Содержание
Растения приспосабливаются к новым опылителям всего за несколько поколений • Сергей Лысенков • Новости науки на «Элементах» • Эволюция, Экология
Швейцарские ученые изучили дивергентную эволюцию репы под действием двух разных опылителей — шмелей и мух-журчалок. Уже девять поколений отбора дали значимые результаты. Опыляемые шмелями растения стали выше и более пахучими, и шмели стали охотнее их посещать. А растения, опыляемые мухами, внешне почти не изменились и не стали более привлекательными для опыляющих их насекомых, но развили способность к размножению без опыления. Таким образом, адаптация к разным опылителям может происходить довольно быстро, а результатом может являться изменение системы размножения растений.
О важной роли животных-опылителей в эволюции писал еще Чарлз Дарвин, посвятивший антэкологическим исследованиям (антэкология — «цветочная экология», сейчас обычно под этим термином понимается экология опыления) несколько лет своей плодотворной жизни и написавший по их итогам два объемных труда — «Опыление у орхидных» (Fertilization of orchids) и «Действие перекрестного опыления и самоопыления в растительном мире» (The Effects of Cross- and Self-Fertilization in the Vegetable Kingdom). С тех пор адаптации цветков к разным группам опылителей стали хрестоматийными примерами коэволюции (совместной эволюции), а многие примеры дивергенции — расхождения признаков под действием отбора — также относятся к взаимоотношениям растений и насекомых (впрочем, не только насекомых: опылять могут и колибри, и летучие мыши, и некоторые другие животные).
С тех пор адаптации цветков к разным группам опылителей стали хрестоматийными примерами коэволюции (совместной эволюции), а многие примеры дивергенции — расхождения признаков под действием отбора — также относятся к взаимоотношениям растений и насекомых (впрочем, не только насекомых: опылять могут и колибри, и летучие мыши, и некоторые другие животные).
Обычно думают, что эволюционные процессы идут неспешно и занимают много времени, но иногда такое расхождение признаков может происходить довольно быстро. Например, в эксперименте на синюхе клейкой Polemonium viscosum, произрастающей в горах Северной Америки, было показано, что новые признаки могут возникнуть уже за одно поколение. Синюхи, растущие на границе леса, опыляются разнообразными насекомыми (мухами, пчелами, шмелями) и имеют узкий венчик. Цветы, растущие в высокогорных тундрах, опыляются в основном шмелями, и их венчик широкий (чтобы производить больше привлекательного нектара). На экспериментальном участке растения с узкими венчиками опылялись предварительно пойманными шмелями. В результате цветки следующего поколения уже имели более широкие венчики, чем родительские растения (см. C. Galen, 1996. Rates of floral evolution: adaptation to bumblebee pollination in an alpine wildflower, Polemonium viscosum).
В результате цветки следующего поколения уже имели более широкие венчики, чем родительские растения (см. C. Galen, 1996. Rates of floral evolution: adaptation to bumblebee pollination in an alpine wildflower, Polemonium viscosum).
Однако эта и подобные ей работы проводились в природных популяциях, где действуют и другие факторы — поэтому остается лазейка для скептика: а в опылителях ли дело? Может, различия связаны с чем-то другим, например, особенностями почвы, климата или патогенами?
Чтобы устранить эту неоднозначность и выявить непосредственное влияние разных групп опылителей на эволюцию растений, ученые из Цюрихского университета провели эксперимент для изучения дивергенции репы Brassica rapa под воздействием разных опылителей — шмелей и мух-журчалок. Репа была выбрана как представитель растений-генералистов (см. generalist and specialist species), опыляемых в природе разными группами насекомых. Семена были взяты из одного источника, чтобы исключить возможные эффекты из-за разных сортов, но они не были чистой линией и имели достаточное генетическое разнообразие, чтобы от них можно было ожидать быстрого ответа на отбор.
В эксперименте растения разделили на три группы: одна опылялась только земляными шмелями Bombus terrestris, вторая — журчалками Episyrphus balteatus, иногда называемыми «мармеладными мухами» (рис. 1), а третью опыляли сами исследователи (это была контрольная группа, не подвергавшаяся отбору на эффективность опыления насекомыми). Эксперимент с каждым типом опыления повторяли три раза, всегда используя по 36 растений для каждого типа (то есть в каждом повторе эксперимента выращивалось по 108 растений). Растения росли в одинаковых условиях (идентичная почва, освещение, условия полива).
Эксперимент длился на протяжении одиннадцати поколений. Семена для каждого следующего поколения отбирались пропорционально продуктивности растений: если одна репа произвела в два раза больше семян, чем другая, то при посадке растений для следующего поколения от нее брали в два раза больше семян. При этом общее число растений в поколениях сохранялось за счет того, что неопыленные растения не давали семян.
Ученые измеряли большое количество признаков растений (высоту, число цветков, размеры отдельных частей цветка, а также количество отдельных пахучих веществ, выделяемых при цветении) каждое нечетное поколение, чтобы отследить не только конечный результат эксперимента, но и динамику изменений. Кроме того, через связь признаков с семенной продуктивностью растений оценивалась сила отбора, действующая на тот или иной признак.
Группы опыляемых разными способами растений из последнего, одиннадцатого поколения существенно различались. Причем больше, чем растения между собой внутри каждой группы (это подтвердилось двумя независимыми статистическими методами). Это говорит в пользу того, что полученные результаты возникли под действием именно отбора, а не генетического дрейфа (см. Genetic drift).
К концу опыта изменились репы как из «шмелиной», так и из «мушиной» группы. Причем изменения оказались разными, что как раз и говорит об адаптивной дивергенции растений под действием отбора, вызванного наличием разных опылителей.
Опыляемые шмелями репы стали выше (рис. 2, а), более пахучими (общее количество выделяемых ими пахучих веществ удвоилось), а отражающая ультрафиолет зона на лепестках увеличилась (хотя в целом окраска (спектр отражения) цветков осталась той же). Изменился и запах, то есть состав пахучих веществ (рис. 2, c, d). Интересно, что шесть из восьми поддержанных отбором признаков (как морфологических, так и химических) положительно коррелируют с количеством нектара, то есть могут служить «честными указателями» на качество цветка. Самое же главное — через 11 поколений отбора репы из шмелиной группы стали гораздо более привлекательны для шмелей, чем таковые из мушиной группы (рис. 3, а). Но что касается общего числа производимых семян — ничего не изменилось (причем на рис. 2, b видно, что в течение всего опыта происходили колебания вокруг одного и того же значения). Еще одно заметное изменение в шмелиной группе — значительно увеличилась доля растений-обманщиков, не производящих нектар. Их стало около трети, в то время как в первом поколении, с которого начинался опыт, таковых вообще не было (рис. 3, b)!
3, b)!
С репами в мушиной группе произошли совсем другие изменения. Во-первых, у них увеличилась семенная продуктивность (рис. 2, b). Это можно было бы объяснить повышением эффективности привлечения журчалок, но мармеладные мухи не выказывали никакого особого предпочтения к растениям ни из шмелиной, ни из мушиной группы (рис. 3, a), да и число посещений на один цветок не изменилось. Безнектарные растения появились и здесь, но в незначительном числе (так же, как и в контрольной группе). Каких-либо заметных изменений в общем облике растений не произошло, они стали только в 1,2 раза ниже. Зато у них возросла роль «вспомогательных репродуктивных технологий» — самосовместимости (способности опыляться собственной пыльцой) и апомиксиса (способности образовывать семена без опыления), что в 15 раз увеличило число плодов, производимых растениями, вообще не посещенными опылителями! С этим соотносятся и некоторые иные тренды изменений, замеченные в этой группе: уменьшение длины пестика и продукции трех пахучих веществ (впрочем, выделение еще одного вещества, бензонитрила (benzonitrile), увеличилось).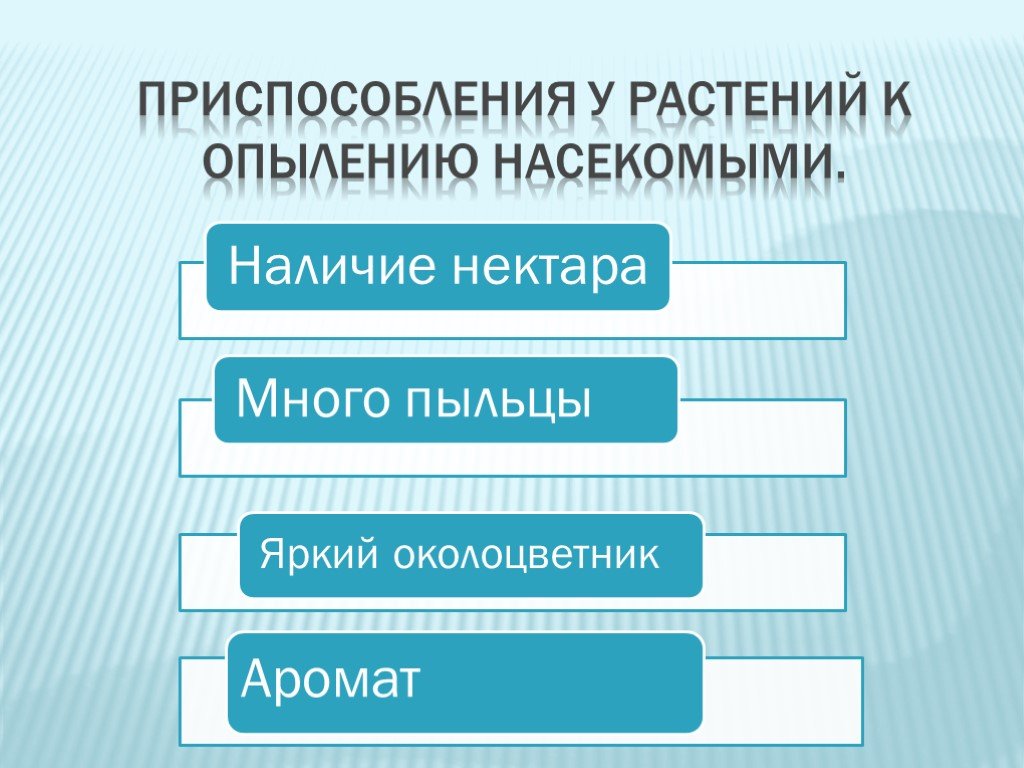 Эти признаки соотносятся с так называемым «синдромом самоопыления», то есть свойственны самоопыляющимся растениям. Впрочем, другие признаки (количество нектара и размер цветка обычно меньше у тех растений, которые полагаются на себя, а не на насекомых) не изменились в этом направлении — авторы связывают это с тем, что мухи-журчалки все-таки играют роль в опылении и способствуют отбору на на сохранение этих признаков.
Эти признаки соотносятся с так называемым «синдромом самоопыления», то есть свойственны самоопыляющимся растениям. Впрочем, другие признаки (количество нектара и размер цветка обычно меньше у тех растений, которые полагаются на себя, а не на насекомых) не изменились в этом направлении — авторы связывают это с тем, что мухи-журчалки все-таки играют роль в опылении и способствуют отбору на на сохранение этих признаков.
Таким образом, разные насекомые вызвали существенное расхождение признаков у растений. Причем, что важно, репа — это неспециализированное растение, приспособленное к посещению разными группами опылителей — как пчелами, так и мухами (и не только журчалками). И если в шмелиной группе приспособление шло именно в сторону лучшего опыления посещающими их насекомыми, то в мушиной отбор привел к развитию альтернативных способов размножения, при которых растение полагается лишь на себя, — по-видимому, мармеладные мухи оказались в целом малоэффективны в опылении цветков репы. Резкое возрастание числа растений-обманщиков, не производящих нектар, авторы тоже связывают с опылением шмелями, так как эти насекомые собирают пыльцу для выкармливания своего потомства и потому готовы посещать и такие цветки. Мухи-журчалки тоже питаются пыльцой, но нектар для них важнее — впрочем, авторы не утверждают этого строго и приводят лишь косвенные доводы в пользу этого предположения: мармеладные мухи могут различать разную концентрацию сахаров, но не количество пыльцы на искусственных цветках (J. P. Sutherland et al., 1999. The influence of floral character on the foraging behaviour of the hoverfly, Episyrphus balteatus)
Мухи-журчалки тоже питаются пыльцой, но нектар для них важнее — впрочем, авторы не утверждают этого строго и приводят лишь косвенные доводы в пользу этого предположения: мармеладные мухи могут различать разную концентрацию сахаров, но не количество пыльцы на искусственных цветках (J. P. Sutherland et al., 1999. The influence of floral character on the foraging behaviour of the hoverfly, Episyrphus balteatus)
Таким образом, изменение состава опылителей даже у растений, исходно «настроенных» на опыление разнообразными животными, может довольно быстро повлиять на облик и способ размножения. Причем, как показал этот эксперимент, не всегда эволюция пойдет по пути повышения эффективности опыления наиболее частым посетителем — если последний явно недостаточен для нормального размножения (как, видимо, произошло, с довольно маленькой журчалкой, лишенной заметного опушения, помогающего переносить пыльцу), отбор может привести к развитию других способов производства семян.
Источник: Daniel D. L. Gervasi, Florian P. Schiestl. Real-time divergent evolution in plants driven by pollinators // Nature Communications. 2017. DOI: 10.1038/ncomms14691.
L. Gervasi, Florian P. Schiestl. Real-time divergent evolution in plants driven by pollinators // Nature Communications. 2017. DOI: 10.1038/ncomms14691.
См. также об адаптации растений к разным опылителям:
1) Длина пестика у цветка Nerine humilis определяется сообществом насекомых-опылителей, «Элементы», 02.10.2015.
2) Конкуренция за опылителей делает совместно цветущие виды пасленовых непохожими по окраске, «Элементы», 16.05.2014.
3) Чтобы избежать межвидовой гибридизации, орхидеям достаточно по-разному пахнуть, «Элементы», 15.06.2011.
Сергей Лысенков
описание, особенности, этапы и виды
В качестве конечной задачи типичного цветка выступает формирование плодов и семян. Для этого необходимо два процесса. Первым является опыление цветков растений. После него происходит собственно оплодотворение — появляются плоды и семена. Рассмотрим далее, какие существуют виды опыления растений.
Общие сведения
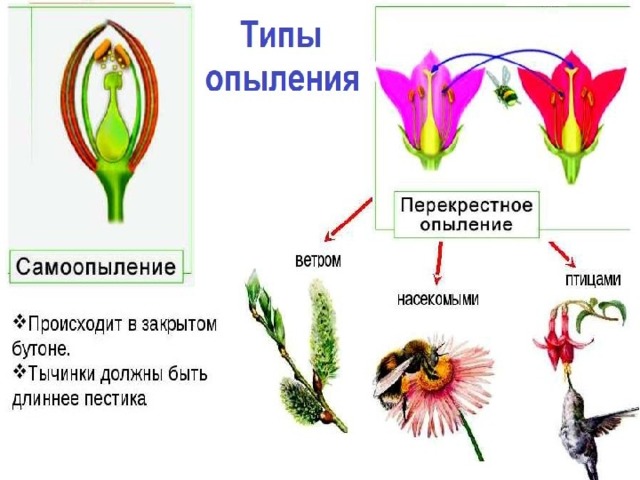
Опыление растений – этап, на котором осуществляется перенос мелких зерен с тычинок на рыльце. Оно тесно связано с другой стадией развития культур — формированием органа размножения. Учеными было установлено два вида опыления: аллогамия и автогамия. При этом первая может осуществляться двумя путями: гейтоногамией и ксеногамией.
Оно тесно связано с другой стадией развития культур — формированием органа размножения. Учеными было установлено два вида опыления: аллогамия и автогамия. При этом первая может осуществляться двумя путями: гейтоногамией и ксеногамией.
Характеристики
Автогамия – опыление растений путем переноса зерен с тычинок на рыльце одного органа размножения. Другими словами, одна система самостоятельно осуществляет необходимый процесс. Аллогамия – перекрестный перенос зерен с тычинок одного органа на рыльце другого. Гейтоногамия предполагает опыление между цветками одной, а ксеногамия – разных особей. Первая генетически схожа с автогамией. В этом случае имеет место только перекомбинация гамет в одной особи. Как правило, такое опыление характерно для многоцветковых соцветий.
Самой благоприятной по своему генетическому эффекту считается ксеногамия. Такое опыление цветковых растений способствует увеличению возможностей перекомбинации генетических данных. Это, в свою очередь, обеспечивает повышение внутривидового разнообразия, последующей приспособительной эволюции. Между тем, автогамия обладает немаловажным значением для стабилизации видовых признаков.
Между тем, автогамия обладает немаловажным значением для стабилизации видовых признаков.
Способы
Метод опыления зависит от агентов переноса зерен и структуры цветка. Аллогамия и автогамия могут осуществляться с помощью одних и тех же факторов. Ими, в частности, выступают ветер, животные, человек, вода. Наибольшим разнообразием отличаются способы при аллогамии. Выделяют следующие их группы:
- Биологические – опыление растений осуществляется с помощью живых организмов. В этой группе выделяют несколько подгрупп. Классификация осуществляется в зависимости от переносчика. Так, осуществляется опыление растений насекомыми (энтомофилия), птицами (орнитофилия), летучими мышами (хироптерофилия). Существуют и другие способы – с помощью моллюсков, млекопитающих и пр. Однако они выявляются в природе достаточно редко.
- Абиотические – опыление растений связано с влиянием небиологических факторов. В этой группе различают перенос зерен с помощью ветра (анемофилию), воды (гидрофилию).

Способы, которыми осуществляется опыление растений, считаются адаптациями к конкретным окружающим условиям. В генетическом плане они менее важны, чем типы.
Приспособление растений к опылению
Рассмотрим первую группу способов. В природе, как правило, встречается энтомофилия. Эволюция растений и переносчиков пыльцы проходила параллельно. Энтомофильные особи легко выделяются среди прочих. У растений и переносчиков есть взаимные приспособления. В некоторых случаях они настолько узкие, что культура не в состоянии самостоятельно существовать без своего агента (или наоборот). Насекомых привлекает:
- Цвет.
- Пища.
- Запах.
Кроме этого, некоторые насекомые используют цветки как убежище. Например, они прячутся там ночью. Температура в цветке выше, чем у внешней среды, на несколько градусов. Существуют насекомые, которые сами размножаются в культурах. Например, осы-хальциды используют для этого цветки.
Орнитофилия
Опыление птицами наблюдается преимущественно в тропических районах. В редких случаях орнитофилия имеет место в субтропиках. К признакам цветков, привлекающих птиц, можно отнести:
В редких случаях орнитофилия имеет место в субтропиках. К признакам цветков, привлекающих птиц, можно отнести:
- Отсутствие запаха. У птиц достаточно слабое обоняние.
- Венчик имеет в основном оранжевый или красный цвет. В редких случаях отмечается синяя или фиолетовая окраска. Стоит сказать, что птицы легко отличают эти цвета.
- Большое количество слабоконцентрированного нектара.
Птицы зачастую не садятся на цветок, а опыляют, зависая рядом с ним.
Хироптерофилия
Летучие мыши опыляют преимущественно тропические кустарники и деревья. В редких случаях они участвуют в переносе зерен на травы. Летучие мыши опыляют цветки ночью. К признакам культур, которые привлекают этих животных, относят:
- Наличие флуоресцентной белой или желто-зеленой окраски. Она также может быть коричневатой, в редких случаях фиолетовой.
- Наличие специфического запаха. Он напоминает секреты и выделения мышей.
- Цветки распускаются ночью либо вечером.
- Крупные части свисают с ветвей на длинных цветоножках (баобаб) либо развиваются непосредственно на стволах деревьев (какао).

Анемофилия
Опыление приблизительно 20 % растений умеренной полосы осуществляется с помощью ветра. На открытых площадях (в степях, пустынях, полярных территориях) этот показатель значительно выше. Анемофильные культуры обладают следующими признаками:
- Мелкие невзрачные цветки, имеющие желтоватый или зеленоватый оттенок, часто не имеющие околоцветника. Если же он присутствует, то представлен в форме пленок и чешуй.
- Наличие многоцветковых соцветий. Такой «букет» может быть представлен повислой осью – сережкой.
- Наличие пыльников на тычиночных тонких нитях.
- Достаточно крупные и зачастую перистые рыльца, выступающие за пределы цветка.
- Культуры одно- или двудомные.
- Образование большого количества пыльцы. Она сухая, мелкая, гладкая. Зерна могут иметь дополнительные приспособления (воздушные мешки, например).

Анемофильные культуры часто формируют большие скопления. Это значительно увеличивает шансы на опыление. В качестве примеров выступают березовые рощи, дубравы, заросли бамбука.
Гидрофилия
Такое опыление достаточно редко встречается в природе. Это обуславливается тем, что вода не является обычной средой обитания культур. У многих растений цветки находятся над поверхностью и опыляются преимущественно насекомыми либо с помощью ветра. К признакам гидрофильных культур можно отнести:
- Цветки мелкие и невзрачные. Они развиваются поодиночке или собираются в небольшие «букеты».
- Как правило, цветки однополые. В качестве примеров выступают валлиснерия, элодея.
- В пыльниках стенка тонкая. В них отсутствует эндотеций. Часто пыльники имеют нитевидную форму. У некоторых культур они оплетают рыльце. Это способствует быстрому проникновению и прорастанию пыльцы.
- В зернах нет экзины. Это обусловлено тем, что пыльца находится в воде и ей не требуется защита от высыхания.

Автогамия
У 75 % растений присутствуют обоеполые цветки. Это обеспечивает самостоятельный перенос зерен без внешних носителей. Автогамия нередко бывает случайной. Это имеет место особенно при неблагоприятных условиях для переносчиков.
Автогамия базируется на принципе «лучше самостоятельное опыление, чем вообще никакого». Такой тип переноса зерен известен у многих культур. Как правило, они развиваются в неблагоприятных условиях, на территориях, где сильно холодно (тундра, горы) или очень жарко (пустыня) и отсутствуют переносчики.
В природе, между тем, встречается и регулярная автогамия. Она постоянна и крайне важна для культур. К примеру, самоопыляются такие растения, как горох, арахис, пшеница, лен, хлопчатник и прочие.
Подтипы
Автогамия может быть:
- Контактной. При движении тычиночных нитей пыльники непосредственно касаются рыльца. Такая автогамия характерна для копытня, седмичника.
- Гравитационной. В этом случае пыльца попадает на рыльце с пыльников, располагающихся выше. При гравитационной автогамии, таким образом, действует сила тяжести. Это характерно для вересковых, грушанковых культур.
- Клейстогамной. В этом случае опыление осуществляется в бутоне или закрытом цветке. Клейстогамия считается крайней степенью автогамии. Она может обуславливаться неблагоприятными факторами (высокая влажность или засуха). Клейстогамия может быть и регулярной, генетически закрепленной. К примеру, у фиалки удивительной в весеннее время появляются сначала нормальные цветки, но опыление в них не происходит, соответственно, не появляется плодов и семян. Впоследствии появляются клейстогамные органы размножения. Они не раскрываются и представлены в форме бутонов. Прорастание пыльцы происходит непосредственно в пыльниках. Трубка проходит сквозь стенку и достигает рыльца. В результате формируется коробочка с семенами.
 При гравитационной автогамии, таким образом, действует сила тяжести. Это характерно для вересковых, грушанковых культур.
При гравитационной автогамии, таким образом, действует сила тяжести. Это характерно для вересковых, грушанковых культур.Клейстогамия обнаруживается в разных систематических группах культур (в некоторых злаках, например).
Укажите четыре адаптации цветков, опыляемых насекомыми.

Ответить
Проверено
214,8 тыс.+ просмотров
Подсказка: Опыление насекомыми называется энтомофилией. Наиболее распространен среди наземных растений. Энтомофильные цветы специально приспособлены для привлечения насекомых.
Полный ответ:
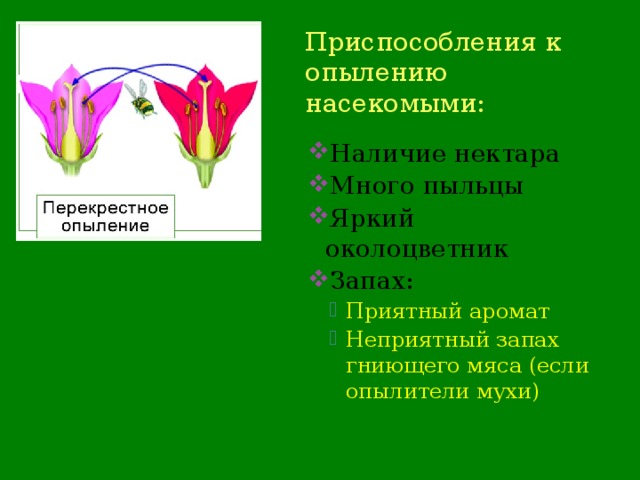
Некоторые приспособления энтомофильных цветков следующие;
1) Очень заметные цветки с ярко окрашенными лепестками и околоцветниками
2) Маленькие цветки собираются в соцветия, чтобы стать заметными.
3) Цветы обычно пахнут для привлечения насекомых.
4) Цветы имеют нектарники для производства и хранения нектара, который служит пищей для насекомых.
5) Пыльцевые зерна обычно имеют шероховатую поверхность и липкие, поэтому они легко прилипают к телу насекомого.
Насекомые могут переносить пыльцевые зерна с пыльников одного цветка и откладывать их точно на рыльце пестика другого цветка. Насекомые посещают цветы, чтобы питаться пыльцой и нектаром, откладывать яйца, в поисках убежища или по неизвестным причинам. Для привлечения насекомых энтомофильные цветы имеют крупные и ярко окрашенные лепестки. Если цветки сгруппированы в заметные соцветия с ароматом. Насекомые-посетители отдают предпочтение определенным цветам. Пчелы, как правило, предпочитают синий цвет, бабочки предпочитают красный цвет, а некоторые другие мухи предпочитают коричневый или фиолетовый цвет.
Для привлечения насекомых энтомофильные цветы имеют крупные и ярко окрашенные лепестки. Если цветки сгруппированы в заметные соцветия с ароматом. Насекомые-посетители отдают предпочтение определенным цветам. Пчелы, как правило, предпочитают синий цвет, бабочки предпочитают красный цвет, а некоторые другие мухи предпочитают коричневый или фиолетовый цвет.
Дополнительная информация:
— Мотыльки, бабочки, жуки являются обычными насекомыми-опылителями. Опыление мотыльками называется фаленофилией, бабочками — психофилией, пчелами — меллитофилией, жуками — кантарофилией.
— Насекомые посещают цветы для получения нектара. Нектарники обычно находятся внутри трубки венчика или шпорца.
— Некоторые цветы выделяют сочные или липкие вещества, благодаря которым пыльца прилипает к телу насекомых
Примечание:
— Иногда венчики цветков опыления насекомыми очень маленькие и незаметные, и в таких случаях некоторые другие части цветка становятся большими или привлекательными.
-Помимо цвета, насекомые ориентируются на цветы по их запаху.
— Некоторые цветы не имеют запаха в дневное время, но издают сильный запах ночью, и такие цветы опыляются ночными бабочками.
Недавно обновленные страницы
Большинство эубактериальных антибиотиков получают из биологии Rhizobium класса 12 NEET_UG
Саламиновые биоинсектициды были извлечены из биологии класса 12 А NEET_UG
Какое из следующих утверждений относительно бакуловирусов класса 12 биологии NEET_UG
Канализационные или городские канализационные трубы не должны быть непосредственно классом биологии 12 NEET_UG
Очистка сточных вод осуществляется путем микроочистки сточных вод B Удобрения 12-го класса биологии NEET_UG
Иммобилизация ферментов — это A Преобразование активного фермента 12-го класса биологии NEET_UG
Большинство эубактериальных антибиотиков получают из A Rhizobium класса 12 биологии NEET_UG
Саламиновые биоинсектициды были извлечены из биологии класса 12 А NEET_UG
Какое из следующих утверждений относительно бакуловирусов класса 12 биологии NEET_UG
Канализационные или городские канализационные трубы не должны быть непосредственно классом биологии 12 NEET_UG
Очистка сточных вод осуществляется путем микроочистки сточных вод B Удобрения 12-го класса биологии NEET_UG
Иммобилизация ферментов — это A Превращение активного фермента 12-го класса биологии NEET_UG
Трендовые сомнения
Студенты также читают
Разница между цветами, опыленными насекомыми и опыленными ветром,
Адаптация полетов
Адаптация на пастбищах для растений и животных
Опыление птицами
Опыление Уликой
Осполов.
Опыление
Искусственное опыление
Видео с вопросами: Вспоминая типичные цветочные адаптации растений, опыляемых насекомыми
Стенограмма видео
На представленной диаграмме показано, как
опылитель опыляет растение, а значит, переносит мужские половые клетки
содержится в пыльце от пыльника цветка до рыльца того же или
другой цветок. Поэтому это растение является
насекомоопыляемое растение. Что из перечисленного не
опишите адаптацию, полезную для растения, опыляемого насекомыми? а) образующаяся пыльца липкая
или остроконечный. (Б) Цветы содержат сладкий сахар
жидкость, называемая нектаром. (С) Цветки крупные и яркие
цветные лепестки. Или (Г) пыльники и рыльце висят
свободно снаружи цветка.
Насекомые-опылители, такие как пчелы и
бабочки, могут быть очень полезны для многих видов цветковых растений, так как они
способны довольно эффективно переносить пыльцу на большие расстояния. Пыльцевые зерна, содержащие
Пыльцевые зерна, содержащие
мужские половые клетки цветкового растения образуются в частях цветка, называемых
пыльники. Стигма является частью
женский репродуктивный орган цветка. Рыльце ведет вниз к яичнику
который содержит одну или несколько женских половых клеток, яйцеклеток.
Как сказано в вопросе,
опыление – это процесс переноса пыльцы с пыльника на рыльце пестика. Это позволяет мужской половой клетке
в конечном итоге оплодотворяют женскую половую клетку в яичнике. Давайте посмотрим на некоторые из
типичные приспособления цветков, опыляемых насекомыми, чтобы мы могли выяснить, какие из
варианты ответов не будут полезны для этого способа опыления.
Некоторые цветы, опыляемые насекомыми
производят сладкую жидкость, называемую нектаром, которая привлекает насекомых к цветку,
особенно пчелы, которые используют нектар для производства меда. Когда насекомые-опылители, такие как
Когда насекомые-опылители, такие как
пчела на изображении, предоставленном нам в вопросе, посещает цветы, чтобы получить доступ к нектару,
часть пыльцы обычно стирается с пыльников на тело насекомого. Когда насекомое посещает другое
цветок, пыльца, которая, как вы помните, содержит мужские половые клетки, может стереться
тело опылителя и на рыльце этого второго цветка. С тем, что мы узнали до сих пор,
мы можем исключить вариант ответа (Б): цветы содержат сладкую сахаристую жидкость, называемую
нектар. Мы знаем, что это приспособление цветка
полезен для некоторых растений, опыляемых насекомыми.
Откуда насекомое знает, что
цветок может содержать нектар? Ну и насекомоопыляемые цветы
как правило, имеют особенно большие и ярко окрашенные лепестки, что свидетельствует о том, что
у этого цветка может быть нектар для насекомого. С помощью этой информации мы можем
С помощью этой информации мы можем
исключить вариант ответа (С): цветы имеют большие и ярко окрашенные лепестки.
В цветках, опыляемых насекомыми,
пыльники и рыльца часто находятся внутри цветка, перед нектаром. Это позволяет пыльникам откладывать
как можно больше пыльцы попадает на тела опылителей, когда они вступают в контакт с
цветок. Это также делает более вероятным, что
пыльца, которая уже находится на теле насекомого, попадет на рыльце пестика. Это говорит о том, что правильный
ответ, скорее всего, будет вариантом (D): пыльники и рыльце свободно свисают снаружи
цветок. Это не помогло бы в
откладывание пыльцы на тело насекомого, также не было бы полезно откладывать пыльцу
со своего тела на клеймо. Вместо этого насекомые могут обойти эти
структуры целиком, если они расположены вне цветка.
Проверим вариант ответа (А) —
произведенная пыльца липкая или колючая — чтобы подтвердить наш выбор.