
Фотосинтез: что такое, определение, фазы. Растения фотосинтез
что такое, определение, фазы :: SYL.ru
Важнейшим органическим процессом, без которого существование всех живых существ нашей планеты было бы под вопросом, является фотосинтез. Что такое фотосинтез? известно всем со школы. Грубо говоря, это процесс образования органических веществ из углекислого газа и воды, который происходит на свету и сопровождается выделением кислорода. Более сложное определение звучит следующим образом: фотосинтез – процесс преобразования световой энергии в энергию химических связей веществ органического происхождения при участии фотосинтетических пигментов. В современной практике под фотосинтезом обычно понимают совокупность процессов поглощения, синтеза и использования света в ряде эндергонических реакций, одной из которых является превращение углекислого газа в органические вещества. А теперь давайте узнаем подробнее, как протекает фотосинтез и на какие фазы этот процесс делится!

Общая характеристика
Хлоропласты, которые есть у каждого растения, отвечают за фотосинтез. Что такое хлоропласты? Это овальные пластиды, в которых содержится такой пигмент, как хлорофилл. Именно хлорофилл определяет зеленую окраску растений. У водорослей данный пигмент представлен в составе хроматофор – пигментсодержащих светоотражающих клеток разной формы. Бурые и красные водоросли, которые обитают на значительных глубинах, куда плохо попадает солнечный свет, имеют иные пигменты.
Вещества фотосинтеза входят в состав автотроф – организмов, способных синтезировать из неорганических веществ органические. Они являются самой нижней ступенью пищевой пирамиды, поэтому входят в рацион всех живых организмов планеты Земля.
Польза фотосинтеза
Зачем же нужен фотосинтез? Кислород, который выделяется из растений во время фотосинтеза, поступает в атмосферу. Поднимаясь в ее верхние слои, он образует озон, который защищает земную поверхность от сильного солнечного излучения. Именно благодаря озоновому экрану живые организмы могут комфортно находиться на суше. Кроме того, как известно, кислород нужен для дыхания живых организмов.

Ход процесса
Все начинается с того, что в хлоропласты попадает свет. Под его влиянием органеллы вытягивают из почвы воду, а также делят ее на водород и кислород. Таким образом, имеют место два процесса. Фотосинтез растений начинается в момент, когда листья уже впитали воду и углекислый газ. Световая энергия аккумулируется в тилакоидах – специальных отсеках хлоропластов, и делит молекулу воды на две составляющие. Часть кислорода уходит на дыхание растения, а оставшаяся часть - в атмосферу.
Затем углекислый газ попадает в пиреноиды – белковые гранулы, окруженные крахмалом. Сюда же поступает водород. Смешавшись друг с другом, эти вещества образуют сахар. Эта реакция также проходит с выделением кислорода. Когда сахар (обобщающее название простых углеводов) смешивается с азотом, серой и фосфором, поступающими в растение из почвы, образуется крахмал (сложный углевод), белки, жиры, витамины и прочие вещества, необходимые для жизни растений. В абсолютном большинстве случаев фотосинтез происходит в условиях природного освещения. Однако искусственное освещение также может в нем поучаствовать.
Классы фотосинтеза
Вплоть до 60-х годов двадцатого века науке был известен один механизм восстановления углекислого газа – по С3-пентозофосфатному пути. Недавно австралийские ученые доказали, что у некоторых видов растений данный процесс может протекать по циклу С4-дикарбоновых кислот.
У растений, которые восстанавливают углекислый газ по С3 пути, фотосинтез лучше всего проходит при умеренной температуре и слабой освещенности, в лесах или темных местах. К таковым растениям можно отнести львиную долю культурных растений и почти все овощи, которые составляют основу нашего рациона.

У второго класса растений фотосинтез активнее всего протекает в условиях высокой температуры и сильной освещенности. В эту группу входят растения, которые произрастают в тропическом и теплом климате, к примеру кукуруза, сахарный тростник, сорго и так далее.
Метаболизм растений, кстати говоря, был обнаружен весьма недавно. Ученым удалось выяснить, что некоторые растения имеют специальные ткани для сохранения запасов воды. Углекислый газ у них скапливается в виде органических кислот и переходит в углеводы лишь через 24 часа. Этот механизм дает растениям возможность сэкономить воду.
Как проходит процесс?
Мы уже знаем в общих чертах, как протекает процесс фотосинтеза и какой фотосинтез бывает, теперь давайте познакомимся с ним глубже.
Начинается все с того, что растение поглощает свет. Ей в этом помогает хлорофилл, который в виде хлоропластов располагается в листьях, стеблях и плодах растения. Основное количество данного вещества сконцентрировано именно в листьях. Все дело в том, что благодаря своей плоской структуре, лист притягивает много света. А чем больше света, тем больше энергии для фотосинтеза. Таким образом, листья в растении выступают своеобразными локаторами, улавливающими свет.
Когда свет поглощен, хлорофилл пребывает в возбужденном состоянии. Он передает энергию другим органам растения, которые участвуют в следующей стадии фотосинтеза. Второй этап процесса протекает без участия света и состоит в химической реакции с участием воды, получаемой из почвы, и углекислого газа, получаемого из воздуха. На этой стадии синтезируются углеводы, которые крайне необходимы для жизни любого организма. В данном случае они не только питают само растение, но и передаются животным, которые его съедают. Люди также получают эти вещества, употребив продукты растительного или животного происхождения.

Фазы процесса
Будучи довольно сложным процессом, фотосинтез делится на две фазы: световую и темновую. Как можно понять из названия, для первой фазы обязательно наличие солнечного излучения, а для второй - нет. Во время световой фазы хлорофилл поглощает квант света, образуя молекулы АТФ и НАДН, без которых невозможен фотосинтез. Что такое АТФ и НАДН?
АТФ (аденозитрифосфат) – нуклеиновый кофермент, которые содержит высокоэнергетические связи и служит источником энергии в любом органическом превращении. Соединение часто называют энергетической волютой.
НАДН (никотинамидадениндинуклеотид) – источник водорода, который используется для синтеза углеводов с участием углекислого газа во второй фазе такого процесса, как фотосинтез.
Световая фаза
Хлоропласты содержат много молекул хлорофилла, каждая из которых поглощает свет. Его поглощают и другие пигменты, но они не способны к фотосинтезу. Процесс проходит лишь в части молекул хлорофилла. Остальные молекулы образуют антенные и светособирающие комплексы (ССК). Они накапливают кванты светового излучения и передают их в реакционные центры, которые также называют ловушками. Реакционные центры располагаются в фотосистемах, которых у фотосинтезирующего растения две. Первая содержит молекулу хлорофилла, способную поглощать свет с длиной волны 700 нм, а вторая – 680 нм.
Итак, два типа молекул хлорофилла поглощают свет и возбуждаются, что способствует переходу электронов на более высокий энергетический уровень. Возбужденные электроны, обладающие большим количеством энергии, отрываются и поступают в цепь переносчиков, расположенную в мембранах тилакоидов (внутренние структуры хлоропластов).

Переход электронов
Электрон из первой фотосистемы переходит от хлорофилла Р680 к пластохинону, а электрон из второй системы – к ферредоксину. При этом на месте отрыва электронов в молекуле хлорофилла образуется свободное место.
Для восполнения недостачи молекула хлорофилла Р680 принимает электроны из воды, образуя ионы водорода. А вторая молекула хлорофилла восполняет недостачу через систему переносчиков от первой фотосистемы.
Так протекает световая фаза фотосинтеза, суть которой состоит в переносе электронов. Параллельно электронному транспорту проходит перемещение ионов водорода через мембрану. Это приводит к их накоплению внутри тилакоида. Накапливаясь в большом количестве, они высвобождаются наружу с помощью сопрягающего фактора. Результатом транспорта электронов является образование соединения НАДН. А перенос иона водорода приводит к образованию энергетической валюты АТФ.
По окончании световой фазы в атмосферу поступает кислород, а внутри лепестка образуются АТФ и НАДН. Затем начинается темновая фаза фотосинтеза.

Темновая фаза
Для этой фазы фотосинтеза необходим углекислый газ. Растение постоянно поглощает его из воздуха. С этой целью на поверхности листка есть устьица – специальные структуры, которые при открывании всасывают углекислый газ. Поступая вовнутрь листка, он растворяется в воде и участвует в процессах световой фазы.
Во время световой фазы в большинстве растений углекислый газ связывается с органическим соединением, которое содержит 5 атомов углерода. В результате образуется пара молекул трехуглеродного соединения под названием 3-фосфоглицериновая кислота. Именно из-за того, что первичным результатом процесса является данное соединение, растения с таким типом фотосинтеза называют С3-растениями.
Дальнейшие процессы, проходящие в хлоропластах, весьма сложны для неискушенных обывателей. В конечном итоге получается шестиуглеродное соединение, синтезирующее простые или сложные углеводы. Именно в виде углеводов растение скапливает энергию. Небольшая часть веществ остается в листе и выполняет его нужды. Остальные углеводы циркулируют по всему растению и поступают в те места, где они больше всего нужны.
Фотосинтез зимой

Многие хотя бы раз в жизни задавались вопросом о том, откуда берется кислород в холодное время года. Во-первых, кислород вырабатывается не только лиственными растениями, но и хвойными, а также морскими растениями. И если лиственные растения зимой замирают, то хвойные продолжают дышать, хотя и менее интенсивно. Во-вторых, содержание кислорода в атмосфере не зависит от того, скинули ли деревья свои листья. Кислород занимает 21 % атмосферы, в любой точки нашей планеты в любое время года. Эта величина не меняется, так как воздушные массы перемещаются очень быстро, а зима наступает не одновременно во всех странах. Ну и, в-третьих, зимой в нижних слоях воздуха, которые мы вдыхаем, содержание кислорода даже больше, чем летом. Причина этого явления – низкая температура, из-за которой кислород становится плотнее.
Заключение
Сегодня мы вспомнили, что такое фотосинтез, что такое хлорофил, и как растения выделяют кислород, поглощая углекислый газ. Безусловно, фотосинтез является важнейшим процессом в нашей жизни. Он напоминает нам о необходимости бережного отношения к природе.
www.syl.ru
Что такое фотосинтез или почему трава зелёная?
Жизнь человека, как и всего живого на Земле невозможна без дыхания. Мы вдыхаем из воздуха кислород, а выдыхаем углекислый газ. Но почему же кислород не кончается? Оказывается, воздух в атмосфере непрерывно подпитывается кислородом. И происходит это насыщение именно благодаря фотосинтезу.
Фотосинтез - просто и понятно!
Каждый человек обязан понимать, что такое фотосинтез. Для этого совсем не нужно писать сложные формулы, достаточно понять всю важность и волшебство этого процесса.
Главную роль в процессе фотосинтеза играют растения – трава, деревья, кустарники. Именно в листьях растений на протяжении миллионов лет происходит удивительное превращение углекислого газа в кислород, так необходимый для жизни любителям дышать. Попробуем разобрать весь процесс фотосинтеза по порядку.
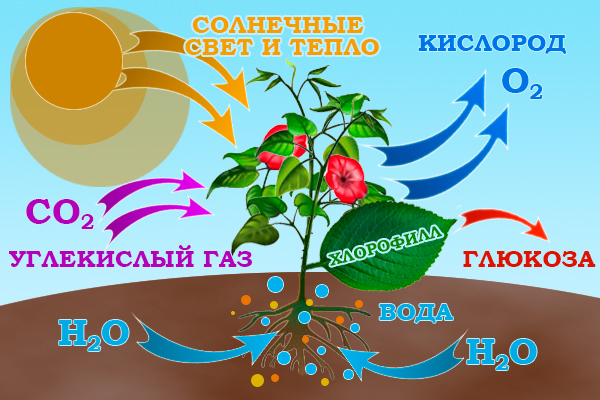
1. Растения берут из почвы воду с растворенными в ней минеральными веществами – азот, фосфор, марганец, калий, различные соли – всего больше 50 различных химических элементов. Это необходимо растениям для питания. Но из земли растения получают лишь 1/5 часть необходимых веществ. Остальные 4/5 они получают из воздуха!
2. Из воздуха растения поглощают углекислый газ. Тот самый углекислый газ, который мы выдыхаем каждую секунду. Углекислым газом растения дышат, как мы с вами дышим кислородом. Но и этого мало.
3. Незаменимый компонент в природной лаборатории - солнечный свет. Солнечные лучи в листьях растений пробуждают необычайную химическую реакцию. Как же это происходит?
4. В листьях растений есть удивительное вещество – хлорофилл. Хлорофилл способен улавливать потоки солнечного света и неутомимо перерабатывать полученные воду, микроэлементы, углекислый газ в органические вещества, необходимые каждому живому существу нашей планеты. В этот момент растения выделяют в атмосферу кислород! Именно эту работу хлорофилла ученые называют сложным словом – фотосинтез.
Презентацию по теме Фотосинтез можно скачать на образовательном портале Учителя.com
Так почему трава зелёная?
Теперь, когда мы знаем, что в клетках растений, содержится хлорофилл, на этот вопрос ответить очень легко. Недаром с древнегреческого языка хлорофилл переводится как «зелёный лист». Для фотосинтеза хлорофилл использует все лучи солнечного света, кроме зеленого. Мы видим траву, листья растений зелеными именно потому, что хлорофилл получается зеленым.

Значение фотосинтеза.
Значение фотосинтеза невозможно переоценить - без фотосинтеза в атмосфере нашей планеты накопилось бы слишком много углекислого газа, большинство живых организмов просто не смогли бы дышать и погибли. Наша Земля превратилась бы в безжизненную планету. Для того чтобы этого не допустить каждому человеку планеты Земля нужно помнить, что мы очень обязаны растениям.
Именно поэтому так важно в городах делать как можно больше парков и зелёных насаждений. Беречь от уничтожения тайгу и джунгли. Или просто посадить дерево рядом с домом. Или не ломать ветки. Только участие каждого человека планеты Земля поможет сохранить жизнь на родной планете.
Но важность фотосинтеза не ограничивается переработкой углекислого газа в кислород. Именно в результате фотосинтеза сформировался озоновый слой в атмосфере, защищающий планету от губительных лучей ультрафиолета. Растения это пища для большинства живых существ на Земле. Пища необходимая и полезная. Питательность растений это тоже заслуга фотосинтеза.
С недавнего времени хлорофилл стали активно использовать в медицине. Люди издавна знали, что больные животные инстинктивно едят зеленые листья, чтобы вылечиться. Ученые выяснили, что хлорофилл сходен с веществом в клетках крови человека и способен творить настоящие чудеса.
ya-uznayu.ru
Что такое фотосинтез?
Человеческому организму для нормального функционирования необходима пища, вода и кислород. Воздух, которым мы дышим, на двадцать процентов состоит из кислорода. Мы знаем, что кислород вырабатывается в процессе фотосинтеза. Однако знаете ли вы, что такое фотосинтез?
Ниже описаны механика фотосинтеза, его разновидности и значение для нашего мира.
Механизм фотосинтеза
Фотосинтезом называют процесс генерации питательных веществ зеленых растений. Синтез питательных веществ из углекислого газа происходит в хлорофиллах (зеленых пигментах растений) под воздействием света. Важную роль в этом процессе играют хлоропласты, свет, углекислый газ, вода и температура.
Ход процесса:
- Свет, участвующий в процессе фотосинтеза, поглощается хлорофиллами.
- В хлорофиллах содержатся хлоропласты – внутриклеточные органоиды зеленого пигмента. Под действием солнечного света хлоропласты вытягивают воду из почвы, разделяя ее на водород и кислород.
- Во время фотосинтеза протекают два процесса. Фотосинтез в листьях начинается после того, как они впитают воду и углекислый газ.
- Световая энергия собирается в специальные отсеки хлоропластов, называемые тилакоиды, а затем делит молекулу воды на кислород и водород.
- Часть кислорода вырабатывается в атмосферу, а часть идет на дыхание растения. После чего углекислый газ в пиреноидах (белковых гранулах, окруженных крахмалом) смешивается с водородом и образует молекулы сахара. В результате этой реакции также выделяется кислород.
- Смешивая сахар с азотом, серой и фосфором, добываемыми из почвы, зеленые растения производят крахмал, жиры, белки, витамины и другие сложные соединения, необходимые для жизни.
Хотя в абсолютном большинстве случаев фотосинтез протекает под воздействием солнечного света, в нем также может участвовать и искусственное освещение.
Виды фотосинтеза
Имеет смысл говорить не о различных видах, а скорее о различных механизмах фотосинтеза. Дело в том, что до 60-ых гг. прошлого века ученым был известен только один механизм фиксации углекислого газа - по С3-пентозофосфатному пути. Однако недавно группа австралийских ученых смогла доказать, что у некоторых растений восстановление углекислого газа происходит по циклу C4-дикарбоновых кислот:
- У растений с реакцией С3 фотосинтез наиболее активно происходит в условиях умеренной температуры и освещенности, в основном, в лесах и в темных местах. К таким растениям относятся почти все культурные растения и большая часть овощей. Они составляют основу рациона человека.
- У растений с реакцией С4 фотосинтез наиболее активно происходит в условиях высоких температура и освещенности. К таким растениям относятся, например, кукуруза, сорго и сахарный тростник, которые произрастают в теплом и тропическом климате.
- Сам метаболизм растений был обнаружен совсем недавно, когда удалось выяснить, что у некоторых растений, имеющих специальные ткани для запаса воды, углекислый газ накапливается в форме органических кислот и фиксируется в углеводах лишь спустя сутки. Такой механизм помогает растениям экономить запасы воды.
Значение фотосинтеза для нашей планеты
Раньше на нашей планете Земля кислорода не было. Его появлению мы обязаны фотосинтезирующим клеткам. Благодаря им в атмосферу планеты стал активно вырабатываться кислород и появилось кислородное дыхание – самый выгодный способ энергетического обмена. Кислород стал причиной образования защитного озонового слоя вокруг Земли, который защищает планету от вредных солнечных радиоактивных лучей. Это позволило живым организмам выйти из океана на сушу, что предопределило дальнейший эволюционный путь развития живых существ.
Но не только выработка кислорода важна в процессе фотосинтеза. Немаловажен и тот факт, что растения поглощают из атмосферы углекислый газ. Переизбыток углекислого газа может вызвать парниковый эффект, что крайне негативно сказалось бы на всех живых организмах нашей планеты. За последние несколько десятилетий содержание углекислого газа в атмосфере порядком увеличилось. Виной тому стала промышленная революция и дальнейший технический прогресс. Ученые продолжают спорить, к каким последствиям это приведет, однако уже сегодня призывают мировое сообщество обратить внимание на данную проблему, пока не стало слишком поздно.
Читайте также статью Как происходит фотосинтез.
elhow.ru
Фотосинтез • Джеймс Трефил, энциклопедия «Двести законов мироздания»
Растения превращают солнечный свет в запасенную химическую энергию в два этапа: сначала они улавливают энергию солнечного света, а затем используют ее для связывания углерода с образованием органических молекул.
Зеленые растения — биологи называют их автотрофами — основа жизни на планете. С растений начинаются практически все пищевые цепи. Они превращают энергию, падающую на них в форме солнечного света, в энергию, запасенную в углеводах (см. Биологические молекулы), из которых важнее всего шестиуглеродный сахар глюкоза. Этот процесс преобразования энергии называется фотосинтезом. Другие живые организмы получают доступ к этой энергии, поедая растения. Так создается пищевая цепь, поддерживающая планетарную экосистему.
Кроме того, воздух, которым мы дышим, благодаря фотосинтезу насыщается кислородом. Суммарное уравнение фотосинтеза выглядит так:
вода + углекислый газ + свет → углеводы + кислород
Растения поглощают углекислый газ, образовавшийся при дыхании, и выделяют кислород — продукт жизнедеятельности растений (см. Гликолиз и дыхание). К тому же, фотосинтез играет важнейшую роль в круговороте углерода в природе.
Кажется удивительным, что при всей важности фотосинтеза ученые так долго не приступали к его изучению. После эксперимента Ван Гельмонта, поставленного в XVII веке, наступило затишье, и лишь в 1905 году английский физиолог растений Фредерик Блэкман (Frederick Blackman, 1866–1947) провел исследования и установил основные процессы фотосинтеза. Он показал, что фотосинтез начинается при слабом освещении, что скорость фотосинтеза возрастает с увеличением светового потока, но, начиная с определенного уровня, дальнейшее усиление освещения уже не приводит к повышению активности фотосинтеза. Блэкман показал, что повышение температуры при слабом освещении не влияет на скорость фотосинтеза, но при одновременном повышении температуры и освещения скорость фотосинтеза возрастает значительно больше, чем при одном лишь усилении освещения.
На основании этих экспериментов Блэкман заключил, что происходят два процесса: один из них в значительной степени зависит от уровня освещения, но не от температуры, тогда как второй сильно определяется температурой независимо от уровня света. Это озарение легло в основу современных представлений о фотосинтезе. Два процесса иногда называют «световой» и «темновой» реакцией, что не вполне корректно, поскольку оказалось, что, хотя реакции «темновой» фазы идут и в отсутствии света, для них необходимы продукты «световой» фазы.
Фотосинтез начинается с того, что излучаемые солнцем фотоны попадают в особые пигментные молекулы, находящиеся в листе, — молекулы хлорофилла. Хлорофилл содержится в клетках листа, в мембранах клеточных органелл хлоропластов (именно они придают листу зеленую окраску). Процесс улавливания энергии состоит из двух этапов и осуществляется в раздельных кластерах молекул — эти кластеры принято называть Фотосистемой I и Фотосистемой II. Номера кластеров отражают порядок, в котором эти процессы были открыты, и это одна из забавных научных странностей, поскольку в листе сначала происходят реакции в Фотосистеме II, и лишь затем — в Фотосистеме I.
Когда фотон сталкивается с 250-400 молекулами Фотосистемы II, энергия скачкообразно возрастает и передается на молекулу хлорофилла. В этот момент происходят две химические реакции: молекула хлорофилла теряет два электрона (которые принимает другая молекула, называемая акцептором электронов) и расщепляется молекула воды. Электроны двух атомов водорода, входивших в молекулу воды, возмещают два потерянных хлорофиллом электрона.
После этого высокоэнергетический («быстрый») электрон перекидывают друг другу, как горячую картофелину, собранные в цепочку молекулярные переносчики. При этом часть энергии идет на образование молекулы аденозинтрифосфата (АТФ), одного из основных переносчиков энергии в клетке (см. Биологические молекулы). Тем временем немного другая молекула хлорофилла Фотосистемы I поглощает энергию фотона и отдает электрон другой молекуле-акцептору. Этот электрон замещается в хлорофилле электроном, прибывшим по цепи переносчиков из Фотосистемы II. Энергия электрона из Фотосистемы I и ионы водорода, образовавшиеся ранее при расщеплении молекулы воды, идут на образование НАДФ-Н, другой молекулы-переносчика.
В результате процесса улавливания света энергия двух фотонов запасается в молекулах, используемых клеткой для осуществления реакций, и дополнительно образуется одна молекула кислорода. (Отмечу, что в результате еще одного, значительно менее эффективного процесса с участием одной лишь Фотосистемы I, также образуются молекулы АТФ.) После того как солнечная энергия поглощена и запасена, наступает очередь образования углеводов. Основной механизм синтеза углеводов в растениях был открыт Мелвином Калвином, проделавшим в 1940-е годы серию экспериментов, ставших уже классическими. Калвин и его сотрудники выращивали водоросль в присутствии углекислого газа, содержащего радиоактивный углерод-14. Им удалось установить химические реакции темновой фазы, прерывая фотосинтез на разных стадиях.
Цикл превращения солнечной энергии в углеводы — так называемый цикл Калвина — сходен с циклом Кребса (см. Гликолиз и дыхание): он тоже состоит из серии химических реакций, которые начинаются с соединения входящей молекулы с молекулой-«помощником» с последующей инициацией других химических реакций. Эти реакции приводят к образованию конечного продукта и одновременно воспроизводят молекулу-«помощника», и цикл начинается вновь. В цикле Калвина роль такой молекулы-«помощника» выполняет пятиуглеродный сахар рибулозодифосфат (РДФ). Цикл Калвина начинается с того, что молекулы углекислого газа соединяются с РДФ. За счет энергии солнечного света, запасенной в форме АТФ и НАДФ-H, сначала происходят химические реакции связывания углерода с образованием углеводов, а затем — реакции воссоздания рибулозодифосфата. На шести витках цикла шесть атомов углерода включаются в молекулы предшественников глюкозы и других углеводов. Этот цикл химических реакций будет продолжаться до тех пор, пока поступает энергия. Благодаря этому циклу энергия солнечного света становится доступной живым организмам.
В большинстве растений осуществляется описанный выше цикл Калвина, в котором углекислый газ, непосредственно участвуя в реакциях, связывается с рибулозодифосфатом. Эти растения называются C3-растениями, поскольку комплекс «углекислый газ—рибулозодифосфат» расщепляется на две молекулы меньшего размера, каждая из которых состоит из трех атомов углерода. У некоторых растений (например, у кукурузы и сахарного тростника, а также у многих тропических трав, включая ползучий сорняк) цикл осуществляется по-другому. Дело в том, что углекислый газ в норме проникает через отверстия в поверхности листа, называемые устьицами. При высоких температурах устьица закрываются, защищая растение от чрезмерной потери влаги. В C3-растения при закрытых устьицах прекращается и поступление углекислого газа, что приводит к замедлению фотосинтеза и изменению фотосинтетических реакций. В случае же кукурузы углекислый газ присоединяется к трехуглеродной молекуле на поверхности листа, затем переносится во внутренние участки листа, где углекислый газ высвобождается и начинается цикл Калвина. Благодаря этому довольно сложному процессу фотосинтез у кукурузы осуществляется даже в очень жаркую, сухую погоду. Растения, в которых происходит такой процесс, мы называем C4-растениями, поскольку углекислый газ в начале цикла транспортируется в составе четырехуглеродной молекулы. C3-растения — это в основном растения умеренного климата , а C4-растения в основном произрастают в тропиках.
Гипотеза Ван Ниля
Процесс фотосинтеза описывается следующей химической реакцией:
СО2 + Н2О + свет → углевод + О2
В начале XX века считалось, что кислород, выделяющийся в процессе фотосинтеза, образуется в результате расщепления углекислого газа. Эту точку зрения опроверг в 1930-е годы Корнелис Бернардус Ван Ниль (Van Niel, 1897–1986), в то время аспирант Стэнфордского университета в штате Калифорния. Он занимался изучением пурпурной серобактерии (на фото), которая нуждается для осуществления фотосинтеза в сероводороде (h3S) и выделяет в качестве побочного продукта жизнедеятельности атомарную серу. Для таких бактерий уравнение фотосинтеза выглядит следующим образом:
СО2 + Н2S + свет → углевод + 2S.
Исходя из сходства этих двух процессов, Ван Ниль предположил, что при обычном фотосинтезе источником кислорода является не углекислый газ, а вода, поскольку у серобактерий, в метаболизме которых вместо кислорода участвует сера, фотосинтез возвращает эту серу, являющуюся побочным продуктом реакций фотосинтеза. Современное подробное объяснение фотосинтеза подтверждает эту догадку: первой стадией процесса фотосинтеза (осуществляемой в Фотосистеме II) является расщепление молекулы воды.
elementy.ru
Фотосинтез | Наука | FANDOM powered by Wikia

лист растения
Фотоси́нтез — фотохимические реакции или Фотохимия — это процесс образования органического вещества из углекислого газа и воды на свету при участии фотосинтетических пигментов (хлорофилл у растений, бактероихлорофилл и бактериородопсин у бактерий). В современной физиологии растений под фотосинтезом чаще понимается фотоавтотрофная функция — совокупность процессов поглощения, превращения и использования энергии квантов света в различных эндэргонических реакциях, в том числе превращения углекислого газа в органические вещества.
Различают оксигенный и аноксигенный типы фотосинтеза. Оксигенный гораздо более широко распространён, его осуществляют растения, цианобактерии и прохлорофиты. В данной статье описан только он, аноксигенному фотосинтезу пурпурных и зелёных бактерий, а также геликобактерий посвящена отдельная статья.
Выделяют три этапа фотосинтеза: фотофизический, фотохимический и химический. На первом этапе происходит поглощение квантов света пигментами, их переход в возбуждённое состояние и передача энергии к другим молекулам фотосистемы. На втором этапе происходит разделение зарядов в реакционном центре, перенос электронов по фотосинтетической электронотранспортной цепи, что заканчивается синтезом АТФ и НАДФН. Первые два этапа вместе называют светозависимой стадией фотосинтеза. Третий этап происходит уже без обязательного участия света и включает в себя биохимические реакции синтеза органических веществ с использованием энергии, накопленной на светозависимой стадии. Чаще всего в качестве таких реакций рассматривается цикл Кальвина и глюконеогенез, образование сахаров и крахмала из углекислого газа воздуха.
Пространственная локализация  Править
Править

Хлоропласты в клетках листа
Фотосинтез растений осуществляется в хлоропластах, обособленных двухмембранных органеллах клетки. Хлоропласты могут быть в клетках плодов, стеблей, однако основным органом фотосинтеза, анатомически приспособленным к его ведению, является лист. В листе наиболее богата хлоропластами ткань палисадной паренхимы. У некоторых суккулентов с вырожденными листьями (например, кактусы) основная фотосинтетическая активность связана со стеблем.
Свет для фотосинтеза захватывается более полно благодаря плоской форме листа, обеспечивающей большое отношение поверхности к объёму. Вода доставляется из корня по развитой сети сосудов (жилок листа). Углекислый газ поступает отчасти посредством диффузии через кутикулу и эпидермис, однако большая его часть диффундирует в лист через устьица и по листу по межклеточному пространству. Растения, осуществляющие С4 и CAM фотосинтез сформировали особые механизмы для активной ассимиляции углекислого газа.
Внутреннее пространство хлоропласта заполнено бесцветным содержимым (стромой) и пронизано мембранами (ламеллами), которые соединяясь друг с другом образуют тилакоиды, которые в свою очередь группируются в стопки, называемые граны. Внутритилакоидное пространство отделено и не сообщается с остальной стромой, предполагается также что внутреннее пространство всех тилакоидов сообщается между собой. Световые стадии фотосинтеза приурочены к мембранам, автотрофная фиксация CO2 происходит в строме.
В хлоропластах имеются свои ДНК, РНК, рибосомы (70s типа), идёт синтез белка (хотя этот процесс и контролируется из ядра). Они не синтезируются вновь, а образуются путём деления предшествующих. Всё это позволило считать их предшественниками свободные цианобактерии, вошедшие в состав эукариотической клетки в процессе симбиогенеза.
Цианобактерии, таким образом, как бы сами являются хлоропластом и в их клетке фотосинтетический аппарат не вынесен в особую органеллу. Их тилакоиды, однако, не образуют стопок, а формируют различные складчатые структуры (у единственной цианобактерии Gloeobacter violaceus тилакоиды отсутствуют вовсе, а весь фотосинтетический аппарат находится в цитоплазматической мембране, не образующей впячиваний). У них и растений также есть различия в светособирающем комплексе (см. ниже) и пигментном составе.
Световая (светозависимая) стадия Править
В ходе световой стадии фотосинтеза образуется высокоэнергетические продукты: АТФ, служащий в клетке источником энергии, и НАДФН, использующийся как восстановитель. В качестве побочного продукта выделяется кислород. В общем роль световых реакций фотосинтеза заключается в том, что в световую фазу синтезируются молекула АТФ и молекулы-переносчики протонов, т.е. НАДФ Н2.
Фотохимическая суть процесса Править
Хлорофилл имеет два уровня возбуждения (с этим связано наличие двух максимумов на спектре его поглощения): первый связан с переходом на более высокий энергетический уровень электрона системы сопряжённых двойных связей, второй — с возбуждением неспаренных электронов азота и кислорода порфиринового ядра. При неизменном спине электрона формируются синглетные первое и второе возбуждённое состояние, при изменённом — триплетное первое и второе.
Второе возбуждённое состояние наиболее высокоэнергетично, нестабильно и хлорофилл за 10-12 сек переходит с него на первое, с потерей 100 кДж/моль энергии только в виде теплоты. Из первого синглетного и триплетного состояний молекула может переходить в основное с выделением энергии в виде света (флуоресценция) или тепла, с переносом энергии на другую молекулу, либо, поскольку электрон на высоком энергетическом уровне слабо связан с ядром, с переносов электрона на другое соединение.
Вторая возможность реализуется в светособирающих комплексах, первая — в реакционных центрах, где переходящий в возбужденное состояние под воздействием кванта света хлорофилл становится донором электрона (восстановителем) и передаёт его на первичный акцептор. Чтобы предотвратить возвращение электрона на положительно заряженный хлорофилл, первичный акцептор передаёт его вторичному. Кроме того, время жизни полученных соединений выше чем у возбуждённой молекулы хлорофилла. Происходит стабилизация энергии и разделения зарядов. Для дальнейшей стабилизации вторичный донор электронов окисляет положительно заряженный хлорофилл, первичным донором же является в случае оксигенного фотосинтеза вода.
Проблемой, с которой сталкиваются при этом проводящие оксигенный фотосинтез организмы, является различие окислительно-восстановительных потенциалов воды (для полуреакции h30 → O2 E0=+0,82 В) и НАДФ+ (E0=-0,32 В). Хлорофилл при этом должен иметь в основном состоянии потенциал больший +0,82 В чтобы окислять воду, но при этом иметь в возбуждённом состоянии потенциал меньший чем −0,32 В чтобы восстанавливать НАДФ+. Одна молекула хлорофилла не может отвечать обоим требованиям. Поэтому сформировалось две фотосистемы и для полного проведения процесса необходимо два кванта света и два хлорофилла разных типов.
Светособирающие комплексы Править
Хлорофилл выполняет две функции: поглощения и передачу энергии. Более 90 % всего хлорофилла хлоропластов входит в состав светособирающих комплексов (ССК), выполняющих роль антенны, передающей энергию к реакционному центру фотосистем I или II. Помимо хлорофилла в ССК имеются каротиноиды, а у некоторых водорослей и цианобактерий — фикобилины, роль которых заключается в поглощении света тех длин волн, которые хлорофилл поглощает сравнительно слабо.
Передача энергии идёт резонансным путём (механизм Фёрстера) и занимает для одной пары молекул 10-10-10-12 сек., расстояние на которое осуществляется перенос составляет около 1 нм. Передача сопровождается некоторыми потерями энергии (10 % от хлорофилла a к хлорофиллу b, 60 % от каратиноидов к хлорофиллу), из-за чего возможна только от пигмента с максимумом поглощения при меньшей длине волны к пигменту с большей. Именно в таком порядке взаимно локализуются пигменты ССК, причём наиболее длинноволновые хлорофиллы находятся в реакционных центрах. Обратный переход энергии невозможен.
ССК растений расположен в мембранах тилакоидов, у цианобактерий основная его часть вынесена за пределы мембран в прикреплённые к ним фикобилисомы — палочковидные полипептидно-пигментные комплексы, в которых находятся различные фикобилины: на периферии фикоэритрины (с максимумом поглощения при 495—565 нм), за ними фикоцианины (550—615 нм) и аллофикоцианины (610—670 нм), последовательно передающие энергию на хлорофилл a (680—700 нм) реакционного центра.
Основные компоненты электронтранспортной цепи Править
Фотосистема II Править
Фотосистема — совокупность ССК, фотохимического реакционного центра и переносчиков электрона. Светособирающий комплекс II содержит 200 молекул хлорофилла a, 100 молекул хлорофилла b, 50 молекул каротиноидов и 2 молекулы феофитина. Реакционный центр фотосистемы II представляет собой пигмент-белковый комплекс, расположенный в тилакоидных мембранах и окружённый ССК. В нём находится димер хлорофилла a с максимумом поглощения при 680 нм (П680). На него в конечном счёте передаётся энергия кванта света из ССК, в результате чего один из электронов переходит на более высокое энергетическое состояние, связь его с ядром ослабляется и возбуждённая молекула П680 становится сильным восстановителем (E0=-0,7 В).
П680 восстанавливает феофитин, в дальнейшем электрон переносится на хиноны, входящие в состав ФС II и далее на пластохиноны, транспортируемые в восстановленной форме к b6f комплексу. Одна молекула пластохинона переносит 2 электрона и 2 протона, которые берутся из стромы.
Заполнение электронной вакансии в молекуле П680 происходит за счёт воды. В состав ФС II входит водоокисляющий комплекс, содержащий в активном центре ионы марганца в количестве 4 штук. Для образования одной молекулы кислорода требуется две молекулы воды, дающие 4 электрона. Поэтому процесс проводится в 4 такта и для его полного осуществления требуется 4 кванта света. Комплекс находится со стороны внутритилакоидного пространства и полученные 4 протона выбрасываются в него.
Таким образом, суммарный результат работы ФС II — это окисление 2 молекул воды с помощью 4 квантов света с образованием 4 протонов во внутритилакоидном пространстве и 2 восстановленных пластохинонов в мембране.
b6f или b/f-комплекс Править
b6f комплекс является насосом, перекачивающим протоны из стромы во внутритилакоидное пространство и создающий градиент их концентрации за счёт выделяющейся в окислительно-восстановительных реакциях электронтранспортной цепи энергии. 2 пластохинона дают перекачку 4 протонов. В дальнейшем трансмембранный протонный градиент (pH стромы около 8, внутритилакоидного пространства — 5) используется для синтеза АТФ трансмембранным ферментом АТФ-синтазой.
Фотосистема I Править
Светособирающий комплекс I содержит примерно 200 молекул хлорофилла.
В реакционном центре первой фотосистемы находится димер хлорофилла a с максимумом поглощения при 700 нм (П700). После возбуждения квантом света он восстанавливает первичный акцептор — хлорофилл a, тот — вторичный (витамин K1), после чего электрон передаётся на филлохинон, от него на ферридоксин, который и восстанавливает НАДФ с помощью фермента ферридоксин-НАДФ-редуктазы.
Белок пластоцианин, восстановленный в b6f комплексе, транспортируется к реакционному центру первой фотосистемы со стороны внутритилакоидного пространства и передаёт электрон на окисленный П700.
Циклический и псевдоциклический транспорт электрона Править
Помимо полного нециклического пути электрона, описанного выше, обнаружены циклический и псевдоциклический.
Суть циклического пути заключается в том, что ферридоксин вместо НАДФ восстанавливает пластохинон, который переносит его назад на b6f комплекс. В результате образуется больший протонный градиент и больше АТФ, но не возникает НАДФН.
При псевдоциклическом пути ферридоксин восстанавливает кислород, который в дальнейшем превращается в воду и может быть использован в фотосистеме II. При этом также не образуется НАДФН.
Темновая стадия Править
В темновой стадии с участием АТФ и НАДФН происходит восстановление CO2 до глюкозы. Хотя свет не требуется для осуществления данного процесса, он участвует в его регуляции.
С3 — фотосинтез, цикл Кальвина Править
Цикл Кальвина или восстановительный пентозофосфатный цикл состоит из трёх стадий:
- Карбоксилирования
- Восстановления
- Регенерация акцептора CO2
На первой стадии к рибулозо-1,5-бифосфату присоединяется CO2 под действием фермента рибулозобисфосфат-карбоксилаза/оксигеназа (Rubisco). Этот белок составляет основную фракцию белков хлоропласта и предположительно наиболее распространённый фермент в природе. В результате образуется промежуточное неустойчивое соединение, распадающееся на две молекулы 3-фосфоглицериновой кислоты (ФГК).
Во второй стадии ФГК в два этапа восстанавливается. Сначала она фосфорилируется АТФ под действием фосфороглицерокиназы, затем НАДФН при воздействии триозофосфатдегидрогеназы её карбоксильная группа окисляется до альдегидной и она становится углеводом (ФГА).
В третьей стадии участвуют 5 молекул ФГА, которые через образование 4-, 5-, 6- и 7-углеродных соединений объединяются в 3 5-углеродных рибулозо-1,5-бифосфата, для чего необходимы 3АТФ.
Наконец, две ФГА необходимы для синтеза глюкозы. Для образования одной её молекулы требуется 6 оборотов цикла, 6 CO2, 12 НАДФН и 18 АТФ.
С4 — фотосинтез Править
При низкой концентрации растворённого в строме CO2 Rubisco катализирует реакцию окисления рибулёзо-1,5-бифосфата и его распад на 3-фосфоглицериновую кислоту и фосфогликолевую кислоту, которая вынужденно используется в процессе фотодыхания.
Для увеличения концентрации CO2 растения С4 типа изменили анатомию листа. Цикл Кальвина у них локализуется в клетках обкладки проводящего пучка, в клетках мезофилла же под действием ФЕП-карбоксилазы фосфоенолпируват карбоксилируется с образованием щавелеуксусной кислоты, которая превращается в малат или аспартат и транспортируется в клетки обкладки, где декарбоксилируется с образованием пирувата, возвращаемого в клетки мезофилла.
С4 фотосинтез практические не сопровождается потерями рибулёзо-1,5-бифосфата из цикла Кальвина, поэтому более эффективен. Однако он требует не 18, а 30 АТФ на синтез 1 молекулы глюкозы. Это оправдывает себя в тропиках, где жаркий климат требует держать устьица закрытыми, что препятствует поступлению CO2 в лист, а также при рудеральной жизненной стратегии.
САМ фотосинтез Править
При CAM (Crassulaceae acid metabolism) фотосинтезе происходит разделение ассимиляции CO2 и цикла Кальвина не в пространстве как у С4, а во времени. Ночью в вакуолях клеток по аналогичному вышеописанному механизму при открытых устьицах накапливается малат, днём при закрытых устьицах идёт цикл Кальвина. Этот механизм позволяет максимально экономить воду, однако уступает в эффективности и С4, и С3. Он оправдан при стресстолерантной жизненной стратегии.
Значение фотосинтеза Править
Фотосинтез является основным источником биологической энергии, фотосинтезирующие автотрофы используют её для синтеза органических веществ из неорганических, гетеротрофы существуют за счёт энергии, запасённой автотрофами в виде химических связей, высвобождая её в процессах дыхания и брожения. Энергия получаемая человечеством при сжигании ископаемого топлива (уголь, нефть, природный газ, торф) также является запасённой в процессе фотосинтеза.
Фотосинтез является главным входом неорганического углерода в биологический цикл. Весь кислород атмосферы биогенного происхождения и является его побочным продуктом. Формирование окислительной атмосферы полностью изменило состояние земной поверхности, сделало возможным появление дыхания, а в дальнейшем, после образования озонового слоя, позволило жизни выйти на сушу.
Изучение фотосинтеза Править
Первые опыты по фотосинтезу были проведены Джозефом Пристли в 1770-1780-х годах, когда он обратил внимание на «порчу» воздуха в герметичном сосуде горящей свечой (воздух переставал быть способен поддерживать горение, помещённые в него животные задыхались) и «исправление» его растениями. Пристли сделал вывод что растения выделяют кислород, который необходим для дыхания и горения, однако не заметил что для этого растениям нужен свет. Это показал вскоре Ян Ингенхауз.
Позже было установлено что помимо выделения кислорода растения поглощают углекислый газ и при участии воды синтезируют на свету органическое вещество. В 1842 Роберт Майер на основании закона сохранения энергии постулировал что растения преобразуют энергию солнечного света в энергию химических связей. В 1877 В. Пфеффер назвал этот процесс фотосинтезом.
Хлорофиллы был впервые выделены в 1818 П. Ж. Пелетье и Ж. Кавенту. Разделить пигменты и изучить их по отдельности удалось М. С. Цвету с помощью созданного им метода хроматографии. Спектры поглощения хлорофилла были изучены К. А. Тимирязевым, он же, развивая положения Майера, показал что именно поглощенные позволяют повысить энергию системы, создав вместо слабых связей С-О и О-Н высокоэнергетические С-С (до этого считалось что в фотосинтезе используются жёлтые лучи, не поглощаемые пигментами листа). Сделано это было благодаря созданному им методу учёта фотосинтеза по поглощённому CO2, в ходе экспериментов по освещению растения светом разных длин волн (разного цвета) оказалось что интенсивность фотосинтеза совпадает со спектром поглощения хлорофилла.
Окислительно-восстановительную сущность фотосинтеза (как оксигенного, так и аноксигенного) постулировал Корнелис ван Ниль. Это означало что кислород в фотосинтезе образуется полностью из воды, что экспериментально подтвердил в 1941 А. П. Виноградов в опытах с изотопной меткой. В 1937 Роберт Хилл установил что процесс окисления воды (и выделения кислорода), а также ассимиляции CO2 можно разобщить. В 1954-1958 Д. Арнон установил механизм световых стадий фотосинтеза, а сущность процесса ассимиляции CO2 была раскрыта Мельвином Кальвином с использованием изотопов углерода в конце 1940-х, за эту работу в 1961 ему была присуждена Нобелевская премия.
В 1955 была выделена и очищена Rubisco. С4 фотосинтез был описан Ю. С. Карпиловым в 1960 и М. Д. Хэтчем и К. Р. Слэком в 1966.
- Холл Д., Рао К. Фотосинтез: Пер. с англ. — М.: Мир, 1983.
ru.science.wikia.com
Фотосинтез - это... Что такое Фотосинтез?

 Лист растения
Лист растения 
Фотосинтез (от др.-греч. φῶς — свет и σύνθεσις — соединение, складывание, связывание, синтез) — процесс образования органических веществ из углекислого газа и воды на свету при участии фотосинтетических пигментов (хлорофилл у растений, бактериохлорофилл и бактериородопсин у бактерий).
В современной физиологии растений под фотосинтезом чаще понимается фотоавтотрофная функция — совокупность процессов поглощения, превращения и использования энергии квантов света в различных эндэргонических реакциях, в том числе превращения углекислого газа в органические вещества.Типы фотосинтеза
Бесхлорофилльный фотосинтез
Осуществляется археями рода Halobacterium, является наиболее примитивным типом фотосинтеза, кванты света поглощаются белком-бактериородопсином, имеющим сходство с родопсином в виде наличия ретиналя, этот тип фотосинтеза отличается отсутствием электрон-транспортной цепи, синтез АТФ осуществляется через создание электрохимического градиента протонов или ионов хлора при помощи бактериородопсиновой и галородопсиновой ионной помпы.
Хлорофилльный фотосинтез
Аноксигенный
Осуществляется пурпурными и зелёными бактериями, а также геликобактериями.
Оксигенный
Оксигенный фотосинтез распространён гораздо шире. Осуществляется растениями, цианобактериями и прохлорофитами.
Этапы фотосинтеза:
- фотофизический;
- фотохимический;
- химический.
На первом этапе происходит поглощение квантов света пигментами, их переход в возбуждённое состояние и передача энергии к другим молекулам фотосистемы. На втором этапе происходит разделение зарядов в реакционном центре, перенос электронов по фотосинтетической электронотранспортной цепи, что заканчивается синтезом АТФ и НАДФН. Первые два этапа вместе называют светозависимой стадией фотосинтеза. Третий этап происходит уже без обязательного участия света и включает в себя биохимические реакции синтеза органических веществ с использованием энергии, накопленной на светозависимой стадии. Чаще всего в качестве таких реакций рассматривается цикл Кальвина и глюкогенез, образование сахаров и крахмала из углекислого газа воздуха.
Первые процессы фотосинтеза у цианобактерий появились ещё в архейскую эру.
Пространственная локализация
 Хлоропласты в клетках листа
Хлоропласты в клетках листа Фотосинтез растений осуществляется в хлоропластах: обособленных двухмембранных органеллах клетки. Хлоропласты могут быть в клетках плодов, стеблей, однако основным органом фотосинтеза, анатомически приспособленным к его ведению, является лист. В листе наиболее богата хлоропластами ткань палисадной паренхимы. У некоторых суккулентов с вырожденными листьями (например, кактусы) основная фотосинтетическая активность связана со стеблем.
Свет для фотосинтеза захватывается более полно благодаря плоской форме листа, обеспечивающей большое отношение поверхности к объёму. Вода доставляется из корня по развитой сети сосудов (жилок листа). Углекислый газ поступает отчасти посредством диффузии через кутикулу и эпидермис, однако большая его часть диффундирует в лист через устьица и по листу по межклеточному пространству. Растения, осуществляющие CAM фотосинтез, сформировали особые механизмы для активной ассимиляции углекислого газа.
Внутреннее пространство хлоропласта заполнено бесцветным содержимым (стромой) и пронизано мембранами (ламеллами), которые соединяясь друг с другом образуют тилакоиды, которые в свою очередь группируются в стопки, называемые граны. Внутритилакоидное пространство отделено и не сообщается с остальной стромой, предполагается также что внутреннее пространство всех тилакоидов сообщается между собой. Световые стадии фотосинтеза приурочены к мембранам, автотрофная фиксация CO2 происходит в строме.
В хлоропластах имеются свои ДНК, РНК, рибосомы (70s типа), идёт синтез белка (хотя этот процесс и контролируется из ядра). Они не синтезируются вновь, а образуются путём деления предшествующих. Всё это позволило считать их потомками свободных цианобактерий, вошедших в состав эукариотической клетки в процессе симбиогенеза.
Цианобактерии, таким образом, как бы сами являются хлоропластом и в их клетке фотосинтетический аппарат не вынесен в особую органеллу. Их тилакоиды, однако, не образуют стопок, а формируют различные складчатые структуры (у единственной цианобактерии Gloeobacter violaceus тилакоиды отсутствуют вовсе, а весь фотосинтетический аппарат находится в цитоплазматической мембране, не образующей впячиваний). У них и растений также есть различия в светособирающем комплексе (см. ниже) и пигментном составе.
Световая (светозависимая) стадия
В ходе световой стадии фотосинтеза образуются высокоэнергетические продукты: АТФ, служащий в клетке источником энергии, и НАДФН, использующийся как восстановитель. В качестве побочного продукта выделяется кислород. В общем, роль световых реакций фотосинтеза заключается в том, что в световую фазу синтезируются молекула АТФ и молекулы-переносчики протонов, то есть НАДФ Н2.
Фотохимическая суть процесса
Хлорофилл имеет два уровня возбуждения (с этим связано наличие двух максимумов на спектре его поглощения): первый связан с переходом на более высокий энергетический уровень электрона системы сопряжённых двойных связей, второй — с возбуждением неспаренных электронов азота и магния порфиринового ядра. При неизменном спине электрона формируются синглетные первое и второе возбуждённое состояние, при изменённом — триплетное первое и второе.
Второе возбуждённое состояние наиболее высокоэнергетично, нестабильно и хлорофилл за 10-12 сек переходит с него на первое, с потерей 100 кДж/моль энергии только в виде теплоты. Из первого синглетного и триплетного состояний молекула может переходить в основное с выделением энергии в виде света (флуоресценция и фосфоресценция соответственно) или тепла, с переносом энергии на другую молекулу, либо, поскольку электрон на высоком энергетическом уровне слабо связан с ядром, с переносом электрона на другое соединение.
Первая возможность реализуется в светособирающих комплексах, вторая — в реакционных центрах, где переходящий в возбуждённое состояние под воздействием кванта света хлорофилл становится донором электрона (восстановителем) и передаёт его на первичный акцептор. Чтобы предотвратить возвращение электрона на положительно заряженный хлорофилл, первичный акцептор передаёт его вторичному. Кроме того, время жизни полученных соединений выше, чем у возбуждённой молекулы хлорофилла. Происходит стабилизация энергии и разделения зарядов. Для дальнейшей стабилизации вторичный донор электронов восстанавливает положительно заряженный хлорофилл, первичным донором же является в случае оксигенного фотосинтеза вода.
Проблемой, с которой сталкиваются при этом проводящие оксигенный фотосинтез организмы, является различие окислительно-восстановительных потенциалов воды (для полуреакции h3O → O2 (E0=+0,82 В) и НАДФ+ (E0=-0,32 В). Хлорофилл при этом должен иметь в основном состоянии потенциал больший +0,82 В чтобы окислять воду, но при этом иметь в возбуждённом состоянии потенциал меньший чем −0,32 В чтобы восстанавливать НАДФ+. Одна молекула хлорофилла не может отвечать обоим требованиям. Поэтому сформировались две фотосистемы, и для полного проведения процесса необходимы два кванта света и два хлорофилла разных типов.
Светособирающие комплексы
Хлорофилл выполняет две функции: поглощения и передачи энергии. Более 90 % всего хлорофилла хлоропластов входит в состав светособирающих комплексов (ССК), выполняющих роль антенны, передающей энергию к реакционному центру фотосистем I или II. Помимо хлорофилла в ССК имеются каротиноиды, а у некоторых водорослей и цианобактерий — фикобилины, роль которых заключается в поглощении света тех длин волн, которые хлорофилл поглощает сравнительно слабо.
Передача энергии идёт резонансным путём (механизм Фёрстера) и занимает для одной пары молекул 10−10—10−12 с, расстояние на которое осуществляется перенос составляет около 1 нм. Передача сопровождается некоторыми потерями энергии (10 % от хлорофилла a к хлорофиллу b, 60 % от каротиноидов к хлорофиллу), из-за чего возможна только от пигмента с максимумом поглощения при меньшей длине волны к пигменту с большей. Именно в таком порядке взаимно локализуются пигменты ССК, причём наиболее длинноволновые хлорофиллы находятся в реакционных центрах. Обратный переход энергии невозможен.
ССК растений расположен в мембранах тилакоидов, у цианобактерий основная его часть вынесена за пределы мембран в прикреплённые к ним фикобилисомы — палочковидные полипептидно-пигментные комплексы, в которых находятся различные фикобилины: на периферии фикоэритрины (с максимумом поглощения при 495—565 нм), за ними фикоцианины (550—615 нм) и аллофикоцианины (610—670 нм), последовательно передающие энергию на хлорофилл a (680—700 нм) реакционного центра.
Основные компоненты электронтранспортной цепи
Основная статья: Цепь переноса электронов фотосинтеза
Фотосистема II
Фотосистема — совокупность ССК, фотохимического реакционного центра и переносчиков электрона. Светособирающий комплекс II содержит 200 молекул хлорофилла a, 100 молекул хлорофилла b, 50 молекул каротиноидов и 2 молекулы феофитина. Реакционный центр фотосистемы II представляет собой пигмент-белковый комплекс, расположенный в тилакоидных мембранах и окружённый ССК. В нём находится димер хлорофилла a с максимумом поглощения при 680 нм (П680). На него в конечном счёте передаётся энергия кванта света из ССК, в результате чего один из электронов переходит на более высокое энергетическое состояние, связь его с ядром ослабляется и возбуждённая молекула П680 становится сильным восстановителем (E0=-0,7 В).
П680 восстанавливает феофитин, в дальнейшем электрон переносится на хиноны, входящие в состав ФС II и далее на пластохиноны, транспортируемые в восстановленной форме к b6f комплексу. Одна молекула пластохинона переносит 2 электрона и 2 протона, которые берутся из стромы.
Заполнение электронной вакансии в молекуле П680 происходит за счёт воды. В состав ФС II входит водоокисляющий комплекс, содержащий в активном центре ионы марганца в количестве 4 штук. Для образования одной молекулы кислорода требуется две молекулы воды, дающие 4 электрона. Поэтому процесс проводится в 4 такта и для его полного осуществления требуется 4 кванта света. Комплекс находится со стороны внутритилакоидного пространства и полученные 4 протона выбрасываются в него.
Таким образом, суммарный результат работы ФС II — это окисление 2 молекул воды с помощью 4 квантов света с образованием 4 протонов во внутритилакоидном пространстве и 2 восстановленных пластохинонов в мембране.
b6f или b/f-комплекс
b6f комплекс является насосом, перекачивающим протоны из стромы во внутритилакоидное пространство и создающий градиент их концентрации за счёт выделяющейся в окислительно-восстановительных реакциях электронтранспортной цепи энергии. 2 пластохинона дают перекачку 4 протонов. В дальнейшем трансмембранный протонный градиент (pH стромы около 8, внутритилакоидного пространства — 5) используется для синтеза АТФ трансмембранным ферментом АТФ-синтазой.
Фотосистема I
Светособирающий комплекс I содержит примерно 200 молекул хлорофилла.
В реакционном центре первой фотосистемы находится димер хлорофилла a с максимумом поглощения при 700 нм (П700). После возбуждения квантом света он восстанавливает первичный акцептор — хлорофилл a, тот — вторичный (витамин K1 или филлохинон), после чего электрон передаётся на ферредоксин, который и восстанавливает НАДФ с помощью фермента ферредоксин-НАДФ-редуктазы.
Белок пластоцианин, восстановленный в b6f комплексе, транспортируется к реакционному центру первой фотосистемы со стороны внутритилакоидного пространства и передаёт электрон на окисленный П700.
Циклический и псевдоциклический транспорт электрона
Помимо полного нециклического пути электрона, описанного выше, обнаружены циклический и псевдоциклический.
Суть циклического пути заключается в том, что ферредоксин вместо НАДФ восстанавливает пластохинон, который переносит его назад на b6f комплекс. В результате образуется больший протонный градиент и больше АТФ, но не возникает НАДФН.
При псевдоциклическом пути ферредоксин восстанавливает кислород, который в дальнейшем превращается в воду и может быть использован в фотосистеме II. При этом также не образуется НАДФН.
Темновая стадия
В темновой стадии с участием АТФ и НАДФН происходит восстановление CO2 до глюкозы (C6h22O6). Хотя свет не требуется для осуществления данного процесса, он участвует в его регуляции.
С3-фотосинтез, цикл Кальвина
Цикл Кальвина или восстановительный пентозофосфатный цикл состоит из трёх стадий:
- карбоксилирования;
- восстановления;
- регенерация акцептора CO2.
На первой стадии к рибулозо-1,5-бифосфату присоединяется CO2 под действием фермента рибулозобисфосфат-карбоксилаза/оксигеназа. Этот белок составляет основную фракцию белков хлоропласта и предположительно наиболее распространённый фермент в природе. В результате образуется промежуточное неустойчивое соединение, распадающееся на две молекулы 3-фосфоглицериновой кислоты (ФГК).
Во второй стадии ФГК в два этапа восстанавливается. Сначала она фосфорилируется АТФ под действием фосфороглицерокиназы с образованием 1,3-дифосфоглицериновой кислоты (ДФГК), затем при воздействии триозофосфатдегидрогеназы и НАДФН ацил-фосфатная группа ДФГК дефосфорилируется и восстанавливается до альдегидной и образуется глицеральдегид-3-фосфат — фосфорилированный углевод (ФГА).
В третьей стадии участвуют 5 молекул ФГА, которые через образование 4-, 5-, 6- и 7-углеродных соединений объединяются в 3 5-углеродных рибулозо-1,5-бифосфата, для чего необходимы 3АТФ.
Наконец, две ФГА необходимы для синтеза глюкозы. Для образования одной её молекулы требуется 6 оборотов цикла, 6 CO2, 12 НАДФН и 18 АТФ.
С4-фотосинтез
Основные статьи: Цикл Хетча-Слэка-Карпилова, С4-фотосинтез
При низкой концентрации растворённого в строме CO2 рибулозобифосфаткарбоксилаза катализирует реакцию окисления рибулозо-1,5-бифосфата и его распад на 3-фосфоглицериновую кислоту и фосфогликолевую кислоту, которая вынужденно используется в процессе фотодыхания.
Для увеличения концентрации CO2 растения С4 типа изменили анатомию листа. Цикл Кальвина у них локализуется в клетках обкладки проводящего пучка, в клетках мезофилла же под действием ФЕП-карбоксилазы фосфоенолпируват карбоксилируется с образованием щавелеуксусной кислоты, которая превращается в малат или аспартат и транспортируется в клетки обкладки, где декарбоксилируется с образованием пирувата, возвращаемого в клетки мезофилла.
С4 фотосинтез практически не сопровождается потерями рибулозо-1,5-бифосфата из цикла Кальвина, поэтому более эффективен. Однако он требует не 18, а 30 АТФ на синтез 1 молекулы глюкозы. Это оправдывает себя в тропиках, где жаркий климат требует держать устьица закрытыми, что препятствует поступлению CO2 в лист, а также при рудеральной жизненной стратегии.
САМ фотосинтез
При CAM (англ. Crassulaceae acid metabolism — кислотный метаболизм толстянковых) фотосинтезе происходит разделение ассимиляции CO2 и цикла Кальвина не в пространстве как у С4, а во времени. Ночью в вакуолях клеток по аналогичному вышеописанному механизму при открытых устьицах накапливается малат, днём при закрытых устьицах идёт цикл Кальвина. Этот механизм позволяет максимально экономить воду, однако уступает в эффективности и С4, и С3. Он оправдан при стресстолерантной жизненной стратегии.
Значение фотосинтеза
Фотосинтез является основным источником биологической энергии, фотосинтезирующие автотрофы используют её для синтеза органических веществ из неорганических, гетеротрофы существуют за счёт энергии, запасённой автотрофами в виде химических связей, высвобождая её в процессах дыхания и брожения. Энергия, получаемая человечеством при сжигании ископаемого топлива (уголь, нефть, природный газ, торф), также является запасённой в процессе фотосинтеза.
Фотосинтез является главным входом неорганического углерода в биологический цикл. Весь свободный кислород атмосферы — биогенного происхождения и является побочным продуктом фотосинтеза. Формирование окислительной атмосферы (кислородная катастрофа) полностью изменило состояние земной поверхности, сделало возможным появление дыхания, а в дальнейшем, после образования озонового слоя, позволило жизни выйти на сушу.
История изучения фотосинтеза
Первые опыты по фотосинтезу были проведены Джозефом Пристли в 1770—1780-х годах, когда он обратил внимание на «порчу» воздуха в герметичном сосуде горящей свечой (воздух переставал быть способен поддерживать горение, помещённые в него животные задыхались) и «исправление» его растениями. Пристли сделал вывод, что растения выделяют кислород, который необходим для дыхания и горения, однако не заметил, что для этого растениям нужен свет. Это показал вскоре Ян Ингенхауз.
Позже было установлено, что помимо выделения кислорода растения поглощают углекислый газ и при участии воды синтезируют на свету органическое вещество. В 1842 Роберт Майер на основании закона сохранения энергии постулировал, что растения преобразуют энергию солнечного света в энергию химических связей. В 1877 В. Пфеффер назвал этот процесс фотосинтезом.
Хлорофиллы были впервые выделены в 1818 П. Ж. Пельтье и Ж. Кавенту. Разделить пигменты и изучить их по отдельности удалось М. С. Цвету с помощью созданного им метода хроматографии. Спектры поглощения хлорофилла были изучены К. А. Тимирязевым, он же, развивая положения Майера, показал, что именно поглощенные лучи позволяют повысить энергию системы, создав вместо слабых связей С-О и О-Н высокоэнергетические С-С (до этого считалось что в фотосинтезе используются жёлтые лучи, не поглощаемые пигментами листа). Сделано это было благодаря созданному им методу учёта фотосинтеза по поглощённому CO2: в ходе экспериментов по освещению растения светом разных длин волн (разного цвета) оказалось, что интенсивность фотосинтеза совпадает со спектром поглощения хлорофилла.
Окислительно-восстановительную сущность фотосинтеза (как оксигенного, так и аноксигенного) постулировал Корнелис ван Ниль. Это означало, что кислород в фотосинтезе образуется полностью из воды, что экспериментально подтвердил в 1941 А. П. Виноградов в опытах с изотопной меткой. В 1937 г. Роберт Хилл установил, что процесс окисления воды (и выделения кислорода), а также ассимиляции CO2 можно разобщить. В 1954—1958 Д. Арнон установил механизм световых стадий фотосинтеза, а сущность процесса ассимиляции CO2 была раскрыта Мельвином Кальвином с использованием изотопов углерода в конце 1940-х, за эту работу в 1961 ему была присуждена Нобелевская премия.
В 1955 году был выделен и очищен фермент рибулозобисфосфат-карбоксилаза/оксигеназа. С4 фотосинтез был описан Ю. С. Карпиловым в 1960 и М. Д. Хэтчем и К. Р. Слэком в 1966.
Прочие факты
- Морской слизень Elysia chlorotica ассимилирует хлоропласты водоросли Vaucheria litorea в клетки пищеварительного тракта. Хлоропласты способны фотосинтезировать в организме слизня в течение нескольких месяцев, что позволяет слизню жить за счет глюкозы, полученной в результате фотосинтеза. Геном слизня кодирует некоторые белки, необходимые хлоропластам для фотосинтеза.[1]
- Предполагается, что в естественной среде фотосинтезирующие бактерии могут использовать не только свет Солнца, но и другие источники света, а потому могут находиться в местах, не подвергающихся облучению звезды[2] В 2005 году Томас Битти из университета Британской Колумбии и Роберт Блейкеншип из университета Аризоны в глубоководных пробах, взятых в окрестностях глубоководного термального источника у побережья Коста Рики обнаружили серобактерию GSB1, сходную с серобактериями родов Chlorobium и Prosthecochloris, содержащую бактериохлорофилл. Они предположили, что вероятность контаминации образца невелика и, следовательно, GSB1 использует для фотосинтеза не солнечный свет (который не проникает сквозь 2,4-километровую толщу моря), а тусклый длинноволновый (~750 нм) свет, испускаемый гидротермальными источниками[2].
- По состоянию на конец 70-х годов XX века, мощность солнечной энергии, перерабатываемой земной растительностью при фотосинтезе, всего лишь на порядок превосходила мощность всех электростанций в мире.[3]
См. также
Литература
- Холл Д., Рао К. Фотосинтез: Пер. с англ. — М.: Мир, 1983.
- Физиология растений / под ред. проф. Ермакова И. П. — М.: Академия, 2007
- Молекулярная биология клетки / Альбертис Б., Брей Д. и др. В 3 тт. — М.: Мир, 1994
- Рубин А. Б. Биофизика. В 2 тт. — М.: Изд. Московского университета и Наука, 2004.
- Чернавская Н. М., Чернавский Д. С. Туннельный транспорт электронов в фотосинтезе. М., 1977.
- В. Любименко. Влияние света на усвоение органических веществ зелёными растениями // Известия Императорской Академии наук. VI серия. — 1907. — № 12. — С. 395—426, с 6 табл.
- Медведев С. С. Физиология растений — СПб,: СПбГУ, 2004
Примечания
- ↑ Rumpho ME, Worful JM, Lee J, et al. (November 2008). «Horizontal gene transfer of the algal nuclear gene psbO to the photosynthetic sea slug Elysia chlorotica». Proc. Natl. Acad. Sci. U.S.A. 105 (46): 17867–17871. DOI:10.1073/pnas.0804968105. PMID 19004808. Проверено 2008-11-24.
- ↑ 1 2 Beatty, J. Thomas; Jörg Overmann, Michael T. Lince, Ann K. Manske, Andrew S. Lang, Robert E. Blankenship, Cindy L. Van Dover, Tracey A. Martinson, F. Gerald Plumley (2005). «An obligately photosynthetic bacterial anaerobe from a deep-sea hydrothermal vent». Proceedings of the National Academy of Sciences of the United States of America 102 (26): 9306 -9310. DOI:10.1073/pnas.0503674102. Проверено 2011-02-10.
- ↑ Углекислый газ в атмосфере земли, Б. М. Смирнов, Объединенный институт высоких температур РАН, 126 (11) (1978), Москва
Ссылки
dic.academic.ru
Фотосинтез растений
Фотосинтез - это уникальный физико-химический процесс, осуществляемый на Земле всеми зелеными растениями и некоторыми бактериями и обеспечивающий преобразование электромагнитной энергии солнечных лучей в энергию химических связей различных органических соединений. Основа фотосинтеза — последовательная цепь окислительно-восстановительных реакций, в ходе которых осуществляется перенос электронов от донора — восстановителя (вода, водород) к акцептору — окислителю (СО2, ацетат) с образованием восстановленных соединений (углеводов) и выделением O2, если окисляется вода
Фотосинтез играет ведущую роль в биосферных процессах, приводя в глобальных масштабах к образованию органического вещества из неорганического.
Фотосинтезирующие организмы, используя солнечную энергию в реакциях фотосинтеза, осуществляют связь жизни на Земле со Вселенной и определяют в конечном итоге всю ее сложность и разнообразие. Гетеротрофные организмы — животные, грибы, большинство бактерий, а также бесхлорофилльные растения и водоросли — обязаны своим существованием автотрофным организмам — растениям-фотосинтетикам, создающим на Земле органическое вещество и восполняющим убыль кислорода в атмосфере. Человечество все более осознает очевидную истину, впервые научно обоснованную К.А. Тимирязевым и В.И. Вернадским: экологическое благополучие биосферы и существование самого человечества зависит от состояния растительного покрова нашей планеты.
Процессы, происходящие в листе
Лист осуществляет три важных процесса – фотосинтез, испарение воды и газообмен. В процессе фотосинтеза в листьях из воды и двуокиси углерода под действием солнечных лучей синтезируются органические вещества. Днем, в результате фотосинтеза и дыхания, растение выделяет кислород и двуокись углерода, а ночью – только двуокись углерода, образующуюся при дыхании.
Большинство растений способно синтезировать хлорофилл при слабом освещении. При прямом солнечном освещении хлорофилл синтезируется быстрее. Необходимая для фотосинтеза световая энергия в известных пределах поглощается тем больше, чем меньше затемнен лист. Потому у растений в процессе эволюции выработалась способность поворачивать пластину листа к свету так, чтобы на нее падало больше солнечных лучей. Листья на растении располагаются так, чтобы не притеснять друг друга. Тимирязев доказал, что источником энергии для фотосинтеза служат преимущественно красные лучи спектра. На это указывает спектр поглощения хлорофилла, где наиболее интенсивная полоса поглощения наблюдается в красной, и менее интенсивное – в сине-фиолетовой части.
 Фото: Nat Tarbox
Фото: Nat Tarbox
Интенсивность фотосинтеза также зависит от фазы развития растения. Максимальная интенсивность фотосинтеза наблюдается в фазе цветения. Обычное содержание углекислоты в воздухе составляет 0,03% по объему. Уменьшение содержания углекислоты в воздухе снижает интенсивность фотосинтеза. Повышение содержания углекислоты до 0,5% увеличивает интенсивность фотосинтеза почти пропорционально. Однако при дальнейшем повышении содержания углекислоты, интенсивность фотосинтеза не возрастает, а при 1% - растение страдает.
Растения испаряют или трансперируют очень большое количество воды. Испарение воды является одной из причин восходящего тока. Вследствие испарения воды растением в нем накапливаются минеральные вещества, и происходит полезное для растения понижение температуры во время солнечного нагрева. Растение регулирует процесс испарения воды посредством работы устьиц. Отложение кутикулы или воскового налета на эпидерме, образование его волосков и другие приспособления направлены к сокращению нерегулируемой трансперации.
Процесс фотосинтеза и постоянное протекающее дыхание живых клеток листа требуют газообмена между внутренними тканями листа и атмосферой. В процессе фотосинтеза из атмосферы поглощается ассимилируемый углекислый газ и возвращается в атмосферу кислородом. Применение изотопного метода анализа показало, что кислород, возвращаемый в атмосферу 16O принадлежит воде, а не углекислому газу воздуха, в котором приобладает другой его изотоп - 15О. При дыхании живых клеток (окисление свободным кислородом органических веществ внутри клетки до углекислого газа и воды) необходимо поступление из атмосферы кислорода и возвращение углекислоты. Этот газообмен также в основном осуществляется через устьичный аппарат.
Процесс фотосинтеза состоит из двух последовательных и взаимосвязанных этапов: светового (фотохимического) и темнового (метаболического). На первой стадии происходит преобразование поглощенной фотосинтетическими пигментами энергии квантов света в энергию химических связей высокоэнергетического соединения АТФ и универсального восстановителя НАДФН — собственно первичных продуктов фотосинтеза, или так называемой «ассимиляционной силы». В темновых реакциях фотосинтеза происходит использование образовавшихся на свету АТФ и НАДФН в цикле фиксации углекислоты и ее последующего восстановления до углеводов. У всех фотосинтезирующих организмов фотохимические процессы световой стадии фотосинтеза происходят в особых энергопреобразующих мембранах, называемых тилакоидными, и организованы в так называемую электрон-транспортную цепь. Темновые реакции фотосинтеза осуществляются вне тилакоидных мембран (в цитоплазме у прокариот и в строме хлоропласта у растений). Таким образом, световая и темновая стадии фотосинтеза разделены в пространстве и во времени.
Интенсивность фотосинтеза древесных растений широко варьирует в зависимости от взаимодействия многих внешних и внутренних факторов, причем эти взаимодействия изменяются во времени и различны у разных видов.
Фотосинтетическую способность иногда оценивают по чистому приросту сухой массы. Такие данные имеют особое значение, потому что прирост представляет собой среднее истинное увеличение массы за большой промежуток времени в условиях внешней среды, включающих обычные периодически наступающие стрессы. Некоторые виды покрытосеменных эффективно осуществляют фотосинтез как при низкой, так и при высокой интенсивности света. Многие голосеменные гораздо более продуктивны при высокой освещенности. Сравнение этих двух групп при низкой и высокой интенсивности света часто дает различное представление о фотосинтетической способности с точки зрения накопления питательных веществ. Кроме того, голосеменные часто накапливают некоторое количество сухой массы в период покоя, тогда как листопадные покрытосеменные теряют ее вследствие дыхания. Поэтому голосеменное растение с несколько более низкой интенсивностью фотосинтеза, чем листопадное покрытосеменное во время периода роста, может накапливать в течение года столько же или даже больше общей сухой массы благодаря гораздо большей продолжительности периода фотосинтетической активности.
Первые опыты по фотосинтезу были проведены Джозефом Пристли в 1770-1780-х годах, когда он обратил внимание на "порчу" воздуха в герметичном сосуде горящей свечой (воздух переставал быть способен поддерживать горение, помещённые в него животные задыхались) и "исправление" его растениями. Пристли сделал вывод что растения выделяют кислород, который необходим для дыхания и горения, однако не заметил что для этого растениям нужен свет. Это показал вскоре Ян Ингенхауз. Позже было установлено что помимо выделения кислорода растения поглощают углекислый газ и при участии воды синтезируют на свету органическое вещество. В 1842 Роберт Майер на основании закона сохранения энергии постулировал что растения преобразуют энергию солнечного света в энергию химических связей. В 1877 В. Пфеффер назвал этот процесс фотосинтезом.
biofile.ru










