
Содержание
какие типы есть, что значит очередное, стадии развития листа
Содержание:
-
Листорасположение у растений — основные термины и понятия -
Морфология и структура -
Принципы листорасположения, какие виды бывают -
Видоизменения листьев, примеры растений
Содержание
-
Листорасположение у растений — основные термины и понятия -
Морфология и структура -
Принципы листорасположения, какие виды бывают -
Видоизменения листьев, примеры растений
Листорасположение у растений — основные термины и понятия
Каждое растение характеризуется не только индивидуальными размерами, формой листьев и цветов. Систематическим признаком выступает также листорасположение (phyllotaxis). Оно определяется порядком закладки зачатков листа в районе конуса нарастания.
Систематическим признаком выступает также листорасположение (phyllotaxis). Оно определяется порядком закладки зачатков листа в районе конуса нарастания.
Лист – незаменимый орган растения, с помощью которого осуществляется газообмен, транспирация и процессы фотосинтеза. Именно поэтому у него пластинчатая структура, позволяющая максимально дать возможность клеткам «насытиться» солнечным светом. Для растения это – орган дыхания, в процессе которого происходит гуттация и испарения влаги. В то же время в листовых пластинах происходит задержка жидкости и растворенных в ней питательных веществ.
В основе классификации представителей растительного мира лежит много свойств, в том числе особенность расположения листьев на стебле. Существует строгий порядок их закладки, целью которого является обеспечение доступа к солнечному свету. Одни листья располагаются спиралевидно по часовой стрелке, другие – против.
Осторожно! Если преподаватель обнаружит плагиат в работе, не избежать крупных проблем (вплоть до отчисления).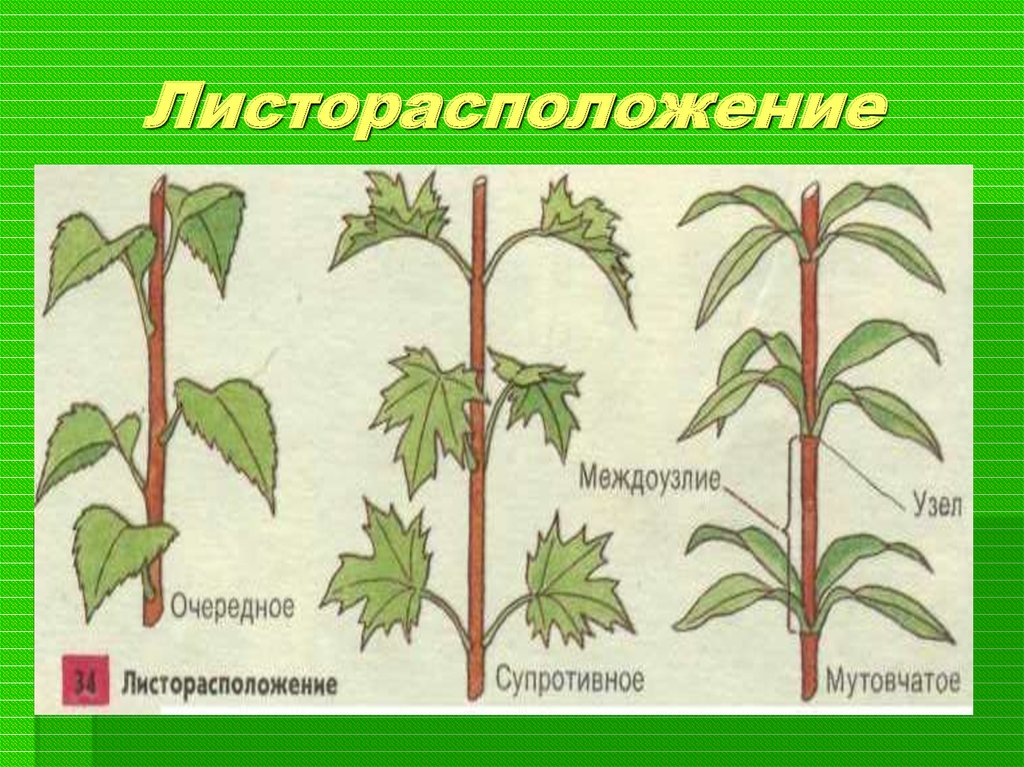 Если нет возможности написать самому, закажите тут.
Если нет возможности написать самому, закажите тут.
Ботаники заметили и математически выразили даже определенную закономерность – последовательность Фибоначчи, – касающуюся угла их расхождения. Листы могут располагаться под углом 180 градусов (тогда в обороте помещается 2 листа), либо 125 градусов (в обороте — 3 листа), при 154 градусах – 5 листьев, при 115 – 8 листьев.
Общепринятая характеристика листорасположения опирается на три понятия:
- очередное;
- супротивное;
- мутовчатое;
- розеточное.
Источник: present5.com
Ученые связывают столь строгую закономерность расположения листьев на стебле с размерами конуса нарастания, а также особенностями взаимовлияния листовых зачатков друг на друга.
Примечание
Существует также гипотеза, согласно которой каждый зарождающийся листочек создает вокруг себя невидимое энергетическое поле. Оно способно тормозить близкое развитие прочих листовых пластин и стимулировать их развитие на определенном расстоянии.
Зрительно строгость phyllotaxis не всегда заметна. Чаще листва выглядит единым целым. Однако, если в стеблевом узле залегает более одного листа, появляется закономерная «правильность». Так, попарно расположенные (один напротив другого) листья называются противоположными (по-научному, супротивными). Разновидностью являются варианты чередования расположения пар. Такая особенность характерна, например, для клена, сирени, мяты, шалфея. Листья у них – накрест сидящие.
Стеблевой узел может содержать три листовых пластины (олеандр). Тогда тройные кольца чередуются между собой. Такая же особенность характерна для растений с четырьмя, шестью, десятью листьями в узле. Чередование ближайших кружков обязательно. Рассмотренный вариант получил название кольчатого (или кольчаторасположенного).
Интересно, что при мысленном проведении линий, соединяющих все кольчаторасположенные листья между собой, вырисовывается несколько вертикалей, для которых введен специальный термин – ортостихи.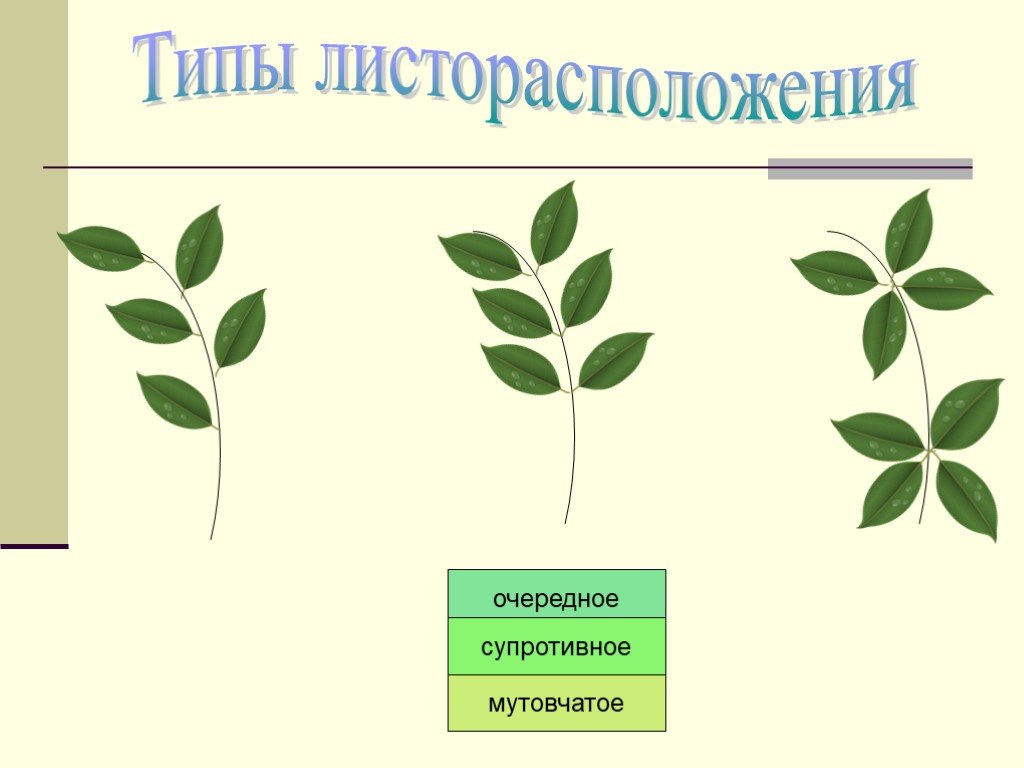
Бывают варианты, что листья просто разбросаны, т.е. в листовом узле он содержится лишь один. Для таких случаев также существует закономерность: при проведении от любого, например, расположенного снизу, листа поочередно соединяющей мысленной линии, она примет винтовой характер. Проекция такой линии на плоскость будет выглядеть спиралью. Эта закономерность обосновывает термин – спиральнорасположенные.
При следовании по спирали снизу вверх неизменно достигается виток, расположенный над первым. Для одних растений, например, липы, такой лист третий по счету (над 2-м находится 4-тый и т.д.). Для других, например, ольхи, четко над первым листом находится четвертый, над вторым – пятый и т.д.
Вертикальные линии, мысленно соединяющие такие прикрывающие друг друга листья, по своей численности будут четко повторять число листьев, которые располагаются между парами перекрывающих.
Между ортостихами можно замерить также горизонтальное расстояние. Для каждого растения оно индивидуально. Длина соединяющего отрезка будет равна кусочку спирали, пролегающему между двумя прикрывающими листьями. Такой отрезок имеет свое название – полный цикл.
Длина соединяющего отрезка будет равна кусочку спирали, пролегающему между двумя прикрывающими листьями. Такой отрезок имеет свое название – полный цикл.
Следовательно, любой вариант листорасположения определяется числом рядов размещения листьев (ортостих).
Морфология и структура
Исходя из возложенных на лист функций, объясняются особенности его строения. Так, клетки эпидермиса предохраняют от излишнего испарения и являются механической защитой целостности паренхимы. Мезофил (паренхима) – ткань, содержащая хлорофилл. Она – место протекания фотосинтеза.
Пронизывающие листовую пластину жилки необходимы для равномерного распределения воды, питательных веществ, механических элементов во внутренней листовой среде. Имеются также устьица – особые группы клеток с нижней стороны пластины, через которые уходит излишняя влага и осуществляется газообмен.
Такие характеристики являются универсальными для любого листика, большого либо малого, фигурного либо просто обтекаемой формы.
Морфология растений лежит в основе их классификации и систематики. Для первых растений на Земле была необходима водная среда. Состояли они из одной клетки и часто имели слабо ветвящиеся ленты. В современной флоре с ними сходы водоросли.
По мере переселения на сушу, растения претерпевали эволюционные изменения и начинали расчленяться на органы. Каждый из них выполнял свою функцию. Так сформировался стебель, функцией которого было вынести листья к свету. Питание растения осуществлялось с помощью корня, который, наравне со стеблем и листьями, был обязательным элементом зародыша.
Приспособленческие процессы привели к метаморфозам, отразившимся как на цветении, так и на листорасположении. Чем более развит и приспособлен к борьбе за существование был стебель, тем больше листьев на нем появлялось. Для стебля свойственно экзогенное ветвление и наличие побегов первого, второго и т.д. порядка.
В процессе своего роста стебель образует побеги. В них закладываются узел, междоузлие, листовой рубец, пазуха листа и листовой след. Там, где к нему крепится лист – узел. Стебель между узлами – междоузлие. Для крепления листа к стеблю служит черешок, на месте которого после опадания листвы остается рубец.
Там, где к нему крепится лист – узел. Стебель между узлами – междоузлие. Для крепления листа к стеблю служит черешок, на месте которого после опадания листвы остается рубец.
Каждый вид растений имеет характерные форму, размер таких рубцов, а также количество следов от листьев. Например, рубец конского каштана имеет 5-7 листовых следов. Ива – три.
Между черешком и стеблем располагается листовая пазуха. Растения, относящиеся к древесным, имеют в каждой пазухе по одной почке (клен, береза, дуб). Бывает и 2 почки (орех) или три (жимолость). Другое название почки – зачаточный побег. На побегах бывают верхушечные почки, боковые, придаточные.
Покрытосеменные растения всегда имеют черешок, прилистники и листовые пластины. Это черешковые виды. Такое строение помогает выбрать оптимальное положение по отношению к свету. Черешок может отсутствовать, например, у алоэ, цикория, орхидеи, огурца.
Там, где черешок примыкает к стеблю, образуется влагалище, а угол между ним и междоузлием получил название пазухи.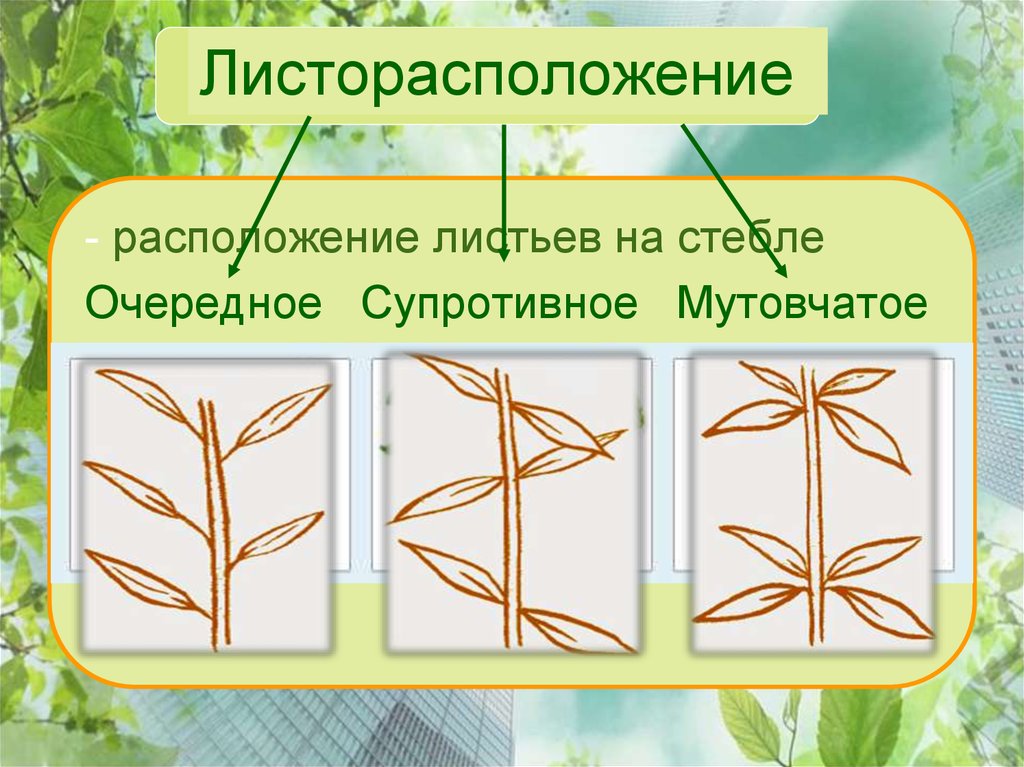 Это – место образования почки, цветка (соцветия).
Это – место образования почки, цветка (соцветия).
Принципы листорасположения, какие виды бывают
Способ расположения листьев на стебле определен для любого вида растений. Общеприняты следующие варианты:
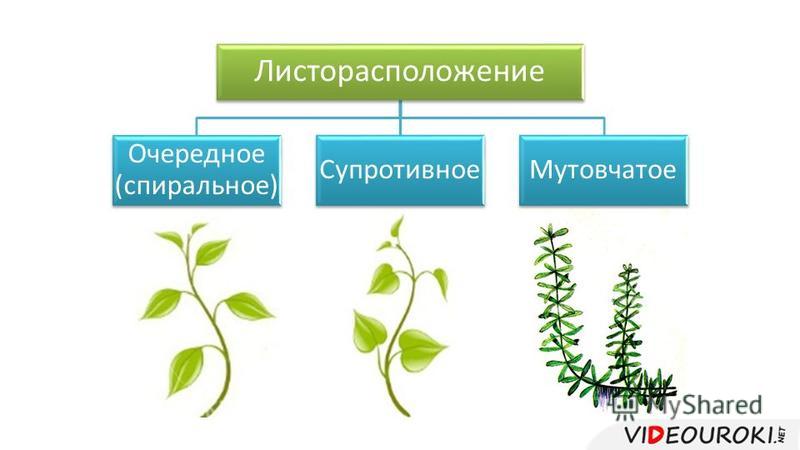
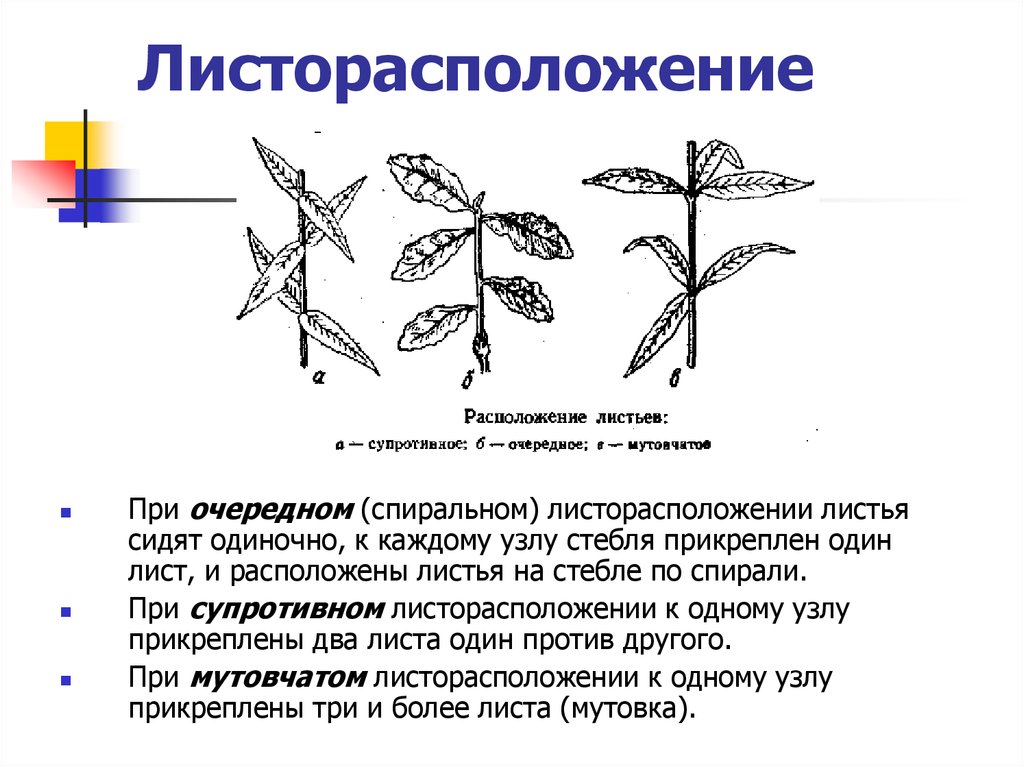
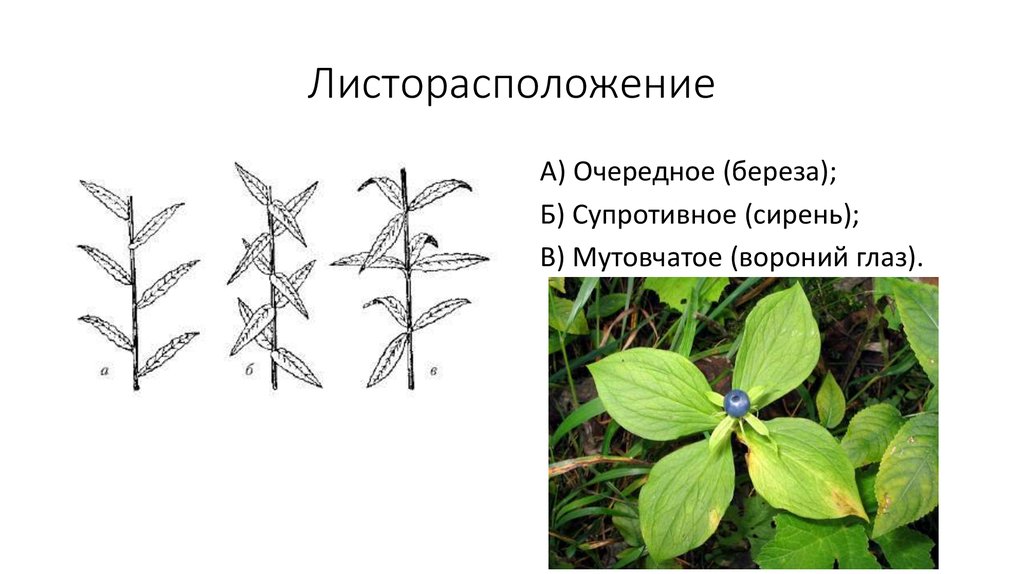
- Очередное. Характерно для растений, у которых листочки располагаются друг за другом и выходят из всех узлов на стебле. Такой тип наблюдается у пшеницы, березы, яблони, розы, пеларгонии.
- Супротивное. В этом варианте в узле располагается два листа. Возможна их ориентация перекрестно-попарно, т.е. каждый последующий лист располагается под прямым углом к предыдущему. Допускается и расположение рядов без поворотов. Пример: фуксия, жасмин, сирень.
- Мутовчатое. В узле располагаются три либо более листьев. Характерно перекрестное расположение, возможны сдвиги на пол-оборота каждого последующего по отношению к предыдущему. При этом вариант расположения напоминает мозаику, как, например, у олеандра или элодеи. В этом варианте мутовки (узлы) могут находиться на верхушке и образовывать супротивно-мутовчатые вариации.

- Розеточное листорасположение. В данном варианте присутствует укороченный стебель, поэтому листья максимально приближаются друг к другу. Междоузлия, как и черешки, практически незаметны, иногда вовсе отсутствуют. Розетки пышные, имеют общую центральную точку, как, например, у агавы, камнеломки или одуванчика.
Видоизменения листьев, примеры растений
Исходя из функций, возложенных на листья, как на орган растения, многое зависит от формы их пластины. Чем крупнее и ровнее свободная поверхность, тем эффективнее фотосинтез и испарение влаги. Поэтому у большинства представителей флоры листья гладкие, округлые, в меру тонкие и упругие.
Однако в ходе эволюции некоторые виды должны были приспособиться к индивидуальным условиям своего произрастания. Они изменились настолько, что стали вовсе не похожими на типичный лист. Причины данного явления следующие:
- В засушливой местности растение вынуждено сохранить свою влагу внутри. Поэтому ему необходимо уменьшить испарение в поверхности. При таких погодных условиях листовая пластина в процессе своей эволюции настолько сокращается, что превращается в колючку, что практически исключает испарение влаги. Кроме того, колючие видоизмененные органы исполняют дополнительную функцию – защитную, например, от съедения животными. Типичный пример – виды кактусов. При этом в кактусе не исключается фотосинтез. Просто он протекает в клетках стебля, которые располагаются практически на поверхности.
- Встречается и частичное превращение листьев в колючки, например, у барбариса, шиповника, розы и др.
- Другой способ приспособления растения к засушливой местности заключается в превращении листьев в надежные запасники влаги. Такие толстые и сочные органы появились у алоэ, агавы и других. Запасы влаги надежно защищены снаружи восковым налетом на листьях. Здесь гораздо меньше устьиц, чем у обычного листа.
- Видоизменение листа в усик – еще один вариант приспособленческой реакции растения на среду обитания. Усики необходимы тем растениям, которые для своего роста вынуждены цепляться за опоры. С помощью этого их стебель поднимается вверх, удерживаясь в вертикальном положении весь период роста. Пример: горох, чина, многие представители семейства бобовых.
- Полезные функции исполняются и такими видоизмененными листьями, как чешуйки. В них – необходимый запас питательных веществ. У отдельных растений чешуйки прячутся внутри почки, обеспечивая их защиту. Типичным примером растения с чешуйками вместо листьев является репчатый лук.
- Видоизмененная форма листьев помогает отдельным видам защититься от поедания насекомыми. В таком случае они превращаются в аппараты для ловли вредителя. Типичный пример – росянка.
 Поэтому ему необходимо уменьшить испарение в поверхности. При таких погодных условиях листовая пластина в процессе своей эволюции настолько сокращается, что превращается в колючку, что практически исключает испарение влаги. Кроме того, колючие видоизмененные органы исполняют дополнительную функцию – защитную, например, от съедения животными. Типичный пример – виды кактусов. При этом в кактусе не исключается фотосинтез. Просто он протекает в клетках стебля, которые располагаются практически на поверхности.
Поэтому ему необходимо уменьшить испарение в поверхности. При таких погодных условиях листовая пластина в процессе своей эволюции настолько сокращается, что превращается в колючку, что практически исключает испарение влаги. Кроме того, колючие видоизмененные органы исполняют дополнительную функцию – защитную, например, от съедения животными. Типичный пример – виды кактусов. При этом в кактусе не исключается фотосинтез. Просто он протекает в клетках стебля, которые располагаются практически на поверхности.
 Усики необходимы тем растениям, которые для своего роста вынуждены цепляться за опоры. С помощью этого их стебель поднимается вверх, удерживаясь в вертикальном положении весь период роста. Пример: горох, чина, многие представители семейства бобовых.
Усики необходимы тем растениям, которые для своего роста вынуждены цепляться за опоры. С помощью этого их стебель поднимается вверх, удерживаясь в вертикальном положении весь период роста. Пример: горох, чина, многие представители семейства бобовых.
У росянки верхнее покрытие листа представлено волосками, между которыми выделяется вязкая жидкость. Цветок своими лепестками привлекает насекомых. Садясь на лист, оно приклеивается лапками к жидкости, после чего лист сворачивается.
Внутри такого свертка выделяется специальный фермент, помогающий переваривать добычу.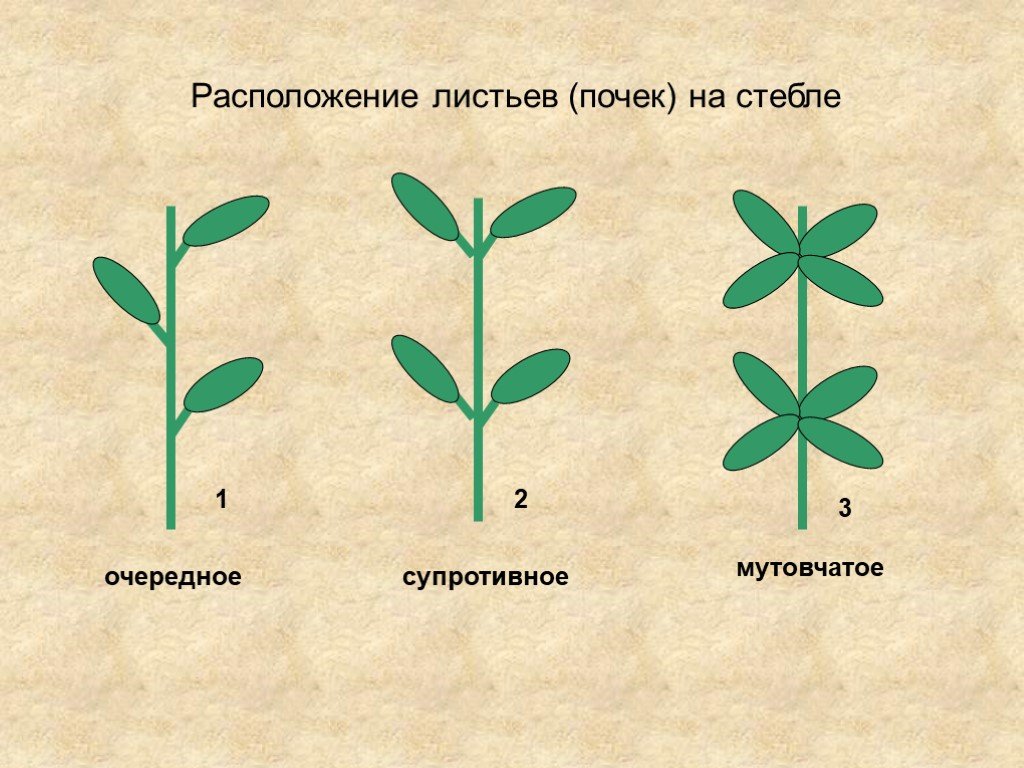 Разложение тела насекомого способствует обогащению тканей растения азотом и прочими органическими веществами. Такое видоизменение органов растения не только обеспечивает защитную функцию, но и улучшает его питание.
Разложение тела насекомого способствует обогащению тканей растения азотом и прочими органическими веществами. Такое видоизменение органов растения не только обеспечивает защитную функцию, но и улучшает его питание.
Насколько полезной была для вас статья?
Рейтинг: 1.00 (Голосов: 1)
Модель растения
Елена Клещенко
«Химия и жизнь» №10, 2014
Талант тоже строго ограничен рамками. Ощущение этих рамок навевает легкую
грусть. И в то же время как-то непроизвольно вызывает умиление. Это как
если поймешь, что бамбук — это бамбук, а дикий виноград — дикий виноград.
Акутагава Рюноскэ
Статья написана по материалам лекции доктора биологических наук В. В. Чуба (кафедра физиологии растений биофака МГУ) «Как создать виртуальное растение» на зимней научной школе «Современная биология и биотехнология будущего» в январе 2014 года.
Оставим пока в стороне вопрос о том, почему таланты, да и простые смертные, не похожи друг на друга и обладают индивидуальностью. Но почему, в самом деле, бамбук — это бамбук, а дикий виноград — дикий виноград? И почему, когда оплодотворенная зародышевая клетка в завязи начинает делиться, образуется не бесформенная клеточная масса, а корешок, стебелек и листья, а потом появляются цветок и плод с семенами, и все это именно такое, как предписали данному виду природа и Карл Линней?
Но почему, в самом деле, бамбук — это бамбук, а дикий виноград — дикий виноград? И почему, когда оплодотворенная зародышевая клетка в завязи начинает делиться, образуется не бесформенная клеточная масса, а корешок, стебелек и листья, а потом появляются цветок и плод с семенами, и все это именно такое, как предписали данному виду природа и Карл Линней?
Морфогенез, «рождение форм», — один из святых граалей биологии. Казалось бы, давно покончено с преформизмом, сторонники которого считали, что при развитии организма структуры не образуются de novo, а развиваются из миниатюрных, почти невидимых зачатков. (А в зачатках половых органов — зачатки будущих организмов, а в тех — еще, и так до субатомных структур… эта дурная бесконечность преформизм и погубила.) Все знают про ДНК, гены, программы развития. И все равно то и дело где-то на просторах Сети или в дружеском застолье раздается вопль: да не может такого быть, чтобы вся необходимая информация содержалась в ДНК, объясните, ученые, или признайте, что клеткам подсказывают высшие силы, куда расти! Ученые ищут объяснения и будут искать еще долго. Участие высших сил пока не доказано, но интересные результаты имеются. Работ, посвященных морфогенезу растений, сравнительно немного. Тема увлекательная, но, с одной стороны, без явной практической значимости, с другой — достаточно сложная, поскольку междисциплинарная: морфология и физиология растений тут встречаются с генетикой и компьютерными науками. Чтобы разобраться в программах развития, необходим программист.
Участие высших сил пока не доказано, но интересные результаты имеются. Работ, посвященных морфогенезу растений, сравнительно немного. Тема увлекательная, но, с одной стороны, без явной практической значимости, с другой — достаточно сложная, поскольку междисциплинарная: морфология и физиология растений тут встречаются с генетикой и компьютерными науками. Чтобы разобраться в программах развития, необходим программист.
Почему листья у одних растений простые и узенькие, а у других пальчато-лопастные, об этом как-нибудь в другой раз. Для начала поставим элементарный вопрос, в который, однако, уместится значительная доля всего растительного многообразия: как регулируется расположение листьев на стебле, чашелистиков, лепестков, тычинок и пестиков в цветке? У хрена и сурепки четыре лепестка, за что их и зовут крестоцветными. У розоцветных, например яблони, — пять, однако у садовых роз лепестков много, «бессчетное количество». Все перечисленные — двудольные цветковые растения, а вот у однодольных (нарцисса, лилии, тюльпана.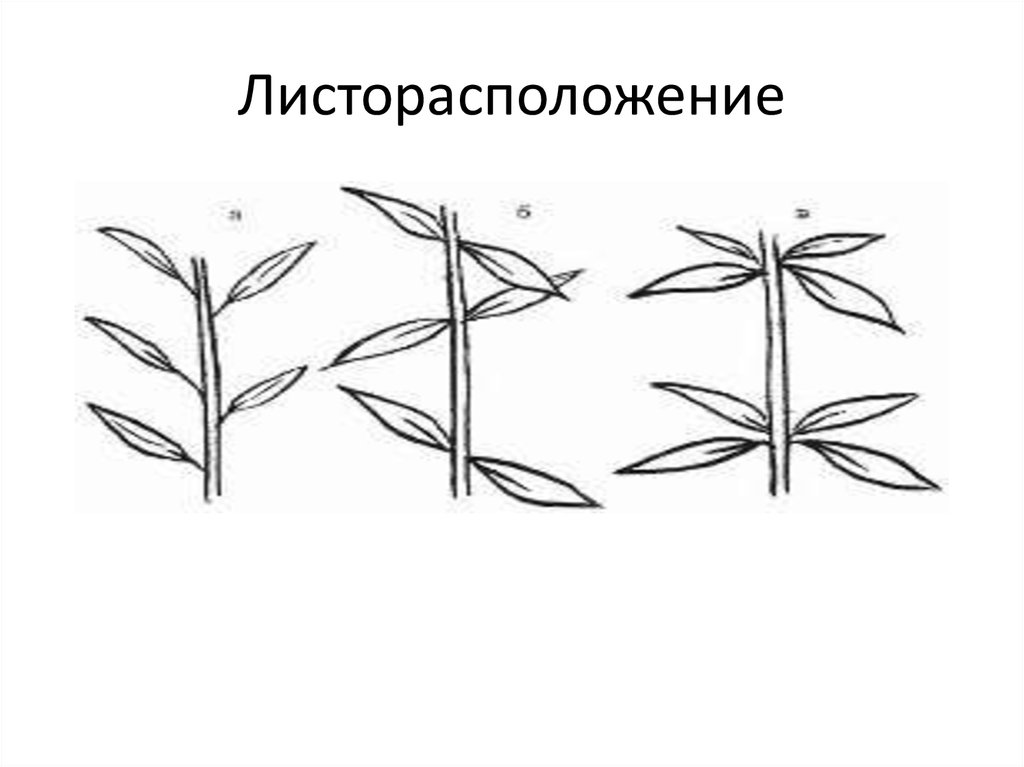 ..) шесть листочков околоцветника, похожих на лепестки, причем расположены они в два круга: три в нижнем, три в верхнем. Что касается листьев, даже те, кто не очень любили ботанику, наверное, помнят со школьных лет: очередное листорасположение, супротивное (по два листа в каждом узле), мутовчатое (больше двух, как у вороньего глаза). И как это получается? Какая сила располагает органы растения именно так, а не иначе?
..) шесть листочков околоцветника, похожих на лепестки, причем расположены они в два круга: три в нижнем, три в верхнем. Что касается листьев, даже те, кто не очень любили ботанику, наверное, помнят со школьных лет: очередное листорасположение, супротивное (по два листа в каждом узле), мутовчатое (больше двух, как у вороньего глаза). И как это получается? Какая сила располагает органы растения именно так, а не иначе?
Спирали Фибоначчи, кривые Гаусса и генные сети
Научно-популярные журналисты любят рассказывать о таинственной связи листорасположения и чисел Фибоначчи. Этот ряд чисел, каждый следующий член которого — сумма двух предыдущих (1, 1, 2, 3, 5, 8, 13, 21, 34, 55, 89, 144) обладает многими замечательными свойствами. Например, если замостить плоскость квадратами с длинами сторон, равными числам Фибоначчи, приставляя больший к меньшему, как показано на рисунке, а затем соединить их противоположные углы дугами с радиусом, равным длине стороны, получится красивая спираль. Подобные спирали мы часто видим в природе: вдоль спиральных линий выстраиваются чешуйки на шишке или цветки, образующие сердцевину ромашки. (Еще одна истина из школьного курса ботаники: ромашка относится к сложноцветным, то, что мы называем ее цветами, на самом деле соцветия из желтых простых цветков в середине и белых язычковых цветков по краям.) Интересно, что в сердцевине «цветка» можно увидеть спирали, закрученные по часовой стрелке и против часовой, при этом количества тех и других относятся друг к другу как числа Фибоначчи. Листья на побеге, ветки на стволе дерева тоже часто располагаются по спирали. Можно представить такой побег как подобие корзинки ромашки, вытянутой по вертикали, с одним или несколькими спиральными рядами листьев вместо цветков.
Подобные спирали мы часто видим в природе: вдоль спиральных линий выстраиваются чешуйки на шишке или цветки, образующие сердцевину ромашки. (Еще одна истина из школьного курса ботаники: ромашка относится к сложноцветным, то, что мы называем ее цветами, на самом деле соцветия из желтых простых цветков в середине и белых язычковых цветков по краям.) Интересно, что в сердцевине «цветка» можно увидеть спирали, закрученные по часовой стрелке и против часовой, при этом количества тех и других относятся друг к другу как числа Фибоначчи. Листья на побеге, ветки на стволе дерева тоже часто располагаются по спирали. Можно представить такой побег как подобие корзинки ромашки, вытянутой по вертикали, с одним или несколькими спиральными рядами листьев вместо цветков.
Тем не менее легко указать примеры, которые на спираль Фибоначчи не укладываются. Есть, например, такие растения, у которых листорасположение хотя и очередное, однако каждый новый нечетный лист располагается точно под предыдущим, и никакие спирали с золотыми сечениями им не указ. Да и со спиральным расположением, наблюдаемым в природе, не всегда получается гладко. К тому же едва ли в растении есть устройство, которое суммирует соседние члены ряда Фибоначчи и откладывает соответствующий угол между листьями. Мало разглядеть порядок, необходимо понять, что его формирует. Что у растения точно есть, так это морфогены — сигнальные вещества, которые распространяются в объеме ткани. Но при этом, очевидно, их концентрация изменяется плавно. Как же эта плавность преобразуется в дискретный сигнал, указывающий листу расти из строго определенной точки, а из соседних — не расти?
Да и со спиральным расположением, наблюдаемым в природе, не всегда получается гладко. К тому же едва ли в растении есть устройство, которое суммирует соседние члены ряда Фибоначчи и откладывает соответствующий угол между листьями. Мало разглядеть порядок, необходимо понять, что его формирует. Что у растения точно есть, так это морфогены — сигнальные вещества, которые распространяются в объеме ткани. Но при этом, очевидно, их концентрация изменяется плавно. Как же эта плавность преобразуется в дискретный сигнал, указывающий листу расти из строго определенной точки, а из соседних — не расти?
Один из возможных механизмов, с помощью которого один морфоген может направлять развитие клеток по различным путям, первым предложил в 70–80-е годы ХХ века Льюис Вольперт, британский специалист по биологии развития, — так называемую модель трехцветного флага. Концентрация сигнального вещества при удалении от центра, в котором оно синтезируется, убывает плавно, но клетки реагируют только тогда, когда она превышает определенные пороговые значения.
Если таких порогов два, одно и то же вещество может направить развитие клетки по трем путям, включая различные варианты генной активности. Важно, что команда, полученная клеткой, определяется ее положением относительно зоны, посылающей сигнал: например, ближние участки, где концентрация морфогена максимальна, станут красными, самые удаленные — синими, промежуточные — желтыми. Таким образом, у нас появляется понятие о позиционной информации: судьба клетки, роль, которую она сыграет в развитии, определяется ее положением.
Экспериментальные подтверждения для этой гипотезы получил сам Вольперт, правда, не на растительной, а на животной модели — изучая развитие конечности цыпленка. Рядом с группой клеток, из которой будет развиваться крыло, была найдена так называемая зона поляризующей активности, которая посылает химический сигнал, определяющий номера пальцев. Эксперименты по пересадке этой зоны подтвердили и сам эффект, и его зависимость от концентрации. Куда бы ни пересаживали этот маленький участок, там вырастали и пальцы с самыми большими номерами.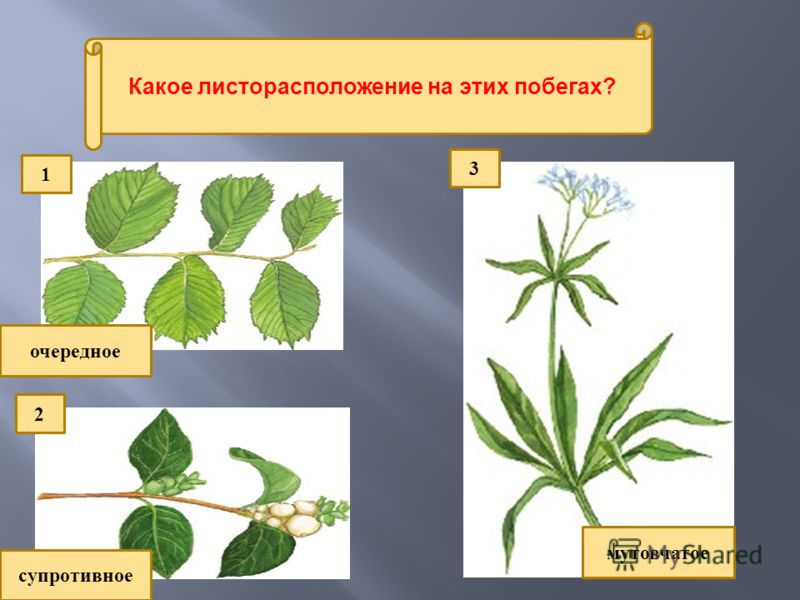
Кроме позитивных сигналов могут быть и негативные — останавливающие (ингибирующие) развитие, причем их может рассылать та же самая группа клеток. Вещество-активатор запускает развитие органа, выделение ингибитора его останавливает. При моделировании можно так подобрать скорость синтеза и распространения сигнальных веществ, что концентрация активатора превысит концентрацию ингибитора лишь на небольшом участке. Для чего это нужно, понятно: чтобы органы в своем развитии и дальнейшем существовании не мешали друг другу. Именно по этой схеме формируется волосок на листе растения: активатор запускает его рост, потом ингибитор движется в соседние клетки и запрещает им отращивать волоски. Для некоторых растений уже известны продукты генов, участвующих в этом процессе.
Самые важные активаторы морфогенеза у растений — фитогормоны ауксины, а наиболее распространенный ауксин — индолилуксусная кислота (ИУК). В тех группах клеток, где концентрация ауксина выше, идут клеточные деления, рост растяжением. Потоки ауксина могут направлять сами клетки: ауксин проникает внутрь клетки пассивно, а выкачивают его наружу белки-насосы PIN. Эти потоки размечают будущие формы растения: там, куда они стремятся, развиваются зачатки листьев или органов цветка. На какой стороне клетки появляются PIN (а растительная клетка, как большинство биообъектов, очень изменчива), туда и направляется ауксин.
Потоки ауксина могут направлять сами клетки: ауксин проникает внутрь клетки пассивно, а выкачивают его наружу белки-насосы PIN. Эти потоки размечают будущие формы растения: там, куда они стремятся, развиваются зачатки листьев или органов цветка. На какой стороне клетки появляются PIN (а растительная клетка, как большинство биообъектов, очень изменчива), туда и направляется ауксин.
Чтобы лучше понять, как работает тот или иной механизм, ученые смотрят, что происходит, когда он сломан. У любимого модельного растения генетиков Arabidopsis thaliana (по-русски резуховидка Таля, но с легкой руки «лабораторных» биологов его и по-русски все чаще называют арабидопсисом) есть мутация pin1-1. Такие мутанты растут только вверх, давая длинные побеги странного вида, без цветов и листьев.
У них поврежден тот самый белок PIN1, который должен был активно выкачивать ИУК из клеток, направляя его в нужные стороны. Собственно, по этой мутации, от английского pin-formed — «похожий на булавку», белок и получил название. Но если нанести на кончик побега немного пасты, содержащей ИУК, в этой точке начнет формироваться лист или цветок.
Но если нанести на кончик побега немного пасты, содержащей ИУК, в этой точке начнет формироваться лист или цветок.
Логично спросить, почему и как самоорганизуются потоки ауксина. Так, была предложена компьютерная программа (Runions A. et al., ACM Trans. Graph., 2006, 24, 702–711), которая позволяет моделировать жилкование на листьях — хоть сетчатое, хоть параллельное. Результат зависит от начального размещения точек синтеза ауксина, которые распределяются по листу равномерно.
Дальше развивающиеся жилки пытаются использовать ауксин из ближайшей точки, а те, кто «не успел», поворачивают в другую сторону. Таких простых начальных условий оказалось достаточно для получения сложных рисунков жилкования, очень близких к реальности.
Эти программы сразу оценили создатели компьютерных игр — они не могут, подобно герою рассказа Толкиена, рисовать всю жизнь один лист, им надо прямо сейчас запустить персонажа в лес, шелестящий сотнями индивидуальных листьев. Но в настоящем растении, конечно, все происходит не так. Ауксин не может быть рассыпан отдельными крупинками, его источники располагаются совершенно иначе.
Ауксин не может быть рассыпан отдельными крупинками, его источники располагаются совершенно иначе.
Известно, что ауксин движется по растению в целом сверху вниз: от апикальной меристемы (верхушечного конуса нарастания; меристемами вообще называются недифференцированные, активно делящиеся ткани растения) к корням. Известно также, что потоки ауксина имеют тенденцию притягиваться друг к другу и сливаться: если клетка «чувствует» неподалеку мощный поток ауксина, она перенаправляет в ту же сторону и свой. (Садоводам и огородникам хорошо знаком эффект апикального доминирования: верхушечная точка роста подавляет боковые, и, если верхушку побега не отщипнуть, растение будет расти ввысь и слабо ветвиться.) Помимо деления ауксин стимулирует рост клеток, вытягивание их в длину с размягчением клеточной стенки. Растения — удивительные существа: они растут, и порой очень быстро, хотя состоят из жестких структурных единиц, их клетки покрыты целлюлозной оболочкой и достаточно прочно связаны с соседями.
Что касается закладки листьев и лепестков, самое интересное происходит в зоне апикальной меристемы. Именно там задается взаимное расположение листьев — по мере вытягивания стебля увеличится только расстояние между узлами. Анатомия этой зоны у цветковых растений неплохо изучена. Верхушку покрывают несколько слоев клеток (обычно три), которые называют туникой, под ними — не так строго ориентированные клетки корпуса. В тунике выделяют также центральную и периферическую зоны меристемы. Их клетки различаются морфологически, кроме того, опыты с введением в клетки красителей показали, что межклеточные контакты открыты в пределах центральной зоны и периферической зоны, а между этими зонами — закрыты, то есть они не обмениваются сигнальными веществами. Под центральной зоной находится организационный центр, в котором клетки делятся слабо, поскольку основная его функция — управление ростом меристемы. Если какие-то клетки меристемы повреждены, организационный центр может пополнить их запас; поэтому раньше его называли «меристемой ожидания».
Клетки меристемы постоянно ведут «переговоры», пересылая друг другу сигнальные белки, короткие пептиды и РНК. В этом процессе множество участников, каждый со своей важной ролью, и подробное их описание мы пропустим, тем более что у разных видов картина различается. Генные сети чем-то похожи на сериалы-эпопеи вроде некогда знаменитой «Санта-Барбары» или «Игр престолов» по Джорджу Мартину. Кажется, что невозможно выучить всех этих малоразличимых героев и злодеев, их врагов и родичей, но проходит первый сезон, и мы уже спорим, кто из них плохой, а кто хороший, кто кому приходится незаконным сыном и кто что замышляет. В сложных взаимоотношениях сигнальных веществ меристемы тоже можно разобраться, но это требует времени.
В качестве «трейлера к сериалу» можно сказать, например, что в организационном центре у арабидопсиса экспрессируется ген WUSCHEL (WUS), активирующий ген CLAVATA 3 (CLV 3) в центральной зоне, а его продукты, в свою очередь, сложным путем подавляют экспрессию WUS (отрицательная обратная связь). Продукт гена ARGONAUTE 10 — он же PINHEAD (PNH), он же ZWILLE (ZLL) — активирует работу гена WUS, который поддерживает клетки в недифференцированном состоянии, и он же участвует в определении «верхней» и «нижней» сторон листа. Гены WUS и PNH/ZLL, возможно, также контролируют потоки ауксина и создают позиционные сигналы для будущих листьев, отмечая места их закладки. Клетки в этой области начинают делиться в плоскости, параллельной поверхности, — начинается рост наружу, появляется выступ, который затем становится листом.
Продукт гена ARGONAUTE 10 — он же PINHEAD (PNH), он же ZWILLE (ZLL) — активирует работу гена WUS, который поддерживает клетки в недифференцированном состоянии, и он же участвует в определении «верхней» и «нижней» сторон листа. Гены WUS и PNH/ZLL, возможно, также контролируют потоки ауксина и создают позиционные сигналы для будущих листьев, отмечая места их закладки. Клетки в этой области начинают делиться в плоскости, параллельной поверхности, — начинается рост наружу, появляется выступ, который затем становится листом.
Зачатки органов растений ботаники называют «примордиями» — это слово ужасно смешит неспециалистов, но мы его будем иногда использовать. В примордиях листьев также активен ген ANT, а гены CUC 1, CUC 2, UFO задают границу примордия.
Теперь перейдем к математическим моделям, которые покажутся нам простыми после нередуцируемой биологической сложности. Как сказал один физик, изучавший теплообмен живых объектов: «Сначала мы считали лосей шарами, но теперь рассматриваем их как цилиндры» (А.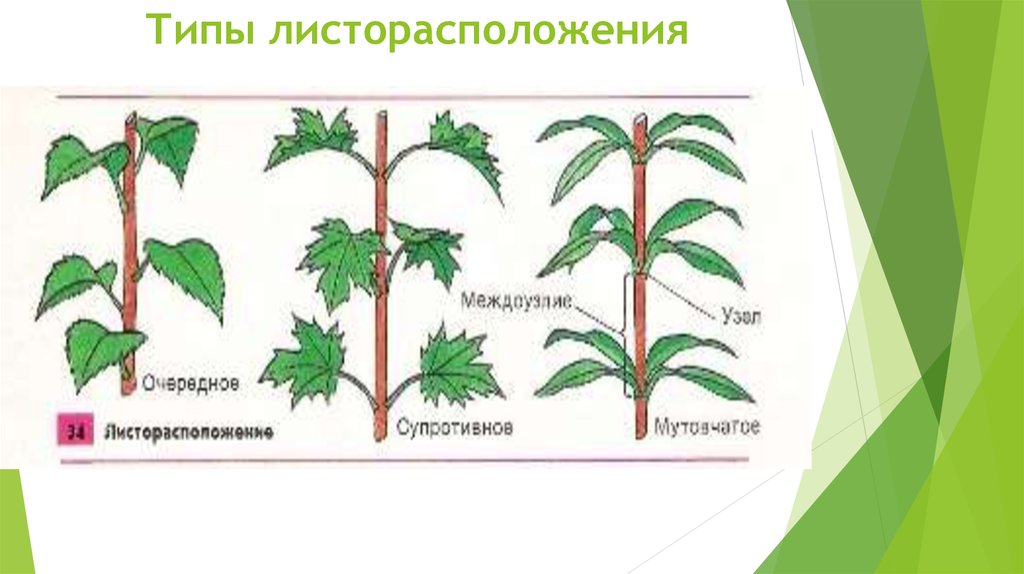 Н. Горбань, Р. Г. Хлебопрос. Демон Дарвина. Идея оптимальности и естественный отбор. Москва: Наука, 1988). Биологи над физиком смеялись, и напрасно: если считать лосей лосями, расчет может затянуться надолго, и еще неизвестно, насколько улучшится результат.
Н. Горбань, Р. Г. Хлебопрос. Демон Дарвина. Идея оптимальности и естественный отбор. Москва: Наука, 1988). Биологи над физиком смеялись, и напрасно: если считать лосей лосями, расчет может затянуться надолго, и еще неизвестно, насколько улучшится результат.
План побега
Вспомним еще раз, что нам известно о закладке листьев. В апикальной меристеме (верхушечном конусе нарастания) есть участки, синтезирующие сигнальные вещества, и, что не менее важно, есть участки, к этим веществам восприимчивые. Бывают сигналы, активирующие рост листа в данной области, и другие, отменяющие действие активаторов (ингибиторы), в зоне действия которых лист не сможет вырасти. Очевидно, что зона активации роста должна быть окружена зоной ингибирования, чтобы листья не наезжали друг на друга: лист, уже начавший расти, сообщает соседним клеткам, что место занято, размечено под будущий лист. Логично предположить, что второй лист вырастет настолько близко к первому, насколько позволит зона ингибирования: природа не любит пустоты. Примордии листьев рассылают сигналы, клетки реагируют на них, включая генные сети, и таким образом определяется, что и где вырастет, — осуществляется разметка будущих органов.
Примордии листьев рассылают сигналы, клетки реагируют на них, включая генные сети, и таким образом определяется, что и где вырастет, — осуществляется разметка будущих органов.
Предпосылки будто бы простые, но построенная на их основе модель оказалась удивительно эффективной. О математической модели, которая с помощью соответствующего программного обеспечения генерирует побеги и цветки in silico, рассказал на зимней школе в Звенигороде доктор биологических наук Владимир Викторович Чуб (кафедра физиологии растений биологического факультета МГУ). В работе участвовали также кафедра генетики, кафедра математической теории интеллектуальных систем мехмата МГУ и Центр биоинженерии РАН. Кафедра высших растений биофака МГУ помогала консультациями, подбором объектов для моделирования и ценными замечаниями.
Пусть растущий побег — это цилиндр с полусферой на верхнем основании, которая изображает меристему. Какие параметры здесь важны? Во-первых, размеры меристемы — ее радиус R. Во-вторых, размеры зоны, восприимчивой (компетентной) к воздействию сигналов, — зоны, где будут размечаться листья. Как мы видели на предыдущих рисунках, это некое кольцо вокруг верхушки побега, которое можно изобразить окружностями с радиусами Rmin и Rmax. Между этими окружностями возникают зачатки органов — примордии. Точка Р — медиана примордия. Нужно также обозначить размеры минимального пространства, необходимого для разметки органа (d0), радиус сферической зоны ингибирования D, в пределах которой примордии не возникают, и скорость роста побега dH.
Как мы видели на предыдущих рисунках, это некое кольцо вокруг верхушки побега, которое можно изобразить окружностями с радиусами Rmin и Rmax. Между этими окружностями возникают зачатки органов — примордии. Точка Р — медиана примордия. Нужно также обозначить размеры минимального пространства, необходимого для разметки органа (d0), радиус сферической зоны ингибирования D, в пределах которой примордии не возникают, и скорость роста побега dH.
Теперь запустим программу, заставим побег расти и формировать листья. Выясняется изумительная вещь: тип листорасположения зависит от соотношения всего двух параметров — радиуса меристемы R и радиуса ингибирующей зоны D, причем разметка примордиев в компетентной зоне полностью определяет облик побега, который формируется в дальнейшем. Меняя эти параметры, можно получить известные в природе типы листорасположения, а также переходные формы между ними.
Возьмем случай, когда D существенно больше R — более чем в два раза. В этом случае модель предсказывает двурядный филлотаксис, или строгую дистихию, — в каждом узле находится один лист, при этом каждый образует с предыдущим угол в 180°, так что ветвь с листьями выглядит плоской. Почему так выходит, легко догадаться. Первая же зона ингибирования накрывает всю меристему, поэтому второй лист на этом уровне возникнуть не может. А возникнет он, естественно, на противоположной стороне: как только зона компетентности, поднимаясь по мере роста, высвободит из зоны ингибирования окошко не меньше d0. Такое листорасположение наблюдается у комнатного растения дихоризандры королевской Dichorisandra reginae, как, впрочем, и у многих других традесканциевых.
Почему так выходит, легко догадаться. Первая же зона ингибирования накрывает всю меристему, поэтому второй лист на этом уровне возникнуть не может. А возникнет он, естественно, на противоположной стороне: как только зона компетентности, поднимаясь по мере роста, высвободит из зоны ингибирования окошко не меньше d0. Такое листорасположение наблюдается у комнатного растения дихоризандры королевской Dichorisandra reginae, как, впрочем, и у многих других традесканциевых.
Если радиус ингибирующей зоны D будет чуть меньше, чем 2R, на смену строгой дистихии приходит спиродистихия — прямые линии листорасположения перекручиваются, листья располагаются по спиралям. Оказалось, что такая модель — две спирали, наложенные друг на друга, — лучше всего описывает реально наблюдаемое листорасположение. Традиционное представление об одной спирали, подчиняющейся дробям Фибоначчи, здесь явно не подходит. Филлотаксис, предсказанный этой моделью, можно наблюдать у видов рода Сordyline и других объектов.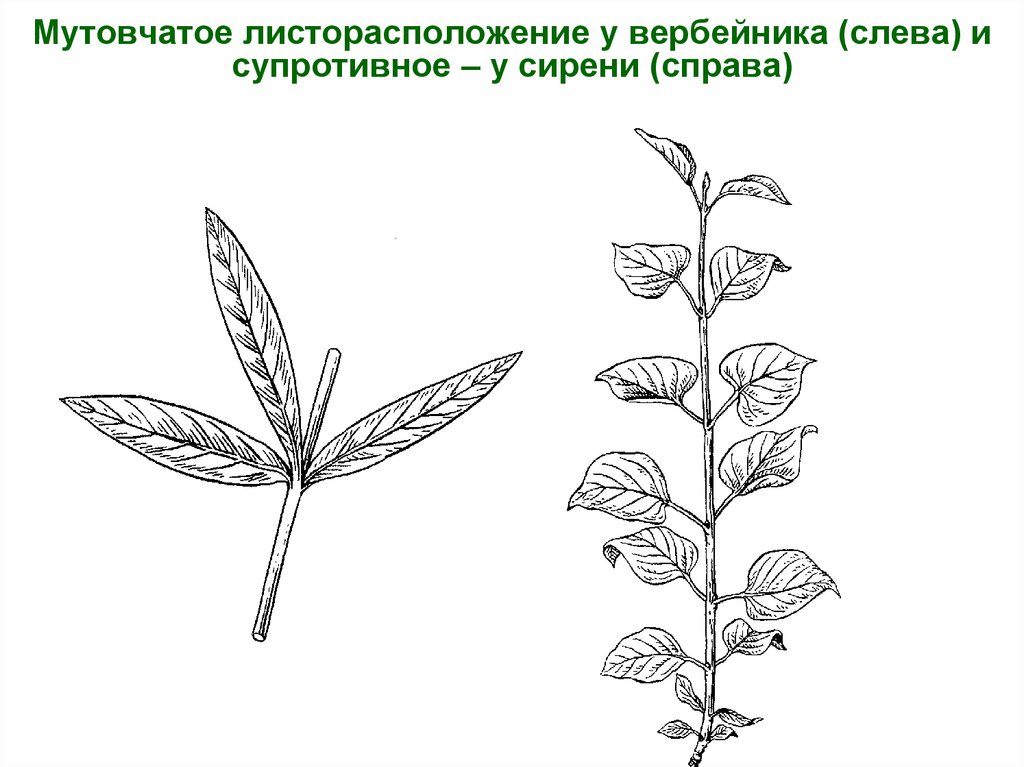
При дальнейшем уменьшении ингибирующей зоны она перестанет перекрывать всю меристему на своем уровне, и два листа смогут возникнуть друг напротив друга. В школьных учебниках это называется супротивным листорасположением, оно наблюдается, например, у сирени. Почему каждая новая пара листьев повернута относительно предыдущей на 90°, догадайтесь сами. Если ингибирующие зоны станут еще меньше, то на одном уровне смогут возникнуть сразу три (при D = 1,2R) или четыре листа (при D = R) — реализуется мутовчатое листорасположение.
А что хорошо для мутовок листьев, то хорошо и для лепестков с чашелистиками: очевидно, эти органы могут размечаться точно так же. Этот подход позволил авторам обратить внимание на странные цветки с нетипичным для данного вида расположением лепестков — следствием мутации или сбоя программы на физиологическом уровне. Все мы знаем «счастливые» пятилепестковые цветки сирени, которые встречаются среди четырехлепестковых не то чтобы часто, но и не так редко, чтобы надоело искать.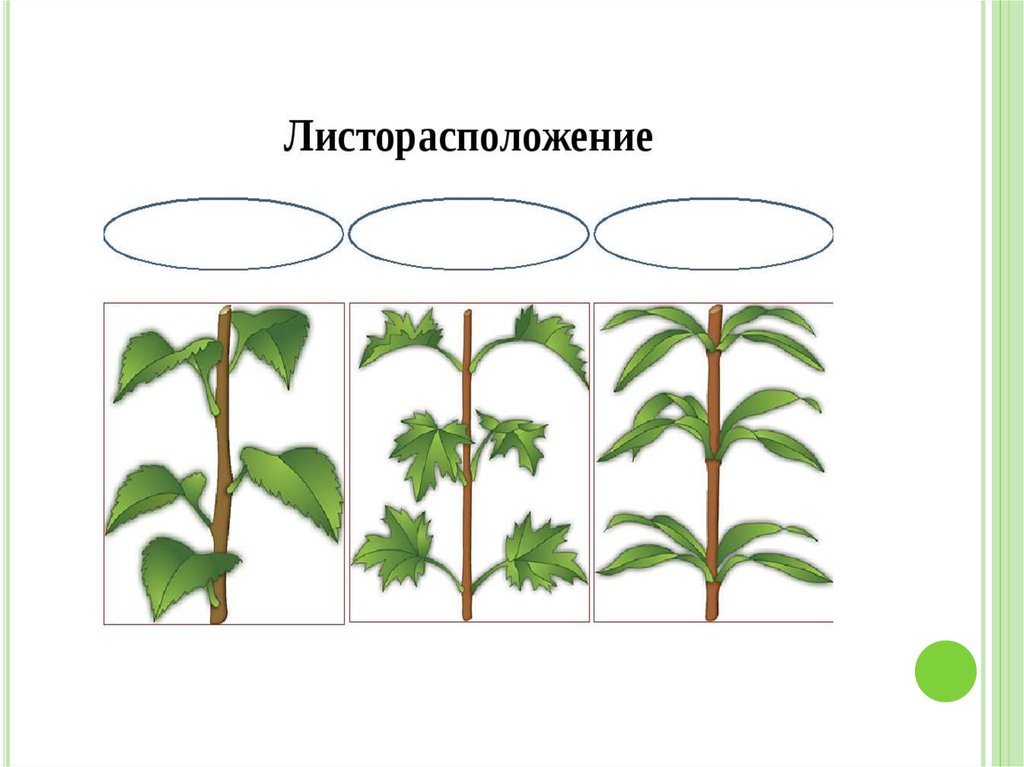 Изредка попадаются нарциссы с пятью «лепестками» вместо шести (как мы уже упоминали, у нарцисса это на самом деле не лепестки, а листочки околоцветника). Такие цветки иногда бывают некрасивыми, кривоватыми.
Изредка попадаются нарциссы с пятью «лепестками» вместо шести (как мы уже упоминали, у нарцисса это на самом деле не лепестки, а листочки околоцветника). Такие цветки иногда бывают некрасивыми, кривоватыми.
Математическая модель предлагает хорошее объяснение: промежуточные величины диаметров зоны ингибирования, из-за чего в одном круге размещаются, например, не пять лепестков, а всего четыре. Или «четыре с половиной»: углы между четырьмя слишком велики, а пятому недостаточно свободного места. Цветок нарцисса обладает трехлучевой симметрией (не шестилучевой, потому что его шесть листочков околоцветника расположены не в одном круге, а двумя кругами по три). Поэтому пятилисточковый нарцисс логично было назвать «2,5-мерным». По словам Владимира Чуба, ботаников сначала возмутил этот термин, однако в конце концов они признали его право на существование.
Изучение аномальных цветков интересно еще и потому, что оно дает ключ к некоторым спорным моментам эволюции цветка. Так, математическое моделирование показывает, что пятимерные цветки едва ли могли возникнуть из шестилепестковых тримерных через потерю одного лепестка и сведение оставшихся в один круг. Скорее всего, трехлепестковый тримерный цветок сначала стал тетрамерным, с четырьмя осями симметрии, а потом уже пентамерным.
Скорее всего, трехлепестковый тримерный цветок сначала стал тетрамерным, с четырьмя осями симметрии, а потом уже пентамерным.
На следующем рисунке представлены переходы от одного вида симметрии к другому в пределах рода. Традесканцию многие выращивают дома или в офисе, но не каждый видел ее цветок; в норме он имеет три лепестка. Хионодокса из семейства гиацинтовых полагается шесть листочков околоцветника в двух кругах, как и лилейнику.
Когда в модели уменьшали радиус ингибирующей зоны D от 1,2 до 1,0 (если принять R за единицу), тримерный цветок через ряд последовательных нетипичных» форм превращался в тетрамерный (восьмилепестковый или восьмилисточковый). Любопытным оказалось строение цветка гортензий. В их стерильных цветках задачу привлечения опылителей выполняют чашелистики, а не лепестки. Один из лепестков, которые в норме мельче и закладываются выше, становился в круг чашелистиков. Полный ряд форм удалось отыскать в природе у гортензии древовидной Hydrangea cinerea.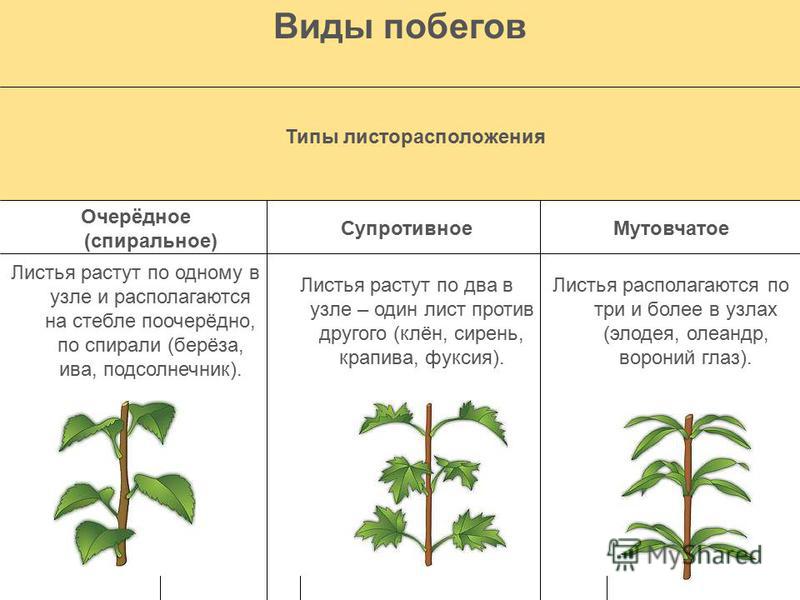
Можно не только увеличивать число лепестков в круге, но и увеличивать число самих кругов — «тянуть за макушку» цветок, заставляя его расти и надстраивать новые ярусы органов; на языке модели — вводя скорость роста dH, не равную нулю. Так можно получить некоторые махровые цветки. В предельном случае цветок растет вверх до исчерпания клеточного материала, который весь превращается в лепестки; у такого цветка не будет генеративных органов — пестиков и тычинок.
О пестиках и тычинках
Цветок — это все же не побег с листьями, и рост вверх у него часто бывает ограничен генетическими факторами. Поэтому, чтобы показать, каким образом апикальная меристема приобретает структуру цветка, в модель необходимо внести коррективы. Скорость роста вверх, то есть dH, принимаем равной нулю, а возможность расширяться обозначим как dR.
Рост вверх в какой-то момент остановится, значит, разметить нужно всю меристему. Поэтому была введена возможность смещения верхней границы разметки органов вверх (dRmin) и верхушечная зона разметки с радиусом R3 и возможностью расширения — dR3.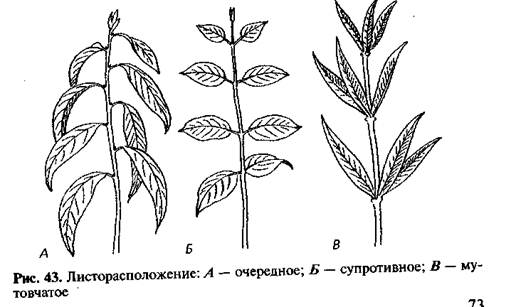 Круговую зону авторы модели назвали базипетальной, кольцевую — акропетальной (базипетальный у ботаников означает «направленный от верхушки к основанию стебля», акропетальный — «направленный к верхушке»).
Круговую зону авторы модели назвали базипетальной, кольцевую — акропетальной (базипетальный у ботаников означает «направленный от верхушки к основанию стебля», акропетальный — «направленный к верхушке»).
Зачем понадобилась базипетальная зона? Ведь известно, что органы цветка закладываются снизу вверх: чашелистики, лепестки, тычинки, пестик или пестики. Однако не исключено, что разметка органов, то есть сигналы «здесь расти, а здесь не расти», выстраивается заранее, не в той же самой последовательности, в которой органы начинают формироваться. А проверить это важно: как мы убедились, разметка первых органов определяет расположение всех последующих.
Конечно, всем интересно, как это происходит у арабидопсиса: его генетика хорошо изучена и было бы заманчиво привязать разметку и закладку органов цветка к активности генных сетей. «Классический», описанный в учебнике цветок арабидопсиса имеет формулу, достаточно типичную для крестоцветных: Ч4 Л4 Т4+2 П2.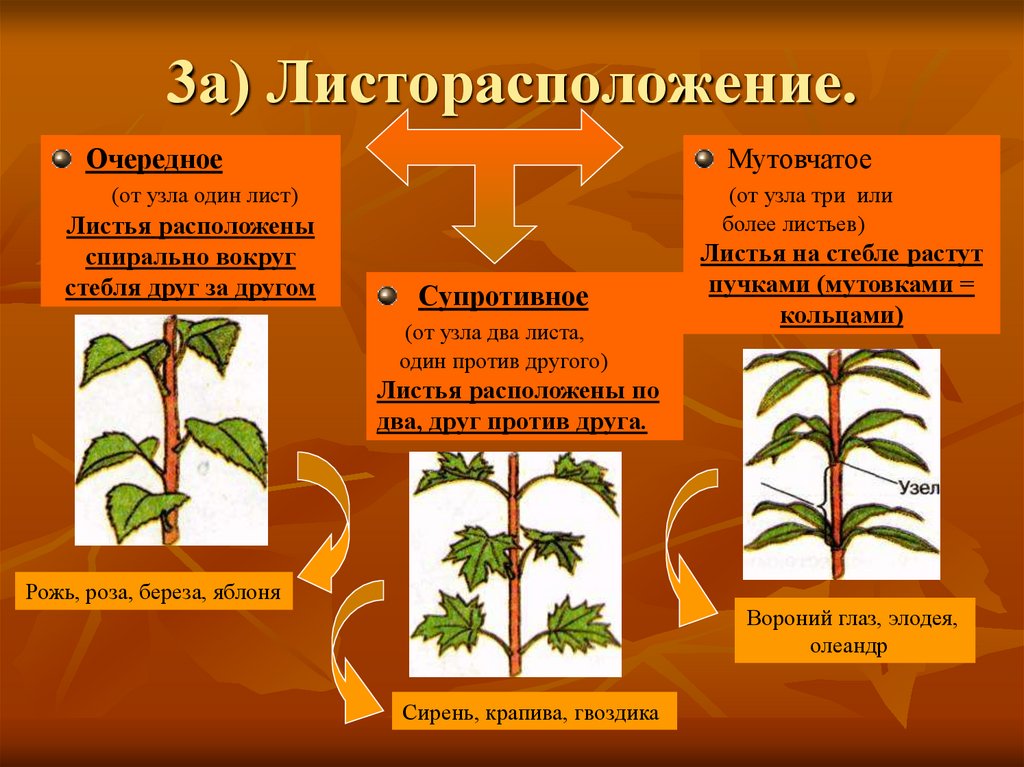 Буквы обозначают соответственно чашелистики, лепестки, тычинки и пестики, а «4+2» — что четыре тычинки у него длинные, а две короткие. Но попадаются цветки арабидопсиса без одной короткой тычинки (Ч4 Л4 Т4+1 П2), реже без обеих коротких тычинок (Ч4 Л4 Т4 П2) или даже с одной дополнительной длинной и без коротких (Ч4 Л4 Т5 П2). Среднее число тычинок в цветках арабидопсиса дикого типа всегда меньше шести!
Буквы обозначают соответственно чашелистики, лепестки, тычинки и пестики, а «4+2» — что четыре тычинки у него длинные, а две короткие. Но попадаются цветки арабидопсиса без одной короткой тычинки (Ч4 Л4 Т4+1 П2), реже без обеих коротких тычинок (Ч4 Л4 Т4 П2) или даже с одной дополнительной длинной и без коротких (Ч4 Л4 Т5 П2). Среднее число тычинок в цветках арабидопсиса дикого типа всегда меньше шести!
В семействе крестоцветных вариации числа органов — не редкость. В целом можно сказать, что у них наиболее стабильны регионы гинецея (из него вырастет пестик) и чашелистиков, а самый вариабельный — регион коротких тычинок. Можно предположить, что стабильные участки — одинаковые в большинстве цветков — размечаются первыми, а вариабельные — последними, «по остаточному принципу», отчего, они и оказываются вариабельными. Тогда введение базипетальной зоны, расширяющейся навстречу акропетальной, выглядит вполне логичным. А между ними попадают тычинки, размечаемые в последнюю очередь. Участки меристемы, не занятые лепестками и пестиками, размечаются, «как получится», подобно остаткам теста для пирожков: когда выйдет шесть штук, когда пять или четыре… Такая вариабельность для цветка не катастрофична: лучше уж пусть не хватает какой-то тычинки, чем пестика.
А между ними попадают тычинки, размечаемые в последнюю очередь. Участки меристемы, не занятые лепестками и пестиками, размечаются, «как получится», подобно остаткам теста для пирожков: когда выйдет шесть штук, когда пять или четыре… Такая вариабельность для цветка не катастрофична: лучше уж пусть не хватает какой-то тычинки, чем пестика.
Действительно, математическая модель с такими исходными данными позволяла создавать как типичные, так и «аномальные» цветки арабидопсиса. Сначала размечалась одна пара чашелистиков, а сверху плодолистики (будущий пестик), затем вторые два чашелистика, четыре лепестка, длинные тычинки и, наконец, короткие. Короткие тычинки, размечаемые последними, утрачивались чаще всего.
Модель проверили на цветках обыкновенного ревеня Rheum из семейства гречишных. Ревень в данном случае хорош обилием материала — у него огромное множество мелких цветочков. На этот раз исследователи не придумывали модель, наиболее похожую на реальный цветок (что всегда немного напоминает подгонку), а пошли от противного: построили все возможные модели разметки меристемы, чтобы проверить, какая из них ближе к реальности.
Моделей получилось четыре. Первая, обозначенная буквой А (от «акропетальная»), предполагала разметку органов снизу вверх, в порядке наблюдаемого их формирования, от чашелистиков к плодолистикам. Модель Б2 соответствовала данным об активности генов меристемы: в ней чашелистики и лепестки размечались снизу, а плодолистики и тычинки — сверху. Модели Б1 (тычинки размечаются вместе с околоцветником) и Б3 (часть тычинок размечается сверху, часть снизу) представляли остальные теоретически возможные варианты. Компьютерные модели цветков предъявляли на оценку экспертам-ботаникам, чтобы те оценили их реалистичность.
Все модели, кроме Б1, дали хорошие решения для «правильного» цветка ревеня. Но лучшей оказалась модель Б3, где тычинки размечались последними, и сверху, и снизу. Прелесть ее была не только в том, что цветок получался каким надо при самом широком варьировании параметров, — даже когда появлялись отклонения, они соответствовали «неправильным» цветам, наблюдаемым в природе. Другие же модели даже при меньших отклонениях очень сильно меняли положения тычинок.
Проверить экспериментально результаты, полученные на виртуальных растениях (например, обнаружить включение синтеза и распространение неких сигнальных веществ именно в такой геометрии), пока не удалось. Процессы, связанные с разметкой органов, протекают стремительно, «остановить мгновение» непросто. Но ценно уже то, что модель подсказывает, что искать, в каком направлении двигаться дальше. «С этой моделью играли ботаники, и после этого они смотрели на цветок совершенно другим взглядом, — рассказывает Владимир Чуб. — Видят в нем зоны разметки, точки, из которых вырастут органы. Пытаются это разглядеть на сканирующих фотографиях, начинают это обсуждать. Что еще может сделать эта модель? Иногда попадается цветок уникальной архитектуры, и второго такого нет на всей плантации. Хочется понять, как он развивался, и эта модель позволяет «отмотать назад» процесс его развития, показать, как бы это могло быть. Моделирование любого процесса в компьютере позволяет ответить на главный вопрос: правильно ли мы представляем себе процесс морфогенеза у растений, и если правильно, то каких данных не хватает, какие еще наблюдения нужно провести? И конечно же это экономит силы, средства и нервы экспериментаторам».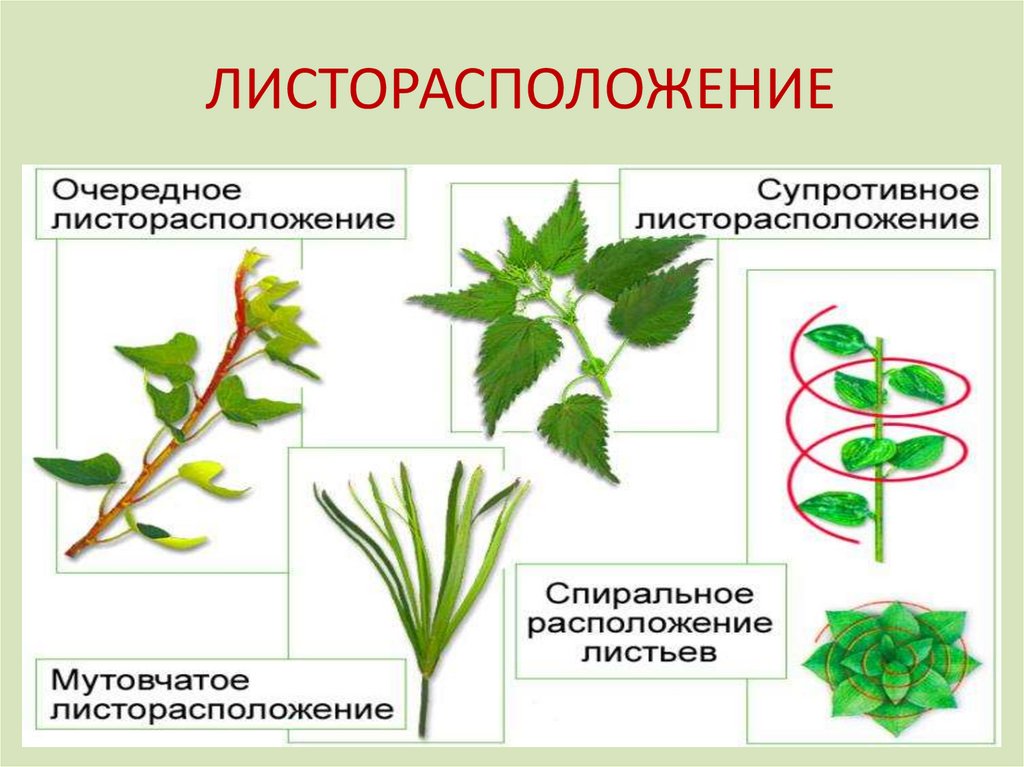
Литература:
Чуб В.В., Юрцева О.В. Математическое моделирование формирования цветка у представителей семейства Polygonaceae. «Ботанический журнал», 2007, 92, 1, 114–134.
Скрябин К.Г., Алексеев Д.В., Ежова Т.А., Козлов В.Н., Кудрявцев В.Б., Носов М.В., Пеннин А.А., Чуб В.В., Шестаков С.В., Шульга О.А. Определение типа и положения органов цветка: динамическая модель развития. «Известия АН. Серия биологическая», 2006, 33, 6, 523–535.
Чуб В.В. Роль позиционной информации в регуляции развития органов цветка и листовых серий побегов. М.: Бином, 2010.
Деревья с супротивными листьями — Sibley Guides
В прошлом справочники по идентификации деревьев подчеркивали наличие противоположных листьев как один из наиболее важных полевых признаков. В Руководстве Сибли по деревьям я использовал более целостный подход, такой как современная идентификация птиц, придавая равный вес всем частям дерева. Дерево может привлечь ваше внимание своими цветами, ветками, корой, плодами и т.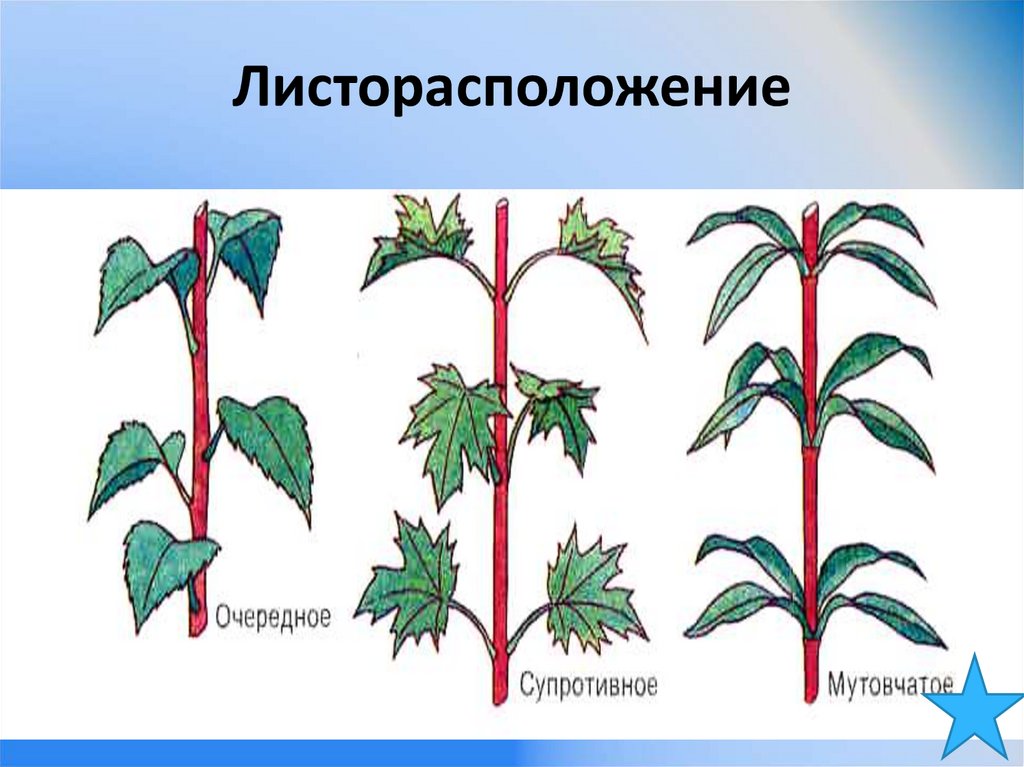 д., и я не думаю, что будет полезно, если полевой гид затем попросит вас проверить, расположены ли листья попеременно или супротивно. Например, лучше узнать все отличительные признаки дубов и кленов, чтобы узнавать их по множеству признаков, а не проверять расположение листьев.
д., и я не думаю, что будет полезно, если полевой гид затем попросит вас проверить, расположены ли листья попеременно или супротивно. Например, лучше узнать все отличительные признаки дубов и кленов, чтобы узнавать их по множеству признаков, а не проверять расположение листьев.
Однако в некоторых случаях, когда вы сталкиваетесь с запутанным деревом, наличие противоположных листьев может быть полезным указателем, помогающим сузить диапазон возможностей. Поэтому ниже я составил список деревьев с противоположными листьями. Интересно, что почти все эти виды встречаются на последних 80 страницах справочника.
Определение из стр. xxiv из The Sibley Guide to Trees :
Листья, которые растут поодиночке от ветки, без другого листа непосредственно поперек ветки, называются очередными, так как они обычно растут с противоположных сторон ветки. . Листья, растущие попарно с противоположных сторон веточки на одном уровне, называются супротивными.
 Если три или более листьев растут с одного уровня на ветке, они называются мутовчатыми.
Если три или более листьев растут с одного уровня на ветке, они называются мутовчатыми.Эти закономерности еще легче обнаружить зимой, так как почки и рубцы на ветвях соответствуют расположению листьев (супротивным или очередным), а ветвление также имеет тенденцию быть супротивным или очередным. Также обратите внимание, что ветки с чередующимися листьями, как правило, растут зигзагообразно, в то время как деревья с противоположными листьями обычно имеют более прямые ветки.
Подсчет видов включает только те, которые показаны в справочнике.
Североамериканские деревья с противоположными листьями
- Katsura, род Cercidophyllum (стр. 101) – один интродуцированный вид [единственный вид в своем семействе]
- Eucalyptus, род Eucalyptus (стр. 109) – у трех интродуцированных видов ювенильная листва супротивная, но взрослая листва чередуется [креп-мирт в том же семействе имеет очередные листья]
- Шелковица бумажная, род Brousonettia (стр. 248) – один интродуцированный вид [у других родов семейства Тутовых нет супротивных листьев]
- Lyontree, род Lyonothamnus (стр. 293) – один аборигенный вид [другие роды семейства розовых не имеют супротивных листьев]
- Beebee Tree, род Evodia (стр. 321) – один интродуцированный вид [связанный с Corktree, но другие виды семейства Rue не имеют супротивных листьев]
- Corktree, род Phellodendron (стр. 323) – один интродуцированный вид [связанный с Beebee Tree, но другие виды семейства Rue не имеют супротивных листьев]
- Buckeyes, род Aesculus (стр. 326-331) – шесть местных и один интродуцированный вид [связанный с кленами, но другие роды семейства мыльных не имеют супротивных листьев]
- Клены, род Acer (стр. 332-351) — 13 местных и десять интродуцированных видов [связанных с каштановыми, но другие роды семейства мыльных не имеют супротивных листьев]
- Кизилы, род Cornus (стр. 355-358) – три (из четырех) местных и один интродуцированный вид имеют супротивные листья [голубиное дерево в том же семействе не имеет]
- Buttonbush, род Cephalanthus (стр. 374) — один местный вид [единственное североамериканское дерево в семействе марены]
- Ясень, род Fraxinus (стр. 376-385) — 17 местных и четыре интродуцированных вида [все остальные роды семейства оливковых также имеют супротивные листья]
- Сирень японская, род Syringa (стр. 386) – один интродуцированный вид [все остальные роды семейства маслиновых также имеют супротивные листья]
- Американское бахромчатое дерево, род Chionanthus (стр. 387) – один аборигенный вид [у всех других родов семейства маслиновых листья супротивные]
- Маслина обыкновенная, род Olea (стр. 388) – один интродуцированный вид [все другие роды семейства маслиновых также имеют супротивные листья]
- Devilwood, род Osmanthus (стр. 388) – один аборигенный вид [у всех остальных родов семейства маслиновых листья супротивные]
- Ива пустынная, род Chilopsis (стр. 389) – один аборигенный вид с супротивными или мутовчатыми листьями [связан с Catalpas, другие роды семейства трубообразных также имеют супротивные листья]
- Catalpas, род Catalpa (стр. 390-391) – два местных и один интродуцированный вид имеют супротивные или мутовчатые листья [родственники пустынной ивы, другие роды семейства трубчатых ползучих также имеют супротивные листья]
- Royal Paulownia, род Paulownia (стр. 392) – один интродуцированный вид [единственный вид в семействе]
- Elders, род Sambucus (стр. 397) – многие аборигенные виды, в основном кустарники [связанные с Viburnums, все остальные роды семейства Muskroot имеют супротивные листья]
- Viburnum, Genus Viburnum (стр. 397) – многие местные виды, в основном кустарники [относящиеся к Elders, все остальные роды семейства Muskroot имеют супротивные листья]
 248) – один интродуцированный вид [у других родов семейства Тутовых нет супротивных листьев]
248) – один интродуцированный вид [у других родов семейства Тутовых нет супротивных листьев] 374) — один местный вид [единственное североамериканское дерево в семействе марены]
374) — один местный вид [единственное североамериканское дерево в семействе марены]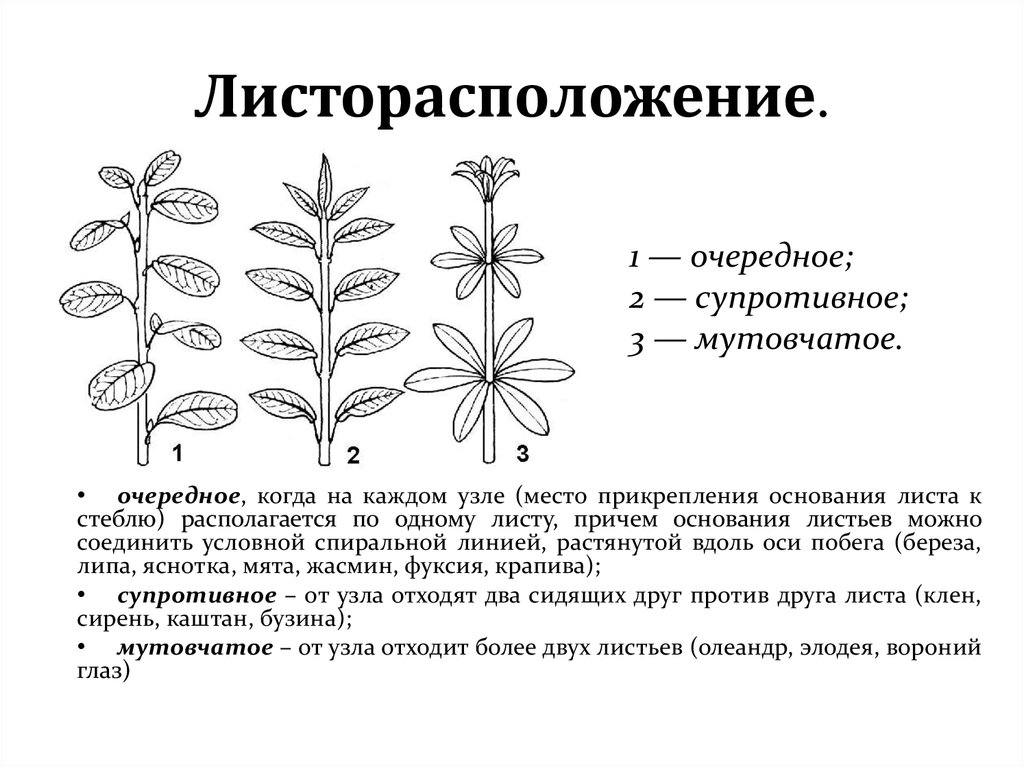 390-391) – два местных и один интродуцированный вид имеют супротивные или мутовчатые листья [родственники пустынной ивы, другие роды семейства трубчатых ползучих также имеют супротивные листья]
390-391) – два местных и один интродуцированный вид имеют супротивные или мутовчатые листья [родственники пустынной ивы, другие роды семейства трубчатых ползучих также имеют супротивные листья]Использование местных растений Джорджии: альтернатива или противоположность
Когда я говорю с людьми об идентификации растений, первое, с чего я начинаю, это расположение листьев. И хотя я обычно говорю о древесных растениях (деревьях, кустарниках и виноградных лозах), расположение листьев на многолетниках также используется для идентификации.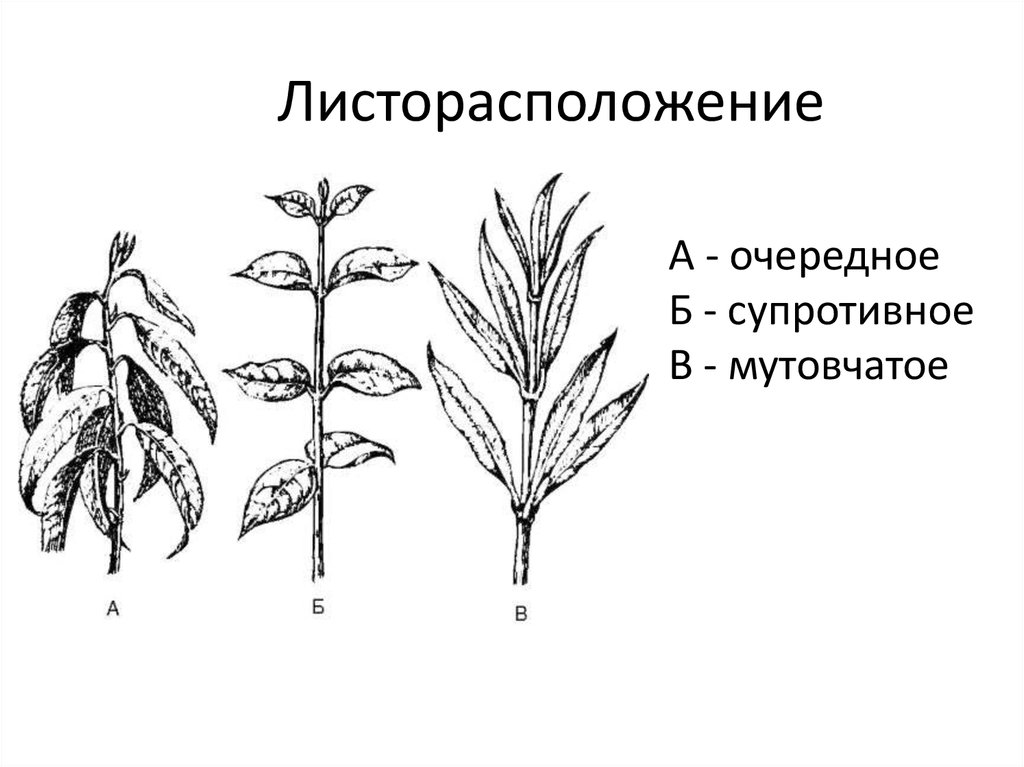 Чтобы упростить концепцию, этот пост будет посвящен древесным растениям.
Чтобы упростить концепцию, этот пост будет посвящен древесным растениям.
| Calycanthus floridus , супротивное расположение листьев |
Расположение листьев на растении либо супротивное, либо очередное: листья могут располагаться супротивно друг другу на стебле или очередно вдоль стебля.
Иногда встречаются растения с листьями, расположенными мутовкой, состоящей из 3 и более листьев вместе. Лилия ( Lilium ) — хороший пример растения с листьями в мутовке.
| Калина с супротивными ветвями |
В зимнее время года вы можете подумать, что вы не можете различить опадающее расположение листьев на лиственных растениях. Подсказки все еще там, даже когда листья исчезли. У более молодых деревьев или кустарников вы можете изучить ветви — они следуют той же схеме, что и листья.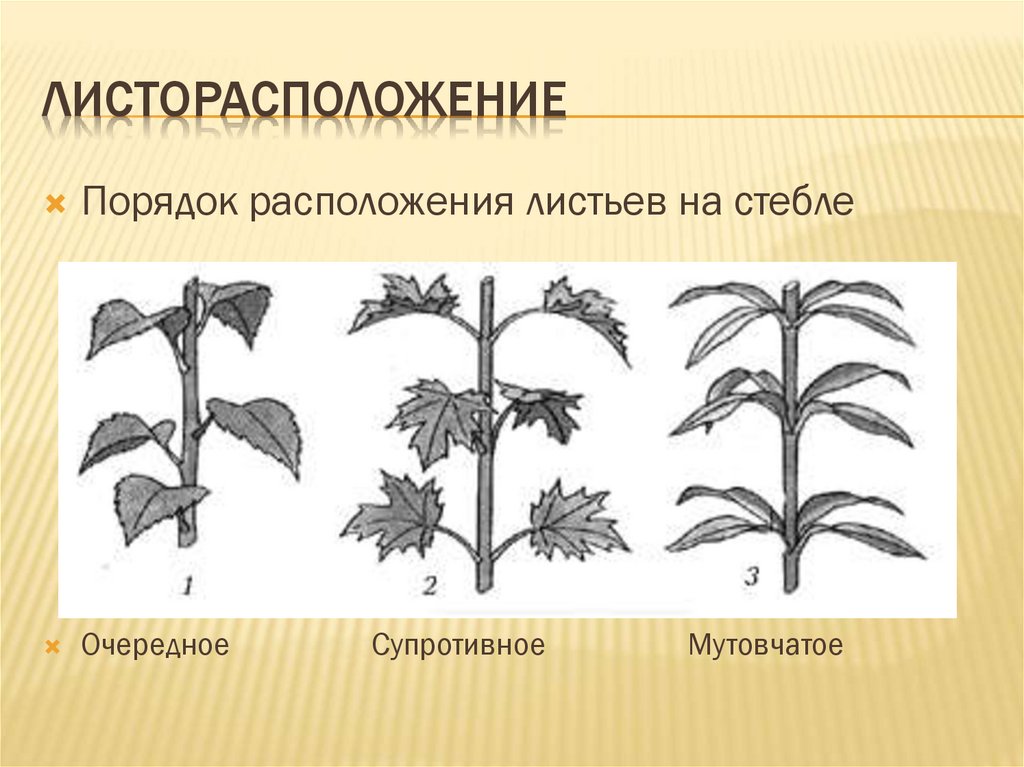 Старые деревья могут не иметь всех исходных ветвей, поэтому лучше осмотреть веточки.
Старые деревья могут не иметь всех исходных ветвей, поэтому лучше осмотреть веточки.
Ветки — это концы ветвей; они представляют собой рост за последний год. У медленно растущего растения веточка может быть очень короткой, а у быстро растущего растения может быть несколько дюймов веточки. Свежий рост на ветке — хорошее место для проверки листовых почек.
| Боярышник ( боярышник ) листовые почки, чередующиеся |
Листовые почки – это листья будущего года. Они могут быть большими и выступающими, как американский бук (9).0007 Fagus grandifolia ), или они могут быть настолько маленькими, что кажутся скрытыми, как сладкий кустарник ( Calycanthus floridus ). В обоих случаях вы все еще можете различить расположение.
Большинство листовых почек достаточно большие, чтобы их можно было увидеть зимой. Обычно вы видите листовую почку, а под ней — рубец (там, где был прикреплен прошлогодний лист). Лишь изредка я чувствую необходимость использовать ручной объектив с 10-кратным увеличением, чтобы выяснить расположение. Еще один совет: ветки с противоположным расположением листьев довольно прямые, в то время как ветки с очередным расположением листьев могут иметь зигзагообразный вид.
Обычно вы видите листовую почку, а под ней — рубец (там, где был прикреплен прошлогодний лист). Лишь изредка я чувствую необходимость использовать ручной объектив с 10-кратным увеличением, чтобы выяснить расположение. Еще один совет: ветки с противоположным расположением листьев довольно прямые, в то время как ветки с очередным расположением листьев могут иметь зигзагообразный вид.
| Листья бука чередующиеся |
Листовые почки не видны весной и летом, потому что они еще не сформировались, но это не имеет значения, потому что у нас есть листья, чтобы помочь нам увидеть расположение. Иногда некоторые листья отсутствуют, поэтому внимательно посмотрите, прежде чем решить, что расположение чередуется; это может быть наоборот с отсутствующими листьями.
Теперь, когда вы определили расположение листьев, вы можете перейти к идентификации растения. Большинство растений имеют чередующееся расположение листьев, поэтому, если у вас противоположное, у вас будет меньший набор вариантов.
Большинство растений имеют чередующееся расположение листьев, поэтому, если у вас противоположное, у вас будет меньший набор вариантов.
Я отказался от использования мнемотехники для запоминания общих противоположных растений; каждая версия, которую я слышал, упускает некоторые из них. Вот распространенные в Грузии: клен ( Acer ), калина ( Aesculus ), калина , ясень ( Fraxinus ), кизил ( Cornus , кроме одного вида чередующийся), бузина ( Sambucus ), бахрома ( Chionanthus ), красавица ягода ( Callicarpa ), Бересклет , жимолость ( Lonicera ), сладкий кустарник ( Calycanthus ) и бутоньерка ( Cephalanthus ).
Большинство книг по идентификации предлагают «ключ», которому вы должны следовать. Ключ проведет вас через серию вопросов, пока растение не будет идентифицировано. Зимние ключи будут сосредоточены на других характеристиках веток, таких как шрамы от листьев, шрамы пучков, верхушечные почки и характеристики почек и наличие чечевичек (здесь вам может понадобиться эта ручная линза).