
Содержание
Ткани организма и их виды
У всех многоклеточных организмов — растений и животных — клетки организованы в ткани, ткани — в органы, а органы — в системы органов. И каждая из этих систем представляет собой целостную структуру, работающую на осуществление жизнедеятельности данного организма как целостности.
Обычно у многоклеточных организмов имеются группы клеток, сходных по происхождению, одинаковых по строению и выполняемым функциям. Они расположены рядом друг с другом, связаны между собой межклеточным веществом и специализированы для выполнения определённых функций, такие клетки называют тканями.
Ткани возникли в ходе эволюционного развития органического мира вместе с многоклеточностью, так как специализация клеток и группирование их в специализированные ткани и органы позволяют лучше обеспечивать процессы жизнедеятельности целостного многоклеточного организма.
В процессе эволюции у организмов сформировалось много различных тканей. Например, у червей насчитывают 10-12 специализированных тканей. Известно, что у цветковых растений насчитывается до 80 разных типов клеток, формирующих десятки различных тканей. В теле пресноводной гидры специализированных тканей немного, но имеется 10 разных типов клеток (эпителиально-мускульные, пищеварительные, нервные, стрекательные, железистые, половые и др.).
Например, у червей насчитывают 10-12 специализированных тканей. Известно, что у цветковых растений насчитывается до 80 разных типов клеток, формирующих десятки различных тканей. В теле пресноводной гидры специализированных тканей немного, но имеется 10 разных типов клеток (эпителиально-мускульные, пищеварительные, нервные, стрекательные, железистые, половые и др.).
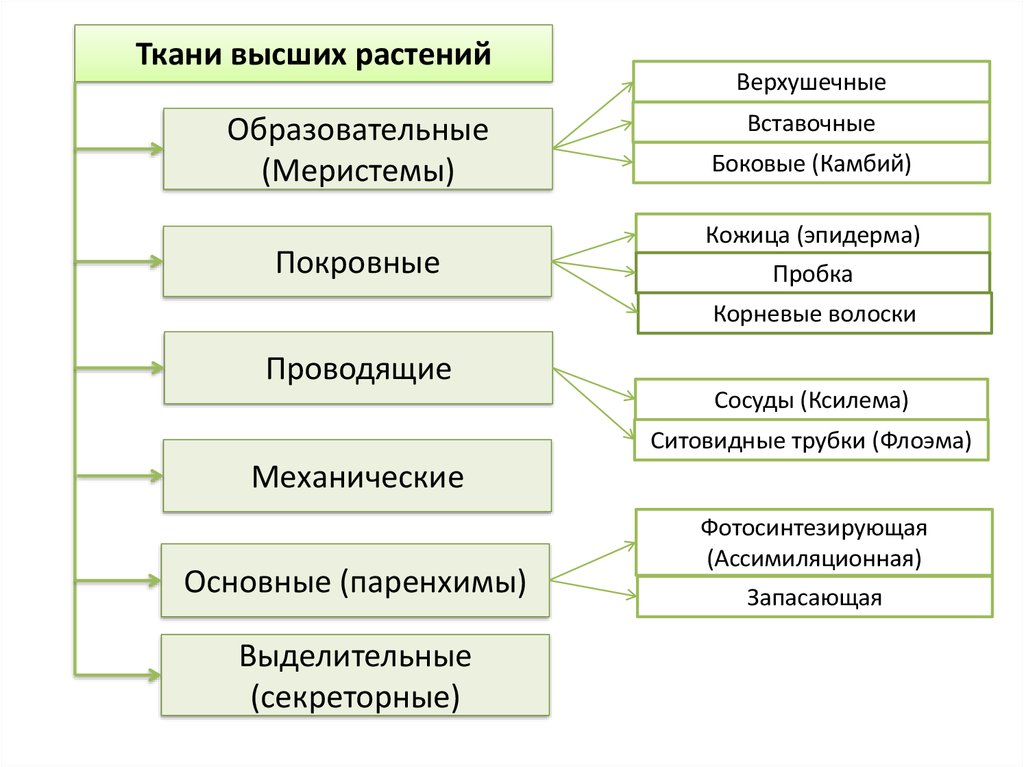
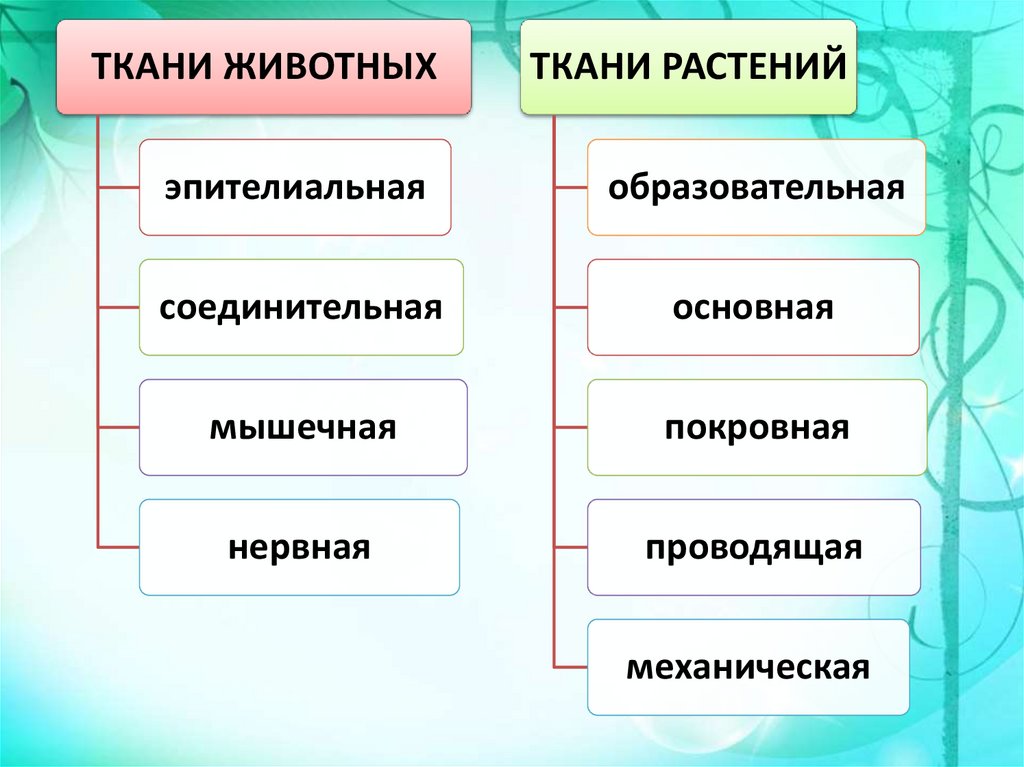
Обычно у животных различают четыре группы тканей (рис. 29):
- эпителиальную,
- соединительную,
- мышечную,
- нервную.
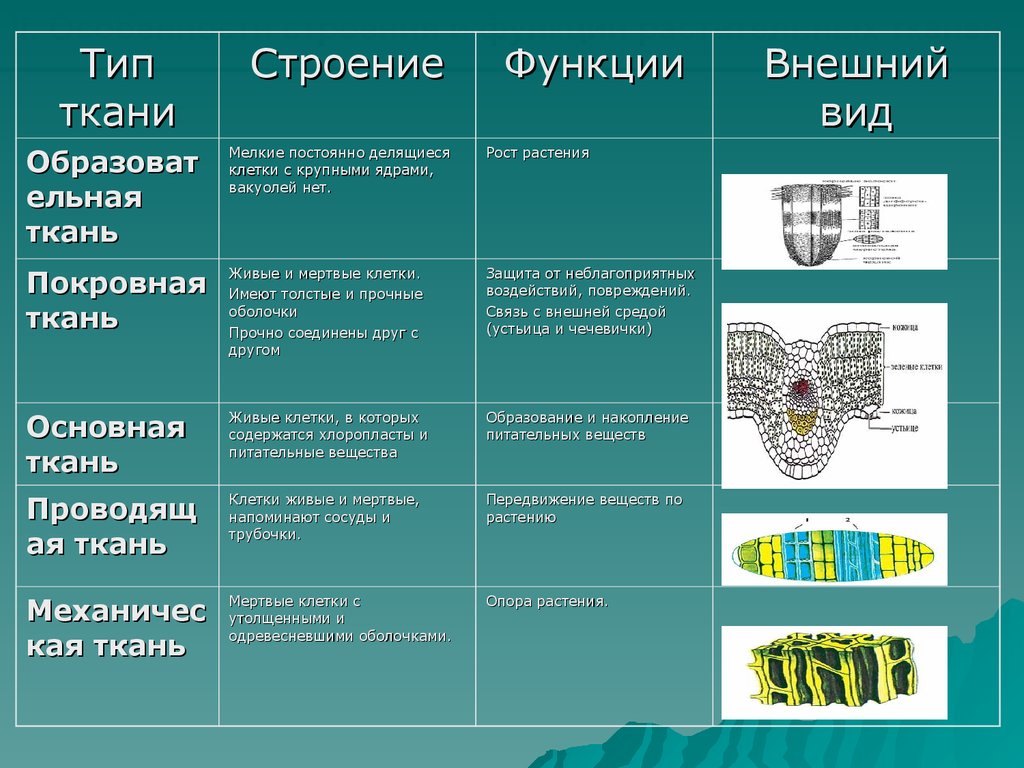
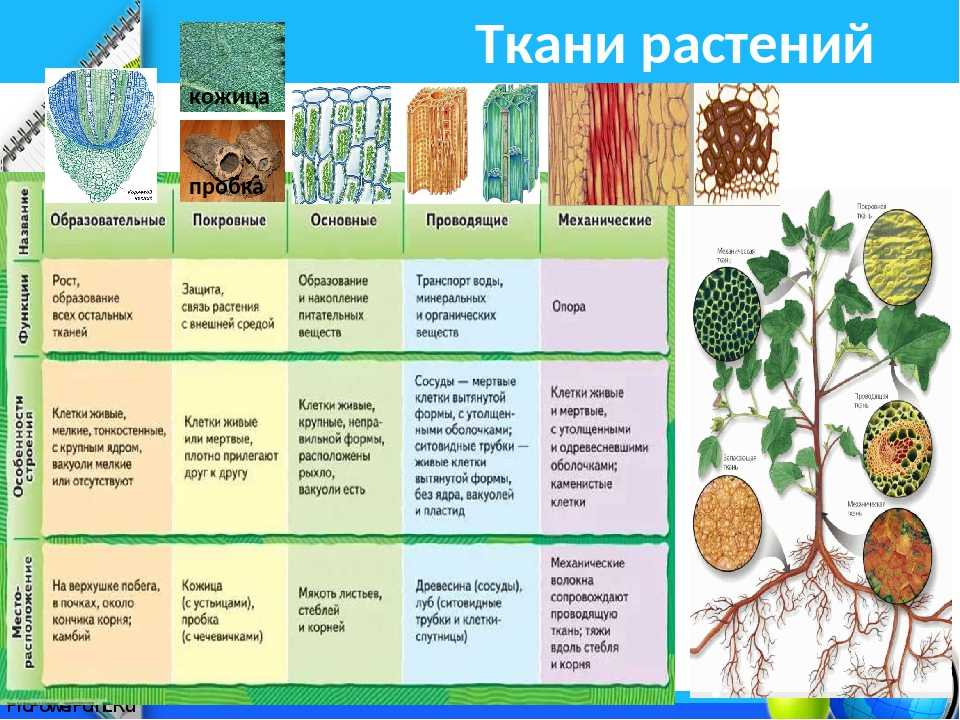
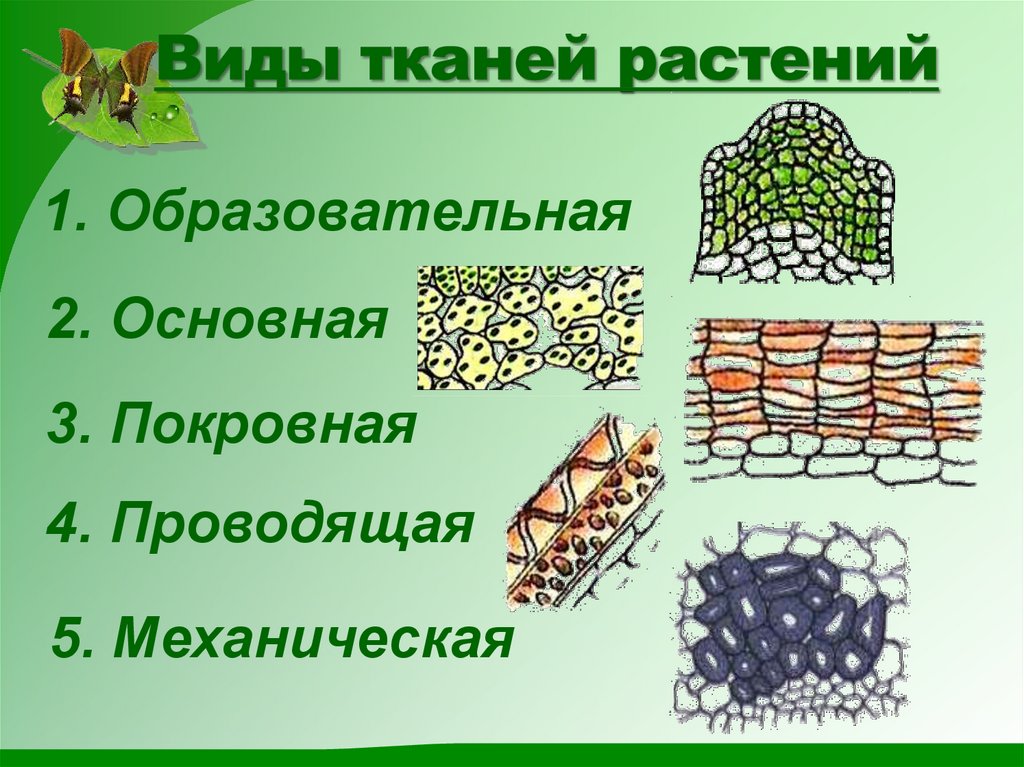
У растений групп тканей больше:
- покровная,
- проводящая,
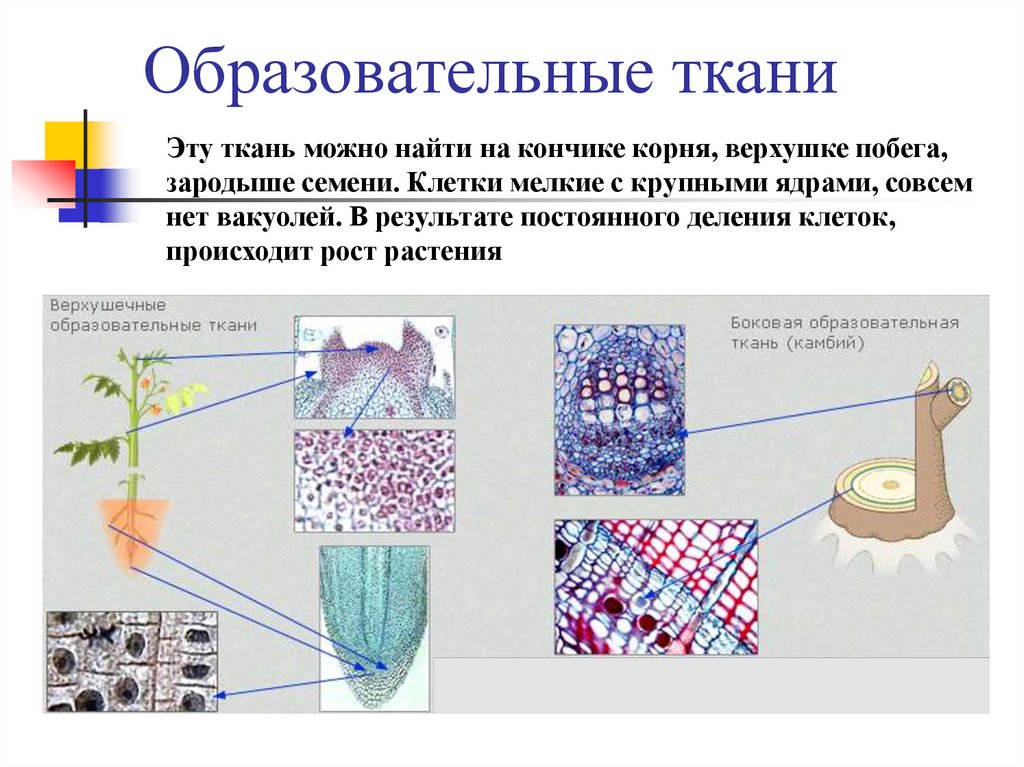
- образовательная (меристема),
- выделительная,
- механическая, Материал с сайта http://doklad-referat.ru
- основные (ассимиляционная, запасающая, воздухоносная).
Загрузка…
Некоторые ткани приобретали новые, необычные свойства. Например, нервная ткань электрического ската выполняет функцию электрозарядного устройства, а часть мышечной ткани этой хрящевой рыбы в процессе эволюции превратилась в мощные электрические аккумуляторы.
Например, нервная ткань электрического ската выполняет функцию электрозарядного устройства, а часть мышечной ткани этой хрящевой рыбы в процессе эволюции превратилась в мощные электрические аккумуляторы.
Несмотря на большое разнообразие форм, клетки разных типов обладают удивительным сходством в своих главных структурных и функциональных особенностях.
На этой странице материал по темам:
Тест по гистологии по растительным тканям
Краткий конспект на тему ткани
Реферат по теме ткани растений и животных
Доклад на тему ткани растении и их виды
Краткие конспекты по гистологии
Вопросы по этому материалу:
Материал с сайта http://Doklad-Referat.ru
Загрузка…
КУЛЬТУРА КЛЕТОК И ТКАНЕЙ • Большая российская энциклопедия
Авторы: О.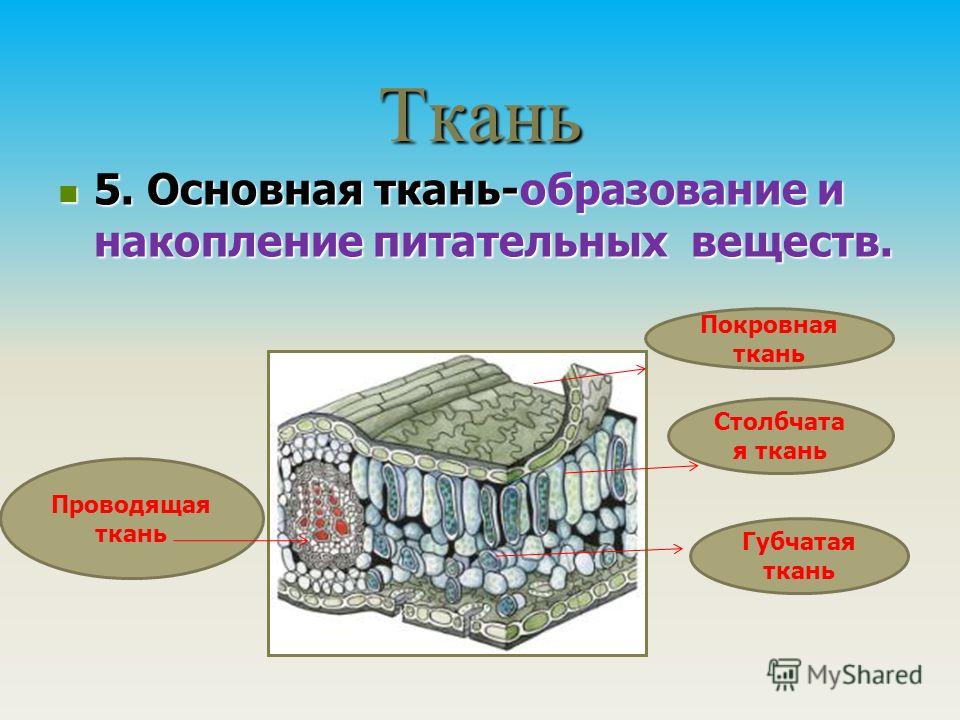 П. Кисурина-Евгеньева
П. Кисурина-Евгеньева
КУЛЬТУ́РА КЛЕ́ТОК И ТКА́НЕЙ, клетки, кусочки тканей или зачатков органов, выращенные вне организма (in vitro). В основе выращивания клеток и тканей лежит строгое соблюдение стерильности и использование спец. питат. сред, обеспечивающих поддержание жизнедеятельности культивируемых клеток и максимально сходных со средой, с которой клетки взаимодействуют в организме. Метод получения К. к. и т. является одним из важнейших в эксперим. биологии. К. к. и т. могут быть заморожены и сохраняться длительное время при темп-ре жидкого азота (–196 °C). Основополагающий эксперимент по культивированию клеток животных провёл амер. учёный Р. Гаррисон в 1907, поместив кусочек зачатка нервной системы зародыша лягушки в сгусток лимфы. Клетки зачатка оставались живыми неск. недель, из них вырастали нервные волокна. Со временем метод был усовершенствован А.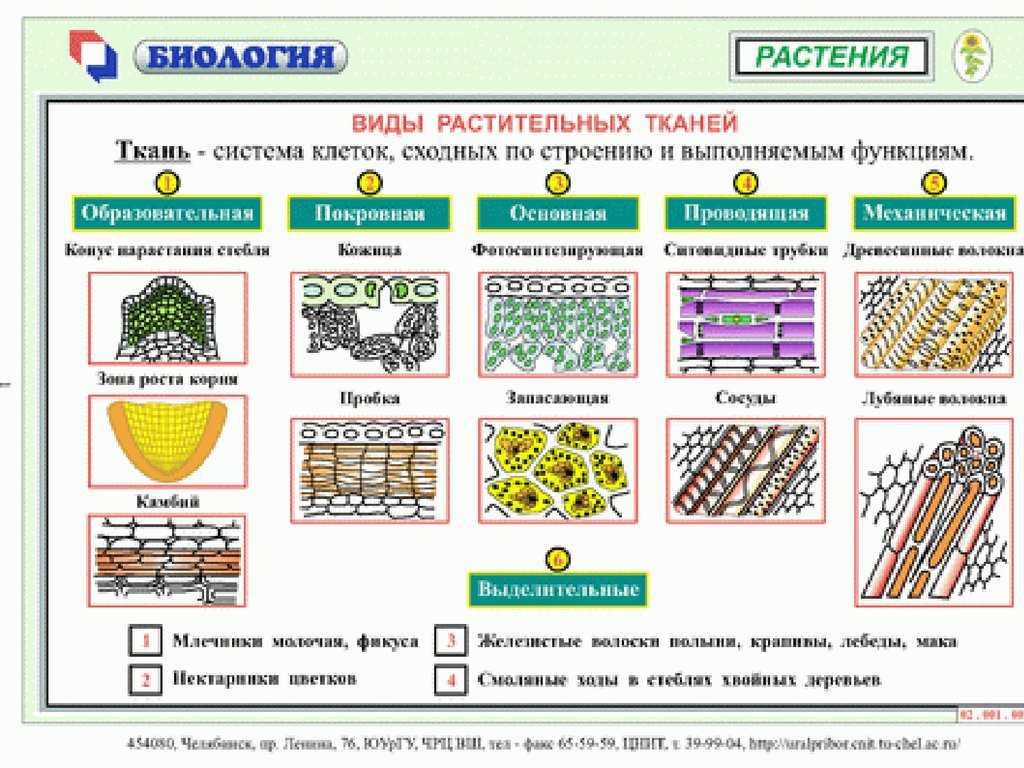 Каррелем (Франция), М. Берроузом (США), А. А. Максимовым (Россия) и др. учёными, использовавшими в качестве питат. среды плазму крови и вытяжку из тканей зародыша. В дальнейшем успехи в получении К. к. и т. были связаны с разработкой сред определённого химич. состава для культивирования разл. типов клеток. Обычно они содержат соли, аминокислоты, витамины, глюкозу, факторы роста, антибиотики, предупреждающие заражение культуры бактериями и микроскопич. грибами. Начало созданию метода К. к. и т. у растений (на кусочке флоэмы моркови) положено Ф. Стюардом (США) в 1958.
Каррелем (Франция), М. Берроузом (США), А. А. Максимовым (Россия) и др. учёными, использовавшими в качестве питат. среды плазму крови и вытяжку из тканей зародыша. В дальнейшем успехи в получении К. к. и т. были связаны с разработкой сред определённого химич. состава для культивирования разл. типов клеток. Обычно они содержат соли, аминокислоты, витамины, глюкозу, факторы роста, антибиотики, предупреждающие заражение культуры бактериями и микроскопич. грибами. Начало созданию метода К. к. и т. у растений (на кусочке флоэмы моркови) положено Ф. Стюардом (США) в 1958.
Для культивирования клеток животных и человека могут быть использованы клетки разного происхождения: эпителиальные (печень, лёгкие, молочная железа, кожа, мочевой пузырь, почка), соединительнотканные (фибробласты), скелетные (кость и хрящи), мышечные (скелетные, сердечная и гладкие мышцы), нервной системы (глиальные клетки и нейроны), железистые клетки, секретирующие гормоны (надпочечники, гипофиз, клетки островков Лангерганса), меланоциты и разл. типы опухолевых клеток. Выделяют 2 направления их культивирования: культура клеток и органная культура (культура органов и тканей). Для получения культуры клеток – генетически однородной быстро пролиферирующей популяции – кусочки ткани (обычно ок. 1 мм3) извлекают из организма, обрабатывают соответствующими ферментами (для разрушения межклеточных контактов) и образующуюся суспензию помещают в питат. среду. Культуры, полученные из эмбриональных тканей, характеризуются лучшей выживаемостью и более активным ростом (из-за низкого уровня дифференцировки и наличия стволовых клеток-предшественников в эмбрионах) по сравнению с соответствующими тканями, взятыми из взрослого организма. Нормальные ткани дают начало культурам с ограниченным временем жизни (т. н. предел Хейфлика), тогда как культуры, полученные из опухолей, способны пролиферировать неограниченно долгое время.
типы опухолевых клеток. Выделяют 2 направления их культивирования: культура клеток и органная культура (культура органов и тканей). Для получения культуры клеток – генетически однородной быстро пролиферирующей популяции – кусочки ткани (обычно ок. 1 мм3) извлекают из организма, обрабатывают соответствующими ферментами (для разрушения межклеточных контактов) и образующуюся суспензию помещают в питат. среду. Культуры, полученные из эмбриональных тканей, характеризуются лучшей выживаемостью и более активным ростом (из-за низкого уровня дифференцировки и наличия стволовых клеток-предшественников в эмбрионах) по сравнению с соответствующими тканями, взятыми из взрослого организма. Нормальные ткани дают начало культурам с ограниченным временем жизни (т. н. предел Хейфлика), тогда как культуры, полученные из опухолей, способны пролиферировать неограниченно долгое время. Однако даже в культуре нормальных клеток некоторые клетки спонтанно иммортализуются, т. е. становятся бессмертными. Они выживают и дают начало клеточным линиям с неограниченным сроком жизни. Исходно клеточная линия может быть получена из популяции клеток или из отд. клетки. В последнем случае линию называют клоновой, или клоном. При длительном культивировании под воздействием разл. факторов свойства нормальных клеток изменяются, происходит трансформация, осн. признаками которой являются нарушения морфологии клеток, изменение числа хромосом (анеуплоидия). При высокой степени трансформации введение таких клеток животному может вызывать образование опухоли. В органной культуре сохраняются структурная организация ткани, межклеточные взаимодействия, поддерживается гистологич. и биохимич. дифференцировка. Ткани, зависимые от гормонов, сохраняют чувствительность к ним и характерные ответы, железистые клетки продолжают секретировать специфич.
Однако даже в культуре нормальных клеток некоторые клетки спонтанно иммортализуются, т. е. становятся бессмертными. Они выживают и дают начало клеточным линиям с неограниченным сроком жизни. Исходно клеточная линия может быть получена из популяции клеток или из отд. клетки. В последнем случае линию называют клоновой, или клоном. При длительном культивировании под воздействием разл. факторов свойства нормальных клеток изменяются, происходит трансформация, осн. признаками которой являются нарушения морфологии клеток, изменение числа хромосом (анеуплоидия). При высокой степени трансформации введение таких клеток животному может вызывать образование опухоли. В органной культуре сохраняются структурная организация ткани, межклеточные взаимодействия, поддерживается гистологич. и биохимич. дифференцировка. Ткани, зависимые от гормонов, сохраняют чувствительность к ним и характерные ответы, железистые клетки продолжают секретировать специфич.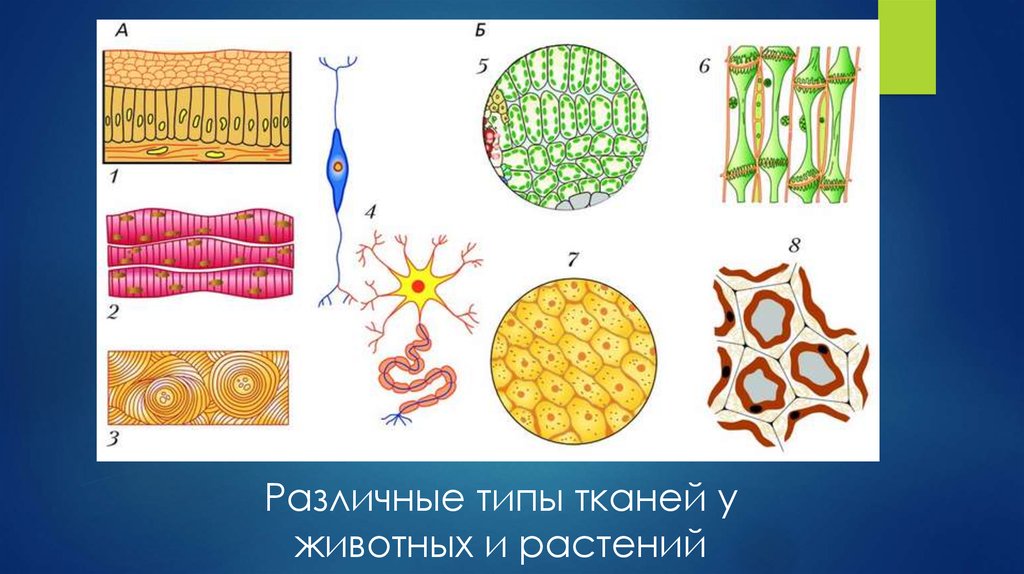 гормоны и т. д. Такие культуры выращивают в культуральном сосуде на плотиках (бумажных, миллипоровых) или на металлич. сетке, плавающих на поверхности питат. среды.
гормоны и т. д. Такие культуры выращивают в культуральном сосуде на плотиках (бумажных, миллипоровых) или на металлич. сетке, плавающих на поверхности питат. среды.
У растений культивирование клеток основано в общем на тех же принципах, что и у животных. Различия же в способах культивирования определяются структурными и биологич. особенностями клеток растений. Большинство клеток растит. тканей обладают тотипотентностью: из одной такой клетки при определённых условиях может развиться полноценное растение. Для получения культуры растит. клеток используется кусочек любой ткани (напр., каллуса) или органа (корня, стебля, листа), в котором присутствуют живые клетки. Его помещают на питат. среду, содержащую минер. соли, витамины, углеводы и фитогормоны (чаще всего цитокины и ауксины). Растит. культуры поддерживают при темп-рах от 22 до 27 °C, в темноте или при освещении.
К. к. и т. находят широкое применение в разных областях биологии и медицины. Культивирование соматич. клеток (все клетки органов и тканей за исключением половых) вне организма определило возможность развития новых способов изучения генетики высших организмов с использованием, наряду с методами классич. генетики, методов молекулярной биологии. Наибольшее развитие получила молекулярная генетика соматич. клеток млекопитающих, что связано с появившейся возможностью постановки прямых экспериментов с клетками человека. К. к. и т. используют при решении таких общебиологич. проблем, как выяснение механизмов экспрессии генов, раннего эмбрионального развития, дифференцировки и пролиферации, взаимодействия ядра и цитоплазмы, клеток со средой, адаптации к разл. химич. и физич. воздействиям, старения, злокачественной трансформации и др. , её применяют для диагностики и лечения наследств. заболеваний. В качестве тест-объектов клеточные культуры являются альтернативой использованию животных при испытании новых фармакологич. средств. Они необходимы при получении трансгенных растений, клонального размножения. Важную роль клеточные культуры играют в биотехнологии при создании гибридов, произ-ве вакцин и биологически активных веществ. См. также Клеточная инженерия.
, её применяют для диагностики и лечения наследств. заболеваний. В качестве тест-объектов клеточные культуры являются альтернативой использованию животных при испытании новых фармакологич. средств. Они необходимы при получении трансгенных растений, клонального размножения. Важную роль клеточные культуры играют в биотехнологии при создании гибридов, произ-ве вакцин и биологически активных веществ. См. также Клеточная инженерия.
Стволовые клетки растений и животных: похожие, но разные
Обзор
. 2014 май; 15(5):301-12.
doi: 10.1038/nrm3790.
Ренце Хейдстра
1
, Сабрина Сабатини
2
Принадлежности
- 1 Биология развития растений, Вагенингенский университет и исследовательский центр, Droevendaalsesteeg 1, 6708 PB Wageningen, Нидерланды.

- 2 Отделение биологии и биотехнологии, Лаборатория функциональной геномики и протеомики модельных систем, Римский университет Ла Сапиенца, Виа деи Сарди 70, 00185 Рим, Италия.
PMID:
24755933
DOI:
10.1038/nrm3790
Обзор
Renze Heidstra et al.
Nat Rev Mol Cell Biol.
2014 май.
. 2014 май; 15(5):301-12.
doi: 10.1038/nrm3790.
Авторы
Ренце Хейдстра
1
, Сабрина Сабатини
2
Принадлежности
- 1 Биология развития растений, Вагенингенский университет и исследовательский центр, Droevendaalsesteeg 1, 6708 PB Wageningen, Нидерланды.
- 2 Отделение биологии и биотехнологии, Лаборатория функциональной геномики и протеомики модельных систем, Римский университет Ла Сапиенца, Виа деи Сарди 70, 00185 Рим, Италия.
- PMID:
24755933
DOI:
10.1038/nrm3790
Абстрактный
Удивительно долгая жизнь растений и их способность к регенерации зависят от активности стволовых клеток растений. Как и у животных, стволовые клетки находятся в нишах стволовых клеток, которые производят сигналы, регулирующие баланс между самообновлением и образованием дочерних клеток, которые дифференцируются в новые ткани. Ниши стволовых клеток растений расположены в меристемах, которые представляют собой организованные структуры, ответственные за большую часть постэмбрионального развития. Непрерывное производство органов, характерное для роста растений, требует надежной регуляторной сети для поддержания баланса между плюрипотентными стволовыми клетками и дифференцирующимся потомством. Компоненты этой сети в настоящее время выяснены и предоставляют уникальную возможность для сравнения стратегий, которые были разработаны в царствах животных и растений и которые лежат в основе логики поведения стволовых клеток.
Непрерывное производство органов, характерное для роста растений, требует надежной регуляторной сети для поддержания баланса между плюрипотентными стволовыми клетками и дифференцирующимся потомством. Компоненты этой сети в настоящее время выяснены и предоставляют уникальную возможность для сравнения стратегий, которые были разработаны в царствах животных и растений и которые лежат в основе логики поведения стволовых клеток.
Похожие статьи
Ниши стволовых клеток растений.
Айхингер Э., Корнет Н., Фридрих Т., Ло Т.
Айхингер Э. и соавт.
Annu Rev Plant Biol. 2012;63:615-36. doi: 10.1146/annurev-arplant-042811-105555. Epub 2012 9 февраля.
Annu Rev Plant Biol. 2012.PMID: 22404469
Обзор.
Поддержание стволовых клеток растений включает прямую транскрипционную репрессию программы дифференцировки.
Ядав Р.К., Пералес М., Груэль Дж., Оно С., Хейслер М., Гирке Т., Йонссон Х., Редди Г.В.
Ядав Р.К. и др.
Мол Сист Биол. 2013;9:654. doi: 10.1038/msb.2013.8.
Мол Сист Биол. 2013.PMID: 23549482
Бесплатная статья ЧВК.ULTRAPETALA1 поддерживает нишу корневых стволовых клеток арабидопсиса независимо от ARABIDOPSIS TRITHORAX1.
Орнелас-Аяла Д., Вега-Леон Р., Петроне-Мендоса Э., Гарай-Арройо А., Гарсия-Понсе Б., Альварес-Буйя Э.Р., Санчес М.П.
Орнелас-Аяла Д. и соавт.
Новый Фитол. 2020 фев; 225(3):1261-1272. дои: 10.1111/nph.16213. Эпаб 201928 окт.
Новый Фитол. 2020.PMID: 31545512
Полученный от организатора сигнал WOX5 поддерживает корневые стволовые клетки колумеллы посредством хроматин-опосредованной репрессии экспрессии CDF4.
Пи Л., Айхингер Э., Ван дер Грааф Э., Ллавата-Перис С.И., Вейерс Д., Хенниг Л., Грут Э., Ло Т.
Пи Л и др.
Ячейка Дев. 2015 8 июня; 33 (5): 576-88. doi: 10.1016/j.devcel.2015.04.024. Epub 2015 28 мая.
Ячейка Дев. 2015.PMID: 26028217
Основные пути, контролирующие поддержание меристемы побегов.
Ли К., Кларк С.Э.
Ли С и др.
Wiley Interdiscip Rev Dev Biol. 2013 сен-октябрь;2(5):671-84. doi: 10.1002/wdev.110. Epub 2013 27 февраля.
Wiley Interdiscip Rev Dev Biol. 2013.PMID: 24014453
Обзор.

Посмотреть все похожие статьи
Цитируется
Профилирование отдельных клеток освещает различные клеточные траектории в растениях.
Чжэн Х.
С., Ву Ф.Х., Ли С.М., Чжан Х.С., Суй Н.
Чжэн Х.С. и др.
Абиотех. 2 апр. 2021 г.; 2(1):64–78. doi: 10.1007/s42994-021-00040-7. Электронная коллекция 2021 март.
Абиотех. 2021.PMID: 36304478
Бесплатная статья ЧВК.Обзор.
Инициация и поддержание стволовых клеток растений в апикальных меристемах корней и побегов.
Ван Дж, Су Ю, Конг Х, Дин Зи, Чжан ХС.
Ван Дж. и др.
Абиотех. 2020 4 мая; 1(3):194-204. doi: 10.1007/s42994-020-00020-3. электронная коллекция 2020 июль.
Абиотех. 2020.PMID: 36303567
Бесплатная статья ЧВК.Обзор.
NAC1 поддерживает активность корневой меристемы путем подавления транскрипции E2Fa в Arabidopsis .
Се С., Дин З.
Се С и др.
Int J Mol Sci. 2022 14 октября; 23 (20): 12258. дои: 10.3390/ijms232012258.
Int J Mol Sci. 2022.PMID: 36293114
Бесплатная статья ЧВК.Опосредованная аутофагией регуляция различных меристем у растений.
Ченг С., Ван К., Мангвар Х., Лю Ф.
Ченг С. и др.
Int J Mol Sci. 2022 2 июня; 23 (11): 6236. дои: 10.3390/ijms23116236.
Int J Mol Sci. 2022.PMID: 35682913
Бесплатная статья ЧВК.Обзор.
Динамика роста клеток двух типов апикальных меристем гаметофитов папоротников.
Ву С, Ян А, Ян Х, Бэнкс Дж А, Чжан С, Чжоу Ю.
Ву С и др.
Плант Дж. 2022 июль; 111 (1): 149-163. doi: 10.1111/tpj.15784. Epub 2022 12 мая.
Завод Дж. 2022.PMID: 35451138
Бесплатная статья ЧВК.
 С., Ву Ф.Х., Ли С.М., Чжан Х.С., Суй Н.
С., Ву Ф.Х., Ли С.М., Чжан Х.С., Суй Н.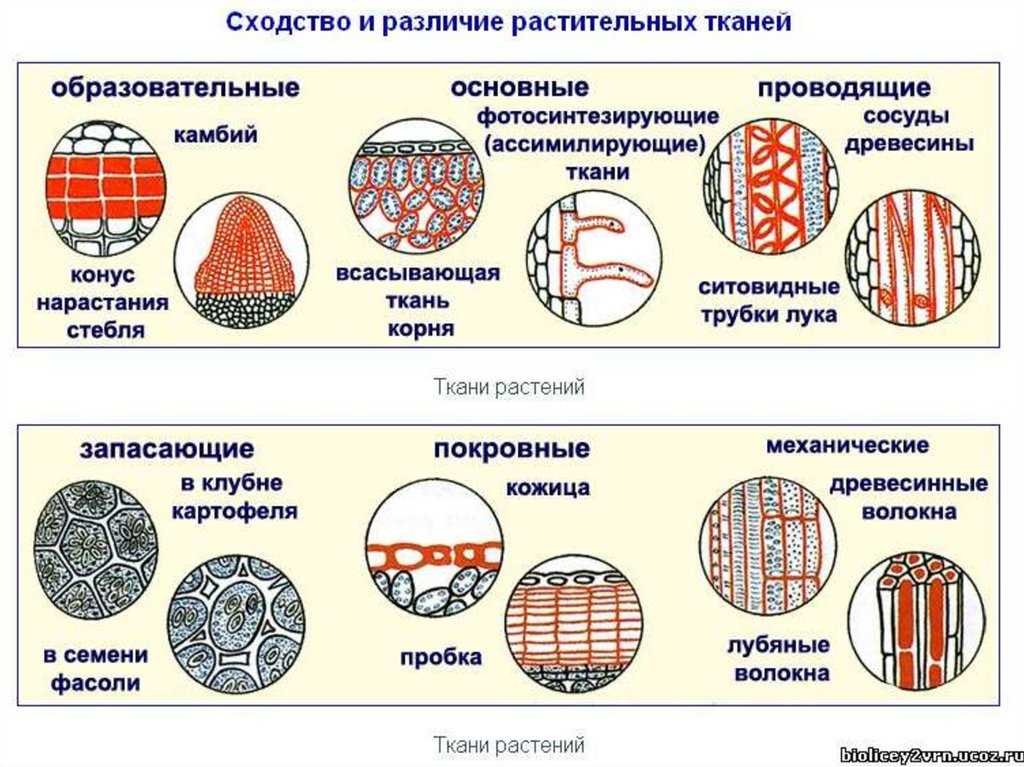 2022 14 октября; 23 (20): 12258. дои: 10.3390/ijms232012258.
2022 14 октября; 23 (20): 12258. дои: 10.3390/ijms232012258.Просмотреть все статьи «Цитируется по»
использованная литература
Растительная клетка.
2008 апр; 20 (4): 934-46—
пабмед
Proc Natl Acad Sci U S A. 31 марта 2009 г .; 106 (13): 5424-9.
—
пабмед
Клетка. 2011 15 апреля; 145(2):242-56
—
пабмед
Биохим Биофиз Акта. 2013 июль; 1830 (7): 3917-27
—
пабмед
Геномика BMC.
2013 ноя 20;14:809—
пабмед
 2008 апр; 20 (4): 934-46
2008 апр; 20 (4): 934-46 2013 ноя 20;14:809
2013 ноя 20;14:809 Типы публикаций
термины MeSH
Басби, С.Х., и Ганнинг, Б.Е.С., 1980, Наблюдения за префазными полосами микротрубочек в однорядных волосках, устьичных комплексах сахарного тростника и корневых меристемах Cyperus, Eur. Дж. Клеточная биология.
21 : 214–223.ПабМед
КАСGoogle ученый
Clayton, L.
C., and Lloyd, C.W., 1984, Связь между плоскостью деления и геометрией веретена в клетках Allium, обработанных CIPC и гризеофульвином: исследование антитубулина, Eur. Дж. Селл. биол.
34 : 248–253.ПабМед
КАСGoogle ученый
Clayton, L.C., and Lloyd, C.W., 1985, Организация актина во время клеточного цикла в меристематических растительных клетках. Актин присутствует в цитокинетическом фрагмопласте, Exp. Сотовый рез.
156 : 231–238.Перекрёстная ссылка
пабмед
КАСGoogle ученый
Clayton, L.C., Black, C.M., and Lloyd, C.W., 1985, Места зарождения микротрубочек в клетках высших растений, идентифицированные аутоантителами против перицентриолярного материала, J. Cell Biol.
101 : 319–324.Перекрёстная ссылка
пабмед
КАСGoogle ученый
Галатис, Б.
, Апостоланос, П., и Катсарос, К., 1983, Синхронная организация двух препрофазных полос микротрубочек и окончательное расположение клеточных пластинок в материнских клетках вспомогательных клеток некоторых видов Triticum, Protoplasma
117 : 24–39.Перекрёстная ссылка
Google ученый
Goosen-De Roo, L., Bahuizen, R., Van Spronsen, P.C., and Libbenga, K.R., 1984, Наличие протяженных фрагмосом, содержащих элементы цитоскелета, в веретеновидных камбиальных клетках Fraxinus excelsior L., Protoplasma
122 : 145–152.Перекрёстная ссылка
Google ученый
Грин, П.Б., 1963, О механизмах удлинения, в: Цитодифференцировка и макромолекулярный синтез (М. Локк, изд.), стр. 203–234, Academic Press, Нью-Йорк.
Google ученый
Грин, П.
Б., и Ланг, Дж. М., 1981, На пути к биофизическому объяснению органогенеза: наблюдения двойного лучепреломления на регенерирующих листьях суккулентного Graptopetalum paraguayense E. Walther, Planta
151 : 413–426.Google ученый
Gunning, B.E.S., 1982a, Корень водяного папоротника Azolla . Клеточная основа развития и множественные роли кортикальных микротрубочек, в: Порядок развития: , его происхождение и регулирование (С. Субтельны и П. Б. Грин, ред.), стр. 379–421, Алан Р. Лисс, Нью-Йорк.
Google ученый
Ганнинг, Б.Е.С., 1982b, Цитокинетический аппарат: его развитие и пространственная регуляция, в: Цитоскелет в росте и развитии растений (C.W. Lloyd, ed.), стр. 229–292, Academic Press, Нью-Йорк.
Google ученый
Gunning, B.E.S., Hughes, J.E., and Hardham, A.
R., 1978a, Формирующие и пролиферативные клеточные деления, дифференцировка клеток и изменения в развитии меристемы корней Azolla, Planta
143 : 121–144.Перекрёстная ссылка
Google ученый
Ганнинг, Б.Е.С., Хардхэм, А.Р., и Хьюз, Дж.Э., 19 лет78b, Препрофазные полосы микротрубочек во всех категориях формирующего и пролиферативного деления клеток в корнях Azolla, Planta
143 : 145–160.Перекрёстная ссылка
Google ученый
Gunning, B.E.S., Hardham, A.R., and Hughes, J.E., 1978c, Доказательства инициации микротрубочек в дискретных областях клеточной коры в клетках кончиков корней Azolla и гипотеза о развитии кортикальных массивов микротрубочек, Планта
143 : 161–179.Перекрёстная ссылка
Google ученый
Hardham, A.
R., and McCully, M.E., 1982, Перепрограммирование клеток после повреждения корней гороха (Pisum sativum L.). I. Деление клеток и дифференцировка новых сосудистых элементов, Протоплазма
112 : 143–151.Перекрёстная ссылка
Google ученый
Hardham, A.R., Green, P.B., and Lang, J.M., 1980, Реорганизация кортикальных микротрубочек и отложение целлюлозы во время формирования листьев у Graptopetalum paraguayense, Planta
149 : 181–195.Перекрёстная ссылка
КАСGoogle ученый
Heath, I.B., 1974, Единая гипотеза о роли связанных с мембраной ферментных комплексов и микротрубочек в синтезе клеточной стенки растений, Ж. Теор. биол.
48 : 445–449.Перекрёстная ссылка
пабмед
КАСGoogle ученый
Heath, I.
B., and Seagull, R.W., 1982, Ориентированные целлюлозные фибриллы и цитоскелет: критическое сравнение моделей, в: The Cytoskeleton in Plant Growth and Development (C.W. Lloyd, ed.), стр. 163 –182, Academic Press, Нью-Йорк.Google ученый
Herth, W., 1983, Массивы «розеток» плазматических мембран, участвующих в образовании микрофибрилл целлюлозы Spirogyra, Planta
159 : 347–356.Перекрёстная ссылка
Google ученый
Itoh, T., and Shimaji, K., 1976, Ориентация микротрубочек и микротрубочек в клетках паренхимы коры тополя при росте растяжением, Bot. Маг. Токио
89 : 291–308.Перекрёстная ссылка
Google ученый
Ланг Дж. М., Эйзингер В. Р. и Грин П. Б., 1982, Влияние этилена на ориентацию микротрубочек и целлюлозных микрофибрилл эпикотильных клеток гороха с полиламеллярными клеточными стенками, Protoplasma
110 : 5–14.Перекрёстная ссылка
КАСGoogle ученый
Ledbetter, M.C., and Porter, K.R., 1963, «Микротрубочки» в тонкой структуре растительных клеток, J. Cell Biol.
19 : 239–250.Перекрёстная ссылка
пабмед
КАСGoogle ученый
Lintilhac, P.M., 1974, Дифференциация, органогенез и тектоника ориентации клеточных стенок. III. Теоретические соображения механики клеточных стенок, Am. Дж. Бот.
61 902:26: 230–237.Перекрёстная ссылка
Google ученый
Lloyd, C.W., 1983, Спиральные микротрубчатые массивы в корневых волосках лука, Nature (Lond.)
305 : 311–313.Перекрёстная ссылка
КАСGoogle ученый
Lloyd, C.W., 1984, К динамической спиральной модели влияния микротрубочек на структуру стенок растений, Int.
Преподобный Цитол.
86 : 1–51.Перекрёстная ссылка
Google ученый
Lloyd, C.W., and Barlow, P.W., 1982, Координация клеточного деления и удлинения: роль цитоскелета, в: The Cytoskeleton in Plant Growth and Development (C.W. Lloyd, ed.), стр. 203–228, Academic Press, Нью-Йорк.
Google ученый
Ллойд, К.В., Лоу, С.Б., и Пис, Г.В., 19 лет80, Способ действия 2,4-D в противодействии удлинению клеток моркови, выращенных в культуре, J. Cell Sci.
45 : 257–268.ПабМед
КАСGoogle ученый
Ллойд К.В., Слабас А.Р., Пауэлл А.Дж. и Лоу С.Б., 1980, Микротрубочки, протопласты и форма растительных клеток. Иммунофлуоресцентное исследование, Planta
147 : 500–506.перекрестная ссылка
Google ученый
Миллер, Дж.
Х., 1980, Ориентация плоскости деления клеток в гаметофитах папоротника: роли формы клеток и стресса, Am. Дж. Бот.
67 : 534–542.Перекрёстная ссылка
Google ученый
Монтезинос, Д., 1982, Роль плазматической мембраны в сборке микрофибрилл целлюлозы, в: Цитоскелет в росте и развитии растений (CW Lloyd, изд.), стр. 147–162, Academic Press, Нью-Йорк.
Google ученый
Палевиц, Б.А., 1982, Устьичный комплекс как модель участия цитоскелета в дифференцировке клеток, в: Цитоскелет в росте и развитии растений (CW Lloyd, ed.), стр. 346–376, Academic Press , Нью-Йорк.
Google ученый
Палевиц Б.А. и Хеплер П.К., 19 лет76, Ориентация микрофибрилл целлюлозы и формирование клеток в развивающихся замыкающих клетках Allium: роль микротрубочек и накопление ионов, Planta
132 : 71–93.Перекрёстная ссылка
Google ученый
Pickett-Heaps, J.D., and Northcote, D.H., 1966, Деление клеток при формировании устьичного комплекса молодых листьев пшеницы, J. Cell Sci.
1 902:26: 121–128.ПабМед
КАСGoogle ученый
Престон, Р. Д., 1961, Целлюлозо-белковые комплексы в клеточных стенках растений, в: Макромолекулярные комплексы (М. В. Эддс, изд.), стр. 229–253, Рональд Пресс, Нью-Йорк.
Google ученый
Престон, Р. Д., 1974, Физическая биология клеточных стенок растений , Чепмен и Холл, Лондон.
Google ученый
Престон, Р. Д., 1982, Случай мультисетевого роста в растущих стенках растительных клеток, Planta
155 : 356–363.Перекрёстная ссылка
Google ученый
Ray, P.
M., 1967, Радиоавтографическое исследование отложения клеточных стенок в растущих растительных клетках, J. Cell Biol.
35 : 659–674.Перекрёстная ссылка
пабмед
КАСGoogle ученый
Робертс, И. Н., Ллойд, К. В., и Робертс, К., 1985, Этилен-индуцированная переориентация микротрубочек: опосредование спиральными массивами, Planta
164 : 439–447.Перекрёстная ссылка
Google ученый
Робинсон Д.Г. и Квадер Х., 1982, Синдром микротрубочек и микрофибрилл, в: Цитоскелет в росте и развитии растений (CW Lloyd, ed.), стр. 109–126, Academic Press, Нью-Йорк.
Google ученый
Roelofsen, A., 1958, Структура клеточной стенки в связи с поверхностным ростом, Acta Bot. Нерл.
7 : 77–89.Google ученый
Roelofsen, P.
A., 1965, Ультраструктура стенки растущих клеток и ее связь с направлением роста, Доп. Бот. Рез.
2 : 68–149.Google ученый
Roelofsen, P.A., и Houwink, A.L., 1953, Структура и рост первичной стенки в волосках некоторых растений и спорангиофоре Phycomyces, Acta Bot. Нерл.
2 : 218–225.Google ученый
Роланд, Дж. К., и Виан, Б., 19 лет79, Стенка растущей растительной клетки: ее трехмерная организация, Int. Преподобный Цитол.
61 : 129–166.Перекрёстная ссылка
Google ученый
Sargent, C., 1978, Дифференциация поперечно-фибриллярной наружной эпидермальной стенки во время роста в длину у Hordium vulgare L., Protoplasma
95 : 309–320.Перекрёстная ссылка
Google ученый
Sawhney, V.
K., and Srivastava, L.M., 1974, Гибберелловая кислота индуцирует удлинение гипокотилей салата и его ингибирование колхицином, Can. Дж. Бот.
52 : 259–264.Перекрёстная ссылка
КАСGoogle ученый
Зингер, С.Дж., и Николсон, Г.Л., 1972, Жидкостно-мозаичная модель структуры клеточных мембран, Наука
175 : 720–731.Перекрёстная ссылка
пабмед
КАСGoogle ученый
Sinnott, E.W., and Bloch, R., 1940, Цитоплазматическое поведение при делении вакуолярных растительных клеток, Proc. Натл. акад. науч. США
26 : 223–227.Перекрёстная ссылка
пабмед
КАСGoogle ученый
Стэхелин Л. А. и Гиддингс Т. Х., 19 лет82, Мембрано-опосредованный контроль микрофибриллярного порядка клеточной стенки, в: Порядок развития: его происхождение и регуляция (S.
 C., and Lloyd, C.W., 1984, Связь между плоскостью деления и геометрией веретена в клетках Allium, обработанных CIPC и гризеофульвином: исследование антитубулина, Eur. Дж. Селл. биол.
C., and Lloyd, C.W., 1984, Связь между плоскостью деления и геометрией веретена в клетках Allium, обработанных CIPC и гризеофульвином: исследование антитубулина, Eur. Дж. Селл. биол. 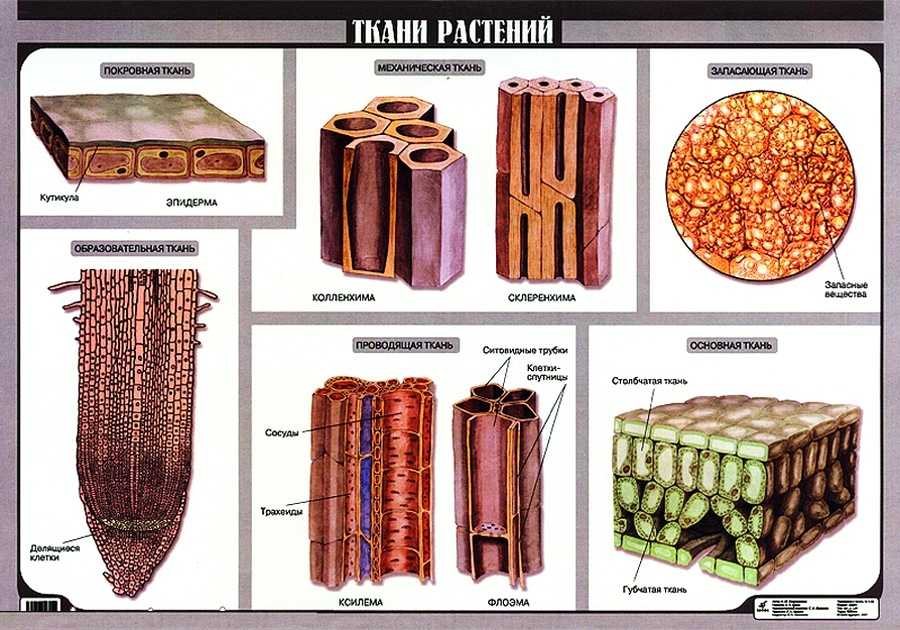 , Апостоланос, П., и Катсарос, К., 1983, Синхронная организация двух препрофазных полос микротрубочек и окончательное расположение клеточных пластинок в материнских клетках вспомогательных клеток некоторых видов Triticum, Protoplasma
, Апостоланос, П., и Катсарос, К., 1983, Синхронная организация двух препрофазных полос микротрубочек и окончательное расположение клеточных пластинок в материнских клетках вспомогательных клеток некоторых видов Triticum, Protoplasma 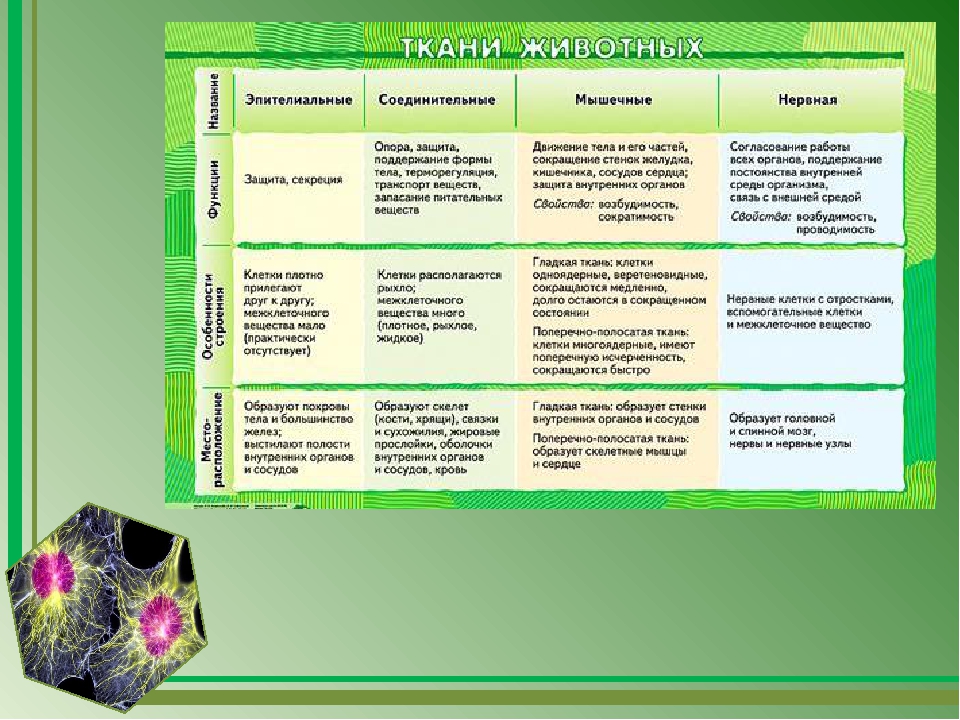 Б., и Ланг, Дж. М., 1981, На пути к биофизическому объяснению органогенеза: наблюдения двойного лучепреломления на регенерирующих листьях суккулентного Graptopetalum paraguayense E. Walther, Planta
Б., и Ланг, Дж. М., 1981, На пути к биофизическому объяснению органогенеза: наблюдения двойного лучепреломления на регенерирующих листьях суккулентного Graptopetalum paraguayense E. Walther, Planta  R., 1978a, Формирующие и пролиферативные клеточные деления, дифференцировка клеток и изменения в развитии меристемы корней Azolla, Planta
R., 1978a, Формирующие и пролиферативные клеточные деления, дифференцировка клеток и изменения в развитии меристемы корней Azolla, Planta 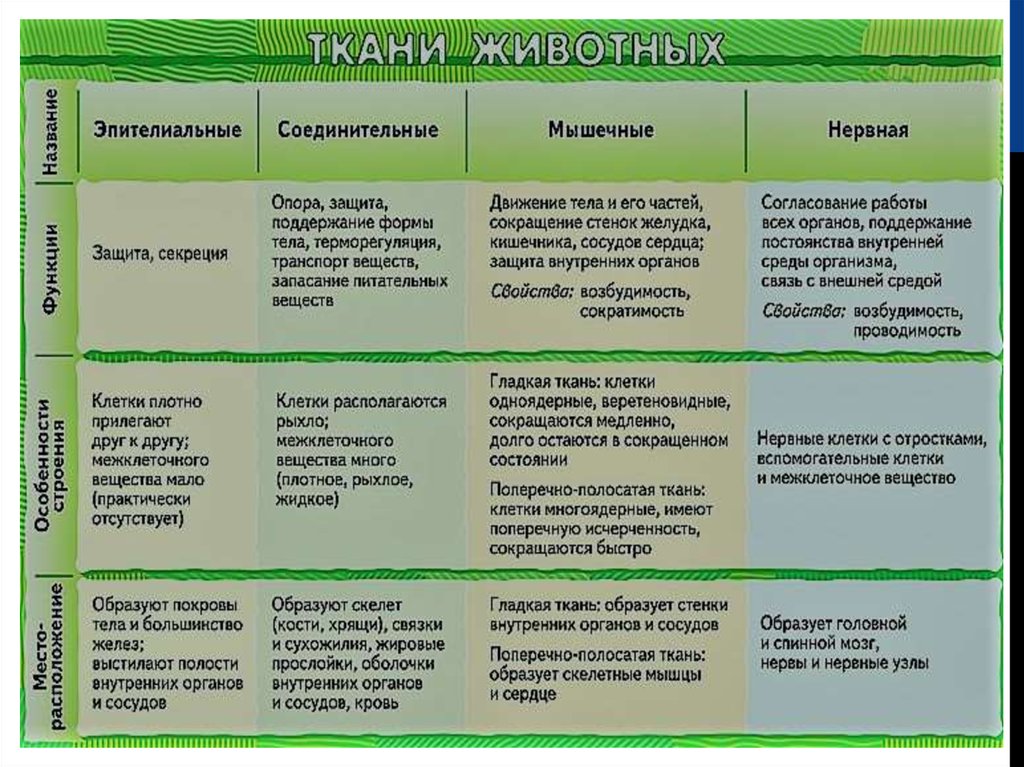 R., and McCully, M.E., 1982, Перепрограммирование клеток после повреждения корней гороха (Pisum sativum L.). I. Деление клеток и дифференцировка новых сосудистых элементов, Протоплазма
R., and McCully, M.E., 1982, Перепрограммирование клеток после повреждения корней гороха (Pisum sativum L.). I. Деление клеток и дифференцировка новых сосудистых элементов, Протоплазма 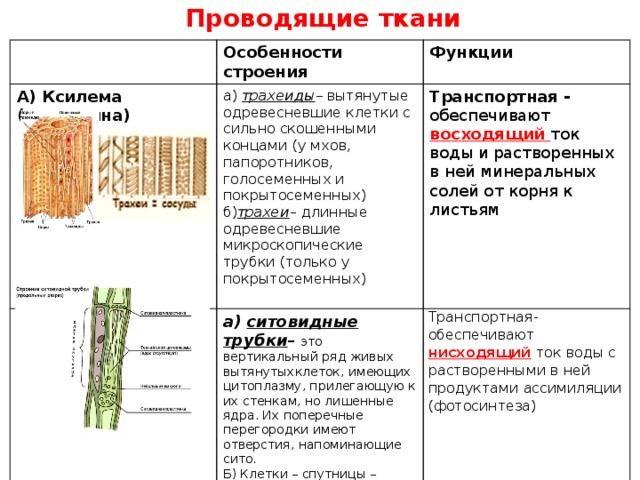 B., and Seagull, R.W., 1982, Ориентированные целлюлозные фибриллы и цитоскелет: критическое сравнение моделей, в: The Cytoskeleton in Plant Growth and Development (C.W. Lloyd, ed.), стр. 163 –182, Academic Press, Нью-Йорк.
B., and Seagull, R.W., 1982, Ориентированные целлюлозные фибриллы и цитоскелет: критическое сравнение моделей, в: The Cytoskeleton in Plant Growth and Development (C.W. Lloyd, ed.), стр. 163 –182, Academic Press, Нью-Йорк.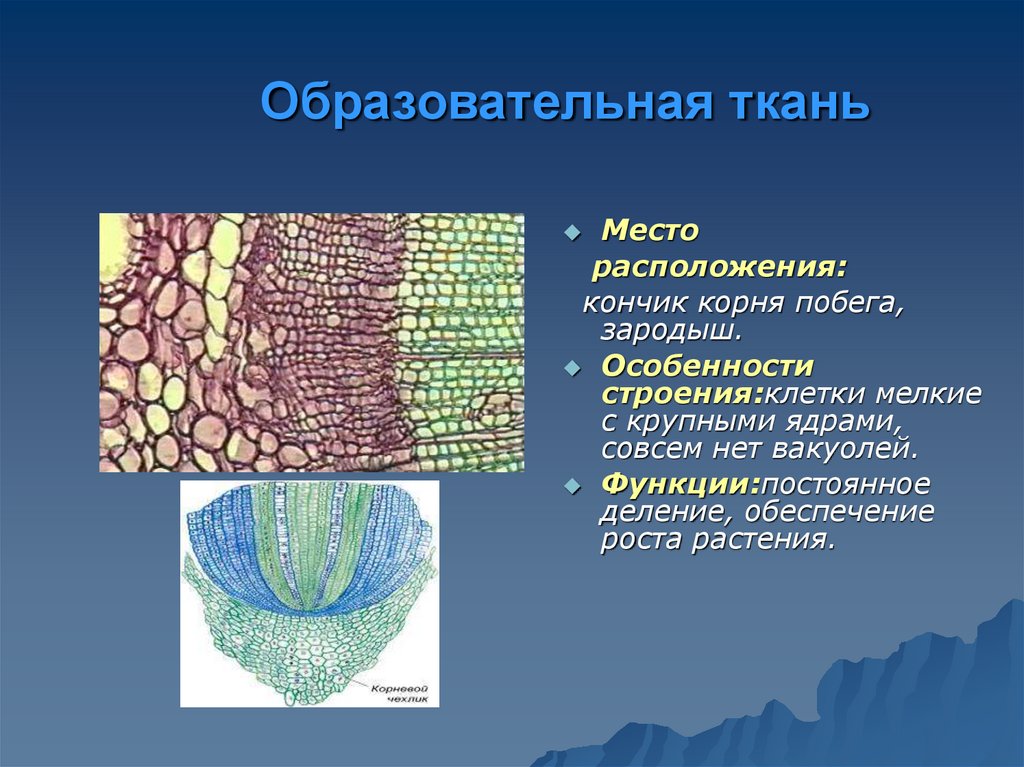
 Преподобный Цитол.
Преподобный Цитол.  Х., 1980, Ориентация плоскости деления клеток в гаметофитах папоротника: роли формы клеток и стресса, Am. Дж. Бот.
Х., 1980, Ориентация плоскости деления клеток в гаметофитах папоротника: роли формы клеток и стресса, Am. Дж. Бот. 
 M., 1967, Радиоавтографическое исследование отложения клеточных стенок в растущих растительных клетках, J. Cell Biol.
M., 1967, Радиоавтографическое исследование отложения клеточных стенок в растущих растительных клетках, J. Cell Biol. 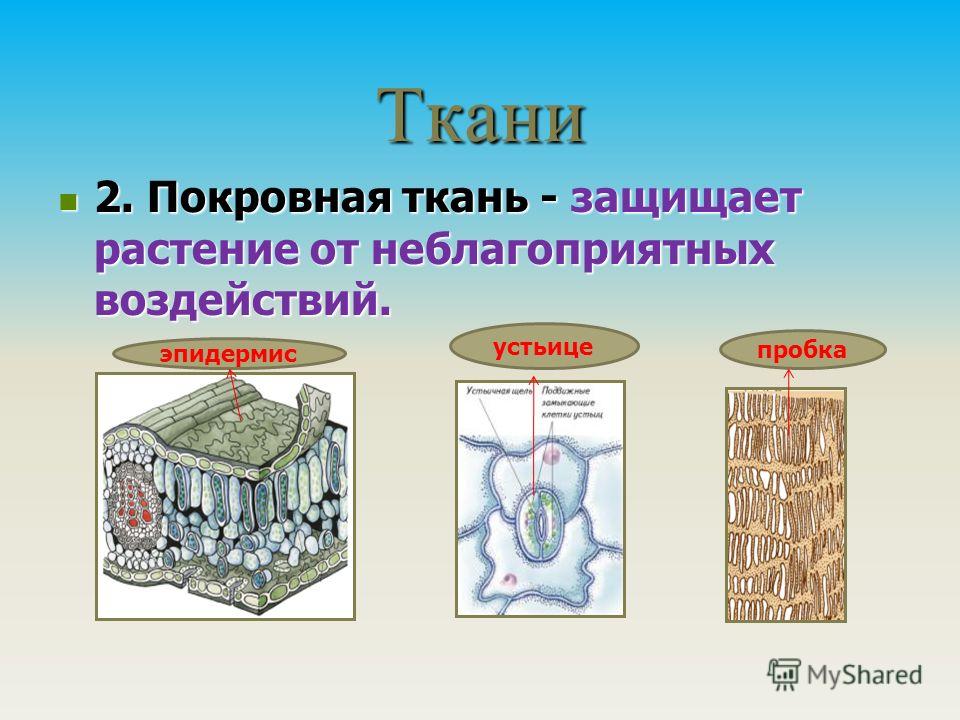 A., 1965, Ультраструктура стенки растущих клеток и ее связь с направлением роста, Доп. Бот. Рез.
A., 1965, Ультраструктура стенки растущих клеток и ее связь с направлением роста, Доп. Бот. Рез.  K., and Srivastava, L.M., 1974, Гибберелловая кислота индуцирует удлинение гипокотилей салата и его ингибирование колхицином, Can. Дж. Бот.
K., and Srivastava, L.M., 1974, Гибберелловая кислота индуцирует удлинение гипокотилей салата и его ингибирование колхицином, Can. Дж. Бот. 