
Содержание
Опыление насекомыми и ветром
Признаки насекомоопыляемых растений
1. Имеются нектарники, железы в основании пестика, которые вырабатывают сладкий нектар. На эту вкусную жижицу, да еще «приправленную» пыльцой, охотно слетаются насекомый. Как следует покопошившись в цветке, опылив его, они, все в пыльце, летят на другие цветки и тоже их опыляют.
2. Цветки обоеполые, пыльца липкая, клейкая, на оболочке пыльцы имеются выросты. Обоеполость цветков нужна, чтобы опыление насекомым происходило сразу, на этом же цветке. А клейкость пыльцы и выросты — для прикрепления к покрову насекомого.
3. Цветки зачастую крупные, их околоцветник ярко, привлекательно окрашен. Это приспособление выработано в рекламных целях — яркие цветки больше привлекают насекомых.
4. Если цветки мелкие, то они собраны в крупные соцветия — для тех же целей.
5. Цветки имеют выраженные запахи. Неизвестно, какие запахи больше нравятся насекомым, но человек различает среди них приятные сладкие запахи (липа, сирень, гиацинт, роза) и отталкивающие, — например, раффлезия Арнольда распространяет запах гнилого мяса, а аморфофаллус титанический (наибольший цветок в мире) «благоухает» протухшими яйцами и рыбой. Однако же и на таких красавцев находятся свои опылители! Но нам все-таки приятнее какой-нибудь душистый табак, чей чудесный аромат разливается в летнюю ночь.
Неизвестно, какие запахи больше нравятся насекомым, но человек различает среди них приятные сладкие запахи (липа, сирень, гиацинт, роза) и отталкивающие, — например, раффлезия Арнольда распространяет запах гнилого мяса, а аморфофаллус титанический (наибольший цветок в мире) «благоухает» протухшими яйцами и рыбой. Однако же и на таких красавцев находятся свои опылители! Но нам все-таки приятнее какой-нибудь душистый табак, чей чудесный аромат разливается в летнюю ночь.
Приспособленность цветков к опылению определенными насекомыми
Многие растения «накрывают стол» для любых насекомых, у них пыльца и нектар доступны всем, кто прилетит, как, например, у мака. Но некоторые виды опыляются только определенными насекомыми. Так, сердцевина цветка львиного зева труднодоступна — в него еще пролезть нужно! И сделать это могут только крупные увесистые насекомые: шмели и пчелы. Они своим тельцем приоткрывают цветок и проникают внутрь. А вот упомянутый уже душистый табак опыляют крупные ночные бабочки бражники, — их очень длинный и тонкий, как игла, хоботок дотягивается до нектара и пыльцы. В тропиках опылением занимаются даже летучие мыши и птицы — например, крошечные колибри.
В тропиках опылением занимаются даже летучие мыши и птицы — например, крошечные колибри.
Признаки ветроопыляемых растений
1. Деревья и кустарники цветут еще до появления первых листьев. Пока нет листвы, леса и парки голые, пыльце легче циркулировать, нестись за ветром. Так ведут себя, например, шелковица, ольха и орешник.
2. Не имеют околоцветника, а если он есть, то развит слабо (злаки).
3. Цветки мелкие, зачастую собраны в соцветия (ива, пшеница).
4. При таком виде опыления пыльца нередко расходуется впустую, улетает без толку, поэтому растение вырабатывает ее с большим избытком. Крупные и клейкие пылинки также не имеют смысла — прилипнут неподалеку, и поэтому здесь пыльца очень мелкая, сухая, летучая.
5. Одиноко стоять на равнине и ждать опыления очень грустно, поэтому ветроопыляемые растения предпочитают образовывать скопления (березовые рощи, тростниковые заросли).
Искусственное опыление
1. Этот вид опыления, перенос пыльцы с женских цветков на мужские, вручную делает человек в определенных целях — выведение новых сортов, подъем урожайности.
2. Как производится опыление? С мужских цветков удаляют пыльники, подсушивают пыльцу и наносят ее на женские цветки в определенное время — причем обрабатывать вообще все цветки в теплице, например, не требуется, достаточно охватить десяток процентов. Еще один способ — аккуратно срезать мужские цветки и потрясти их пыльниками над женскими, стараясь не повреждать последние. Например, однодомную кукурузу, имеющую мужские соцветия-метелки и женские початки, можно опылить так, не ожидая милостей от природы: собрать в бумажную воронку пыльцу и рассеять ее над рыльцами пестиков.
Хочешь сдать экзамен на отлично? Жми сюда — подготовка к ОГЭ по биологии (9 класс)
Специализированные опылители способствуют сохранению редких растений • Елена Устинова • Новости науки на «Элементах» • Ботаника, Экология, Энтомология
В многовидовых сообществах одновременно сосуществует большое количество видов, у многих из которых низкая численность. Несмотря на это они устойчиво воспроизводятся в сообществе, выдерживая конкурентную борьбу с видами с высокой численностью. Ученые пока не до конца разобрались в механизмах, благоприятствующих таким малочисленным видам. Между тем, их понимание является ключом к поддержанию разнообразия растительных сообществ. Комплексное исследование сети взаимодействий растений и опылителей в масштабах многовидового сообщества в американском штате Калифорния позволило понять, как особенности опыления могут способствовать сохранению редких видов. Похоже, что ключевую роль здесь играют специализированные насекомые, которые поддерживают узкие экологические ниши для «своих» редких видов растений и не дают им проиграть конкуренцию более многочисленным видам.
Несмотря на это они устойчиво воспроизводятся в сообществе, выдерживая конкурентную борьбу с видами с высокой численностью. Ученые пока не до конца разобрались в механизмах, благоприятствующих таким малочисленным видам. Между тем, их понимание является ключом к поддержанию разнообразия растительных сообществ. Комплексное исследование сети взаимодействий растений и опылителей в масштабах многовидового сообщества в американском штате Калифорния позволило понять, как особенности опыления могут способствовать сохранению редких видов. Похоже, что ключевую роль здесь играют специализированные насекомые, которые поддерживают узкие экологические ниши для «своих» редких видов растений и не дают им проиграть конкуренцию более многочисленным видам.
Природные сообщества (биоценозы) как правило состоят из многих видов, численность которых сильно разнится. Обычно в сообществе есть один или несколько доминантных видов, преобладающих по численности и занимающих большую часть биотопа. Кроме видов-доминантов любой биоценоз включает множество более редких видов, которые, однако, очень важны для него. Они увеличивают видовое богатство, которое позволяет каждому виду поддерживать высокое разнообразие связей с другими компонентами биоценоза — животными, грибами, бактериями. В результате повышается эффективность использования ресурсов, растет продуктивность сообщества, а также снижается вероятность вымирания отдельных видов (см. новость В разнообразном сообществе у животных меньше шансов вымереть («Элементы», 13.05.2009) и статью Э. Лекявичус, 2018. Биологическое разнообразие — зачем и почему? I. Поиски функционального объяснения). Кроме того, малочисленные виды могут служить резервом для пополнения и замещения доминантов, что придает биоценозу устойчивость и обеспечивает надежность его функционирования в разных условиях.
Они увеличивают видовое богатство, которое позволяет каждому виду поддерживать высокое разнообразие связей с другими компонентами биоценоза — животными, грибами, бактериями. В результате повышается эффективность использования ресурсов, растет продуктивность сообщества, а также снижается вероятность вымирания отдельных видов (см. новость В разнообразном сообществе у животных меньше шансов вымереть («Элементы», 13.05.2009) и статью Э. Лекявичус, 2018. Биологическое разнообразие — зачем и почему? I. Поиски функционального объяснения). Кроме того, малочисленные виды могут служить резервом для пополнения и замещения доминантов, что придает биоценозу устойчивость и обеспечивает надежность его функционирования в разных условиях.
Итак, малочисленные виды очень важны для поддержания видового разнообразия, однако остается непонятным, как множество малочисленных видов сосуществует и выдерживает конкурентную борьбу с многочисленными видами. При перекрестном опылении растениям с малой численностью затруднительно обмениваться пыльцой, ведь рядом может не оказаться других растений этого вида. Случайная гибель одной особи (например, от растительноядных животных или случайно упавшего дерева) для редких видов может быть критична и привести к тому, что воспроизведение популяции уже окажется невозможным. Популяция малочисленных видов не может дать столько же семян, сколько производит популяция многочисленных видов, а значит у редких видов намного меньше шансов проявиться в сообществе в следующем поколении.
Случайная гибель одной особи (например, от растительноядных животных или случайно упавшего дерева) для редких видов может быть критична и привести к тому, что воспроизведение популяции уже окажется невозможным. Популяция малочисленных видов не может дать столько же семян, сколько производит популяция многочисленных видов, а значит у редких видов намного меньше шансов проявиться в сообществе в следующем поколении.
Взаимодействие растений и опылителей считается одним из наиболее важных факторов биоразнообразия на Земле. Сопряженная эволюция цветковых растений и насекомых обуславливает огромную диверсификацию в обеих группах: появление новых видов растений создает потенциальные ниши для опылителей и наоборот (см. новости Коэволюция насекомых и растений началась задолго до появления цветков («Элементы», 11.11.2009) и Экспериментально доказано, что разнообразие растительных сообществ поддерживается расхождением видов по разным нишам («Элементы», 23.09.2009)). Подавляющее большинство цветковых растений (примерно 80%) опыляются животными. Без опылителей у более половины из них заметно снизится число семян — несмотря на то, что большинство из них способны к самооплодотворению.
Без опылителей у более половины из них заметно снизится число семян — несмотря на то, что большинство из них способны к самооплодотворению.
Авторы недавней статьи в журнале Nature исследовали богатую видами систему, расположенную на серпентиновых почвах (см. Serpentine soil) в природном заповеднике Маклафлин в Калифорнии, США (McLaughlin Natural Reserve, рис. 1). Серпентиновые почвы очень неблагоприятны для жизни растений, так как обеднены питательными веществами (азотом, фосфором и калием), а также содержат высокие концентрации тяжелых металлов, включая хром, железо, кобальт и никель. Многие произрастающие здесь растения являются эндемиками (примерно 20%), которые являются редкими в регионе видами. Из-за близкого расположения грунтовых вод на поверхность выходят небольшие эфемерные ручейки с высоким содержанием бикарбонатов, что приводит к заболачиванию территории в конце лета. Это ограничивает время цветения и плодоношения произрастающих тут растений, обеспечивая значительное перекрытие сроков цветения, что обостряет конкуренцию за опылителей. В этой системе опыление является важным фактором, влияющим на успешное размножение и, вероятно, сосуществование.
В этой системе опыление является важным фактором, влияющим на успешное размножение и, вероятно, сосуществование.
Ученые предположили, что взаимодействие между растениями и опылителями в масштабах сообщества может способствовать сохранению редких видов растений, которые подвергаются большему риску исчезновения, чем многочисленные виды. Для объяснения того, как это происходит, они выдвинули четыре гипотезы.
Успешное опыление происходит при попадании на рыльце пестика пыльцы растения того же самого вида (см. задачу Кто какой цветок опыляет?). Напротив, когда на рыльце пестика попадает пыльца другого вида растений, происходит снижение репродуктивного успеха, так как, во-первых, произойдет бессмысленная трата пыльцы, а, во-вторых, «чужая» пыльца может закрыть поверхность рыльца, мешая возможному последующему опылению пыльцой своего вида.
Конкуренция растений за услуги опылителей приводит к разграничению состава опылителей: при одновременном цветении в сообществе разных видов растений отбор благоприятствует тому, чтобы спектр их опылителей перекрывался как можно меньше (см. новость Конкуренция за опылителей делает совместно цветущие виды пасленовых непохожими по окраске, «Элементы», 16.05.2014). Это не только снижает конкуренцию за опылителей, но и увеличивает шансы получения своей пыльцы. В результате более специализированные растения, приспособленные к опылению конкретной группой насекомых, получают преимущество по сравнению с растениями, у которых широкий круг опылителей. Первая гипотеза предполагает, что разграничение состава опылителей может благоприятствовать малочисленным видам растений из-за их большей специализации, чем у многочисленных видов (рис. 2, А). Для оценки этой гипотезы авторы исследования сначала проверили, различаются ли спектры опылителей у разных растений в изучаемом сообществе. Затем они выяснили, являются ли редкие виды растений более специализированными, чем многочисленные виды.
новость Конкуренция за опылителей делает совместно цветущие виды пасленовых непохожими по окраске, «Элементы», 16.05.2014). Это не только снижает конкуренцию за опылителей, но и увеличивает шансы получения своей пыльцы. В результате более специализированные растения, приспособленные к опылению конкретной группой насекомых, получают преимущество по сравнению с растениями, у которых широкий круг опылителей. Первая гипотеза предполагает, что разграничение состава опылителей может благоприятствовать малочисленным видам растений из-за их большей специализации, чем у многочисленных видов (рис. 2, А). Для оценки этой гипотезы авторы исследования сначала проверили, различаются ли спектры опылителей у разных растений в изучаемом сообществе. Затем они выяснили, являются ли редкие виды растений более специализированными, чем многочисленные виды.
Стоит отметить, что растения с широким кругом опылителей имеют свои преимущества при отсутствии большого количества совместно цветущих видов, поскольку самые разные группы насекомых (и не только) могут посещать их цветки, что потенциально увеличивает общую численность опылителей. Если других цветущих растений в это время нет, то все эти посетители (даже те, которые не обладают высокой специализацией) будут осуществлять перенос пыльцы на растения того же вида. К тому же большое разнообразие опылителей позволяет растениям не зависеть от колебаний численности и благополучия популяции конкретного вида опылителей.
Если других цветущих растений в это время нет, то все эти посетители (даже те, которые не обладают высокой специализацией) будут осуществлять перенос пыльцы на растения того же вида. К тому же большое разнообразие опылителей позволяет растениям не зависеть от колебаний численности и благополучия популяции конкретного вида опылителей.
Вторая гипотеза касается так называемой асимметрии взаимодействий (рис. 2, Б). Когда растения имеют общих опылителей, редкие виды извлекают выгоду из того, что опылителей привлекают растущие рядом растения многочисленных видов (см. новость Асимметрия взаимовыгодных отношений способствует высокому биоразнообразию, «Элементы», 26.04.2006). Хотя редкие виды также могут получать чужую пыльцу, привлекая общих с многочисленными видами опылителей, за счет большего числа опылителей они могут получать больше своей пыльцы, чем получали бы, произрастая изолированно. А многочисленные виды, наоборот, при этом теряют впустую больше своей пыльцы, чем в ситуации, когда редких видов нет. Таким образом, асимметричность опылительной сети может потенциально увеличивать приспособленность редких видов, но снижать приспособленность многочисленных видов из-за растрачивания пыльцы.
Таким образом, асимметричность опылительной сети может потенциально увеличивать приспособленность редких видов, но снижать приспособленность многочисленных видов из-за растрачивания пыльцы.
Чтобы проверить эту гипотезу, авторы оценили, получают ли редкие виды большее количество своей пыльцы (вместе с большим количеством чужой пыльцы) по сравнению с многочисленными видами при совместном использовании опылителей. Также авторы выясняли, выше ли у многочисленных видов трата своей пыльцы (по количеству пыльцевых зерен на рыльцах чужих видов растений), когда опылители, которые в основном посещают многочисленные виды, переходят к редким видам.
Третья гипотеза касается механизма, опылителями не опосредованного: редкие виды потенциально могут поддерживать свою численность за счет способности к самоопылению, когда передача пыльцы от пыльника к рыльцу происходит внутри цветка без помощи опылителя. Самоопыление более вероятно при небольшом физическом расстоянии между рыльцами и пыльниками. Оно, вероятно, позволяет редким видам выиграть в приспособленности, когда опылителей мало. Чтобы оценить данную гипотезу, авторы выясняли, приводит ли более короткое расстояние между пыльниками и рыльцем к повышению репродуктивного успеха, и имеют ли редкие виды более короткие расстояния между пыльниками и рыльцем.
Оно, вероятно, позволяет редким видам выиграть в приспособленности, когда опылителей мало. Чтобы оценить данную гипотезу, авторы выясняли, приводит ли более короткое расстояние между пыльниками и рыльцем к повышению репродуктивного успеха, и имеют ли редкие виды более короткие расстояния между пыльниками и рыльцем.
Наконец, четверная гипотеза касается массовых видов: они, вероятно, меньше нуждаются в самоопылении, так как за счет своей высокой численности в основном получают пыльцу своего вида просто в силу высокой численности независимо от вида опылителей, поскольку их пыльцы в сообществе в любом случае большинство, и она рано или поздно будет перенесена на рыльце. Данная гипотеза была проверена путем выявления связи численности растения с количеством своей пыльцы на рыльцах.
Все четыре гипотезы были проверены с использованием моделирования структурными уравнениями с учетом филогенетических взаимодействий между видами. Моделирование структурными уравнениями (см. Structural equation modeling) представляет собой необычайно мощную технику многомерного анализа.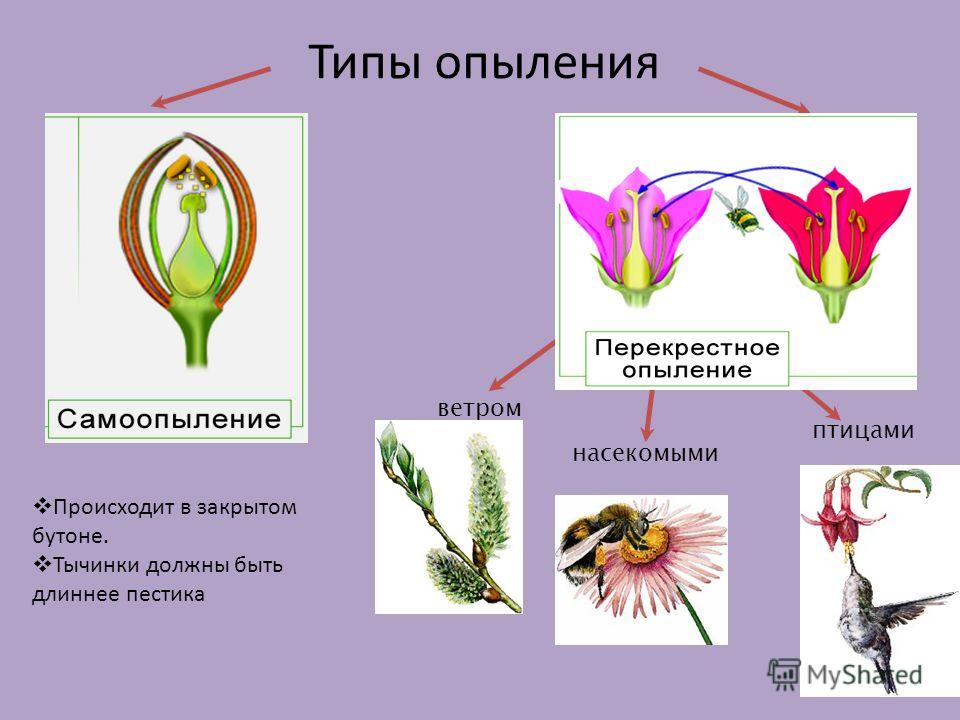 Сперва строится модель, предполагающая, что различные аспекты изучаемого явления связаны друг с другом статистическими и причинно-следственными связями и формируют некоторую структуру. Эта структура описывается системой уравнений, но обычно ее можно представить в виде блок-схемы. Далее на основе экспериментальных данных или наблюдений программа определяет, насколько хорошо данная модель соответствует входным данным. Авторы построили несколько разных моделей, включая и исключая разные гипотезы, и выбрали ту, которая наилучшим образом соответствовала собранным данным из сети взаимодействия растений и опылителей. Это позволило определить, какие из выдвинутых гипотез реализуются в исследуемом сообществе.
Сперва строится модель, предполагающая, что различные аспекты изучаемого явления связаны друг с другом статистическими и причинно-следственными связями и формируют некоторую структуру. Эта структура описывается системой уравнений, но обычно ее можно представить в виде блок-схемы. Далее на основе экспериментальных данных или наблюдений программа определяет, насколько хорошо данная модель соответствует входным данным. Авторы построили несколько разных моделей, включая и исключая разные гипотезы, и выбрали ту, которая наилучшим образом соответствовала собранным данным из сети взаимодействия растений и опылителей. Это позволило определить, какие из выдвинутых гипотез реализуются в исследуемом сообществе.
Для сбора необходимых для данных авторы наблюдали взаимодействия растений и опылителей в течение двух последовательных сезонов цветения на участке 10 000 м2, разбитом на пять площадок на расстоянии 0,3–5 км друг от друга. Каждую площадку посещали один раз в неделю в пик сезона цветения (апрель — июнь): в общей сложности получалось 9–10 недель каждый год. Наблюдения проводились с 8:00 до 17:00 одновременно двумя-тремя людьми. Поскольку авторы стремились собрать равное количество опылителей для каждого вида растений (150 в среднем) равномерно по площадкам и годам, за каждым видом растений наблюдали в течение 20–30 минут в день, увеличивая время для редко посещаемых растений.
Наблюдения проводились с 8:00 до 17:00 одновременно двумя-тремя людьми. Поскольку авторы стремились собрать равное количество опылителей для каждого вида растений (150 в среднем) равномерно по площадкам и годам, за каждым видом растений наблюдали в течение 20–30 минут в день, увеличивая время для редко посещаемых растений.
В итоге ученые зарегистрировали 7324 опылителя, которые посетили 79 видов совместно цветущих растений (62 рода из 29 семейств). Эти опылители относились к 416 видам: 192 вида пчел (4951 особь, перепончатокрылые), 131 вид мух (1409 особей, двукрылые), 35 видов жуков (428 особей, жесткокрылые), 30 видов бабочек и мотыльков (244 особи, чешуекрылые), 14 видов ос (104 особи, перепончатокрылые), 3 вида муравьев (22 особи, перепончатокрылые), 10 других видов насекомых (25 особей) и один вид колибри (141 особь).
В результате полевых наблюдений была составлена сеть растений и опылителей, которая выявила существенные различия в спектре опылителей среди разных видов растений. Были и сверхспециализированные растения, взаимодействующие только с одним видом опылителей, и растения с очень широким кругом опылителей, включающим до 77 видов. Оказалось, что растения, которые цветут в основном одновременно, имеют меньше общих видов-опылителей, чем их должно быть при случайном распределении. Это говорит о разграничении опылителей между видами. Как и предполагалось, малочисленные виды были более специализированными, чем многочисленные.
Были и сверхспециализированные растения, взаимодействующие только с одним видом опылителей, и растения с очень широким кругом опылителей, включающим до 77 видов. Оказалось, что растения, которые цветут в основном одновременно, имеют меньше общих видов-опылителей, чем их должно быть при случайном распределении. Это говорит о разграничении опылителей между видами. Как и предполагалось, малочисленные виды были более специализированными, чем многочисленные.
Чтобы отслеживать приспособленность сообщества на стадии опыления, авторы определили 3,1 млн пыльцевых зерен, которые были расположены на рыльцах совместно цветущих видов (по 54 рыльца каждого из 66 видов) за два года наблюдений за опылителями. Это позволило количественно оценить потерю приспособленности из-за растраты своей пыльцы на рыльцах сосуществующих чужих растений и получения чужой пыльцы.
Оказалось, что лучше всего полученным данным соответствует модель, которая включает в себя только разграничение состава опылителей и асимметричное взаимодействие, но не рассматривает механизмы гарантированного опыления из-за самоопыления или высокой численности. К тому же у малочисленных видов расстояние между рыльцем и пыльником было в среднем больше, что препятствует самоопылению. Этот результат опровергает роль самоопыления в поддержании малочисленных видов по крайней мере в данном сообществе с высоким разнообразием опылителей.
К тому же у малочисленных видов расстояние между рыльцем и пыльником было в среднем больше, что препятствует самоопылению. Этот результат опровергает роль самоопыления в поддержании малочисленных видов по крайней мере в данном сообществе с высоким разнообразием опылителей.
Растения с широким кругом опылителей получали больше чужой пыльцы, что снижало их приспособленность по сравнению со специализированными растениями. Хотя повышенный транспорт пыльцы в целом увеличивал количество и чужой, и своей пыльцы, разнообразие опылителей снижало приспособленность, возможно, из-за потребления пыльцы или других механизмов потери пыльцы во время транспортировки. Таким образом, более специализированные редкие виды действительно получают преимущество. Однако вопреки предсказаниям авторов, растения с широким кругом опылителей не испытывали снижение приспособленности из-за растраты своей пыльцы. Это говорит о том, что разнообразие опылителей и количество посещений, возможно, менее важны, чем такие факторы как эффективность опылителей в переносе пыльцы.
В соответствии с гипотезой асимметричного взаимодействия, редкие виды имели тенденцию получать немного больше чужой пыльцы от сосуществующих растений, чем более многочисленные виды. Тем не менее, опылители, которые поставляют чужую пыльцу, также увеличивают доставку своей пыльцы, что способствует поддержанию этих редких видов. Кроме того, более высокая численность приводит к большей потере приспособленности из-за растраты пыльцы.
Результаты данного исследования подтверждают гипотезу о том, что взаимодействие растений и опылителей играет важную роль в поддержании малочисленных видов в богатых сообществах растений. Однако вопрос о том, универсальны ли данные механизмы для любых сообществ, требует дальнейших исследований.
Очень важно проводить учеты взаимодействий между растениями и опылителями в масштабах всего сообщества для выявления механизмов, которые влияют на успех опыления. Авторы, однако, полностью осознают, что для полного понимание механизмов, поддерживающих разнообразие, требуется дальнейшее изучение того, какие процессы сильнее всего влияют на эффективность размножения растений.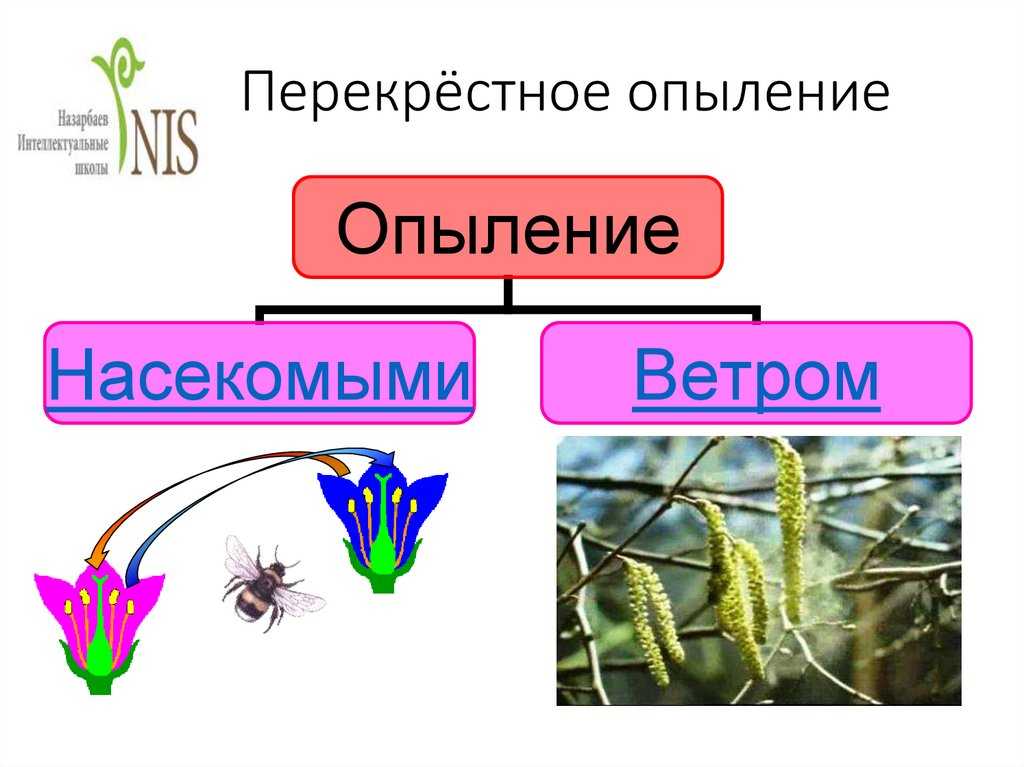 Ведь размножение растений может зависеть не только и не столько от опыления, а, например, от дальнейшего развития семязачатков и доступности необходимых для развития ресурсов. Тем не менее, показанная в обсуждаемой работе роль опыления в поддержании разнообразия растений указывает на необходимость сохранения функционирующих сообществ растений и опылителей, а не только видов растений. Эти соображения как никогда актуальны для сохранения биоразнообразия, поскольку изменения во взаимодействиях растений и опылителей становятся обычным явлением. Сокращение числа опылителей может усилить конкуренцию растений и сделать это в ущерб редким видам, которые являются более специализированными, чем многочисленные виды.
Ведь размножение растений может зависеть не только и не столько от опыления, а, например, от дальнейшего развития семязачатков и доступности необходимых для развития ресурсов. Тем не менее, показанная в обсуждаемой работе роль опыления в поддержании разнообразия растений указывает на необходимость сохранения функционирующих сообществ растений и опылителей, а не только видов растений. Эти соображения как никогда актуальны для сохранения биоразнообразия, поскольку изменения во взаимодействиях растений и опылителей становятся обычным явлением. Сокращение числа опылителей может усилить конкуренцию растений и сделать это в ущерб редким видам, которые являются более специализированными, чем многочисленные виды.
Источник: Na Wei, Rainee L. Kaczorowski, Gerardo Arceo-Gómez, Elizabeth M. O’Neill, Rebecca A. Hayes & Tia-Lynn Ashman. Pollinators contribute to the maintenance of flowering plant diversity // Nature. 2021. DOI: 10.1038/s41586-021-03890-9.
Елена Устинова
Цветочные признаки и адаптация растений к насекомым-опылителям: подход адвоката дьявола
Andersson, S. and B. Widén. 1993. Опосредованный опылителями отбор по цветочным признакам в синтетической популяции Senecio integrifolius (Asteraceae). Ойкос , 66: 72–79.
and B. Widén. 1993. Опосредованный опылителями отбор по цветочным признакам в синтетической популяции Senecio integrifolius (Asteraceae). Ойкос , 66: 72–79.
Перекрёстная ссылка
Google ученый
Арнольд С.Дж. и М. Дж. Уэйд. 1984. Об измерении естественного и полового отбора: Теория. Эволюция , 38: 709–719.
Перекрёстная ссылка
Google ученый
Бейкер Х.Г., 1961. Адаптация цветковых растений к ночным и сумеречным опылителям. кв. преподобный биол. , 36: 64–73.
Перекрёстная ссылка
Google ученый
Бейкер Х. Г. 1963. Эволюционные механизмы в биологии опыления. Наука , 139: 877–883.
перекрестная ссылка
пабмед
КАС
Google ученый
Бейкер, Х. Г. 1983. Очерк истории антэкологии или биологии опыления. В Биология опыления (изд. LA Real), Academic Press, Орландо, Флорида, стр. 7–28.
В Биология опыления (изд. LA Real), Academic Press, Орландо, Флорида, стр. 7–28.
Google ученый
Бейкер Х.Г. и П.Д. Херд. 1968. Внутрифлорная экология. Энн. Преп. Энтомол ., 13: 385–414.
перекрестная ссылка
Google ученый
Битти, А. 1974. Цветочная эволюция в Виола . Энн. Миссури Бот. Гард. , 61: 781–793.
Перекрёстная ссылка
Google ученый
Бертин Р.И. и М.Ф. Уилсон. 1980. Эффективность дневного и ночного опыления двух молочая. Кан. Дж. Бот. , 58: 1744–1746.
Перекрёстная ссылка
Google ученый
Bosch, J. 1986. Insectos floricolas y polinización en un matorral de romero . Неопубликовать диссертация, унив. Барселона, Барселона.
Google ученый
Bosch, J. 1992. Цветочная биология и опылители трех совместно встречающихся видов Cistus (Cistaceae). Бот. Дж. Линн. Soc ., 109: 39–55.
1992. Цветочная биология и опылители трех совместно встречающихся видов Cistus (Cistaceae). Бот. Дж. Линн. Soc ., 109: 39–55.
Перекрёстная ссылка
Google ученый
Бройлс, С.Б. и Р. Вятт. 1990. Анализ отцовства в естественной популяции Asclepias exaltata : множественное отцовство, функциональный пол и «гипотеза донорства пыльцы». Эволюция , 44: 1454–1468.
Перекрёстная ссылка
Google ученый
Кэмпбелл, Д.Р. 1989. Измерения отбора гермафродитного растения: различия в успешности опыления самцов и самок. Эволюция , 43: 318–334.
Перекрёстная ссылка
Google ученый
Дарвин, К. 1862. О различных приспособлениях, с помощью которых британские и зарубежные орхидеи оплодотворяются насекомыми . Мюррей, Лондон.
Google ученый
Darwin, C. 1877. Различные формы цветов на растениях одного и того же вида . Мюррей, Лондон.
1877. Различные формы цветов на растениях одного и того же вида . Мюррей, Лондон.
Google ученый
Дэвис, Массачусетс, 1987. Роль посетителей цветов во взрывном опылении Thalia geniculata (Marantaceae), коста-риканского болотного растения. Бык. Торри Бот. Клуб , 114: 134–138.
Перекрёстная ссылка
Google ученый
Девлин Б. и Н. К. Элстранд. 1990. Изменчивость мужской и женской фертильности у дикой редьки, гермафродита. утра. Нац. , 136: 87–107.
Перекрёстная ссылка
Google ученый
Дудаш М.Р. 1991. Влияние размера растения на женские и мужские функции у гермафродитов Sabatia angularis (Gentianaceae). Экология , 72: 1004–1012.
Перекрёстная ссылка
Google ученый
Экхарт В.М. 1991. Влияние экспозиции цветков на посещаемость опылителей различается среди популяций Phacelia linearis (Hydrophyllaceae). Эволюция. Экол. , 5: 370–384.
Влияние экспозиции цветков на посещаемость опылителей различается среди популяций Phacelia linearis (Hydrophyllaceae). Эволюция. Экол. , 5: 370–384.
перекрестная ссылка
Google ученый
Экхарт В.М. 1992. Пространственно-временная изменчивость численности и изменчивость кормового поведения опылителей гинодиеобразных Phacelia linearis (Hydrophyllaceae). Ойкос , 64: 573–586.
Перекрёстная ссылка
Google ученый
Эндлер, Дж.А. 1986. Естественный отбор в дикой природе . Принстонский университет Press, Принстон, Нью-Джерси.
Google ученый
Фаэгри, К. и Л. ван дер Пейл. 1966. Принципы экологии опыления . Пергамон, Оксфорд.
Google ученый
Фельзенштейн, Дж. 1985. Филогенез и сравнительный метод. утра. Нац. , 125: 1–15.
утра. Нац. , 125: 1–15.
Перекрёстная ссылка
Google ученый
Фенстер, CB 1991. Отбор колибри по морфологии цветков. Биотропика , 23: 98–101.
Перекрёстная ссылка
Google ученый
Футуйма, Д.Дж. 1979. Эволюционная биология . Синауэр, Сандерленд, Массачусетс.
Google ученый
Гален, К. 1989. Измерение опосредованного опылителями отбора по морфометрическим признакам: шмели и альпийский небесный пилот, Polemonium viscosum . Эволюция , 43: 882–890.
Перекрёстная ссылка
Google ученый
Гален, К. и М.Е.А. Ньюпорт. 1988. Качество опыления, набор семян и признаки цветка у Polemonium viscosum : дополнительные эффекты вариации аромата и размера цветка. утра. Дж. Бот. , 75: 900–905.
Перекрёстная ссылка
Google ученый
Гандерс, Ф.Р. 1979. Биология гетеростилии. Н. Рвение. Дж. Бот. , 17: 607–635.
Google ученый
Гизелин, М.Т. 1984. Предисловие к «Различные приспособления, с помощью которых орхидеи оплодотворяются насекомыми », 2-е изд., К. Дарвин. ун-т Chicago Press, Чикаго, Иллинойс (переиздание).
Google ученый
Гиттлман, Дж.Л. и Х.К. Лух. 1992. О сравнении сравнительных методов. Энн. Преподобный Экол. Сист. , 23: 383–404.
Перекрёстная ссылка
Google ученый
Гулд С.Дж. и Э.С. Врба. 1982. Exaptation — недостающий термин в науке о форме. Палеобиология , 8: 4–15.
Google ученый
Гитиан П., Х. Гитиан и Л. Наварро. 1993. Перенос пыльцы и дневное опыление по сравнению с ночным у Lonicera etrusca 9.0005 . Acta Oecol. , 14: 219–227.
1993. Перенос пыльцы и дневное опыление по сравнению с ночным у Lonicera etrusca 9.0005 . Acta Oecol. , 14: 219–227.
Google ученый
Хардер, Л.Д. и С.Ч.Х. Баррет. 1993. Удаление пыльцы с трехцветного растения Pontederia cordata : Влияние положения пыльника и специализации опылителя. Экология , 74: 1059–1072.
Перекрёстная ссылка
Google ученый
Харви, П.Х. и М. Д. Пейгель. 1991. Сравнительный метод в эволюционной биологии . Оксфордский университет Пресс, Оксфорд.
Google ученый
Эррера, К.М. 1986. Рассеянные позвоночные растения: почему они ведут себя не так, как должны. В Frugivores and Seed Dispersing (А. Эстрада и Т. Х. Флеминг, ред.), Junk, Dordrecht, стр. 5–18.
Перекрёстная ссылка
Google ученый
Эррера, К. М. 1987 и . Компоненты «качества» опылителей: сравнительный анализ разнообразного сообщества насекомых. Ойкос , 50: 79–90.
М. 1987 и . Компоненты «качества» опылителей: сравнительный анализ разнообразного сообщества насекомых. Ойкос , 50: 79–90.
Перекрёстная ссылка
Google ученый
Эррера, К.М. 1988. Вариация взаимности: пространственно-временная мозаика сообщества опылителей. биол. Дж. Линн. Soc ., 35: 95–125.
Перекрёстная ссылка
Google ученый
Эррера, К.М. 1992. Исторические эффекты и процессы сортировки как объяснение современных экологических закономерностей: синдромы признаков у средиземноморских древесных растений. утра. Нац. , 140: 421–446.
Перекрёстная ссылка
Google ученый
Эррера, К.М. 1993. Селекция по морфологии цветков и экологическим детерминантам плодовитости фиалки, опыляемой бражником. Экол. моногр. , 63: 251–275.
Перекрёстная ссылка
Google ученый
Herrera, J. 1985. Biologia reproductiva del matorral de Doñana . Неопубликовать диссертация, унив. Севилья, Севилья.
1985. Biologia reproductiva del matorral de Doñana . Неопубликовать диссертация, унив. Севилья, Севилья.
Google ученый
Эррера, Дж. 1988. Взаимоотношения опыления в южных испанских средиземноморских кустарниках. Дж. Экол. , 76: 274–287.
Перекрёстная ссылка
Google ученый
Хорвиц, К.К. и Д.В. Щемске. 1990. Пространственно-временные вариации у насекомых-мутуалистов неотропической травы. Экология , 71: 1085–1097.
Перекрёстная ссылка
Google ученый
Хоу, Х. Ф. 1984. Ограничения эволюции взаимности. утра. Нац. , 123: 764–777.
Перекрёстная ссылка
Google ученый
Иноуэ, Д.В. 1979. Образцы трубчатого венчика цветков шмелей с двух континентов. Мэриленд Агрик. Эксп. Ста. Спец. Разное Опубл. , 1: 461–463.
Google ученый
Иноуэ, Д.В. и Г.Х. Пайк. 1988. Биология опыления в Снежных горах Австралии: сравнение с горным штатом Колорадо, США. австр. Дж. Экол. , 13: 191–218.
Перекрёстная ссылка
Google ученый
Дженнерстен, О. и Д. Х. Морс. 1991. Качество опыления дневными и ночными насекомыми, посещающими молочай обыкновенный, Asclepias syriaca . утра. Средний Нац. , 125: 18–28.
Перекрёстная ссылка
Google ученый
Джордано, П. 1990. Биология размножения трех видов рода Lonicera (Caprifoliaceae) в Сьерра-де-Касорла. Анал. Жард. Бот. Мадрид , 48: 31–52.
Google ученый
Калин Арройо, М.Т., Р. Примак и Дж. Арместо. 1982. Общественные исследования экологии опыления в Андах с умеренным климатом в центральной части Чили. I. Механизмы опыления и высотная изменчивость. утра. Дж. Бот. , 69: 82–97.
I. Механизмы опыления и высотная изменчивость. утра. Дж. Бот. , 69: 82–97.
Перекрёстная ссылка
Google ученый
Кохмер, Дж.П. и С.Н. Гендель. 1986. Ограничения и конкуренция в эволюции фенологии цветения. Экол. моногр. , 56: 303–325.
Перекрёстная ссылка
Google ученый
Кон, Дж.Р. и С.Ч.Х. Баррет. 1992. Экспериментальные исследования функционального значения гетеростилии. Эволюция , 46: 43–55.
Перекрёстная ссылка
Google ученый
Ланде, Р. и С.Дж. Арнольд. 1983. Измерение отбора по коррелирующим признакам. Эволюция , 37: 1210–1226.
Перекрёстная ссылка
Google ученый
Лессиос, Х.А. 1990. Адаптация и филогения как детерминанты размера яиц у иглокожих с двух сторон Панамского перешейка. утра. Нац. , 135: 1–13.
Нац. , 135: 1–13.
Перекрёстная ссылка
Google ученый
Масиор Л.В. 1971. Коэволюция растений и животных — систематические выводы из взаимодействия растений и насекомых. Таксон , 20: 17–28.
Перекрёстная ссылка
Google ученый
МакКолл, С. и Р. Б. Примак. 1992. Влияние характеристик цветков, погоды, времени суток и времени года на посещаемость насекомых в трех растительных сообществах. утра. Дж. Бот. , 79: 434–442.
Перекрёстная ссылка
Google ученый
Милтон С.Дж. и Э.Дж. Молл. 1982. Фенология австралийских акаций ЮЗ. Кейп, Южная Африка, и его последствия для управления. Бот. Дж. Линн. Soc ., 84: 295–327.
Перекрёстная ссылка
Google ученый
Мёллер, А.П. и Т.Р. Беркхед. 1992. Попарный сравнительный метод, иллюстрируемый частотой копуляций у птиц. утра. Нац. , 139: 644–656.
утра. Нац. , 139: 644–656.
Перекрёстная ссылка
Google ученый
Монтальво, А.М. и Дж. Д. Акерман. 1986. Относительная эффективность опылителя и эволюция цветочных признаков у Spathiphyllum friedrichsthalii (Araceae). утра. Дж. Бот. , 73: 1665–1676.
Перекрёстная ссылка
Google ученый
Моттен, А.Ф., Д.Р. Кэмпбелл, Д.Э. Александр и Л. Х. Миллер. 1981. Эффективность опыления посетителей-специалистов и универсалов в популяции Северной Каролины Claytonia virginica . Экология , 62: 1278–1287.
Перекрёстная ссылка
Google ученый
Nilsson, L.A. 1988. Эволюция цветков с глубокими трубками венчика. Природа , 334: 147–149.
Перекрёстная ссылка
Google ученый
Nilsson, L.A., L. Jonsson, L. Rason и E. Randrianjohany. 1985. Механизмы монофилии и опыления у Angraecum arachnites Schltr. (Orchidaceae) в гильдии длинноязычных бражников (Sphingidae) на Мадагаскаре. биол. Дж. Линн. Соц ., 26: 1–19.
Rason и E. Randrianjohany. 1985. Механизмы монофилии и опыления у Angraecum arachnites Schltr. (Orchidaceae) в гильдии длинноязычных бражников (Sphingidae) на Мадагаскаре. биол. Дж. Линн. Соц ., 26: 1–19.
Перекрёстная ссылка
Google ученый
Нильссон, Л.А., Л. Йонссон, Л. Ралисон и Э. Рандрианджохани. 1987. Ангрекоидные орхидеи и бражники в центральном Мадагаскаре: специализированные системы опыления и универсальные собиратели. Биотропика , 19: 310–318.
Перекрёстная ссылка
Google ученый
Олесен, Дж. М. 1985. Макаронезийский элемент «птица-цветок» и его связь с птицами и пчелами-оппортунистами. Бот. Дж. Линн. Soc ., 91: 395–414.
Перекрёстная ссылка
Google ученый
Олесен, Дж. М. 1988. Цветочная биология канарских островов Echium wildpretii : птичий цветок или водный ресурс для пустынных пчел? Акта Бот. Нерл. , 37: 509–513.
Нерл. , 37: 509–513.
Google ученый
Персиваль, М. 1965. Биология цветов . Пергамон, Оксфорд.
Google ученый
Персиваль, М. 1974. Цветочная экология прибрежных кустарников на юго-востоке Ямайки. Биотропика , 6: 104–129.
Перекрёстная ссылка
Google ученый
Петтерссон, М. В. 1991. Опыление гильдией колеблющихся популяций мотыльков: вариант неспециализации в Silene vulgaris . Дж. Экол. , 79: 591–604.
Перекрёстная ссылка
Google ученый
Подолер Х., И. Галон и С. Газит. 1984. Роль нитидуловых жуков в естественном опылении анноны в Израиле. Acta Oecol., Оэкол. заявл. , 5: 369–381.
Google ученый
Цена, м.в. и Н. М. Васер. 1979. Распространение пыльцы и оптимальный ауткроссинг у Delphinium nelsonii . Природа , 277: 294–297.
Распространение пыльцы и оптимальный ауткроссинг у Delphinium nelsonii . Природа , 277: 294–297.
Перекрёстная ссылка
Google ученый
Primack, RB 1983. Опыление насекомыми горной флоры Новой Зеландии. Н. Рвение. Дж. Бот. , 21: 317–333.
Google ученый
Примак, Р. Б. и Х. Канг. 1989. Измерение приспособленности и естественного отбора в популяциях диких растений. Энн. Преподобный Экол. Сист. , 20: 367–396.
Перекрёстная ссылка
Google ученый
Проктор М. и П. Йео. 1973. Опыление цветов . Коллинз, Лондон.
Google ученый
Ramsey, M.W. 1988. Различия в эффективности опылителей прилетающих птиц и насекомых Banksia menziesii (Proteaceae). Экология (Берл.) , 76: 119–124.
Google ученый
Рив, Х. К. и П.В. Шерман. 1993. Адаптация и цели эволюционных исследований. кв. преподобный биол. , 68: 1–32.
К. и П.В. Шерман. 1993. Адаптация и цели эволюционных исследований. кв. преподобный биол. , 68: 1–32.
Перекрёстная ссылка
Google ученый
Рик, К.М. 1950. Отношения опыления Lycopersicon esculentum в местных и зарубежных регионах. Эволюция , 4: 110–122.
Перекрёстная ссылка
Google ученый
Робертсон, Дж. Л. и Р. Вятт. 1990. Доказательства экотипов опыления желтобахромчатой орхидеи, Platanthera ciliar is . Эволюция , 44: 121–133.
Перекрёстная ссылка
Google ученый
Шемске, Д.В. 1983. Пределы специализации и коэволюции мутуализма растений и животных. В Coevolution (MH Nitecki, ed.), Chicago Univ. Press, Чикаго, Иллинойс. стр. 67–109.
Google ученый
Шемске, Д.В. и К.С. Хорвиц. 1984. Различия среди цветочных посетителей в способности к опылению: предварительное условие специализации мутуализма. Наука , 225: 519–521.
1984. Различия среди цветочных посетителей в способности к опылению: предварительное условие специализации мутуализма. Наука , 225: 519–521.
Перекрёстная ссылка
пабмед
КАС
Google ученый
Шемске, Д.В. и К.С. Хорвиц. 1988. Взаимодействие растений и животных и производство плодов у неотропических трав: анализ пути. Экология , 69: 1128–1137.
Перекрёстная ссылка
Google ученый
Шемске, Д.В. и К.С. Хорвиц. 1989. Временная вариация отбора по цветочному признаку. Эволюция , 43: 461–465.
Перекрёстная ссылка
Google ученый
Шлютер Д. 1988. Оценка формы естественного отбора по количественному признаку. Эволюция , 42: 849–861.
Перекрёстная ссылка
Google ученый
Смит, Дж.М.Б. 1988. Prunus (Amygdalaceae) в Новом Южном Уэльсе. Телопеа , 3: 145–157.
Телопеа , 3: 145–157.
Google ученый
Сноу, А.А. и Д.В. Рубик. 1987. Осаждение и удаление пыльцы пчелами, посещающими два вида деревьев в Панаме. Биотропика , 19: 57–63.
перекрестная ссылка
Google ученый
Сокаль Р.Р. и К.А. Брауманн. 1980. Тесты значимости для коэффициентов вариации и профилей изменчивости. Сист. Зоол. , 29: 50–66.
Перекрёстная ссылка
Google ученый
Стеббинс Г.Л. 1970. Адаптивная радиация репродуктивных характеристик покрытосеменных растений. I. Механизмы опыления. Энн. Преподобный Экол. Сист. , 1: 307–326.
Перекрёстная ссылка
Google ученый
Сагден, Э.А. 1986. Антекология и эффективность опыления Styrax officinale subsp. redivivum (Стираковые). утра. Дж. Бот. , 73: 919–930.
Дж. Бот. , 73: 919–930.
Перекрёстная ссылка
Google ученый
Фогель С., К. Вестеркамп, Б. Тиль и К. Гесснер. 1984. Орнитофилия на Канаришен Инзельн. Завод Сист. Эвол. , 146: 225–248.
Перекрёстная ссылка
Google ученый
Waser, N.M. 1983 a . Адаптивный характер цветочных признаков: идеи и доказательства. В Oplination Biology (LA Real, ed.), Academic Press, Орландо, Флорида, стр. 241–285.
Google ученый
Waser, N.M. 1983 b . Конкуренция за опыление и различия в характере цветков среди симпатрических видов растений: обзор доказательств. В Справочник по экспериментальной биологии опыления (CE Jones и RJ Little, ред.), Van Nostrand Reinhold, New York, стр. 277–293.
Google ученый
Васер Н. М. и М.В. Цена. 1990. Эффективность опыления и эффективность посещения шмелями и колибри Delphinium nelsonii . Собрать. Бот. (Барселона) , 19:9–20.
М. и М.В. Цена. 1990. Эффективность опыления и эффективность посещения шмелями и колибри Delphinium nelsonii . Собрать. Бот. (Барселона) , 19:9–20.
Перекрёстная ссылка
Google ученый
Уильямс, Г.К. 1966. Адаптация и естественный отбор . Принстонский университет Press, Принстон, Нью-Джерси.
Google ученый
Williamson, M. 1972. Анализ биологических популяций . Арнольд, Лондон.
Google ученый
Уилсон П. и Дж. Д. Томсон. 1991. Неоднородность среди цветочных посетителей приводит к несоответствию между удалением и отложением пыльцы. Экология , 72: 1503–1507.
Перекрёстная ссылка
Google ученый
Wolda, H. 1983. «Долгосрочная» стабильность популяций тропических насекомых. Рез. Поп. Экол. , Доп. 3: 112–126.
, Доп. 3: 112–126.
Перекрёстная ссылка
Google ученый
Вулф Л.М. и С.Ч.Х. Баррет. 1989. Закономерности удаления и отложения пыльцы у трехцветных растений Pontederia cordata L. (Pontederiaceae). Биол. Дж. Линн. Soc ., 36: 317–329.
Перекрёстная ссылка
Google ученый
Циммерман, Дж.К., Д.В. Рубик и Дж. Д. Акерман. 1989. Асинхронная фенология неотропической орхидеи и ее пчелы-опылителя эуглоссина. Экология , 70: 1192–1195.
Перекрёстная ссылка
Google ученый
Адаптация к опылению
Адаптация к опылению
Цветы нуждаются в опылении. Опыление – это процесс перемещения пыльцевого зерна с тычинки тычинки на рыльце пестика плодолистика. Есть несколько цветов, которые могут самоопыляться сами по себе, но это ограничивает их инбридинг. Большинство видов полагаются на какой-либо вектор опыления для опыления. Переносчиком может быть любой агент, способный перемещать пыльцу с пыльника на рыльце пестика.
Переносчиком может быть любой агент, способный перемещать пыльцу с пыльника на рыльце пестика.
Имеются данные о том, что вода и ветер являются переносчиками опыления у некоторых видов, но многие виды не зависят от случайных или расположенных только вниз по течению путей опыления, предлагаемых этими переносчиками. Действительно, такие переносчики полезны только в ситуациях, когда присутствуют большие популяции очень ограниченного числа видов.
Большинство цветов эволюционировали, чтобы использовать вектор «умной бомбы» или «волшебной пули»… животные! У этих переносчиков есть органы чувств, чтобы находить цветы, у них есть способность передвигаться, чтобы добраться до цветов, несмотря на большие промежутки между особями, и у них достаточно интеллекта, чтобы помнить, что они могут рассчитывать на вознаграждение, если они неоднократно посещают один конкретный вид.
Чтобы эффективно использовать вектор опыления животными, цветок должен привлечь животное при первом посещении. Это можно сделать одним из двух способов: визуальными сигналами и обонятельными сигналами.
Это можно сделать одним из двух способов: визуальными сигналами и обонятельными сигналами.
Визуальные подсказки
Некоторые виды используют цветовой узор, известный как «бычий глаз», чтобы отметить свое положение в окружающей среде… чтобы выделиться на фоне зеленой листвы. Очевидно, что не так уж много цветков, опыляемых животными, имеют зеленый цвет листьев!
Ниже представлена черноглазая сьюзен… напомню, что существует более 100 видов неродственных черноглазых сьюзан… одна из проблем общих названий. Этот из рода Рудбекия . Обратите внимание на его очевидный внешний вид. Бычий глаз черный и окружен желтым кольцом, что делает его заметным.
Лилейник ( Hemerocallis ) ниже также имеет заметный бычий глаз, но обратите внимание на то, что цветовой узор перевернут. Бычий глаз имеет светлый цвет, а кольцо вокруг него очень темное. Эффект тот же, однако цветок будет очевиден для потенциального опылителя, летящего над головой.
Важно, чтобы любой используемый цветовой узор находился в пределах видимого диапазона цветов, наблюдаемых опылителем. Цвета, очевидные для одного животного, могут быть невидимы для другого. Растение, показанное ниже, называется Gaillardia (окрашенная ромашка). Этот вид, очевидно, использует желтый бычий глаз с окружающими его красными и желтыми кольцами. При взгляде на него создается впечатление мишени для дартса. Наблюдаемый опылитель — бабочки. Бабочек определенно привлекают красно-желтые узоры, и им нужна большая посадочная площадка, обеспечиваемая этим широким цветком.
С другой стороны, Gaillardia не ограничивает свое опыление только бабочками. Послеобеденное наблюдение покажет вам, что медоносные пчелы совершают еще больше визитов! Пчелы не способны воспринимать красный цвет, так как же этот красно-желтый узор выглядит для пчелы? Это требует некоторого мысленного перевода и специальной фотографии, чтобы человек мог оценить то, что может наблюдать пчела.
Сначала нам нужно убрать красный цвет с изображения. Этого можно добиться только с помощью причудливой видеоработы или черно-белой фотографии. Вот две нарисованные ромашки на типичном черно-белом фото:
Обратите внимание, как красно-желтый узор теперь отображается в темном/светлом узоре. Это черно-белый вид цветка в человеческом видении.
Ниже приведена фотография тех же двух ромашек, сделанная через несколько секунд с красным фильтром на объективе камеры:
Обратите внимание, как этот красный фильтр удалил большую часть темного рисунка, вызванного красным пигментом. Таким образом, фотография выше показывает, как выглядят цветы в остальной части спектра человеческого зрения (оранжевый, желтый, зеленый, синий, фиолетовый). В основном почти все отражает эти цвета света.
Пчелиное зрение не включает красный цвет, но оно распространяется на ультрафиолет. Другими словами, пчелы видят невидимые нам цвета! Наша пленка не очень чувствительна к ультрафиолету, потому что она была разработана для демонстрации изображений, видимых человеку.
Чтобы исправить это и увидеть ультрафиолетовые узоры в этих цветах, нам нужно использовать еще один фильтр. Этот фильтр блокирует видимый свет и позволяет ультрафиолетовому излучению подвергать пленку воздействию. Ниже приведено ультрафиолетовое изображение тех же двух маргариток, сделанное через несколько секунд с обоими фильтрами:
Обратите внимание, как центр каждой ромашки полностью поглощает ультрафиолетовый свет. Лепестки по периметру отражают ультрафиолет. Таким образом, бычий глаз отражает все цвета пчелиного зрения, кроме УФ. Поскольку УФ-излучение является пчелиным эквивалентом человеческого фиолетового, бычий глаз можно рассматривать как глубокий пчелино-желтый… почти пчелино-оранжевый. Кольцо по периметру отражает все цвета пчелиного зрения, включая УФ, поэтому кольцо кажется пчелино-белым.
Таким образом, красно-желтая маргаритка, видимая человеку и бабочке, кажется пчеле бело-желтой маргариткой. Таким образом, цветок может использовать любого из двух опылителей: бабочек или пчел.
 Чтобы исправить это и увидеть ультрафиолетовые узоры в этих цветах, нам нужно использовать еще один фильтр. Этот фильтр блокирует видимый свет и позволяет ультрафиолетовому излучению подвергать пленку воздействию. Ниже приведено ультрафиолетовое изображение тех же двух маргариток, сделанное через несколько секунд с обоими фильтрами:
Чтобы исправить это и увидеть ультрафиолетовые узоры в этих цветах, нам нужно использовать еще один фильтр. Этот фильтр блокирует видимый свет и позволяет ультрафиолетовому излучению подвергать пленку воздействию. Ниже приведено ультрафиолетовое изображение тех же двух маргариток, сделанное через несколько секунд с обоими фильтрами:
Еще один способ привлечь опылителей — использовать направляющие для нектара, как у дикой гвоздики, показанной ниже:
Проводники нектара — это цветовые узоры, исходящие из источника нектарной награды. Подобно краске на взлетно-посадочной полосе в аэропорту, они направляют опылителя к награде.
Обонятельные сигналы
Некоторые опылители более обонятельные, чем зрительные, и, конечно, некоторые используют оба чувства. Цветы развили ароматы, и это приводит к эффективному привлечению опылителей. Опять же, разные животные имеют разную чувствительность и чувствительность к ароматам.
Существует растение под названием Sauromatum или лилия вуду, у которого стебель цветка завёрнут в пурпурно-чёрный лист. Он эволюционировал, чтобы опыляться мухами. Наверняка черноватый цвет листа напоминает гниющую плоть или навоз (пищевые и яйцекладущие предметы). Аромат, испускаемый этим аппаратом, очень силен… людям на ум приходит коровник.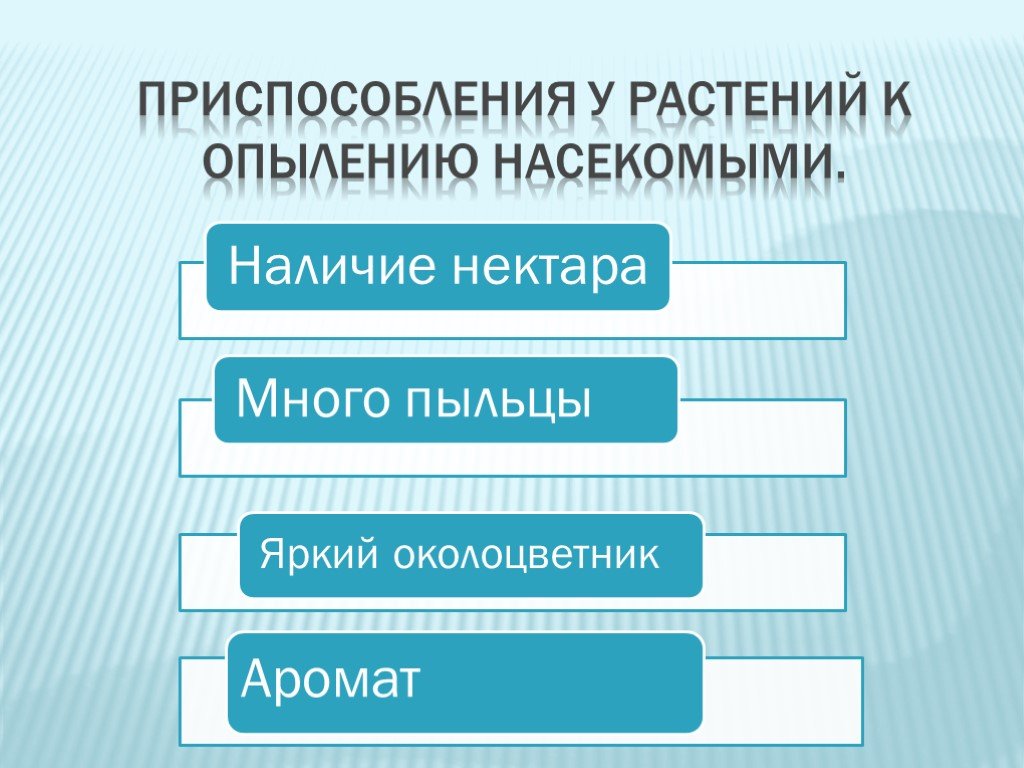 Мухам очень нравится этот аромат, но он не привлекает (и даже не отталкивает) многих других видов.
Мухам очень нравится этот аромат, но он не привлекает (и даже не отталкивает) многих других видов.
Люди не так чувствительны к ароматам, как многие другие животные, и мы, конечно, не разработали очень хорошую терминологию для описания ароматов. Тем не менее, мы ценим некоторые ароматы, такие как гераниол из герани и розы. Бабочки и птицы не очень обоняние. Они гораздо более визуальны в поведении. Пчел привлекают определенные запахи, особенно те, которые мы можем описать как сладкие или пряные. Мотыльки и летучие мыши очень обонятельны и не слишком хорошо ориентируются визуально. Цветы, опыляемые летучими мышами, обычно издают сильный фруктовый или мускусный аромат, в то время как цветы, опыляемые мотыльками, издают очень пьянящие сладкие ароматы.
Как только опылитель совершил первую посадку, цветок должен вознаградить животное, чтобы оно восприняло награду в результате своего визита. Затем его интеллект позволит ему решить посетить похожие цветы поблизости, чтобы получить дополнительные награды. .. перенося пыльцу с растения на растение в процессе. Наградой может быть нектар, пыльца, поведение или их комбинация.
.. перенося пыльцу с растения на растение в процессе. Наградой может быть нектар, пыльца, поведение или их комбинация.
Награды за нектар
Нектар в основном экссудат из флоэмы. Он вырабатывается в структурах цветков, называемых нектарниками. Нектарник может быть на любой части цветка и представляет собой просто участок эпидермиса, состоящий из множества постоянно открытых устьиц. Под эпидермисом нектарника находятся окончания жилок, которые постоянно выгружают сахар из флоэмы. Вода вытекает путем осмоса, и результатом является «кровотечение» сладкой жидкости через устьица. Нектар может накапливаться в основании цветка или, возможно, даже в длинном мешочке, называемом нектарной шпорой:
Фиалки выше показывают сине-желтый узор с направляющими для нектара. Отверстие в «бычьем глазу» соединяется с мешочком в одном из лепестков… нектарной шпорой. Это видно в цветке, который сбоку (слева) на фотографии выше. За основными лепестками видна заостренная шпорца. Эта шпора содержит каплю нектара. Пчела должна откачать каплю со дна шпорца, вставив свой хоботок в центр цветка и вниз к основанию шпорца нектара.
Эта шпора содержит каплю нектара. Пчела должна откачать каплю со дна шпорца, вставив свой хоботок в центр цветка и вниз к основанию шпорца нектара.
Вот некоторые цветы водосбора Aquilegia :
Вы, наверное, заметили, что цветки водосбора имеют красно-желтую окраску, привлекающую бабочек или птиц. Кроме того, вы можете заметить, что цветок наклоняется вниз. Опылитель должен находиться ниже цветка и направлять свой хоботок вверх, чтобы вставить его в очень длинные шпорцы нектара. Очевидно, что бабочки так не парят, но колибри ДОЛЖНЫ! Это кивающее положение цветка с длинным путем нектара характерно для цветов колибри.
Какой длины самая длинная нектарная шпора? Ну, есть орхидея Habenaria , обнаруженная Дарвином, у которой есть 11-дюймовая шпора! Дарвин предположил, что существовала некая моль (цветок ароматный и с белыми цветками) с 11-дюймовым хоботком, который мог бы стать опылителем. Разочарованный, он так и не нашел мотылька. Пятьдесят лет спустя мотылек был найден, его хоботок действительно 11 дюймов в длину, и он действительно опыляет орхидею!
Награды за пыльцу
Цветы, особенно цветы, опыляемые жуками, должны обеспечивать дополнительную пыльцу для пищевых нужд опылителей. Пчелы также питаются пыльцой. В то время как нектар в основном состоит из сахара и очень калорийной пищи для пчел и колибри, пыльца является источником белка, витаминов и минералов. К счастью для растений, опылители питаются пыльцой, но не привередливы в уходе за собой, поэтому в конечном итоге они оставляют хотя бы несколько пыльцевых зерен на рыльце пестика. Один взгляд на цветок мака показывает, сколько пыльцы необходимо, чтобы удовлетворить жука!
Пчелы также питаются пыльцой. В то время как нектар в основном состоит из сахара и очень калорийной пищи для пчел и колибри, пыльца является источником белка, витаминов и минералов. К счастью для растений, опылители питаются пыльцой, но не привередливы в уходе за собой, поэтому в конечном итоге они оставляют хотя бы несколько пыльцевых зерен на рыльце пестика. Один взгляд на цветок мака показывает, сколько пыльцы необходимо, чтобы удовлетворить жука!
Награды за поведение
Иногда цветок может сойти с рук с минимальным вознаграждением… позволив животному стать вознаграждением. Это некоторые орхидеи-башмачки ( Cypripedium ).
Обратите внимание на желтый лепесток, который превратился в своеобразный мешочек. Этот мешочек удерживает внутри химический ароматизатор, перед которым мухи не могут устоять. Мухи приземляются рядом с отверстием мешка и перелезают через край, попадая внутрь. Аромат опьяняет мух. Они спотыкаются и так далее в течение некоторого времени. Они взбираются по задней части сумки к двум «окнам», через которые в сумку льется свет. На выходе мешочки с пыльцой прилипают к их спине. Они так хорошо провели время, они посещают второй мешочек с орхидеей, и во время этого посещения мешочки с пыльцой из первого посещения удаляются у мухи с помощью рыльца … опыление достигнуто!
Они взбираются по задней части сумки к двум «окнам», через которые в сумку льется свет. На выходе мешочки с пыльцой прилипают к их спине. Они так хорошо провели время, они посещают второй мешочек с орхидеей, и во время этого посещения мешочки с пыльцой из первого посещения удаляются у мухи с помощью рыльца … опыление достигнуто!
Возможно, самая причудливая адаптация цветов к поведению животных обнаружена у орхидеи Ophrys (извините, этого слайда нет… крысы!). Эта орхидея выглядит как самка осы и даже пахнет ею. Аромат орхидеи собирают самки ос и используют в качестве полового аттрактанта для самцов (фермомон). Первыми зацветают орхидеи. Затем из куколок выходят осы-самцы. Они пахнут женщинами! Они летят к источнику аромата… вот они! Они пытаются спариваться с «самками», но это всего лишь цветы. Пыльцевые мешочки прилипают к самцу осы. Разочарованный самец переходит к «псевдокопуляции» с другими цветами. Пыльцевые мешочки удаляются, и достигается опыление. Наконец появляются настоящие осы-самки, посещают увядающие цветы орхидеи, чтобы собрать феромон, и начинают «появляться» в ароматическом профиле ос-самцов. Он спаривается с настоящей самкой, это работает! И осы тоже размножаются. Если подумать, эти два вида изящно синхронизированы, химически и структурно связаны, так что ни один из них не может правильно воспроизводиться без другого. Удивительно!
Он спаривается с настоящей самкой, это работает! И осы тоже размножаются. Если подумать, эти два вида изящно синхронизированы, химически и структурно связаны, так что ни один из них не может правильно воспроизводиться без другого. Удивительно!
Другие примеры, когда форма имеет значение, включают обычный тропический цветок гибискуса с лучшим сочетанием цветов для привлечения птиц:
На виде спереди вы видите выравнивание «бычий глаз». Колибри должен совместить красный и желтый бычий глаз с темно-красным бычим глазом в основании цветка, чтобы направить свой клюв к награде. Пять темно-красных объектов, торчащих дальше всего, — это пять стигм. Желтые объекты — это пыльники множества сросшихся тычинок.
Вид сбоку показывает вам подход, который должен предпринять колибри, чтобы получить награду и добиться опыления. Птица парит перед цветком. Когда он приближается, его голова ударяется о рыльца. Он подходит ближе, голова трется о пыльники и покрывается пыльцой. Птица забирает награду. Он переходит к следующему цветку. Его голова ударяется о рыльца… откладывая пыльцу с предыдущего цветка… затем он приближается, собирая пыльцу со второго цветка на свою голову… и получает награду. Это продолжается так, что каждый кивающий цветок опыляет следующий, помещая пыльцу в головные перья этой парящей птицы!
Птица забирает награду. Он переходит к следующему цветку. Его голова ударяется о рыльца… откладывая пыльцу с предыдущего цветка… затем он приближается, собирая пыльцу со второго цветка на свою голову… и получает награду. Это продолжается так, что каждый кивающий цветок опыляет следующий, помещая пыльцу в головные перья этой парящей птицы!
Аналогичное пространственное расположение имеет и этот лилейник. Рыльце выходит прямо наружу и первым поражает опылителя. Тычинки гравитропны и изгибаются вверх, так что пыльца попадает на опылителя после рыльцевого контакта. Награда находится в основании яблочка.
Ниже приведен еще один пример опыления птицами… в Tillandsia , бромелиевой родственнице ананаса.
Обратите внимание, что Тилландсия цветки не кивают, а смотрят вверх! Цветовой узор подходит птицам, но колибри не сможет собирать нектар с этого цветка. Угол неправильный! Вместо этого его опыляют птицы-птицы.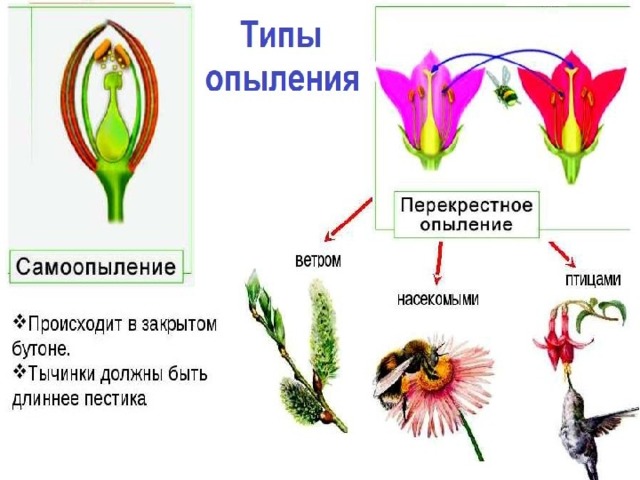 Конечно, вам нужны насесты, так как сидящие птицы не могут (или не парят) в воздухе. Красноватые листья вокруг пурпурных цветов очень жесткие и обеспечивают необходимые опоры для птиц. Птицы собирают нектар внутри трубчатого цветка и опыляют его так же, как колибри. Обратите внимание на то же относительное положение белых рыльцов и желтых пыльников, но, конечно, эта птица наклоняет голову вниз из положения сидя.
Конечно, вам нужны насесты, так как сидящие птицы не могут (или не парят) в воздухе. Красноватые листья вокруг пурпурных цветов очень жесткие и обеспечивают необходимые опоры для птиц. Птицы собирают нектар внутри трубчатого цветка и опыляют его так же, как колибри. Обратите внимание на то же относительное положение белых рыльцов и желтых пыльников, но, конечно, эта птица наклоняет голову вниз из положения сидя.
Теперь о некоторых проблемах формы бабочки:
Вышеупомянутое растение — молочай ( Asclepias ). Опыляется бабочками. Вы заметите, что цветок, показанный с точки зрения приземления, имеет очень небольшую «посадочную платформу», но использует мотив красно-розовой бабочки:
Чтобы бабочка приземлилась здесь, ее лапки должны пройти между чашеобразными лепестками, на каждом из которых находится капелька нектара. Там, где лапки цепляются за стебель, между лепестками находится кусочек ткани (пыльник), покрытый каким-то ботаническим «суперклеем». Бабочка пьет из чашечек, но, улетая, вырывает из цветка пыльцевые мешочки. Они приклеены к его ногам!
Бабочка пьет из чашечек, но, улетая, вырывает из цветка пыльцевые мешочки. Они приклеены к его ногам!
Вид сбоку выше показывает зеленоватые подушечки, где находятся клей и пыльцевые мешочки. Когда бабочка садится на следующий цветок, мешочки с пыльцой отрываются от рыльца пестика и происходит опыление.
В Коннектикуте нашим государственным цветком является горный лавр ( Kalmia ). У него прекрасный механизм опыления!
Ниже приведены некоторые цветочные бутоны лавра на разных стадиях развития:
Обратите внимание на куполообразную форму бутонов посередине с десятью мешочками по краям. Внутри этих бутонов находится десять тычинок. Они сильно разрастаются до раскрытия цветка. По мере роста нитей пыльники ударяются о вершину купола закрытой почки. Продолжающийся рост нити направляет пыльники между гребнями в куполе, так что в конечном итоге пыльники оказываются в десяти мешочках. Этот изгиб нити обеспечивает силы растяжения/сжатия в нити. Затем раскрывается бутон цветка:
Затем раскрывается бутон цветка:
Развертывание околоцветника, очевидно, дополнительно увеличивает силы растяжения/сжатия в нитях с пыльниками, захваченными карманами. Если внимательно присмотреться к цветку ближе к центру, то можно увидеть красное кольцо вокруг «бычьего глаза». Вы можете увидеть белый столбик с зеленым рыльцем, выходящим из середины, и десять дугообразных нитей с темными пыльниками, захваченными мешочками. Этот цветок раскрылся, но еще не был посещен. На этот цветок садится опылитель. Вы можете себе представить, что происходит! Потенциальная энергия, выраженная как силы натяжения/сжатия в нитях, преобразуется в кинетическую энергию, когда вес опылителя сотрясает пыльники, высвобождаемые из мешочков.
Бах, бах, бах, бах, бах, бах, бах, бах, бах, бах! Опылителя бьют десятью тычинками со всех сторон и со всех сторон тщательно опыляют пыльцой! Когда он приземляется на другой только что распустившийся цветок, его тело сначала касается выступающего рыльца, и происходит опыление.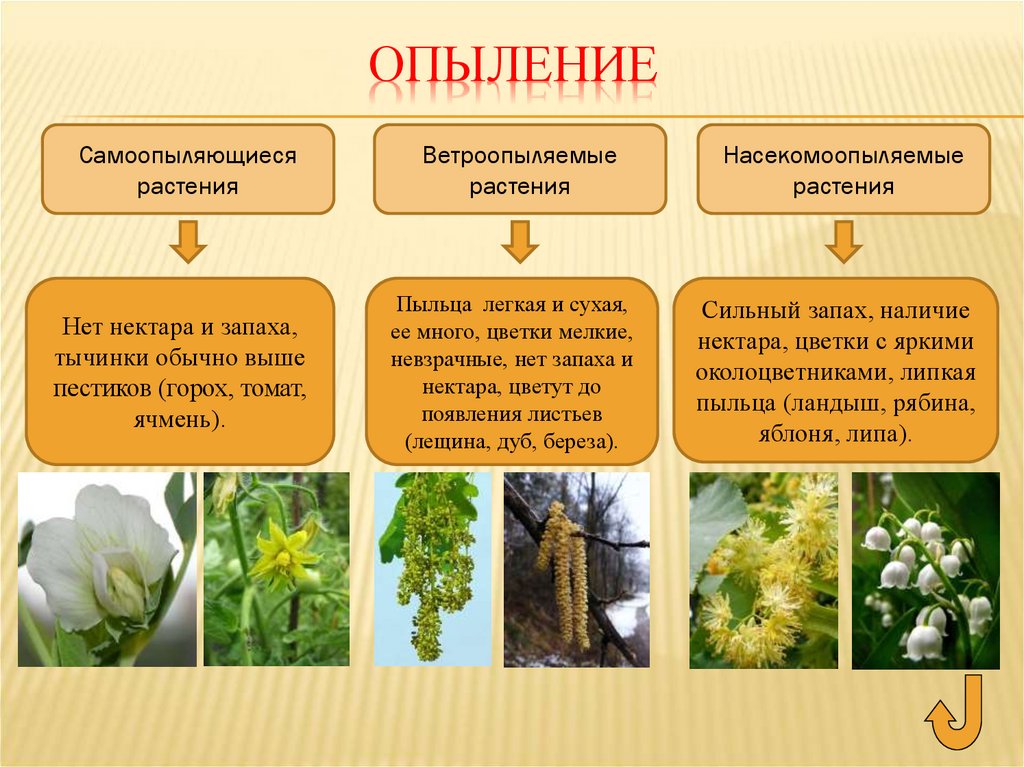
Вы могли заметить, что несколько других цветов на фотографии выше были посещены. Как вы можете сказать?
В следующий раз, когда вы увидите горный лавр в цвету (май-июнь), поднимитесь и ткните пальцем в только что распустившийся цветок и посмотрите, что произойдет! Это довольно весело, но не забывайте также выполнять свою работу по опылению!
В качестве прощального выстрела я решил показать вам пример растения, опыляемого ветром, а не животным.
Помните, что ветер переносит пыльцу только по ветру, так что это не «волшебная пуля». Гравитация также будет воздействовать на пыльцу, поэтому опыление вниз более вероятно, чем опыление вверх. Нет способа привлечь ветер… так что цвета и ароматы будут пустой тратой времени. Такие награды, как нектар или поведение, безусловно, не имеют значения. Вам не нужны большие посадочные платформы, шпоры для нектара, мешочки, насесты или другие приспособления, связанные с комфортом существ.
На фото выше кукурузное поле имеет растения с мужскими цветками на верхушке растения. Они называются кисточками. Женские цветки находятся в «ушках» внизу на стебле. Это было бы нисходящим направлением опыления. Обратите внимание, что кукуруза эволюционировала, чтобы расти большими популяциями, поэтому, вероятно, рыльце будет с подветренной стороны от пыльника. Конечно, важно, чтобы мы выращивали кукурузу таким же образом. Если вы сажаете в своем саду только несколько растений кукурузы, помните, что один ряд будет хорошо опыляться только в том случае, если преобладающий ветер дует по всей длине ряда. Поскольку вы не можете быть в этом уверены, лучше сажать кукурузу прямоугольными блоками в несколько коротких рядов (миниатюрная версия поля, показанного выше), а не в один длинный ряд.
Они называются кисточками. Женские цветки находятся в «ушках» внизу на стебле. Это было бы нисходящим направлением опыления. Обратите внимание, что кукуруза эволюционировала, чтобы расти большими популяциями, поэтому, вероятно, рыльце будет с подветренной стороны от пыльника. Конечно, важно, чтобы мы выращивали кукурузу таким же образом. Если вы сажаете в своем саду только несколько растений кукурузы, помните, что один ряд будет хорошо опыляться только в том случае, если преобладающий ветер дует по всей длине ряда. Поскольку вы не можете быть в этом уверены, лучше сажать кукурузу прямоугольными блоками в несколько коротких рядов (миниатюрная версия поля, показанного выше), а не в один длинный ряд.
Вот крупный план мужских цветков. Это стебель с несколькими мужскими цветками. У каждого цветка по три свисающие вниз тычинки. Нити очень тонкие и шевелятся при малейшем ветерке (тем лучше распределяется пыльца из пыльников). Пыльники большие и производят большое количество желтых пыльцевых зерен. Зерна сухие (не липкие), поэтому они плавают в воздухе по отдельности, а не слипаются гроздьями и не падают на землю.
Зерна сухие (не липкие), поэтому они плавают в воздухе по отдельности, а не слипаются гроздьями и не падают на землю.
На кукурузных полях и поблизости разносится столько пыльцы, что ваша машина покрыта… буквально пылью… пыльцы. Слава богу, пыльца кукурузы, по-видимому, не очень аллергенна!
Рыльце женских цветков называется шелком. Внизу початок кукурузы (внутри зеленые листья, называемые шелухой). На концах шелухи выступают желтые рыльца. Они очень длинные, липкие и пушистые… чтобы лучше ловить любые плавающие зерна пыльцы. На каждое зернышко кукурузного початка приходится одно рыльце! Каждое рыльце должно получить пыльцевое зерно, чтобы его ядро могло правильно развиваться.
В следующий раз, когда перед вами будет початок кукурузы, вы можете подумать о том, сколько цветов нужно, чтобы сделать один початок кукурузы. Сколько шелков пришлось снять с него? Сколько пыльцевых зерен нужно было собрать? Удивительно!
Эта страница © Росс Э.