
Содержание
Биология. Растения, бактерии, грибы, лишайники. 6 класс. Подготовка к ЕГЭ. Контрольные и самостоятельные работы
книги
Детская литература
Старше 12 лет
Учебная литература
Пособия и справочники для сдачи экзаменов и поступления в ВУЗы
ЕГЭ
Код 521514
- ISBN: 978-5-699-20022-1
- 160 страниц
- июль 2007
- Эксмо
- Мастер-класс для учителя
- 120 г
Бумажная книга
Нет в продаже
Вместе с этой книгой покупают
Волшебники страны Однаждыбумажная книга
Крессида Коуэлл
13. 6 USD
6 USD
В корзину
Модульное вязание на спицах. 100 круговых…бумажная книга
Ники Эпштейн
8.74 USD
В корзину
Избранные сочинениябумажная книга
Александр Чехов
11.48 USD
В корзину
Попугаи для «чайников»бумажная книга
Никки Мустаки
Нет в продаже
Почему они не работают? Новый взгляд на…бумажная книга
Сьюзен Фаулер
Нет в продаже
-23%
Горький квест. Том первыйбумажная книга
Александра Маринина
15. 19 USD 11.69 USD
19 USD 11.69 USD
В корзину
Видеоурок по биологии «Бактерии. Простейшие. Грибы и лишайники»
Все растения подразделяют на низшие и высшие. Высшие растения подразделяют на споровые и семенные.
Покрытосеменные растения относятся к высшим семенным растениям.
Покрытосеменные очень разнообразны. Среди них имеются вечнозелёные и листопадные деревья, а также кустарники и полукустарники, однолетние и многолетние травы.
Все покрытосеменные, несмотря на своё многообразие, имеют общий план строения.
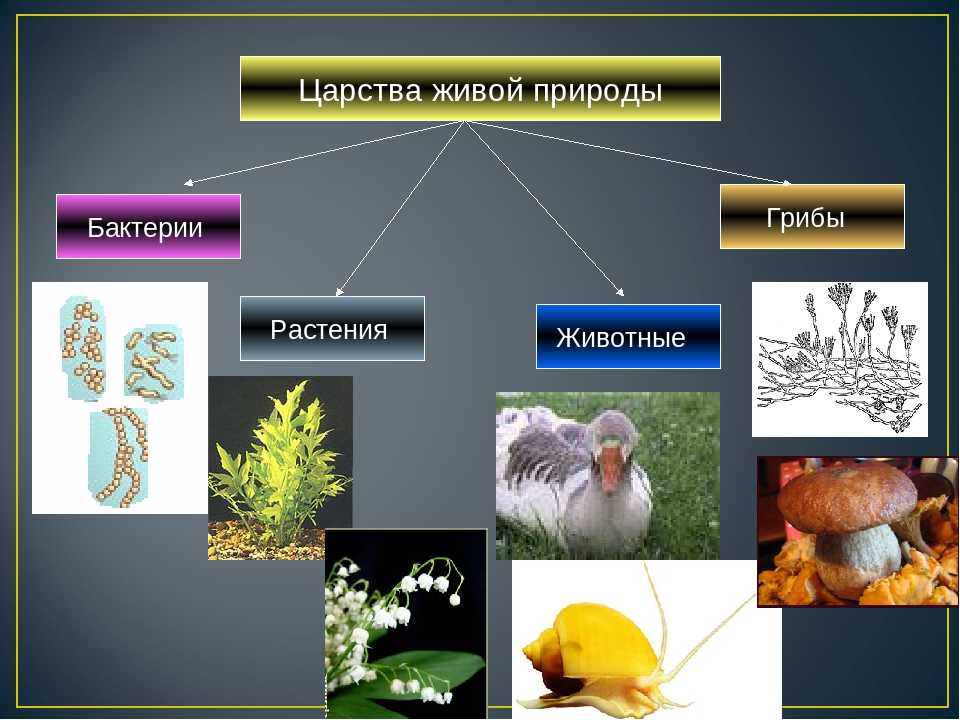
Все живые организмы, обитающие на Земле, делят на две империи — это неклеточные организмы и клеточные организмы.
А также на два надцарства Прокариоты ― живые организмы, состоящие из клеток, которые не имеют клеточного ядра и мембранных органелл.
Эукариоты ― живые организмы, клетки которых содержат ядро, а также мембранные органеллы.
Отдельно выделяют царства Вирусы и Бактериофаги — неклеточные формы жизни.
А также царство Дробянки (это бактерии и цианобактерии), которые относятся к надцарству Прокариоты.
А надцарство Эукариоты включает царство Растения, Грибы и Животные.
Бактерии — это относительно просто устроенные микроскопические одноклеточные организмы. Они имеют разные формы.
Некоторые бактерии для передвижения используют реснички и жгутики, которые дают им возможность перемещаться в жидкой среде в поисках более благоприятных условий.
Вспомним строение бактерий. В отличие от клеток растений, животных и грибов, клетки бактерий имеют упрощённое строение и не имеют многих органелл.
Как известно, бактерии очень жизнеспособны, могут выдержать как высокие, так и низкие температуры. И все это благодаря плотной клеточной стенке, которая сохраняет постоянную форму.
Сверху клеточную стенку многих бактерий окружает слизистая структура — капсула бактерий.
Под клеточной стенкой располагается цитоплазматическая мембрана, которая отделяет цитоплазму от клеточной стенки.
Ядра, отделённого от цитоплазмы ядерной оболочкой, в бактериальной клетке нет. Поэтому наследственный материал бактерий располагается прямо в цитоплазме и представлен одной хромосомой — кольцевой молекулой ДНК.
Бактериям, как и любым другим живым организмам, необходимо получение энергии. Большинство бактерий питается готовыми органическими веществами.
И лишь некоторые из них, например синезелёные, или цианобактерии, способны создавать органические вещества из неорганических.
По способу питания бактерии, питающиеся готовыми органическими веществами, делят на две группы: сапротрофы, которые получают органические вещества из отмерших организмов или выделений живых организмов, и паразиты, которые питаются органическими веществами живых организмов.
Бактерии способны быстро делиться. Размножаются они делением одной клетки на две.
В неблагоприятных условиях у некоторых видов бактерий (при недостатке пищи, влаги, резких изменениях температуры) образуются особые клетки — споры.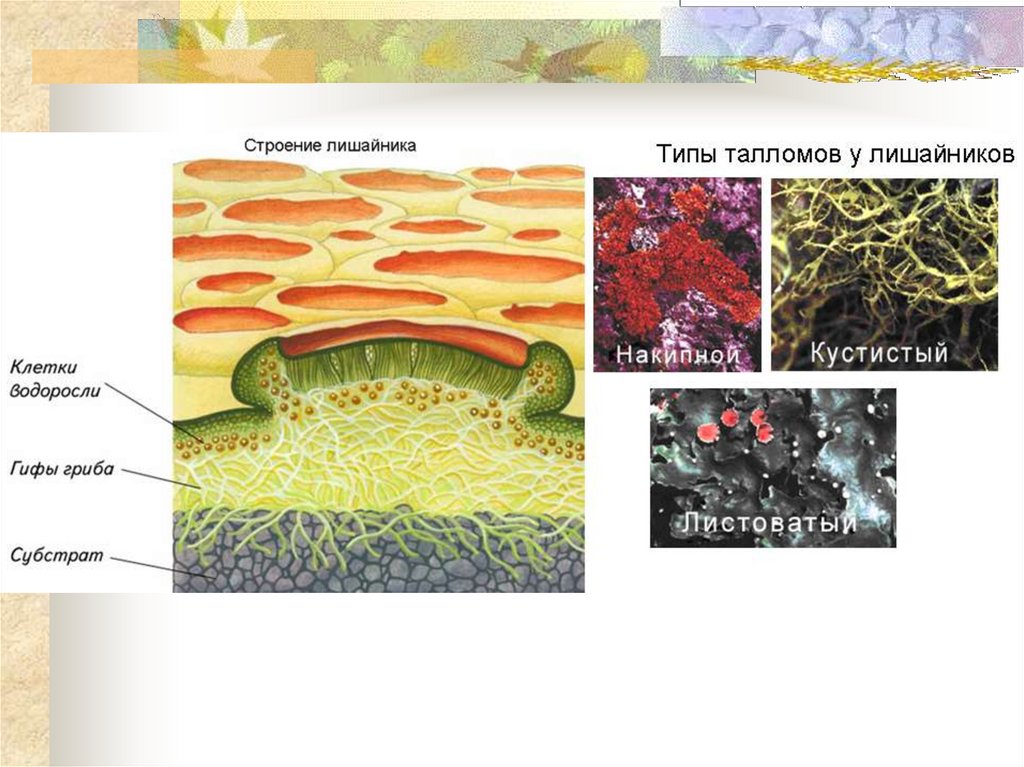
В благоприятных условиях спора прорастает и становится жизнедеятельной бактерией.
Царство Животные подразделяется на два подцарства:
Одноклеточные, или Простейшие, тело которых состоит из одной клетки, и Многоклеточные, тело которых состоит из множества клеток.
В подцарстве выделяют следующие типы: Саркожгутиконосцы, Инфузории, Эвгленозои и Споровики.
Например, к саркожгутиконосцам относится амёба обыкновенная; к инфузориям — инфузория-туфелька; к эвгленозоям — эвглена зелёная; к споровикам — грегарина.
Как мы уже сказали, тело простейшего организма состоит из одной клетки. Эта клетка представляет собой целый организм, который способен самостоятельно существовать, то есть передвигаться, дышать, питаться, размножаться.
Снаружи клетка покрыта цитоплазматической мембраной. Основные компоненты клетки одноклеточных — это ядро и цитоплазма.
В цитоплазме содержатся все органоиды, характерные для животной клетки, — это митохондрии, рибосомы, лизосомы, комплекс Гольджи, эндоплазматическая сеть.
Кроме этого, у простейших имеются органоиды специального назначения. Функцию пищеварения, например, выполняет пищеварительная вакуоль, а функцию выделения — сократительные вакуоли.
Одноклеточные с постоянной формой тела имеют клеточный рот, клеточную глотку, а также орган выделения — порошицу — отверстие, через которое выводятся непереваренные остатки пищи.
Органоидами движения у простейших могут быть ложноножки, жгутики, реснички.
Большинство простейших имеет гетеротрофный тип питания — они используют готовые органические вещества. Питаются другими простейшими, бактериями и водорослями.
Но некоторые способны к фотосинтезу. Так, эвглена зелёная на свету способна к автотрофному способу питания, а в темноте — к гетеротрофному.
Простейшие, не имеющие постоянной формы тела, способны захватывать пищу всей его поверхностью с помощью фагоцитоза и пиноцитоза. Фагоцитоз — это захват твёрдых частиц пищи, а пиноцитоз — захват капелек жидкости с помощью ложноножек.
Дыхание одноклеточных простейших осуществляется всей поверхностью тела. В организм простейшего постоянно поступает вода, содержащая кислород, и она удаляется вместе с углекислым газом через сократительную вакуоль.
Размножение простейших происходит преимущественно бесполым способом. Сначала надвое делится ядро, затем делится цитоплазма. Простейшие с наступлением неблагоприятных условий способны образовывать цисту. В цистах процессы жизнедеятельности клетки практически прекращаются, но они могут оставаться жизнеспособными в течение десятков лет до наступления благоприятных условий.
Хотя грибы и выделяют в отдельное царство, они имеют схожие признаки с растениями и животными.
Всё многообразие грибов делится на группы. Наиболее известные — это шляпочные грибы. У них есть шляпка и ножка. А всё вместе — плодовое тело гриба.
Шляпка и ножка состоят из плотно прилегающих друг к другу нитей грибницы. В ножке все нити одинаковы, а в шляпке они образуют два слоя — верхний, покрытый кожицей, окрашенной в разные цвета, и нижний.
По ножке поступают питательные вещества к шляпке, на которой по мере развития созревают споры. С помощью спор грибы размножаются.
В спорах заключена вся наследственная информация о грибе. Они служат для размножения.
Листьев, стебля, корневой системы у грибов нет. Нет у них и хлорофилла. Грибы, в отличие от растений, питаются уже готовыми органическими веществами.
Шляпочные грибы разделяют на пластинчатые, у которых нижний слой плодовых тел образован пластинками (пластинки — это спорообразующий слой), и трубчатые грибы, у которых нижний слой плодовых тел образован многочисленными трубочками. В трубочках, как в кладовых, хранятся миллиарды спор.
Созревшие споры высыпаются в богатую перегноем почву и прорастают. Из них развиваются тонкие ветвящиеся белые нити грибницы. Грибница внешне похожа на корень и состоит из ветвящихся нитей.
Эти нити состоят из клеток с цитоплазмой и ядром.
Многочисленные нити грибницы пронизывают лесную подстилку и верхний слой почвы, разлагают растительные остатки и получают необходимые питательные вещества. Такие грибы называются сапрофиты.
Такие грибы называются сапрофиты.
Между определёнными видами деревьев и грибов устанавливается тесная связь, полезная как одному, так и другому организму. Такая связь называется симбиозом. Гриб снабжает дерево водой и минеральными солями. А дерево даёт грибу органические вещества, например углеводы, которые гриб из-за отсутствия хлорофилла вырабатывать не может.
Таким образом, нити грибницы плотно оплетают корень дерева и даже проникают внутрь его, образуя грибокорень, или микоризу.
Помимо шляпочных грибов существуют и плесневые грибы, дрожжи, грибы-паразиты.
Плесени — это микроскопические грибы. По способу питания они сапрофиты, то есть питаются уже готовыми органическими веществами.
Пожалуй, наиболее часто встречается плесневый гриб мукор, или как его ещё называют белая головчатая плесень. Этот гриб вы наверняка видели.
Он появляется на хлебе, который уже полежал в тёплом влажном месте.
В процессе развития мукора на нитях грибницы появляются головки. Их называют спорангиями, так как они наполнены миллионами спор. Головки созревают, лопаются, и споры вылетают. А попав в благоприятные условия, споры прорастают в грибницу.
Их называют спорангиями, так как они наполнены миллионами спор. Головки созревают, лопаются, и споры вылетают. А попав в благоприятные условия, споры прорастают в грибницу.
Плесень — гриб пеницилл. На него было обращено внимание биологов.
Если посмотреть на гриб под микроскопом, то можно увидеть на концах нитей грибницы целые кисти спор. Споры у пеницилла расположены не в головках, как у мукора, а на концах нитей грибницы в мелких кисточках.
Гриб пеницилл выделяет особое вещество — пенициллин. Пенициллин — это антимикробный препарат, который приводит к гибели бактерий, используется в медицине.
Дрожжевые грибы — это бесцветные одноклеточные организмы, в каждой клетке которых имеется ядро. Значит, дрожжи не бактерии, а грибы, у которых только нет грибницы. Многие дрожжевые грибы размножаются почкованием.
Среди грибов немало паразитов. Они вызывают различные болезни растений, животных и человека. Вспомним некоторых из них. Картофельный гриб, или фитофтора, поражает листья картофеля и проникает в клубни.
Опасный паразит злаков — гриб головня, который вызывает пыльную головню. Разные виды этого гриба могут поражать хлебные злаки: пшеницу, ячмень, кукурузу и др. Чёрная пыль на колосьях — это споры гриба.
Споры грибов-трутовиков проникают в дерево через трещины, раны, появляющиеся в коре при поломке ветвей. Поселяясь на ветвях и стволах деревьев и питаясь содержимым их клеток, многие трутовики губят деревья.
Долгое время лишайники относили к царству Растения, сейчас их относят к царству Грибы.
Лишайники — это группа симбиотических организмов. Тело лишайника — слоевище — состоит из грибов и микроскопических водорослей или цианобактерий.
Гифы гриба плотно оплетают водоросли. А в некоторых местах гифы прорастают внутрь клетки водоросли.
И гриб, и водоросль получают от совместного существования пользу.
Гриб от водоросли получает питательные вещества, в основном глюкозу, производимые водорослью в результате фотосинтеза, а также азотсодержащие вещества. А водоросль получает от гриба воду и минеральные вещества. Также гриб создаёт водоросли благоприятный микроклимат. Он защищает её от высыхания, закрывая от ультрафиолетового излучения солнца.
А водоросль получает от гриба воду и минеральные вещества. Также гриб создаёт водоросли благоприятный микроклимат. Он защищает её от высыхания, закрывая от ультрафиолетового излучения солнца.
По внешнему виду различают лишайники накипные листоватые и кустистые.
Лишайники, как и любые другие живые организмы, размножаются. Размножение их происходит главным образом кусочками слоевища, а также особыми группами клеток гриба и водоросли. Группы клеток образуются внутри тела лишайника, которое затем под давлением разрывается, и клетки разносятся ветром и дождевыми потоками.
Грибковое разнообразие лишайников: от экстремальной толерантности к взаимодействию с водорослями
1. Хоксворт Д.Л., Онеггер Р. Таллом лишайника: симбиотический фенотип грибов, специализирующихся на питании, и его ответ на продуценты желчи. В: Williams MAJ, редактор. Специальный том Ассоциации систематики. Кларендон Пресс; Оксфорд, Великобритания: 1994. стр. 77–98. [Google Scholar]
2. Cardinale M.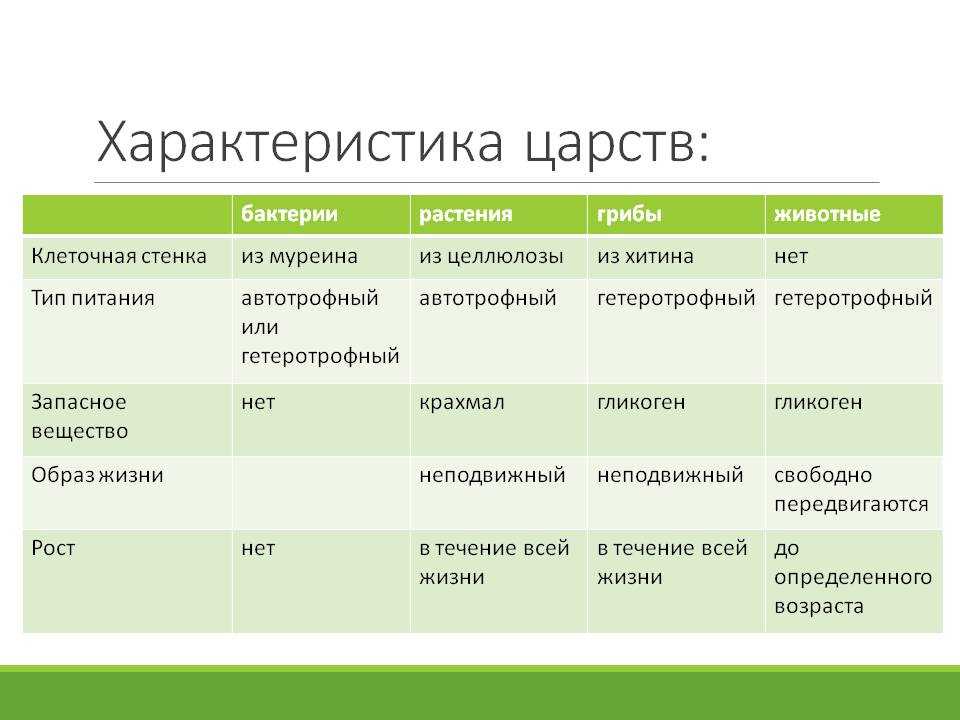 , Vieira de Castro J., Jr., Müller H., Berg G., Grube M. In situ анализ бактериального сообщества, связанного с лишайником северного оленя Cladonia arbuscula выявляет преобладание Alphaproteobacteria. ФЭМС микробиол. Экол. 2008; 66: 63–71. doi: 10.1111/j.1574-6941.2008.00546.x. [PubMed] [CrossRef] [Google Scholar]
, Vieira de Castro J., Jr., Müller H., Berg G., Grube M. In situ анализ бактериального сообщества, связанного с лишайником северного оленя Cladonia arbuscula выявляет преобладание Alphaproteobacteria. ФЭМС микробиол. Экол. 2008; 66: 63–71. doi: 10.1111/j.1574-6941.2008.00546.x. [PubMed] [CrossRef] [Google Scholar]
3. Grube M., Cardinale M., Vieira De Castro J., Jr., Müller H., Berg G. Видоспецифичное структурное и функциональное разнообразие бактериальных сообществ лишайников. симбиоз. ISME J. 2009; 3: 1105–1115. doi: 10.1038/ismej.2009.63. [PubMed] [CrossRef] [Google Scholar]
4. Grube M., Cernava T., Soh J., Fuchs S., Aschenbrenner I., Lassek C., Wegner U., Becher D., Riedel K., Sensen C.W. и соавт. Изучение функциональных контекстов симбиотической устойчивости внутри лишайниковых бактерий с помощью сравнительной омики. ИСМЕ Дж. 2015;9: 412–424. doi: 10.1038/ismej.2014.138. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
5. Бейтс С.Т., Кропси Г. В., Капорасо Дж.Г., Найт Р., Фиерер Н. Бактериальные сообщества, связанные с симбиозом лишайников. заявл. Окружающая среда. микробиол. 2011;77:1309–1314. doi: 10.1128/AEM.02257-10. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
В., Капорасо Дж.Г., Найт Р., Фиерер Н. Бактериальные сообщества, связанные с симбиозом лишайников. заявл. Окружающая среда. микробиол. 2011;77:1309–1314. doi: 10.1128/AEM.02257-10. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
6. Бейтс С.Т., Берг-Лайонс Д., Лаубер К.Л., Уолтерс В.А., Найт Р., Фиерер Н. Предварительное исследование эукариот, связанных с лишайниками, с использованием пиросеквенирования. . лихенолог. 2012;44:137–146. дои: 10.1017/S0024282911000648. [CrossRef] [Google Scholar]
7. Ашенбреннер И., Чернава Т., Берг Г., Грубе М. Понимание многовидовых микробных симбиозов. Фронт. микробиол. 2016;7:180. doi: 10.3389/fmicb.2016.00180. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
8. Моя П., Молинс А.А., Анез-Альберола Ф.М., Муджиа Л., Баррено Э. Неожиданное связанное разнообразие микроводорослей в лишайнике Ramalina farinacea обнаруживаются пиросеквенированием. ПЛОС ОДИН. 2017;12:e0175091. doi: 10.1371/journal.pone. 0175091. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
0175091. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
9. Molins A., Moya P., García-Breijo F.J., Reig-Armiñana J., Barreno E. Мультиинструментальный подход к оценке разнообразия микроводорослей у лишайников: изоляция, секвенирование по Сэнгеру, HTS и ультраструктурные корреляции. лихенолог. 2018;50:123–138. doi: 10.1017/S0024282917000664. [CrossRef] [Google Scholar]
10. Лоури Дж. Д., Дидерих П. Лишайниковые грибы: взаимодействие, эволюция и биоразнообразие. Бриолог. 2003; 106: 80–120. дои: 10.1639/0007-2745(2003)106[0080:LFIEAB]2.0.CO;2. [CrossRef] [Google Scholar]
11. Арнольд А.Е., Мядликовска Дж., Хиггинс К.Л., Сарвате С.Д., Гуггер П., Уэй А., Хофстеттер В., Кауфф Ф., Луцони Ф. Филогенетическая оценка сетей трофического перехода для аскомицетов: являются ли лишайники колыбелью диверсификации симбиотрофных грибов? Сист. биол. 2009; 58: 283–297. doi: 10.1093/sysbio/syp001. [PubMed] [CrossRef] [Google Scholar]
12. Muggia L., Grube M. Грибковый состав талломов лишайников, оцененный по одноцепочечному конформационному полиморфизму.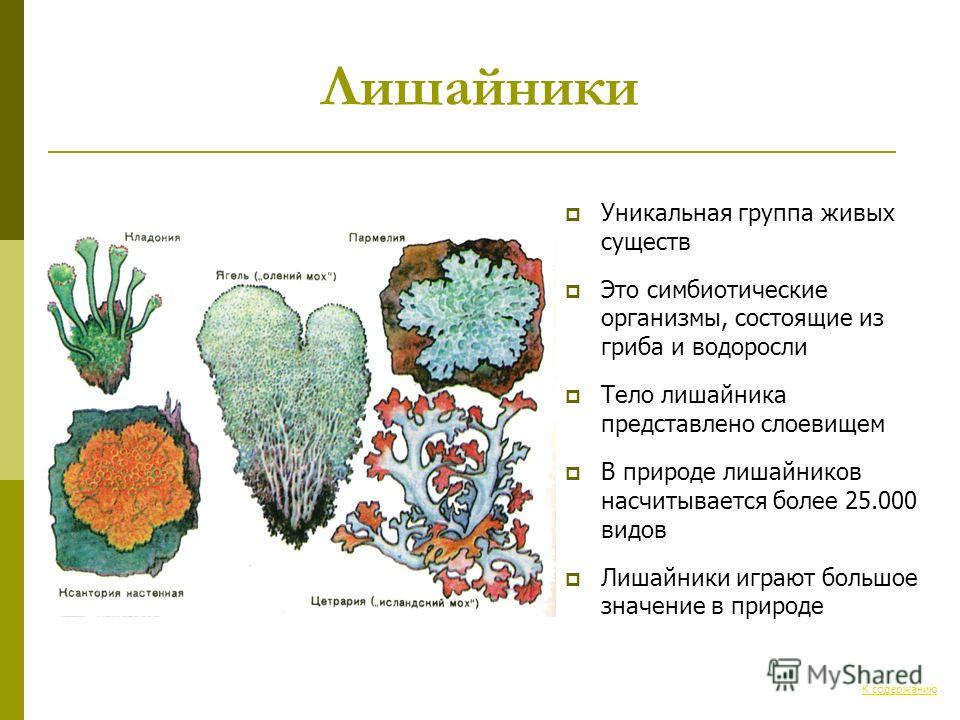 лихенолог. 2010; 42:1–13. дои: 10.1017/S0024282909990752. [CrossRef] [Google Scholar]
лихенолог. 2010; 42:1–13. дои: 10.1017/S0024282909990752. [CrossRef] [Google Scholar]
13. У’Рен Дж. М., Лутцони Ф., Мядликовска Дж., Арнольд А. Е. Анализ сообществ показывает близкое сходство между эндофитными и эндолихенными грибами во мхах и лишайниках. микроб. Экол. 2010;60:340–353. doi: 10.1007/s00248-010-9698-2. [PubMed] [CrossRef] [Google Scholar]
14. Fleischhacker A., Grube M., Kopun T., Muggia L. Анализ сообщества выявил большое разнообразие лихенофильных грибов в альпийских местообитаниях. микроб. Экол. 2015;70:348–360. doi: 10.1007/s00248-015-0579-6. [PubMed] [CrossRef] [Google Scholar]
15. Muggia L., Fleischhacker A., Kopun T., Grube M. Экстремотолерантные грибы из альпийских скальных лишайников и их филогенетические взаимоотношения. Грибные водолазы. 2016;76:119–142. doi: 10.1007/s13225-015-0343-8. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
16. Spribille T., Tuovinen V., Resl P., Vanderpool D., Wolinski H., Aime M.C. , Schneider K., Stabentheiner E., Тум-Хеллер М., Тор Г. и др. Базидиомицетовые дрожжи в коре аскомицетов макролишайников. Наука. 2016; 353: 488–492. doi: 10.1126/science.aaf8287. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
, Schneider K., Stabentheiner E., Тум-Хеллер М., Тор Г. и др. Базидиомицетовые дрожжи в коре аскомицетов макролишайников. Наука. 2016; 353: 488–492. doi: 10.1126/science.aaf8287. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
17. Spribille T. Относительный симбионтный вклад и лишайниковый симбиотический результат. Курс. мнение биол. растений 2018;44:57–63. doi: 10.1016/j.pbi.2018.02.007. [PubMed] [CrossRef] [Google Scholar]
18. Tulasne L.-R. Mémoire pour servir à l’histoire organographique et physiologique des lichens. Анна. науч. Нац. 1852; 17: 153–249. [Google Scholar]
19. Lawrey J.D., Diederich P. Lichenicolous Fungi — всемирный контрольный список, включая доступные изолированные культуры и последовательности. [(по состоянию на 1 марта 2018 г.)]; 2018 г. Доступно в Интернете: http://www.lichenicolous.net
20. Флейшхакер А. Магистерская диссертация. Университет Граца; Грац, Австрия: декабрь 2011 г. Лихенолезные грибы, вторгающиеся в Xanthoria parietina.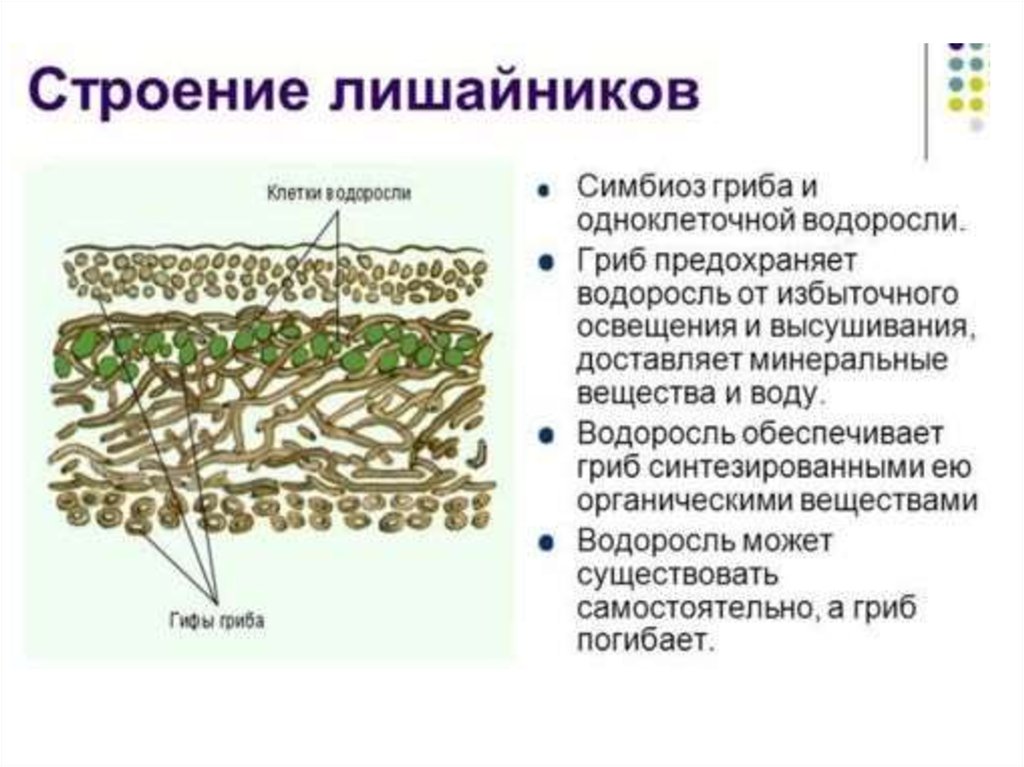 [Google Scholar]
[Google Scholar]
21. Цуриков А., Этайо Ж. Capronia suijae (Herpotrichiellaceae, Eurotiomycetes), новый гриб на Xanthoria parietina из Беларуси, с определителем лихенофильных видов, произрастающих на Xanthoria s. ул. лихенолог. 2017; 49:1–12. doi: 10.1017/S0024282916000530. [Перекрестная ссылка] [Академия Google]
22. Hahn C. Pilze an Xanthoria — Weltschlüssel. [(по состоянию на 8 января 2018 г.)]; 2017 Доступно в Интернете: http://forum.pilze-bayern.de/index.php/topic,1556.0.html
. В: Фриш А., Ланге У., Стайгер Б., редакторы. Lichenologische Nebenstunden. Вклад в таксономию и экологию лишайников в честь Клауса Калба. Лихенологическая библиотека. Том 96. Дж. Крамер в der Gebrüder Borntraeger Verlagsbuchhandlung; Берлин, Германия: Штутгарт, Германия: 2007. стр. 103–128. [Академия Google]
24. Grube M., Lücking R. Тонкие структуры листовидных лишайников и их лишайниковых грибов, изученные методом эпифлуоресценции. Симбиоз. 2002; 32: 229–246. [Google Scholar]
2002; 32: 229–246. [Google Scholar]
25. Грубе М., Хафельнер Дж. Исследования лишайниковых грибов рода Didymella (Ascomycetes, Dothideales) Nova Hedwig. 1990; 51: 283–360. [Google Scholar]
26. De los Ríos A., Ascaso C., Grube M. Изучение механизмов заражения лишайниковыми грибами с помощью различных микроскопических методов. библ. Лихенол. 2002; 82: 153–161. [Академия Google]
27. Арутюнян С., Муджиа Л., Грубе М. Черные грибы в лишайниках из сезонно засушливых местообитаний. Стад. Микол. 2008; 61: 83–90. doi: 10.3114/sim.2008.61.08. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
28. Selbmann L., Grube M., Onofri S., Isola D., Zucconi L. Антарктические эпилитические лишайники как ниши для черных меристематических грибов. Биология. 2013;2:784–797. doi: 10.3390/biology2020784. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
29. Гостинчар С., Грубе М., де Хоог С., Залар П., Гунд-Симерман Н. Экстремотолерантность грибов: Эволюция на грани.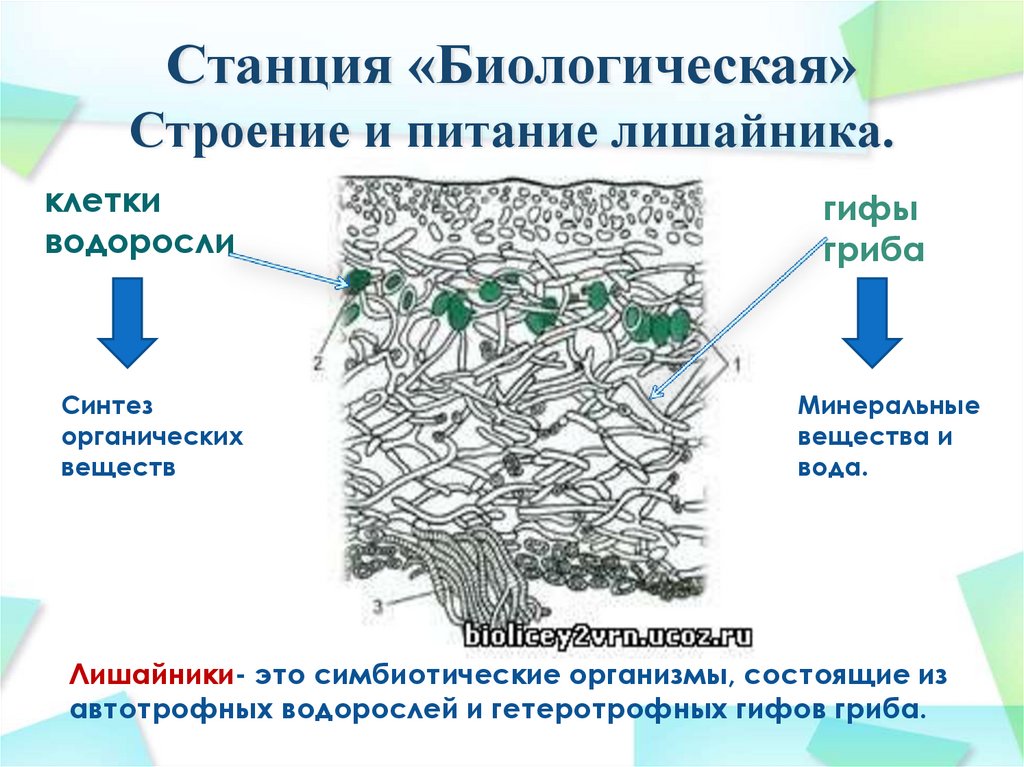 ФЭМС микробиол. Экол. 2010;71:2–11. дои: 10.1111/j.1574-6941.2009.00794.х. [PubMed] [CrossRef] [Google Scholar]
ФЭМС микробиол. Экол. 2010;71:2–11. дои: 10.1111/j.1574-6941.2009.00794.х. [PubMed] [CrossRef] [Google Scholar]
30. Зельбманн Л., Зуккони Л., Изола Д., Онофри С. Каменные черные грибы: превосходство в экстремальных условиях, от Антарктики до космоса. Курс. Жене. 2015;61:335–345. doi: 10.1007/s00294-014-0457-7. [PubMed] [CrossRef] [Google Scholar]
31. Горбушина А.А. Жизнь на скалах. Окружающая среда. микробиол. 2007; 9: 1613–1631. doi: 10.1111/j.1462-2920.2007.01301.x. [PubMed] [CrossRef] [Google Scholar]
32. Горбушина А.А., Бротон В.Дж. Микробиология границы раздела атмосфера-горная порода (как биологические взаимодействия и физические стрессы регулируют сложную микробную систему. Ann. Rev. Microbiol. 2009).;63:431–450. doi: 10.1146/annurev.micro.091208.073349. [PubMed] [CrossRef] [Google Scholar]
33. Гостинчар С., Муджиа Л., Грубе М. Полиэкстремотолерантные черные грибы: олиготрофизм, адаптивный потенциал и связь с симбиозами лишайников. Фронт. микробиол.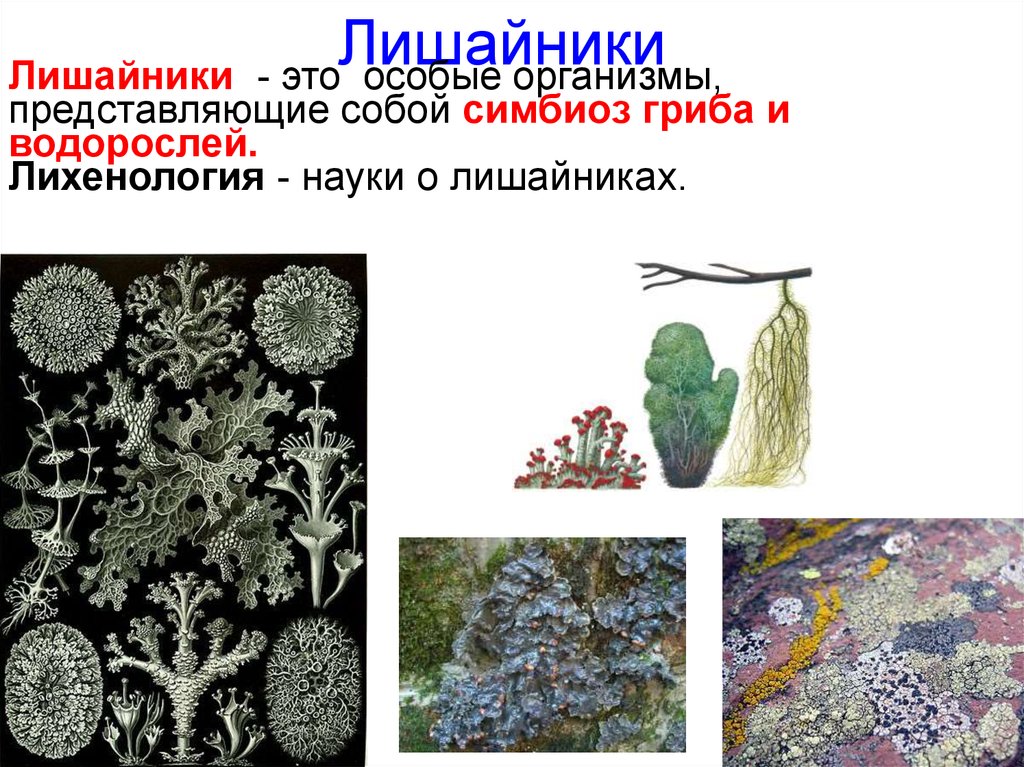 2012;3:390. doi: 10.3389/fmicb.2012.00390. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
2012;3:390. doi: 10.3389/fmicb.2012.00390. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
34. Zhang T., Wie X.L., Zhang Y.Q., Liu H.Y., Yu L.Y. Разнообразие и распространение грибов, связанных с лишайниками, в регионе Ню-Олесунн (Шпицберген, высокая Арктика) по данным 454 пиросеквенирования. науч. Отчет 2015; 5:14850. doi: 10.1038/srep14850. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
35. Fernández-Mendoza F., Fleischhacker A., Kopun T., Grube M., Muggia L. Метабаркодирование ITS1 подчеркивает низкую специфичность микобиомов лишайников в локальном масштабе. Мол. Экол. 2017;26:4811–4830. doi: 10.1111/mec.14244. [PubMed] [CrossRef] [Google Scholar]
36. Люкинг Р., Дал-Форно М., Сикаруди М., Гиллевет П.М., Бунгартц Ф., Монкада Б., Янез-Аябака А., Чавес Дж.Л., Кока Л.Ф. , Лоури Дж. Д. Один макролишайник включает сотни неизвестных видов. проц. Натл. акад. науч. США. 2014;111:11091–11096. doi: 10.1073/pnas.1403517111. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
37. Адамс Р.И., Аменд А.С., Тейлор Дж.В., Брунс Т.Д. Уникальный сигнал искажает восприятие видового богатства и состава при высокопроизводительном секвенировании микробных сообществ. : тематическое исследование грибков в домашней пыли. микроб. Экол. 2013;66:735–741. doi: 10.1007/s00248-013-0266-4. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
Адамс Р.И., Аменд А.С., Тейлор Дж.В., Брунс Т.Д. Уникальный сигнал искажает восприятие видового богатства и состава при высокопроизводительном секвенировании микробных сообществ. : тематическое исследование грибков в домашней пыли. микроб. Экол. 2013;66:735–741. doi: 10.1007/s00248-013-0266-4. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
38. U’Ren J.M., Riddle J.M., Monacell J.T., Carbone I., Miadlikowska J., Arnold AE. Хранение тканей и выбор праймеров влияют на выводы, основанные на пиросеквенировании. разнообразия и состава сообществ эндолихенных и эндофитных грибов. Мол. Экол. Рез. 2014;14:1032–1048. дои: 10.1111/1755-0998.12252. [PubMed] [CrossRef] [Google Scholar]
39. Schoch C.L., Seifert K.A., Huhndorf S., Robert V., Spouge J.L., Levesque C.A., Chen W. Fungal Barcoding Consortium. Ядерная рибосомная внутренняя транскрибируемая спейсерная область (ITS) как универсальный маркер штрих-кода ДНК для грибов. проц. Натл. акад. науч. США. 2012; 109:1–6. doi: 10. 1073/pnas.1117018109. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
1073/pnas.1117018109. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
40. U’Ren J.M., Lutzoni F., Miadlikowska J., Laetsch A.D., Arnold E. Хозяин и географическая структура эндофитных и эндолихенных грибов на континентальном шкала. Являюсь. Дж. Бот. 2012;99: 898–914. doi: 10.3732/ajb.1100459. [PubMed] [CrossRef] [Google Scholar]
41. Марк К., Корнехо С., Келлер С., Флюк Д., Шайдеггер С. Штрихкодирование грибов, образующих лишайники, с использованием пиросеквенирования 454 оспаривается артефактной и биологической изменчивостью последовательностей. Геном. 2016; 59: 685–704. doi: 10.1139/gen-2015-0189. [PubMed] [CrossRef] [Google Scholar]
42. Баззикалупо А.Л., Балинт М., Шмитт И. Сравнение рДНК ITS1 и ITS2 при секвенировании 454 гиперразнообразных грибковых сообществ. Грибковая экол. 2013; 6: 102–109. doi: 10.1016/j.funeco.2012.09.003. [CrossRef] [Google Scholar]
43. Döbbeler P. Moosbewohnende Ascomyceten I. Die Pyrenocarpen, den Gametophyten Besiedelnden Arten. Том 14. Mitt Botanische Staatssammlung; Мюнхен, Германия: 1978. стр. 1–360. [Google Scholar]
Том 14. Mitt Botanische Staatssammlung; Мюнхен, Германия: 1978. стр. 1–360. [Google Scholar]
44. Доббелер П. Биоразнообразие мохообразных аскомицетов. Биодайверы. Консерв. 1977; 6: 721–738. doi: 10.1023/A:1018370304090. [CrossRef] [Google Scholar]
45. Wang Y., Zheng Y., Wang X., Wie X., Wie J. Грибковое сообщество, связанное с лишайниками, в Hypogymnia hypotrypa (Parmeliaceae, Ascomycota) зависит от географического распространения и высоты над уровнем моря. Фронт. микробиол. 2016;7:1231. doi: 10.3389/fmicb.2016.01231. [PMC бесплатная статья] [PubMed] [CrossRef] [Google Scholar]
46. Persoh D., Rambold G. Лишайниково-ассоциированные грибы Letharietum vulpinae. Микол. прог. 2012; 11:1–8. doi: 10.1007/s11557-011-0786-6. [CrossRef] [Google Scholar]
47. Хоксворт Д.Л. Лихенофильные гифомицеты. Бык. бр. Мус. (Nat. Hist.) Bot. сер. 1979;6:183–300. [Google Scholar]
48. Хоксворт Д.Л. Лихенофильные колеомицеты. Бык. бр. Мус. (Nat. Hist.) Bot. сер. 1981; 9: 1–98. [Google Scholar]
сер. 1981; 9: 1–98. [Google Scholar]
49. Lawrey J.D., Diederich P., Nelsen M.P., Sikaroodi M., Gillevet P.M., Brand A.M., van den Boom P. Облигатно лихенический род Lichenoconium представляет собой новую линию дотидеомицетов. Грибковая биол. 2011; 115:176–187. doi: 10.1016/j.funbio.2010.12.002. [PubMed] [CrossRef] [Академия Google]
50. Lawrey J.D., Diederich P., Nelsen M.P., Freebury C., Van den Broeck D., Sikaroodi M., Ertz D. Филогенетическое размещение видов lichenicolous Phoma в Phaeosphaeriaceae (Pleosporales, Dothideomycets) Fungal Divers. 2012;55:195–213. doi: 10.1007/s13225-012-0166-9. [CrossRef] [Google Scholar]
51. Ertz D., Heuchert B., Braun U., Freebury CE, Common R.S., Diederich P. Вклад в филогению и таксономию рода Taeniolella с акцентом на таксоны лихениколезных. Грибковая биол. 2016; 120:1416–1447. doi: 10.1016/j.funbio.2016.05.008. [PubMed] [CrossRef] [Академия Google]
52. Muggia L., Kopun T., Ertz D. Филогенетическое размещение лихеноподобного анаморфного рода Lichenodiplis и его связь с Muellerella -подобными телеоморфами. Грибковая биол. 2015;119:1115–1128. doi: 10.1016/j.funbio.2015.08.011. [PubMed] [CrossRef] [Google Scholar]
Филогенетическое размещение лихеноподобного анаморфного рода Lichenodiplis и его связь с Muellerella -подобными телеоморфами. Грибковая биол. 2015;119:1115–1128. doi: 10.1016/j.funbio.2015.08.011. [PubMed] [CrossRef] [Google Scholar]
53. Gueidan C., Villaseñor C.R., de Hoog G.S., Gorbushina A.A., Untereiner W.A., Lutzoni F. Обитающий в скалах предок мутуалистических и богатых патогенами грибковых линий. Стад. Микол. 2008; 61: 111–119.. doi: 10.3114/sim.2008.61.11. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
54. Gueidan C., Ruibal C., de Hoog GS, Schneider H. Грибы, населяющие скалы, возникли в периоды сухого климата в позднем и среднем девоне. Триас. Грибковая биол. 2011; 115:987–996. doi: 10.1016/j.funbio.2011.04.002. [PubMed] [CrossRef] [Google Scholar]
55. Kirk P.M., Cannon P.F., Minter D.W., Stalpers J.A. Словарь грибов Эйнсворт и Бисби. 10-е изд. КАБ Интернэшнл; Уоллингфорд, Великобритания: 2008 г. [Google Scholar] 9.0003
56. Schoch C.L., Crous P.W., Groenewald J.Z., Boehm E.W.A., Burgess T.I., De Gruyter J., De Hoog G.S., Dixon L.J., Grube M., Gueidan C., et al. Общеклассовая филогенетическая оценка дотидеомицетов. Стад. Микол. 2009; 64:1–15. doi: 10.3114/sim.2009.64.01. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
Schoch C.L., Crous P.W., Groenewald J.Z., Boehm E.W.A., Burgess T.I., De Gruyter J., De Hoog G.S., Dixon L.J., Grube M., Gueidan C., et al. Общеклассовая филогенетическая оценка дотидеомицетов. Стад. Микол. 2009; 64:1–15. doi: 10.3114/sim.2009.64.01. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
57. Onofri S., Selbmann L., Zucconi L., de Hoog GS, de los Rios A., Ruisi S., Grube M. Грибковые ассоциации на холодном краю жизни. В: Секбах Дж., редактор. Водоросли и цианобактерии в экстремальных условиях. Спрингер; Доротех, Нидерланды: 2007. стр. 735–757. [Академия Google]
58. Зельбманн Л., де Хоог Г.С., Маццаглия А., Фридманн Э.И., Онофри С. Грибы на краю жизни: криптоэндолитические черные грибы из антарктической пустыни. Стад. Микол. 2005; 51:1–32. [Google Scholar]
59. Egidi E., de Hoog S., Isola D., Onofri S., Quaedvlieg W., de Vries M., Verkley G.J.M., Stielow J.B., Zucconi L., Selbmann L. Филогения и таксономия меристематических черных грибов, населяющих скалы, у Dothideomycetes на основе многолокусной филогении. Грибные водолазы. 2014;65:127–165. doi: 10.1007/s13225-013-0277-y. [Перекрестная ссылка] [Академия Google]
Грибные водолазы. 2014;65:127–165. doi: 10.1007/s13225-013-0277-y. [Перекрестная ссылка] [Академия Google]
60. Selbmann L., Isola D., Egidi E., Zucconi L., Gueidan C., de Hoog G.S., Onofri S. Горные вершины как резервуары для новых каменных грибов: Saxomyces gen. ноябрь и четыре новых вида из Альп. Грибные водолазы. 2014;65:167–182. doi: 10.1007/s13225-013-0234-9. [CrossRef] [Google Scholar]
61. Muggia L., Kocourkova J., Knudsen K. Выделение комплекса из видов Lichenothelia из каменных сообществ пустыни. Микология. 2015;107:1233–1253. дои: 10.3852/15-021. [PubMed] [CrossRef] [Академия Google]
62. Гунде-Цимерман Н., Залар П., де Хоог С., Племениташ А. Гиперсоленые воды в солончаках — естественные экологические ниши для галофильных черных дрожжей. ФЭМС микробиол. Экол. 2000; 32: 235–240. doi: 10.1111/j.1574-6941.2000.tb00716.x. [PubMed] [CrossRef] [Google Scholar]
63. Muggia L., Hafellner J., Wirtz N., Hawksworth D.L., Grube M. Стерильные микрофиламентные лихенизированные грибы Cystocoleus ebeneus и Racodium rupestre являются родственниками растений. патогены и клинически значимые дотивидные грибы. Микол. Рез. 2008; 112: 50–56. doi: 10.1016/j.mycres.2007.08.025. [PubMed] [CrossRef] [Академия Google]
патогены и клинически значимые дотивидные грибы. Микол. Рез. 2008; 112: 50–56. doi: 10.1016/j.mycres.2007.08.025. [PubMed] [CrossRef] [Академия Google]
64. Texteira M.M., Moreno L.F., Stielow B.J., Muszewska A., Hainaut M., Gonzaga L., Abouelleil A., Patané J.S.L., Priest M., Souza R., et al. Изучение геномного разнообразия черных дрожжей и их родственников (Chaetothyriales, Ascomycota) Stud. Микол. 2017; 86:1–28. doi: 10.1016/j.simyco.2017.01.001. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
65. Muggia L., Kopun T., Grube M. Влияние питательной среды на разнообразие культивируемых грибов из лишайников. Молекулы. 2017;22:824. дои: 10.3390/молекулы 22050824. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
, Дай Д.К. и др. Семейства дотидеомицетов. Грибные водолазы. 2013; 63:1–313. doi: 10.1007/s13225-013-0263-4. [CrossRef] [Google Scholar]
67. Gueidan C., Aptroot A., da Silva Cáceres M.E., Badali H., Stenroos S. Переоценка порядков и семейств внутри подкласса Chaetothyriomycetidae (Eurotiomycetes, Ascomycota) Mycol. прог. 2014;13:1027–1039. doi: 10.1007/s11557-014-0990-2. [CrossRef] [Google Scholar]
прог. 2014;13:1027–1039. doi: 10.1007/s11557-014-0990-2. [CrossRef] [Google Scholar]
68. Chen K.H., Miadlikowska J., Molnár K., Arnold A.E., U’Ren J.M., Gaya E., Gueidan C., Lutzoni F. Филогенетический анализ эндофитов евротиомицетов выявил их близкое родство к Chaetothyriales, Eurotiales и новому порядку — Phaeomoniellales. Мол. Филогенет. Эвол. 2015;85:117–130. doi: 10.1016/j.ympev.2015.01.008. [PubMed] [CrossRef] [Google Scholar]
69. Liu J.K., Hyde K.D., Jeewon R., Phillips A.J.L., Maharachchikumbura S.S.N., Ryberg M., Liu Z.Y., Zhao Q. Ранжирование высших таксонов с использованием времени расхождения: тематическое исследование у дотидеомицетов. Грибные водолазы. 2017; 84: 75–99. doi: 10.1007/s13225-017-0385-1. [CrossRef] [Google Scholar]
70. Петрини О., Хейк У., Дрейфус М.М. Анализ грибных сообществ, выделенных из кустистых лишайников. Микология. 1990; 82: 444–451. doi: 10.2307/3760015. [CrossRef] [Google Scholar]
71. Гирланда М., Исокроно Д., Бьянко К. , Луппи-Моска А.М. Два листоватых лишайника как экологические ниши микрогрибов. Микология. 1997; 89: 531–536. дои: 10.2307/3760987. [CrossRef] [Google Scholar]
, Луппи-Моска А.М. Два листоватых лишайника как экологические ниши микрогрибов. Микология. 1997; 89: 531–536. дои: 10.2307/3760987. [CrossRef] [Google Scholar]
72. Crittenden P.D., David J.C., Hawksworth D.L., Campbell F.S. Попытка выделения и успеха в культивировании широкого спектра лихенообразующих и лихенообразных грибов. Новый Фитол. 1995;130:267–297. doi: 10.1111/j.1469-8137.1995.tb03048.x. [CrossRef] [Google Scholar]
73. Йошимура И., Ямамото Ю., Накано Т., Финни Дж. Выделение и культивирование фотобионтов и микобионтов лишайников. В: Краннер И., Беккет Р.П., Варма А.К., редакторы. Протоколы в лихенологии. Культивирование, биохимия, экофизиология и использование в биомониторинге. Спрингер; Берлин/Гейдельберг, Германия: 2002. стр. 3–33. [Google Scholar]
74. Suryanarayanan T.S., Thirunavukkarasu N., Hariharan G., Balaji P. Возникновение необлигатных микрогрибов в слоевищах лишайников. Сидовия. 2005; 57:120. [Академия Google]
75. Stocker-Wörgötter E. Изучение продукции вторичных соединений в микобионтах культивируемых лишайников. В: Краннер И., Беккет Р.П., Варма А.К., редакторы. Протоколы в лихенологии. Культивирование, биохимия, экофизиология и использование в биомониторинге. Спрингер; Берлин/Гейдельберг, Германия: 2002. стр. 296–306. [Google Scholar]
Изучение продукции вторичных соединений в микобионтах культивируемых лишайников. В: Краннер И., Беккет Р.П., Варма А.К., редакторы. Протоколы в лихенологии. Культивирование, биохимия, экофизиология и использование в биомониторинге. Спрингер; Берлин/Гейдельберг, Германия: 2002. стр. 296–306. [Google Scholar]
76. Винаяка К.С., Кришнамурти Ю.Л., Банакар С., Кекуда Т.Р.П. Ассоциация и изменчивость эндофитных грибов среди некоторых макролишайников в центральных Западных Гатах, Южная Индия. Междунар. Дж. Карр. микробиол. заявл. науч. 2016;5:115–124. doi: 10.20546/ijcmas.2016.506.014. [Перекрестная ссылка] [Академия Google]
77. Ruibal C., Gueidan C., Selbmann L., Gorbushina A.A., Crous P.W., Groenewald J.Z., Staley J.T. Филогения обитающих в скалах грибов, родственных Dothideomycetes. Стад. Микол. 2009; 64: 123–133. doi: 10.3114/sim.2009.64.06. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
78. Untereiner WA, Gueidan C., Orr MJ, Diederich P. Филогенетическое положение лишайникового аскомицета Capronia peltigerae . Грибные водолазы. 2011;49:225–233. doi: 10.1007/s13225-011-0097-х. [CrossRef] [Google Scholar]
Грибные водолазы. 2011;49:225–233. doi: 10.1007/s13225-011-0097-х. [CrossRef] [Google Scholar]
79. Diederich P., Ertz D., Lawrey J.D., Sikaroodi M., Untereiner W.A. Молекулярные данные помещают гифомицетные лихениколезные роды Sclerococcum рядом с Dactylospora
6 (Eurotiomyce) и parmeliae
в Cladophialophora (Chaetotyriales) Fungal Divers. 2013;58:61–72. doi: 10.1007/s13225-012-0179-4. [CrossRef] [Google Scholar]
80. Хокинг А.Д., Питт Дж.И. Дихлоран-глицериновая среда для подсчета ксерофильных грибов в продуктах с низким содержанием влаги. заявл. Окружающая среда. микробиол. 1980;39:488–492. [Бесплатная статья PMC] [PubMed] [Google Scholar]
81. Lilly H.L., Barnett V.G. Физиология грибов. 1-е изд. Книжная компания McGraw Hill; Нью-Йорк, штат Нью-Йорк, США: 1951. с. 464. [Google Scholar]
82. Пагано Дж., Левин Дж.Д., Трехо В. Диагностическая среда для дифференциации видов Candida . Антибиот. Анну. 1910; 5: 137–143. [PubMed] [Google Scholar]
[PubMed] [Google Scholar]
83. Kellogg J., Raja H.A. Эндолихеновые грибы: новый источник богатых биологически активными вторичными метаболитами на горизонте. Фитохим. 2017; 16: 271–29.3. doi: 10.1007/s11101-016-9473-1. [CrossRef] [Google Scholar]
84. Calcott M.J., Ackerley D.F., Knight A., Keyzers R.A., Owen J.G. Вторичный метаболизм при симбиозе лишайников. хим. соц. 2018; 47:1730–1760. doi: 10.1039/C7CS00431A. [PubMed] [CrossRef] [Google Scholar]
85. Wang Y., Zheng Z., Liu S., Zhang H., Li E., Guo L., Che Y. Оксэпинохроменоны, фурохроменон и их предполагаемые предшественники из эндолихеновый гриб Coniochaeta sp. Дж. Нат. Произв. 2010;73:920–924. doi: 10.1021/np100071z. [PubMed] [CrossRef] [Google Scholar]
86. Kannangara BTSDP, Rajapaksha RSCG, Paranagama P.A. Природа и биоактивность эндолихеновых грибов у Pseudocyphellaria sp., Parmotrema sp. и Usnea sp. в горном лесу Хакгала на Шри-Ланке. лат. заявл. микробиол. 2009; 48: 203–209. doi: 10.1111/j.1472-765X.2008.02512.x. [PubMed] [CrossRef] [Google Scholar]
2009; 48: 203–209. doi: 10.1111/j.1472-765X.2008.02512.x. [PubMed] [CrossRef] [Google Scholar]
87. He H., Bigelis R., Yang H.Y., Chang LP, Singh M.P. Лихениколины А и В, новые биснафтопироны из неизвестного штамма лихениколезного гриба LL-RB0668. Дж. Антибиот. 2005; 58: 731–736. дои: 10.1038/ja.2005.99. [PubMed] [CrossRef] [Google Scholar]
88. Милланес А.М., Дидерих П., Ведин М. Cyphobasidium gen. nov., новая обитающая в лишайниках линия цистобазидиомицетов (Pucciniomycotina, Basidiomycota, Fungi) Fungal Biol. 2016; 120:1466–1477. doi: 10.1016/j.funbio.2015.12.003. [PubMed] [CrossRef] [Google Scholar]
89. Lopandic K., Molnár O., Prillinger H. Fellomyces mexicanus sp. nov., новый представитель рода дрожжей Fellomyces, выделенный из лишайника Cryptothecia rubrocincta , собранная в Мексике. микробиол. Рез. 2005; 160:1–11. doi: 10.1016/j.micres.2004.09.004. [PubMed] [CrossRef] [Google Scholar]
90. Сингх Н., Упрети Д.К., Гупта В. К., Дай X.Ф., Цзян Ю. Эндолихенные грибы: скрытый резервуар биофармацевтических препаратов следующего поколения. Тенденции биотехнологии. 2017; 35:808–813. doi: 10.1016/j.tibtech.2017.03.003. [PubMed] [CrossRef] [Google Scholar]
К., Дай X.Ф., Цзян Ю. Эндолихенные грибы: скрытый резервуар биофармацевтических препаратов следующего поколения. Тенденции биотехнологии. 2017; 35:808–813. doi: 10.1016/j.tibtech.2017.03.003. [PubMed] [CrossRef] [Google Scholar]
91. Хоксворт Д.Л., Патерсон Р.Р.М., Голос Н. Исследование наличия метаболитов в облигатно лихеновых грибах из тридцати родов. В: Feige GB, Lumbsch HT, редакторы. Фитохимия и хемотаксономия лихенизированных аскомицетов — Festschrift в честь Зигфрида Хунека. Bibliotheca Lichenologica, Дж. Крамер; Берлин, Германия: Штутгарт, Германия: 1993. С. 101–108. [Google Scholar]
92. Виджератне Е.М.К., Башял Б.П., Гунатилака М.К., Арнольд А.Е., Гунатилака А.А.Л. Увеличение химического разнообразия метаболитов грибов: биогенетически родственные гептакетиды эндолихенового гриба Corynespora sp. Дж. Нат. Произв. 2010;73:1156–1159. дои: 10.1021/np
4v. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
93. Падхи С., Тайунг К. Антимикробный потенциал эндолихенных грибов in vitro, выделенных из талломов Parmelia лишайник против некоторых патогенов человека. Университет Бени-Суэф. J. Основное приложение. науч. 2015; 4: 299–306. doi: 10.1016/j.bjbas.2015.11.006. [CrossRef] [Google Scholar]
Университет Бени-Суэф. J. Основное приложение. науч. 2015; 4: 299–306. doi: 10.1016/j.bjbas.2015.11.006. [CrossRef] [Google Scholar]
94. Goers L., Freemont P., Polizzi K.M. Системы и технологии совместного культивирования: вывод синтетической биологии на новый уровень. Дж. Р. Соц. Интерфейс. 2014;11:20140065. doi: 10.1098/rsif.2014.0065. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
95. Muggia L., Kraker S., Gößler T., Grube M. Насильственный симбиоз грибов и водорослей в альгинатных сферах. ФЭМС микробиол. лат. 2018 год: 10.1093/женский/fny115. [PubMed] [CrossRef] [Google Scholar]
96. Hom E.F.Y., Murray A.W. Нишевая инженерия демонстрирует скрытую способность грибково-водорослевого мутуализма. Наука. 2014; 345:94–98. doi: 10.1126/science.1253320. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
97. Горбушина А.А., Бек А., Шульте А. Микроколониальные породы, населяющие грибы и фотобионты лишайников: доказательства мутуалистических взаимодействий. Микол. Рез. 2005; 109:1288–1296. doi: 10.1017/S0953756205003631. [PubMed] [CrossRef] [Академия Google]
Микол. Рез. 2005; 109:1288–1296. doi: 10.1017/S0953756205003631. [PubMed] [CrossRef] [Академия Google]
98. Brunauer G., Blaha J., Hager A., Turk R., Stocker-Wörgötter E., Grube M. Выделенный лихеноподобный гриб образует лихеноидные структуры при совместном культивировании с различными кокковидными водорослями. Симбиоз. 2007; 44: 127–136. [Google Scholar]
99. Аметрано К.Г., Зельбманн Л., Маггиа Л. Стандартизированный подход к совместному культивированию дотидеевых грибов, населяющих скалы, и фотобионтов лишайников in vitro. Симбиоз. 2017;73:35–44. doi: 10.1007/s13199-017-0479-2. [CrossRef] [Google Scholar]
100. Хоксворт Д.Л. Lichenothelia , новый род для группы Microthelia aterrima . лихенолог. 1981; 13: 141–153. doi: 10.1017/S0024282981000182. [CrossRef] [Google Scholar]
101. Muggia L., Gueidan C., Knudsen K., Perlmutter G., Grube M. Связи черных грибов с лишайниками. Микопатология. 2013; 175: 523–535. doi: 10.1007/s11046-012-9598-8. [PubMed] [CrossRef] [Google Scholar]
[PubMed] [CrossRef] [Google Scholar]
Симбиоз: водоросли и грибы движутся из прошлого в будущее
- Паола Бонфанте,
- Туринский университет, Италия;
Попросите студента-биолога описать связь между водорослями и грибами, и он обязательно объяснит вам, как грибковые структуры, называемые гифами, могут окружать клетки водорослей, образуя совершенно новый организм с собственным метаболизмом, называемый лишайником (Honegger, 1991; рис. 1А). ). Благодаря обмену питательными веществами между водорослями и грибами, а также адаптивным механизмам, насчитывающим около 415 миллионов лет, лишайники могут выживать в самых экстремальных условиях.
Грибно-водорослевой симбиоз.
( A ) В лишайниках клетки водорослей (зеленые) окружены гифами грибов (оранжевые), образуя новый организм со своим собственным метаболизмом и свойствами. Грибковые гифы агрегируют, образуя грибковые псевдоткани. Когда водоросль N. oceanica растет в присутствии почвенного грибка M. elongata , водоросли сначала собираются и вступают в контакт с поверхностью гифы ( B ). В конце концов водоросли попадают в гифу, которая меняет цвет на зеленый из-за присутствия внутри нее фотосинтетически активных водорослей ( C ).
Грибковые гифы агрегируют, образуя грибковые псевдоткани. Когда водоросль N. oceanica растет в присутствии почвенного грибка M. elongata , водоросли сначала собираются и вступают в контакт с поверхностью гифы ( B ). В конце концов водоросли попадают в гифу, которая меняет цвет на зеленый из-за присутствия внутри нее фотосинтетически активных водорослей ( C ).
При всех известных взаимодействиях между водорослями и грибами клетки водорослей остаются вне гиф гриба. Теперь в eLife Кристоф Беннинг, Грегори Бонито и их коллеги, в том числе Чжи-Ян Ду из Мичиганского государственного университета в качестве первого автора, сообщают, как при определенных условиях клетки водорослей могут проникать в грибок (Du et al., 2019).). Эксперименты проводились с Nannochloropsis oceanica, — видом водорослей, обитающим в морской и пресной воде, и Mortierella elongata — грибом, обитающим в почве. Эксперименты с изотопными индикаторами выявили обмен питательными веществами, включая углерод и азот, между двумя партнерами. Более того, оба растения оставались физиологически активными в течение двух месяцев совместного культивирования, при этом клетки водорослей продолжали расти, делиться и оставаться фотосинтетически активными в гифах (рис. 1В, С).
Более того, оба растения оставались физиологически активными в течение двух месяцев совместного культивирования, при этом клетки водорослей продолжали расти, делиться и оставаться фотосинтетически активными в гифах (рис. 1В, С).
Симбиозы между микробами и растениями или животными часто используются в качестве примеров коэволюции между царствами: окаменелости предоставляют прямые доказательства существования симбиоза в прошлом, а филогенетический анализ может показать, когда появились симбиотические партнеры (Lutzoni et al., 2018). ; Колер и др., 2015). Например, окаменелости, датированные девонской эрой (около 450 миллионов лет назад), обнаруживают модели колонизации грибами, очень похожие на те, что производятся сегодня Glomeromycotina (Remy et al., 19).94): эти облигатные симбионты (то есть симбионты, которые зависят от хозяина, чтобы выжить) проникают в корневые клетки растений, образуя структуры, называемые арбускулами, в широко распространенной форме симбиоза, которая сейчас называется арбускулярной микоризой.
Однако окаменелости и филогенетика не могут сказать нам, как на самом деле сформировались различные формы симбиоза, которые мы наблюдаем сегодня. Филогенетический анализ подтверждает идею о том, что Glomeromycotina являются представителями Mucoromycota, раннего дивергирующего типа грибов (Spatafora et al., 2016), как и Мортиерелла . Однако неясно, произошли ли грибы, которые сегодня ответственны за арбускулярную микоризу, из сапротрофных грибов (которые питаются мертвым или разлагающимся веществом), поскольку нет никаких доказательств того, что такие грибы эволюционировали, чтобы стать обязательными симбионтами. Напротив, доступны элегантные эксперименты по эволюции азотфиксирующих бактерий: перенос плазмиды sym в штамм Ralstonia ясно демонстрирует, что патоген может эволюционировать в симбиотическую бактерию (Clerissi et al., 2018).
Исследование Du et al. основывается на предыдущей работе, которая показала, что одноклеточная зеленая водоросль Chlamydomonas reinhardtii и дрожжи могут взаимодействовать в определенных физиологических условиях (Hom and Murray, 2014). Однако тот факт, что N. oceanica и M. elongata в основном сохраняют свой фенотип при совместном культивировании, довольно удивителен. Напротив, грибы в лишайниках агрегируют и образуют псевдоткани, в которые внедряются клетки водорослей (а также бактерии) (Cardinale et al., 2008; Рисунок 1A).
Однако тот факт, что N. oceanica и M. elongata в основном сохраняют свой фенотип при совместном культивировании, довольно удивителен. Напротив, грибы в лишайниках агрегируют и образуют псевдоткани, в которые внедряются клетки водорослей (а также бактерии) (Cardinale et al., 2008; Рисунок 1A).
Явление, наблюдаемое Du et al. началось с того, что N. oceanica флокулировали (то есть слипались) вокруг грибка. Очевидный вопрос: какой сигнал вызывает начало процесса флокуляции? В частности, обнаруживает ли N. oceanica и реагирует ли он на молекулы, выделяемые M. elongata ? Поскольку геномы обоих партнеров были секвенированы, различные подходы к транскриптомии и метаболомике должны помочь исследователям ответить на эти вопросы.
Также будет интересно выяснить, играет ли MEP α, ген, кодирующий белок, переносящий аммиак (NH 3 ) в растениях и бактериях, роль в обмене азота между N. oceanica и . М. elongata . Известно, что ген MEP α был перенесен от прокариот к Leotiomyceta, которые являются предками грибов, встречающихся в лишайниках, а также к зеленым растениям (Lutzoni et al. , 2018), и до сих пор обнаруживается у всех таких грибов. и растения. Mortierella и Glomeromycotina оба очень богаты липидами (которые содержат много углерода), но последний не может синтезировать липиды, полагаясь вместо этого на свое растение-хозяин. Тип обмена липидов, наблюдаемый у N. oceanica и M. elongata , может найти применение в биотехнологии. Действительно, Ду и соавт. ранее показали, что использование генетических методов для сверхэкспрессии гена, называемого DGTT5, приводит к увеличению накопления липидов у M. elongata 9.0290, что может увеличить выход биотоплива из микроводорослей из этой системы (Du et al., 2018).
, 2018), и до сих пор обнаруживается у всех таких грибов. и растения. Mortierella и Glomeromycotina оба очень богаты липидами (которые содержат много углерода), но последний не может синтезировать липиды, полагаясь вместо этого на свое растение-хозяин. Тип обмена липидов, наблюдаемый у N. oceanica и M. elongata , может найти применение в биотехнологии. Действительно, Ду и соавт. ранее показали, что использование генетических методов для сверхэкспрессии гена, называемого DGTT5, приводит к увеличению накопления липидов у M. elongata 9.0290, что может увеличить выход биотоплива из микроводорослей из этой системы (Du et al., 2018).
Наконец, известно, что M. elongata является хозяином для различных видов эндобактерий (Uehling et al., 2017; Desirò et al., 2018). Поскольку эти бактерии являются отличительными эволюционными маркерами древних грибов Mucoromycota (Bonfante and Desirò, 2017), дополнительные сведения о M. elongata могут также помочь нам понять биологические свойства Mucoromycota, которые делают их склонными к инвазии как прокариотами, так и эукариотами. .
.
Ссылки
Статья и информация об авторе
Сведения об авторе
Паола Бонфанте
Паола Бонфанте работает на факультете наук о жизни и системной биологии Туринского университета, Турин, Италия
Для корреспонденции
Конкурирующие интересы
Конкурирующие интересы не заявлены
«Этот ORCID iD идентифицирует автора этой статьи:»
0000-0003-3576-8530
История публикаций
- Версия записи опубликована: 16 июля 2019 г. (версия 1)
Авторское право
© Bonfante, 2019 г.
Показатели
Число цитирований статей, полученное путем опроса самых высоких значений из следующих источников: Crossref, PubMed Central, Scopus.
Ссылки для скачивания
Список ссылок, состоящий из двух частей, для загрузки статьи или частей статьи в различных форматах.