Первичная кора начинается клетками пластинчатой хлорофиллоносной колленхимы, затем идут хлоренхима и паренхима. Крахмалоносное влагалище выражено нечетко. К первичной коре примыкает перицикл. Над флоэмными участками он состоит из небольших групп одревесневших волокон склеренхимы.
Благодаря деятельности камбия происходят глубокие вторичные изменения. Камбий наращивает внутрь мощные слои вторичной ксилемы (древесины). Наружу камбий образует широкий слой вторичной коры, которая состоит из флоэмы (луба) и сердцевинных лучей. По мере утолщения стебля клетки тонкостенной хлорофиллоносной паренхимы сердцевинных лучей сильно растягиваются. На срезах они выделяются в виде светлых треугольников, чередующихся с трапециевидными участками флоэмы. Сердцевинные лучи продолжаются в ксилеме в виде узких полос, доходя до сердцевины. Это первичные сердцевинные лучи. Они служат для передвижения веществ в радиальном направлении. К осени в них откладываются крахмал и масло, расходуемые весной. По мере нарастания коры и древесины из камбия один за другим возникают новые сердцевинные лучи (вторичные). Длина каждого луча зависит от его возраста.
Флоэма липы неоднородна. Камбий откладывает чередующиеся полоски — твердого (лубяные волокна) и мягкого (тонкостенные живые элементы) луба. Мягкий луб состоит из ситовидных трубок с клетками-спутницами и лубяной паренхимы. Ситовидные трубки относятся к собственно проводящей системе, по ним перемещаются органические вещества, выработанные в листьях; лубяная паренхима служит для накопления запасных питательных веществ (крахмала, глюкозы, масла). Весной они гидролизуются и расходуются, осенью откладываются вновь. Твердый луб (лубяные волокна) — это флоэмная склеренхима. Волокна твердого луба - мертвые клетки с утолщенными одревесневшими стенками. Длина их невелика — 0,87..0,26 мм. Между приростами луба, образуемого камбием из года в год, нет ясно видимых границ. Вторичная кора (от камбия до эндодермы) вместе с первичной (от эндодермы до перидермы) образуют кору стебля. На срезе дерева кора занимает незначительную площадь 1/10 объема.
Кора липы легко отделяется от ствола, особенно весной, когда начинается сокодвижение и клетки камбия, лежащие глубже, интенсивно делятся. По камбию снимают с липы кору, которую раньше использовали для плетения лаптей (лыко), изготовления коробов, коробок-лубянок, кровли и т. д.
Камбий расположен между корой и глубжележащей древесиной. Состоит он из узких четырехгранных призматических тонкостенных клеток с крупными ядрами и цитоплазмой почти без вакуолей. Клетки камбия делятся параллельно поверхности органа, образуя правильные радиальные ряды. За счет камбия стебель растет в толщину.
Внутрь от камбия идет древесина, составляющая 9/10 объема ствола. Древесина (ксилема) включает сосуды (трахеи), трахеиды, древесинную паренхиму и древесинную склеренхиму (либриформ). Общая особенность всех элементов ксилемы — одревеснение клеточных стенок. Древесина характеризуется наличием годичных слоев (годичных колец), четко выделяющихся на поперечном срезе. Происхождение их связано с периодичностью в деятельности камбия. К зиме камбий вступает в период покоя. Весной, с началом сокодвижения в период распускания листьев, камбий активно делится, откладывая широкопросветные и тонкостенные крупные сосуды. К сосудам обычно примыкают клетки древесинной паренхимы. С приближением осени, когда деятельность камбия ослабевает, возникают узкопросветные сосуды и трахеиды, древесинные волокна. Отличия весенней и осенней древесины создают резкую границу между годичными кольцами. По числу годичных колец можно определить возраст дерева.
В центре стебля находится сердцевина, окруженная небольшим числом спиральных и кольчатых сосудов первичной ксилемы.
На ширину годичных колец в значительной степени влияют условия произрастания. У сосны и лиственницы, прозябающих на болоте, образуются такие узкие слои, что в возрасте 20...30 лет диаметр ствола не превышает 2 см. Влияют также погодные условия и затенение. Широкие кольца соответствуют благоприятным годам жизни, узкие — неблагоприятным. Анализ годичных колец позволяет установить колебания климатических условий за многие века. (Дендрохроноло́гия (от греч. dendron — дерево, chronos — время, logos — учение) — научная дисциплина о методах датирования событий, природных явлений, археологических находок и древних предметов, основанная на исследовании годичных колец древесины. Используется для датирования деревянных предметов и фрагментов древесных стволов (например, в постройках), а также в биологии — при изучении биологических изменений за последние тысячелетия. Существует направление в дендрохронологии — дендроклиматология, занимающаяся изучением закономерностей сложения годичных слоев древесных пород для установления климата в прошлые геологические эпохи.) Во влажных тропиках, где не существует резкой смены времен года, годичные кольца не обнаруживаются, так как древесина нарастает равномерно.
Особенности строения стебля хвойных.
У хвойных (ель, сосна и др.) строение стебля сходно со строением его у древесных двудольных.
Отличительные особенности строения стебля хвойных:
1. Наличие вертикальных и горизонтальных смоляных ходов в коре и в древесине.
2. Флоэма состоит только из ситовидных клеток и лубяной паренхимы.
3. Ксилема состоит в основном из трахеид с окаймленными порами (сосудов нет).
По стволам хвойных вода поднимается гораздо медленнее, чем по стволам лиственных древесных пород.
Похожие статьи:
poznayka.org
Листья хвойных растений
Категория: Анатомия растений
Листья хвойных растений
Листья многих хвойных растений живут в течение нескольких лет. Они приспособлены к недостаточному водоснабжению, особенно в зимнее время, и резким колебаниям летних и зимних температур. Поэтому листья большинства хвойных имеют ксероморфную структуру: они жесткие, мелкие, с малой испаряющей поверхностью. С анатомическим строением листьев хвойных можно ознакомиться на примере сосны.
Строение листа (хвои) сосны обыкновенной (Pinus sylvestris L.)
У сосны обыкновенной жесткие игольчатые листья (хвоя) располагаются парами на укороченных побегах.
Хвою фиксируют спиртом, который частично растворяет содержащуюся в них смолу. Чтобы легче получить поперечные срезы, пары хвоинок зажимают между кусочками сердцевины бузины или втыкают в сердцевину. Тонкие срезы обрабатывают раствором флороглюцина и соляной кислотой.

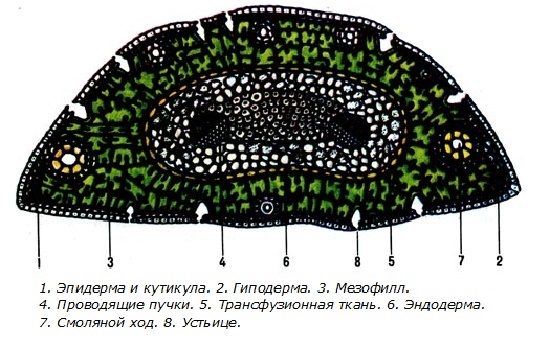
Рис. 1. Поперечный разрез хвои сосны (схема): эп — эпидермис, у — устьица, г — гиподерма, с. к. — смоляной канал, с. м. — складчатый мезофилл, энд — эндодерма, п. п. — проводящий пучок, т. т. — трансфузионная ткань, кс — ксилема, фл — флоэма, скл — склеренхима
Поперечный срез листа имеет полукруглые очертания (рис. 89). Снаружи расположен эпидермис с толстой кутикулой. Клетки эпидермиса почти квадратные. Наружные, боковые и внутренние стенки клеток сильно утолщены, у наиболее старых листьев они нередко одревесневают. От небольшой округлой внутренней полости к углам клетки отходят узкие щелевид-ные поровые каналы. Под эпидермисом находится гиподерма, состоящая из одного, а в углах — из двух-трех слоев волокон с утолщенными одревесневшими стенками.
Устьица расположены по всей поверхности листа. Их замыкающие клетки находятся на уровне гиподермы, под околоустьичными клетками. Околоустьичные клетки очень крупные, с сильно утолщенными наружными стенками. Стенки замыкающих и околоустьичных клеток в утолщенных местах одревесневают. Устьичная щель ведет в подустьичную воздушную полость, окруженную клетками мезофилла.

Рис. 2. Часть поперечного среза хвои сосны: эп — эпидермис, з. к. — замыкающая клетка, о. к. — околоустьичная клетка, пд. п — подустьичная полость, г — гиподерма, с. к. — смоляной канал, э. к.—эпителиальные клетки, скл — склеренхима, с. м. — складчатый мезофилл, энд — эндодерма с крахмальными зернами, к. т. п. — клетка транс-фузионной паренхимы с зернами крахмала, к. т. т. — клетка трансфузион-ной трахеиды с окаймленными порами, я — ядро, хл — хлоропласты
Мезофилл однородный, складчатый. Складки возникают вследствие врастания внутренних слоев оболочки в полость клетки, которая при этом приобретает лопастные очертания. За счет складок увеличивается поверхность постенного слоя цитоплазмы, содержащего хлоропласты. Клетки мезофилла соединены плотно, межклетники между ними очень малы.
В мезофилле непосредственно под гиподермой или несколько глубже расположены схизогенные смоляные каналы. Они проходят вдоль листа и заканчиваются слепо вблизи его верхушки. Снаружи смоляной канал имеет обкладку из толстостенных неодревеснев-ших волокон. Внутри он выстлан тонкостенными живыми клетками эпителия, выделяющими смолу.
Проводящая система представлена двумя коллатеральными закрытыми пучками, расположенными в центре хвои под углом один к другому. Ксилема, состоящая из трахеид с узкими полостями, обращена к плоской стороне листа, флоэма — к выпуклой. Таким образом, плоская сторона хвои представляет собой морфологически верхнюю, а выпуклая — морфологически нижнюю стороны листа.
Снизу между пучками находится тяж волокон с толстыми, слегка одревесневшими стенками. Проводящие пучки и примыкающие к ним механические элементы окружены трансфузионной тканью, состоящей из клеток двух типов. Возле ксилемы клетки несколько удлинены, содержимого в них нет, их одревесневшие стенки имеют окаймленные поры. Эти клетки называют трансфузионными трахеидами. Остальные клетки живые, паренхимные, тонкостенные. Они содержат смолистые вещества, нередко в них встречаются зерна крахмала. Трансфузионная ткань, по-видимому, участвует в перемещении веществ между проводящими пучками и мезофиллом.
Проводящие пучки вместе с окружающей их трансфузионной тканью отделены от мезофилла эндодермой, представляющей собой однорядный слой паренхимных клеток с пятнами Каспари на радиальных стенках.
Задание. 1. При малом увеличении микроскопа зарисовать схему строения листа, отметив эпидермис с устьицами, гиподерму, складчатый мезофилл, смоляные каналы, эндодерму, проводящие пучки, механические волокна и трансфузионную ткань.2. При большом увеличении зарисовать участок хвои с эпидермисом, гиподермой, складчатым мезофиллом, смоляным каналом и эндодермой с пятнами Каспари на
Кроме сосны складчатый мезофилл и смоляные каналы встречаются у ели (виды Picea), кедра (виды Ced-rus), листья которых содержат по одному проводящему пучку.
Один проводящий пучок встречается и у так называемых пятихвойных сосен, например у сибирской (Pinus sibirica (Rupr.) Мауг.) и веймутовой (P. strobus L.), у которых укороченные побеги несут не по две хвоинки, как у обыкновенной сосны, а по пяти.
Лист тиса (Taxus baccata L.) более широкий, складчатого мезофилла в нем нет. На верхней стороне листа хлорофиллоносные клетки несколько вытянуты по вертикали и более узкие, чем на нижней стороне. Смоляной канал без механической обкладки расположен близ флоэмной части единственного проводящего пучка, с двух сторон от которого находится трансфузионная ткань.
Анатомия растений - Листья хвойных растенийgardenweb.ru
Стебли древесных двудольных и хвойных растений
Если мы будем рассматривать срез только что сформировавшегося стебля древесного растения (вблизи точки роста), то мы его не сможем отличить от первичной структуры стебля травянистого двудольного растения с непучковым типом строения. Но уже к середине лета, особенно к осени мы обнаружим эти различия. Во-первых, у древесных растений (деревьев и кустарников) всегда формируется непучковый тип строения, во-вторых, покровная ткань, уже в первый год жизни меняется на перидерму, а с возрастом на древесном стволе образуется корка (ритидом). Следующей особенностью является наличие годичных колец. Мощное развитие ксилемы, ежегодное образование с помощью камбия слоя весенней (крупнопросветной) и осенней (толстостенной и мелкопросветной) ксилемы, именуемой у деревьев древесиной, создаются годичные кольца, Толщина годичного кольца зависит от климатических условий, состава, влажности почвы и других эколого-географических факторов. Например, у 40-летней карликовой березы (Betula nana), выросшей в Гренландии ствол имел 4 мм в диаметре, а средняя ширина годичного кольца не более 0,05 мм, а у экземпляров того же вида, выращенного в ботаническом саду Средней Европы, средняя ширина годичного кольца древесины составляла уже около 16 мм. Древесина еще при жизни дерева подвергается с течением времени значительным изменениям. Сосуды закупориваются тиллами - выростами стенок соседних с сосудами паренхимных клеток, проникающих сквозь поры в оболочках сосудов. Процесс закупорки просвета сосудов предохраняет древесину от повреждения различными грибковыми заболеваниями. Тиллы в большом количестве характерны старой древесине. Закупоренные сосуды прекращают функционировать в качестве проводящих воду элементов. Живые клетки этих слоев ксилемы отмирают и выполняют лишь механическую функцию, как и либриформ – обеспечение прочности ствола. Более старая часть древесины (ближе к сердцевине) резко выделяется своей окраской. Это ядровая древесина. Живая, более молодая древесина (расположена ближе к камбию) называется у работников лесного хозяйства заболонью. Она имеет светлую окраску и очень активно функционирует в качестве проводящей воду ткани. Ядровая древесина имеет различную окраску из-за локализации в ней различных, чаще всего фенольных соединений. У ольхи ядровая древесина окрашена в оранжевый цвет, у барбариса – в желтый, у красного дерева (Caesalpinia brasiliensis) - в красный, у грецкого ореха - в бурый, у тисса - темно-красный, у кампешевого дерева - в кроваво-красный. Из кампешевого дерева извлекают очень популярный в гистологии и цитологии краситель гематоксилин (рис. 91).
Строение стебля хвойных растений
Стебель хвойных растений имеет общую со стеблем лиственных растений структуру - непучковый тип строения (рис. 92), ксилема и флоэма которых располагается сплошным кольцом. Между флоэмой и ксилемой располагается слой камбиальных клеток, обеспечивающих рост стебля в толщину. В древесине хорошо выражены годичные кольца. В центре - сердцевина. Покровная ткань - перидерма или корка. Отличительные особенности в структуре стебля хвойных растений заключаются в том, что проводящие элементы ксилемы представлены только трахеидами с различными типами утолщений, от чего ксилема является однородной. Трахеиды не закупориваются тиллами - они заполняются смолой, кроме того этот процесс происходит значительно позднее, чем у лиственных пород. Хвойным характерно наличие смоляных ходов. В стебле хвойных смоляные ходы двух типов – однослойные и боченковидные. Их можно обнаружить и в древесине и в коровой части, рассматривая срезы стебля в тангентальном или радиальном направлении и на поперечных срезах. У хвойных во флоэме отсутствуют клетки - спутницы.
Для облегчения работы по диагностике типов осевых органов нами предлагается определитель (табл. ) осевых органов по поперечным срезам. Из указанной таблицы видно, что основным диагностическим признаком стебля и его видоизменений, а так же отличием от корня является наличие сердцевины. Характер распределения проводящих тканей лежит в основе более детальной диагностики осевых вегетативных органов.
studfiles.net
Стебель хвойных растений
Категория: Анатомия растений
Стебель хвойных растений
Строение молодого стебля сосны обыкновенной (Pinus sylvestris L.)
Для изучения анатомического строения стебля могут быть использованы 3—4-летние удлиненные побеги сосны, выросшей в благоприятных условиях освещения. Лучше всего собирать материал во второй половине лета, когда прирост побегов в длину закончен. Материал фиксируют в спирте.
Поперечные срезы последовательно обрабатывают флороглюцином, соляной кислотой и заключают в глицерин. До проведения реакции на одревеснение срезы можно обработать раствором иода в водном растворе йодистого калия. Для работы пригодны срезы небольших размеров; важно только, чтобы в них были и периферические, и внутренние участки стебля, включающие сердцевину. Анатомическую топографию стебля изучают при малом увеличении, некоторые детали строения— при большом увеличении микроскопа.
Стебель сосны в поперечном сечении более или менее округлый.
Вокруг нее расположена слабо развитая первичная древесина, элементы которой нередко оказываются более одревесневшими, чем примыкающие к ним снаружи клетки вторичной древесины. Во вторичной древесине можно различить кольца прироста. Древесина состоит из толстостенных трахеид, расположенных радиальными рядами. В каждом кольце прироста видны схизогенные смоляные каналы, окруженные обкладкой из неодревесневших тонкостенных паренхимных клеток. Радиальные ряды трахеид последнего, самого молодого кольца прироста постепенно переходят в камбиальную зону, состоящую из нескольких слоев тонкостенных узко-просветных клеток, вытянутых в тангентальном направлении.

Рис. 1. Схема строения трехлетней ветки сосны: пд — перидерма, л. к. — первичная кора, в. с. «. — вертикальные смоляные каналы, в. фл. — вторичная флоэма, кмб— камбий, в. др.— вторичная древесина, г. г. п. — границы годичных приростов, л. др. — первичная древесина, сц — сердцевина, л. з. — перимедуллярная зона, л. лд. л. — первичные и в. лд. л. — вторичные лубо-древесинные лучи, з. см. — замкнутые смоловместилища
Вокруг камбиальной зоны узким кольцом расположен вторичный луб, или вторичная флоэма. Первичная флоэма, находящаяся на границе с первичной корой, практически неразличима.
Вторичный луб состоит из очень мелких светлых таблитчатых в поперечном сечении ситовидных элементов и более или менее округлых паренхимных клеток, составляющих короткие тангентальные цепочки. На срезе обработанном раствором иода в водном растворе йодистого калия, в паренхимных клетках можно обнаружить запасной крахмал. Все элементы луба расположены радиальными рядами.
В радиальном направлении древесина и луб пересечены узкими, обычно однорядными лучами разной протяженности. Формирование лучей начинается в период первичной дифференцировки анатомической структуры стебля, когда центральный цилиндр имеет еще пучковое строение. При вторичном утолщении нарастание лучей в радиальном направлении осуществляется за счет камбия, производные которого раздвигают концевые участки лучей, расположенные в зонах хорошо сохраняющейся в стеблях первичной древесины и плохо заметной первичной флоэмы. Хотя в стеблях с вторичным утолщением почти весь луч составлен клетками вторичного (камбиального) происхождения, за этими лучами можно сохранить название первичных, как это было принято в старой учебной анатомической литературе. Первичные лучи располагаются между первичной корой и сердцевиной. Этим, вероятно, и объясняется также то, что в русской и зарубежной литературе лучи называют сердцевинными, применяя этот термин и к лучам вторичного происхождения, возникающим в любом возрасте побега вследствие деления коротких камбиальных клеток. Вторичные лучи по сравнению с первичными имеют меньшую протяженность в радиальном направлении, никогда не доходят до первичной коры и сердцевины, но по строению не отличаются от первичных лучей.
В лубе лучи строго радиальны лишь вблизи камбиальной зоны, по мере удаления от нее они слегка искривляются. Некоторые лучи на конце расширены и вдаются в первичную кору. Расширения представляют собой замкнутые смоловместилища шаровидной или мешковидной формы.
Первичная кора довольно узкая, наружная часть ее состоит из клеток, более или менее вытянутых в тангентальном направлении. В первичной коре хорошо заметны округлые или овальные в поперечном сечении вертикальные смоляные каналы, расположенные вокруг центрального цилиндра.
Первичная кора окружена перидермой, все элементы которой расположены радиальными рядами. Наружная часть перидермы состоит из нескольких слоев клеток пробки, внутренняя — из феллогена и трех-четы-рех слоев клеток феллодермы. Снаружи от перидермы видны сухие, легко отрывающиеся от стебля чешуйки, представляющие собой остатки отмершего эпидермиса и нескольких наружных слоев клеток первичной коры.
Задание. При малом увеличении микроскопа зарисовать схему строения многолетней ветки, отметив сердцевину, первичную древесину, вторичную древесину с кольцами прироста, камбиальную зону, луб, лубо-древесин-ные (сердцевинные) лучи, первичную кору со смоляными каналами и перидерму.
Строение проводящих тканей
Для детального изучения строения проводящих тканей—древесины и луба (флоэмы)—лучше использовать древесину и кору стволов взрослых (не моложе 10—15 лет) деревьев, так как в стволе структурные особенности элементов, слагающих проводящую систему, выражены лучше, чем в ветвях.
Древесина и луб представляют собой сложные ткани, состоящие из элементов, различных по строению, функции и расположению. Такие ткани изучают на срезах, проведенных в трех взаимно перпендикулярных направлениях: поперечном, перпендикулярном продольной оси органа, продольном радиальном, плоскость которого проходит по радиусу перпендикулярно кольцам прироста древесины, и продольном, тангентальном, проведенном по касательной к ним, перпендикулярно сердцевинным лучам (рис. 1).

Рис. 2. Образец древесины, подготовленный для изготовления срезов: г. к. — годичные кольца древесины, Я — плоскость поперечного среза, Р — плоскость продольного радиального среза, Т — плоскость тангентального среза
Небольшие кусочки стволовой коры снимают вместе с камбиальной зоной и периферическим кольцом древесины Место, с которого снята кора, следует замазать садовым варом. Материал собирают летом до окончания вегетационного периода и фиксируют в спирте.
Кусочки древесины хранят в спирте с глицерином. Перед изготовлением срезов древесину для размягчения кипятят в воде в течение 1—2 ч и режут в горячем состоянии, смазывая глицерином поверхность среза, которую предварительно выравнивают скальпелем, ориентируясь по кольцам прироста. При изготовлении поперечных срезов бритву следует вести не поперек, а вдоль этих колец. В таком направлении древесину резать легче. Срез должен захватить два-три кольца прироста. Для получения продольных срезов необходимо сделать небольшие поперечные зарубки на расстоянии 5—7 мм от торца образца. Это предохранит от поранения, если бритва сорвется.
Строение древесины сосны обыкновенной (Pinus sylvestris L.)
Древесина состоит из продольных трахеид, обеспечивающих восходящий ток воды и растворенных в ней минеральных веществ, а также выполняющих механическую функцию; лучевых трахеид, обусловливающих перемещение водных растворов в горизонтальном (радиальном) направлении; паренхимных клеток, входящих в состав лучей и обкладки смоляных каналов; эпителия, выстилающего полость канала и участвующего в выделении смол, эфирных масел и бальзамов.
Срезы древесины рассматривают в глицерине.
На поперечном срезе хорошо выражены кольца прироста (рис. 48). Внутренняя часть каждого кольца состоит из довольно широкопросветных, в очертании многоугольных трахеид, слагающих раннюю, или весеннюю, древесину. Периферическая часть кольца прироста, образующаяся во второй половине вегетационного периода, представлена более толстостенными уз-копросветными трахеидами, составляющими позднюю, или летнюю, древесину. Обычно она темнее ранней.
В пределах каждого кольца переход от ранней древесины к поздней постепенный. Трахеиды расположены радиальными рядами. Весенние (ранние) трахеиды выполняют главным образом функцию проведения водных растворов. На их радиальных стенках видны крупные окаймленные поры. Они отличаются от простых пор тем, что вторичные оболочки трахеид, образующие стенки камеры поры, сводообразно приподняты над замыкающей пленкой, или мембраной поры. В середине свода находится отверстие, соединяющее камеру поры с полостью трахеиды.

Рис. 3. Поперечный срез древесины сосны: г г. п.— граница годичного прироста, т. р. д.— трахеиды ранней древесины, т. п. д. — трахеиды поздней древесины, о. га. — окаймленные поры, т — торус, п. га.— полуокаймленная пора, д. л.—древесинный луч, л. т. — лучевая трахеида, л. га. — лучевая паренхима в. с. к. — вертикальный смоляной канал, п. с. к. — паренхимная обкладка смоляного канала, э. «. — эпителиальные клетки, см. — капли смолы
Если разрез прошел через боковые части свода, то пора выглядит как линзовидное расширение клеточной стенки. Стенки трахеид летней (поздней) древесины имеют небольшое число пор. Основная функция этих трахеид —механическая. Между радиальными рядами трахеид расположены древесинные лучи, состоящие обычно из одного ряда клеток, структурные особенности которых лучше рассматривать на радиальном срезе.
Смолов ы делительная система древесины представлена вертикальными и горизонтальными смоляными канала-м и. Вертикальные смоляные каналы обычно расположены в поздней древесине кольца прироста. На срезе они видны в поперечном сечении. Канал окружен обкладкой из паренхимных клеток. Полость канала выстлана тонкостенными эпителиальными клетками. При изготовлении среза нежные эпителиальные Радиальный
клетки нередко разрушаются, срез древесины дает представление о трахеидах как очень длинных, тых в продольном направлении прозенхимных клетках со слегка закругленными окончаниями. Между концами трахеид, расположенными на одном уровне, ‘вклиниваются трахеиды, принадлежащие другому ярусу.
Окаймленные поры, находящиеся на радиальных тенках трахеид, видны в плане. Они представляют со-ои совокупность трех концентрических окружностей, самая внутренняя из которых соответствует отверстию камеры поры, средняя — очертаниям торуса, внешняя — контурам окаймления.

Рис. 3. Продольный радиальный срез древесины сосны: д. л. — древесинный луч, л. п. — лучевая паренхима, л. т. — лучевые трахеиды, т. Р. д. — трахеиды ранней древесины, т. п. д. — трахеиды поздней древесины, о. тр. — окончания трахеид, о. п. — окаймленные поры, от — отверстие поры, ок — окаймление поры, т — торус, ок. п. — окновидные (оконцевые) поры
Древесинные лучи на радиальном срезе имеют вид широких горизонтальных полос, состоящих из нескольких рядов клеток. Внутренняя часть луча сложена тонкостенными клетками лучевой паренхимы, вытянутыми в радиальном направлении. Клетки содержат крахмал и капли масла. В старой древесине протопласты клеток лучевой паренхимы отмирают. В месте пересечения с каждой продольной трахеи-дой в стенке паренхимной клетки формируется по одной крупной округло-четырехугольной простой поре. Эти поры называют оконцевыми или ок-новидными. Общее число пор в клетке зависит от числа трахеид, с которыми данная клетка соприкасается. Со стороны трахеиды поры имеют небольшое окаймление. Такие полуокаймленные поры можно было видеть на поперечном срезе древесины, если его плоскость прошла через середину луча. По краям луча располагаются лучевые трахеиды, или трахеидальные клетки. Это мертвые клетки, обеспечивающие перемещение воды в радиальном направлении. Обычно они короче клеток лучевой паренхимы и часто имеют неправильные очертания. Внутренняя поверхность их клеточных стенок зубчатая. Многочисленные мелкие окаймленные поры расположены на всех стенках. Лучи, состоящие из клеток функционально и морфологически неоднородных, называют гетерогенными.

Рис. 4. Продольный тангентальный срез древесины сосны: о. тр. — окончания трахеид, о. п. — окаймленные поры, о. л. — однорядный луч, м. л. — многорядный луч, с. к. — смоляной канал, э. к.—эпителиальные клетки, л. т.— лучевые трахеиды, л. п.— лучевая паренхима, ок. п. — оконцевая пора, т — торус
На тангентальном срезе вертикальные трахеиды имеют заостренные окончания. Поры на их радиальных стенках видны в разрезе. Древесинные лучи вклинивающиеся между трахеидами, перерезаны поперек. Они представляют собой цепочки коротких округлых или квадратно-прямоугольных клеток. Средняя часть луча занята клетками запасающей паренхимы, краевые клетки луча представлены лучевыми трахеидами. Наряду с однорядными лучами высотой 1 —12 клеток встречаются сложные многорядные веретеновидные лучи, в расширенной средней части которых находится по одному смоляному каналу, принадлежащему горизонтальной смоловыделительной системе. На продольных срезах могут быть и вертикальные смоляные каналы в виде широких полос, состоящих либо из эпителиальных клеток, либо из клеток паренхимной обкладки. Если срез пройдет через середину полости канала, то срез распадется на две части.
Задание. 1. При большом увеличении микроскопа зарисовать участок поперечного среза на границе двух колец прироста, обратив внимание на характер поперечного сечения трахеид ранней и поздней древесины; отметить поры на радиальных стенках, изобразить древесинный луч и смоляной канал в поздней древесине.2. Зарисовать участок радиального среза, отметив особенности строения продольных трахеид с окаймленными порами, показать луч, состоящий из лучевых трахеид с мелкими окаймленными порами и клеток паренхимы с порами в стенках, смежных со стенками продольных трахеид.3. Зарисовать строение древесины на тангентальном срезе, отметив трахеиды с цепочками окаймленных пор на перерезанных радиальных стенках; изобразить простои, однорядный, и сложный, многорядный, лучи.
Строение вторичного луба сосны обыкновенной (Pinus sylvestris L.)
Вторичный луб (флоэму), так же как и древесину, изучают на поперечных, радиальных и тангентальных срезах.
В проведении продуктов ассимиляции участвует лишь самая молодая часть луба, прилегающая к камбию. Ширина ее обычно не превышает 0,3 мм. Снаружи от нее расположена более старая и более широкая зона, в которой проводящие элементы уже не функционируют. Эту зону называют непроводящей. У сосны непроводящий луб выполняет функции запаса питательных веществ и выделения. Луб окружен чешуйчатой коркой (см. с. 143).
Вторичный луб, или флоэма, состоит из ситовидных элементов, проводящих растворы органических соединений, продольных тяжей паренхимных клеток, лучевой паренхимы и смоловыделитель-ных эпителиальных клеток.
Функциональные различия между проводящей и непроводящей зонами луба коррелируют с их структурными особенностями.
На поперечном срезе элементы проводящей зоны расположены радиальными рядами. Ситовидные клетки тонкостенные, широкопросвет-ные, квадратных или прямоугольных очертаний.

Рис. 4. Схема строения стволовой коры сосны: к — корка, ч. к. — чешуи корки, пд — перидерма,, н. фл. — непроводящая флоэма, п. фл. — проводящая флоэма, кмб — камбий, др — древесина, лд. л. — лубо-древесинные лучи, смв — смоловместилища, т. р. п. — тангентальные ряды паренхимных клеток

Рис. 5. Поперечный срез вторичного луба сосны: л. — лубо-древесинный луч, с. к. — ситовидные клетки, с. п. — ситовидные поля, п. к,— паренхимные клетки, к. з. — крахмальные зерна, кр — кристалл оксалата кальция, кмб — камбий, др — древесина
Лучи преимущественно однорядные, реже состоят из нескольких рядов вытянутых в радиальном направлении паренхимных клеток с цитоплазмой, ядром, иногда с зернами крахмала. В проводящей зоне лучи строго радиальны, в непроводящей они извилисты, некоторые лучи местами расширены. Мешковидные или головчатые расширения лучей представляют собой замкнутые смо-ловместилища. Они формируются в результате преобразования лучевой паренхимы в эпителиальные смоло-выделительные клетки.
В непроводящей зоне слои широкопросветных ситовидных клеток обычно чередуются со слоями сильно деформированных, сжатых в радиальном направлении очень тонкостенных ситовидных клеток, расположенных косыми рядами. Слои этих клеток, дифференцирующихся, по-видимому, весной, определяют границы годичных приростов. Если материал собран с деревьев, выросших в неблагоприятных условиях, годичная слоистость луба не видна.
Во внутренних частях непроводящей зоны паренхим-ные клетки составляют рыхлые тангентальные цепочки. Одни клетки округлые, значительно крупнее проводящих элементов, с зернами запасного крахмала (запасающая паренхима), другие — мелкие, угловатые, с бурым содержимым и кристаллическими включениями (кри-сталлоносная паренхима).
В периферической зоне луба, примыкающей к корке, объем паренхимы значительно увеличивается вследствие деления клеток и их последующего растяжения. Это обусловливает разрастание (дилатацию) луба в танген-тальном направлении. Из паренхимных клеток наружных слоев непроводящей зоны в результате двух последовательных делений тангентальными перегородками вычленяются клетки пробкового камбия (феллогена), образующего перидерму. На поперечном срезе внутренние перидермы располагаются отдельными дугами.
На продольном радиальном срезе видно, что ситовидные элементы по строению сходны с трахеидами. Это очень длинные клетки с притуплёнными концами. На их радиальных стенках расположены в один ряд округлые ситовидные поля с очень мелкими прободениями. На срезе, включающем проводящий луб, камбиальную зону и древесину, можно проследить постепенное формирование ситовидных полей, которые обычно находятся почти на одном уровне с окаймленными порами в трахеидах.
Паренхимные клетки, находящиеся между ситовидными элементами, вытянуты в продольном направлении. Они составляют тяжи, конечные клетки которых имеют клиновидные очертания. Такую паренхиму называют тяжевой. В проводящей зоне паренхимные клетки по ширине почти не отличаются от ситовидных элементов, в непроводящей — они более широкие. На периферии этой зоны правильность их расположения продольными тяжами постепенно исчезает, клетки округляются, между ними возникают межклетники. Кроме клеток запасающей паренхимы в тяже могут быть и кристаллоносные клетки с одиночными призматическими кристаллами оксалата кальция, окруженными матриксом из дубильных веществ, смол и других соединений, обусловливающих бурую окраску содержимого этих клеток. Кристаллоносные клетки обычно мертвые.

Рис. 5. Продольные радиальные срезы проводящей (Л) и непроводящей (Б) зоны вторичной флоэмы сосны: с. к. — ситовидные клетки, с., п. — ситовидные поля, о. с. к. — окончания ситовидных клеток, л. л. — лубяной луч, л. к.— внутренние лежачие клетки луча, о. к. — «белковые» клетки, п. к. — паренхимные клетки, ст. к. — краевые стоячие клетки луча, к. п. — крахмалоносная паренхима, з. л. п. — запасающие клетки лучевой паренхимы, кр. п. — кристаллоносная паренхима, кр — кристалл оксалата кальция, к. з. — крахмальные зерна, я — ядра
Лубяные лучи представляют собой горизонтальные полосы, состоящие из нескольких рядов клеток.
На срезах непроводящего луба лучи часто прерывисты, так как в этой зоне они проходят не строго ради-ально. Лучи в лубе сосны гетерогенные. Внутренние их клетки, вытянутые в радиальном направлении, называют лежачими. В них можно видеть цитоплазму и ядро,
в непроводящем лубе эти -ил клетки содержат запасной крахмал. Краевые клетки, вытянутые в вертикальном направлении, называют стоячими. Те из них, которые соприкасаются с тяжевой паренхимой, выполняют функцию запаса, а клетки, контактирующие с ситовидными элементами через мелкие ситовидные поля, имеют цитоплазму, ядро, но никогда не содержат запасного крахмала. По функции они сходны с сопровождающими клетками флоэмы цветковых растений. Эти клетки называют «белковыми» или клетками Страсбургера. «Белковые» клетки после прекращения деятельности ситовидных элементов отмирают, поэтому в непроводящем лубе они очень мелкие, деформированные и выглядят пустыми в отличие от крупных округлых запасающих клеток лучевой паренхимы. Если срез прошел через смоловместилище, то внутри луча можно видеть полость, окруженную многочисленными тонкостенными эпителиальными клетками.
Тангентальный срез проводящего луба расположением элементов напоминает тангентальный срез древесины. На радиальных стенках ситовидных элементов ситовидные поля видны в разрезе. Хорошо выражена тяжевая паренхима, состоящая из крахмалоносных и кристаллоносных клеток. Лубяные лучи двух типов: однорядные, сложенные округлыми или округло-четырехугольными клетками, и более высокие много-рядные с перерезанным поперек смоляным каналом, который представляет собой продолжение горизонтального смоляного канала древесины. В непроводящей зоне луба смоляные каналы расширяются, увеличивается число выстилающих их эпителиальных клеток. Остальные клетки луча неравномерно разрастаются, и некоторые из них по внешнему виду почти не отличаются от клеток тяжевой паренхимы.

Рис. 5. Продольный тангентальный срез вторичной флоэмы сосны: с. к. — ситовидные клетки, с. п. — ситовидные поля, к. п. — клетки крис-таллоносной паренхимы, з. п. — клетки запасающей паренхимы, о. л. — однорядный луч, м. л. — многорядный луч, б. к. — «белковая» клетка, см. к. — смоляной канал, я — ядро, к. з. — крахмальные зерна, кр — кристаллы оксалата кальция, э. к. — эпителиальные клетки
Строение перидермы и корки сосны обыкновенной (Pinus sylvestris L.)
Стволы 10—15-летних и более старых деревьев имеют корку — комплекс мертвых наружных участков луба, разделенных перидермами, которые на поперечных срезах имеют вид дуг. Такую корку называют чешуйчатой.
Перидермы, разделяющие чешуи корки, состоят из нескольких слоев клеток, расположенных радиальными рядами. Три-четыре самых внутренних слоя представляют собой фел-лодерму. В самой внутренней перидерме, расположенной на границе с жизнедеятельным лубом, клетки феллодермы живые, со слегка утолщенными
стенками. Кнаружи от нее располагается феллоген, или пробковый камбий, в виде однорядного слоя таблитчатых клеток. В наружных перидермах, в которых деятельность феллогена прекращена, его клетки неотличимы от клеток феллодермы. Периферические клетки перидермы составляют пробку, или феллему. У сосны она состоит из трех типов клеток. Внутренние клетки имеют слегка утолщенные стенки, содержащие суберин. Это типичные клетки пробки. К ним примыкают прозрачные клетки с очень тонкими стенками. Радиальные стенки их обычно извилисты. Это так называемая губчатая пробка. Наружные слои перидермы представлены клетками с очень толстыми одревесневшими слоистыми стенками, в которых обычно хорошо видны многочисленные узкие норовые каналы. Эти клетки по строению сходны с каменистыми клетками. Их называют феллоидами. Так как связь между клетками-феллоидами и губчатой пробкой очень непрочна, между ними легко возникают разрывы, способствующие опадению чешуй корки.

Рис. 6. Строение перидермы сосны: фд — феллодерма, фг — феллоген, т. п.— типичная пробка, г. п. — губчатая пробка, к. п. — каменистая пробка, к. крк. — клетки корки, п — поры, л. п. — лубяная паренхима, я —ядро
Задание. 1. При малом увеличении микроскопа зарисовать схему строения вторичного луба и корки, отметив проводящую и непроводящую зоны, однорядные лучи и лучи со смоловместилищами.2. При большом увеличении микроскопа зарисовать участки поперечного, радиального и тангентального срезов, показав на рисунках расположение и общий вид ситовидных клеток с ситовидными полями на радиальных стенках, клетки крахмалоносной и кристаллоносной тяжевой паренхимы; на радиальном срезе непроводящей зоны луба обратить внимание на строение луча, отметить в нем отмершие «белковые» клетки и клетки запасающей паренхимы.3. При большом увеличении микроскопа зарисовать три типа клеток пробки.
Вместо сосны со строением стебля хвойных растений можно ознакомиться на примере лиственницы (Larix) или ели (Picea).
В молодых 3—4-летних стеблях лиственницы вертикальных смоляных каналов в первичной коре может не быть, так как содержащая их наружная часть первичной коры рано сбрасывается вследствие более глубокого, чем у сосны, заложения феллогена и образования перидермы. Древесина лиственницы и ели отличается от древесины сосны толстостенными эпителиальными клетками, выстилающими полости смоляных каналов. Поры на радиальных стенках трахеид лиственницы иногда располагаются в два ряда. На смежных стенках между вертикальными трахеидами и паренхимными клетками образуются не оконцевые, а мелкие окаймленные поры, у лиственницы иногда встречается тяжевая древесинная паренхима.
В непроводящей зоне вторичной флоэмы этих деревьев наряду с ситовидными клетками, запасающей и кри-гтяллоносной папенхимой имеются механические элементы. У ели это группы каменистых клеток, у лиственницы — волокнистые склереиды — длинные прозенхимные толстостенные клетки, которые на поперечном срезе имеют округлые очертания.

Рис. 7. Поперечный срез луба можжевельника сибирского (Juniperus sibirica Burgsd.): л. л. — лубяные лучи, с. к. — ситовидные клетки, с. п. — ситовидное поле, п. к. — паренхимные клетки, л. в. — лубяные волокна, я — ядро
Интересно строение древесины и луба тиса (Taxus), кипариса (Сир-ressus) и можжевельника (Juniperus). В их древесине нет смоляных каналов, но хорошо развита тяжевая паренхима. Тра-хеиды тиса кроме пор имеют спиральные утолщения, образующиеся, видимо, за счет третичной оболочки. Древесинные лучи у этих пород сложены однородными клетками. Такие лучи называют гомогенными.
У кипариса и можжевельника луб состоит из чередующихся тангентальных однорядных слоев проводящих, паренхимных и механических элементов, которые в свою очередь располагаются радиальными рядами. Больших различий в строении проводящей и непроводящей зон луба нет. На поперечном срезе в радиальном ряду между двумя механическими элементами, представляющими собой очень длинные волокна с одревесневшими стенками, расположены примыкающие к ним ситовидные клетки, разделенные одной, очень редко двумя паренхимными клетками. Кристаллоносной паренхимы в лубе этих древесных пород нет. В ситовидных клетках после прекращения их деятельности откладывается мелкий кристаллический песок.
Анатомия растений - Стебель хвойных растенийgardenweb.ru
Основные болезни хвойных
22 Апреля 2013 г.

Белошапкина Ольга Олеговна, доктор сельскохозяйственных наук
Фото автора
Опубликовано в журнале "Настоящий хозяин" (№ 4/2012)
Хвойные деревья и кустарники круглый год не теряют своей привлекательности, особенно, если хорошо растут, развиваются и не болеют. Для своевременного выявления болезней необходимо регулярно проводить фитопатологический мониторинг. Затем по его результатам, оценив конкретную ситуацию, степень поражения и целесообразность проведения защитных мероприятий, а также метеорологические условия, выбирают защитные меры против конкретных заболеваний.
Визуальное диагностирование большинства болезней хвойных довольно проблематично, что связано с явлением т.н. фитопатологической конвергенции, когда одинаковые симптомы возникают в результате различных причин. К таким общим симптомам в первую очередь относится усыхание ветвей, пожелтение, побурение и опадение или отмирание хвои.
При их появлении следует начать общепрофилактические мероприятия: удалять хвою, вырезать пораженные ветви и попытаться создать благоприятные условия для роста и развития растения, включая обработки иммуномодуляторами и внекорневые и корневые подкормки удобрениями для хвойных. Нередко требуются консультации специалиста по защите растений.
Развитие болезней зачастую зависит от здоровья посадочного материала, наличия у него механических травм, повреждений насекомыми, а также правильности посадки и дальнейшего ухода. Молодые растения в целом менее устойчивы к комплексу неинфекционных и инфекционных заболеваний, с возрастом их устойчивость повышается.
Будьте внимательны при покупке саженцев. Кора должна быть равномерно характерно окрашенная, без трещин и наплывов. Концы веток и корни – эластичные, не сухие. Почки и хвоя здоровых растений живые, а не высохшие; под корой виден зеленоватый слой живой ткани; на срезе сосуды побегов – светлые, равномерно окрашенные.
СОЗДАЙТЕ ПОДХОДЯЩИЕ УСЛОВИЯ
Отрицательное влияние на рост и развитие хвойных оказывают неблагоприятные условия окружающей среды. Избыточное увлажнение, связанное с естественным заболачиванием почвы, поднятием уровня грунтовых вод, обильными осенними осадками или чрезмерными поливами контейнерных растений, приводит к пожелтению и некротизации хвои. Такие же симптомы часто появляются из-за недостатка влаги в почве и низкой влажности воздуха.
|
Низкие температуры зимой и весенние заморозки вызывают подмерзание кроны и корней, при этом хвоя может приобретать красноватый цвет, становится сухой, отмирает, кора побегов растрескивается. Весной в солнечные часы, когда почва еще не оттаяла полностью и корни не функционируют, часто наблюдается побурение, ожог хвои туи и можжевельников. По возможности, такие растения в феврале-апреле следует притенять. Для защиты от солнечных ожогов и отлупа коры, ее можно побелить известью или специальной побелкой ранней весной или в конце осени. В первый год после посадки молодые растения желательно в вечерние часы опрыскивать водой и притенять в жару.
Многие хвойные теневыносливы, при выращивании на открытых солнечных местах они могут отставать в росте, хвоя их может желтеть и даже отмирать. С другой стороны, светолюбивые сосны, лиственницы и даже можжевельники не выносят сильного затенения. |
.jpg) Ожог туи Ожог туи
|
ПОДКОРМКИ УКРЕПЛЯЮТ ИММУНИТЕТ
Состояние и внешний вид растений во многом зависят от обеспеченности элементами питания и их сбалансированности. Так, недостаток в почве железа приводит к пожелтению и даже побелению хвои на отдельных побегах; при дефиците фосфора молодая хвоя приобретает красно-фиолетовый оттенок; при нехватке азота растения заметно хуже растут, становятся хлоротичными.
Рекомендуется проводить корневые и внекорневые подкормки, лучше специальными удобрениями, предназначенными для хвойных. Имеется положительный опыт применения биологически активных препаратов, в т.ч. регуляторов роста, повышающих устойчивость растений к неблагоприятным факторам и погрешностям в уходе. Такие препараты, как супер гумисол, циркон, эпин-экстра, силиплант, никфан, иммуноцитофит, используемые в рекомендуемых производителями концентрациях для опрыскиваний и полива под корень, повышают приживаемость саженцев, укрепляют иммунитет растений к температурным, водным и даже пестицидным стрессам, улучшают потребление элементов питания.
ФУЗАРИОЗ И КОРНЕВЫЕ ГНИЛИ
Хвойные не часто поражаются инфекционными болезнями, хотя в ряде случаев могут очень страдать от них. Значительные выпады или задержку роста молодых контейнерных растений и сеянцев в школках вызывают виды почвообитающих грибов, чаще родов Pythium и Rhizoctonia, приводящих к постепенному побурению, отмиранию корней и полеганию всходов.
Саженцы и молодые растения хвойных пород подвержены также фузариозному усыханию (возбудители – анаморфные грибы рода Fusarium). Это заболевание называют еще трахеомикозным увяданием. Возбудитель из почвы проникает в корни, которые буреют, частично загнивают; затем гриб проникает в сосудистую систему и заполняет ее своей биомассой, затрудняя доступ питательных веществ. При этом на поперечном срезе пораженной ветви хорошо заметно сплошное, а чаще прерывистое потемнение кольца ксилемы и сердцевины. Хвоя желтеет, краснеет и опадает, крона частично редеет, а сами растения постепенно усыхают. Первое время заболевание может протекать в скрытой форме.
 |
 |
| Фузариоз ели | Выделение Fusarium из хвои и побегов ели |
Факторы риска. Возбудитель сохраняется в растениях, в пораженных растительных остатках и часто распространяется с зараженным посадочным материалом из питомников или с инфицированной почвой.
 Фузариоз можжевельника Фузариоз можжевельника |
Меры защиты. Лечение пораженных деревьев практически невозможно, через несколько лет они погибают. Для предупреждения корневых гнилей и фузариоза необходимо использовать здоровый посадочный материал; своевременно удалять все засохшие экземпляры с корнями и пораженные растительные остатки. В профилактических целях также замачивают молодые растения с открытой корневой системой в растворе одного из препаратов: фитоспорин-М, витарос, максим. При первых симптомах проливают почву растворами биопрепаратов: фитоспорина-М, агата-25К, гамаира, можно использовать фунгицид фундазол. |
Рекомендуется и предпосевная обработка семян этими препаратами, а также ТМТД.
АЛЬТЕРНАРИОЗ, ПЛЕСЕНЬ И УСЫХАНИЕ ВЕТВЕЙ
Серая плесень, или гниль (возбудитель – гриб Botrytis cinerea) и альтернариоз (возбудители – грибы
 рода Alternaria) поражают надземные части молодых растений можжевельника, туи. Побеги становятся серо - коричневыми или черноватыми, будто слоем пыли покрытыми конидиями, которые перезаражают растения в течение вегетации. Растения ослабляются, теряется их декоративность.
рода Alternaria) поражают надземные части молодых растений можжевельника, туи. Побеги становятся серо - коричневыми или черноватыми, будто слоем пыли покрытыми конидиями, которые перезаражают растения в течение вегетации. Растения ослабляются, теряется их декоративность.
Факторы риска. Эти заболевания особенно часто развиваются на непроветриваемых участках при сильном загущении посадок и недостаточном освещении.
Меры защиты. В качестве защитных мер рекомендуется своевременная прореживающая обрезка и вырезка пораженных ветвей, дезинфекция всех срезов раствором медного купороса и обработка их масляной краской на натуральной олифе или замазкой типа раннет. Эффективны профилактические опрыскивания весной и осенью бордоской смесью, абига-пик, скором, чистоцветом. При сильном поражении в летнее время опрыскивания повторяют.
У туи и можжевельника часто возникает инфекционное усыхание ветвей. Вызывается оно несколькими возбудителями из отдела анаморфные грибы. Кора усыхает, и на ней образуются многочисленные плодовые тела – пикниды, бурого и черного цвета в виде точек и бугорков. Хвоя желтеет и опадает, ветви кустов буреют и усыхают. Инфекция сохраняется в коре пораженных ветвей и неубранных растительных остатках. Развитию болезни способствуют загущенные посадки растений и использование зараженного посадочного материала. Меры борьбы аналогичны защите от серой гнили.
ШЮТТЕ – БУРОЕ, СНЕЖНОЕ, НАСТОЯЩЕЕ
У хвойных есть заболевания, характерные только для этих пород. В первую очередь, это шютте, возбудителями которого являются некоторые виды грибов- аскомицетов.
На можжевельнике признаки поражения шютте (возбудитель – Lophodermium juniperinum) появляются в начале лета на прошлогодней хвое, приобретающей грязно-желтую или бурую окраску. С конца лета на поверхности хвоинок заметны круглые черные до 1,5 мм плодовые тела (апотеции), в которых сохраняется сумчатое спороношение гриба. Болезнь интенсивно развивается на ослабленных растениях, во влажных условиях может привести к их гибели.
Бурое шютте, или бурая снежная плесень (грибы рода Herpotrichia), кроме можжевельника, поражает сосны, пихту, ели, кедры, кипарисовик, туи. Встречается чаще в питомниках, молодняках, на самосеве и молодом подросте. Болезнь развивается под снегом при температуре не ниже 0,5°С. Поражение обнаруживается после схода снега: на бурой отмершей хвое заметен черно-серый паутинистый налет мицелия, а затем и точечные плодовые тела гриба-возбудителя. Хвоя долго не опадает, тонкие ветви отмирают.
 |
 |
| Шютте на ели | Хвоя ели с мицелием возбудителя шютте |
Факторы риска. Развитию болезни способствуют высокая влажность, наличие впадин на посевных площадях, загущенность растений. Вредоносность шютте возрастает при высоком снежном покрове и его длительном таянии.
Настоящее шютте, возбудителем которого является гриб Lophodermium seditiosum – одна из главных причин преждевременного опадения хвои у сосны. В основном поражаются молодые растения, в т.ч. в открытом грунте питомников, и ослабленные деревья, что может привести к их отмиранию из-за сильного опадания хвои. В течение весны и раннего лета хвоя становится бурой и опадает. Осенью на хвое заметны маленькие желтоватые точки, постепенно разрастающиеся и буреющие; позд-нее на отмершей осыпавшейся хвое образуются точечные черные плодовые тела – апотеции, которыми гриб сохраняется.
Сходные симптомы и цикл развития имеет гриб Lophodermium pinastri, возбудитель обыкновенного шютте сосны. Осенью или чаще весной следующего года хвоя желтеет или становится красновато-бурой и отмирает. Затем на хвоинках образуются плодовые тела гриба в виде мелких чёрных штрихов или точек, чернеющие и увеличивающиеся к осени.
Факторы риска. Рассеиванию спор и заражению хвои способствует умеренно теплая погода, моросящие дожди и росы. Чаще поражаются и погибают ослабленные растения в питомниках и хвойные до 3-летнего возраста, а также самосев сосны.
Снежное шютте вызывается грибом Phlacidium infestans, поражающим в основном виды сосны. Особенно он вредоносен в многоснежных районах, где иногда полностью уничтожает возобновление сосны обыкновенной; развивается под снежным покровом, даже при температуре около 0°С. Мицелий растет от хвоинки к хвоинке и к соседним растениям. После таяния снега погибшая хвоя и нередко побеги буреют и отмирают, покрываются сероватым, быстро исчезающим налетом мицелия. В течение лета хвоя отмирает, становится красновато- рыжей, позднее светло-серой. Она крошится, но почти не опадает. К осени на ней становятся видны плодовые тела – апотеции, в виде маленьких темных точек. Аскоспоры из них распространяются воздушными течениями на живую хвою непосредственно перед установлением снежного покрова.
Факторы риска. Развитию гриба благоприятствуют моросящие дожди, выпадение и таяние снега осенью, мягкая многоснежная зима, затяжная весна.
Защитные меры от шютте должны выполняться в комплексе. Необходимо удалять опавшую больную хвою; по возможности, отряхивать снег с нижних веток. Не допускается произрастание вблизи питомника даже отдельных взрослых деревьев сосны, ели. Хотя сейчас во главу угла ставится сохранение здоровья растений с помощью экологически и экономически обоснованных мер защиты, фунгицидные обработки против шютте в питомниках применяют обязательно. Опрыскивания медьсодержащими препаратами, скором, чистоцветом в течение лета эффективно снижают развитие заболеваний.
Наиболее подвержены шютте затененные и ослабленные экземляры, поэтому необходимо придать растениям как можно большую устойчивость, что возможно при более широком использовании иммуномодуляторов. Эффективно сочетание фунгицидных обработок с биологически активными препаратами и микроудобрениями.
Вредоносность шютте довольно сильно различается по способности к заражению определенных видов и сортов, поэтому необходимо иметь информацию о таких устойчивых формах, отдавая им предпочтение при посадке.
В тех местностях, где шютте повреждает сосну обыкновенную, можно использовать сосну скрученную или ель европейскую, которые поражаются крайне редко. В лесах и парках, вместо естественного возобновления, рекомендуются посадки саженцев необходимого происхождения, они более равномерно распределяются по площади, затрудняя заражение мицелием одного растения от другого, и быстрее достигают высоты больше критического уровня.
ОПАСНЫЕ РЖАВЧИННЫЕ БОЛЕЗНИ
Особое значение для хвойных имеют ржавчинные заболевания, вызываемые грибами отдела Базидиомикота, класса Урединомицеты. Возбудители поражают чаще всего хвою и кору побегов, фактически все они являются разнохозяйными и с хвойных переходят на другие растения. Ниже приводим описание наиболее распространенных.
Ржавчину хвои сосны вызывают нескольких видов грибов рода Coleosporium. Они поражают в основном 2-хвойные виды сосен, главным образом в питомниках и молодняках. Эциостадия гриба развивается весной на хвое сосны в виде жёлтых пузыревидных пустул, расположенных в беспорядке на обеих сторонах хвоинок. При сильном распространении болезни хвоя преждевременно желтеет и опадает, а растения теряют декоративность. Урединио- и телиоспоры образуются на мать-и-мачехе, крестовнике, осоте, колокольчике и других травянистых растениях.
Смоляной рак сосны, рак серянка (Cronartium flaccidium и Peridermium pini). В развитии первого гриба участвуют промежуточные хозяева – мятлик болотный и недотрога, на листьях которых развиваются урединио- и телиостадии. Второй гриб распространяется только в эциальной стадии от сосны к сосне. Заражение дерева происходит через сучья, откуда мицелий распространяется в ствол. Грибы поражают кору молодых деревьев, либо вершины и ветви старых сосен, там, где гладкая и тонкая кора. Мицелий проникает в клетки древесины и смоляные ходы, разрушает их. Пораженная часть обильно пропитывается живицей и приобретает серовато-черную окраску. Развиваясь в клетках камбия, грибница прекращает рост древесины через 2–3 года после заражения.
Разнохозяйный гриб Cronartium ribicola вызывает сосновый вертун, пузырчатую ржавчину, или столбчатую ржавчину смородины. Вначале происходит заражение хвои, постепенно гриб распространяется в кору и древесину ветвей и стволов сосны кедровой, веймутовой (5-хвойных). Стволики сеянцев изгибаются. У более взрослых растений в местах поражения кора растрескивается, из разрывов наблюдается выделение смолы и выступают эции в виде желто-оранжевых пузырьков. Под воздействием грибницы образуется утолщение, вышележащая часть побега засыхает или искривляется. Промежуточным хозяином является смородина и крыжовник, у которых сильно поражаются листья.
 Возбудители ржавчины можжевельника (грибы рода Gymnosporangium) поражают кизильник, боярышник, яблоню, грушу, айву, которые являются промежуточными хозяевами. Весной заболевание развивается на листьях, вызывая образование желтоватых выростов (пустул) на их нижней стороне; а с верхней заметны круглые оранжевые с черными точками пятна (эциальная стадия). С осени, иногда и весной, на хвое и ветвях можжевельника появляются желто-оранжевые студенистые массы телиоспор гриба-возбудителя. Пораженные части побегов веретенообразно-вздутые.
Возбудители ржавчины можжевельника (грибы рода Gymnosporangium) поражают кизильник, боярышник, яблоню, грушу, айву, которые являются промежуточными хозяевами. Весной заболевание развивается на листьях, вызывая образование желтоватых выростов (пустул) на их нижней стороне; а с верхней заметны круглые оранжевые с черными точками пятна (эциальная стадия). С осени, иногда и весной, на хвое и ветвях можжевельника появляются желто-оранжевые студенистые массы телиоспор гриба-возбудителя. Пораженные части побегов веретенообразно-вздутые. Меры защиты. В качестве защитных мероприятий от ржавчинных заболеваний можно рекомендовать пространственную изоляцию от поражаемых растений, имеющих общего возбудителя болезни. Так, не стоит выращивать рядом с соснами тополь и осину, 5-хвойные сосны следует изолировать от посадок черной смородины.
Уменьшают распространенность болезней опрыскивания препаратами фитоспорин М, абига-пик. Проводят вырезку пораженных побегов. Повышение иммунитета растений, возможное за счет применения микроудобрений и иммуностимуляторов, существенно снижают вредоносность ржавчин.
www.ruspitomniki.ru
|
Хвойные растения являются самыми древними из существующих растений на нашей планете. Их возраст исчисляется сотнями миллионов лет. Эволюция практически не повлияла на анатомическое строение хвои и шишек. При сравнении листьев хвойных, которые в простонародье называют иголками, с листьями цветковых растений, можно заметить, что при относительном однообразии хвоинок они имеют различную форму, размеры, окраску, а у некоторых видов совсем не выглядят, как привычная хвоя. ХвояХвоя внешне походит на узкие игольчатые листья. Характерно наличие плотной кожицы, которую покрывает восковидная субстанция. Это необходимо для уменьшения испарения влаги голосеменными. К примеру, у ели хвоинки четырехгранные, но зачастую грани практически незаметны, и хвоинки выглядят уплощенными. Рисунок. Поперечный разрез хвои сосны обыкновенной Если разрезать хвоинку, она имеет форму неправильного ромба, причем самый плоский угол направлен книзу. Здесь расположена срединная жилка листа. Вдоль других граней иголочки заметны полосы белого цвета, образованные устьицами – дыхательными отверстиями, через которые осуществляется дыхание растений. Также устьица служат для испарения влаги, которую дерево впитывает из грунта даже в лютую стужу. Это объясняет тот факт, что ели, как и другие хвойные, нельзя пересаживать в осеннее время, так как корни не могут крепко укорениться, и вода практически не поднимается по стеблю к хвое, хотя дыхание происходит в том же режиме.
Важным отличием хвойных пород от лиственных заключается в том, что у них черешок прочно связан с веткой и остается на ней, даже после отмирания иголки. Иголки отпадают через 6-7 лет. Они хорошо защищены от воздействия неблагоприятных факторов внешней среды толстым слоем воскового налета – кутикулой. Причем у многих видов налет настолько толстый, что хвоинки приобретают голубой оттенок. У хвойных нет истинных плодов и цветов. Они принадлежат к отделу голосеменных.Их семена прикреплены непосредственно к семенным чешуям, а те из них, которые собраны в женские шишки снабжены особыми крылышками. Покидая шишку, они планируют на крыльях, при вращении напоминая маленькие вертолетики. Это помогает им перемещаться подальше от материнского растения. Хвойные шишкиВнешний вид шишек хвойных разнообразен и специфичен. Они могут отличаться по длине, форме, размещению в пространстве, цвету, структуре и форме спорофиллов, способу распространения семян, т. д. Но принципиальная структура шишек одинакова. Все шишки в основании имеют ось, которая обособлена от вегетативной части дерева и является коротким побегом с расположенными на нем спороносными листьями – спорофиллами.
Различают женские и мужские шишки. Подавляющее большинство хвойных относятся к однодомным. У них и женские, и мужские шишки развиваются на одном растении. В большинстве случаев мужские шишки группами сосредоточены в пазухах листьев, иногда – на верхушках боковых побегов. Шишки женского пола отличаются компактным расположением, изредка они расположены поодиночке. Похожие материалы: |


beaplanet.ru
Строение древесины хвойных пород
Древесина хвойных пород отличается довольно простым и правильным строением.
В центре ствола хвойных пород находится так называемая сердцевина, имеющая вид неправильного круга диаметром 1—4 мм. Сердцевина состоит из паренхимных тонкостенных и одревесневших клеток. В молодом деревце или вершинном побеге, где еще нет законченной дифференциации тканей древесины, клетки сердцевины выполняют функции запасающих тканей и содержат крахмал, жиры и другие органические продукты. По мере роста дерева и выделения специализированных тканей, надобность в запасающих клетках сердцевины отпадает, они отмирают и заполняются воздухом.
На поперечном и продольном разрезах видно, что у сосны, лиственницы и кедра центральная часть ствола отличается от периферической более темным цветом, а в свежесрубленном состоянии, кроме того, и меньшим содержанием воды.
Темноокрашенная центральная часть древесины называется ядром, а периферическая светлая — заболонью.
У ели и пихты центральная часть ствола отличается от периферической только меньшим содержанием воды и называется спелой древесиной.
Ядровая и спелая часть древесины у растущих деревьев выполняет механические функции, не участвуя в обмене веществ.
С возрастом деревьев процентное содержание ядра или спелой древесины увеличивается.
В условиях Севера протяжение заболони по радиусу деревьев в возрасте 140 лет (на высоте 1,3 м) неодинаково для различных пород.
Ядро и спелая древесина образуются тогда, когда окаймленные поры трахеид закупориваются и выключаются из водопроводящей системы дерева, а клеточные оболочки пропитываются особыми ядерными веществами.
Водопроводящие и механические функции у хвойных деревьев выполняют трахеиды, они являются основным элементом древесины и занимают 90—95% общего ее объема.
Трахеиды расположены радиальными рядами и на поперечном срезе имеют вид четырехугольных или шестиугольных клеток, а на радиальном и тангентальном разрезах — вид волокон с косо срезанными концами. Радиальные ряды состоят из повторяющихся в каждом годичном кольце двух типов трахеид — ранних и поздних. Ранние трахеиды, выполняя проводящие функции, имеют широкую внутреннюю полость и тонкие стенки с многочисленными порами. Поздние трахеиды, образующие так называемую позднюю древесину, выполняют механические функции, поэтому имеют утолщенные стенки и сравнительно узкие полости.
Характерная особенность трахеид — окаймленные поры, сгруппированные преимущественно на радиальных стенках у концов, которыми каждая трахеида вклинивается между соседними, образуя плотное соединение.
Окаймленные поры — это не простые отверстия, соединяющие полости двух соседних клеток, с поверхности они прикрыты тонким первичным слоем клеточной оболочки. Поры соседних трахеид всегда находятся одна против другой и разделены общей для двух клеток срединной пластинкой (мембраной). Следовательно, окаймленная пора образуется двумя соседними трахеидами. Мембрана окаймленной поры в центре имеет утолщение, называемое торусом. Над первичным слоем и торусом вторичные слои соседних клеток нависают в виде купола, как бы окаймляя их, отсюда и название — окаймленная пора.
При выходе из полости клеток диаметр пор примерно равен 4—5 μ, затем сильно расширяется и в месте примыкания к срединной пластинке достигает 12—25. Диаметр торуса всегда несколько превышает внутренний диаметр поры.
Первичные слои клеток, образующие мембрану окаймленной поры, в своей неутолщенной части имеют тончайшие отверстия, облегчающие проникновение веществ из клетки в клетку. В живых клетках эти мельчайшие отверстия заполнены плазматическими нитями, при помощи которых плазма всех живых клеток организма соединена в одно целое.
При отклонении мембраны в ту или другую сторону торус закрывает выходное отверстие из полости клетки, прекращая этим дальнейшее функционирование поры.
У трахеид, расположенных в зоне ядра или спелой древесины, подавляющее большинство пор закупорено торусом, и они выключаются из водопроводящей системы. В заболони число пор, закрытых торусом, незначительно, и практически они не влияют на передвижение воды в древесине.
Число окаймленных пор в трахеидах довольно велико. Так, количество их у ранних трахеид 80—100, а у поздних — 10—20.
Размеры трахеид у хвойных деревьев изменяются не только в зависимости от породы, но также и от возраста, условий местопроизрастания и других факторов. Основываясь на обширных исследованиях ученых, можно принять для сосны, ели, лиственницы и пихты следующие размеры трахеид: длина 2—4 мм, ширина или наружный диаметр 0,2—0,04 мм, толщина стенок 0,005—0,008 мм. У поздних трахеид длина и толщина стенок больше, а ширина меньше.
Клеточные стенки трахеид состоят из целлюлозы, лигнина, гемицеллюлоз и полиуронидов. Полости этих клеток заполнены водой и воздухом.
Вторым элементом хвойных пород являются сердцевинные лучи, идущие между трахеидами в строго радиальном направлении.
Длина сердцевинных лучей увеличивается с каждым новым вегетационным периодом, по мере удаления камбиального слоя от центра ствола.
Сердцевинные лучи состоят из паренхимных живых клеток и предназначены для хранения питательных веществ. Клетки сердцевинных лучей значительно меньше трахеид, длина их примерно 0,04—0,08 мм, ширина 0,01—0,03 мм. Сердцевинные лучи у хвойных пород по ширине обычно образуются одним рядом клеток, если же в сердцевинном луче залегает горизонтальный смоляной ход, — то и несколькими рядами.
По высоте в луче всегда несколько рядов клеток: у сосны, кедра, лиственницы и ели верхний и нижний ряды образованы горизонтальными трахеидами с мелкими окаймленными порами, внутренние ряды — паренхимными клетками с простыми порами.
Выполняя функции запасающих тканей, сердцевинные лучи должны быть постоянно связаны с лубом, по которому идут питательные вещества от листьев. Эта связь осуществляется через камбий, который образует сердцевинные лучи как в сторону древесины, так и в сторону луба.
Стенки клеточной оболочки сердцевинных лучей сравнительно тонки и состоят в основном из целлюлозы, что делает их непрочными в механическом отношении. Полости клеток заполнены протопластом, содержащим в себе живую часть — протоплазму и ядро, а также неживые включения вроде зерен крахмала, жиров и т. п.
Протоплазма клеток сердцевинных лучей через ультрамикроскопические отверстия в мембране пор соединена в одно целое, а также соединена с протоплазмой клеток камбиального слоя и живых клеток луба. Благодаря этому клетки сердцевинных лучей могут выполнять функцию запасающих тканей и передавать запасы органических веществ камбию и другим живым клеткам.
В ядровой и спелой древесине сердцевинные лучи, как и все другие ткани, отмирают и перестают выполнять запасающие функции.
У хвойных пород, кроме сердцевинных лучей, есть еще одна ткань, которая выполняет запасающие функции, — это древесная паренхима. Клетки древесной паренхимы разбросаны между трахеидами или сгруппированы около смоляных ходов, стенки их сравнительно тонки и состоят из целлюлозы, гемицеллюлоз и лигнина.
В ядровой и спелой древесине клетки паренхимы отмирают и заполняются воздухом и водой. Количество древесной паренхимы в хвойных породах незначительно и обычно не превышает 1 % всей древесины.
Характерной особенностью древесины хвойных пород являются смоляные ходы, за исключением пихты, у которой они наблюдаются только иногда и то единично. Каждый ход состоит из канала, образуемого паренхимными клетками путем их дифференциации.
Смоляные ходы бывают вертикальные, тянущиеся параллельно оси ствола, и горизонтальные, проходящие по сердцевинным лучам.
Диаметр вертикальных смоляных ходов в среднем у сосны равен 0,1 мм, у ели и лиственницы — 0,06 мм. Длина ходов колеблется от 10 до 100 см.
Горизонтальные смоляные ходы имеют диаметр 0,03—0,04 мм и длину не выше сердцевинного луча. На 1 см2 поперечного среза сосны и ели насчитывается от 200 до 300 вертикальных смоляных ходов.
Горизонтальные смоляные ходы своими каналами всегда соединены с вертикальными, так что вся система образует одно целое. Такое строение дает возможность обильно заливать смолой ранения в любой части ствола.
Кроме того, через горизонтальные смоляные ходы живые клетки соединены с другими живыми тканями дерева.
В ядровой и спелой древесине горизонтальные смоляные ходы зарастают тиллами, часть ходов в ядровой древесине перестает функционировать и отключает от общей системы вертикальные смоляные ходы.
Смоляные ходы в заболони, вследствие смоляного давления, способны выделять терпентин непосредственно после их перерезания. Смоляные ходы ядровой древесины этим свойством не обладают, и терпентин в них находится как бы в хранилище.
При растрескивании древесины или других разрушениях смоляных ходов в ядре терпентин, разливаясь, не кристаллизуется, а пропитывает мертвые клетки. Как показывают исследования, смола и терпены обладают слабой токсичностью по отношению к дереворазрушающим грибам, поэтому большая стойкость смолистой древесины против гниения объясняется не столько антисептическими, сколько физическими их свойствами.
Смоляные ходы располагаются, главным образом, в позднем слое годичного кольца.
Обобщая все вышеизложенное, можно сделать следующее заключение о строении древесины хвойных пород.
Ткани, составляющие древесину, располагаются в стволе от периферии к центру в определенном порядке. Снаружи ствол покрывает пробковый слой коры, состоящий из мертвых пробковых и пробковидных клеток, очень устойчивых против микроорганизмов и насекомых. Клетки пробкового слоя у сосны и лиственницы содержат воды около 20%, у ели — 30%. К пробковым клеткам примыкает очень тонкий слой феллодермы, состоящий из клеток пробкового камбия, а также паренхимных клеток коры. Затем идет лубяной слой, состоящий из ситовидных трубок, сердцевинных лучей, лубяных волокон и лубяной паренхимы.
В феллодерме и лубе много живых клеток, заполненных плазмой и содержащих жиры и крахмал. Ситовидные трубки заполнены органически ми веществами, идущими от листьев. Все это делает феллодерму и луб весьма благоприятным питательным материалом для развития микроорганизмов и насекомых. Клетки луба и феллодермы имеют в среднем влажность у ели 141%, у сосны 152, у лиственницы 200% от абсолютно сухого веса; при высыхании они изменяют объем, что приводит к растрескиванию луба и постепенному отпаду коры.
За лубом идет камбиальный слой, состоящий из живых клеток, при жизни очень стойких против грибов, а в отмершем-состоянии, после рубки дерева, являющихся хорошей питательной средой для микроорганизмов.
За камбием следуют слои собственно древесины. В заболони, площадь которой колеблется в широких пределах в зависимости от диаметра деревьев, 50—60% клеток первого годичного слоя, считая от камбия, — живые, содержат протопласт, а также некоторые другие органические вещества. В последующих годичных кольцах заболони количество живых клеток резко уменьшается, они сохраняются в сердцевинных лучах, в смоляных ходах и в древесной паренхиме около лучей и смоляных каналов.
Следовательно, весь поверхностный слой заболони — это весьма хорошая питательная среда для микроорганизмов, в последующих слоях эти благоприятные условия для развития грибов остаются только в сердцевинных лучах и клетках древесной паренхимы.
Ядровая и спелая древесина состоит целиком из мертвых клеток, полости которых заполнены воздухом, а стенки пропитаны смолистыми и ядерными веществами. Поэтому ядро и спелая древесина могут служить питательной средой только для более высокоорганизованных видов грибов — базидиомицетов.
В круглых лесных сортиментах, непосредственно после валки деревьев, древесина по всей боковой поверхности прикрыта корой и обнажена только с торцов. От поражения ее грибами до некоторой степени механически предохраняет пробковый слой коры, а камбиальный слой до отмирания клеток сохраняет способность противостоять грибам и препятствует их проникновению в древесину. Влажность заболони также неблагоприятна для развития грибов.
Таким образом, в теплое время года, в первые 10—15 дней после рубки, древесина в круглых лесных сортиментах не подвергается серьезным изменениям и при надлежащих защитных мероприятиях может долго сохранять свое качество. Это в известной мере относится к пиломатериалам и другим сортиментам, о чем будет подробно сказано в последующих разделах.
Если вы нашли ошибку, пожалуйста, выделите фрагмент текста и нажмите Ctrl+Enter.
www.activestudy.info










