
Семейство протейные (Proteaceae). Протейные растения
Протейные растения
Семейство протейные (Proteaceae) из порядка протейные (Proteales) представлено деревьями и кустарниками с кожистыми цельными или рассеченными очередными листьями без прилистников. Обычна гетерофиллия. Цветки в разнообразных соцветиях, в частности в головках, окруженных окрашенными в яркие цвета прицветниками, напоминающими обертку сложноцветных. Энтомофилия и орнитофилия. Известно также опыление сумчатыми млекопитающими. Цветки обоеполые, обычно правильные. Околоцветник простой 4-членный, венчиковидный, в нижней части образующий более или менее длинную трубку. Лопасти обычно закручены назад. Тычинок 4, супротивных листочкам околоцветника, тычиночные нити, приросшие к трубке околоцветника. Завязь верхняя, 1-гнездная, часто на ножке. Стилодий 1, длинный. При основании завязи нектароносный диск. Семязачатки весьма различны по количеству и строению. Интегументов 2, в некоторых случаях 3. Плоды вскрывающиеся и невскрывающиеся, сухие и сочные.
Семена крылатые. Семядоли различны по величине, иногда их более 2 (до 8). В эмбриогенезе нередко образование эндоспермальных гаусториев. 60 родов и около 1400 видов в сухих областях Южного полушария, в особенности в Австралии и Южной Африке. Немногие роды проникли в тропики.
Протейные - характерное семейство флоры Южной Африки (так называемой капской флоры) и субтропических районов Австралии. У них вырабатываются различные приспособления к перенесению засушливого климата. У видов австралийского рода хакея (Hakea) боковые сегменты перисто-рассеченных листьев превращаются в колючие зубцы, округлые в поперечном сечении, с всесторонне развитой палисадной тканью. В палисадной ткани содержатся остеобласты. Губчатая паренхима отсутствует. Вообще, для большинства протейных характерны жестколистность и резко выраженная ксероморфная структура. Многие виды - пирофиты - растения, приспособившиеся к пожарам, часто вспыхивающим в различных районах Австралии. У них, в частности, выражена способность к образованию обильной корневой поросли. Дело доходит до того, что листовки у видов рода банксия (Banksia), обладающие чрезвычайно твердым околоплодником, открываются лишь после сильного нагревания (вспомним сосну Банкса).
 Roupala montana. Фото: Joao Medeiros
Roupala montana. Фото: Joao Medeiros
 Alloxylon flammeum. Фото: Tatiana Gerus
Alloxylon flammeum. Фото: Tatiana Gerus
Протейные охотно разводят в прохладных оранжереях, а в сухих субтропиках - в открытом грунте, поскольку их соцветия и листва часто декоративны. В качестве примера можно привести южноафриканское серебряное дерево (Leucadendron argenteum), отличающееся обильной листвой особенного серебристо-белого цвета. В сухих тропиках и субтропиках часто разводят и австралийскую гревиллею (Grewillea) с характерным образом рассеченными листьями, доставляющую исключительно ценную древесину. Соцветия многих протейных особенно эффектны благодаря почти одновременному распусканию цветков.
Протейные - изолированно стоящее семейство. Для выяснения родственных связей очень важна интерпретация нектарных чешуек, расположенных при основании завязи. Раньше их считали за остатки второго круга околоцветника, и таким образом цветок выглядел как вторично упрощенный. Сейчас нектарные чешуйки или диск чаще считают новообразованиями, адаптировавшими цветки к зоофилии.
Самым замечательным растением своего континента после эвкалиптов и акаций австралийцы считают телопею. Она тоже из протейных. Но не всякий австралиец видел живую телопею. Браконьеры уничтожили телопею в лесах так основательно, что она встречается редко.
За сто миль от Сиднея нашли редкий эвкалиптовый лес, недавно горевший. Забрались на скалу, с которой видны окрестные холмы, и оттуда увидели вдали темно- красную точку. Телопея! Ростом она оказалась метра четыре. Стебель прямой, как у иван-чая. И так же обрамлен листьями. На верхушке крупное, как кочан, красное соцветие. Но росла одиноко.
В Южной Африке есть другой уникум из протейных — серебряное дерево, леукадендрон серебряный. Растет по прибрежным холмам. Когда с моря набегает легкий бриз, длинные листочки леукадендрона, покрытые шелковистыми волосками, блестят, как серебряные. У леукадемдрона все необычно. Не только листья. Ствол снизу доверху покрыт рубцами от старых опавших листьев. У двудольных деревьев такой феномен — явление редкое. Эти рубцы, как у пальм, не зарастают и не исчезают даже у очень старых деревьев.
Женские цветки собраны в соцветия-головки размером с небольшое яблоко. Когда отцветут, соцветие превращается в подобие шишки. Глубоко под чешуями упрятаны плодики, в каждом — по одному семечку. Снаружи, как перья, торчат только остатки от венчиков — по четыре штуки крест-накрест. Ветер дунет, перистый хохолок надуется, как вытяжной парашютик, и извлечет плодик из камеры. И потом, качаясь под перистым куполом, он поплывет по воздушным волнам.
Самый видный представитель протейных — протей цинаровый, или «сахарный буш». Деревце маленькое, намного меньше других протейных. Зато соцветие крупное, как корзинка подсолнуха, и даже чуточку похоже по форме. Красоты необычайной. Внутри снежно-белые бутоны. По периферии золотистые цветки. Все это сооружение оторочено крупными прилистниками, снаружи белыми, внутри металлически-красными. Нектара дает столько, что, наклонив его 20-сантиметровую корзинку, можно нацедить полную ложку.
Неожиданно для всех африканцев протей стал страдать от натиска своей родственницы хакеи. Страдают протеи и от огня, те, что имеют тонкую кору. Самое забавное деревце из протейных — ксиломелюм грушевидный. Выделяется среди сородичей формой плодов. Они довольно точная копия груш, только прикрепляются к плодоножкам не острым, а тупым концом, так сказать, задом наперед. Груши совершенно деревянистые и, конечно, несъедобные.
Основные растения семейства:
Протея «Кинг» Сорт с крупным головчатым соцветием, листочки обертки сиреневато-розовые, заостренные, многорядные.
Протея «Финус» Соцветие розоватое, слегка вытянутое, хорошо смотрится в зимних букетах, придавая им оригинальность и нежность.
Леукоспермум «Танго» Яркое, броское соцветие; сочные краски, устойчивость в срезке делают это растение одним из самых привлекательных при создании парадных букетов.
Протея «Блэк Коун» Флористы используют цветонос со следами от опавших соплодий, украсив его другими растениями, например цветками орхидей.
Протея ползучая Один из самых нетребовательных видов, отличается вытянутым желто-красным соцветием и узкими сизыми листьями.
Банксия Бакстера Вид с жесткими, сизоватыми зубчатыми листьями и коротким колосовидным соцветием. Эта серебристо-серая банксия обычно используется в рождественской аранжировке.
Протея «Сильвия» Сорт с крупным розовым соцветием, укрытым овальными жесткими листьями. Хорошо смотрится в букетах.
Леукадендрон ивовый Эффектный кустарник с темно-бордовыми плотными ланцетными листьями, особенно привлекателен в зимний период.
Протея олеандровая Розово-красное соцветие, листочки-обертки овальные, на верхушке темно-красные, что придает ему некоторую мрачноватость.
biofile.ru
Протейные — WiKi
Распространение
 Ксиломелюм Xylomelum
Ксиломелюм Xylomelum Подавляющее большинство видов — обитатели сухих субтропических районов, лишь немногие виды произрастают в тропических дождевых лесах (виды родов Roupala, Helicia, Гревиллея).
Все виды семейства произрастают почти исключительно в Южном полушарии.
В Австралии и Тасмании около 800 видов. Особенно здесь характерны представители родов Банксия, Hakea, Dryandra, Гревиллея. Второй центр распространения протейных лежит в Капской области (около 400 видов). Здесь наиболее часто встречаются представители родов Протея и Leucadendron. Отдельные роды проникают на север до Эфиопии. Так же протейные многочисленны в Новой Каледонии. Относительно небольшое количество видов произрастает на Мадагаскаре, в Южной Индии, Шри-Ланке, Восточной и Юго-восточной Азии, Новой Гвинее, Новой Зеландии и на островах Тихого океана. Несколько видов обитают в Центральной и Южной Америке.
Дальше всего на север проникает Helicia lancifolia[3].
Биологическое описание
Листья, как правило, очередные, без прилистников, отличаются крайним разнообразием формы и размеров — от игольчатых и шиловидных, плоских цельных цельнокрайних и зубчатых до глубоко многократно рассечённых. Вариабельность листьев часто проявляется в пределах одного вида и в зависимости от возраста побегов — одного и того же экземпляра.
Цветки, сидящие по 1 или чаще 2 в пазухах брактей (прицветников), собраны в кисти, колосья, реже зонтики или головчатые соцветия. Соцветия часто окружены обёрткой из многочисленных ярко окрашенных стерильных прицветников или верхушечных листьев и напоминают соцветия сложноцветных, а после созревания плодов шишку голосеменных.
Цветки обоеполые (раздельнополые у представителей рода Leucadendron), правильные или зигоморфные, 4-членные с простым венчиковидным сростнолистным околоцветником. Тычинок 4. Гинецей образован единственным плодолистиком.
Плод — листовка, коробочка, орешек или костянка. Семена многочисленные или 1—2, как правило без эндосперма, часто крылатые. Плоды и семена у многих видов отличаются исключительной жароустойчивостью. Плоды некоторых видов раскрываются только под воздействием высоких температур, что является адаптацией к сезонным пожарам[3].
Подсемейства
Список родов
|
|
В культуре
В коллекциях ботанических садов протейные — сравнительно редкая группа. В ГБС коллекцию протейных начали собирать с конца 60-х годов XX века. В 1991 году в коллекции насчитывалось более 120 видов, относящихся к 18 родам. Большая часть растений выращена из семян полученных из ботанических садов Австралии, Южной Африки и Европы. Гревиллея крупная и Stenocarpus sinuatus получены в 1948 году из ГДР[3].
Большая часть культивируемых в комнатах и оранжереях протейних относятся к холодной и умеренной температурной группе. Размножение семенами и черенками[4].
Галерея
Примечания
- ↑ Об условности указания класса двудольных в качестве вышестоящего таксона для описываемой в данной статье группы растений см. раздел «Системы APG» статьи «Двудольные».
- ↑ 1 2 Proteaceae по данным сайта The Plant List (англ.) (лат.) (Проверено 25 августа 2014)
- ↑ 1 2 3 Порубиновская Г. В. Определитель видов семейства Proteaceae Juss. Оранжерейные растения: Таблицы для определения: Сб. науч. ст. М.:Наука, 1991. — 256 с.
- ↑ Сааков С. Г. Оранжерейные и комнатные растения и уход за ними. — Москва «Наука», 1985
Литература
Ссылки
ru-wiki.org
Семейство протейные (Proteaceae)
Семейство протейных, насчитывающее более 70 родов и, вероятно, не менее 1400 видов, распространено главным образом в южном полушарии. Наибольшая концентрация протейных наблюдается в Южной Африке (преимущественно в Капском флористическом царстве), где встречается 14 родов и почти 400 видов, и особенно в Австралии вместе с Тасманией (45 родов и около 800 видов). Однако ареал семейства гораздо шире и охватывает тропическую Африку от Эфиопии на севере, Мадагаскар, Южную Индию и остров Шри-Ланка, Восточную и Юго-Восточную Азию, Новую Гвинею, Новую Каледонию, Новую Зеландию и многие острова Тихого океана. Из всех протейных дальше всех на север заходят некоторые виды рода хелиция (Helicia, рис. 180), ареал которого простирается от Индокитая и Тайваня до Японии (остров Хонсю).
Очень велика роль протейных в растительном мире Капского флористического царства, где они поражают множеством форм, разнообразием листьев и исключительной красотой соцветий. Наиболее характерны виды левкадендрона (Leucadendron) и протеи (Protea, табл.45). Значительно меньшую роль играют протейные в тропической Африке, но и здесь они местами принимают заметное участие в растительности, например в Северной Гвинее. Довольно заметную роль играют протейные на Мадагаскаре. Но наиболее характерны они для растительного мира Австралии, особенно для Юго-Западной Австралии, Квинсленда, Нового Южного Уэльса и Тасмании. Здесь бросаются в глаза многочисленные виды гревиллеи (Grevillea), виды эндемичного рода банксия (Banksia, табл. 46, рис. 181) и многие другие. В Юго-Западной Австралии обычны виды эндемичного рода дриандра (Dryandra, рис. 182), так же как виды австралийско-тасманийского рода хакея (Hakea, табл. 46 и 47). Значительна роль протейных также в растительном мире Новой Каледонии. Что касается Азии и Америки, то протейные играют там менее заметную роль.
Протейные чаще всего кустарники и кустарнички, реже деревья, иногда очень высокие. Есть среди них также полукустарники и даже корневищные травы. Для многих протейных характерна жестколистность. Нередки среди них относительно мелколистные формы. Весь облик большинства протейных говорит о приспособлении к временной сухости муссонного климата и к бедным почвам. Как у южноафриканских, так и у австралийских протейных часто имеются так называемые лигнотуберусы — деревянистые подземные запасающие органы стеблевого происхождения. Однако в дождевых лесах встречаются деревья и с более или менее мезоморфными листьями. Протейные ясно выраженного жестколистного характера чаще всего кустарники, но бывают среди них и деревья высотой от 6 до 20 м. Очень многие типично жестколистные виды произрастают на почвах, бедных питательными веществами, особенно фосфором. Листья протейных характеризуются исключительно широким размахом изменчивости как размеров, формы и строения, так и типов жилкования. Не случайно Линней назвал род протея именем Протея, который, согласно древнегреческому мифу, мог по желанию обрести любую форму и любой образ.
Цветки протейных обычно в различного рода бокоцветных соцветиях: кистях, зонтиках, колосьях или головках (простых или сложных), редко одиночные в пазухах листьев (результат редукции соцветия). Очень характерна ясно выраженная в семействе тенденция к образованию головчатых соцветий, часто внешне удивительно напоминающих соцветия многих сложноцветных. Головчатые соцветия возникают независимо у разных представителей семейства. Особенно характерны такие соцветия для рода протея в Южной и тропической Африке и родов телопеа (Telopea), дриандра и других в Австралии. Цветки протейных актиноморфные или более или менее зигоморфные, обычно обоеполые, но иногда однополые и у некоторых родов, как, например, у левкадендрона, двудомные. Околоцветник представлен венчиковидной, обычно сростнолистной, 4-члениой чашечкой. Интересно, что в бутоне чашечка обычно трубчатая, но при распускании цветка чашелистики более или менее отделяются друг от друга и в той или иной степени загибаются книзу, а иногда закручиваются. Что касается венчика, то он редуцирован, хотя у разных родов в неодинаковой степени. Вместо нормально развитого венчика в цветке обычно имеется более или менее развитый 4-членный подпестичный круг желёзок или чешуек или же чашевидный диск, как правило, несущие секреторную функцию. Другими словами, лепестки у протейных в той или иной степени превращены в нектарники. В то время как в актиноморфных цветках можно видеть полный 4-членный круг желёзок, каждая из которых соответствует лепестку, в зигоморфных цветках круг этот неполный: абаксиальные желёзки исчезают и сохраняются только 3 или даже только 2 адаксиальные желёзки. Иногда этот процесс идет дальше, и в цветке не сохраняется никаких следов венчика. У австралийского рода франкландия (Franklandia) сохранилось еще определенное сходство с венчиком, но 4 лопасти его сростнолепестного венчика уже функционируют как нектарники. В тех случаях, когда лепестки сведены к желёзкам и чешуйкам, эти последние обычно более или менее сросшиеся между собой, что, возможно, свидетельствует о том, что венчик у протейных был сростнолепестный. Тычинок всегда 4, чередующихся с чашелистиками, с широкими нитями, обычно более или менее (иногда почти полностью) приросшими к чашелистикам. Лишь редко нити бывают полностью свободными, как у относительно примитивного монотипного тасманийского рода беллендена (Bellendena). Пыльники иногда с ясно выраженным надсвязником, особенно хорошо развиты у монотипного новокаледонского рода бопреопсис (Beaupreopsis). Гинецей одноплодолистиковый, но, несмотря на общую довольно высокую специализированность цветка протейных, плодолистик относительно примитивный, еще в сущности кондупликатный. У некоторых родов, имеющих рудиментарный венчик, плодолистик на более ранней стадии своего развития еще не вполне замкнутый и в нижней своей части (в области завязи), а также в районе рыльца еще более или менее открытый. У некоторых протейных свободные брюшные поверхности несут железистые волоски, как это наблюдается у кондупликатных плодолистиков некоторых примитивных цветковых растений. Однако в строении плодолистика протейных ясно выражены также определенные признаки довольно высокой специализации. Столбик нитевидный и прямой или же согнутый, у большинства видов с различной формы косым или боковым рыльцем. Функционирующая рыльцевая поверхность может быть ограничена маленькой точкой на верхушке столбика или в центре верхушечного диска. У многих протейных с наличием рудиментарного венчика завязь на ножке (гинофоре), что характерно для орнитофильных видов. Семязачатков в каждом плодолистике от одного до многих. Плод — листовка, орешек, крылатка или костянка. За исключением рода боллендена, семена без эндосперма.
Цветки протейных обычно протандричны. Опыление производится различными представителями животного мира — насекомыми, птицами (нектарниками, белоглазками, австралийско-новогвинейскими медососами и родственными им южноафриканскими сахарными птицами и американскими колибри), рукокрылыми (крыланами и летучими мышами) и мелкими нелетающими млекопитающими (грызунами, мелкими сумчатыми из семейства лазящих сумчатых, или кускусов, — Phalangeridae).
У протейных наблюдается довольно большое разнообразие приспособлений к перекрестному опылению, особенно хорошо изученное у австралийских протейных (Р. Каролин, 1908). У некоторых из них механизм опыления относительно простой, как, например, у австралийской персоонии игольчатой (Persoonia acerosa), цветки которой посещают главным образом пчелы. В только что раскрывшемся цветке столбик с еще незрелым рыльцем окружен тесно расположенными вокруг него длинными пыльниками, которые вскоре вскрываются продольными щелями. Когда насекомое просовывает свой хоботок между пыльниками и столбиком, часть пыльцы откладывается на хоботке и теле. На следующей женской фазе развития цветка пыльники закручиваются назад, а рыльце становится клейким и готово к восприятию пыльцы. Иной механизм опыления наблюдается у другого австралийского протейного — изопогона укрополистного (Isopogon anethifolius). Пыльники изопогона также тесно прижаты к столбику, но, в отличие от персоонии, они открываются еще в бутоне. Непосредственно под пыльниковым каналом на столбике имеется вздутие, преграждающее пыльцу от падения вниз. Затем сегменты околоцветника вместе с прикрепленными к ним тычинками закручиваются назад, оставляя в центре цветка покрытый пыльцой свободно возвышающийся столбик. Таким образом, в отличие не только от персоонии, но и от подавляющего большинства цветковых растений, подача пыльцы опылителю осуществляется не самими пыльниками, а через посредство столбика. Эту же модель, но в более или менее видоизмененной форме, мы находим и у целого ряда других родов протейных. Так, у австралийского рода ламбертия (Lambertia) пыльца откладывается в бутоне на клейкой верхней части столбика. У другого австралийского протейного — петрофилы окрашенной (Petrophila fucifolia) — оседанию пыльцы на столбике способствуют покрывающие ее короткие волоски, представляющие собой, таким образом, аналог «пыльцеснимателя» колокольчиковых (см. ниже, семейство колокольчиковых). У всех этих растений рыльце созревает и бывает готово к восприятию пыльцы лишь после окончания мужской фазы, когда большая часть пыльцы уже выгружена опылителями или засохла.
У гревиллеи, хакеи, телопей, ломатии и некоторых других родов механизм опыления несколько иной. Верхушка столбика у них уплощена и расширена в форме прямого, косого или бокового диска, в центре которого в виде маленькой выпуклости расположено рыльце. Пыльники вскрываются в бутоне, и пыльца откладывается на дисковидной верхушке столбика, в том числе и на самом рыльце, которое на, этой мужской стадии еще совершенно не восприимчиво к ней; Некоторое время дисковидная верхушка столбика остается зажатой в верхней части еще полностью не раскрывшегося околоцветника. У некоторых протейных, например, у рода протеи, диск не высвобождается до тех пор, пока этой части цветка не коснется опылитель. Но уже при малейшем прикосновении сразу же срабатывает этот удивительный триггерный механизм, в котором столбик играет роль пружины изгиба, и диск с силой вырывается из объятий околоцветника и разбрасывает накопившуюся на нем пыльцу, осыпая ею тело опылителя. Благодаря этому механизму достигается двоякая цель: до высвобождения диска пыльца бывает защищена от превратностей внешнего мира, а внезапное ее высвобождение способствует разбрасыванию пыльцы. После освобождения диска начинается женская стадия — крошечное рыльце в центре диска увеличивается в размере и становится готовым к восприятию пыльцы, принесенной опылителем с другого цветка.
Совершенно иную модель механизма опыления мы наблюдаем у некоторых других родов. Особенно интересны в этом отношении австралийские роды симфионема (Symphyonema) и коноспермум (Conospermum). В мелких желтых цветках симфионемы пыльники расположены тесно вокруг столбика. Когда пыльники созревают, сегменты околоцветника закручиваются назад. В поисках нектара опылитель (в данном случае обычно бражники) обязательно коснется пыльников, которые внезапно отделяются друг от друга, разбрасывая при этом пыльцу во всех направлениях. Часть этой пыльцы попадает и на опылителя. А в крупных цветках коноспермума эллиптического (С. ellipticum) пыльники соединены вокруг столбика, причем некоторые из них заканчиваются длинной щетинкой, или остью. Часть пыльников стерильна. Столбик согнут в форме лебединой шеи с рыльцем, направленным в сторону от фертильных пыльников. Малейшее прикосновение к столбику или к щетинкам пыльников приводит к быстрому движению столбика и внезапному отделению пыльников друг от друга. Верхняя часть столбика поворачивается в сторону пустых пыльников, и рыльце с силой ударяет по опылителю (в данном случае обычно по мухе). Если на опылителе уже есть пыльца с другого цветка, то часть этой пыльцы прилипнет к рыльцу. Пыльники отделяются почти сразу же за движением столбика, поднимая маленькое облачко пыльцы, часть которой попадает на муху.
Очень многие протейные приспособлены к опылению исключительно или главным образом птицами, особенно в Австралии и Южной Африке. Птицы опыляют виды протейных с крупными красными, оранжевыми, желтыми или желтовато-зелеными, часто контрастно окрашенными верхушечными соцветиями, отдельные цветки которых обильно выделяют нектар и обычно лишены запаха, столь важного для привлечения насекомых и млекопитающих. Цветки некоторых видов банксии выделяют так много нектара, что он используется в качестве пищи аборигенами Австралии.
Взаимное приспособление протейных и птиц в некоторых случаях принимает форму взаимной зависимости. Классическим примером является южноафриканская сахарная птица промеропс (Promerops cafer), которая настолько приспособилась к опылению некоторых видов протеи, что почти не может существовать без этих растений. У сахарной птицы клюв и язык длинные и тонкие, а соцветия этих видов протеи, окруженные плотно прилегающими друг к другу листочками обертки, представляют собой как бы естественную чашу, из которой птица пьет нектар. Сахарная птица настолько сжилась с видами протеи, что даже гнезда свои строит из листочков обертки и из старых цветков, заключающих зрелые семена, и тем самым содействует диссеминации. В качестве примера рассмотрим опыление у кустарниковой протеи крупноголовой (P. macrocephala). Листочки обертки этого вида светло-зеленые, но хохолок, образованный волосками на кончиках листочков, имеет посередине красный круг. Внутренние листочки обертки длиннее наружных, и их шерстисто опушенные верхушки отогнуты внутрь, к центру соцветия, и образуют удобную посадочную площадку для птиц. Чашечка состоит из одного свободного узкого чашелистика, несущего стерильную тычинку, и трех сросшихся чашелистиков, несущих каждый по одной фертильной тычинке. Пыльники заканчиваются маленьким ярко-красным надсвязником, который лежит на рыльце и тем самым ограждает его от собственной пыльцы, т. е. препятствует самоопылению. Та часть столбика, которая прилегает к пыльникам, имеет своеобразное строение. Она характеризуется восемью мозолистыми гребнями, в промежутках между которыми лежат гнезда пыльников. Когда в дальнейшем столбик удлиняется, в канавках между ребрами остается освободившаяся из пыльников пыльца. Крошечное рыльце на верхушке столбика не может опыляться этой пыльцой. Опыление происходит при участии птиц. Стоя на листочках обертки соцветия, птица погружает голову в полное нектара соцветие и при этом касается последовательно рылец и покрытых пыльцой частей столбиков. Наряду с сахарной птицей способствует опылению также нектарница (Nectarinia shalybea), но сахарная птица является более частой посетительницей. Не удивительно, что соцветия протеи привлекают также многочисленных насекомых, в частности двукрылых и жуков, которые, несомненно, также принимают какое-то участие в опылении. Аналогичный способ опыления может наблюдаться у протеи ползучей (P. repens) — национального цветка Южно-Африканской Республики, а также у некоторых других видов.
Птицами опыляются также некоторые протейные Австралии. Многие виды гревиллеи и банксии, также как виды дриандры, аденантоса (Aclenanthos), хакеи и ламбертии опыляются медососовыми. Птицами опыляется также новозеландская рева-рева (Knightia excelsa). У дриандры снежной (D. nivea) соцветие имеет чашевидную форму. Нектар, вытекающий из многочисленных цветков, накапливается на дне этой чаши; обильные его запасы привлекают птиц-опылителей.
Как впервые убедительно показал Л. Карпентер (1978), значительную роль в опылении некоторых родов протейных Австралии играют нелетающие млекопитающие — грызуны и сумчатые. Некоторые виды банксии с крючковидными столбиками опыляются почти исключительно нелетающими млекопитающими, включая местный вид крысы (Rattus fuscipes) и различные роды мелких сумчатых. В Австралии и Новой Гвинее соцветия банксии посещают виды сумчатого рода петаурус (Petaurus). Опыление протейных некоторыми мелкими млекопитающими наблюдалось также в Африке. Опыление нелетающими млекопитающими характерно для низкорослых, нередко корневищных протейных, у которых цветки, собранные в более или менее чашевидные головчатые соцветия, развиваются невысоко над землей или же пробиваются через песок прямо от подземных корневищ (Дж. Рурке и Д. Вине, 1977; Д. Вине и Дж. Рурке, 1978). Такие «геофлорные», как их иногда называют, формы характерны для рода протея, а также для ряда видов дриандры, банксии и некоторых других. Цветки многих из этих видов обильно производят нектар и часто имеют острый запах. В Южной Африке грызуны (в данном случае особый вид полевой мыши) особенно активно посещают некоторые виды протеи. Их привлекают сочные и сладкие листочки обертки соцветия, так же как сочные столбики. Пыльца собирается в шерсти, покрывающей голову мыши, и таким образом переносится с одного растения на другое. Таким же путем переносится пыльца австралийским мышевидным сумчатым — хоботно-головым кускусом (Tarsipes spencerae), питающимся нектаром протейных. Разрушительная деятельность этих животных компенсируется огромным числом цветков в соцветии (у банксии, вероятно, превышающим 4000 цветков в колосе). В Австралии нелетающие млекопитающие посещают также многие крупные кустарниковые и даже древесные протейные, хотя «геофлорные» виды, видимо, все же лучше приспособлены к опылению ими. Эти геофлорные виды (многие виды банксии) имеют запах и выделяют обильный нектар, что привлекает многих насекомых. Как указывает австралийский ботаник М. К. Моркомб (1968), обилие ночных насекомых, посещающих соцветия банксии, в свою очередь привлекает нелетающих млекопитающих. Однако, принимая во внимание высокую приспособленность хоботкоголового кускуса к питанию нектаром, трудно предположить, что его привлекают только насекомые.
Способы распространения плодов и семян протейных очень разнообразны. Многие роды, у которых все или некоторые виды произрастают в дождевых лесах, характеризуются плодами-листовками, содержащими различное число обычно крылатых семян. Но еще чаще плоды нераскрывающиеся. Некоторые из них настоящие костянки, хорошо приспособленные для эндозоохорного распространения. Они также характерны для дождевых лесов. Интересно, что примитивная костянка монотипного новокаледонского рода гарниерия (Garnieria) содержит несколько семян. У тасманийского рода беллендена, так же как у многих других родов, плод сухой, нераскрывающийся и односемянный. Протейные с такими плодами обычно склерофильны и растут в открытых лесах, редколесьях, кустарниковых зарослях (скрэбах) и верещатниках, хотя имеются и исключения. Такие сухие нераскрывающиеся плоды часто бывают снабжены пучком волосков или, как у франкландии трехостной (Franklandia triaristata), перистыми остями. Они распространяются ветром, насекомыми, птицами (как у некоторых видов протеи) и, возможно, также млекопитающими.
Обычно семейство протейные делят на два подсемейства — гревиллеевые (Grevilleoideae), характеризующиеся цветками большей частью в пазухе одного кроющего листа и обычно раскрывающимися плодами, и протейные (Proteoideae), у которых цветки большей частью одиночные в пазухах кроющих листьев, а плоды нераскрывающиеся, орешкообразные или костянковидные. Эта система имеет ряд недостатков, и поэтому австралийские ботаники Л. Джонсон и Б. Бриггс предложили в 1975 г. свою систему. По новой системе семейство делится уже не на два, а на пять подсемейств. Но эта новая система также страдает недостатками, на которых здесь нет возможности останавливаться. Создание вполне удовлетворительной филогенетической системы протейных, как, впрочем, и многих других семейств, — дело будущего.
Одним из самых примитивных протейных является монотипный род плакоспермум (Placospermum), встречающийся в дождевых лесах Северного Квинсленда. Это дерево с кожистыми, перистонервными листьями. Плодолистики с многочисленными (15 и более) семязачатками, а плоды — кожистые листовки, содержащие несколько крылатых семян. В некоторых отношениях примитивен также монотипный тасманийский род беллендена (Bellendena), характеризующийся совершенно свободными тычинками "и семенами с эндоспермом. Ио в то же время рудиментарный венчик у беллендены полностью отсутствует. Оба рода относятся к подсемейству протейных.
К тому же подсемейству протейных относится небольшой род эмботриум (Embothrium, 8 видов), отличающийся очень разорванным ареалом, включающим Южную Америку (Эквадор, Перу и Чили), Новую Гвинею вместе с островами Ару и Австралию. Это типичные орнитофильные деревья и кустарники, произрастающие преимущественно в дождевых лесах. Виды эмботриума очень красивы, и некоторые из них успешно культивируются в Юго-Западной Англии и в Ирландии. К эмботриуму близок небольшой род телопея (Telopea), насчитывающий всего 4 вида в Восточной Австралии и на острове Тасмания. Это высокие кустарники с красивыми, красными, слегка зигоморфными цветками, собранными в густые верхушечные кисти. Плоды — листовки, содержащие крылатые семена. Телопея прекрасная (Т. speciosissima) — национальный цветок Австралии.
Во многих отношениях очень интересен род макадамия (Macadamia), также относящийся к подсемейству протейных. В него входит 12 видов, один из которых растет на острове Сулавеси, 5 видов встречаются в Австралии (Квинсленд и Новый Южный Уэльс) и 6 видов — в Новой Каледонии. Деревья и кустарники с обоеполыми цветками в верхушечных или пазушных кистях. Плод костянковидный, с одним или двумя семенами внутри твердого эндокардия. Семена съедобны, очень питательны и отличаются хорошим вкусом, несколько напоминающим вкус лещины. Как орехоплодное растение макадамия хорошо известна аборигенам Австралии, которые называют ее «кинуал-кинуал». Одна из разновидностей макадамии тройчатолистной (М. ternifolia), иногда выделяемая в отдельный вид (макадамия цельнолистная — М. integrifolia), широко культивируется в Австралии, Юго-Западной Азии, на Гавайских островах и в некоторых других местах. На Гавайских островах квинслендский орех является одним из важнейших культурных растений и занимает второе место после кофе. Перспективна для культуры: также эндемичная для Сулавеси макадамия Гильдебранда (М. hildebrandii), которая оказалась лучше приспособленной к влажному тропическому климату, чем австралийский вид. К макадамии близок монотипный мадагаскарский род малагасия (Malagasia), о котором, к сожалению, известно очень мало.
Род гревиллея (Grevillea) — самый большой в семействе. Он включает около 250 видов, распространенных главным образом в Австралии, 3 вида в Новой Каледонии, 1 вид на Новых Гебридах и 4 вида в Восточной Малезии и Новой Гвинее. Деревья и кустарники с обоеполыми цветками в кистях и плодами-листовками содержащими 1 или 2 плоских семени. Некоторые виды гревиллеи доставляют ценную древесину. Австралийская гревиллея мощная, или «шелковый дуб» (G. robusta), и некоторые другие виды широко используются в тропиках как растения, дающие хорошую тень на кофейных плантациях и ценную древесину. Гревиллея мощная используется для облесения и для придорожных аллей.
 ,
, 
Следующий по числу видов австралийский род хакея (Hakea, табл. 46 и 47) относится к тому же подсемейству и к той же трибе, что и гревиллея. В нем более 100 видов, произрастающих в Австралии и Тасмании, главным образом в Юго-Западно-Австралийской флористической провинции и в южной части Эремейской провинции. Ксерофильные кустарники или редко небольшие деревья, отличающиеся очень твердой древесиной. Плод сухой, с твердым, часто деревянистым околоплодником, раскрывающимся у верхушки на 2 створки, с 2 или 1 семенем. Древесину хакеи светлокрылой (Н. leucoptera) используют для производства фанеры, курительных трубок, мундштуков.
 ,
, Род банксия (Banksia, рис. 181, табл. 46) заключает около 50 видов, распространенных в Австралии и Тасмании, за исключением одного вида банксия зубчатая (В. dentata), произрастающего также в Новой Гвинее, на островах Ару. Ксерофильные кустарники и деревья с жесткими зубчатыми листьями и цветками в густых продолговатых колосьях. Плоды сухие, сплющенные, раскрывающиеся двумя твердыми, часто деревянистыми створками, расположенными перпендикулярно оси колоса. Все соплодие очень похоже на шишку какого-нибудь голосеменного растения. Семян обычно 2, с верхушечным перепончатым крылом. Цветки в большом количестве производят нектар, привлекающий насекомых и птиц, относящихся к семействам нектарницевые, белоглазковые и медососовые. На соцветиях банксии нередко можно наблюдать карликовую сумчатую летягу (Petaurus breviceps), которая высасывает нектар своим длинным, тонким, червеобразным языком. Древесина банксии твердая, используют ее для оконных рам, при строительстве лодок и судов, для изготовления мебели и для тонких столярных работ.
 ,
, К банксии близок род дриандра (Dryandra, рис. 182, табл. 46), насчитывающий более 50 видов, распространенных во внетропических областях Западной Австралии. Виды дриандры — кустарники, иногда низкие и цветущие близ основания. Листья дриандр довольно характерны — колючезубчатые или перистолопастные. Цветки обоеполые, в густых головках, окруженных многочисленными листочками обертки. Плод — коробочка, раскрывающаяся двумя кожистыми или деревянистыми створками и несущая 2 или 1 семя. Семена крылатые. Отпечатки листьев дриандры неоднократно указывались для третичных отложений Евразии, но все эти определения ошибочны. В большинстве случаев отпечатки, относившиеся к дриандре, оказываются отпечатками листьев комптонии из семейства мириковых (Myricaceae).
К другой группе протейных относится африканский род протея (Protea, табл. 45). В роде протея насчитывается более 100 видов, большинство которых сосредоточено в юго-западной части Капского флористического царства. Некоторые виды доходят до Центральной Африки. Многие виды протеи необычайно красивы. Особенно эффектна протея артишоковидная (P. cynaroides) с ее крупными (до 30 см в диаметре) головчатыми соцветиями. Протея олеандровидная (P. neriifolia) с листьями, похожими на листья олеандра, очень характерна для кустарниковых зарослей юго-западного побережья Южной Африки. Виды протеи — небольшие деревья и кустарники с многочисленными обоеполыми зигоморфными цветками, собранными в головчатые соцветия, окруженные многочисленными, часто различным образом окрашенными листочками обертки. Плод — односемянный орех.

К протее близок южноафриканский род левкадендрон (Leucadendron, табл. 45), заключающий около 100 видов, большинство которых встречаются в юго-западной части Капского флористического царства. Левкадендрон — кустарники или деревья с двудомными цветками и односемянными орехообразными плодами. Как и виды протеи, они играют значительную роль в растительном покрове Капского царства. Наиболее известный представитель рода — знаменитое «серебряное дерево» (L. argenteum), густо покрытое тонкими шелковистыми волосками и сверкающее на склонах блестящими серо-зелеными листьями, как посеребренный ивовый куст.
Поделитесь на страничкеslovar.wikireading.ru
Семейство протейные (Proteaceae) - это... Что такое Семейство протейные (Proteaceae)?
Семейство протейных, насчитывающее более 70 родов и, вероятно, не менее 1400 видов, распространено главным образом в южном полушарии. Наибольшая концентрация протейных наблюдается в Южной Африке (преимущественно в Капском флористическом царстве), где встречается 14 родов и почти 400 видов, и особенно в Австралии вместе с Тасманией (45 родов и около 800 видов). Однако ареал семейства гораздо шире и охватывает тропическую Африку от Эфиопии на севере, Мадагаскар, Южную Индию и остров Шри-Ланка, Восточную и Юго-Восточную Азию, Новую Гвинею, Новую Каледонию, Новую Зеландию и многие острова Тихого океана. Из всех протейных дальше всех на север заходят некоторые виды рода хелиция (Helicia, рис. 180), ареал которого простирается от Индокитая и Тайваня до Японии (остров Хонсю).
Очень велика роль протейных в растительном мире Капского флористического царства, где они поражают множеством форм, разнообразием листьев и исключительной красотой соцветий. Наиболее характерны виды левкадендрона (Leucadendron) и протеи (Protea, табл.45). Значительно меньшую роль играют протейные в тропической Африке, но и здесь они местами принимают заметное участие в растительности, например в Северной Гвинее. Довольно заметную роль играют протейные на Мадагаскаре. Но наиболее характерны они для растительного мира Австралии, особенно для Юго-Западной Австралии, Квинсленда, Нового Южного Уэльса и Тасмании. Здесь бросаются в глаза многочисленные виды гревиллеи (Grevillea), виды эндемичного рода банксия (Banksia, табл. 46, рис. 181) и многие другие. В Юго-Западной Австралии обычны виды эндемичного рода дриандра (Dryandra, рис. 182), так же как виды австралийско-тасманийского рода хакея (Hakea, табл. 46 и 47). Значительна роль протейных также в растительном мире Новой Каледонии. Что касается Азии и Америки, то протейные играют там менее заметную роль.
Протейные чаще всего кустарники и кустарнички, реже деревья, иногда очень высокие. Есть среди них также полукустарники и даже корневищные травы. Для многих протейных характерна жестколистность. Нередки среди них относительно мелколистные формы. Весь облик большинства протейных говорит о приспособлении к временной сухости муссонного климата и к бедным почвам. Как у южноафриканских, так и у австралийских протейных часто имеются так называемые лигнотуберусы — деревянистые подземные запасающие органы стеблевого происхождения. Однако в дождевых лесах встречаются деревья и с более или менее мезоморфными листьями. Протейные ясно выраженного жестколистного характера чаще всего кустарники, но бывают среди них и деревья высотой от 6 до 20 м. Очень многие типично жестколистные виды произрастают на почвах, бедных питательными веществами, особенно фосфором. Листья протейных характеризуются исключительно широким размахом изменчивости как размеров, формы и строения, так и типов жилкования. Не случайно Линней назвал род протея именем Протея, который, согласно древнегреческому мифу, мог по желанию обрести любую форму и любой образ.
Цветки протейных обычно в различного рода бокоцветных соцветиях: кистях, зонтиках, колосьях или головках (простых или сложных), редко одиночные в пазухах листьев (результат редукции соцветия). Очень характерна ясно выраженная в семействе тенденция к образованию головчатых соцветий, часто внешне удивительно напоминающих соцветия многих сложноцветных. Головчатые соцветия возникают независимо у разных представителей семейства. Особенно характерны такие соцветия для рода протея в Южной и тропической Африке и родов телопеа (Telopea), дриандра и других в Австралии. Цветки протейных актиноморфные или более или менее зигоморфные, обычно обоеполые, но иногда однополые и у некоторых родов, как, например, у левкадендрона, двудомные. Околоцветник представлен венчиковидной, обычно сростнолистной, 4-члениой чашечкой. Интересно, что в бутоне чашечка обычно трубчатая, но при распускании цветка чашелистики более или менее отделяются друг от друга и в той или иной степени загибаются книзу, а иногда закручиваются. Что касается венчика, то он редуцирован, хотя у разных родов в неодинаковой степени. Вместо нормально развитого венчика в цветке обычно имеется более или менее развитый 4-членный подпестичный круг желёзок или чешуек или же чашевидный диск, как правило, несущие секреторную функцию. Другими словами, лепестки у протейных в той или иной степени превращены в нектарники. В то время как в актиноморфных цветках можно видеть полный 4-членный круг желёзок, каждая из которых соответствует лепестку, в зигоморфных цветках круг этот неполный: абаксиальные желёзки исчезают и сохраняются только 3 или даже только 2 адаксиальные желёзки. Иногда этот процесс идет дальше, и в цветке не сохраняется никаких следов венчика. У австралийского рода франкландия (Franklandia) сохранилось еще определенное сходство с венчиком, но 4 лопасти его сростнолепестного венчика уже функционируют как нектарники. В тех случаях, когда лепестки сведены к желёзкам и чешуйкам, эти последние обычно более или менее сросшиеся между собой, что, возможно, свидетельствует о том, что венчик у протейных был сростнолепестный. Тычинок всегда 4, чередующихся с чашелистиками, с широкими нитями, обычно более или менее (иногда почти полностью) приросшими к чашелистикам. Лишь редко нити бывают полностью свободными, как у относительно примитивного монотипного тасманийского рода беллендена (Bellendena). Пыльники иногда с ясно выраженным надсвязником, особенно хорошо развиты у монотипного новокаледонского рода бопреопсис (Beaupreopsis). Гинецей одноплодолистиковый, но, несмотря на общую довольно высокую специализированность цветка протейных, плодолистик относительно примитивный, еще в сущности кондупликатный. У некоторых родов, имеющих рудиментарный венчик, плодолистик на более ранней стадии своего развития еще не вполне замкнутый и в нижней своей части (в области завязи), а также в районе рыльца еще более или менее открытый. У некоторых протейных свободные брюшные поверхности несут железистые волоски, как это наблюдается у кондупликатных плодолистиков некоторых примитивных цветковых растений. Однако в строении плодолистика протейных ясно выражены также определенные признаки довольно высокой специализации. Столбик нитевидный и прямой или же согнутый, у большинства видов с различной формы косым или боковым рыльцем. Функционирующая рыльцевая поверхность может быть ограничена маленькой точкой на верхушке столбика или в центре верхушечного диска. У многих протейных с наличием рудиментарного венчика завязь на ножке (гинофоре), что характерно для орнитофильных видов. Семязачатков в каждом плодолистике от одного до многих. Плод — листовка, орешек, крылатка или костянка. За исключением рода боллендена, семена без эндосперма.
Цветки протейных обычно протандричны. Опыление производится различными представителями животного мира — насекомыми, птицами (нектарниками, белоглазками, австралийско-новогвинейскими медососами и родственными им южноафриканскими сахарными птицами и американскими колибри), рукокрылыми (крыланами и летучими мышами) и мелкими нелетающими млекопитающими (грызунами, мелкими сумчатыми из семейства лазящих сумчатых, или кускусов, — Phalangeridae).
У протейных наблюдается довольно большое разнообразие приспособлений к перекрестному опылению, особенно хорошо изученное у австралийских протейных (Р. Каролин, 1908). У некоторых из них механизм опыления относительно простой, как, например, у австралийской персоонии игольчатой (Persoonia acerosa), цветки которой посещают главным образом пчелы. В только что раскрывшемся цветке столбик с еще незрелым рыльцем окружен тесно расположенными вокруг него длинными пыльниками, которые вскоре вскрываются продольными щелями. Когда насекомое просовывает свой хоботок между пыльниками и столбиком, часть пыльцы откладывается на хоботке и теле. На следующей женской фазе развития цветка пыльники закручиваются назад, а рыльце становится клейким и готово к восприятию пыльцы. Иной механизм опыления наблюдается у другого австралийского протейного — изопогона укрополистного (Isopogon anethifolius). Пыльники изопогона также тесно прижаты к столбику, но, в отличие от персоонии, они открываются еще в бутоне. Непосредственно под пыльниковым каналом на столбике имеется вздутие, преграждающее пыльцу от падения вниз. Затем сегменты околоцветника вместе с прикрепленными к ним тычинками закручиваются назад, оставляя в центре цветка покрытый пыльцой свободно возвышающийся столбик. Таким образом, в отличие не только от персоонии, но и от подавляющего большинства цветковых растений, подача пыльцы опылителю осуществляется не самими пыльниками, а через посредство столбика. Эту же модель, но в более или менее видоизмененной форме, мы находим и у целого ряда других родов протейных. Так, у австралийского рода ламбертия (Lambertia) пыльца откладывается в бутоне на клейкой верхней части столбика. У другого австралийского протейного — петрофилы окрашенной (Petrophila fucifolia) — оседанию пыльцы на столбике способствуют покрывающие ее короткие волоски, представляющие собой, таким образом, аналог «пыльцеснимателя» колокольчиковых (см. ниже, семейство колокольчиковых). У всех этих растений рыльце созревает и бывает готово к восприятию пыльцы лишь после окончания мужской фазы, когда большая часть пыльцы уже выгружена опылителями или засохла.
У гревиллеи, хакеи, телопей, ломатии и некоторых других родов механизм опыления несколько иной. Верхушка столбика у них уплощена и расширена в форме прямого, косого или бокового диска, в центре которого в виде маленькой выпуклости расположено рыльце. Пыльники вскрываются в бутоне, и пыльца откладывается на дисковидной верхушке столбика, в том числе и на самом рыльце, которое на, этой мужской стадии еще совершенно не восприимчиво к ней; Некоторое время дисковидная верхушка столбика остается зажатой в верхней части еще полностью не раскрывшегося околоцветника. У некоторых протейных, например, у рода протеи, диск не высвобождается до тех пор, пока этой части цветка не коснется опылитель. Но уже при малейшем прикосновении сразу же срабатывает этот удивительный триггерный механизм, в котором столбик играет роль пружины изгиба, и диск с силой вырывается из объятий околоцветника и разбрасывает накопившуюся на нем пыльцу, осыпая ею тело опылителя. Благодаря этому механизму достигается двоякая цель: до высвобождения диска пыльца бывает защищена от превратностей внешнего мира, а внезапное ее высвобождение способствует разбрасыванию пыльцы. После освобождения диска начинается женская стадия — крошечное рыльце в центре диска увеличивается в размере и становится готовым к восприятию пыльцы, принесенной опылителем с другого цветка.
Совершенно иную модель механизма опыления мы наблюдаем у некоторых других родов. Особенно интересны в этом отношении австралийские роды симфионема (Symphyonema) и коноспермум (Conospermum). В мелких желтых цветках симфионемы пыльники расположены тесно вокруг столбика. Когда пыльники созревают, сегменты околоцветника закручиваются назад. В поисках нектара опылитель (в данном случае обычно бражники) обязательно коснется пыльников, которые внезапно отделяются друг от друга, разбрасывая при этом пыльцу во всех направлениях. Часть этой пыльцы попадает и на опылителя. А в крупных цветках коноспермума эллиптического (С. ellipticum) пыльники соединены вокруг столбика, причем некоторые из них заканчиваются длинной щетинкой, или остью. Часть пыльников стерильна. Столбик согнут в форме лебединой шеи с рыльцем, направленным в сторону от фертильных пыльников. Малейшее прикосновение к столбику или к щетинкам пыльников приводит к быстрому движению столбика и внезапному отделению пыльников друг от друга. Верхняя часть столбика поворачивается в сторону пустых пыльников, и рыльце с силой ударяет по опылителю (в данном случае обычно по мухе). Если на опылителе уже есть пыльца с другого цветка, то часть этой пыльцы прилипнет к рыльцу. Пыльники отделяются почти сразу же за движением столбика, поднимая маленькое облачко пыльцы, часть которой попадает на муху.
Очень многие протейные приспособлены к опылению исключительно или главным образом птицами, особенно в Австралии и Южной Африке. Птицы опыляют виды протейных с крупными красными, оранжевыми, желтыми или желтовато-зелеными, часто контрастно окрашенными верхушечными соцветиями, отдельные цветки которых обильно выделяют нектар и обычно лишены запаха, столь важного для привлечения насекомых и млекопитающих. Цветки некоторых видов банксии выделяют так много нектара, что он используется в качестве пищи аборигенами Австралии.
Взаимное приспособление протейных и птиц в некоторых случаях принимает форму взаимной зависимости. Классическим примером является южноафриканская сахарная птица промеропс (Promerops cafer), которая настолько приспособилась к опылению некоторых видов протеи, что почти не может существовать без этих растений. У сахарной птицы клюв и язык длинные и тонкие, а соцветия этих видов протеи, окруженные плотно прилегающими друг к другу листочками обертки, представляют собой как бы естественную чашу, из которой птица пьет нектар. Сахарная птица настолько сжилась с видами протеи, что даже гнезда свои строит из листочков обертки и из старых цветков, заключающих зрелые семена, и тем самым содействует диссеминации. В качестве примера рассмотрим опыление у кустарниковой протеи крупноголовой (P. macrocephala). Листочки обертки этого вида светло-зеленые, но хохолок, образованный волосками на кончиках листочков, имеет посередине красный круг. Внутренние листочки обертки длиннее наружных, и их шерстисто опушенные верхушки отогнуты внутрь, к центру соцветия, и образуют удобную посадочную площадку для птиц. Чашечка состоит из одного свободного узкого чашелистика, несущего стерильную тычинку, и трех сросшихся чашелистиков, несущих каждый по одной фертильной тычинке. Пыльники заканчиваются маленьким ярко-красным надсвязником, который лежит на рыльце и тем самым ограждает его от собственной пыльцы, т. е. препятствует самоопылению. Та часть столбика, которая прилегает к пыльникам, имеет своеобразное строение. Она характеризуется восемью мозолистыми гребнями, в промежутках между которыми лежат гнезда пыльников. Когда в дальнейшем столбик удлиняется, в канавках между ребрами остается освободившаяся из пыльников пыльца. Крошечное рыльце на верхушке столбика не может опыляться этой пыльцой. Опыление происходит при участии птиц. Стоя на листочках обертки соцветия, птица погружает голову в полное нектара соцветие и при этом касается последовательно рылец и покрытых пыльцой частей столбиков. Наряду с сахарной птицей способствует опылению также нектарница (Nectarinia shalybea), но сахарная птица является более частой посетительницей. Не удивительно, что соцветия протеи привлекают также многочисленных насекомых, в частности двукрылых и жуков, которые, несомненно, также принимают какое-то участие в опылении. Аналогичный способ опыления может наблюдаться у протеи ползучей (P. repens) — национального цветка Южно-Африканской Республики, а также у некоторых других видов.
Птицами опыляются также некоторые протейные Австралии. Многие виды гревиллеи и банксии, также как виды дриандры, аденантоса (Aclenanthos), хакеи и ламбертии опыляются медососовыми. Птицами опыляется также новозеландская рева-рева (Knightia excelsa). У дриандры снежной (D. nivea) соцветие имеет чашевидную форму. Нектар, вытекающий из многочисленных цветков, накапливается на дне этой чаши; обильные его запасы привлекают птиц-опылителей.
Как впервые убедительно показал Л. Карпентер (1978), значительную роль в опылении некоторых родов протейных Австралии играют нелетающие млекопитающие — грызуны и сумчатые. Некоторые виды банксии с крючковидными столбиками опыляются почти исключительно нелетающими млекопитающими, включая местный вид крысы (Rattus fuscipes) и различные роды мелких сумчатых. В Австралии и Новой Гвинее соцветия банксии посещают виды сумчатого рода петаурус (Petaurus). Опыление протейных некоторыми мелкими млекопитающими наблюдалось также в Африке. Опыление нелетающими млекопитающими характерно для низкорослых, нередко корневищных протейных, у которых цветки, собранные в более или менее чашевидные головчатые соцветия, развиваются невысоко над землей или же пробиваются через песок прямо от подземных корневищ (Дж. Рурке и Д. Вине, 1977; Д. Вине и Дж. Рурке, 1978). Такие «геофлорные», как их иногда называют, формы характерны для рода протея, а также для ряда видов дриандры, банксии и некоторых других. Цветки многих из этих видов обильно производят нектар и часто имеют острый запах. В Южной Африке грызуны (в данном случае особый вид полевой мыши) особенно активно посещают некоторые виды протеи. Их привлекают сочные и сладкие листочки обертки соцветия, так же как сочные столбики. Пыльца собирается в шерсти, покрывающей голову мыши, и таким образом переносится с одного растения на другое. Таким же путем переносится пыльца австралийским мышевидным сумчатым — хоботно-головым кускусом (Tarsipes spencerae), питающимся нектаром протейных. Разрушительная деятельность этих животных компенсируется огромным числом цветков в соцветии (у банксии, вероятно, превышающим 4000 цветков в колосе). В Австралии нелетающие млекопитающие посещают также многие крупные кустарниковые и даже древесные протейные, хотя «геофлорные» виды, видимо, все же лучше приспособлены к опылению ими. Эти геофлорные виды (многие виды банксии) имеют запах и выделяют обильный нектар, что привлекает многих насекомых. Как указывает австралийский ботаник М. К. Моркомб (1968), обилие ночных насекомых, посещающих соцветия банксии, в свою очередь привлекает нелетающих млекопитающих. Однако, принимая во внимание высокую приспособленность хоботкоголового кускуса к питанию нектаром, трудно предположить, что его привлекают только насекомые.
Способы распространения плодов и семян протейных очень разнообразны. Многие роды, у которых все или некоторые виды произрастают в дождевых лесах, характеризуются плодами-листовками, содержащими различное число обычно крылатых семян. Но еще чаще плоды нераскрывающиеся. Некоторые из них настоящие костянки, хорошо приспособленные для эндозоохорного распространения. Они также характерны для дождевых лесов. Интересно, что примитивная костянка монотипного новокаледонского рода гарниерия (Garnieria) содержит несколько семян. У тасманийского рода беллендена, так же как у многих других родов, плод сухой, нераскрывающийся и односемянный. Протейные с такими плодами обычно склерофильны и растут в открытых лесах, редколесьях, кустарниковых зарослях (скрэбах) и верещатниках, хотя имеются и исключения. Такие сухие нераскрывающиеся плоды часто бывают снабжены пучком волосков или, как у франкландии трехостной (Franklandia triaristata), перистыми остями. Они распространяются ветром, насекомыми, птицами (как у некоторых видов протеи) и, возможно, также млекопитающими.
Обычно семейство протейные делят на два подсемейства — гревиллеевые (Grevilleoideae), характеризующиеся цветками большей частью в пазухе одного кроющего листа и обычно раскрывающимися плодами, и протейные (Proteoideae), у которых цветки большей частью одиночные в пазухах кроющих листьев, а плоды нераскрывающиеся, орешкообразные или костянковидные. Эта система имеет ряд недостатков, и поэтому австралийские ботаники Л. Джонсон и Б. Бриггс предложили в 1975 г. свою систему. По новой системе семейство делится уже не на два, а на пять подсемейств. Но эта новая система также страдает недостатками, на которых здесь нет возможности останавливаться. Создание вполне удовлетворительной филогенетической системы протейных, как, впрочем, и многих других семейств, — дело будущего.
Одним из самых примитивных протейных является монотипный род плакоспермум (Placospermum), встречающийся в дождевых лесах Северного Квинсленда. Это дерево с кожистыми, перистонервными листьями. Плодолистики с многочисленными (15 и более) семязачатками, а плоды — кожистые листовки, содержащие несколько крылатых семян. В некоторых отношениях примитивен также монотипный тасманийский род беллендена (Bellendena), характеризующийся совершенно свободными тычинками "и семенами с эндоспермом. Ио в то же время рудиментарный венчик у беллендены полностью отсутствует. Оба рода относятся к подсемейству протейных.
К тому же подсемейству протейных относится небольшой род эмботриум (Embothrium, 8 видов), отличающийся очень разорванным ареалом, включающим Южную Америку (Эквадор, Перу и Чили), Новую Гвинею вместе с островами Ару и Австралию. Это типичные орнитофильные деревья и кустарники, произрастающие преимущественно в дождевых лесах. Виды эмботриума очень красивы, и некоторые из них успешно культивируются в Юго-Западной Англии и в Ирландии. К эмботриуму близок небольшой род телопея (Telopea), насчитывающий всего 4 вида в Восточной Австралии и на острове Тасмания. Это высокие кустарники с красивыми, красными, слегка зигоморфными цветками, собранными в густые верхушечные кисти. Плоды — листовки, содержащие крылатые семена. Телопея прекрасная (Т. speciosissima) — национальный цветок Австралии.
Во многих отношениях очень интересен род макадамия (Macadamia), также относящийся к подсемейству протейных. В него входит 12 видов, один из которых растет на острове Сулавеси, 5 видов встречаются в Австралии (Квинсленд и Новый Южный Уэльс) и 6 видов — в Новой Каледонии. Деревья и кустарники с обоеполыми цветками в верхушечных или пазушных кистях. Плод костянковидный, с одним или двумя семенами внутри твердого эндокардия. Семена съедобны, очень питательны и отличаются хорошим вкусом, несколько напоминающим вкус лещины. Как орехоплодное растение макадамия хорошо известна аборигенам Австралии, которые называют ее «кинуал-кинуал». Одна из разновидностей макадамии тройчатолистной (М. ternifolia), иногда выделяемая в отдельный вид (макадамия цельнолистная — М. integrifolia), широко культивируется в Австралии, Юго-Западной Азии, на Гавайских островах и в некоторых других местах. На Гавайских островах квинслендский орех является одним из важнейших культурных растений и занимает второе место после кофе. Перспективна для культуры: также эндемичная для Сулавеси макадамия Гильдебранда (М. hildebrandii), которая оказалась лучше приспособленной к влажному тропическому климату, чем австралийский вид. К макадамии близок монотипный мадагаскарский род малагасия (Malagasia), о котором, к сожалению, известно очень мало.
Род гревиллея (Grevillea) — самый большой в семействе. Он включает около 250 видов, распространенных главным образом в Австралии, 3 вида в Новой Каледонии, 1 вид на Новых Гебридах и 4 вида в Восточной Малезии и Новой Гвинее. Деревья и кустарники с обоеполыми цветками в кистях и плодами-листовками содержащими 1 или 2 плоских семени. Некоторые виды гревиллеи доставляют ценную древесину. Австралийская гревиллея мощная, или «шелковый дуб» (G. robusta), и некоторые другие виды широко используются в тропиках как растения, дающие хорошую тень на кофейных плантациях и ценную древесину. Гревиллея мощная используется для облесения и для придорожных аллей.
 ,
, 
Следующий по числу видов австралийский род хакея (Hakea, табл. 46 и 47) относится к тому же подсемейству и к той же трибе, что и гревиллея. В нем более 100 видов, произрастающих в Австралии и Тасмании, главным образом в Юго-Западно-Австралийской флористической провинции и в южной части Эремейской провинции. Ксерофильные кустарники или редко небольшие деревья, отличающиеся очень твердой древесиной. Плод сухой, с твердым, часто деревянистым околоплодником, раскрывающимся у верхушки на 2 створки, с 2 или 1 семенем. Древесину хакеи светлокрылой (Н. leucoptera) используют для производства фанеры, курительных трубок, мундштуков.
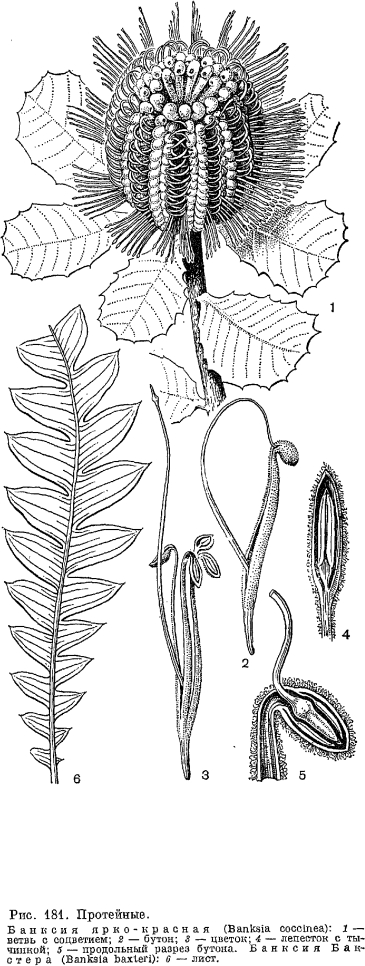 ,
, Род банксия (Banksia, рис. 181, табл. 46) заключает около 50 видов, распространенных в Австралии и Тасмании, за исключением одного вида банксия зубчатая (В. dentata), произрастающего также в Новой Гвинее, на островах Ару. Ксерофильные кустарники и деревья с жесткими зубчатыми листьями и цветками в густых продолговатых колосьях. Плоды сухие, сплющенные, раскрывающиеся двумя твердыми, часто деревянистыми створками, расположенными перпендикулярно оси колоса. Все соплодие очень похоже на шишку какого-нибудь голосеменного растения. Семян обычно 2, с верхушечным перепончатым крылом. Цветки в большом количестве производят нектар, привлекающий насекомых и птиц, относящихся к семействам нектарницевые, белоглазковые и медососовые. На соцветиях банксии нередко можно наблюдать карликовую сумчатую летягу (Petaurus breviceps), которая высасывает нектар своим длинным, тонким, червеобразным языком. Древесина банксии твердая, используют ее для оконных рам, при строительстве лодок и судов, для изготовления мебели и для тонких столярных работ.
 ,
, К банксии близок род дриандра (Dryandra, рис. 182, табл. 46), насчитывающий более 50 видов, распространенных во внетропических областях Западной Австралии. Виды дриандры — кустарники, иногда низкие и цветущие близ основания. Листья дриандр довольно характерны — колючезубчатые или перистолопастные. Цветки обоеполые, в густых головках, окруженных многочисленными листочками обертки. Плод — коробочка, раскрывающаяся двумя кожистыми или деревянистыми створками и несущая 2 или 1 семя. Семена крылатые. Отпечатки листьев дриандры неоднократно указывались для третичных отложений Евразии, но все эти определения ошибочны. В большинстве случаев отпечатки, относившиеся к дриандре, оказываются отпечатками листьев комптонии из семейства мириковых (Myricaceae).
К другой группе протейных относится африканский род протея (Protea, табл. 45). В роде протея насчитывается более 100 видов, большинство которых сосредоточено в юго-западной части Капского флористического царства. Некоторые виды доходят до Центральной Африки. Многие виды протеи необычайно красивы. Особенно эффектна протея артишоковидная (P. cynaroides) с ее крупными (до 30 см в диаметре) головчатыми соцветиями. Протея олеандровидная (P. neriifolia) с листьями, похожими на листья олеандра, очень характерна для кустарниковых зарослей юго-западного побережья Южной Африки. Виды протеи — небольшие деревья и кустарники с многочисленными обоеполыми зигоморфными цветками, собранными в головчатые соцветия, окруженные многочисленными, часто различным образом окрашенными листочками обертки. Плод — односемянный орех.

К протее близок южноафриканский род левкадендрон (Leucadendron, табл. 45), заключающий около 100 видов, большинство которых встречаются в юго-западной части Капского флористического царства. Левкадендрон — кустарники или деревья с двудомными цветками и односемянными орехообразными плодами. Как и виды протеи, они играют значительную роль в растительном покрове Капского царства. Наиболее известный представитель рода — знаменитое «серебряное дерево» (L. argenteum), густо покрытое тонкими шелковистыми волосками и сверкающее на склонах блестящими серо-зелеными листьями, как посеребренный ивовый куст.
Жизнь растений: в 6-ти томах. — М.: Просвещение. Под редакцией А. Л. Тахтаджяна, главный редактор чл.-кор. АН СССР, проф. А.А. Федоров. 1974.
dic.academic.ru
Протейные
С.В. Мельникова
В последнее время в нашей стране появляется все больше магазинов, где можно увидеть в продаже высушенные растения для декоративных цветочных композиций. Есть здесь известные всем плотные бурые початки рогоза (чаще неправильно называемого «камышом»), метелки ковыля с длинными остями, стебли бамбука и кокосовые орехи...
Но, пожалуй, больше всего привлекают внимание покупателей экзотические сухоцветы – крупные, жесткие соцветия в ярких обертках, чем-то напоминающие огромный раскрашенный чертополох или забавные бутылочные ершики. Большинство этих диковинок – растения из семейства протейных. Для многих из нас они в новинку, но европейским любителям причудливых творений природы знакомы уже несколько столетий.
Протейные (Proteaceae) – довольно обширное ботаническое семейство. К нему относят около 70 родов и более 1400 видов растений, распространенных в тропических и субтропических (редко – умеренных) областях главным образом к югу от экватора и преимущественно в Восточном полушарии. Облик протейных может быть достаточно разнообразным – среди них встречаются и многолетние травы, и деревья высотой до 20 м. Но наиболее типичная для семейства жизненная форма – небольшие деревца и кустарники.
Протейные – жители областей с бедной почвой и муссонным климатом, отличающимся периодическими засухами. Такие условия отразились на внешнем облике этих растений – у протейных кожистые листья чаще всего линейной или игольчатой формы, у многих представителей семейства есть особые подземные органы стеблевого происхождения – лигнотуберусы, которые служат вместилищами воды. Настоящим украшением протейных являются их цветки, собранные в эффектные кисти, колосья или головчатые соцветия, похожие на соцветия сложноцветных. Характерная и примечательная черта семейства – приспособленность многих видов к опылению позвоночными животными – птицами, летучими мышами и мелкими лазающими млекопитающими. Цветки таких протейных ярко окрашены в разные оттенки желтого и красного цветов и содержат много сладкого нектара.
Множество удивительных представителей протейных – почти 400 видов относящихся к 14 родам – можно увидеть в Капской провинции на юге Африки. Здесь встречается, например, около 100 видов необычайно красивых растений из рода протея (Protea). Название, впоследствии ставшее названием всего семейства, дал этим растениям Карл Линней. Предполагают, что великий систематик использовал имя древнегреческого морского божества Протея, который мог по своему желанию принимать разный облик, чтобы подчеркнуть большое разнообразие протей – не только разные их виды сильно отличаются друг от друга по внешнему облику, но и листья в пределах одного экземпляра могут быть различными по форме.
Самая эффектная из протей – протея артишоковая (P. cynaroides) с крупными, до 30 см в диаметре, головчатыми соцветиями, которые окружены яркими обертками. Местное население называет это растение «протея-король». Другое название протеи артишоковой – «горшок меда». Действительно, крупные соцветия растения наполнены сладким нектаром, который африканцы используют как лекарство от кашля. Полны сладкого нектара и красивые крупные соцветия протеи ползучей (P.repens), считающейся цветком-символом ЮАР.
Еще один вид южноафриканских протей – протея крупноголовая (P.macrocephala). Она интересна тесной связью с так называемой сахарной птицей (Promerops cafer). Эта птица опыляет протею, извлекая из ее соцветий, обертки которых образуют подобие чаш, нектар с помощью длинного клюва. Она и гнездится в этих кустарниках, используя для постройки гнезд старые цветки и листочки обертки. Крупноголовую протею опыляют и другие пернатые, например нектарницы.
Среди других южноафриканских протейных знаменито так называемое «серебряное дерево», или леукадендрон серебристый (Leucadendron argenteum), – растение, густо покрытое длинными серебристыми волосками. Очень эффектны во время цветения и представители рода леукоспермум (Leucospermum) – небольшие кустарники с плотными и заостренными кожистыми листьями. Из соцветий леукоспермума торчат длинные пестики, придавая всему цветку «ощетинившийся» вид. Местные жители называют эти растения «подушками для булавок».
Еще большим разнообразием протейных отличается Австралия – здесь встречается около 800 видов растений этого семейства. Многие из них также приспособлены к опылению позвоночными – лазающими сумчатыми и грызунами, и их соцветия имеют характерный острый запах. Сильный острый запах присущ представителям рода банксия (Banksia), соцветия которой напоминают своеобразные ершики для мытья посуды, а соплодия похожи на шишки голосеменных растений. Банксии растут в жестколистных тропических лесах, а также в саваннах Австралии и Тасмании вместе с эвкалиптами и акациями. Всего известно около 50 видов этого рода. Эмблемой австралийского штата Новый Южный Уэльс стала другая представительница протейных – телопея красивейшая (Telopea speciosissima). Ее крупные соцветия имеют правильную форму и напоминают соцветия хризантемы.
В Австралии и Южной Африке многие протейные культивируют в садах и парках как декоративные растения, а ряд видов даже выращивают на специальных плантациях – как красивые «сухоцветы», которые потом экспортируют в другие страны, главным образом в Европу. Коммерческие плантации протейных есть и в США. Можно встретить эти растения и в ботанических садах на юге Европы.
В умеренной же зоне Северного полушария протейные в открытом грунте не выживают и увидеть их можно, да и то, к сожалению, нечасто, только в оранжереях ботанических садов и у любителей.
Впрочем, так было не всегда. В XIX в. эти растения были очень популярны в Европе, их выращивали в зимних садах при дворах королей и в домах знати. Лондонский купец Георг Хибберт, чьим именем впоследствии был назван новый род протейных из Новой Зеландии – хиббертия (Hibbertia), собрал в своем саду коллекцию из 150 видов этих растений! Несколько видов протейных с особо красивыми соцветиями – леукадендрон, протея, банксия – часто выращивали в оранжереях Европы XIX в. Они хорошо цвели, их регулярно демонстрировали на выставках. Однако впоследствии внимание европейских любителей переключилось на орхидеи, и протейные понемногу утратили былую популярность.
Выращивать протейные в домашних условиях довольно сложно, а уход за ними должен быть очень тщательным. Эти растения весьма требовательны к освещению и вентиляции.
Сложно достать и семена, но они хороши тем, что сохраняют всхожесть в течение нескольких лет. Сотрудники Главного Ботанического сада РАН рекомендуют высевать семена протейных в смесь песка и верхового торфа с февраля по май. Сеять надо на глубину в 2,5 раза больше размера семени. Посевы выдерживают до прорастания при температуре 20–25о С без избыточного увлажнения. Всходы появляются через 5–6 недель или даже несколько месяцев. Процесс прорастания можно ускорить, проведя предварительную стратификацию – поместив семена во влажный песок и выдержав в холодильнике 7–8 недель при температуре 8–10 оС. Пересаживают сеянцы при появлении у них 2–4 настоящих листьев в смесь бурого рыхлого торфа и сосновой хвои в соотношении 1:1. Хорошо подходит для протейных и готовая земляная смесь для азалий и рододендронов, имеющая показатель рН 5,0–5,5 – при защелачивании почвы растения желтеют и гибнут. Молодые растения не выносят ни пересушивания земли, ни переувлажнения. Поливать протейные надо мягкой подкисленной водой.
К удобрениям протейные нетребовательны, т.к. в природе они растут на очень бедных землях. Для подкормки используют лишь кислые удобрения (готовые смеси для азалий и рододендронов). В зимнее время протейные должны находиться при температуре 5–6о С.
Список литературы
Жизнь растений. – М.: Просвещение, 1981.
Ивченко С. Занимательно о фитогеографии. – М.: Молодая гвардия, 1985.
Якимова Т. Многоликие протейные //В мире растений. 2000. № 5.
mirznanii.com
Семейство протейных. Фото
В древнегреческой мифологии есть такой персонаж — мудрый старец Протей, сын морского владыки Посейдона. Протей обладает способностью принимать образы различных существ и предметов. Он превращается в зверей и птиц, в огонь, в воду, в деревья и цветы. Именно Протея вспомнил знаменитый систематик растений К. Линней, придумывал названия растениям из семейства протейных. Учёного поразило многообразие обликов этих родственных видов. Посудите сами: плотные кожистые листья протейных бывают широкими и узкими, большими и маленькими, с ровными краями или с игольчатыми. Даже на одном экземпляре форма листьев может меняться с возрастом.
Странные цветки протейных собраны в соцветия — в кисти, колосья, зонтики или головки. В одном соцветии может насчитываться до 4000 цветков, как, например, у банксии вересколистной, крупное соцветие которой напоминает ёршик для мытья посуды. Таким же многообразием отличается и форма плодов протейных.
Семейство протейных, насчитывающее более 70 родов и, вероятно, не менее 1400 видов, распространено главным образом в южном полушарии. Наибольшая концентрация протейных наблюдается в Южной Африке (преимущественно в Капском флористическом царстве), где встречается 14 родов и почти 400 видов, и особенно в Австралии вместе с Тасманией (45 родов и около 800 видов). Однако ареал семейства гораздо шире и охватывает тропическую Африку от Эфиопии на севере, Мадагаскар, Южную Индию и остров Шри-Ланка, Восточную и Юго-Восточную Азию, Новую Гвинею, Новую Каледонию, Новую Зеландию и многие острова Тихого океана. Из всех протейных дальше всех на север заходят некоторые виды рода хелиция (Helicia), ареал которого простирается от Индокитая и Тайваня до Японии (остров Хонсю).
 Grevillea baileyana. Фото: Tatiana Gerus
Grevillea baileyana. Фото: Tatiana Gerus
 Faurea galpinii. Фото: Derek Keats
Faurea galpinii. Фото: Derek Keats
 Protea cynaroides. Фото: echoforsberg
Protea cynaroides. Фото: echoforsberg
Очень велика роль протейных в растительном мире Капского флористического царства, где они поражают множеством форм, разнообразием листьев и исключительной красотой соцветий. Наиболее характерны виды левкадендрона (Leucadendron) и протеи (Protea). Значительно меньшую роль играют протейные в тропической Африке, но и здесь они местами принимают заметное участие в растительности, например в Северной Гвинее. Довольно заметную роль играют протейные на Мадагаскаре. Но наиболее характерны они для растительного мира Австралии, особенно для Юго-Западной Австралии, Квинсленда, Нового Южного Уэльса и Тасмании. Здесь бросаются в глаза многочисленные виды гревиллеи (Grevillea), виды эндемичного рода банксия (Banksia) и многие другие. В Юго-Западной Австралии обычны виды эндемичного рода дриандра (Dryandra), так же как виды австралийско-тасманийского рода хакея (Hakea). Значительна роль протейных также в растительном мире Новой Каледонии. Что касается Азии и Америки, то протейные играют там менее заметную роль.
Протейные чаще всего кустарники и кустарнички, реже деревья, иногда очень высокие. Есть среди них также полукустарники и даже корневищные травы. Для многих протейных характерна жестколистность. Нередки среди них относительно мелколистные формы. Весь облик большинства протейных говорит о приспособлении к временной сухости муссонного климата и к бедным почвам. Как у южноафриканских, так и у австралийских протейных часто имеются так называемые лигнотуберусы — деревянистые подземные запасающие органы стеблевого происхождения. Однако в дождевых лесах встречаются деревья и с более или менее мезоморфными листьями. Протейные ясно выраженного жестколистного характера чаще всего кустарники, но бывают среди них и исключения. Многие типично жестколистные виды произрастают на почвах, бедных питательными веществами, особенно фосфором. Листья протейных характеризуются исключительно широким размахом изменчивости как размеров, формы и строения, так и типов жилкования. Не случайно Линней назвал род протея именем Протея, который, согласно древнегреческому мифу, мог по желанию обрести любую форму и любой образ.
Цветки
Цветки протейных обычно в различного рода бокоцветных соцветиях: кистях, зонтиках, колосьях или головках (простых или сложных), редко одиночные в пазухах листьев (результат редукции соцветия). Очень характерна ясно выраженная в семействе тенденция к образованию головчатых соцветий, часто внешне удивительно напоминающих соцветия многих сложноцветных. Головчатые соцветия возникают независимо у разных представителей семейства. Особенно характерны такие соцветия для рода протея в Южной и тропической Африке и родов телопеа (Telopea), дриандра и других в Австралии. Цветки протейных актиноморфные или более или менее зигоморфные, обычно обоеполые, но иногда однополые и у некоторых родов, как, например, у левкадендрона, двудомные.
Околоцветник представлен венчиковидной, обычно сростнолистной, 4-членной чашечкой. Интересно, что в бутоне чашечка обычно трубчатая, но при распускании цветка чашелистики более или менее отделяются друг от друга и в той или иной степени загибаются книзу, а иногда закручиваются. Что касается венчика, то он редуцирован, хотя у разных родов в неодинаковой степени. Вместо нормально развитого венчика в цветке обычно имеется более или менее развитый 4-членный подпестичный круг желёзок или чешуек или же чашевидный диск, как правило, несущие секреторную функцию. Другими словами, лепестки у протейных в той или иной степени превращены в нектарники. В то время как в актиноморфных цветках можно видеть полный 4-членный круг желёзок, каждая из которых соответствует лепестку, в зигоморфных цветках круг этот неполный: абаксиальные желёзки исчезают и сохраняются только 3 или даже только 2 адаксиальные желёзки. Иногда этот процесс идет дальше, и в цветке не сохраняется никаких следов венчика. У австралийского рода франкландия (Franklandia) сохранилось еще определенное сходство с венчиком, но 4 лопасти его сростнолепестного венчика уже функционируют как нектарники. В тех случаях, когда лепестки сведены к желёзкам и чешуйкам, эти последние обычно более или менее сросшиеся между собой, что, возможно, свидетельствует о том, что венчик у протейных был сростнолепестный.
Тычинок всегда 4, чередующихся с чашелистиками, с широкими нитями, обычно более или менее (иногда почти полностью) приросшими к чашелистикам. Лишь редко нити бывают полностью свободными, как у относительно примитивного монотипного тасманийского рода беллендена (Bellendena). Пыльники иногда с ясно выраженным надсвязником, особенно хорошо развиты у монотипного новокаледонского рода бопреопсис (Beaupreopsis). Гинецей одноплодолистиковый, но, несмотря на общую довольно высокую специализиро-ванность цветка протейных, плодолистик относительно примитивный, еще в сущности кондупликатный. У некоторых родов, имеющих рудиментарный венчик, плодолистик на более ранней стадии своего развития еще не вполне замкнутый и в нижней своей части (в области завязи), а также в районе рыльца еще более или менее открытый. У некоторых протейных свободные брюшные поверхности несут железистые волоски, как это наблюдается у кондупликатных плодолистиков некоторых примитивных цветковых растений. Однако в строении плодолистика протейных ясно выражены также определенные признаки довольно высокой специализации. Столбик нитевидный и прямой или же согнутый, у большинства видов с различной формы косым или боковым рыльцем.
Функционирующая рыльцевая поверхность может быть ограничена маленькой точкой на верхушке столбика или в центре верхушечного диска. У многих протейных с наличием рудиментарного венчика завязь на ножке (гинофор), что характерно для орнитофильных видов. Семязачатков в каждом плодолистике от одного до многих. Плод — листовка, орешек, крылатка или костянка. За исключением рода беллендена, семена без эндосперма. Цветки протейных обычно протандричны. Опыление производится различными представителями животного мира — насекомыми, птицами (нектарниками, белоглазками, австралийско-новогвинейскими медососами и родственными им южноафриканскими сахарными птицами и американскими колибри), рукокрылыми (крыланами и летучими мышами) и мелкими нелетающими млекопитающими (грызунами, мелкими сумчатыми из семейства лазящих сумчатых, или кускусов,— Phalangeridae).
Опыление протейных
У протейных наблюдается довольно большое разнообразие приспособлений к перекрестному опылению, особенно хорошо изученное у австралийских протейных. Так, у австралийского рода ламбертия (Lambertia) пыльца откладывается в бутоне на клейкой верхней части столбика. У другого австралийского протейного - петрофилы окрашенной (Petrophila fucit'olia) - оседанию пыльцы на столбике способствуют покрывающие ее короткие волоски, представляющие собой, таким образом, аналог «пыльцеснимателя» колокольчиковых (см. ниже, семейство колокольчиковых). У всех этих растений рыльце созревает и бывает готово к восприятию пыльцы лишь после окончания мужской фазы, когда большая часть пыльцы уже выгружена опылителями или засохла.
У гревиллеи, хакеи, телопеи, ломатии и некоторых других родов механизм опыления несколько иной. Верхушка столбика у них уплощена и расширена в форме прямого, косого или бокового диска, в центре которого в виде маленькой выпуклости расположено рыльце. Пыльники вскрываются в бутоне, и пыльца откладывается на дисковидной верхушке столбика, в том числе и на самом рыльце, которое на этой мужской стадии еще совершенно не восприимчиво к ней. Некоторое время дисковидная верхушка столбика остается зажатой в верхней части еще полностью не раскрывшегося околоцветника. У некоторых протейных, например у рода протеи, диск не высвобождается до тех пор, пока этой части цветка не коснется опылитель. Но уже при малейшем прикосновении сразу же срабатывает этот удивительный триггерный механизм, в котором столбик играет роль пружины изгиба, и диск с силой вырывается из объятий околоцветника и разбрасывает накопившуюся на нем пыльцу, осыпая ею тело опылителя. Благодаря этому механизму достигается двоякая цель: до высвобождения диска пыльца бывает защищена от превратностей внешнего мира, а внезапное ее высвобождение способствует разбрасыванию пыльцы. После освобождения диска начинается женская стадия — крошечное рыльце в центре диска увеличивается в размере и становится готовым к восприятию пыльцы, принесенной опылителем с другого цветка.
Совершенно иную модель механизма опыления наблюдается у некоторых других родов. Особенно интересны в этом отношении австралийские роды симфионема (Symphyonema) и коноспермум (Conospermum). В мелких желтых цветках симфионемы пыльники расположены тесно вокруг столбика. Когда пыльники созревают, сегменты околоцветника закручиваются назад. В поисках нектара опылитель (в данном случае обычно бражники) обязательно коснется пыльников, которые внезапно отделяются друг от друга, разбрасывая при этом пыльцу во всех направлениях. Часть этой пыльцы попадает и на опылителя.
Очень многие протейные приспособлены к опылению исключительно или главным образом птицами, особенно в Австралии и Южной Африке. Птицы опыляют виды протейных с крупными красными, оранжевыми, желтыми или желтовато-зелеными, часто контрастно.окрашенными верхушечными соцветиями, отдельные цветки которых обильно выделяют нектар и обычно лишены запаха, столь важного для привлечения насекомых и млекопитающих. Цветки некоторых видов банксии выделяют так много нектара, что он используется в качестве пищи аборигенами Австралии. Взаимное приспособление протейных и птиц в некоторых случаях принимает форму взаимной зависимости. Классическим примером является южноафриканская сахарная птица промеропс (Promerops cafer), которая настолько приспособилась к опылению некоторых видов протеи, что почти не может существовать без этих растений. У сахарной птицы клюв и язык длинные и тонкие, а соцветия этих видов протеи, окруженные плотно прилегающими друг к другу листочками обертки, представляют собой как бы естественную чашу, из которой птица пьет нектар. Сахарная птица настолько сжилась с видами протеи, что даже гнезда свои строит из листочков обертки и из старых цветков, заключающих зрелые семена, и тем самым содействует диссеминации.
Над цветками протейних трудятся самые разные насекомые, птицы и даже млекопитающие. Летающими млекопитающими (крыланами и летучими мышами) тропические растения опыляются нередко, но протейные смогли приспособиться к опылению и нелетающими зверушками. Австралийские виды банксии опыляются поссумами-медоедами — маленькими сумчатыми млекопитающими. Африканские протейные опыляют грызуны — местные виды крыс и полевых мышей. Растения привлекают зверюшек сладким нектаром, а также сочными частями цветков. Эти крошки переносят на своей шёрстке пыльцу с одного соцветия на другие, осуществляя перекрёстное опыление.
Как бы ни были малы зверьки-опылители, они всё же не подлетают к цветам, а карабкаются по соцветиям и неизбежно губят чать цветов. Но их тысячи в одном соцветии, и растение готово пожертвовать некоторыми чтобы опылить остальные.
Множество удивительных представителей протейных растёт в Капской провинции на юге Африки – узком поясе прибрежных гор. Ландшафт данной провинции разнообразен и популяции естественным образом отделенные друг от друга горными грядами и прочими эколого-географическими факторами, формировали в процессе эволюции значительное видовое разнообразие Капской флоры. Так, например, в данной области произрастает около 100 видов необычайно красивых растений из рода протея (Protea). Это название роду дал Карл Линней (по имени древнегреческого морского божества Протея, который мог по своему желанию принимать разный облик ) из-за большого разнообразия протей: не только разные их виды сильно отличаются друг от друга по внешнему облику, но и листья одного экземпляра могут быть различными по форме.
Самая эффектная из протей - протея артишоковая (P. cynaroides) с крупными (до 30 см в диаметре) головчатыми соцветиями, которые окружены яркими обёртками. Местное население называет это растение "протея-король". Другое название протеи артишоковой - "горшок меда", так как её крупные соцветия наполнены сладким нектаром (его африканцы используют как лекарство от кашля).
biofile.ru
ПРОТЕЙНЫЕ
ПРОТЕЙНЫЕпорядок (Proteales) двудольных растений и единств, его сем. (Proteaceae). П. занимают изолированное положение в системе и их филогенетич. связи не вполне ясны; возможно, что они происходят от предков камнеломковых. Деревья и кустарники, редко кустарнички с цельными или многораздельными листьями. Цветки обычно обоеполые, 4-членные, безлепестные, с венчиковидной чашечкой, часто в крупных головчатых соцветиях, похожих на соцветия сложноцветных, редко одиночные. Гинецей апокарпно-мономерный; завязь полунижняя. Семена с прямым зародышем, без эндосперма. Цветки протандричны, опыляются насекомыми, птицами, летучими мышами, мелкими нелетающими сумчатыми, грызунами. Ок. 1050 видов (по др. данным, до 1400), объединённых в 60 родов, гл. обр. в Юж. полушарии, особенно в Австралии и Юж. Африке. Большинство видов — ксерофиты. Наиб, крупные роды — (Grevillea) (св. 200 видов), (Nakea) (св. 100 видов), (Protea) (ок. 130 видов), (Leucodendron) (ок. 70 видов). Нек-рые виды дают ценную древесину, другие — съедобные семена. Мн. виды из родов (Banksia, Grevillea и др.) разводят как декоративные (в СССР — только в оранжереях).

Протейные. 1 — гревилея мощная (Grevillea robusta): а — ветвь с листьями, б — соцветие, в — бутон (столбик в виде пружины), г — плоды; 2 — протея исключительная (Protea eximia), растение с соцветием; 3 — банксия ярко-красная (Banksia coccinea): a — цветок, б — бутон, в — ветвь с соцветием; 4 — дриандра северная (Dryandra arctotidis): а — цветок.
.(Источник: «Биологический энциклопедический словарь.» Гл. ред. М. С. Гиляров; Редкол.: А. А. Бабаев, Г. Г. Винберг, Г. А. Заварзин и др. — 2-е изд., исправл. — М.: Сов. Энциклопедия, 1986.)
Поделитесь на страничкеslovar.wikireading.ru





