
Содержание
Обмен веществ у растений | 6 класс
Содержание
В этом уроке поговорим про обмен веществ у растений, двумя важнейшими процессами которого являются фотосинтез и дыхание.
Что такое обмен веществ
Для всех процессов жизнедеятельности растений — роста, цветения, плодоношения, размножения и других — нужны органические вещества.
Эти вещества используются как «строительный материал» клетки и ее компонентов и как специальные «механизмы», которые превращают одни вещества в другие и выполняют прочую работу.
Все они («строительный материал» и «механизмы») построены из углерода (C), водорода (H) и кислорода (O) — атомов, которые присутствуют в углекислом газе (CO2) и воде (H2O). А также из макро- и микроэлементов.
1 — внешняя среда; 2 — мембрана; 3 — цитоплазма; 4 — микро- и макроэлементы; 5 — органические молекулы, из которых построены компоненты клетки; 6 — клеточные «механизмы», которые занимаются превращением веществ
Атомы углерода и кислорода растения получают из углекислого газа воздуха во время фотосинтеза.
А атомы водорода и макро-/микроэлементы — с водой (бо́льшая часть которой испаряется) из почвы во время минерального питания.
В процессе фотосинтеза образуются углеводы, из которых синтезируются все остальные органические вещества, необходимые растению.
Энергию растения получают во время дыхания при разложении органических веществ с помощью кислорода.
Органические вещества при этом проходят через серию многочисленных химических реакций, в процессе которых образуются промежуточные продукты.
I — минеральное питание; II — фотосинтез; III — дыхание; IV — остальные химические реакции и процессы жизнедеятельности
Промежуточные продукты тоже участвуют в разных клеточных процессах.
Все эти многочисленные химические реакции, все эти превращения веществ друг в друга называются обменом веществ.
Обмен веществ — это все процессы в организме, во время которых происходит разложение одних веществ и синтез других.
Важнейшие процессы обмена веществ
Обмен веществ обеспечивается дыханием и фотосинтезом.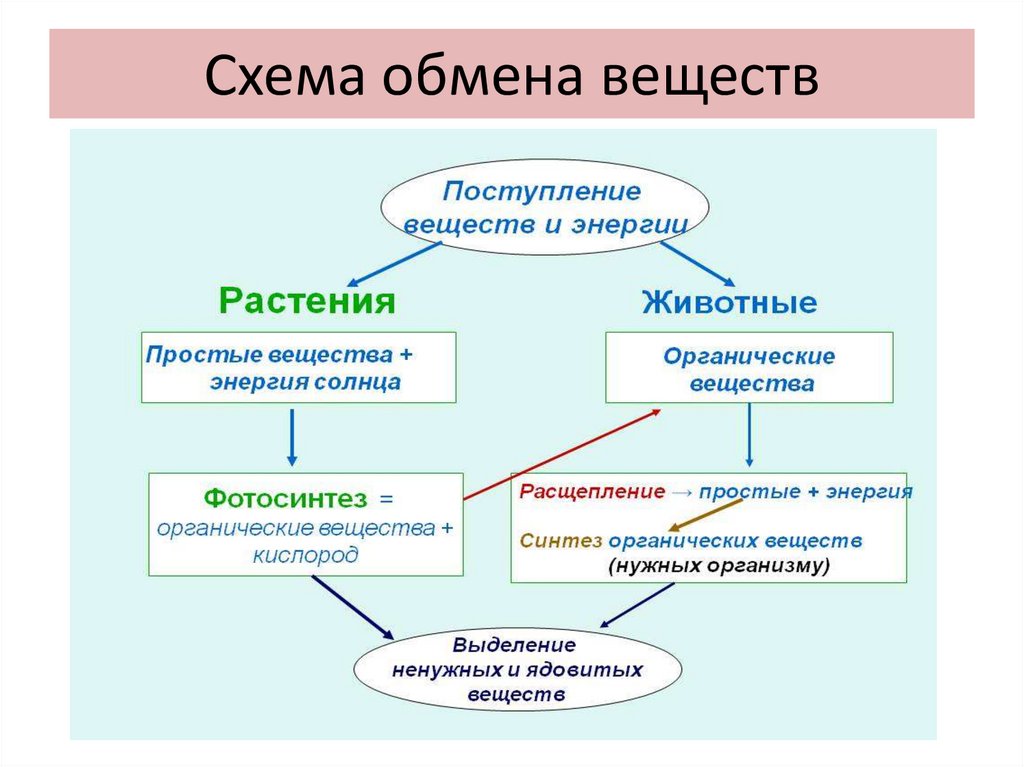 Это два противоположных, но взаимосвязанных процесса.
Это два противоположных, но взаимосвязанных процесса.
Важнейшими процессами обмена веществ являются дыхание и фотосинтез.
| Название процесса | Суть процесса |
| Фотосинтез | Поглощение углекислого газа и воды и синтез органических веществ, в химических связях которых запасается поглощенная хлорофиллом энергия. Этот процесс сопровождается выделением кислорода. |
| Дыхание | Поглощение кислорода и разложение с его помощью органических веществ, что высвобождает накопленную в них энергию. При этом выделяется углекислый газ и вода. |
Зеленый овал — хлоропласт, в котором протекает фотосинтез, красный — митохондрия, в которой проходят некоторые стадии дыхания
Укажем в таблице черты сходства и различия процессов фотосинтеза и дыхания.
| Черты сходства | Черты различия |
Дыхание и фотосинтез — важнейшие процессы обмена веществ, которые сопровождаются поглощением из окружающей среды одних газов и выделением других. В ходе обоих процессов участвует вода, и происходит превращение энергии. | Фотосинтез протекает в зеленых частях растения, в хлоропластах, на свету. Из неорганических веществ синтезируются органические, в химических связях которых накапливается энергия. Дыхание протекает во всех частях растения, в цитоплазме и митохондриях, круглосуточно. |
Какие вещества участвуют в двух взаимосвязанных процессах обмена веществ у растений — дыхания и фотосинтеза? Как они образуются?
Показать ответ
Скрыть ответ
В обоих процессах участвуют:
- неорганические вещества: углекислый газ, вода и кислород;
- органические вещества, в основном углеводы.
Во время фотосинтеза из углекислого газа и воды образуются органические вещества и кислород.
А во время дыхания из органических веществ с помощью кислорода образуются углекислый газ и вода.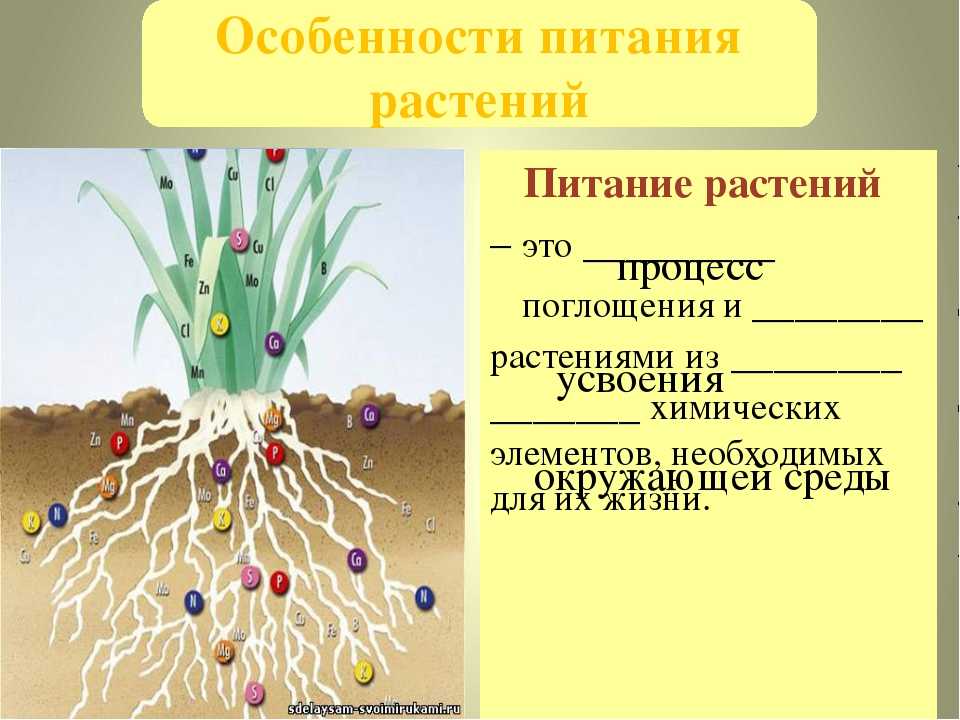
{"questions":[{"content":"Что происходит во время фотосинтеза, дыхания и минерального питания?[[grouper-17]]","widgets":{"grouper-17":{"type":"grouper","labels":["Фотосинтез","Дыхание","Минеральное питание"],"items":[["поглощение углекислого газа","выделение кислорода","синтез углеводов"],["поглощение кислорода","выделение углекислого газа","распад углеводов"],["поглощение воды","поглощение микро- и макроэлементов"]]}}}]}Роль обмена веществ в жизни растений
Благодаря обмену веществ поддерживается жизнь растения, его рост, развитие.
В процессе обмена веществ растение получает необходимые строительные блоки, из которых может образовывать новые клетки и ткани, а также энергию, за счет которой происходит это образование.
Движение воды с растворенными в ней солями — голубые стрелки, движение продуктов фотосинтеза — фиолетовые
Продукты фотосинтеза (органические вещества) по проводящим путям доставляются во все органы, во все части растения — к корням, стеблям, цветкам, плодам и др.
А там эти органические вещества расщепляются с высвобождением энергии и промежуточных продуктов обмена, которые используются для нужд конкретного органа.
1 — молекула органического вещества; 2 — цепочка химических реакций; 3 — промежуточные продукты дыхания
Так с помощью обмена веществ, обеспечивается взаимосвязь всех частей растения.
Когда растение погибает, в нем прекращается обмен веществ. И наоборот, с прекращением обмена веществ растение погибает.
Опишите кратко процессы превращения энергии при фотосинтезе, дыхании, при расщеплении в клетках органических веществ на неорганические и при образовании новых клеток.
Показать ответ
Скрыть ответ
При фотосинтезе — энергия Солнца превращается в химическую энергию и запасается в форме органических веществ.
При дыхании — энергия высвобождается с разрушением химических связей органических веществ.
При расщеплении в клетках органических веществ на неорганические — происходит выделение энергии.
При образовании новых клеток — тратится энергия, запасенная в органических веществах во время фотосинтеза и извлеченная из них во время дыхания.
{"questions":[{"content":"Во время дыхания энергия [[fill_choice-25]], а во время фотосинтеза энергия [[fill_choice-28]].","widgets":{"fill_choice-25":{"type":"fill_choice","options":["запасается","высвобождается"],"answer":1},"fill_choice-28":{"type":"fill_choice","options":["запасается","высвобождается"],"answer":0}}}]}Связь организма растения со средой
Для протекания обмена веществ растение поглощает нужные ему элементы из окружающей среды.
Из почвы во время минерального питания растение всасывает воду и растворенные в ней минеральные соли.
1 — проводящие пути растения; 2 — молекулы воды; 3 — макро- и микроэлементы
В форме минеральных солей в организм поступают макро- и микроэлементы, которые входят в состав многих органических веществ растения и участвуют во множестве клеточных процессов.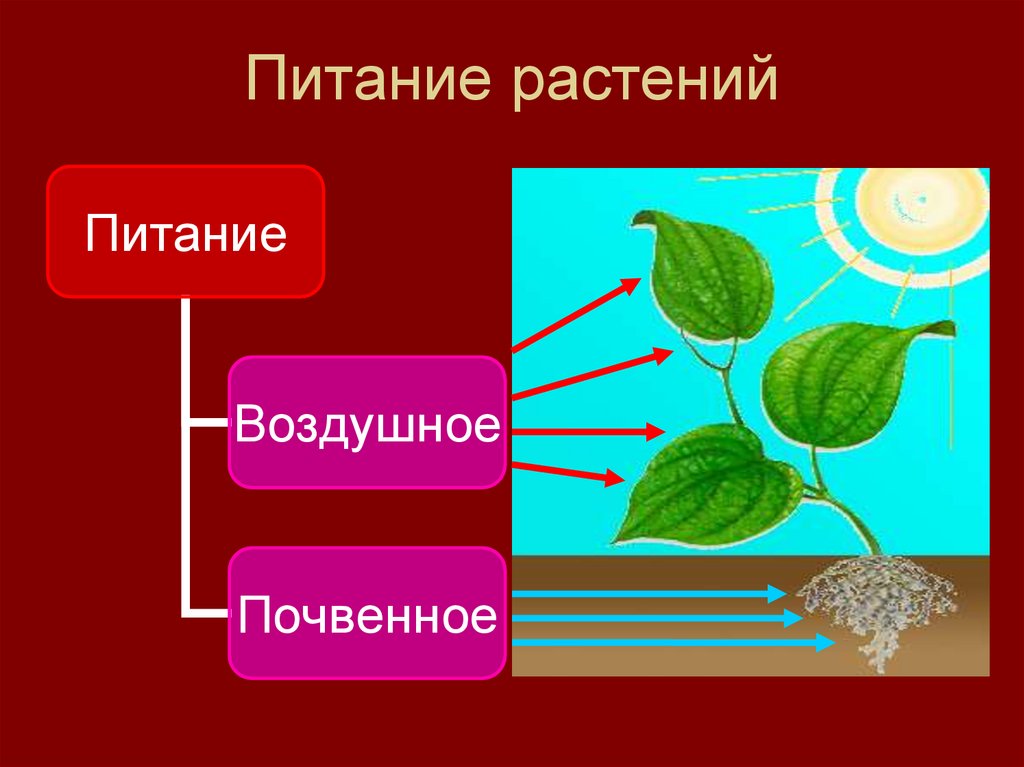
Вода обеспечивает ток этих элементов по растению и накопление их в тканях. Вода участвует и в процессе фотосинтеза.
Для фотосинтеза необходим также углекислый газ, который растение поглощает из воздуха. А побочный продукт фотосинтеза — кислород — растение выделяет обратно в атмосферу.
Чтобы осуществлять дыхание, растение поглощает из окружающей среды кислород и выделяет в нее углекислый газ.
Во время дыхания, а также испарения в окружающую среду постоянно выделяются молекулы воды.
Схема взаимосвязи организма растения со средой
Для всех процессов нужна энергия, источником которой является солнечный свет, поглощаемый хлорофиллом листьев тоже из окружающей среды.
Таким образом, обмен веществ обеспечивает взаимодействие растения с окружающей средой.
Обмен веществ обеспечивает взаимосвязь всех частей растения и связь организма со средой.
Решите биологическую задачу
Весной у березы, сделав разрез в коре, часто берут сладкий березовый сок.
1. От восходящего или нисходящего тока берут этот сок?
2. Что произойдет с березой, если у нее взять очень много сока?
Показать ответы
Скрыть ответы
1. Для ответа на этот вопрос нужно вспомнить строение стебля, функции проводящих тканей и роль фотосинтеза.
Верхний слой стебля — кора, состоит из корки и луба. Луб — это проводящая ткань, по которой переносятся продукты фотосинтеза от листьев к остальным органам растения.
Продуктами фотосинтеза в основном являются углеводы (сахара), которые переносятся в разные части растения для их участия в обмене веществ и дыхании.
Ток продуктов фотосинтеза называется нисходящим. Поэтому ответ: от нисходящего тока.
2. Если продукты, необходимые для обмена веществ и для получения жизненно важной энергии, перестанут поступать в нужном количестве, организм, скорее всего, погибнет.
{"questions":[{"content":"Отметьте правильные утверждения.[[choice-12]]","widgets":{"choice-12":{"type":"choice","options":["Дыхание происходит в клетках всех органов непрерывно только на свету. ","При дыхании поглощается кислород, а выделяется углекислый газ. При фотосинтезе — наоборот.","В процессе дыхания сложные органические вещества распадаются, при этом высвобождается химическая энергия."],"answer":[1,2]}}}]}
","При дыхании поглощается кислород, а выделяется углекислый газ. При фотосинтезе — наоборот.","В процессе дыхания сложные органические вещества распадаются, при этом высвобождается химическая энергия."],"answer":[1,2]}}}]}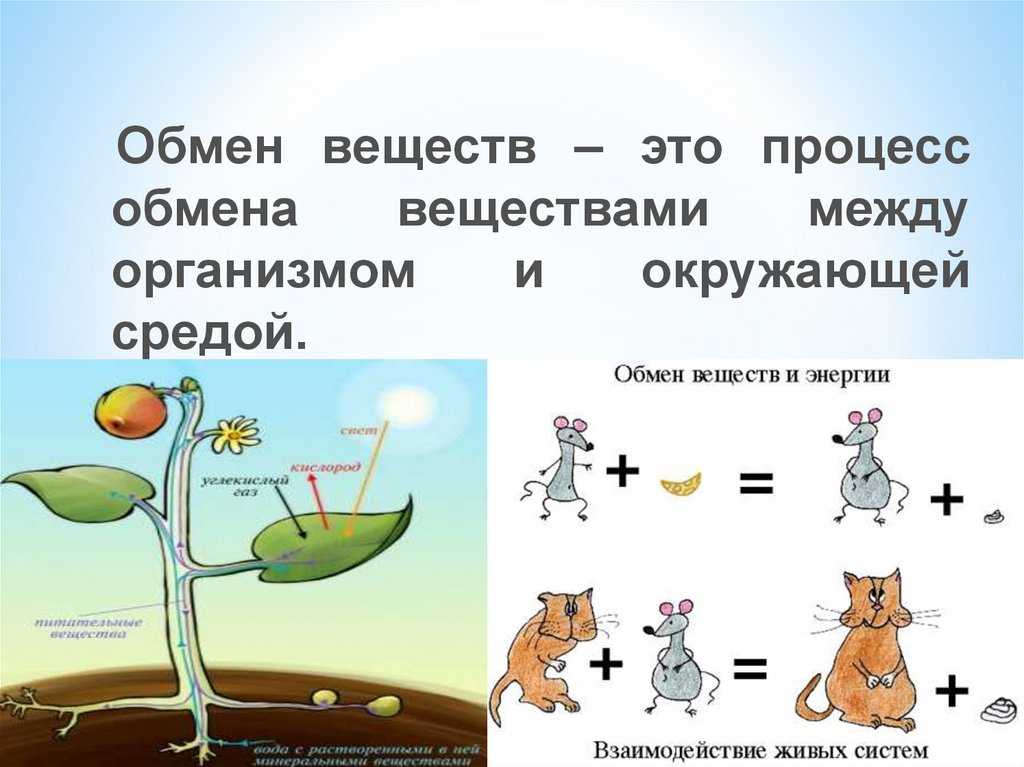 ","При дыхании поглощается кислород, а выделяется углекислый газ. При фотосинтезе — наоборот.","В процессе дыхания сложные органические вещества распадаются, при этом высвобождается химическая энергия."],"answer":[1,2]}}}]}
","При дыхании поглощается кислород, а выделяется углекислый газ. При фотосинтезе — наоборот.","В процессе дыхания сложные органические вещества распадаются, при этом высвобождается химическая энергия."],"answer":[1,2]}}}]}Выделение у растений и животных
ВСПОМНИТЕ
Вопрос 1. Как растения удаляют ненужные вещества?
У растений нет специальной выделительной системы. Многие ненужные для них вещества откладываются в клетках, многоклеточных вместилищах и сохраняются на протяжении всей их жизни. От многих вредных веществ растения освобождаются во время листопада, при слущивании наружных слоёв коры — корки.
Некоторые растения от избытка воды и солей освобождаются через специальные, похожие на устьица группы клеток — водяные устьица, или гидатоды. Обычно они располагаются на верхушке и по краям листа. Выделяемые ими капли (росу) можно видеть на листьях утром в жаркую погоду.
Вопрос 2. Как удаляются ненужные продукты жизнедеятельности у животных?
Животные по-разному избавляются от продуктов обмена. Например, амёба от излишков воды избавляется с помощью сократительной вакуоли, которая, периодически сокращаясь, выталкивает наружу находящуюся в ней жидкость. Всей поверхностью тела удаляются ненужные вещества у гидр, медуз.
Вопрос 1. Где у растений накапливаются продукты обмена веществ?
У растений продукты обмена веществ накапливаются в вакуолях клеток, в специальных хранилищах, например в смоляных ходах у хвойных, млечных ходах у одуванчика и молочая. У многолетних растений они накапливаются в коре, иногда в древесине. Удаление продуктов жизнедеятельности у растений происходит через корни и опавшие листья. Установлено, что к осени в клетках листьев накапливаются вредные для растения вещества, которые удаляются из растения вместе с опадающими листьями.
Вопрос 2. Как происходит выделение вредных веществ у растений?
Как происходит выделение вредных веществ у растений?
Через устьица и чечевички коры, например березы, из растения удаляется углекислый газ.
Выделение сахаров у растений осуществляется специальными образованиями — нектарниками. У большинства растений они находятся в цветках, а у некоторых — на стеблях и листьях. Нектар обладает бактерицидными свойствами и защищает завязь цветка от микроорганизмов. К тому же нектар наряду с окрашенным венчиком и ароматом цветков является важным приспособлением для привлечения насекомых, осуществляющих перекрёстное опыление.
Через специальные железы растений в атмосферу выделяются летучие вещества, в том числе эфирные масла. К эфиромасличным растениям относятся пеларгония, мята, мелисса, эвкалипт. Многие из них используются в лекарственных целях, а также для ароматизации продуктов, изготовления парфюмерной продукции.
Опавшие листья растений содержат неорганические и органические вещества и представляют собой очень ценное удобрение.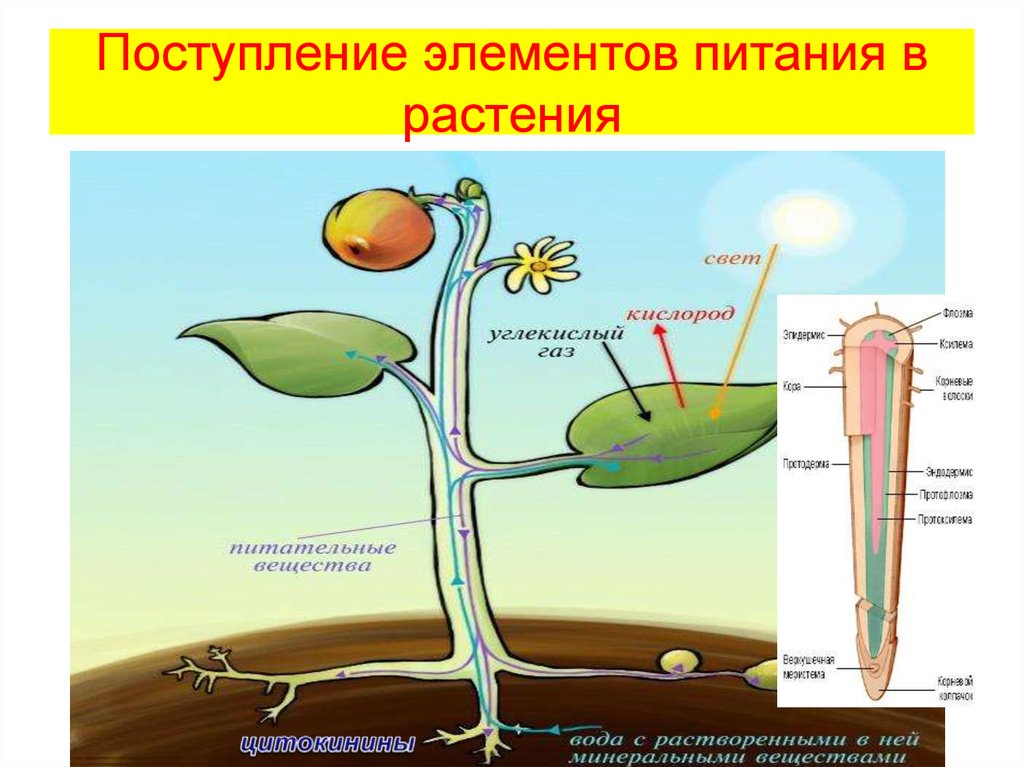 Поэтому садоводы закладывают листья в компостные кучи. Благодаря опавшим листьям почва в лесу ежегодно обогащается перегноем. Вот почему их не надо жечь. Вполне понятно, что сбор опавших листьев и вообще удаление лесной подстилки в лесу отрицательно сказываются па жизни деревьев.
Поэтому садоводы закладывают листья в компостные кучи. Благодаря опавшим листьям почва в лесу ежегодно обогащается перегноем. Вот почему их не надо жечь. Вполне понятно, что сбор опавших листьев и вообще удаление лесной подстилки в лесу отрицательно сказываются па жизни деревьев.
Вопрос 3. Какие продукты обмена веществ выделяются из организма позвоночных животных через лёгкие, кишечник, потовые железы?
Через легкие выводятся из организма углекислый газ вода и летучие вещества. Кишечник выделяет некоторые соли в составе кала, потовые железы — воду, соли, органические вещества.
1. Выпишите из текста параграфа новые понятия, найдите их определения в интернет-источниках, энциклопедических словарях.
Нектарники— медовые желёзки растений, выделяющие сахаристый сок — нектар. Внешние выделительные структуры, обычно расположенные в цветке и выделяющие сладкий сок, служащий приманкой для животных-опылителей, чаще всего насекомых.
Нефридии — органы выделения у беспозвоночных, выполняющие функции осморегуляции, извлечения и удаления из организма вредных продуктов обмена веществ.
Листопад — биологический процесс сбрасывания листвы растениями. В условиях умеренного климата зимой многим растениям не хватает воды. Вода в замёрзшем грунте находится в состоянии льда и не может проникать в клетки корней.
2. Используя интернет-источники, научно-популярную литературу, подготовьте сообщение на тему «Значение процессов выделения у живых организмов».
В процессе обмена веществ в клетках образуются конечные продукты. Среди них могут быть и ядовитые для клеток вещества. Так, при расщеплении аминокислот, нуклеиновых кислот и других азотсодержащих соединений образуются токсические вещества —аммиак, мочевина и мочевая кислота, которые по мере их накопления подлежат выведению из организма. Должны удаляться» кроме того, избыток воды, углекислый газ, яды, которые поступают вместе с вдыхаемым воздухом, поглощаемой пищей и водой, избыток витаминов, гормонов, лекарственные препараты и т.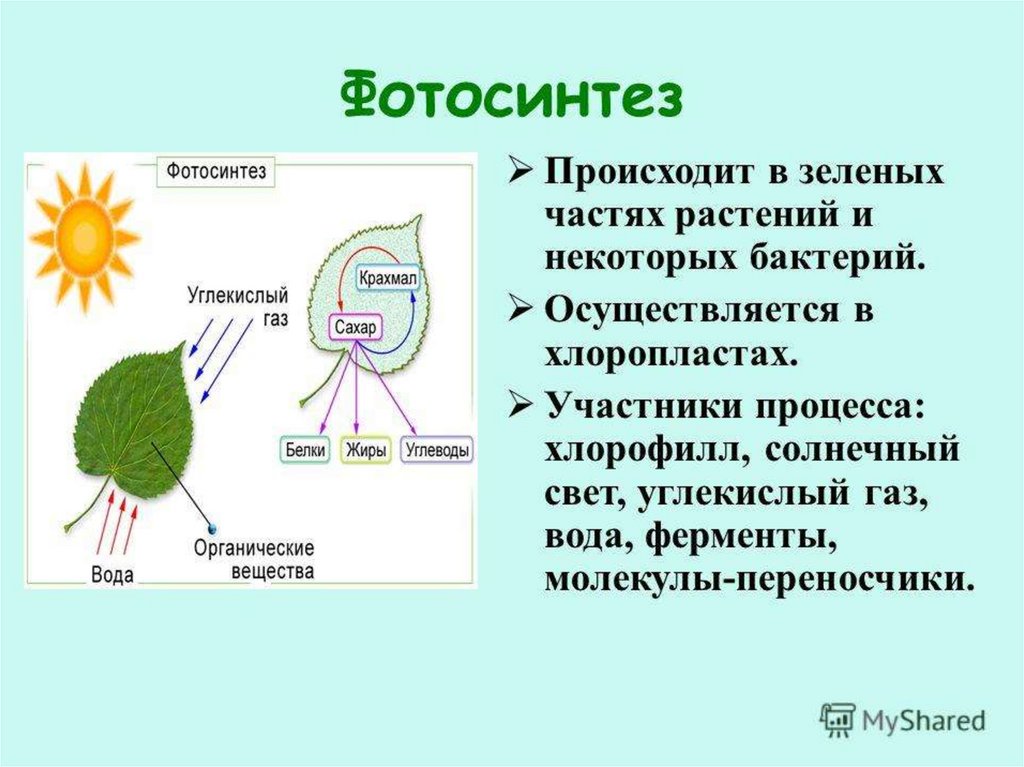 п. При накоплении этих веществ в организме возникает опасность нарушения постоянства состава и объема внутренней среды организма, что может отразиться на здоровье человека.
п. При накоплении этих веществ в организме возникает опасность нарушения постоянства состава и объема внутренней среды организма, что может отразиться на здоровье человека.
Органы выделения и их функции. Выделительную функцию выполняют многие органы. Так, легкие выводят из организма углекислый газ, пары воды, некоторые летучие вещества, например пары эфира, хлороформа при наркозе, пары алкоголя при опьянении. Потовыми железами удаляются вода и соли, небольшие количества мочевины, мочевой кислоты, а при напряженной мышечной работе — молочная кислота. Слюнные и желудочные железы выделяют некоторые тяжелые металлы, ряд лекарственных веществ, чужеродные органические соединения. Важную экскреторную функцию выполняет печень, удаляя из крови гормоны (тироксин, фолликулин), продукты расщепления гемоглобина, азотистого метаболизма и многие другие вещества. Поджелудочная железа и кишечные железы выводят соли тяжелых металлов, лекарственные вещества.
Однако основная роль в процессах выделения принадлежит специализированным органам — почкам. К важнейшим функциям почек относится участие в регуляции: 1) объема крови и других жидкостей внутренней среды, 2) постоянства осмотического давления крови и других жидкостей тела, 3) ионного состава жидкостей внутренней среды и ионного баланса организма, 4) кислотно-щелочного равновесия, 5) выведения из организма конечных продуктов азотистого обмена и чужеродных веществ. Таким образом, почки являются органом, обеспечивающим гомеостаз внутренней среды организма.
К важнейшим функциям почек относится участие в регуляции: 1) объема крови и других жидкостей внутренней среды, 2) постоянства осмотического давления крови и других жидкостей тела, 3) ионного состава жидкостей внутренней среды и ионного баланса организма, 4) кислотно-щелочного равновесия, 5) выведения из организма конечных продуктов азотистого обмена и чужеродных веществ. Таким образом, почки являются органом, обеспечивающим гомеостаз внутренней среды организма.
ПОДУМАЙТЕ
Чем различается выделение веществ у растений и животных? Составьте сравнительную таблицу.
У животных есть органы выделения: у позвоночных — почки, у простейших — сократительная вакуоля. Органы выделения удаляют из организма мочевину, мочевую кислоту и аммиак — продукты распада разных веществ, в том числе и белка, У растений сходных отходов образуется немного. Это объясняется типом их питания и отсутствием мышечной активности. Из растительных организмов аммиак не выводится.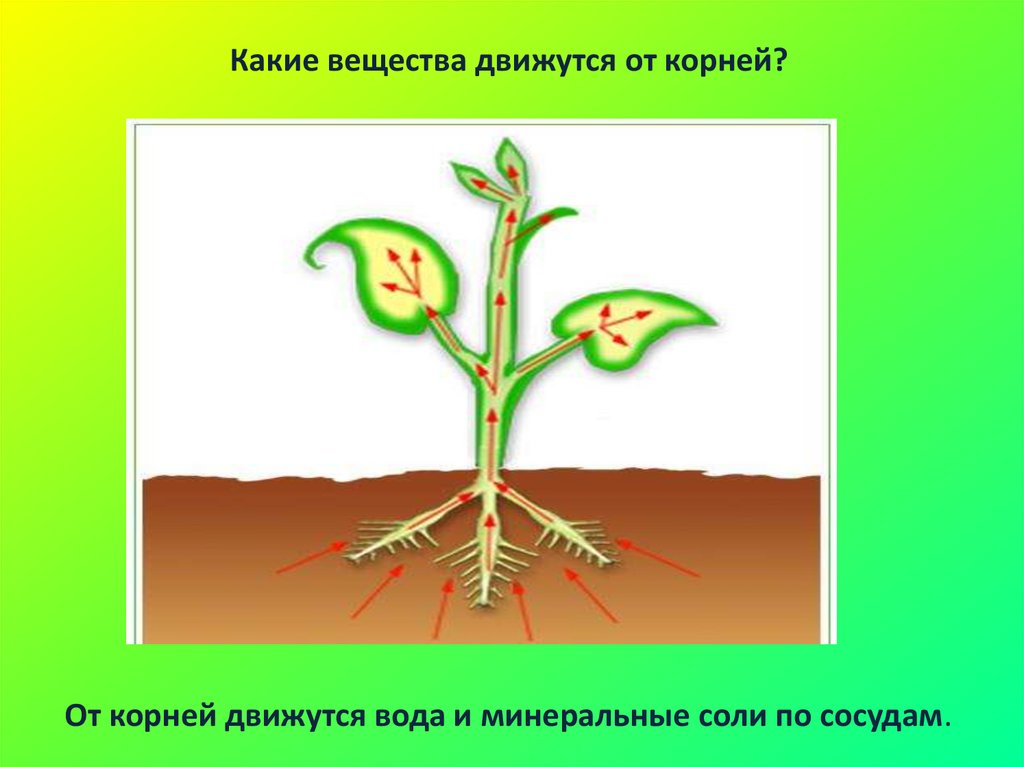 Путем химических превращений он переходит в безвредные для растения вещества — аспарагин и глутамин. Это кладовые азота, который может быть использован растением вновь при биосинтезе азотсодержащих соединений. Некоторые продукты жизнедеятельности накапливаются у растений в листьях и удаляются при листопаде.
Путем химических превращений он переходит в безвредные для растения вещества — аспарагин и глутамин. Это кладовые азота, который может быть использован растением вновь при биосинтезе азотсодержащих соединений. Некоторые продукты жизнедеятельности накапливаются у растений в листьях и удаляются при листопаде.
Растения перерабатывают специализированные продукты метаболизма, чтобы обеспечить питательные вещества для первичного метаболизма
Пожалуйста, включите JavaScript в вашем браузере.
10 августа 2021 г.
Обзор исследований
Биология
Растения расщепляют сложные продукты метаболизма для восстановления и повторного использования питательных веществ
Рисунок 1: Кресс-салат талевый ( Arabidopsis thaliana ) способен перерабатывать специализированные продукты метаболизма для извлечения питательных веществ, необходимых для основного метаболизма. © MAURO FERMARIELLO/НАУЧНАЯ ФОТОБИБЛИОТЕКА
Растения могут повторно использовать специализированные продукты метаболизма и расщеплять их для извлечения питательных веществ, необходимых для основных метаболических процессов, обнаружили ученые RIKEN 1 .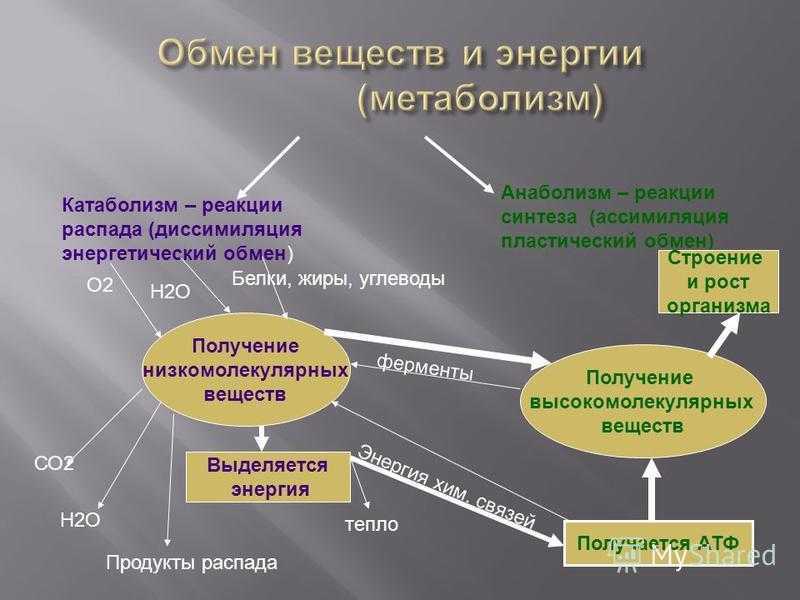 Деактивация фермента, ответственного за этот механизм, может улучшить производство полезных для здоровья химических веществ в сельскохозяйственных культурах в будущем.
Деактивация фермента, ответственного за этот механизм, может улучшить производство полезных для здоровья химических веществ в сельскохозяйственных культурах в будущем.
В дополнение к производству сахаров и других молекул, непосредственно участвующих в их росте и развитии, растения производят ряд вторичных метаболитов — специализированных молекул, которые помогают им адаптироваться к окружающей среде и взаимодействовать с их экосистемой, например, обеспечивая защиту от поедания растений животные. Ученые давно задавались вопросом, могут ли питательные вещества, используемые для синтеза вторичных метаболитов, высвобождаться и повторно использоваться в первичном метаболизме.
Теперь команда под руководством Масами Хираи из Центра устойчивого ресурсоведения RIKEN изучила этот вопрос, изучив, может ли кресс-салат Thale (рис. 1) использовать вторичный метаболит глюкозинолат в качестве источника серы. Глюкозинолаты частично отвечают за остроту таких овощей, как горчица и хрен, а также за питательную ценность и пользу для здоровья брокколи и брюссельской капусты.
Команда выращивала растения в культурах без серы, с разным содержанием серы или с глюкозинолатом. Растения, выращенные без серы, имели серьезные дефекты роста, тогда как растения, выращенные с глюкозинолатом, демонстрировали нормальный рост. Это говорит о том, что растения использовали сульфат в поставляемом глюкозинолате в качестве альтернативного источника серы.
Рёске Сугияма (слева), Масами Хираи (справа) и их коллеги показали, что растения могут перерабатывать специальные продукты метаболизма, чтобы повторно использовать содержащиеся в них питательные вещества в основных метаболических процессах. © 2021 РИКЕН
На самом деле измерения концентрации серы показали, что проростки накопили больше серы, чем растения, выращенные с таким же количеством сульфата. Это заставило команду заподозрить, что растения также использовали другие атомы серы (то есть несульфатные атомы серы) из глюкозинолата.
Чтобы исследовать это, они подготовили глюкозинолаты, в которых атом серы, образующий ключевую связь в молекуле, был более тяжелым изотопом, чтобы они могли отслеживать его в более поздних метаболических процессах. Команда обнаружила, что меченая сера составляет 28–42% различных серосодержащих аминокислот в проростках, что ясно демонстрирует расщепление поступающего глюкозинолата и его повторное включение в первичный метаболизм.
Команда обнаружила, что меченая сера составляет 28–42% различных серосодержащих аминокислот в проростках, что ясно демонстрирует расщепление поступающего глюкозинолата и его повторное включение в первичный метаболизм.
«Я был очень взволнован, увидев это», — говорит Рёске Сугияма, ведущий автор исследования. «Поскольку потенциальная роль глюкозинолата в качестве резервуара серы обсуждалась на основе сульфатной группы, мы были удивлены, что другой распространенный атом серы также доступен в качестве источника. Это подтверждает наличие систематического пути распада в растениях».
«Хотя потенциальная переработка вторичных метаболитов обсуждалась на протяжении десятилетий, прямых доказательств ее физиологических преимуществ не было получено», — добавляет Сугияма. «Наши результаты показывают, что важный вторичный метаболит перерабатывается растениями и, таким образом, может служить резервуаром питательных веществ для первичного метаболизма».
Связанное содержимое
- Обнаружение метаболитов отдельной клетки
- Что в этом растении? Лучшая автоматизированная система поиска потенциальных лекарств
- Овощи
- Как заводы работают с серой
Ссылка
- 1. Сугияма Р., Ли Р., Кувахара А., Накабаяши Р., Сотта Н., Мори Т., Ито Т., Окама-Оцу, Н., Фудзивара, Т., Сайто, К. и др. Ретроградный поток серы от глюкозинолатов к цистеину в Arabidopsis thaliana . Proceedings of the National Academy of Sciences USA 118 , e20178
(2021). doi: 10.1073/pnas.20178
 Сугияма Р., Ли Р., Кувахара А., Накабаяши Р., Сотта Н., Мори Т., Ито Т., Окама-Оцу, Н., Фудзивара, Т., Сайто, К. и др. Ретроградный поток серы от глюкозинолатов к цистеину в Arabidopsis thaliana . Proceedings of the National Academy of Sciences USA 118 , e20178
Сугияма Р., Ли Р., Кувахара А., Накабаяши Р., Сотта Н., Мори Т., Ито Т., Окама-Оцу, Н., Фудзивара, Т., Сайто, К. и др. Ретроградный поток серы от глюкозинолатов к цистеину в Arabidopsis thaliana . Proceedings of the National Academy of Sciences USA 118 , e20178Механизмы устойчивости к самопродуцируемым токсичным вторичным метаболитам в растениях
Ахмед М.С., Эйнли К., Пэриш Дж.Х., Хади С.М. (1994)Фрагментация белков кверцетином, индуцированная свободными радикалами. Канцерогенез 15:1627–1630
Статья
пабмед
КАС
Google ученый
Aimi N, Nishimura M, Miwa A, Hoshino H, Sakai S, Haginiwa J (1989) Пумилозид и дезоксипумилозид; возможные промежуточные продукты биосинтеза камптотецина. Тетраэдр Летт 30:4991–4994
Артикул
КАС
Google ученый
Alcantara J, Bird DA, Franceschi VR, Facchini J (2005) Биосинтез сангвинарина связан с эндоплазматическим ретикулумом в культивируемых клетках опийного мака после обработки элиситором.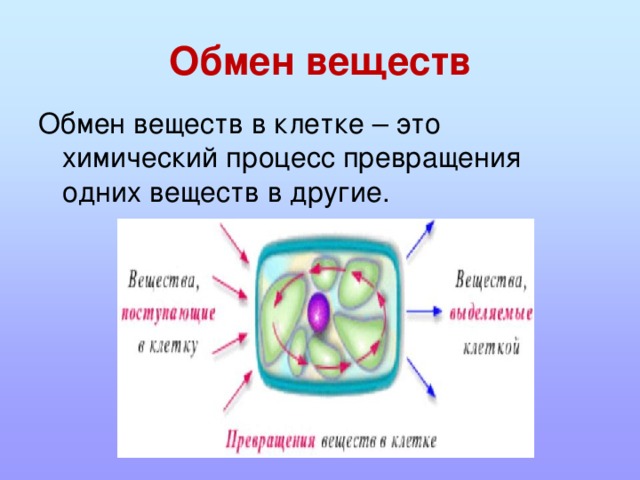 Завод Физиол 138:173–183
Завод Физиол 138:173–183
Статья
пабмед
КАС
Google ученый
Алфенито М.Р., Сауэр Э., Годман К.Д., Бьюэлл Р., Мол Дж., Коес Р., Уолбот В. (1998) Функциональное дополнение секвестрации антоцианов в вакуоли широко расходящейся глутатион-S-трансферазой. Растительная клетка 10:1135–1149
Статья
пабмед
КАС
Google ученый
Bailey JA, Carter GA, Burden RS, Wain RL (1975) Борьба с болезнями ржавчины с помощью дитерпенов из Nicotiana glutinosa . Природа 255:328–329
Статья
КАС
Google ученый
Bajaj NP, McLean MJ, Waring MJ, Smekal E (1990) Селективное по последовательности рН-зависимое связывание с ДНК бензофенантридиновых алкалоидов. J Mol Recognit 3:48–54
Статья
пабмед
КАС
Google ученый
Barnes P, Putnam AR, Burke BA (1986) Аллелопатическая активность ржи ( Secale злаки L. ). В: Патнэм А.Р., Танг К.С. (ред.) Наука об аллелопатии. Wiley-Interscience, Нью-Йорк, стр. 271–286 9.0003
). В: Патнэм А.Р., Танг К.С. (ред.) Наука об аллелопатии. Wiley-Interscience, Нью-Йорк, стр. 271–286 9.0003
Google ученый
Biggins JB, Onwueme KC, Thorson JS (2003) Устойчивость к эндииновым противоопухолевым антибиотикам путем самопожертвования CalC. Science 301:1537–1541
Статья
пабмед
КАС
Google ученый
Cicek M, Esen A (1998) Структура и экспрессия дхурриназы (бета-глюкозидазы) из сорго. Завод Физиол 116:1469–1478
Артикул
пабмед
КАС
Google ученый
Conn EE (1980) Цианогенные глюкозиды. Annu Rev Plant Physiol 31:433–451
Статья
КАС
Google ученый
Cundliffe E (1989) Как организмы, вырабатывающие антибиотики, избегают самоубийства. Annu Rev Microbiol 43:207–233
Статья
пабмед
КАС
Google ученый
Cutler HG, Reid W, Deletang J (1977) Ингибирующие рост растений свойства детерпена из табака.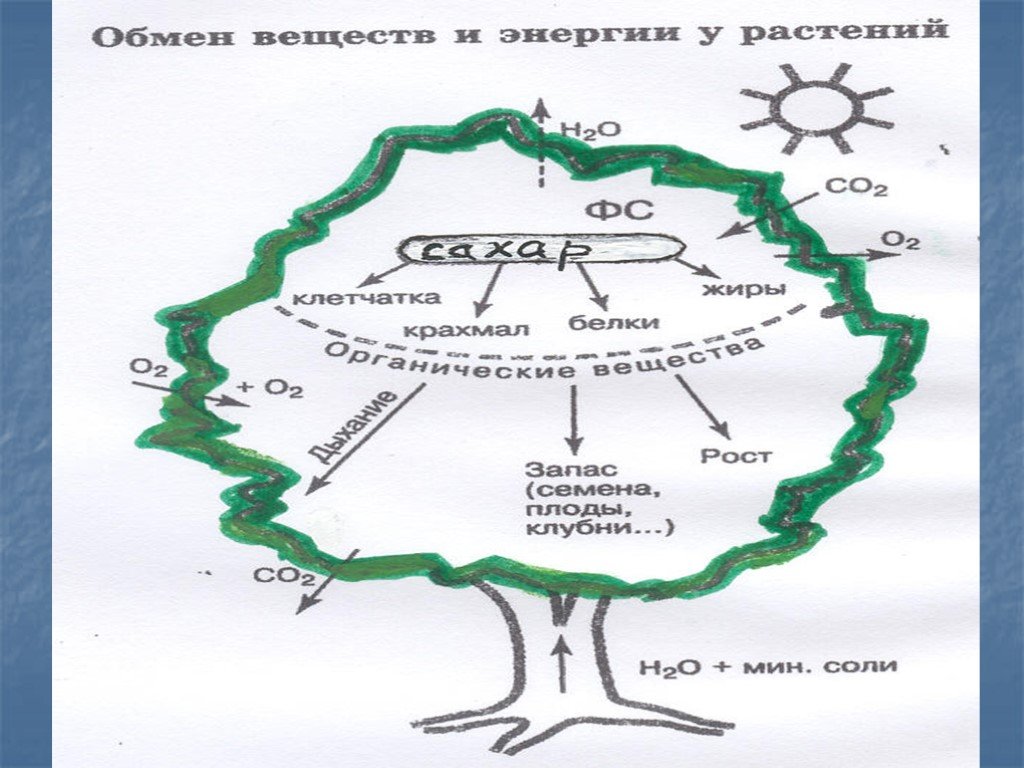 Физиология клеток растений 18:711–714
Физиология клеток растений 18:711–714
КАС
Google ученый
Debaujon I, Peeters AJ, Leon-Kloosterziel KM, Koornneef M (2001) Ген TRANSPARENT TESTA12 арабидопсиса кодирует мультилекарственный вторичный транспортер-подобный белок, необходимый для секвестрации флавоноидов в вакуолях эндотелия семенной кожуры. Растительная клетка 13:853–871
Статья
пабмед
КАС
Google ученый
Деус-Нойман Б., Зенк М.Х. (1984) Высокоселективная система поглощения алкалоидов в вакуолях высших растений. Планта 162:250–260
Артикул
КАС
Google ученый
Deus-Neumann B, Zenk MH (1984) Нестабильность продукции индольных алкалоидов в культурах суспензии клеток Catharanthus roseus . Планта Мед 50:427–431
Статья
пабмед
КАС
Google ученый
Деус-Нойман Б. , Зенк М.Х. (1986) Накопление алкалоидов в вакуолях растений не связано с механизмом ионной ловушки. Планта 167:44–53
, Зенк М.Х. (1986) Накопление алкалоидов в вакуолях растений не связано с механизмом ионной ловушки. Планта 167:44–53
Статья
КАС
Google ученый
Диксон Р.А., Стил С.Л. (1999) Флавоноиды и изофлавоноиды – золотая жила для метаболической инженерии. Trends Plant Sci 4:394–400
Статья
пабмед
Google ученый
Facchini PJ, St-Pierre B (2005) Синтез и торговля ферментами биосинтеза алкалоидов. Curr Opin Plant Biol 8: 657–666
Артикул
пабмед
КАС
Google ученый
Fairbairn JW (1972) Трихомы и железы Cannabis sativa L. Bull Narc 23:29–33
Google ученый
Glöckner C, Wolf H (1984) Механизм естественной устойчивости к кирромициновым антибиотикам у актиномицетов. FEMS Microbiol Lett 25:121–124
Статья
Google ученый
Goodman CD, Casati P, Walbot V (2004) Белок, ассоциированный с множественной лекарственной устойчивостью, участвующий в транспорте антоцианов у Zea mays .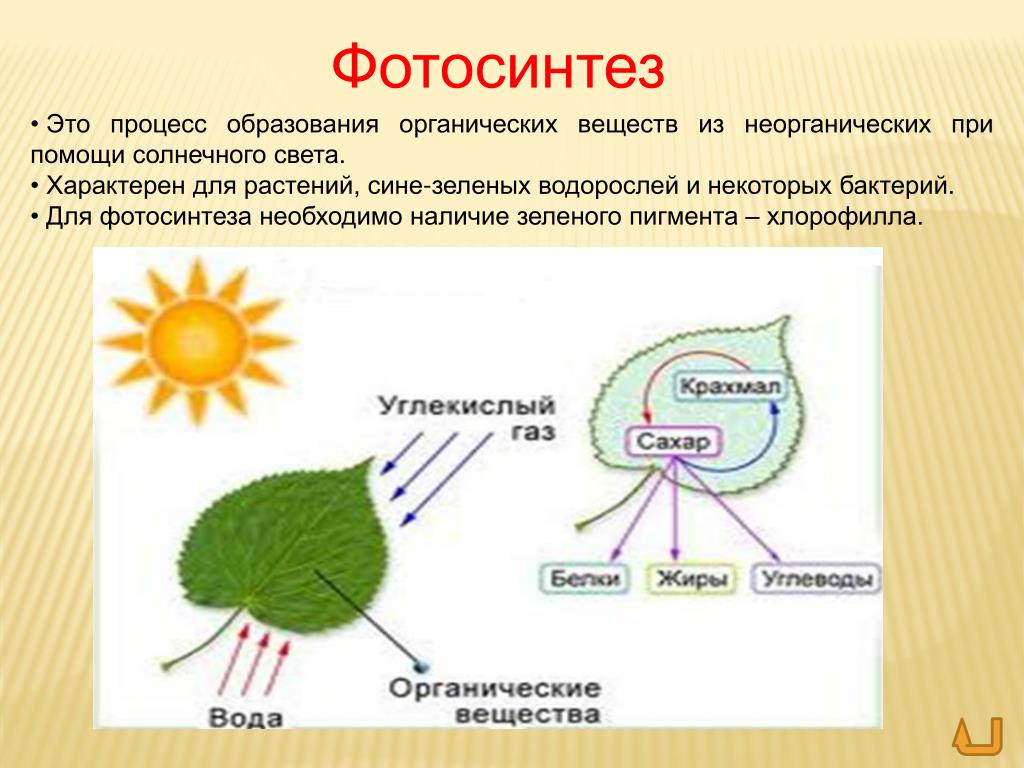 Plant Cell 16:1812–1826
Plant Cell 16:1812–1826
Статья
пабмед
КАС
Google ученый
Gottesman MM, Fojo T, Bates SE (2002)Множественная лекарственная устойчивость при раке: роль АТФ-зависимых транспортеров. Nat Rev Рак 2:48–58
Статья
пабмед
КАС
Google ученый
Govindachari TR, Viswanathan N (1972) Алкалоиды Mappia foetida . Фитохимия 11:3529–3531
Статья
КАС
Google ученый
Gundlach H, Muller MJ, Kutchan TM, Zenk MH (1992) Жасмоновая кислота является преобразователем сигнала в культурах клеток растений, индуцированных элиситором. Proc Natl Acad Sci USA 89:2389–2393
Статья
пабмед
КАС
Google ученый
Guo Z, Wagner GJ (1995) Биосинтез лабдендиола и склареола в бесклеточных экстрактах трихом Nicatiana glutinosa . Планта 197:627–632
Планта 197:627–632
Статья
КАС
Google ученый
Harborne JB, Williams CA (2000) Успехи в исследованиях флавоноидов с 1992 г. Фитохимия 55:481–504
Статья
пабмед
КАС
Google ученый
Hartmann T, Kutchan TM, Strack D (2005) Эволюция метаболического разнообразия. Фитохимия 66:1198–1199
Статья
пабмед
КАС
Google ученый
Hopp W, Seitz HU (1987) Поглощение ацилированного антоцианина изолированными вакуолями из культуры клеточной суспензии Daucus carota . Планта 170:74–85
Статья
КАС
Google ученый
Hopwood DA (2007) Как бактерии, продуцирующие антибиотики, обеспечивают свою саморезистентность до того, как биосинтез антибиотиков выведет их из строя? Мол Микробиол 63:937–940
Артикул
пабмед
КАС
Google ученый
Hsiang YH, Hertzberg R, Hecht S, Liu LF (1985) Камптотецин индуцирует связанные с белками разрывы ДНК с помощью топоизомеразы ДНК млекопитающих I.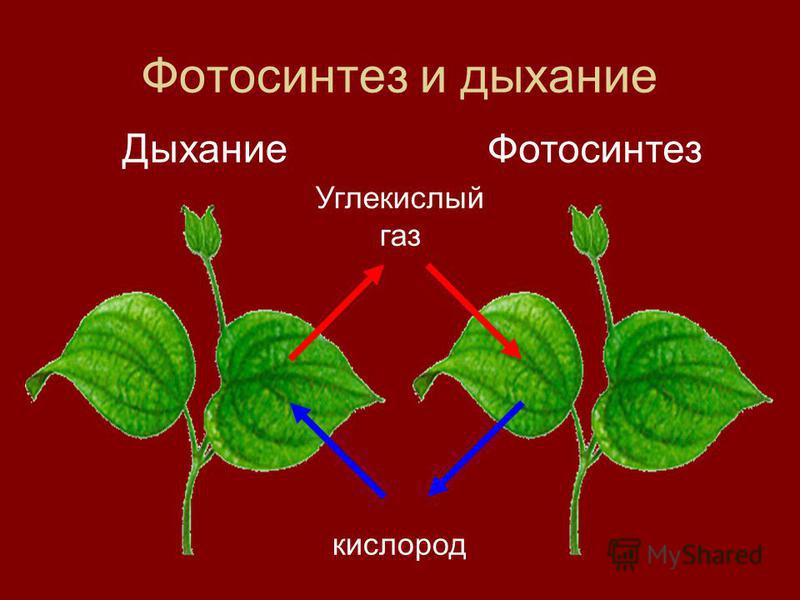 J Biol Chem 260:14873–14878
J Biol Chem 260:14873–14878
PubMed
КАС
Google ученый
Huges MA (1999) Биосинтез и деградация цианогенных гликозидов. В: Бартон Д., Наканиши К. (ред.) Комплексная химия натуральных продуктов, том 1. Elsevier, Амстердам, стр. 881–895
.
Google ученый
Hughes J, Mellows G, Soughton S (1980) Как Pseudomonas fluorescens , организмы, продуцирующие антибиотик псевдомоновую кислоту А, избегают самоубийства? FEBS Lett 122:322–324
Статья
пабмед
КАС
Google ученый
Husebye H, Chadchawan S, Winge P, Thangstad OP, Bones AM (2002)Специфическая экспрессия тиоглюкозидглюкогидролазы 1 (мирозиназы) в защитных клетках и идиобластах флоэмы у арабидопсиса. Завод Физиол 128:1180–1188
Артикул
пабмед
КАС
Google ученый
Inokoshi J, Tomoda H, Hashimoto H, Watanabe A, Takeshima H, Omura S (1994) Устойчивые к церуленину мутанты Saccharomyces cerevisiae с измененным геном синтазы жирных кислот. Mol Gen Genet 244:90–96
Mol Gen Genet 244:90–96
Статья
пабмед
КАС
Google ученый
Jasinski M, Stukkens Y, Degand H, Purnelle B, Marchand-Brynaert J, Boutry M (2001) Переносчик кассетного типа, связывающий АТФ на плазматической мембране растений, участвует в секреции противогрибковых терпеноидов. Растительная клетка 13:1095–1107
Статья
пабмед
КАС
Google ученый
Julsing MK, Koulman A, Woerdenbag HJ, Quax WJ, Kayser O (2006) Комбинаторный биосинтез вторичных метаболитов лекарственных растений. Biomol Eng 23:265–279
Статья
пабмед
КАС
Google ученый
Kakiuchi N, Hattori M, Ishii H, Namba T (1987) Влияние бензо[c]фенантридиновых алкалоидов на обратную транскриптазу и их свойство связывания с нуклеиновыми кислотами. Planta Med 53:22–27
PubMed
КАС
Google ученый
Kavallaris M, Kuo DY, Burkhart CA, Regl DL, Norris MD, Haber M, Horwitz SB (1997) Устойчивые к таксолу эпителиальные опухоли яичников связаны с измененной экспрессией специфических изотипов бета-тубулина. J Clin Invest 100:1282–1293
J Clin Invest 100:1282–1293
Статья
пабмед
КАС
Google ученый
Кавагути А., Томода Х., Окуда С., Авая Дж., Омура С. (1979) Резистентность к церуленину у грибка, продуцирующего церуленин. Арх Биохим Биофиз 197:30–35
Статья
пабмед
КАС
Google ученый
Ким Э.С., Малберг П.Г. (1997)Развитие пластид в дисковых клетках железистых трихом каннабиса (Cannabaceae). Клетки Mol 7:352–359
PubMed
КАС
Google ученый
Kitamura S, Shikazono N, Tanaka A (2004) TRANSPARENT TESTA 19 участвует в накоплении как антоцианов, так и проантоцианидинов в Арабидопсис . Завод J 37:104–114
Артикул
пабмед
КАС
Google ученый
Kjaer A (1976) Глюкозинолаты в крестоцветных. В: Vaughn JG, MacLeod AJ, Jones BMG (ред.) Биология и химия крестоцветных. Academic Press, Лондон, стр. 207–219
Academic Press, Лондон, стр. 207–219
Google ученый
Кляйн М., Вайссенбёк Г., Дюфо А., Гайяр К., Кройц К., Мартинойя Э. (1996) Различные механизмы возбуждения управляют вакуолярным поглощением флавоноидного глюкозида и гербицидного глюкозида. J Biol Chem 271:29666–29671
Статья
пабмед
КАС
Google ученый
Klein M, Martinoia E, Hoffmann-Thoma G, Weissenböck G (2000) Зависимый от мембранного потенциала ABC-подобный транспортер опосредует вакуолярное поглощение глюкуронидов флавонов ржи: регуляция поглощения глюкуронидов глутатионом и его конъюгатами. Растение Дж 21:289–304
Артикул
пабмед
КАС
Google ученый
Klein M, Burla B, Martinoia E (2006) Подсемейство белка, ассоциированного с множественной лекарственной устойчивостью (MRP/ABCC), АТФ-связывающих переносчиков кассет в растениях. FEBS Letter 580:1112–1122
FEBS Letter 580:1112–1122
Статья
пабмед
КАС
Google ученый
Kliebenstein DJ, Kroymann J, Mitchell-Olds T (2005) Система глюкозинолат-мирозиназа в экологическом и эволюционном контексте. Curr Opin Plant Biol 8: 264–271
Артикул
пабмед
КАС
Google ученый
Kojima M, Poulton JE, Thayer SS, Conn EE (1979) Распределение дхуррина и ферментов, участвующих в его метаболизме, в тканях листьев Sorghum bicolor . Физиол растений 63:1022–1028
PubMed
КАС
Google ученый
Королева О.А., Дэвис А., Дикен Р., Торп М.Р., Томос А.Д., Хедрич Р. (2000) Идентификация нового типа клеток, богатых глюкозинолатом, в цветочном стебле арабидопсиса. Растение Физиол 124:599–608
Артикул
пабмед
КАС
Google ученый
Li S, Yi Y, Wang Y, Zhang Z, Beasley RS (2002) Накопление камптотецина и вариации в камптотеке.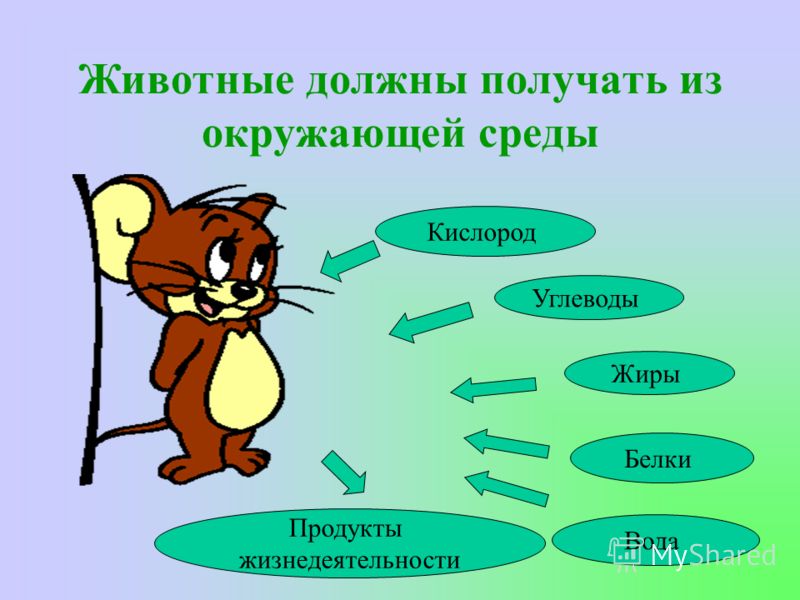 Планта Мед 68:1010–1016
Планта Мед 68:1010–1016
Статья
пабмед
КАС
Google ученый
Marinova K, Pourcel L, Weder B, Schwarz M, Barron D, Routaboul JM, Debaujon I, Klein M (2007a) Транспортер MATE TT12 арабидопсиса действует как вакуолярный флавоноид/H+-антипортер, активный в клетках, накапливающих проантоцианидин семенной кожуры. Растительная клетка 19:2023–2038
Артикул
пабмед
КАС
Google ученый
Марионова К.И., Кляйншмидт К., Вайссенбёк Г., Кляйн М. (2007b) Биосинтез флавоноидов в первичных листьях ячменя требует присутствия вакуоли и контролирует активность вакуолярного транспорта флавоноидов. Завод Физиол 144:432–444
Артикул
КАС
Google ученый
Маррс К.А., Алфенито М.Р., Ллойд А.М., Уолбот В. (1995) Глутатион-S-трансфераза, участвующая в вакуолярном переносе, кодируемом геном кукурузы Bronze-2 . Природа 375:397–400
Природа 375:397–400
Статья
пабмед
КАС
Google ученый
Martinoia E, Massonneau A, Frangne N (2000) Процессы переноса растворенных веществ через вакуолярную мембрану высших растений. Plant Cell Physiol 41:1175–1186
Статья
пабмед
КАС
Google ученый
Mende P, Wink M (1987) Поглощение хинолизидинового алкалоида лупанина протопластами и изолированными вакуолями культивируемых в суспензии клеток Lupinus polyphyllus . Диффузионный или переносчик-опосредованный транспорт? J Plant Physiol 129:229–242
CAS
Google ученый
Мендизабал В.Е., Адлер-Гращинский Э. (2007)Каннабиноиды как терапевтические средства при сердечно-сосудистых заболеваниях: рассказ о страстях и иллюзиях. Бр Дж. Фармакол 151:427–440
Артикул
пабмед
КАС
Google ученый
Мейерманс Х. , Моррил К., Лапьер С., Полле Б., Де Брюйн А., Буссон Р., Хердевейн П., Девриз Б., Ван Беумен Дж., Марита Дж.М., Ральф Дж., Чен С., Бурггрейв Б., Ван Монтегю М., Мессенс E, Boerjan W (2000) Модификации лигнина и накопление фенольных глюкозидов в ксилеме тополя при подавлении кофеилкоэнзима A O -метилтрансферазы, фермента, участвующего в биосинтезе лигнина. Дж Биол Хим 275:36899–36909
, Моррил К., Лапьер С., Полле Б., Де Брюйн А., Буссон Р., Хердевейн П., Девриз Б., Ван Беумен Дж., Марита Дж.М., Ральф Дж., Чен С., Бурггрейв Б., Ван Монтегю М., Мессенс E, Boerjan W (2000) Модификации лигнина и накопление фенольных глюкозидов в ксилеме тополя при подавлении кофеилкоэнзима A O -метилтрансферазы, фермента, участвующего в биосинтезе лигнина. Дж Биол Хим 275:36899–36909
Артикул
пабмед
КАС
Google ученый
Morimoto S, Tanaka Y, Sasaki K, Tanaka H, Fukamizu T, Shoyama Y, Shoyama Y, Taura F (2007) Идентификация и характеристика каннабиноидов, которые вызывают гибель клеток посредством перехода митохондриальной проницаемости в клетках листьев Cannabis . J Biol Chem 282:20739–20751
Статья
пабмед
КАС
Google ученый
Mueller LA, Goodman CD, Silady RA, Walbot V (2000) AN9, глутатион петунии S -трансфераза, необходимая для секвестрации антоцианов, представляет собой белок, связывающий флавоноиды. Завод Физиол 123:1561–1570
Завод Физиол 123:1561–1570
Артикул
пабмед
КАС
Google ученый
Osbourn AE (1996) Готовые противомикробные соединения и защита растений от грибковых атак. Растительная клетка 8: 1821–1831
Артикул
пабмед
КАС
Google ученый
Otani M, Shitan N, Sakai K, Martinoia E, Sato F, Yazaki K (2005) Характеристика вакуолярного транспорта эндогенного алкалоида берберина в Coptis japonica . Завод Физиол 138:1939–1946
Статья
пабмед
КАС
Google ученый
Pasqua G, Monacelli B, Valletta A (2004) Клеточная локализация противоракового препарата камптотецина в Camptotheca acuminata Decne (Nyssaceae). Eur J Histochem 48: 321–327
ПабМед
Google ученый
Pommier Y, Pourquier P, Fan Y, Strumberg D (1998) Механизм действия топоизомеразы I эукариотической ДНК и препаратов, нацеленных на фермент.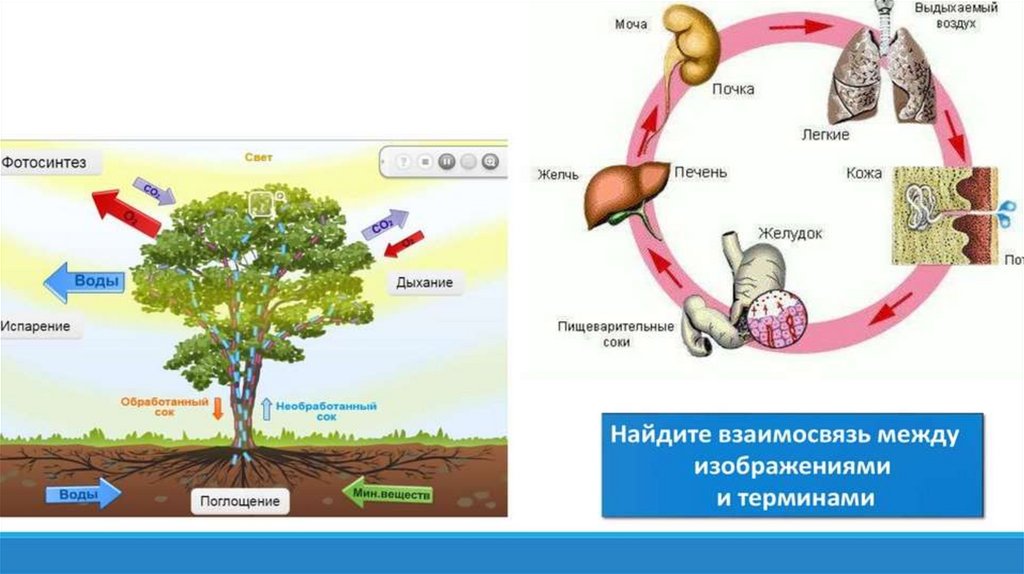 Biochim Biophys Acta 1400:83–105
Biochim Biophys Acta 1400:83–105
PubMed
КАС
Google ученый
Ranganathan S, Benetatos CA, Colarusso PJ, Dexter DW, Hudes GR (1998) Измененная экспрессия изотипа бета-тубулина в клетках карциномы предстательной железы человека, устойчивых к паклитакселу. Бр Дж Рак 77: 562–566
ПабМед
КАС
Google ученый
Раск Л., Андреассон Э., Экбом Б., Эрикссон С., Понтоппидан Б., Мейер Дж. (2000) Мирозиназа: эволюция семейства генов и защита травоядных у Brassicaceae. Растение Мол Биол 42:93–113
Артикул
пабмед
КАС
Google ученый
Rea PA (2007) Растительные АТФ-связывающие кассетные транспортеры. Annu Rev Plant Biol 58:347–375
Артикул
пабмед
КАС
Google ученый
Riis L, Bellotti AC, Bonierbale M, O’Brien GM (2003) Цианогенный потенциал маниоки и его влияние на травоядных насекомых общего профиля Cyrtomenus bergi (Hemiptera: Cydnidae).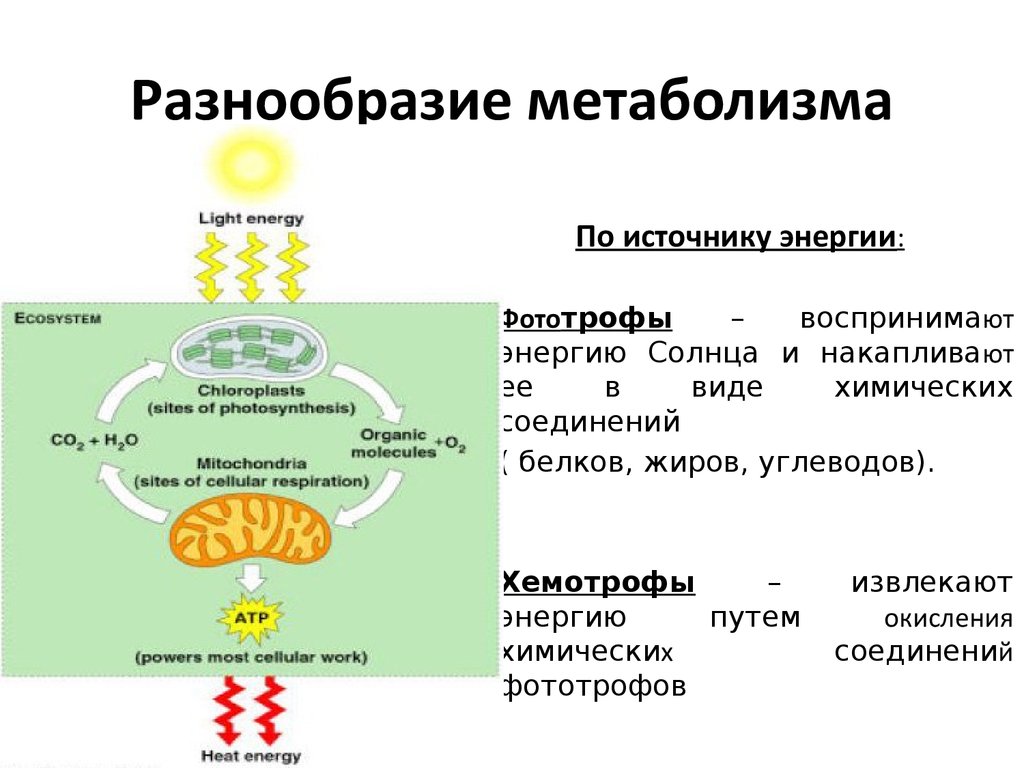 J Econ Entomol 96:1905–1914
J Econ Entomol 96:1905–1914
Статья
пабмед
Google ученый
Роос В., Эверс С., Хике М., Чопе М., Шуманн Б. (1998) Сдвиг внутриклеточного распределения рН как часть сигнального механизма, приводящего к выведению бензофенантридиновых алкалоидов. Биосинтез фитоалексина в культивируемых клетках Eschscholtzia californica . Завод Физиол 118:349–364
Артикул
пабмед
КАС
Google ученый
Roytrakul S, Verpoorte R (2007) Роль вакуолярных транспортных белков во вторичном метаболизме растений: Catharanthus roseus 9Культура клеток 0014. Phytochem Rev 6:383–396
Статья
КАС
Google ученый
Rueff J, Gaspar J, Laires A (1995) Структурные требования к мутагенности флавоноидов при нитрозировании. Структурно-деятельностное исследование. Мутагенез 10:325–328
Статья
пабмед
КАС
Google ученый
Сайто К.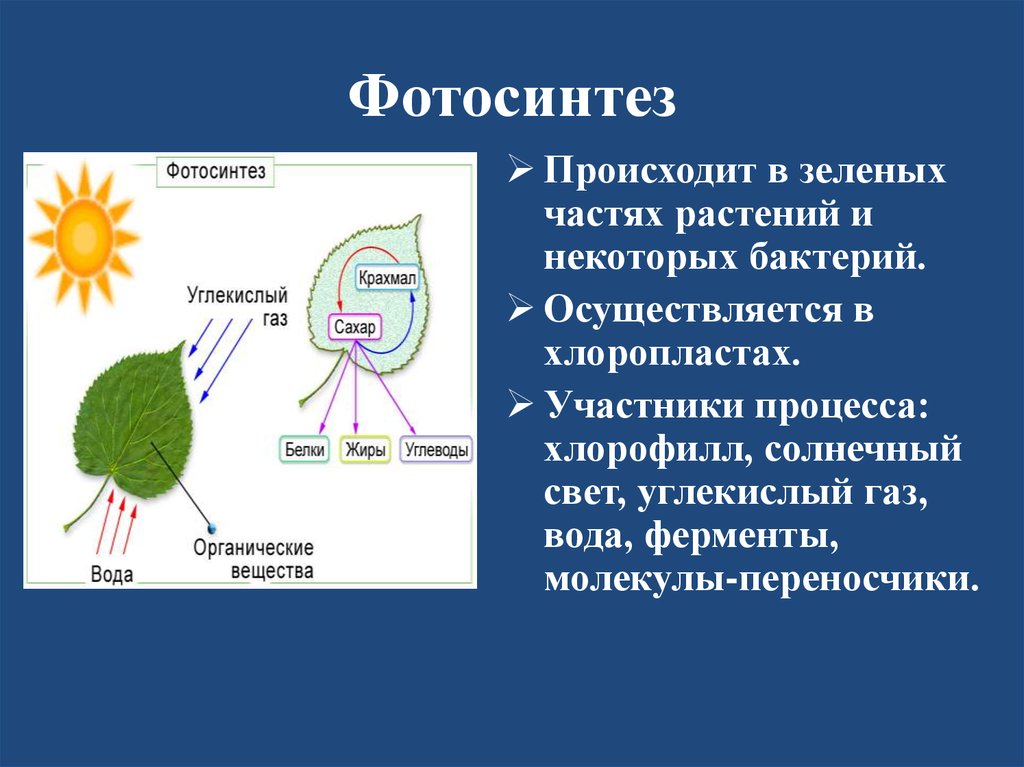 , Судо Х., Ямадзаки М., Косеки-Накамура М., Китадзима М., Такаяма Х., Аими Н. (2001) Возможное производство камптотецина культурой волосатых корней Ophiorrhiza pumila . Plant Cell Rep 20:267–271
, Судо Х., Ямадзаки М., Косеки-Накамура М., Китадзима М., Такаяма Х., Аими Н. (2001) Возможное производство камптотецина культурой волосатых корней Ophiorrhiza pumila . Plant Cell Rep 20:267–271
Статья
КАС
Google ученый
Sakai K, Shitan N, Sato F, Ueda K, Yazaki K (2002) Характеристика транспорта берберина в клетки Coptis japonica и участие белка ABC. J Exp Bot 53:1879–1886
Статья
пабмед
КАС
Google ученый
Сато Х., Тагучи Г., Фукуи Х., Табата М. (1992) Роль яблочной кислоты в растворении избытка берберина, накапливающегося в вакуолях Coptis japonica . Фитохимия 31:3451–3454
Статья
КАС
Google ученый
Scheiner-Bobis G (2001) Сангвинарин индуцирует отток K + из дрожжевых клеток, экспрессирующих натриевые насосы млекопитающих. NS Arch Pharmacol 7:1288–1289
CAS
Google ученый
Schmeller T, Latz-Bruning B, Wink M (1997) Биохимическая активность берберина, пальматина и сангвинарина, опосредующая химическую защиту от микроорганизмов и травоядных.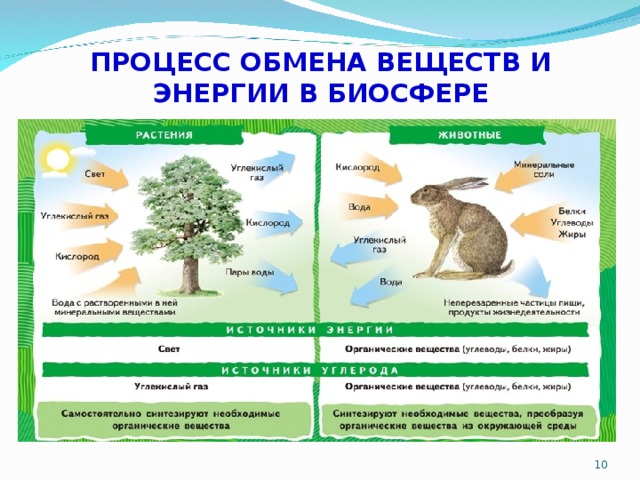 Фитохимия 44:257–266
Фитохимия 44:257–266
Статья
пабмед
КАС
Google ученый
Schulz M, Wieland I (1999) Различия в метаболизме BOA среди видов в различных полевых сообществах – биохимические доказательства коэволюционных процессов в растительных сообществах? Химиоэкология 9:133–141
Артикул
КАС
Google ученый
Schumacher HM, Gundlach H, Fiedler F, Zenk MH (1987) Выявление синтеза бензофенантридиновых алкалоидов в культурах клеток Eschscholzia. Plant Cell Rep 6:410–413
CAS
Google ученый
Sicker D, Frey M, Schulz M, Gierl A (2000) Роль природных бензоксазинонов в стратегии выживания растений. Инт Рев Цитол 198:319–346
Статья
пабмед
КАС
Google ученый
Sicker D, Schneider B, Hennig L, Knop M, Schulz M (2001)Детоксикация карбаматов гликозидов из бензоксазолин-2(3H)-она в экстрактах и экссудатах корней кукурузы. Фитохимия 58:819–825
Фитохимия 58:819–825
Статья
пабмед
КАС
Google ученый
Sirikantaramas S, Taura F, Tanaka Y, Ishikawa Y, Morimoto S, Shoyama Y (2005) Синтаза тетрагидроканнабиноловой кислоты, фермент, контролирующий психоактивность марихуаны, секретируется в полость хранения железистых трихом. Физиология клеток растений 46:1578–158
Артикул
пабмед
КАС
Google ученый
Sirikantaramas S, Taura F, Morimoto S, Shoyama Y (2007a) Последние достижения в исследованиях Cannabis sativa : исследования биосинтеза и его потенциал в биотехнологии. Curr Pharm Biotech 8:237–243
Статья
КАС
Google ученый
Сирикантарамас С., Асано Т., Судо Х., Ямадзаки М., Сайто К. (2007b) Камптотецин: терапевтический потенциал и биотехнология. Карр Фарм Биотех 8:196–202
Артикул
КАС
Google ученый
Sirikantaramas S, Sudo H, Asano T, Yamazaki M, Saito K (2007c) Транспорт камптотецина в волосатых корнях Ophiorrhiza pumila .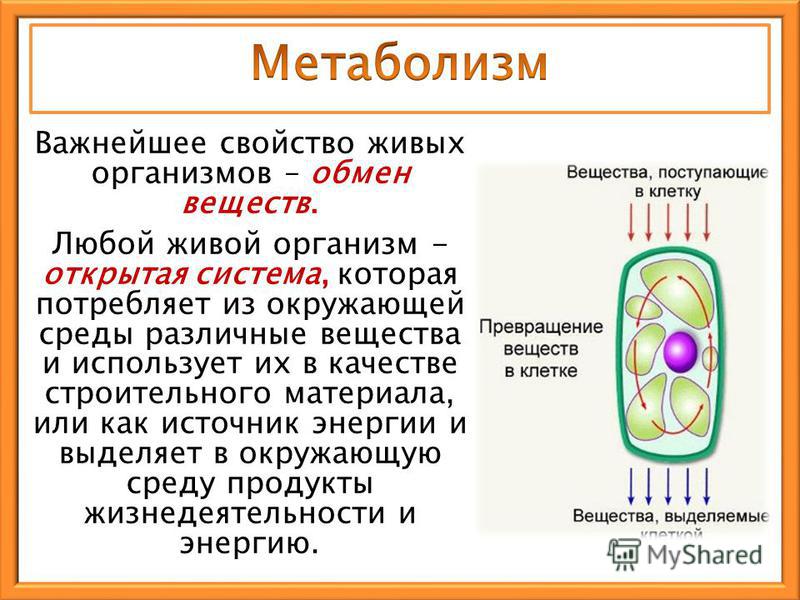 Фитохимия (в печати)
Фитохимия (в печати)
Stukkens Y, Bultreys A, Grec S, Trombik T, Vanham D, Boutry M (2005). большую роль в защите растений от патогенов. Завод Физиол 139:341–352
Артикул
пабмед
КАС
Google ученый
Такатани Х., Ока М., Фукуда М., Нарасаки Ф., Накано Р., Икеда К., Тераши К., Киношита А., Сода Х., Канда Т., Шнайдер Э., Коно С. (1997) Анализ мутаций генов и количественное определение ДНК-топоизомеразы I при ранее не леченных немелкоклеточных карциномах легкого. Jpn J Cancer Res 88: 160–165
PubMed
КАС
Google ученый
Таттерсалл Д.Б., Бак С., Джонс П.Р., Олсен К.Е., Нильсен Дж.К., Хансен М.Л., Хой П.Б., Мёллер Б.Л. (2001) Устойчивость к травоядным за счет искусственного синтеза цианогенного глюкозида. Science 293:1826–1828
Статья
пабмед
КАС
Google ученый
Таура Ф. , Сирикантарамас С., Шояма Ю., Йошикай К., Шояма Ю., Моримото С. (2007) Синтаза каннабидиоловой кислоты, определяющий хемотип фермент в волокнах типа Каннабис посевной . FEBS Lett 581:2929–2934
, Сирикантарамас С., Шояма Ю., Йошикай К., Шояма Ю., Моримото С. (2007) Синтаза каннабидиоловой кислоты, определяющий хемотип фермент в волокнах типа Каннабис посевной . FEBS Lett 581:2929–2934
Статья
пабмед
КАС
Google ученый
Thayer SS, Conn EE (1981)Субклеточная локализация дхуррин-бета-глюкозидазы и гидроксинитриллиазы в клетках мезофилла листовых пластинок сорго. Физиол растений 67:617–622
PubMed
КАС
Google ученый
Thiara AS, Cundliffe E (1988) Клонирование и характеристика гена ДНК-гиразы B из Streptomyces sphaeroides , придающий устойчивость к новобиоцину. EMBO J 7:2255–2259
PubMed
КАС
Google ученый
Томода Х., Кавагути А., Ясухара Т., Накадзима Т., Омура С., Окуда С. (1984) Резистентность к церуленину у грибка, продуцирующего церуленин. III.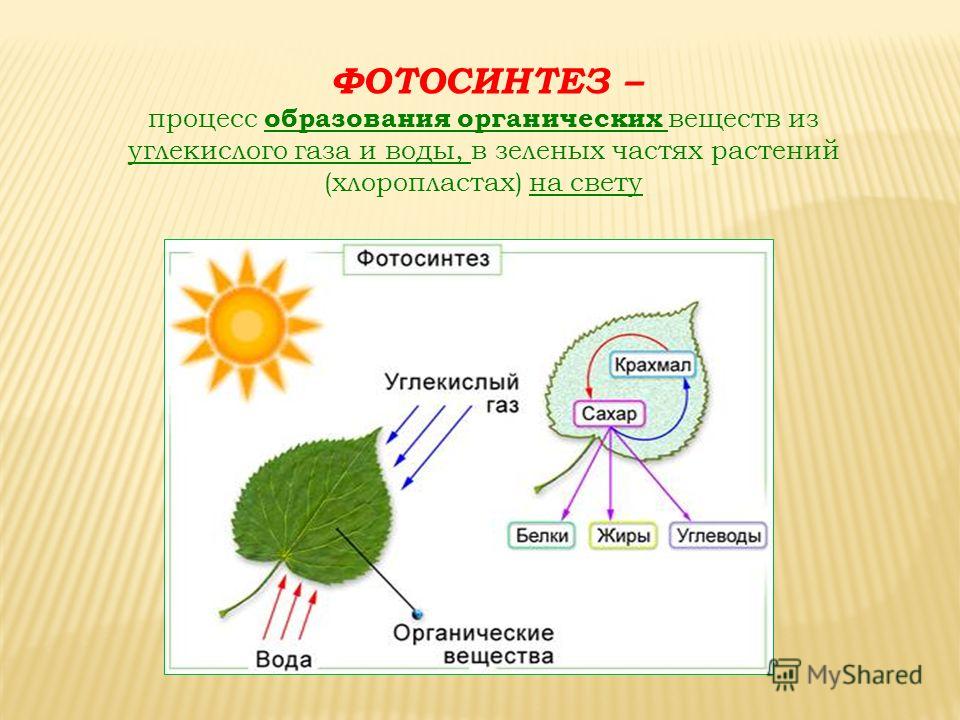 Исследования пептидов активного центра синтетазы жирных кислот из Cephalosporium caerulens . J Biochem 95:1713–1723
Исследования пептидов активного центра синтетазы жирных кислот из Cephalosporium caerulens . J Biochem 95:1713–1723
CAS
Google ученый
Цурутани Дж., Нитта Т., Хирашима Т., Комия Т., Уэдзима Х., Тада Х., Сюничи Н., Тохда А., Фукуока М., Накагава К. (2002) Точечные мутации в гене топоизомеразы I у пациентов с немелкоклеточной рак легких лечили иринотеканом. Рак легких 35:299–304
Статья
пабмед
Google ученый
Ueda K, Cardarelli C, Gottesman MM, Pastan I (1987) Экспрессия полноразмерной кДНК человеческого гена «MDR1» придает устойчивость к колхицину, доксорубицину и винбластину. Proc Natl Acad Sci USA 84:3004–3008
Артикул
пабмед
КАС
Google ученый
van den Brüle S, Müller A, Fleming AJ, Smart CC (2002) Транспортер ABC SpTUR2 придает устойчивость к противогрибковому склареолу. Завод J 30:649–662
Завод J 30:649–662
Артикул
пабмед
Google ученый
Vickery ML (1981) Сахарный обмен. В: Викери М.Л., Викери Б. (ред.) Вторичный метаболизм растений. University Park Press, Балтимор, стр. 32–41
Виллегас М., Соммарин М., Броделиус П.Е. (2000) Влияние ортованадата натрия на образование и распределение бензофенантридиновых алкалоидов в культурах суспензии клеток Eschscholzia californica . Plant Physiol Biochem 38:233–241
Статья
КАС
Google ученый
von Rad U, Huttl R, Lottspeich F, Gierl A, Frey M (2001) Две глюкозилтрансферазы участвуют в детоксикации бензоксазиноидов в кукурузе. Завод J 28: 633–642
Артикул
Google ученый
Wall ME, Wani MC, Cook C, Palmer K, McPhail A, Sim G (1966) Растительные противоопухолевые агенты. I. Выделение и структура камптотецина, нового алкалоидного лейкоза и ингибитора опухолей из Camptotheca acuminata .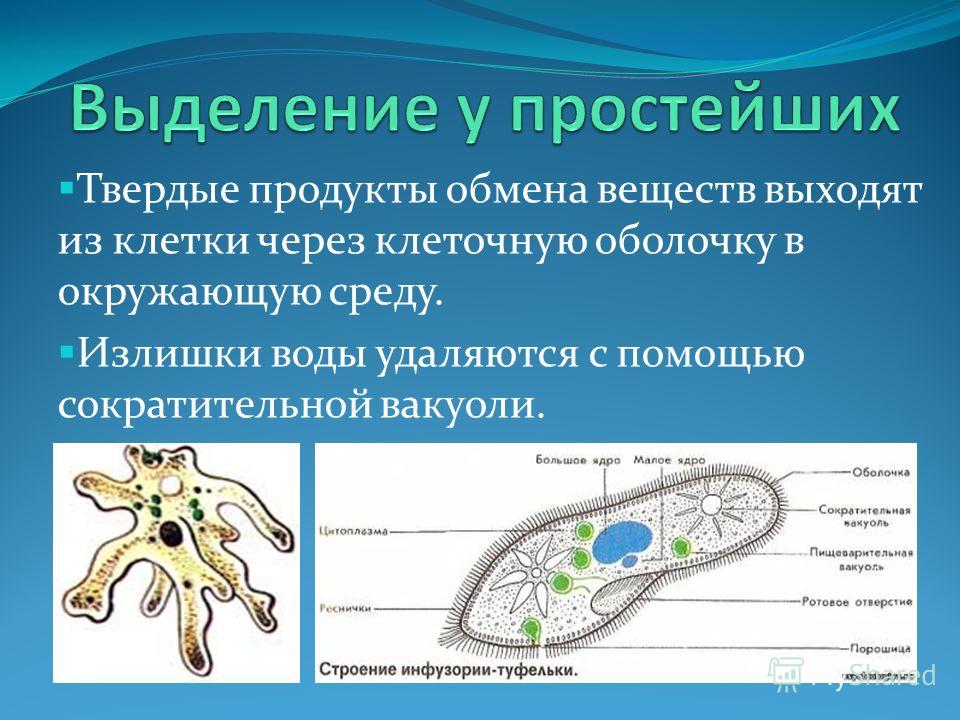 J Am Chem Soc 88:3888–3890
J Am Chem Soc 88:3888–3890
Статья
КАС
Google ученый
Weiss D, Baumert A, Vogel M, Roos W (2006)Сангвинаринредуктаза, ключевой фермент детоксикации бензофенантридина. Окружающая среда растительных клеток 29:291–302
Артикул
пабмед
КАС
Google ученый
Виндзор А.Дж., Райхельт М., Фигут А., Сватос А., Кройманн Дж., Клибенштейн Д.Дж., Гершензон Дж., Митчелл-Олдс Т. (2005)Географическая и эволюционная диверсификация глюкозинолатов среди ближайших родственников Arabidopsis thaliana (Brassicaceae). Фитохимия 66:1321–1333
Статья
пабмед
КАС
Google ученый
Wink M, Roberts MF (1998) Компартментация биосинтеза алкалоидов, транспорт и хранение. В: Робертс М.Ф., Винк М. (ред.) Алкалоиды, биохимия, экология и применение в медицине. Plenum Press, Нью-Йорк, стр. 239–262
Google ученый
Yamamoto K, Takase H, Abe K, Saito Y, Suzuki A (1993) Фармакологические исследования антидиарейных эффектов препарата, содержащего берберин и траву герани.