
Строение цветка растения. Органы репродукции (пестики и тычинки), строение семязачтка. Семязачатки цветковых растений образуются
(жр) Семязачаток цветковых — Zbio
Материал из Zbio
А. Л. ТахтаджянСтроение семязачатков (семяпочек) цветковых растений во многом сходно со строением семязачатков голосеменных. Но в то время как у голосеменных преобладают в общем массивные семязачатки, обычно с толстыми, иногда даже грубыми покровами и со сравнительно большим запасом питательных веществ, у цветковых семязачатки обычно небольшие, часто очень мелкие, в большинстве случаев со слабо развитыми покровами и почти совершенно лишены запасов питательных веществ. Оказавшись в полости завязи, этой своеобразной влажной камере, семязачатки могут формироваться быстро и с минимальной затратой строительного материала, что оказалось исключительно важным биологическим преимуществом. Это обстоятельство особенно подчеркивал известный русский ботаник М. И. Го-лепкии (1927), который в книге «Победители в борьбе за существование» писал следующее: «Уменьшение затраты питательных и строительных веществ на семяпочки до оплодотворения только и позволило, весьма возможно, покрытосеменным развить необычайную, так сказать, деятельность в деле выработки приспособлений, с одной стороны, для защиты зародышей (в семенах и плодах), с другой — для расселения по поверхности земли. В отношении увеличения количества семян, органов размножения, стоит сравнить покрытосеменные опять-таки с голосеменными, например, какой-нибудь тополь с елью или сосной — у первого буквально неисчислимые количества семян, у вторых очень ограниченные. Про травянистые формы и говорить нечего». Очень хорошо сказано. Действительно, экономия материала, используемого на построение семязачатка, несомненно, позволила цветковым растениям существенно повысить семенную продуктивность, или урожайность. В то же время та же экономия материала привела не только к значительному упрощению семязачатка, но и к более быстрому его формированию, чем у голосеменных.
[править] ФОРМА И ОРИЕНТАЦИЯ СЕМЯЗАЧАТКОВ
Прежде чем перейти к строению семязачатка, рассмотрим его основные морфологические типы (рис. 25), установленные еще французским ботаником Ш. Мирбезем в 1829 г.

 Рис. 25. Различные формы семязачатков:1 - ортотропный; 2 - анатропный: 3 - гемитропный; 4 - кампилотропный; 5 - амфитропный.
Рис. 25. Различные формы семязачатков:1 - ортотропный; 2 - анатропный: 3 - гемитропный; 4 - кампилотропный; 5 - амфитропный. У многих цветковых растений микропиле семязачатка находится на одной оси с семяножкой (фуникулусом) и, таким образом, семязачаток расположен перпендикулярно к плаценте. Такой радиально-симметричный семязачаток был назван ортотропным (от греч. orthos — прямой, правильный и tropos — поворот, направление) или прямым. (рис. 25). Нередко его называют атропным (от греч. а — частица отрицания). Ортотропные семязачатки характерны для хлорантовых, савруровых, перцевых, роголистника, платана, мириковых, ореховых, валлиснерии, взморника, рдеста, ксирисовых, эриокауловых, рестиевых и некоторых других родов и семейств.
Гораздо более распространены семязачатки, загнутые вниз (в сторону плаценты) на 180° и приросшие вследствие этого своим боком к удлиненной семяножке. Поэтому микропиле у них обращено к плаценте и расположено рядом с основанием семязачатка. Такие семязачатки называются анатропными (от греч. anatrope — перевернутый, обращенный) или обращенными (рис. 25, 26). Анатропные семязачатки характерны для большинства цветковых, как двудольных, так и однодольных. Известный чешский ботаник Ф. Нетолицкий (1926) пришел к выводу, что анатропный тип является исходным, а ортотропный — производным, что вполне согласуется с современными представлениями.
Наряду с анатропными и ортотропными семязачатками существует еще несколько их «архитектурных» типов, которые, однако, меньше распространены.. Следует прежде всего упомянуть гемитропные (от греч. hemi — половина, или, точнее, «полу»), или полуобращенные, семязачатки, повернутые только на 90° по отношению к плаценте. Они характерны, например, для казуарины, некоторых первоцветных, некоторых бурачниковых, губоцветных, пасленовых и норичниковых, мальпигиевых, некоторых лилейных и ряда других растений. Гемитропные семязачатки представляют собой промежуточный тип между анатропными и ортотропными (рис. 25).
В тех случаях, когда семязачаток вследствие более сильного разрастания одной стороны становится изогнутым, он называется кампилотропным (от греч. kampylos — изогнутый) или полусогнутым. В отличие от анатропного и гемитропного типов здесь изогнуты интегумент и даже мегаспорангий (нуцеллус). Этот тип семязачатка характерен для многих представителей порядка гвоздичных, для каперсовых, некоторых бобовых, для филезиевых, некоторых злаков и ряда других групп. В большинстве случаев кампилотропные семязачатки возникают из анатропных, но не исключено, что иногда они могут возникать и из ортотропных.
В некоторых случаях семязачаток перегибается посередине таким образом, что в продольном разрезе его мегаспорангий имеет подковообразную форму. Это амфитропный (от греч. amphi — оба) семязачаток. Он встречается у лейтнерии, кроссосомы, кнеоровых некоторых акантовых и немногих других растений (рис. 25).
Между всеми этими типами имеются промежуточные формы, и поэтому очень часто бывает трудно определить, какой тип семязачатка имеет то или иное растение. Тем не менее выделение этих типов оказалось очень полезным, так как они имеют определенное систематическое значение.
Как это уже давно указывал датский ботаник Й. Э. Варминг (1913), в систематическом отношении важно также, куда загнуты анатропные или кампилотропные семязачатки — к верхушке (в этом случае они называются эпитропными) или ко дну завязи (они носят название апотропных; рис. 27).
[править] СТРОЕНИЕ СЕМЯЗАЧАТКОВ
Основные черты строения семязачатков мы уже рассматривали в предыдущем томе «Жизни растений». Но напомним еще раз, что семязачаток состоит из двух основных частей — мегаспорангия (нуцеллуса) и окружающего его покрова, называемого интегументом. Интегумент не полностью закрывает мегаспорангий, на верхушке семязачатка его края не срастаются полностью и оставляют отверстие в виде канала для внедрения пыльцевых трубок. Это микропиле, или, иначе, семявход. Нижняя часть семязачатка, т. е. место ее перехода в семяножку (фуникулус), называется халазой. Некоторые авторы, в том числе даже такой известный эмбриолог, как индийский ботаник П. Махешвари (1950), весь семязачаток приравнивают к мегаспорангию. Это большая ошибка. Семязачаток — это не только мегаспорангий, это мегаспорангий плюс интегумент.
 Рис. 26. Анатропный семязачаток смородины золотой (Ribes aureum).
Рис. 26. Анатропный семязачаток смородины золотой (Ribes aureum). Читатель может спросить: почему мегаспорангий семязачатка называют нуцеллусом, если известно, что это мегаспорангий? Дело в том, что долгое время это не было известно, пока в 1851 г. великий немецкий ботаник В. Гофмейстер не установил, что образование, давно известное под названием нуцеллуса, представляет собой не что иное, как мегаспорангий. Но, как часто бывает в науке, даже после замечательных исследований Гофмейстера мегаспорангий семенных растений продолжали называть нуцеллусом, а некоторые ботаники, как, например, американский морфолог А. Имс (1961), подвергали сомнению даже само наличие мегаспорангия у цветковых растений.
Однако большинство ботаников справедливо считают, что наличие у цветковых мегаспорангия, правда очень упрощенного и редуцированного, не вызывает сомнений. Ведь в нуцеллусе имеются два необходимых элемента каждого спорангия — спорогенная ткань (обычно крайне редуцированная) и стенка, которая, однако, гистологически сильно упрощена. Совершенно очевидно, что нуцеллус цветковых морфологически вполне соответствует нуцеллусу голосеменных, т. е. они представляют собой гомологичные образования и являются мегаспорангиями.
Как и у голосеменных, мегаспорангий цветковых окружен интегументом. Однако, в то время как у голосеменных имеется лишь один интегумент, который, согласно гипотезе Маргариты Бенсон (1904), произошел из обеспложенного периферического слоя спорангиев, окружавших центральный фертильный спорангий, у подавляющего большинства цветковых имеются два интегумента. В связи с этим различают битегмальные (от лат. bis — дважды и tegmen — покров) семязачатки, т. е. семязачатки с двумя интегументами, и унитегмальные (от лат. unus — один), т. е. семязачатки с одним интегументом.
В равных линиях эволюции цветковых растений из битегмальных семязачатков произошли унитегмальные. Унитегмальные семязачатки характерны для березовых, мириковых, ореховых, вересковых и близких им семейств, кизиловых, аралиевых, зонтичных, для различных семейств двудольных, характеризующихся спайнолепестным венчиком, и для некоторых однодольных. По-видимому, в большинстве случаев унитегмальные семязачатки произошли из битегмальных в результате слияния обоих покровов, но в ряде случаев они возникли вследствие недоразвития внешнего или чаще внутреннего покрова. Так, в семействе раффлезиевых наблюдается редукция внешнего интегумента, а у рода митрастемон (Mitrastemon) он совершенно подавлен и семязачатки унитегмальные. У родственного семейства гидноровых семязачаток постоянно унитегмальные. В то же время у некоторых представителей семейства розовых, таких, как гравилат, земляника, лапчатка и манжетка, унитегмальный семязачаток возник в результате недоразвития внутреннего покрова. В некоторых семействах, таких, как лютиковые, розовые, бобовые, ивовые, пальмы, орхидные и другие, даже близкие роды отличаются числом интегументов. Это показывает, что унитегмальные семязачатки возникли из битегмальных независимо в самых разных ветвях развития цветковых растений.
У некоторых паразитных двудольных покров семязачатка исчезает полностью и мегаспорангий оказывается, таким образом, голым. Такие атегмальные (от греч. а — частица отрицания) семязачатки характерны для многих, представителей порядка санталовых. У некоторых баланофоровых утрачен не только покров семязачатка, но даже стенка мегаспорангия.
Но откуда появился второй интегумент у битегмальных семязачатков? Этот вопрос оказался одним из наиболее трудных в морфологии растений, и его еще нельзя считать вполне решенным. По всей вероятности, из двух покровов лишь внутренний является настоящим интегументом, соответствующим единственному интегументу голосеменных. Что же касается внешнего интегумента, то скорее всего он имеет другое происхождение. Ряд авторов высказал предположение, что внешний интегумент цветковых растений имеет не спорангиальное, а листовое происхождение. По мнению этих авторов, внешний интегумент произошел из купулы («плюски») семенных папоротников — этих вероятных предков цветковых растений. В видоизмененной форме купула сохранилась и у некоторых примитивных голосеменных. Так, английский палеоботаник Мэри Стоупс (1905) рассматривала наружный мясистый слой семени ныне живущих саговниковых, или саркотесту, как образование, гомологичное купуле типа лагеностомы (Lagenostoma). Но из купулы произошел не только внешний слой покрова семязачатка ряда голосеменных, но, по-видимому, и внешний интегумент цветковых растений. Идея купулярной природы внешнего интегумента цветковых растений была выдвинута независимо французским ботаником А. Госсеном (1946) и английским ботаником Дж. Уолтоном (1953), а сравнительно недавно — американским ботаником Дж. Л. Стеббинсом (1974). Она представляется вполне убедительной.
 Рис. 27. Типы анатропного семязачатка в зависимости от его поворота в гнезде и от направления фуникулуса: в левом ряду - эпитропные, в правом - апотропные.
Рис. 27. Типы анатропного семязачатка в зависимости от его поворота в гнезде и от направления фуникулуса: в левом ряду - эпитропные, в правом - апотропные. [править] МЕГАСПОРАНГИЙ И РАЗВИТИЕ МЕГАСПОРЫ
В семязачатках наблюдаются, два типа строения мегаспорангия и в соответствии с этим различают два типа семязачатков, названные Ф. Ван Тигемом (1898) крассинуцеллятными (от лат. crassus — толстый и nucellus — ядрышко) и тенуинуцеллятными (от лат. tenuis — тонкий). Крассинуцеллятные семязачатки характеризуются относительно мощно развитым мегаспорангием, и поэтому материнская клетка мегаспор (мегаспороцит) отделена от эпидермы мегаспорангия одним или несколькими слоями клеток. У тенуинуцеллятных семязачатков, мегаспорангий которых более редуцирован, материнская клетка мегаспор лежит непосредственно под эпидермой мегаспорангия. Эти два типа не всегда бывают строго разграничены, и между ними известны различные переходы. Первый тип является более примитивным, второй тип возник из первого в результате редукции мегаспорангия. Поэтому крассинуцеллятные семязачатки характерны для относительно более примитивных двудольных и для большинства однодольных, в то время как тенуинуцеллятные семязачатки встречаются у относительно более подвинутых двудольных и лишь у немногих однодольных. Кроме того, крассинуцеллятные семязачатки чаще всего бывают битегмальными, а тенуинуцеллятные, как правило, унитегмальные.
У цветковых растений в мегаспорангии образуется обычно одна материнская клетка мегаспор. Она претерпевает два деления мейоза, в результате чего аналогично появлению тетрады гаплоидных микроспор образуется тетрада гаплоидных мегаспор. Как и при формировании микроспор, появляются каллозовые оболочки, обеспечивающие изоляцию материнской клетки и мегаспор от окружающих клеток.
Мегаспоры тетрады бывают расположены по-разному, но чаще всего образуется линейная тетрада, реже — Т-образная и еще реже — квадратная или тетраэдрическая. У громадного большинства цветковых растений возникают четыре мегаспоры, из которых нижняя (халазальная) и значительно реже верхняя (микропилярная), или промежуточная, увеличиваются в размерах и функционирует как мегаспора, а остальные три отмирают и разрушаются. Очень редко вместо четырех потенциальных мегаспор образуются три, две, а иногда только одна. В последнем случае материнская клетка мегаспоры становится непосредственно мегаспорой.
molbiol.ru
Формирование семязачатка.
Семязачаток
Видоизмененный мегаспорангий , одетый покровами,1 из мегаспор, которого прорастает в женский гаметофит.
Мегаспорангий
Рис 6
1 мегаспорангий
Рис 7
1археспориальная клетка
нуклеус
=мейоз=>
4 мегаспоры
Семязачаток
Рис8
Из 4 мегаспор делится 1 => женский гаметофит
Период формирования семязачатка около 1 часа.
Мужской гаметофит попав на женскую шишку прорастает.
Формирование семени.
После формирования гаметофитов пыльца высыпается и происходит опыление => оплодотворение. В одном из архегониев возникает зигота. Второй архегоний рассасывается. После оплодотворения весь семязачаток превращается в семя. В первую очередь формируется зародыш семени:
Зигота =митоз =>2 клетки =>
Рис9
Одна из них обращена к пыльцевходу, делится чаще в поперечном направлении и из неё формируется, так называемый , «подвесок» -
Рис 10
Вторая клетка проталкивается внутрь эндосперма, => из неё формируется зародыш. На конце образует подвеску, клетка делится поперечно. На противоположном конце деление клетки замедлено, а по бокам оно еще продолжается, и таким образом со стороны подвеска формируется корешок, стебелек, почечку, листочки = семядоли
Рис 11
Этот молодой спорофит – зародыш, способен прервать развитее на стадии молодых органов и уйти в покой – анабиоз, те этот зародыш способен дождаться благоприятных условий для дальнейшего существования.
Параллельно с развитием зародыша клетки нуцеллуса опробковевают => защищают от испарения.
Интегумент одревесневают и превращаются в твердый покров, или кожуру семени.
Пыльцевход закрывается и таким образом семязачаток превращается в семя.
Таким образом семя – зародыш снабженный запасом питательных веществ, и защищенный покровами – твердым снаружи и опропковевшим тонким слоем и состоит из:
зародыш
женский гаметофит – эндосперм (первичный)
тонкая кожура
твердая кожура
Преимущества голосемянных
спорофит доминирует и имеет довольно совершенную структуру.
споры разных размеров формируются на разных шишках и порастают в разедльнопполые гаметофиты.
гаметофиты ♂ и ♀ редуцированы и паразитируют на спорофите (противоположность мхам). Мужские гаметы неподвижные – спермии
половой процесс не зависит от водной среды
встреча гамет обеспечивается особенностями строениями мужского гаметофита, формируется из него пыльцевой трубки и структурной семязачатки
наличие семян обеспечивает выживаемость дочернего спорофита, так как зародыш способен переждать неблагоприятные условия, имея запас питательных веществ и защищен покровами от высыхания и механических повреждений.
наличие семян обеспечивает лучшее расселение растений.
Отдел покрытосеменные
При цветении растений из цветочных почек формируется соответствующие части цветка: наружные бугорки конуса нарастания превращаются в листочки околоцветника (простого или двойного, свободного или сросшегося, актиноморфного или зигоморфного).
Следующий ряд первичных бугорков конуса нарастания цветочной почки дифференцируется в тычинки (андроцей).
Тычинка состоит из тычиночной нити, связника, и дух пыльников. Самые сложные процессы происходят в гнездах пыльника: здесь из микроспор формируются мужские гаметофиты – пыльцевые зерна, каждое пыльцевое зерно это редуцированный до двух клеток мужской гаметофит. Пыльцевые зерна защищены от внешней среды, стенками пыльника, имеющими специфическое строение.
Самые верхние первичные бугорки конуса нарастания почки дифференцируются в плодолистики. Плодолистик - это структурная единица гинецея или пестика. Число плодолистиков в гинецеи (пестика) могут быть различными (от одного до множества), а также они могут быть свободными или сросшимися. Пестик имеет завязь, столбик (иногда отсутствует) и рыльце. В завязи пестика происходят процессы формирования семязачатка, оплодотворения и развитие семени и плода. При формировании семязачатка , как и в пыльниках происходит процесс мегаспорогенеза и развития женского гаметофита –зародышевого мешка. Редуцированный до восьми клеток женский гаметофит достаточно надежно защищен от внешней среды, стенкой мегаспорангия (нуцеллусом) интегументами и многослойными стенками завязи.
Строение и места формирования покрытосеменных гаметофитов способствует гарантированному ( в определенной мере) оплодотворению и возникновению дочернего спорофита – зародыша с большой пластичностью и повышенной выживаемостью.
studfiles.net
Цветок растения. Строение, органы размножения (пестики и тычинки)
Цветок — сложная система органов, обеспечивающая семенное размножение у цветковых растений. Появление цветка в процессе эволюции — ароморфоз, обусловивший широкое распространение на Земле покрытосеменных, или цветковых, растений.
Функции цветка:
- образование тычинок с пыльцевыми зернами плодолистиков (пестиков) с семяпочками;
- опыление;
- сложные процессы оплодотворения;
- формирование семени и плода.
Цветок — это укороченный видоизмененный и ограниченный в росте побег, несущий околоцветник, тычинки, плодолистики (пестики). Строение цветков у всех цветковых растений сходно, а форма — разнообразна. В этом проявляется идиоадаптация — приспособление к различным способам опыления.
 Цветок растения
Цветок растенияНаружное строение цветка
Цветком заканчивается главный стебель или боковые. Безлистная часть стебля под цветком называется цветоножкой. У сидячих цветков цветоножка отсутствует либо сильно укорочена. Цветоножка переходит в укороченную ось цветка, его стеблевую часть — цветоложе. Форма цветоложа может быть удлиненной, выпуклой, плоской, вогнутой. На цветоложе располагаются все части цветка: чашелистики и лепестки, тычинки и пестик (пестики).
Чашелистики и лепестки вместе составляют околоцветник. Чашелистики обычно защищают от повреждений цветок, особенно бутон, но могут выполнять и другие функции. В зеленых чашелистиках, содержащих хлоропласты, происходит фотосинтез. У некоторых растений (тюльпан, ветреница) они становятся лепестковидными и выполняют функции лепестков; могут служить для защиты развивающихся плодов, для их распространения.
Произошли чашелистики из верховых вегетативных листьев. Доказательством этого является их морфологическое сходство с листьями, четко, выраженное у некоторых растений (пион), и спиральное расположение. Совокупность чашелистиков образует чашечку, которая бывает либо раздельнолистной, либо сростнолистной.
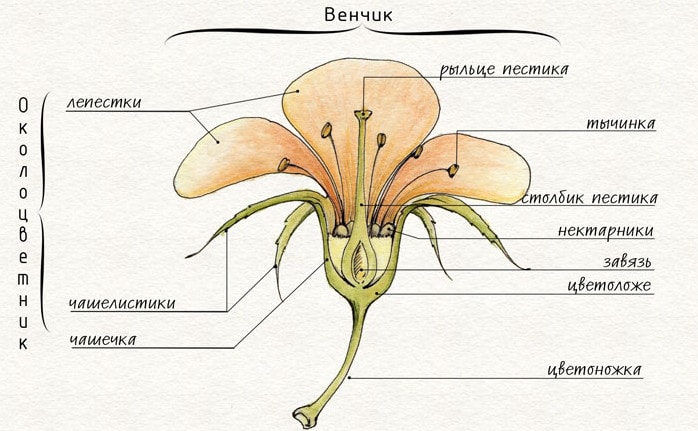 Строение цветка растения
Строение цветка растенияЛепестки выполняют функции привлечения опылителей и содействия успешному опылению. Происхождение лепестков двоякое: у одних растений — это видоизмененные тычинки. Такие лепестки у кувшинки, а также у представителей семейств лютиковых, гвоздичных, маковых и др. Другая группа растений имеет лепестки, как и чашелистики, листового происхождения (пион, магнолиевые).
Совокупность лепестков цветка называется венчиком. Размеры, строение, окраска венчика разнообразны, что связано с биологией опыления. У ветроопыляемых растений венчик либо недоразвит, либо отсутствует. Лепестки могут срастаться краями, образуя скоростнолепестной венчик (вьюнок, петуния). В процессе эволюции такой венчик произошел от свободнолепестного.
При наличии в цветке чашечки и венчика околоцветник называется двойным. Если лепестков нет либо разница между ними неясно выражена, околоцветник называется простым. Простой околоцветник может быть либо венчиковидным с яркой окраской — у тюльпанов, лилий, ландышей, либо чашечковидным, зеленого цвета — у конопли, лебеды, крапивы. Цветки без околоцветника называются голыми — у осоки, ивы.
Тычинки
 Тычинки
ТычинкиВнутри околоцветника ближе к лепесткам располагаются тычинки. Число их различно: от одной до десятка и более. В процессе эволюции тычинка дифференцировалась на тычиночную нить и пыльник. Пыльник состоит из двух половинок, соединенных продолжением тычиночной нити. В каждой половинке пыльника по два спорангия, их называют гнездами пыльника, или пыльцевыми мешками.
 Строение тычинки цветка
Строение тычинки цветкаГнезда заполнены тканью из первичных спорогенных клеток. В результате ряда последовательных митозов из первичных спорогенных клеток образуется множество материнских клеток — микроспор. Затем материнские клетки делятся мейотически, образуя тетрады гаплоидных микроспор. Каждая такая микроспора превращается в пыльцевое зерно. Для этого она увеличивается в размерах и покрывается двойной оболочкой: наружной (экзиной) и внутренней (интиной). Наружная оболочка благодаря основному ее компоненту — спорополленину — характеризуется высокой стойкостью: не растворяется в кислотах и щелочах, выдерживает температуру до 300°С, сохраняется миллионы лет в геологических отложениях.
Внутри пыльцевого зерна формируется мужской гаметофит: гаплоидная микроспора делится митотически, образуя более крупную клетку-трубку (вегетативную) и в ней —маленькую генеративную клетку. Генеративная клетка делится еще раз митотически на две мужские гаметы — спермии.
Пестики
 Пестик
ПестикВнутреннюю часть цветка занимают пестики. Количество их бывает от одного до десятка и более. Каждый пестик образован одним или многими сросшимися плодолистиками.
В нижней части пестика — завязи — находятся семязачатки (семяпочки). Из верхней его части в процессе эволюции образуется столбик, приподнимающий рыльце над пестиком. При отсутствии столбика рыльце называется сидячим. Завязь может быть верхняя, если она располагается на плоском или выпуклом цветоложе, а все остальные части цветка прикрепляются под пестиком. У цветков с нижней завязью вогнутое цветоложе срастается с ее стенкой, околоцветник и тычинки прикрепляются над пестиком.
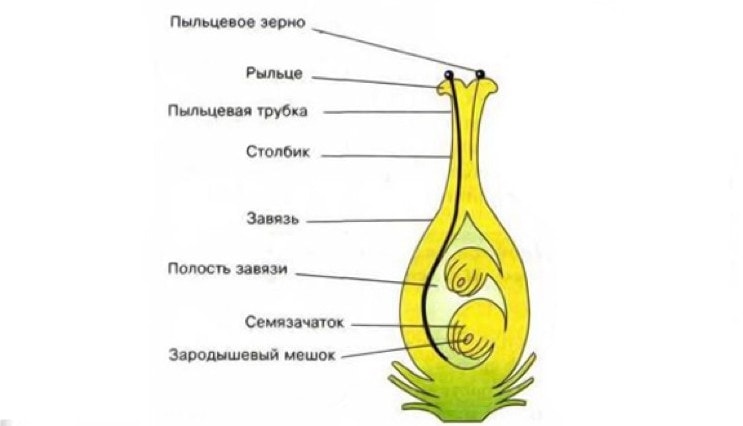 Строение пестика
Строение пестикаУ завязи пестика находится полость — гнездо. Различают одно-, и многогнездную завязи. Многогнездная завязь образуется в результате срастания нескольких плодолистиков. Количество гнезд равно числу сросшихся плодолистиков. В каждом гнезде на стенках завязи формируются семязачатки (семяпочки), либо сидячие, либо на семяножках. Их бывает от одного (слива, вишня) до нескольких тысяч (мак, орхидные).
Строение семязачатка (семяпочки)
При анатомическом исследовании семяпочки различаются следующие составные части:
- Семяножка;
- нуцеллус;
- покровы;
- микропиле;
- зародышевый мешок.
По семяножке в зародышевый мешок поступают питательные вещества и ею семяпочка крепится в завязи. Нуцеллус семяпочки — это паренхиматическая питающая и защитная для мегаспор ткань. Снаружи нуцеллус одет одним или двумя покровами (интегументами). Они покрывают нуцеллус не сплошь. Чаще сверху семяпочки они не соединяются и образуют небольшое отверстие, которое называется микропиле, или пыльцевход.
Самую внутреннюю часть семяпочки занимает зародышевый мешок, который у покрытосеменных представляет собой женский гаметофит.
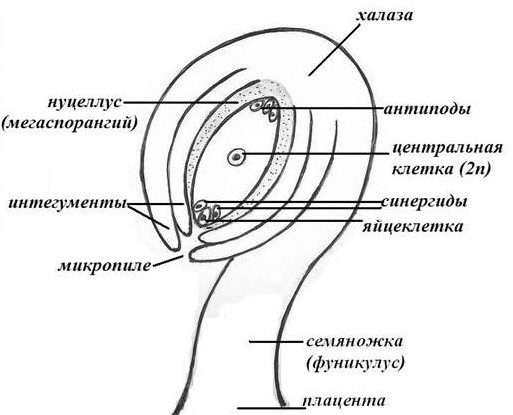 Строение семязачатка
Строение семязачаткаСемязачаток (семяпочка) состоит из макроспорангия и окружающего его покрова. В макроспорангии закладывается одна материнская клетка, из которой путем мейоза образуется тетрада гаплоидных макроспор. Три из них отмирают и разрушаются, а четвертая (дающая начало женскому гаметофиту) макроспора сильно вытягивается в длину, одновременно ее гаплоидное ядро делится митотически. Дочерние ядра расходятся к разным полюсам удлиненной клетки.
Далее, каждое из образовавшихся ядер делится митотически еще дважды и образует по четыре гаплоидных ядра у разных полюсов клетки. Это уже зародышевый мешок, имеющий восемь гаплоидных ядер. Затем от каждой из двух четверок ядер по одному направляются к центру зародышевого мешка, где они сливаются, образуя вторичное диплоидное ядро.
После этого в цитоплазме зародышевого мешка возникают клеточные перегородки между ядрами и он становится семиклеточным.
У одного из полюсов зародышевого мешка размещается яйцевой аппарат, состоящий из более крупной яйцеклетки и двух вспомогательных клеток. У противоположного полюса — три клетки-антипода. Все шесть клеток гаплоидны. В центре находится диплоидная клетка со вторичным ядром.
У большинства растений цветки имеют тычинки и пестики и называются обоеполыми. Бывают цветки и однополыми: тычиночные (мужские) либо пестичные (женские). Мужские и женские цветки могут располагаться на одной особи, такое растение называют однодомным (огурец, кукуруза, дуб, береза), а если на разных особях — двудомным (конопля, ива, тополь). Однополые цветки и двудомные растения — это одно из приспособлений к перекрестному опылению.
Диаграммы и форумулы растений
Для краткого описания цветка пользуются диаграммами и формулами. Диаграмма — это схематическая проекция элементов цветка на плоскость, перпендикулярную к его оси. Все части цветка, прицветник и материнский побег обозначают определенными значками: чашелистики — фигурной скобкой, лепестки — круглой скобкой, тычинки — поперечным разрезом через пыльник, пестик — поперечным разрезом через завязь.
При составлении формулы цветка околоцветник обозначают буквой О, чашелистики — Ч, лепестки — Л, тычинки — Т, пестик — П. Число частей цветка обозначают цифрой, написанной у основания буквы. Если тычинок и пестиков больше 12, ставят значок — ∞. При срастании частей цветка соответствующие цифры берут в скобки. Верхнюю завязь обозначают горизонтальной чертой под числом, нижнюю — над числом пестиков.
animals-world.ru
Биология для студентов - 13. Цветок, его происхождение, части цветка (околоцветник, андроцей гинецей), семязачаток, пыльца

Околоцветник - это стерильная (бесплодная, не имеющая способности к оплодотворению) часть цветка, выполняющая защитную функцию, а также функцию привлечения опылителей. Околоцветник, дифференцированный на чашечку и венчик, называют двойным, а не дифференцированный на чашечку и венчик - простым. Простой чашечковидный околоцветник имеет обычно зеленый цвет, простой венчиковидный окрашен ярко. Цветки, не имеющие околоцветника, называют голыми.
Чашечка. Обычно состоит из небольших зеленых цветолистиков - чашелистиков. Чаще чашелистики образуют один круг. Иногда же под чашечкой расположен круг листовидных образований, называемый подчашием Венчик обычно состоит из цветолистиков более крупных, чем чашелистики, и ярко окрашенных - лепестков. Лепестки могут срастаться между собой, хотя бы частично, или оставаться свободными. Различают два типа венчиков: свободнолепестные (раздельнолепестные) и сростнолепестные (спайнолепестные).
Венчик, как и чашечка, может быть: актиноморфным (правильным, полисимметричным), когда через него можно провести две или больше плоскостей симметрии, зигоморфным (неправильным, моносимметричным), когда можно провести только одну плоскость симметрии, асимметричным, когда нельзя провести ни одной плоскости симметрии (канна, орхидеи).
Андроцей - это совокупность тычинок одного цветка. Число тычинок в андроцее колеблется в больших пределах - от одной до нескольких сотен. Тычинки могут быть или свободными (свободный андроцей), или в различной степени сросшимися (сросшийся андроцей). Если срастаются все тычинки, то их называют однобратственными, если одна тычинка остается свободной - двубратственными, и если тычинки срастаются в несколько групп - многобратственными. По высоте относительно друг друга тычинки могут быть одинаковыми или разными двусильными, когда две тычинки выше других, трехсильными и т. д.

В типичном случае тычинка состоит из тычиночной нити и пыльника. Тычиночные нити у большинства растений простые, неветвящиеся. Но иногда они имеют боковые выросты разнообразной формы или ветвятся. Если тычиночная нить отсутствует, то тычинку называют сидячей.Пыльник обычно состоит из двух половин, называемых теками, соединенных между собой связником.

Иногда связник рассматривают как самостоятельную третью часть тычинки, однако под микроскопом ткань его неотличима от ткани пыльника. В каждой из тек находится по два гнезда, или пыльцевых мешка. Снаружи пыльника имеется однослойная эпидерма. Это все, что осталось от микроспорофилла. Глубже расположенные ткани принадлежат микроспорангию. Субэпидермальный слой называют эндотецием. Клетки его рано теряют живое содержимое, а стенки на обращенной внутрь стороне клетки несут фиброзные пояски, способствующие вскрытию пыльника. За эндотецием в молодом пыльнике расположены средний слой и выстилающий слой, или тапетум. Клетки тапетума крупные, с густой цитоплазмой и с несколькими ядрами. В центре каждого гнезда имеется спорогенная ткань, из клеток которой образуются микроспоры, а затем пыльца. По мере созревания пыльника клетки тапетума и среднего сдоя исчезают, так как их содержимое идет на питание растущих микроспор и пыльцы. Строение пыльцы довольно однообразно, но структура ее покрова - спородермы - отличается разнообразием.
У некоторых видов растений часть тычинок не имеет пыльников и представлена лишь тычиночными нитями. Такие бесплодные тычинки называют стаминодиями (лен). Иногда стаминодии имеют вид окрашенных лепестковидных пластинок (у канны).
Гинецей - это совокупность плодолистиков (мегаспорофиллов) одного цветка, образующих один или несколько пестиков. Гинецей, состоящий из одного плодолистика, образующего один пестик, называют одночленным, а из нескольких плодолистиков - многочленным.

Многочленный гинецей может быть апокарпным, если плодолистики не срастаются между собой и образуют несколько пестиков, и ценокарпным, если плодолистики срастаются в один пестик. Ценокарпный гинецей состоит из стольких плодолистиков, сколько отдельных столбиков несет завязь или лопастей - рыльце и сколько гнезд или швов имеет завязь. При этом надо учитывать всю совокупность перечисленных признаков, так как один из них не всегда точно указывает на число плодолистиков. Лизикарпный гинецей одногнездный гинецей, образованный несколькими сросшимися плодолистиками. Семязачатки находятся на колонке, расположенной в центре завязи, например у представителей сем. Гвоздичные.
Пестик - это закрытое вместилище семязачатков (семяпочек), образовавшееся вследствие срастания краев одного или нескольких плодолистиков. Пестик состоит из завязи, вздутой части, где находятся семязачатки, столбика и рыльца, воспринимающего пыльцу. Иногда столбик отсутствует, тогда рыльце сидячее.
Завязь в зависимости от положения по отношению к другим частям цветка бывает верхняя и нижняя. Верхняя завязь располагается свободно на плоском, выпуклом или вогнутом цветоложе и образуется только плодолистиками. Ее можно легко отделить от цветоложа препаровальной иглой. В образовании нижней завязи, кроме плодолистиков, принимают участие и другие части цветка - чаще основания чашелистиков, лепестков и тычинок, реже цветоложе, с которыми она срастается. Поэтому отделить иглой такую завязь нельзя. Выделяют еще полунижнюю завязь, когда по крайней мере нижняя половина ее срастается с другими частями цветка. В зависимости от числа не сообщающихся между собой гнезд завязи бывают одногнездными (если гнезда так или иначе сообщаются между собой), двугнездными и многогнездными.
Внутри завязи образуются семязачатки.

Место прикрепления семязачатка к стенке завязи называют плацентой (семяносцем). Семязачаток прикрепляется к плаценте фуникулусом (семяножкой). Семязачаток снаружи имеет два покрова - интегумента, которые на верхушке не смыкаются, оставляя отверстие - микропиле (пыльцевход). Основание семязачатка, называемое халазой, находится на противоположной микропиле стороне. Под интегументами сформированного семязачатка расположена многоклеточная ткань - нуцеллус (мегаспорангий). Одна из его клеток становится археспориальной. Из нее образуются четыре мегаспоры. Одна мегаспора дает начало восьмиядерному зародышевому мешку.
Жизненный цикл покрытосеменных.

Образование женского гаметофита - зародышевого мешка - происходит следующим образом. После редукционного деления (мейоза) одной из клеток нуцеллуса (археспориальной клетки) образуются четыре гаплоидные мегаспоры. Этот процесс называют макроспорогенезом. Три мегаспоры вскоре отмирают, а из одной мегаспоры после трех последовательных делений митозом образуется восемь гаплоидных ядер; на их основе формируются клетки: рядом с микропиле - яйцеклетка (женская гамета) и две клетки - синергиды, на противоположном конце семязачатка (в области халазы) - три клетки - антиподы. В центре зародышевого мешка два ядра сливаются - образуется диплоидное центральное ядро (ядро центральной клетки). Так формируется зародышевый мешок — женский гаметофит.
В пыльниках тычинок цветка образуются пыльцевые зерна (пыльца) — мужские гаметофиты. Это происходит следующим образом. В результате редукционного деления диплоидных клеток в гнездах пыльников образуются многочисленные гаплоидные микроспоры. Этот процесс называют микроспорогенезом. Ядро каждой микроспоры делится митозом с образованием генеративной клетки и клетки трубки (сифоногенной клетки). Это и есть пыльцевое зерно. Из генеративной клетки митозом образуются мужские гаметы (гаплоидные половые клетки) - спермии. Спермии не имеют жгутиков: они доставляются к зародышевому мешку и яйцеклетке пыльцевой трубкой, которую образует сифоногенная клетка.
Семяпочка — зачаток семени, скрытый в завязи цветка у высших растений. Семяпочка имеет следующее строение: она состоит из центральной части, так называемого ядра, одетого двумя покровами, наружным и внутренним. Оба покрова оставляют наверху небольшое отверстие — семявход, или микропиле. Внутри ядра, заполненной тканью, находится одна крупная клетка, которая называется зародышевым мешком. В этом зародышевом мешке образуется яйцевая клеточка, которая по оплодотворении дает начало зародышу. Семяпочки развиваются на плодолистиках, т. е. метаморфозированных листьях. У покрытосеменных плодолистики срастаются в замкнутое помещение, т. е. завязь, внутри которой помещаются С. С прикреплены к стенкам завязи по швам плодолистиков (семяносцы стенкоположные).
Иногда семяпочки сидят на дне завязи своим основанием (халязой), обращенные кверху семявходом. Такие семяпочки называются прямыми, или ортотропными. Гораздо чаще они подвешены на особой нити, так называемом семенном канатике (raphe), которая начинается от основания семяпочки, но прирастает к ней сбоку и отделяется лишь около микропиле. Это — обращенные семяпочки, или анатропные.
В семенном канатике проходит сосудистый пучок, по которому совершается приток питательных веществ. По оплодотворении яйцевой клеточки из нее начинает развиваться зародыш, а в зародышевом мешке, кроме того, часто отлагаются питательные вещества, т. е. образуется эндосперм. Таким образом, зародышевый мешок (со своим содержимым) сильно разрастается и вытесняет остальную ткань в ядре семяпочки.
Опыление. После созревания пыльцы пыльники лопаются и пыльца попадает на рыльце пестика. Этот процесс называют опылением. Различают два способа опыления: самоопыление и перекрестное опыление. Самоопыление может осуществляться только в обоеполых цветках. Оно наиболее хорошо выражено у нераскрывающихся (клейстогамных) цветков. Перекрестное опыление в ходе эволюции оказалось более прогрессивным. При перекрестном опылении потомство может совмещать наследственные свойства обоих родителей, а это открывает более широкие возможности в приспособлении к разным условиям существования. Подавляющее большинство растений имеет перекрестное опыление.
По способу переноса пыльцы различают: анемофилию (ветром), гидрофилию (водой), энтомофилию (насекомыми), орнитофилию (птицами), мирмекофилию (муравьями) и др. Хорошо известны приспособления цветков к определенным насекомым-опылителям. Так, виды клевера имеют венчик, сросшийся в длинную трубку, и только шмели и некоторые длиннохоботковые пчелы способны доставать из нее нектар и осуществлять перекрестное опыление. Различают две формы перекрестного опыления: гейтеногамию (соседнее опыление), когда оно осуществляется в пределах одного растения, но пыльца с одного цветка попадает на рыльце другого; и ксеногамию (собственно перекрестное), когда пыльца с цветка одной особи переносится на рыльце цветка другой особи.
Для предотвращения самоопыления у растений выработались разнообразные приспособления. Наиболее широко распространены дихогамия и гетеростилия. Дихогамия - это разновременное созревание тычинок и пестиков в обоеполом цветке. При гетеростилии у одних экземпляров в обоеполых цветках пестики с длинными столбиками, а тычинки с короткими тычиночными нитями, у других - наоборот.
vseobiology.ru
Строение семязачатков
Строение семязачатковОсновные черты строения семязачатков мы уже рассматривали в предыдущем томе «Жизни растений». Но напомним еще раз, что семязачаток состоит из двух основных частой — мегаспорангия (нуцеллуса) и окружающего его покрова, называемого интегументом. Интегумент не полностью закрывает могаспорангий, на верхушке семязачатка его края не срастаются полностью и оставляют отверстие в виде канала для внедрения пыльцевых трубок. Это микропиле, или, иначе, семявход. Нижняя часть семязачатка, т. е. место ее перехода в семяножку (фуникулус), называется халазой. Некоторые авторы, в том числе даже такой известный эмбриолог, как индийский ботаник П. Махешвари (1950), весь семязачаток приравнивают к мегаспорангию. Это большая ошибка. Семязачаток — это не только мегаспорангий, это мегаспорангий плюс интегумент.
Читатель может спросить: почему мегаспорангий семязачатка называют нуцеллусом, если известно, что это мегаспорангий? Дело в том, что долгое время это не было известно, пока в 1851 г. великий немецкий ботаник В. Гофмейстер не установил, что образование, давно известное под названием нуцеллуса, представляет собой не что иное, как мегаспорангий. Но, как часто бывает в науке, даже после замечательных исследований Гофмейстера мегаспорангий семенных растений продолжали называть нуцеллусом, а некоторые ботаники, как, например, американский морфолог А. Имс (1961), подвергали сомнению даже само наличие мегаспорангия у цветковых растений.
Однако большинство ботаников справедливо считают, что наличие у цветковых мегаспорангия, правда очень упрощенного и редуцированного, не вызывает сомнений. Ведь в нуцеллусе имеются два необходимых элемента каждого спорангия — спорогенная ткань (обычно крайне редуцированная) и стенка, которая, однако, гистологически сильно упрощена. Совершенно очевидно, что нуцеллус цветковых морфологически вполне соответствует нуцеллусу голосеменных, т. е. они представляют собой гомологичные образования и являются мегаспорангиями.
Как и у голосеменных, мегаспорангий цветковых окружен интегументом. Однако, в то время как у голосеменных имеется лишь один интегумент, который, согласно гипотезе Маргариты Бенсон (1904), произошел из обеспложенного периферического слоя спорангиев, окружавших центральный фертильный спорангий, у подавляющего большинства цветковых имеются два иитегумента. В связи с этим различают битегмальные (от лат. bis — дважды и legmen — покров) семязачатки, т. е. семязачатки с двумя интегументами, и унитегмальные ( от лат. unus — один), т. е. семязачатки с одним интегументом.
В разных линиях эволюции цветковых растений из битегмальных семязачатков произошли унитегмальные. Унитегмальные семязачатки характерны для березовых, мириковых, ореховых, вересковых и близких им семейств, кизиловых, аралиевых, зонтичных, для различных семейств двудольных, характеризующихся спайнолепестным венчиком, и для некоторых однодольных. По-видимому, в большинстве случаев унитегмальные семязачатки произошли из битегмальных в результате слияния обоих покровов, но в ряде случаев они возникли вследствие недоразвития внешнего или чаще внутреннего покрова. Так, в семействе раффлезиевых наблюдается редукция внешнего интегумента, а у рода митрастемон (Mitvastemon) oн совершенно подавлен и семязачатки унитегмальные. У родственного семейства гидпоровых семязачаток постоянно унитегмальный. В то же время у некоторых представителей семейства розовых, таких, как гравилат, земляника, лапчатка и манжетка, унитегмальный семязачаток возник в результате недоразвития внутреннего покрова. В некоторых семействах, таких, как лютиковые, розовые, бобовые, ивовые, пальмы, орхидные и другие, даже близкие роды отличаются числом интегументов. Это показывает, что унитегмальные семязачатки возникли из битегмальных независимо в самых разных ветвях развития цветковых растений.
У некоторых паразитных двудольных покров семязачатка исчезает полностью и мегаспорангий оказывается, таким образом, голым. Такие атегмальные (от греч. а — частица отрицания) семязачатки характерны для многих, представителей порядка санталовых. У некоторых баланофоровых утрачен не только покров семязачатка, но даже стенка мегаспорангия.
Но откуда появился второй интегумент у битегмальных семязачатков? Этот вопрос оказался одним из наиболее трудных в морфологии растений, и его еще нельзя считать вполне решенным. По всей вероятности, из двух покровов лишь внутренний является настоящим интегументом, соответствующим единственному интегументу голосеменных. Что же касается внешнего интегумента, то скорее всего он имеет другое происхождение. Ряд авторов высказал предположение, что внешний интегумент цветковых растений имеет не спорангиальное, а листовое происхождение. По мнению этих авторов, внешний интегумент произошел из купулы («плюски») семенных папоротников — этих вероятных предков цветковых растений. В видоизмененной форме купула сохранилась и у некоторых примитивных голосеменных. Так, английский палеоботаник Мэри Стоунс (1905) рассматривала наружый мясистый слой семени ныне живущих саговниковых, или саркотесту, как образование, гомологичное купуле типа лагеностомы (Lagenostoma). Но из купулы произошел не только внешний слой покрова семязачатка ряда голосеменных, но, по-видимому, и внешний интегумент цветковых растений. Идея купулярной природы внешнего интегумента цветковых растений была выдвинута независимо французским ботаником Л. Госсеном (1946) и английским ботаником Дж. Уолтоном (1953), а сравнительно недавно — американским ботаником Дж. Л. Стеббинсом (1974). Она представляется вполне убедительной.
Источник: Биологическая энциклопедия
предыдущие статьи
последующие статьи
mirznanii.com
Семязачаток - это... Что такое Семязачаток?
Семязачатки саговниковых отличаются по величине (длиной от 5—6 см у некоторых видов саговника до 5—7 мм у замии карликовой) и по форме. Но при этом они довольно однотипны в основных чертах развития и внутреннего строения. Открыто сидящие на «черешке» листовидного мегаснорофилла у саговников, висячие на нижней стороне щитовидной чешуи у замий, прикрытые более или менее плоской чешуей мегастробила у других родов, семязачатки всегда состоят из нуцеллуса (ядра семязачатка) и одевающего его покрова — интегумента. Толстый покров этот прочно срастается с нуцеллусом, отходя от него лишь на верхушке семязачатка. Здесь, в центре образованного интегументом конусообразного выступа, имеется отверстие — микропиле, а под ним полость — пыльцевая камера.
Главной частью семязачатка является нуцеллус. Это и есть собственно мегаспорангий, который, в отличие от открытых микроснорангиев саговниковых, как и у других голосеменных, заключен в защитный покров.
Образовавшаяся в нуцеллусе в результате мейоза единственная функционирующая мегаспора быстро растет за счет трех остальных, вскоре отмирающих спор тетрады и окружающих клеток нуцеллуса и вырабатывает хорошо выраженную двуслойную оболочку, как это свойственно спорам бессеменных высших растений, переносимым токами воздуха. Но мегаспора саговниковых никогда не покидает мегаспорангия, и назвапная особенность ее строения сохранилась как пережиток прошлого, унаследована от далеких предков, расселявшихся с помощью спор. У саговников наружная оболочка мегаспоры в дополнение пропитывается кутином, что, очевидно, придает ей еще более архаичный характер.
Итак, спорогенез завершен. К этому времени интегумент разросшегося семязачатка уже дифференцирован на три слоя — мясистые наружный и внутренний и твердый средний, состоящий из омертвевших клеток. Оба мясистых слоя пронизапы целой системой проводящих пучков, обеспечивающих разрастающийся семязачаток необходимыми питательными веществами.
Образовавшаяся в нуцеллусе мегаспора тут же и прорастает, образуя женский гаметофит. Процесс этот образно описан еще Ч. Чемберленом. Вслед за первым делением ядра споры происходят многие повторные ядерные деления, при этом не образуется клеточных перегородок (рис. 168, 5). Многочисленные свободные ядра (их число, например у диоона, может доходить до тысячи) оказываются в тонком постенном слое цитоплазмы. В дальнейшем начинают обособляться клетки гаметофита, сперва по периферии, затем все ближе и ближе к центру мегаспоры, пока вся она не заполняется многоклеточной тканью (рис. 168, 6). По общему объему и по числу составляющих кле ток женский гаметофит саговниковых не уступает даже наиболее крупным свободно живущим гаметофитам (заросткам) папоротников.
Женский гаметофит развивается как запасающая ткань (первичный эндосперм). Разрастаясь, он вытесняет нуцеллус (рис. 168, 7), затем потребляет вещества из внутреннего мясистого слоя интегумента, пока этот слой не превращается в тонкую пленку ковнутри от твердой «скорлупы». Клетки эндосперма постепенно заполняются запасными материалами, среди которых преобладает крахмал (до 65— 70% в пересчете на сухую массу у видов макрозамии). Накапливаются в эндосперме также жирные масла (у саговника поникающего их содержание может доходить до 23%), а также белки.
Наконец, в клетках женского гаметофита обнаруживаются лейкопласты. Хотя женский гаметофит саговниковых и утерял в ходе эволюции возможность самостоятельного существования вне семязачатка, он удивительным образом сохранил способность к развитию хлорофилла (позеленению) на свету при извлечении из мегаспорангия. Наблюдали превращение лейкопластов в хлоропласта и на семязачатках, в которых не произошло оплодотворение. В этом случае гаметофит прорастал через микропиле и его выступающий конец зеленел на свету.
В этом отношении интересны опыты по выращиванию эксплантатов (кусочков ткани) от женского гаметофита саговниковых в стерильной культуре. В зависимости от соотношения в питательной среде различных стимуляторов роста растущая клеточная масса гаметофита образует то корни, то стеблевые почки, то, наконец, зародышеподобные образования (эмбриоиды), т. е. проявляет особенности морфогенеза, свойственные спорофиту.
В верхней части нормально сформировавшегося женского гаметофита под микропиле развиваются женские половые органы — архегонии (рис. 168, 7, 16). Материнских клеток архегониев закладывается много, и это еще одна архаическая черта саговниковых. Правда, вполне развитых архегониев у них оказывается обычно не более десяти. Совершенно исключителен в этом отношении микроцикас, у которого образуется несколько десятков архегониев.
Яйцеклетка в архегонии достигает огромных размеров (у микроцикаса она длиной до 6 мм). Необычайно велико и ядро яйцеклетки; будучи иногда диаметром до 500 мкм, оно видимо простым глазом, как точка. Напротив, шейка архегония маленькая и состоит обычно из двух мелких клеток, которые к моменту оплодотворения ослизняются, открывая доступ к яйцеклетке. К этому времени между микропиле и верхней частью гаметофита оказывается довольно большая полость (пыльцевая и архегониальная камеры при прорыве оболочки мегаспоры сливаются), куда и попадают переносимые ветром пыльцевые зерна.
Жизнь растений: в 6-ти томах. — М.: Просвещение. Под редакцией А. Л. Тахтаджяна, главный редактор чл.-кор. АН СССР, проф. А.А. Федоров. 1974.
dic.academic.ru
Происхождение семязачатка - это... Что такое Происхождение семязачатка?
Происхождение семязачатка Происхождение семязачаткаТеперь уместно перейти к вопросу о происхождении семязачатка. Но сначала познакомимся несколько подробнее с его строением. Если проследить развитие семязачатка с начала его заложения, то легко заметить, что мегаспорангий (нуцеллус) возникает в виде бугорка, состоящего из мелких паренхимных клеток. Этот бугорок обрастает затем интегументом, появляющимся в виде кольцевого валика в основании нуцеллуса. Но интегумент не полностью обрастает нуцеллус — его края оставляют на его верхушке открытый узкий канал — микропиле (от греч. micros — малый и pyle — ворота), через который проникают внутрь микроспоры. Семязачаток прикреплен к плаценте посредством более или менее заметной семяножки, или фуникулуса (от лат. funiculus — тонкий канат, веревка). Место в семязачатке, где интегумент переходит в семяножку, т. е. нижний конец семязачатка, называют халазой (от греч. chalaza — бугорок).
Происхождение интегумента долгое время не поддавалось удовлетворительному объяснению. Первоначально принималось происхождение иптегумента из покрывальца (индузия) папоротников. Но «индузиальная» гипотеза происхождения интегумента представляет в настоящее время лишь исторический интерес. Гораздо более правдоподобной является так называемая «синангиальная» гипотеза. Согласно этой гипотезе, выдвинутой впервые английским палеоботаником Маргаритой Бенсон (1908), интегумент представляет собой кольцо подвергшихся стерилизации, сросшихся и слившихся между собой спорангиев, окружающих центральный функционирующий мегаспорангий, а микропиле соответствует первоначальному промежутку между верхушками спорангиев. Другими словами, семязачаток является фактически синангием, у которого все спорангии, кроме одного, стерилизовались и образовали покров (интегумент) единственного, фертильного спорангия.
Хорошим подтверждением «синангиальной» гипотезы являются примитивные семязачатки семенных папоротников, сохранившие часто весьма ясные следы своего синангиального происхождения. Семязачатки ряда семенных папоротников обладали сегментированными интегументами с проводящим пучком в каждом сегменте (камере). Пыльцевая камера физостомы (Physostoma), например, была окружена 12 «щупальцами», представляющими собой свободные концы стерильных спорангиев, или камер интегумента. Каждая продольная камера примитивных многокамерных палеозойских интегументов соответствует одному стерилизованному спорангию. Интересно, однако, что сегментированные интегументы известны не только у примитивных палеозойских голосеменных, но и у мезозойских беннеттитовых и даже у современных саговниковых. Так, у некоторых родов саговниковых (макрозамии, цератозамии, энцефаляртоса и др.) вокруг микропиле видно 7—16 ясных лопастей склеротесты (внутреннего твердого слоя), соответствующих верхушкам сегментов интегумента. Все эти факты говорят о том, что семязачаток есть, в сущности, видоизмененный и сильно специализированный мегасинангий. Другими словами, семязачаток представляет собой мегасинангий, в котором произошло разделение функций между центральным фертильным мегаспорангием и окружающим его кольцом мегаспорангиев, которые стерилизовались и превратились в защитный покров. Однако как центральный, так и окружающие мегаспорангии подверглись стольким структурным и функциональным изменениям, что семязачаток с полным основанием рассматривается как очень крупное эволюционное новообразование.
Жизнь растений: в 6-ти томах. — М.: Просвещение. Под редакцией А. Л. Тахтаджяна, главный редактор чл.-кор. АН СССР, проф. А.А. Федоров. 1974.
.
- Происхождение моховидных
- Происхождение спорангия
Смотреть что такое "Происхождение семязачатка" в других словарях:
Происхождение цветковых растений — Представляют ли цветковые растения одну большую естественную группу, происходящую от одного общего корня, или они состоят из нескольких или даже многих независимых линий развития, имеющих происхождение от разных предков? Исключительно… … Биологическая энциклопедия
Строение семязачатков — Основные черты строения семязачатков мы уже рассматривали в предыдущем томе «Жизни растений». Но напомним еще раз, что семязачаток состоит из двух основных частой мегаспорангия (нуцеллуса) и окружающего его покрова, называемого… … Биологическая энциклопедия
Классификация и филогения — Большинство авторов все семейства хвойных объединяет в один порядок хвойные (Соniferales). Однако уже давно наметилась тенденция к разделению этой, хотя и вполне естественной, но таксономически очень дифференцированной группы на несколько … Биологическая энциклопедия
КЛАСС СЕМЕННЫЕ ПАПОРОТНИКИ ИЛИ ЛИГИНОПТЕРИДОПСИДЫ (LYGNOPTERIDOPSDA ИЛИ PTERIDOSPERMAE) — Изучая палеозойские и раннемезозойские растения, находимые в земных слоях в виде отпечатков и окаменелостей, палеоботаники нередко встречают ископаемые листья, по строению неотличимые от перистых листьев настоящих папоротников, но несущие … Биологическая энциклопедия
Семя1 — Семя цветковых растений по развитию и строению существенно отличается от семени голосеменных. Это объясняется различиями в строении семязачатка голосеменных и цветковых растений и в не меньшей степени в характере оплодотворения и… … Биологическая энциклопедия
Классификация и филогения голосеменных — Подразделение голосеменных на классы, подклассы и порядки, а также таксономический ранг и естественность самой систематической группы в целом является предметом больших разногласий. В этой книге мы рассматриваем голосеменные как… … Биологическая энциклопедия
Общая характеристика5 — Растения, относящиеся к отделу голосеменных (Pinophyta, или Gymnospermae), так же как и к следующему отделу цветковых, или покрытосеменных, отличаются от всех остальных представителей растительного мира тем, Что производят семена. Что же… … Биологическая энциклопедия
Цветковые растения — покрытосеменные (Magnoliophyta, или Angiospermae), отдел высших растений, имеющих цветок. Насчитывает свыше 400 семейств, более 12 000 родов и, вероятно, не менее 235 000 видов. По числу видов Ц. р. значительно превосходят все остальные… … Большая советская энциклопедия
Подкласс гамамелидиды (Hamamelididae) — Подкласс гамамелидид представляет собой одну из крупных ветвей родословного древа цветковых растений, которая берет начало непосредственно от магнолиид, притом, вероятно, от наиболее примитивных и древних их представителей. Среди… … Биологическая энциклопедия
Семейство санталовые (Santalaceae) — В семействе около 35 родов и примерно 425 видов. Санталовые широко распространены в тропических, субтропических и умеренных областях обоих полушарий. Лишь немногие виды встречаются в странах с холодным климатом. Дальше всех на север… … Биологическая энциклопедия
Книги
- Семязачаток цветковых растений. Строение, функции, происхождение, И. И. Шамров. В книге излагаются современные представления о развитии, строении, функциях и происхождении семязачатка цветковых растений. Описаны разработанные автором принципы организации и типизации… Подробнее Купить за 668 грн (только Украина)
- Семязачаток цветковых растений: строение, функции, происхождение, Шамров И.И.. В книге излагаются современные представления о развитии, строении, функциях и происхождении семязачатка цветковых растений. Описаны разработанные автором принципы организации и типизации… Подробнее Купить за 556 руб
- Семязачаток цветковых растений: строение, функции, происхождение, И. И. Шамров. В книге излагаются современные представления о развитии, строении, функциях и происхождении семязачатка цветковых растений. Описаны разработанные автором принципы организации и типизации… Подробнее Купить за 299 руб электронная книга
dic.academic.ru









