Содержание
Двойное оплодотворение цветковых растений, Леннаучфильм, 1969
Фильм демонстрирует формирование яйцеклетки и вторичного ядра зародышевого мешка, а также пыльцевого зерна с вегетативной и генеративной клетками.
Серия Биология
Тема: Двойное оплодотворение цветковых растений.
«Леннаучфильм», автор сценария Л.Зильберберг, режиссер Е.Иванова, 1969 г.
Дительность: 0:9:30
Ещё в разделе
15.01.2012
Опыление и оплодотворение цветковых растений, НаучФильм, 1987
Знакомит с понятием «опыление» и способами опыления: перекрестное насекомыми и ветром, самоопыление; далее — оплодотворение и как оно происходит, развитие плода из семяпочки.
02.12.2015
Аскарида
Фильм знакомит с внешним и внутренним строением аскарид. Показаны возможные пути заражения яйцами аскариды. Ряд кадров посвящен циклу развития аскариды.
Показаны возможные пути заражения яйцами аскариды. Ряд кадров посвящен циклу развития аскариды.
22.11.2015
Жизнь клетки и взаимодействие ее с вирусом
Фильм посвящен основной единице жизни — клетке, а так же воздействиям факторов внешней среды.
22.11.2015
Как видят насекомые?
Как выглядит наш мир в представлении животных? Различают ли они цвета? Распознают ли очертания, формы? Как видят насекомые? На все эти вопросы ответит этот учебный фильм.
21.11.2015
Чем раковые клетки отличаются от здоровых?
Как растут раковые клетки? Как химиотерапия борется с раком и почему вызывает побочные эффекты? Джордж Зайдан объяснит, почему быстрое деление раковых клеток это не только их преимущество, но и главная слабость.
18. 11.2015
11.2015
Обмен веществ и энергии в клетке
Часть 1 состоит из трех фрагментов: «Пластический и энергетический обмен», «Роль ферментов и АТФ в процессах обмена веществ», «Подготовительный этап энергетического обмена». Часть 2 содержит два фрагмента: «Неполное окисление — гликолиз», «Полное (кислородное) окисление».
18.11.2015
Почва
Фильм рассказывает об образовании плодородной почвы, значении дождевых червей, кротов, мышей и сусликов в повышении плодородия почвы. На мультипликационной схеме показаны слои почвы. Также фильм знакомит с удобрением почвы, снегозадержанием и борьбой с образованием оврагов, мероприятиями по орошению и осушению почв.
18.11.2015
Пресмыкающиеся
Фильм состоит из нескольких фрагментов рассказывающих о змеях, ящерицах, черепахах.
18. 11.2015
11.2015
Простейшие
В фильме рассказывается о представителях Типа (на сегодняшний день Подцарства) Простейшие. Классы Корненожки, Радиолярии, Споровики, Инфузории
18.11.2015
Ракообразные
Фильм рассказывает о речном раке — представителе класса ракообразных, об особенностях внешнего строении раков, об их органах чувств, о среде обитания, питании и размножении речного рака, о представителях класса ракообразных — дафниях и циклопах. Уделено внимание особенностям строения тела, расположению глаз, дыханию, размножению этих животных
18.11.2015
Тип кишечнополостные
На примере гидры рассматривается строение, особенности размножения кишечнополостных. Фильм знакомит с представителями типа кишечнополостных -медузой, актинией, коралловым полипом.
Оценка:
Пока комментариев нет
Оплодотворение у животных и растений
Половое размножение организмов связано с их морфологической и физиологической половой дифференциацией (половой диморфизм) и половым процессом.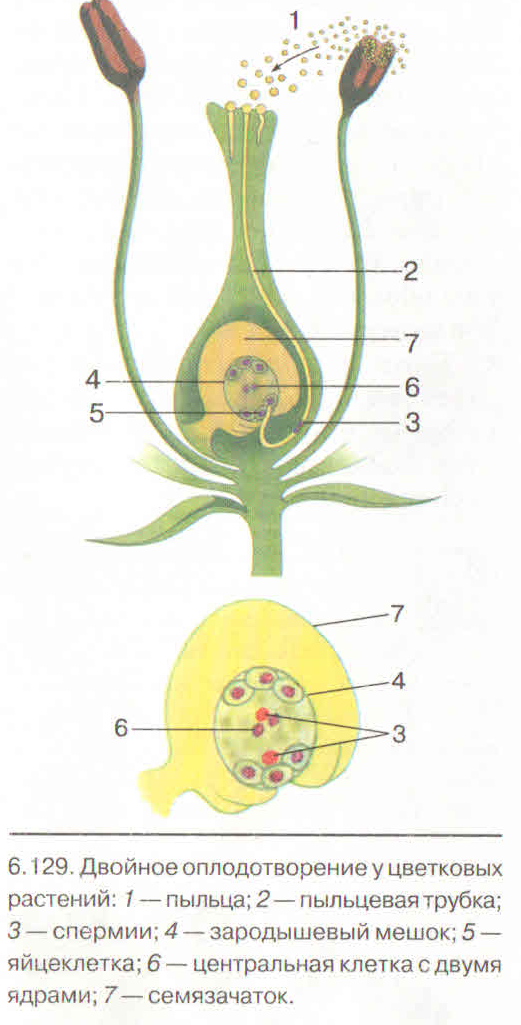
Половой процесс характеризуется системой приспособительных механизмов:
- образованием мужских и женских гамет,
- их слиянием в процессе оплодотворения (сингамия),
- объединением ядер (кариогамия),
- синаупсисогомологивдных хромосом в мейозе и перекомбинацией наследственных факторов.
Цикл полового размножения охватывает период от момента формирования половых клеток до их нового воспроизведения в следующем поколении.
Оплодотворением принято называть побуждение яйца к развитию в результате кариогамии. Оплодотворение представляет собой процесс необратимый — оплодотворенное однажды яйцо не может быть оплодотворено вновь. Сингамия и кариогамия составляют сущность процесса оплодотворения. Однако у некоторых видов воспроизведение нового поколения осуществляется на основе только женской гаметы — яйцеклетки без оплодотворения (девственное размножение). В этом случае половое размножение также заканчивается созреванием гамет. Оба эти способа размножения могут чередоваться у одного и того же вида.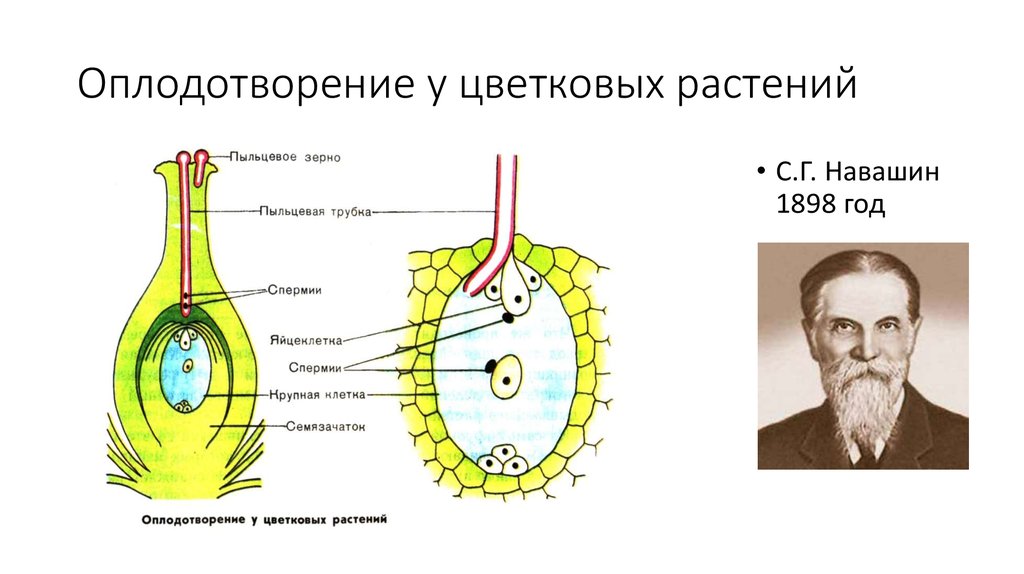
В процессе оплодотворения осуществляются следующие важные генетические явления, необходимые для существования вида:
- восстановление диплоидного набора хромосом, а в пределах диплоидного набора — парности гомологичных (материнских и отцовских) хромосом, разошедшихся в мейозе в процессе образования половых клеток у родительских организмов;
- обеспечение материальной непрерывности между следующими друг за другом поколениями;
- объединение в одном индивидууме наследственных свойств материнского и отцовского организмов.
Для обеспечения оплодотворения необходимо одновременное созревание гамет материнского и отцовского организма. У перекрестноопыляющихся растений созревание мужских и женских половых клеток может не совпадать во времени, и это несоответствие служит приспособительным механизмом, препятствующим самоопылению. Возможно, что несоответствие во времени созревания половых клеток у разных полов одного вида является одним из путей возникновения перекрестного опыления.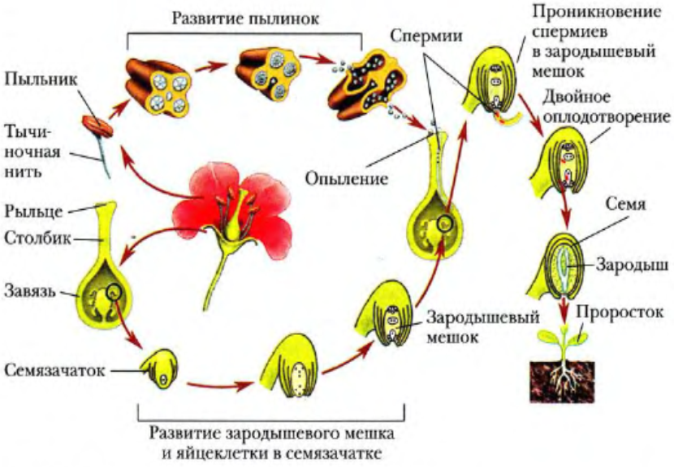
Оплодотворение у животных
Процесс оплодотворения у животных можно разделить на несколько фаз.
Первая фаза начинается с того, что сперматозоид либо прикрепляется к любой точке поверхности яйца, либо проникает в нее через микропиле. Момент соприкосновения головки сперматозоида с яйцом является начальным в цепи химических реакций. Эту фазу называют фазой активации яйца. В норме активацию яйца вызывают сперматозоиды своего вида. В некоторых случаях (у червя Rhabdites monohystera) сперматозоиды могут активировать яйцо, но при этом мужское ядро не сливается с материнским. Такое явление называют псевдогамным оплодотворением.
Вторая фаза процесса оплодотворения начинается после проникновения в яйцо, одного, а у некоторых животных и нескольких сперматозоидов. Проникший сперматозоид «готовится» к слиянию с женским ядром и последующему митозу: ядро сперматозоида постепенно набухает и приобретает вид интерфазного ядра. Такое ядро называют семенным, или мужским, пронуклеусом.
К моменту соприкосновения сперматозоида с яйцом и проникновения его внутрь ядро яйцеклетки у разных животных может находиться на разных стадиях деления созревания. Ядро яйцеклетки, готовое к слиянию с ядром сперматозоида, называют женским пронуклеусом. Собственно оплодотворение, т. е. слияние отцовского и материнского пронуклеусов, возможно лишь после окончания мейоза.
Проникновение сперматозоида может происходить на стадиях:
- ооцита I с покоящимся ядром
- ооцита I в стадии метафазы I
- ооцита II в стадиях мета — или анафазы II
- зрелой яйцеклетки
У иглокожих и кишечнополостных сперматозоид может проникать в яйцеклетку после завершения мейоза. Такое оплодотворение называют оплодотворением типа морского ежа. После проникновения сперматозоида в яйцо его ядро вскоре соединяется с женским ядром; ядро зиготы приступает к первому делению — дроблению яйца.
У бесчерепных (ланцетник) и всех позвоночных проникновение сперматозоида в яйцеклетку происходит, как правило, во время метафазы II.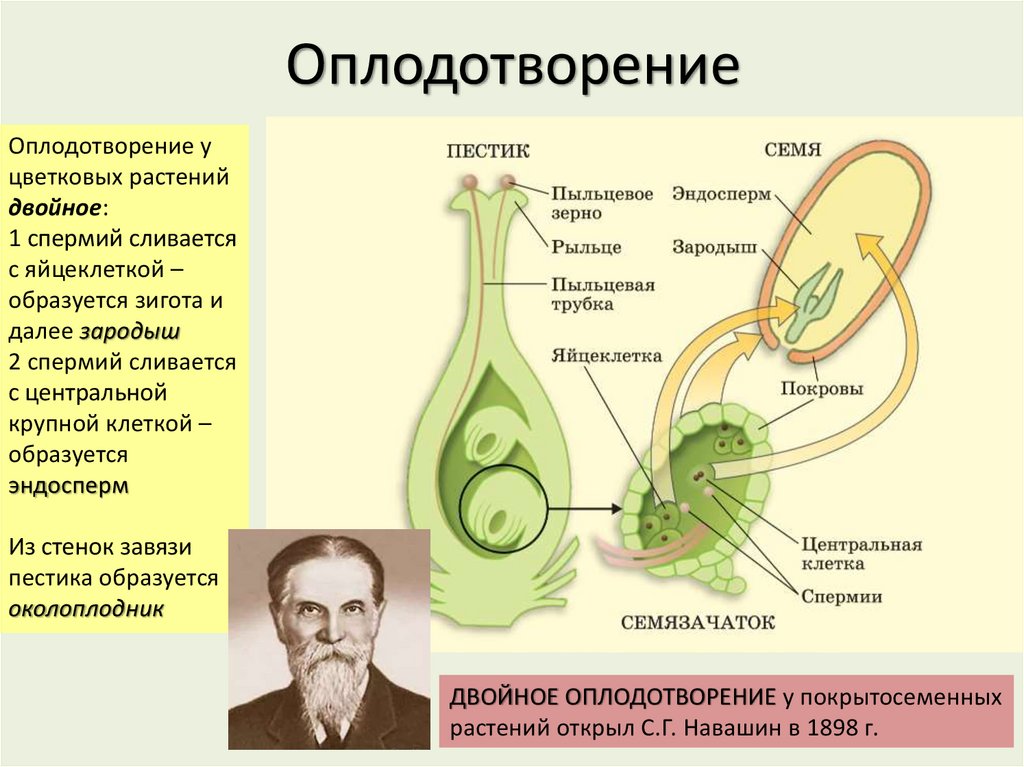 У асцидий, двустворчатых моллюсков и ряда других животных сперматозоид проникает в яйцеклетку на стадии метафазы I, а у губок, аскарид и некоторых других животных — на стадии ооцит I, т. е. до наступления мейоза. Этот тип оплодотворения называют типом аскариды. Проникший в цитоплазму яйца сперматозоид «ожидает» в стадии покоя окончания второго мейотического деления яйца.
У асцидий, двустворчатых моллюсков и ряда других животных сперматозоид проникает в яйцеклетку на стадии метафазы I, а у губок, аскарид и некоторых других животных — на стадии ооцит I, т. е. до наступления мейоза. Этот тип оплодотворения называют типом аскариды. Проникший в цитоплазму яйца сперматозоид «ожидает» в стадии покоя окончания второго мейотического деления яйца.
В акте оплодотворения два гаплоидных пронуклеуса сливаются в одно ядро. Кариогамия дает начало новому качественному процессу — развитию зиготы. Этот момент является кульминационным пунктом процесса полового размножения. В результате кариогамии, гомологичные хромосомы, разошедшиеся в мейозе предыдущего поколения, вновь воссоединяются в одном ядре зиготы.
Для понимания ряда важных генетических явлений необходимо знать, какие элементы сперматозоида проникают в яйцеклетку. Раньше считалось, что цитоплазма сперматозоида и ее органоиды не попадают в яйцеклетку. В настоящее время все больше накапливается фактов в пользу того, что в цитоплазму яйцеклетки у млекопитающих проникает не только головка (ядро) сперматозоида, но и его шейка и даже хвостовая часть. Если это подтвердится, то взгляды на роль цитоплазмы мужского организма в передаче его свойств потомству должны быть пересмотрены. Впрочем, генетических данных на этот счет пока нет; известны лишь факты передачи вирусных заболеваний.
Если это подтвердится, то взгляды на роль цитоплазмы мужского организма в передаче его свойств потомству должны быть пересмотрены. Впрочем, генетических данных на этот счет пока нет; известны лишь факты передачи вирусных заболеваний.
Вместе с ядром сперматозоида в цитоплазму яйцеклетки проникает центриоль, которая через некоторое время образует центросферу, дающую начало веретену дробления.
Приведенное общее описание оплодотворения у животных в деталях может варьировать у разных видов. Вследствие этих изменений процесс оплодотворения у каждого вида может протекать специфично, препятствуя межвидовому скрещиванию.
Оплодотворение у растений
У растений так же, как и у животных, сущность оплодотворения сводится к слиянию двух гаплоидных ядер.
Оплодотворение у растений в принципе сходно с таковым у животных, однако существование у растений гаметофита привело к появлению у них и некоторых особенностей.
Цитологический механизм этого процесса у голосеменных был создан русским ботаником Н.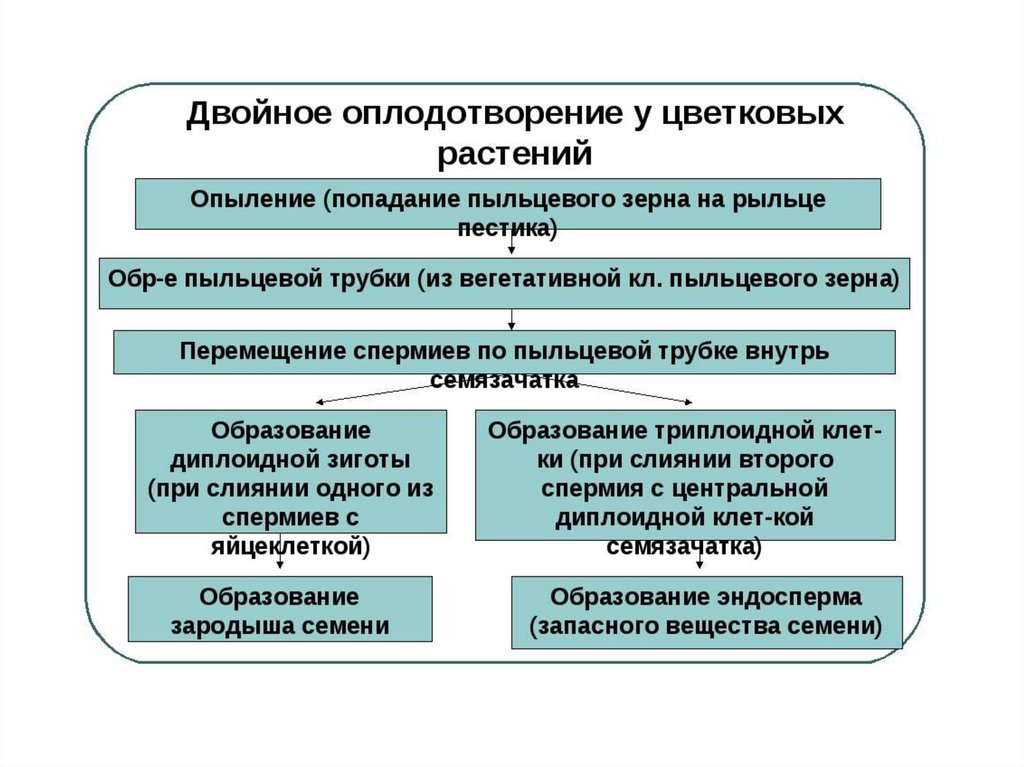 Н. Горожанкиным в 1880 г., а у покрытосеменных — Е. Страсбургером в 1884 г. Е. Страсбургер охарактеризовал оплодотворение у покрытосеменных следующим образом:
Н. Горожанкиным в 1880 г., а у покрытосеменных — Е. Страсбургером в 1884 г. Е. Страсбургер охарактеризовал оплодотворение у покрытосеменных следующим образом:
- процесс оплодотворения включает в себя слияние ядра мужской и женской гамет,
- цитоплазма гамет не имеет отношения к оплодотворению,
- ядро спермия и ядро яйцеклетки суть настоящие ядра.
Слияние спермия с ядром яйцеклетки и является собственно актом оплодотворения, в результате которого образуется зигота с диплоидным набором хромосом.
Выше было сказано, что микрогаметогенез завершается образованием двух спермиев, которые возникают или в пыльцевом зерне, или в пыльцевой трубке при прорастании пыльцевого зерна. Время начала прорастания зерен после попадания их на рыльце у разных растений варьирует в зависимости, от внешних условий и состояния рыльца и пестика. Так, например, у свеклы прорастание пыльцевых зерен начинается через 2 ч, у кок-сагыза — через 5 мин, а у кукурузы, сорго и других растений происходит почти немедленно.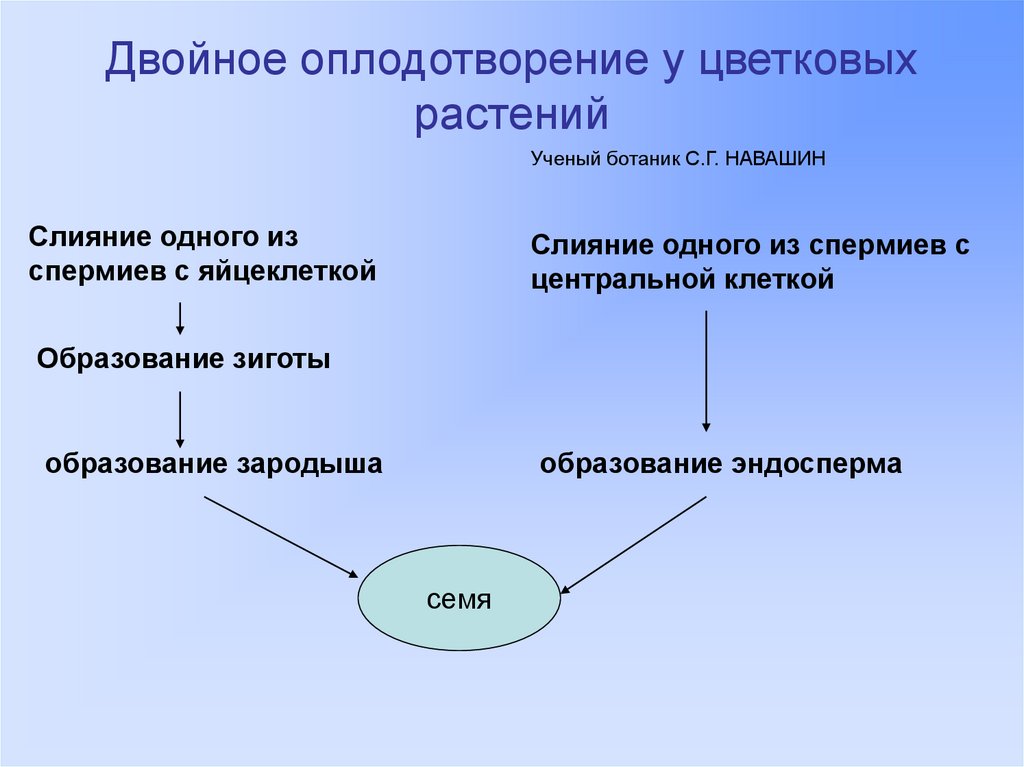
Первым признаком прорастания пыльцевого зерна является увеличение его объема. Обычно из одного пыльцевого зерна образуется одна трубка, но у некоторых растений (мальвовые, тыквенные) из одного зерна образуется несколько трубок, однако полного развития достигает лишь одна из них. Характер роста пыльцевых трубок определяется наследственными свойствами растений. К. Корренсом у MeiaridrTum (дрёма) было обнаружено, что при одновременном прорастании на рыльце нескольких пыльцевых зерен скорость роста пыльцевых трубок нередко зависит от их числа: чем больше их, тем медленнее они прорастают, при этом наблюдается конкуренция.
Пыльцевая трубка, дорастая до микропиле, приходит в соприкосновение с той частью зародышевого мешка, где находится яйцевой аппарат — яйцеклетка и синергиды. Впрочем, у некоторых растений пыльцевая трубка подходит к зародышевому мешку через халазальную часть семяпочки.
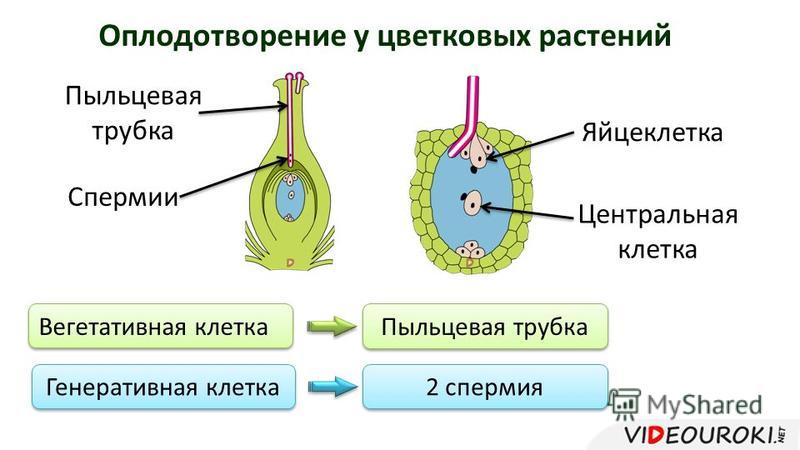
Передвигающиеся по пыльцевой трубке по мере ее роста два генеративных ядра — спермия после разрыва трубки вместе с ее содержимым попадают внутрь зародышевого мешка.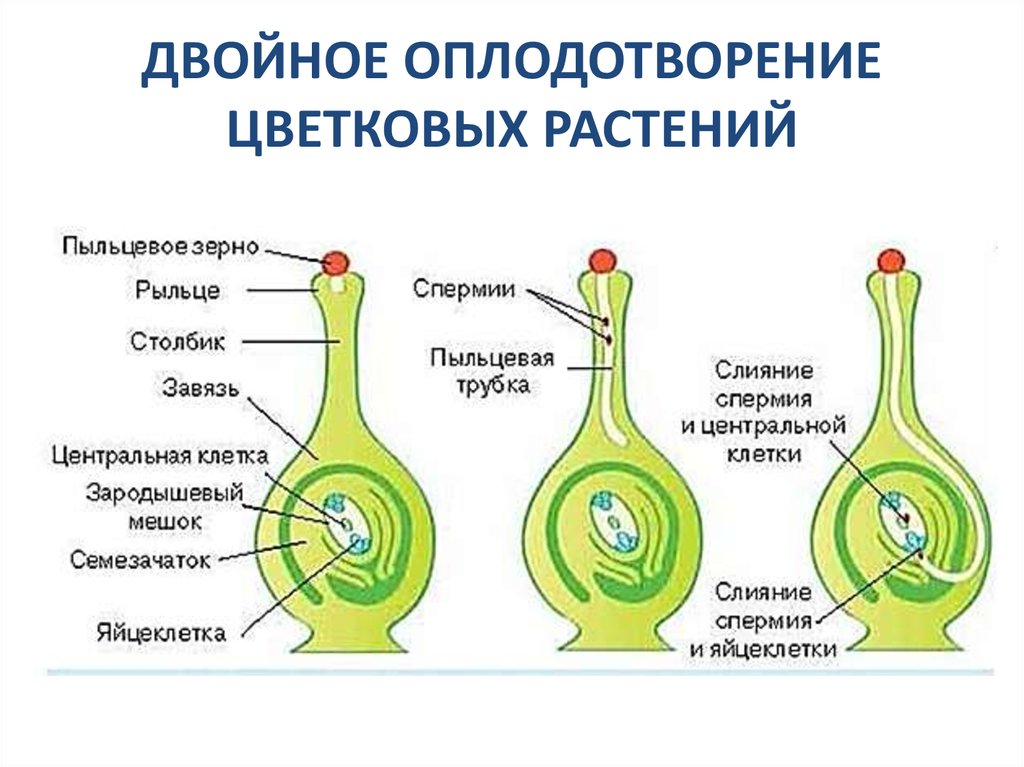 Спермин могут быть округлой, штопорообразной формы, иногда разрыхленные, с видимыми хромосомными нитями и др. Ядра их в этот момент, как правило, находятся в стадии телофазы. Из двух проникших в зародышевый мешок спермиев один спермий внедряется яйцеклетку и сливается с гаплоидным ядром последней. Слияние ядра спермия с ядром яйцеклетки является центральным моментом оплодотворения у растений.
Спермин могут быть округлой, штопорообразной формы, иногда разрыхленные, с видимыми хромосомными нитями и др. Ядра их в этот момент, как правило, находятся в стадии телофазы. Из двух проникших в зародышевый мешок спермиев один спермий внедряется яйцеклетку и сливается с гаплоидным ядром последней. Слияние ядра спермия с ядром яйцеклетки является центральным моментом оплодотворения у растений.
Схема двойного оплодотворения у растений
У растений так же, как и у животных, готовность к слиянию мужского и женского ядер может быть различной. Условно можно считать, что у растений имеются два типа оплодотворения: тип сложноцветных, аналогичный типу морского ежа у животных, и тип лилейных, аналогичный типу аскариды. В первом случае (тип сложноцветных) ядро спермия проникает в зрелую яйцеклетку в состоянии незавершенной телофазы, растворяет оболочку ядра яйцеклетки и переходит в интерфазное состояние. Во втором случае (тип лилейных) спермий проникает в яйцеклетку, находясь на стадии поздней телофазы. Ядро спермия не проникает в ядро яйцеклетки, а остается лежать рядом с ним. Каждое ядро в дальнейшем начинает подготавливаться к делению обособленно, и объединение их хромосом происходит только на стадии метафазы первого митотического деления зиготы. В оплодотворенной яйцеклетке — в зиготе восстанавливается диплоидное число хромосом. Из зиготы развивается зародыш семени.
Ядро спермия не проникает в ядро яйцеклетки, а остается лежать рядом с ним. Каждое ядро в дальнейшем начинает подготавливаться к делению обособленно, и объединение их хромосом происходит только на стадии метафазы первого митотического деления зиготы. В оплодотворенной яйцеклетке — в зиготе восстанавливается диплоидное число хромосом. Из зиготы развивается зародыш семени.
После оплодотворения у покрытосеменных растений развивается дополнительный эмбриональный орган — эндосперм, который представляет собой питательное депо зародыша. Начало развития эндосперма вторым оплодотворением. Второй спермий пыльцевой трубки, попадая в зародышевый мешок» сливается с диплоидным ядром центральной клетки зародышевого мешка. При этом образуется набор хромосом: два одинаковых набора хромосом материнского организма и один набор отцовского.
Слияние одного спермия с яйцеклеткой, а другого — с ядром центральной клетки называют двойным оплодотворением. Честь этого открытия, сделанного в 1898 г. , принадлежит нашему соотечественнику С. Г. Навашину. Триплоидная природа ядер эндосперма впервые была установлена у скерды (Crepis) М. С. Навашиным в 1915 г.
, принадлежит нашему соотечественнику С. Г. Навашину. Триплоидная природа ядер эндосперма впервые была установлена у скерды (Crepis) М. С. Навашиным в 1915 г.
Образование ткани, питающей зародыш, является особенностью растений. У животных эта функция возложена на запасные питательные вещества яйцеклетки и материнский организм, питающий зародыш через плаценту.
Одна из особенностей оплодотворения у растений, вытекающая из наличия у них двойного оплодотворения, представляет собой явление, называемое ксениями. Этот термин был предложен в 1881 г. В. Фоке. Смысл этого явления заключается в прямом влиянии пыльцы на признаки и свойства эндосперма. Например, имеются сорта кукурузы с желтым эндоспермом (желтые семена) и с белым эндоспермом (белые семена). Если женские цветки белозерного сорта опылить пыльцой желтозерного сорта, то, несмотря на то, что эндосперм развивается на растении белозерного сорта, окраска его будет желтой или бледно-желтой. Следовательно, ядро спермия способно изменить окраску эндосперма, ибо эта ткань так же как и ткань зародыша, гибридного происхождения.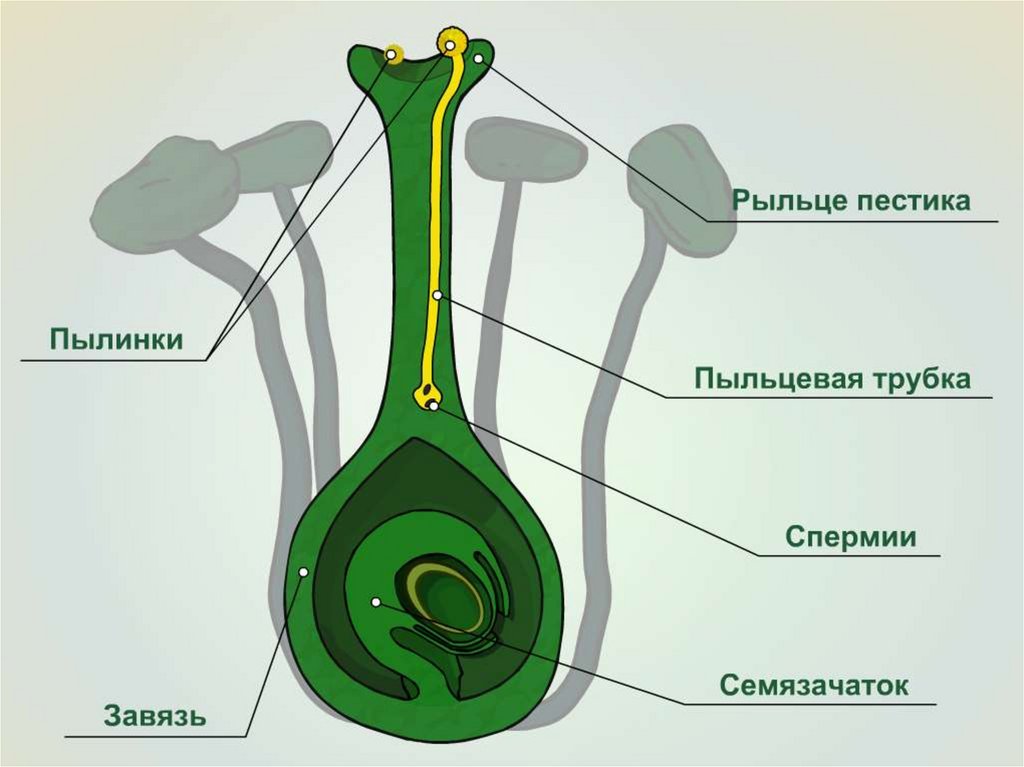
Таков в самых общих чертах процесс оплодотворения у животных и растений. Однако он подвержен приспособительным изменениям в зависимости от особенностей строения половых клеток и биологии размножения, свойственных каждому виду животных и растений.
Биология 2e, Структура и функции растений, Размножение растений, опыление и оплодотворение
После того, как пыльца отложится на рыльце пестика, она должна прорасти и пройти через столбик, чтобы достичь семязачатка. Микроспоры, или пыльца, содержат две клетки: клетку пыльцевой трубки и генеративную клетку. Клетка пыльцевой трубки вырастает в пыльцевую трубку, по которой проходит генеративная клетка. Для прорастания пыльцевой трубки требуются вода, кислород и определенные химические сигналы. Проходя через столбик и достигая зародышевого мешка, рост пыльцевой трубки поддерживается тканями столбика. Тем временем, если генеративная клетка еще не разделилась на две клетки, она теперь делится, образуя два спермия. Пыльцевая трубка направляется химическими веществами, выделяемыми синергидами, присутствующими в зародышевом мешке, и входит в семязачаточный мешок через микропиле.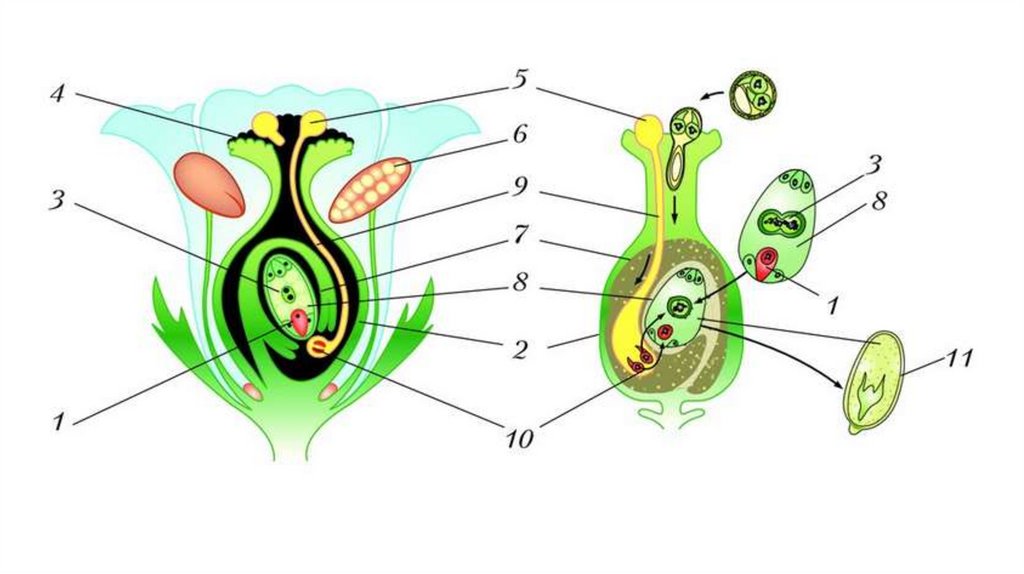 Из двух сперматозоидов один сперматозоид оплодотворяет яйцеклетку, образуя диплоидную зиготу; другой сперматозоид сливается с двумя полярными ядрами, образуя триплоидную клетку, которая развивается в эндосперм . Вместе эти два события оплодотворения у покрытосеменных известны как двойное оплодотворение (рисунок). После завершения оплодотворения никакие другие сперматозоиды не могут проникнуть внутрь. Оплодотворенная семяпочка образует семя, тогда как ткани завязи становятся плодом, обычно обволакивающим семя.
Из двух сперматозоидов один сперматозоид оплодотворяет яйцеклетку, образуя диплоидную зиготу; другой сперматозоид сливается с двумя полярными ядрами, образуя триплоидную клетку, которая развивается в эндосперм . Вместе эти два события оплодотворения у покрытосеменных известны как двойное оплодотворение (рисунок). После завершения оплодотворения никакие другие сперматозоиды не могут проникнуть внутрь. Оплодотворенная семяпочка образует семя, тогда как ткани завязи становятся плодом, обычно обволакивающим семя.
У покрытосеменных один сперматозоид оплодотворяет яйцеклетку с образованием зиготы 2 n , а другой спермий оплодотворяет центральную клетку с образованием эндосперма 3 n .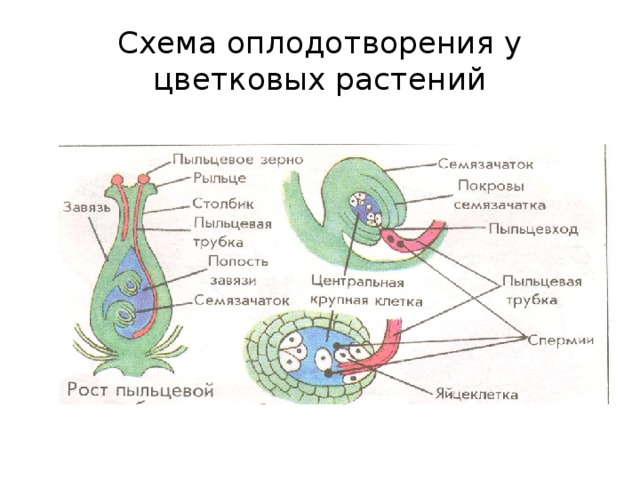 Это называется двойным оплодотворением.
Это называется двойным оплодотворением.
После оплодотворения зигота делится на две клетки: верхнюю, или терминальную, и нижнюю, или базальную. Деление базальной клетки дает начало суспензору , который в конечном итоге соединяется с материнской тканью. Суспензор обеспечивает транспортировку питательных веществ от материнского растения к растущему зародышу. Терминальная клетка также делится, давая начало проэмбриону шаровидной формы (рис. a ). У двудольных (эвдикотовых) развивающийся зародыш имеет форму сердца из-за наличия двух рудиментарных семядолей (рисунок б ). У неэндоспермных двудольных, таких как Capsella bursa , сначала развивается эндосперм, но затем он переваривается, и запасы пищи перемещаются в две семядоли. По мере увеличения зародыша и семядолей им не хватает места внутри развивающегося семени, и они вынуждены изгибаться (рис.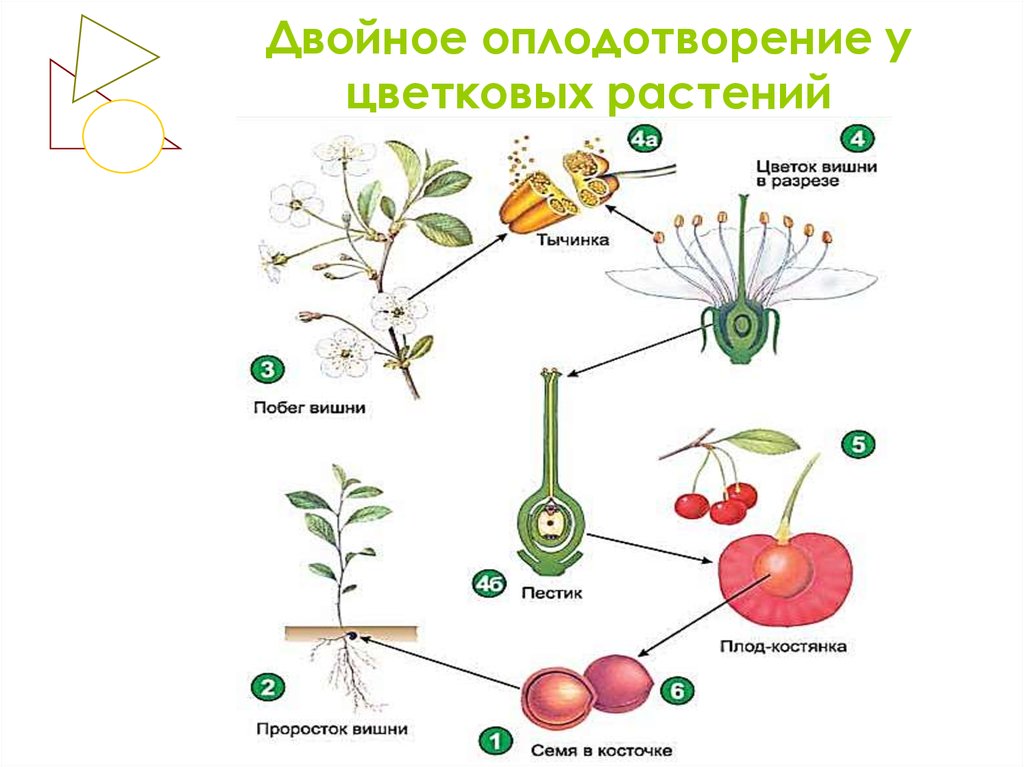 c ). В конечном итоге зародыш и семядоли заполняют семя (рис. d ), и семя готово к рассеиванию. Эмбриональное развитие через некоторое время приостанавливается, и рост возобновляется только при прорастании семени. Развивающийся саженец будет полагаться на запасы пищи, хранящиеся в семядолях, до тех пор, пока первый набор листьев не начнет фотосинтез.
c ). В конечном итоге зародыш и семядоли заполняют семя (рис. d ), и семя готово к рассеиванию. Эмбриональное развитие через некоторое время приостанавливается, и рост возобновляется только при прорастании семени. Развивающийся саженец будет полагаться на запасы пищи, хранящиеся в семядолях, до тех пор, пока первый набор листьев не начнет фотосинтез.
Показаны стадии развития эмбриона в семязачатке пастушьей сумки ( Capsella bursa ). После оплодотворения зигота делится на верхнюю терминальную и нижнюю базальную клетки. (а) На первой стадии развития терминальная клетка делится, образуя глобулярный проэмбрион. Базальная клетка также делится, давая начало суспензору. (б) На второй стадии развивающийся зародыш имеет форму сердца из-за наличия семядолей. (c) На третьей стадии растущий эмбрион выбегает из места и начинает изгибаться. (d) В конце концов, он полностью заполняет семя. (кредит: модификация работы Роберта Р. Уайза; данные масштабной линейки от Мэтта Рассела)
Развитие семени
Зрелая семяпочка превращается в семя. Типичное семя содержит семенную оболочку, семядоли, эндосперм и один зародыш (рисунок).
Типичное семя содержит семенную оболочку, семядоли, эндосперм и один зародыш (рисунок).
Art Connection
Показаны структуры семян двудольных и однодольных растений. Двудольные (слева) имеют две семядоли. Однодольные, такие как кукуруза (справа), имеют одну семядолю, называемую щитком; он направляет питание растущему эмбриону. Зародыши как однодольных, так и двудольных растений имеют перышко, образующее листья, гипокотиль, образующий стебель, и корешок, образующий корень. Эмбриональная ось включает в себя все, что находится между перышком и корешком, за исключением семядолей.
Какое из следующих утверждений верно?
- И однодольные, и двудольные растения имеют эндосперм.
- Корень развивается в корень.
- Перышко является частью эпикотиля.
- Эндосперм является частью зародыша.
Сохранение пищевых запасов в семенах покрытосеменных различается у однодольных и двудольных растений. У однодольных, таких как кукуруза и пшеница, одна семядоля называется щитком ; щитик соединяется непосредственно с зародышем через сосудистую ткань (ксилему и флоэму).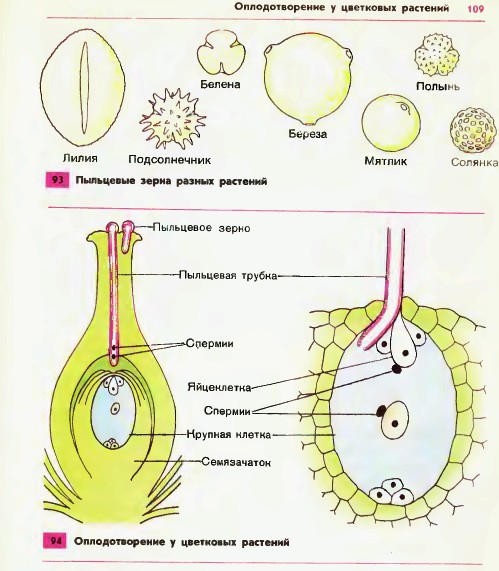 Запасы пищи хранятся в большом эндосперме. При прорастании ферменты выделяются алейрон , один слой клеток внутри семенной оболочки, которая окружает эндосперм и зародыш. Ферменты расщепляют запасенные углеводы, белки и липиды, продукты которых поглощаются щитком и транспортируются через сосудистую нить к развивающемуся эмбриону. Следовательно, щиток можно рассматривать как орган всасывания, а не запасающий орган.
Запасы пищи хранятся в большом эндосперме. При прорастании ферменты выделяются алейрон , один слой клеток внутри семенной оболочки, которая окружает эндосперм и зародыш. Ферменты расщепляют запасенные углеводы, белки и липиды, продукты которых поглощаются щитком и транспортируются через сосудистую нить к развивающемуся эмбриону. Следовательно, щиток можно рассматривать как орган всасывания, а не запасающий орган.
Две семядоли в семени двудольных также имеют сосудистые связи с зародышем. В эндоспермных двудольных запасы пищи хранятся в эндосперме. Таким образом, во время прорастания две семядоли действуют как поглощающие органы, чтобы поглощать высвобождаемые ферментами запасы пищи, как и у однодольных (у однодольных, по определению, также есть эндоспермные семена). Табак ( Nicotiana tabaccum ), томат ( Solanum lycopersicum ) и перец ( Capsicum annuum ) являются примерами эндоспермных двудольных растений.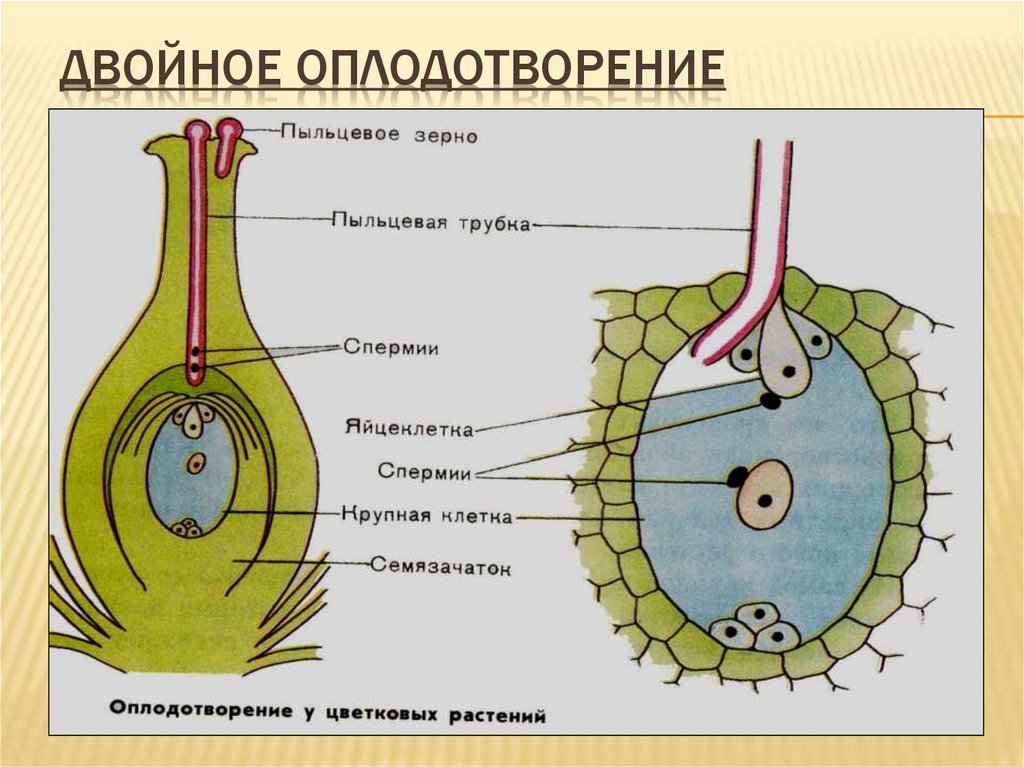 У неэндоспермных двудольных триплоидный эндосперм развивается нормально после двойного оплодотворения, но пищевые запасы эндосперма быстро ремобилизуются и перемещаются в развивающиеся семядоли для хранения. Две половинки семени арахиса ( Arachis hypogaea ) и лущеный горох ( Pisum sativum ) горохового супа представляют собой отдельные семядоли, загруженные пищевыми запасами.
У неэндоспермных двудольных триплоидный эндосперм развивается нормально после двойного оплодотворения, но пищевые запасы эндосперма быстро ремобилизуются и перемещаются в развивающиеся семядоли для хранения. Две половинки семени арахиса ( Arachis hypogaea ) и лущеный горох ( Pisum sativum ) горохового супа представляют собой отдельные семядоли, загруженные пищевыми запасами.
Семя вместе с семязачатком защищено семенной оболочкой, которая образуется из покровов семязачаточного мешка. У двудольных семенная оболочка далее делится на внешнюю оболочку, известную как теста , и внутреннюю оболочку, известную как тегмен .
Зародышевая ось состоит из трех частей: перышка, корешка и гипокотиля. Часть зародыша между точкой прикрепления семядолей и корешком известна как гипокотиль (гипокотиль означает «ниже семядолей»).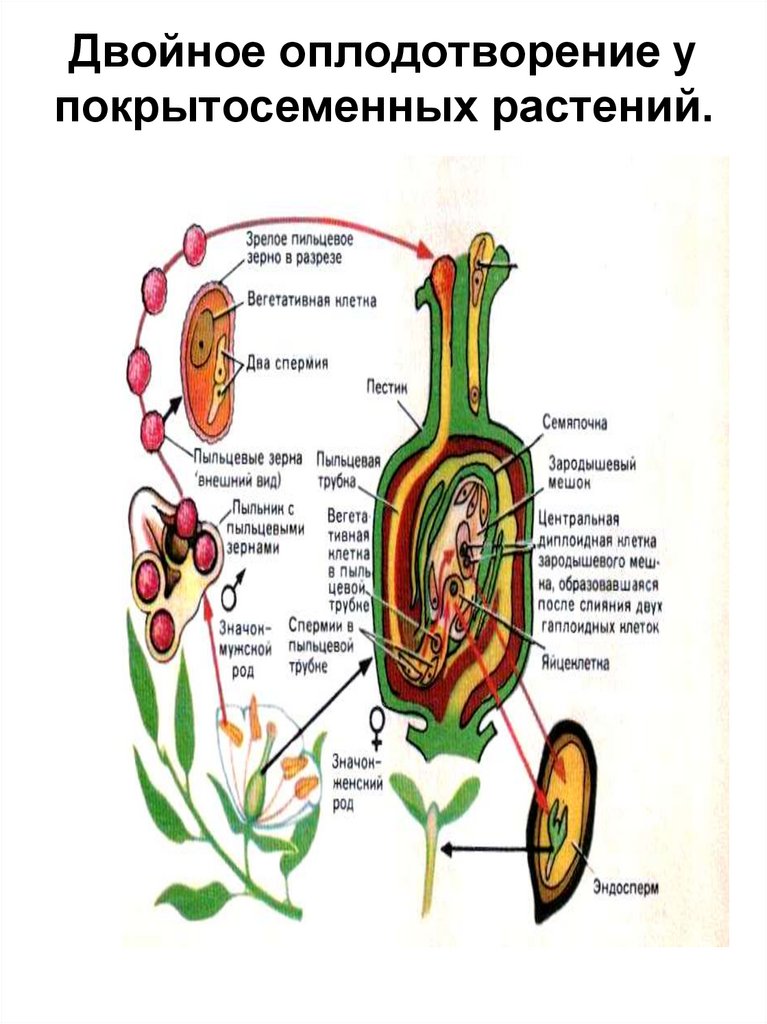 Зародышевая ось заканчивается корнем (эмбриональный корень), который представляет собой область, из которой разовьется корень. У двудольных гипокотильи выступают над землей, образуя стебель растения. У однодольных гипокотиль не виден над землей, потому что у однодольных нет удлинения стебля. Часть зародышевой оси, выступающая над семядолями, известна как эпикотиль . шлейф состоит из эпикотиля, молодых листьев и верхушечной меристемы побега.
Зародышевая ось заканчивается корнем (эмбриональный корень), который представляет собой область, из которой разовьется корень. У двудольных гипокотильи выступают над землей, образуя стебель растения. У однодольных гипокотиль не виден над землей, потому что у однодольных нет удлинения стебля. Часть зародышевой оси, выступающая над семядолями, известна как эпикотиль . шлейф состоит из эпикотиля, молодых листьев и верхушечной меристемы побега.
При прорастании семян двудольных эпикотиль имеет форму крючка с пером, направленным вниз. Эта форма называется перьевым крючком, и она сохраняется до тех пор, пока прорастание происходит в темноте. Поэтому, когда эпикотиль проталкивается через жесткую и абразивную почву, перышко защищено от повреждений. При воздействии света крючок гипокотиля расправляется, молодые лиственные листья обращены к солнцу и расширяются, а эпикотиль продолжает удлиняться.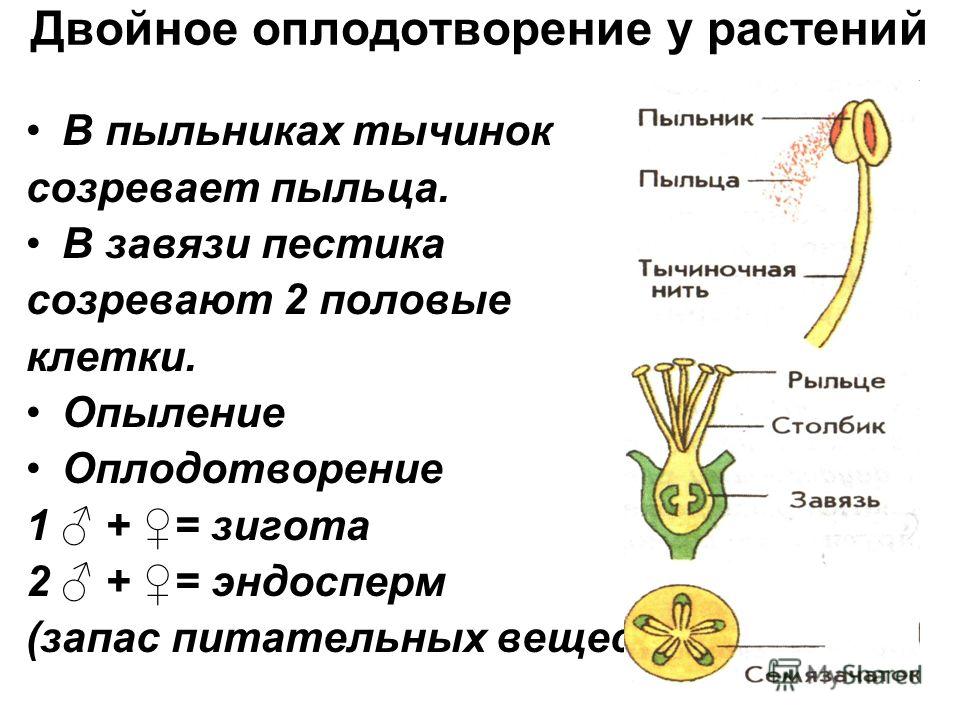 В это время корень также растет и дает первичный корень. По мере того, как он растет вниз, образуя стержневой корень, боковые корни разветвляются во все стороны, образуя типичную стержневую корневую систему двудольных растений.
В это время корень также растет и дает первичный корень. По мере того, как он растет вниз, образуя стержневой корень, боковые корни разветвляются во все стороны, образуя типичную стержневую корневую систему двудольных растений.
У семян однодольных растений (рисунок) раковина и надкрылье семенной кожуры сливаются. По мере прорастания семени появляется первичный корень, защищенный покрытием кончика корня: колеориза . Затем появляется первичный побег, защищенный колеоптилем : покрытием верхушки побега. При воздействии света (т. е. когда перышко вышло из почвы и защитный колеоптиль больше не нужен) удлинение колеоптиля прекращается, листья расширяются и разворачиваются. На другом конце зародышевой оси первичный корень вскоре отмирает, а из основания стебля появляются другие, придаточные корни (корни, не возникающие из обычного места, т.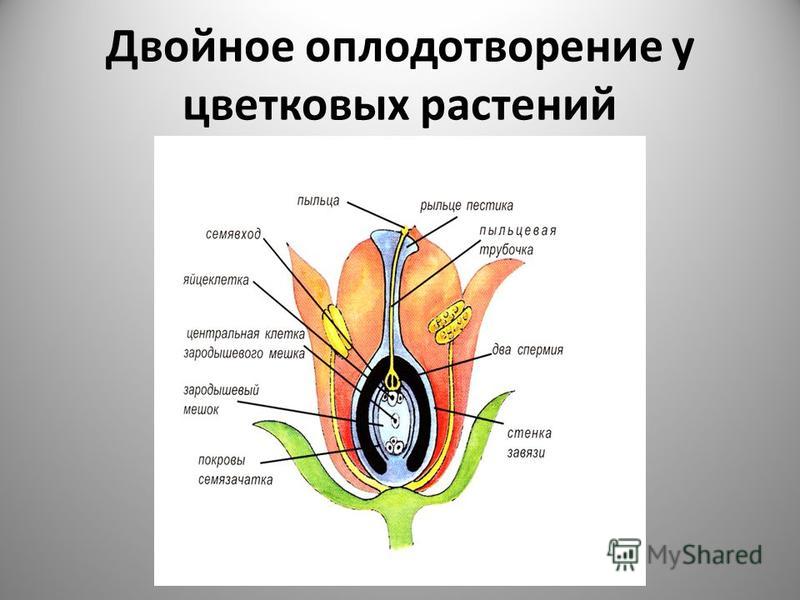 е. корень). Это дает однодольным растениям мочковатую корневую систему.
е. корень). Это дает однодольным растениям мочковатую корневую систему.
При прорастании этого семени однодольной травы первым появляется первичный корень, или корешок, за которым следует первичный побег, или колеоптиль, и придаточные корни.
Прорастание семян
Многие зрелые семена вступают в период бездействия или чрезвычайно низкой метаболической активности: процесс, известный как покой , который может длиться месяцы, годы и даже столетия. Покой помогает сохранить жизнеспособность семян в неблагоприятных условиях. При возвращении к благоприятным условиям происходит прорастание семян. Благоприятные условия могут быть такими разнообразными, как влажность, свет, холод, огонь или химическая обработка. После сильных дождей появляется много новых всходов. Лесные пожары также приводят к появлению новых саженцев. Некоторые семена требуют яровизация (обработка холодом) до того, как они прорастут.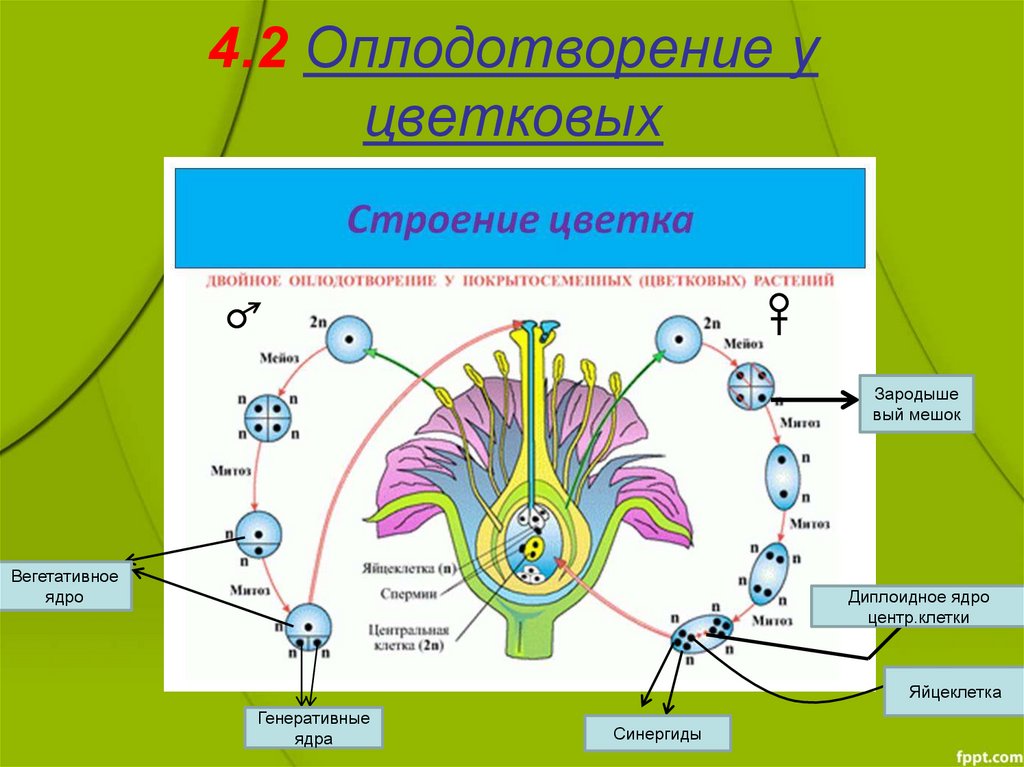 Это гарантирует, что семена, произведенные растениями в умеренном климате, не прорастут до весны. Растения, произрастающие в жарком климате, могут иметь семена, которые нуждаются в термической обработке для прорастания, чтобы избежать прорастания в жаркое и сухое лето. У многих семян наличие толстой семенной кожуры замедляет прорастание. Скарификация , которая включает механические или химические процессы для смягчения оболочки семян, часто применяется перед прорастанием. Также можно использовать предварительное замачивание в горячей воде или прохождение через кислую среду, например пищеварительный тракт животного.
Это гарантирует, что семена, произведенные растениями в умеренном климате, не прорастут до весны. Растения, произрастающие в жарком климате, могут иметь семена, которые нуждаются в термической обработке для прорастания, чтобы избежать прорастания в жаркое и сухое лето. У многих семян наличие толстой семенной кожуры замедляет прорастание. Скарификация , которая включает механические или химические процессы для смягчения оболочки семян, часто применяется перед прорастанием. Также можно использовать предварительное замачивание в горячей воде или прохождение через кислую среду, например пищеварительный тракт животного.
В зависимости от размера семян время появления всходов может варьироваться. У видов с крупными семенами достаточно запасов пищи, чтобы прорасти глубоко под землей, и при этом их эпикотиль простирается до поверхности почвы. Семена мелкосеменных видов обычно требуют света в качестве сигнала прорастания. Это гарантирует, что семена прорастут только на поверхности почвы или рядом с ней (где больше света). Если бы они прорастали слишком глубоко под поверхностью, у развивающегося ростка не было бы достаточных запасов пищи, чтобы добраться до солнечного света.
Если бы они прорастали слишком глубоко под поверхностью, у развивающегося ростка не было бы достаточных запасов пищи, чтобы добраться до солнечного света.
Биология, биологическое разнообразие, семенные растения, покрытосеменные растения
Фаза взрослой особи, или спорофита, является основной фазой жизненного цикла покрытосеменных растений (рисунок). Как и голосеменные, покрытосеменные разноспоровые. Следовательно, они генерируют микроспоры, которые будут генерировать пыльцевые зерна в качестве мужских гаметофитов, и мегаспоры, которые образуют семязачатки, содержащие женские гаметофиты. Внутри микроспорангиев пыльников мужские гаметофиты делятся путем мейоза с образованием гаплоидных микроспор, которые, в свою очередь, подвергаются митозу и дают начало пыльцевым зернам. Каждое пыльцевое зерно содержит две клетки: одну генеративную клетку, которая разделится на два спермия, и вторую клетку, которая станет клеткой пыльцевой трубки.
Показан жизненный цикл покрытосеменных растений.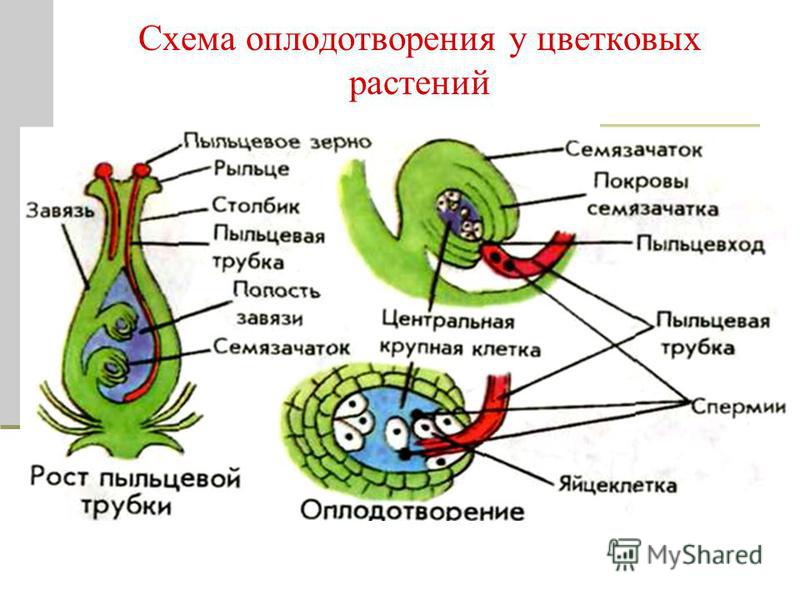 Пыльники и плодолистики — это структуры, которые укрывают настоящие гаметофиты: пыльцевое зерно и зародышевый мешок. Двойное оплодотворение — процесс, уникальный для покрытосеменных растений. (кредит: модификация работы Марианы Руис Вильярреал)
Пыльники и плодолистики — это структуры, которые укрывают настоящие гаметофиты: пыльцевое зерно и зародышевый мешок. Двойное оплодотворение — процесс, уникальный для покрытосеменных растений. (кредит: модификация работы Марианы Руис Вильярреал)
Если бы у цветка не было мегаспорангиев, какие гаметы не образовались бы? Если бы в цветке не было микроспорангиев, какие гаметы не образовались бы?
Семяпочка, защищенная завязью плодолистика, содержит мегаспорангий, защищенный двумя слоями покровов и стенкой завязи. В каждом мегаспорангии мегаспороцит подвергается мейозу, образуя четыре мегаспоры — три маленькие и одну большую. Выживает только крупная мегаспора; он производит женский гаметофит, называемый зародышевым мешком. Мегаспора делится три раза, образуя восьмиклеточную стадию. Четыре из этих клеток мигрируют к каждому полюсу зародышевого мешка; два подходят к экватору и в конечном итоге сливаются, образуя 2 n полярное ядро; три клетки, удаленные от яйца, образуют антиподы, а две ближайшие к яйцу клетки становятся синергидами.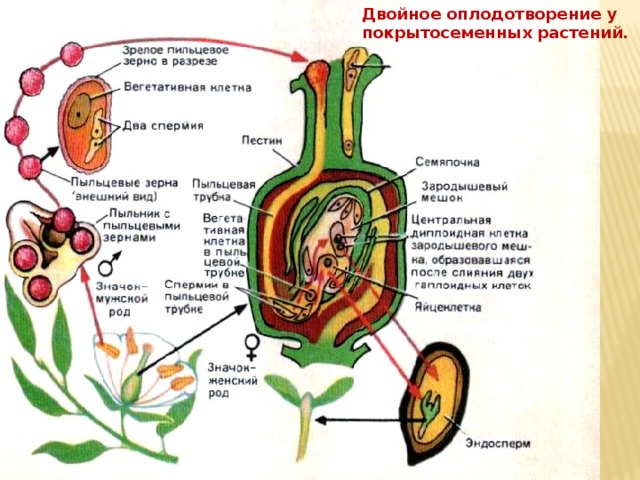
Зрелый зародышевый мешок содержит одну яйцеклетку, две синергиды или «хелперные» клетки, три антиподальные клетки и два полярных ядра в центральной клетке. Когда пыльцевое зерно достигает рыльца, пыльцевая трубка отходит от зерна, растет вниз по столбику и входит через микропиле: отверстие в покровах семязачатка. Два спермия откладываются в зародышевый мешок.
Затем происходит двойное оплодотворение. Один сперматозоид и яйцеклетка объединяются, образуя диплоидную зиготу — будущий эмбрион. Другой сперматозоид сливается с полярными ядрами 2 n , образуя триплоидную клетку, из которой развивается эндосперм — ткань, служащая запасом пищи. Зигота развивается в зародыш с корешком или небольшим корнем и одним (однодольные) или двумя (двудольными) листообразными органами, называемыми семядолями . Это различие в числе зародышевых листьев лежит в основе двух основных групп покрытосеменных: однодольных и эвдикотовых. Запасы пищи семени хранятся вне зародыша в виде сложных углеводов, липидов или белков.