
Особенности растительных тканей. Образовательные ткани. Образовательные ткани растений
38 Образовательные ткани. Их типы, положение в теле растения.
ОБРАЗОВАТЕЛЬНЫЕ ТКАНИ (меристемы), ткани растений, состоящие из клеток, которые длительное время сохраняют способность к делению. Благодаря многократному делению этих клеток происходит рост растений в течение всей их жизни (у некоторых деревьев это сотни и даже тысячи лет). Второе важное свойство клеток меристем заключается в том, что они дают начало специализированным клеткам, образующим постоянные ткани – покровные, основные, проводящие, механические, выделительные.
В зависимости от распределения на теле формирующегося растения выделяют четыре вида меристем.
Верхушечные, или апикальные, меристемы обеспечивают рост побегов и корней в длину.
Боковые, или латеральные, меристемы обусловливают нарастание стеблей и корней в толщину и называются камбием.
Вставочные, или интеркалярные, меристемы временно сохраняются в междоузлиях стебля и в основаниях молодых листьев, обеспечивая рост этих участков, но затем превращаются в постоянные ткани. Раневые, или травматические, меристемы возникают в местах повреждения растения, где образуют защитный каллюс.
Типы меристем:
1)Пластинчатые меристемы, состоят из одного слоя клеток, деление происходит перпендикулярно поверхности органа. Учувствуют в образовании эпидермы.
2)Колончатые или стержневые, кубические и призматические клетки, которые делятся в плоскости, перпендикулярной продольной оси органа.
3)Массивные меристемы, клетки которых делятся в разных направлениях, осуществляя равномерное увеличение объема этой ткани.
39. Понятие об элементарных, годичных и монокарпических побегах.
Побег, вырастающий из почки за один период роста, называется элементарным. Побег, развивающийся в течение одного вегетационного периода называется – годичным побегом, а у многолетних растений годичным приростом. В умеренном климате большинство древесных растений имеет лишь один прирост за год, т. е. элементарный побег у них является = годичному. Но у дуба, например, нередко в середине лета наступает второй период; тогда годичный Побег состоит из двух элементарных. Побег цитрусовых дают в год нередко 3-4 прироста, тропические деревья (какао) - до 7 приростов - это характерно для без сезонного климата. В сезонном климате 1 годичный побег = 1-2 элементарным побегам.
Основной единицей побеговой системы растений является монокарпический побег(МНКП). Это однолетние или многолетние побеги, отмирающие после цветения и образования плодов и семян. МНКП развивается из почки и в течение одного или нескольких лет нарастает моноподиально. Заканчивается цикл развития МНКП цветением и плодоношением. По окончании плодоношения МНКП отмирает, за исключением своей базальной части, где располагаются органы возобновления, дающие начало новым дочерним побегам и придаточным стеблевым корням. Многолетние травянистые растения представляют собой в морфологическом отношении систему последовательно сменяющих друг друга МНКП.
40 Морфологическое разнообразие листьев. Принципы их классификации.
Лист – боковой орган растения с ограниченным верхушечным ростом, выполняющий три функции: фотосинтез, транспирацию и дыхание.
Типичный лист состоит из пластинки, имеющей дорзовентральной строение (верхняя адаксиальная сторона отличается от нижней – абаксиальной), основания, черешка и прилистников.
Листорасположение описывается при помощи следующих терминов:
Очерёдное(последовательное) — листья располагаются по одному (в очередь) на каждый узел.
Супроти́вное— листья располагаются по два на каждом узле и обычно перекрёстно-попарно, то есть каждый последующий узел на стебле развёрнут относительно предыдущего на угол 90°; либо двумя рядами, если не развёрнут, но имеется несколько узлов.
Муто́вчатое— листья располагаются по три и более на каждом особом узле стебля — мутовке. В отличие от супротивных листьев, у мутовчатых каждый последующий завиток может находиться под углом 90° от предыдущего, а может и не находиться, вращаясь на половину угла между листьями в завитке. Следует однако учесть, что супротивные листья могут показаться мутовчатыми на конце стебля.
Розе́точное— листья, расположенные в розетке (пучок листьев, расположенных по кругу из одного общего центра).
По тому, как листовые пластинки разделены, могут быть описаны две основные формы листьев:
Простой листсостоит из единственной листовой пластинки и одного черешка. Хотя он может состоять из нескольких лопастей, промежутки между этими лопастями не достигают основной жилки листа. Простой лист всегда опадает целиком.
Если выемки по краю простого листа не достигают четверти полуширины листовой пластины, то такой простой лист называется цельным.
Сложный листсостоит из несколькихлисточков, расположенных на общем черешке (который называетсярахис). Листочки, помимо своей листовой пластинки, могут иметь свой черешок (который называетсячерешочек, иливторичный черешок) и свои прилистники (который называютсяприлистничками, иливторичными прилистниками). В сложном листе каждая пластинка опадает отдельно. Так как каждый листочек сложного листа можно рассматривать как отдельный лист.
У пальчатыхлистьев все листовые пластинки расходятся по радиусу от окончания корешка подобно пальцам руки. Главный черешок листа отсутствует. Примерами таких листьев может служить конский каштан.
У перистыхлистьев листовые пластинки расположены вдоль основного черешка. В свою очередь, перистые листья могут бытьнепарноперистыми, с верхушечной листовой пластинкой, например, ясень ипарноперистыми, без верхушечной пластинки.
У двуперистыхлистьев листья разделены дважды: пластинки расположены вдоль вторичных черешков, которые в свою очередь прикреплены к главному черешку.
У трёхлистныхлистьев имеется только три пластинки; например, клевер.
studfiles.net
Ткани высших растений
Содержание:
Понятие ткани
Ткань — это устойчивый комплекс клеток, имеющих общее происхождение и сходное строение и выполняющих одинаковые функции.
Виды растительных тканей: основные, образовательные, механические, проводящие, покровные, выделительные.
Простые ткани — ткани, состоящие из одного вида клеток и выполняющие только основные функции (пример: ткани листа, молодого корня).
Сложные ткани — ткани, состоящие из различных по строению клеток, выполняющих, наряду с основными, некоторые дополнительные .функции (пример: клетки ксилемы выполняют проводящую и опорную функции).
Основная ткань (паренхима)
♦Основная ткань (паренхима) — это ткань растений, обеспечивающая многообразные функции (см. ниже) и образованная крупными живыми клетками, среди которых располагаются различные специализированные ткани.
■ Продольные и поперечные размеры клеток паренхимы отличаются не более чем в два раза.
■ Основные функции паренхимы: фотосинтезирующая (ассимилирующая), запасающая, воздухоносная и др.
❖ Виды основной ткани: хлоренхима, запасающая паренхима, аэренхима, водоносная паренхима.
Хлоренхима (хлорофиллоносная, или ассимиляционная паренхима) — ткань растений, в клетках которой в большом количестве содержатся хлоропласты. Состоит из тонкостенных клеток; выполняет функцию фотосинтеза и образуется в зеленых листьях и приповерхностных слоях стеблей растений.
Запасающая паренхима — ткань, в клетках которой откладываются в твердом или растворенном виде запасные питательные вещества (крахмал, сахара, белки), впоследствии использующиеся растением в процессе его жизнедеятельности.
Аэренхима (или воздухоносная паренхима) — воздухоносная ткань растений, образованная клетками разной формы и имеющая хорошо развитые межклетники, по которым циркулируют газы; способствует снабжению растения кислородом или углекислым газом.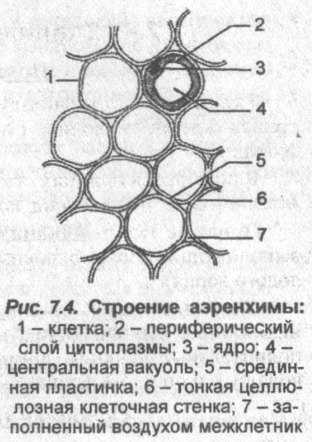
■ Аэренхима развивается в разных органах болотных и водных высших растений (кувшинок и др.) и обеспечивает у них нормальный газообмен в условиях пониженной аэрации.
Водоносная паренхима — особая ткань растений, образованная крупными клетками паренхимы, имеющими тонкие стенки и лишенными хлоропластов, в вакуолях которых содержатся слизистые вещества, удерживающие воду; способствует снабжению клеток растения водой.
■ Водоносная паренхима характерна для высших растений засушливых районов (кактусов, агав, алоэ и др.) и солончаков и обеспечивает у них нормальное водоснабжение в условиях длительного отсутствия влаги.
Образовательные ткани (меристемы)
❖ Образовательная ткань, или меристема, состоит из активно делящихся клеток с интенсивным обменом веществ и обеспечивает рост растения в течение всей его жизни за счет постоянного деления и образования новых клеток.
Особенности клеток образовательной ткани: клетки не дифференцированы, многогранны, плотно прилегают друг к другу, имеют тонкие стенки, крупное, расположенное в центре ядро, густую цитоплазму и небольшие вакуоли; могут делиться в разных направлениях. Одна часть клеток меристем постепенно дифференцируется, превращаясь в клетки различных постоянных тканей растения и формируя его тело, другая их часть задерживается на эмбриональной стадии развития в течение всей жизни растения.
esculappro.ru
Особенности растительных тканей. Образовательные ткани
Особенности растительных тканей
При выделении, изучении и систематизации тканей у растений необходимо учитывать их специфические особенности.1. Образование, строение, топография и функции тканей контролируются генетически. Это объясняет сходство и различие тканей у разных генотипов растений.
2. Ткани не возникают в дифинитивном, т.е. в окончательно завершенном виде. Они развиваются в ходе онтогенеза растений. В процессе онтогенеза химический состав, клеточное строение и функции тканей могут изменяться. Например, у мятликовых оболочки клеток мелкоклеточной паренхимы стебля, примыкающей к склеренхиме, могут пропитываться лигнином, повышая жесткость соломины. У древесных пород по мере старения стебля происходит необратимое разрушение сосудов и преобразование проводящей древесины в ядровую, т.е. непроводящую. Показателен пример изменения структуры проводящих пучков у травянистых двудольных. Исходно они развиваются из прокамбия и состоят из протоксилемы и протофлоэмы, позднее в пучках появляются проводящие элементы первичной метаксилемы и первичной метафлоэмы. С появлением камбия в таких пучках образуются элементы вторичной ксилемы и вторичной флоэмы.
3. Ткани могут быть образованы пространственно разобщенными клетками. Так, в частности, располагаются опорные клетки в листьях чая китайского.
4. Разные ткани могут выполнять одинаковые функции. Например, упругость стебля обеспечивается в первую очередь механическими тканями и существенно дополняется проводящими.
5. У растений можно наблюдать постепенный переход одних тканей в другие. В зонах роста корней, стеблей и других органов отсутствуют четкие границы между образовательными и постоянными тканями.
6. Функционально и структурно сходные ткани могут иметь разное происхождение. Например, механическая ткань склеренхима может образоваться из клеток перицикла и клеток камбия; проводящие ткани у двудольных могут возникнуть из прокамбия и камбия.
7. Различия в клеточном строении одной и той же ткани могут возникнуть в результате гетерохронного, т.е. разновременного, их заложения. Поэтому различаются между собой клетки весенней, летней и осенней древесины одного и того же годичного кольца у деревьев, равно как и ткани разных междоузлий у мятликовых.
8. Количественные показатели тканей могут существенно изменяться под влиянием средовых факторов. Например, в зависимости от режима освещения изменяется плотность расположения устьиц на поверхности листа; субклеточный состав ассимиляционной паренхимы зависит от обеспеченности растений азотом и водой.
При изучении растительных тканей широко используются методы ботаники и других наук. Среди них наиболее результативными считаются методы оптической и электронной микроскопии; гистохимический метод, основанный на специфическом окрашивании разных тканей цитологическими красителями; методы физики – деформационный, поляризационный и интерференционный; биохимические и физиологические методы. Математические методы широко используются для анализа первичной информации о клетках и тканях. Большую перспективу имеют методы экологической анатомии.
Образовательные ткани
Значение и разнообразие образовательных тканей
Отличительной особенностью растений является их способность к неограниченному росту. Рост растений служит основой развития как отдельных органов, так и всего организма, он генетически детерминирован и обеспечивается двумя процессами – делением клеток и их растяжением. Растяжение клеток указывает на начало их дифференциации и формирование постоянных тканей. В этом процессе важная роль принадлежит фитогормонам.
Деление клеток не создает новых структур, но поставляет исходный материал для построения тканей и органов растений, а следовательно, служит исходным процессом для последующего роста и развития. Деление клеток в типичных условиях вегетации является отличительным признаком образовательных тканей растений, или меристем. Выделяют два типа клеток меристем. Одни из них, именуемые инициалями, способны делиться неограниченно многократно, самовоспроизводиться при этом и давать начало клеткам второго типа – производным от инициалей. Производные инициалей делятся ограниченное число раз и преобразуются в постоянные ткани.
 Клетки апикальной меристемы кончика корня лука
Клетки апикальной меристемы кончика корня лука
По происхождению образовательные ткани бывают первичными и вторичными. Первичные возникают при развитии зародыша семени или в результате морфогенетической деятельности конуса нарастания почки. К первичным меристемам относятся апикальные и интеркалярные меристемы побега, прокамбий и перицикл, спорогенная меристема. Из первичных меристем образуются первичные постоянные ткани, но могут возникнуть и вторичные меристемы. Например, из клеток перицикла в корне двудольных могут образоваться камбий и феллоген (пробковый камбий).
Вторичные меристемы могут возникнуть либо из первичных меристем, либо в результате деления дифференцированных клеток основной паренхимы, как это бывает при образовании камбия.
Однако надо иметь в виду, что при использовании современных методов биотехнологии, меристематическая ткань может быть генерирована из любой живой растительной клетки. Особенности клеточного строения меристем связаны с их функцией. Клетки меристем всегда молодые, их развитие задерживается на эмбриональной фазе. Поэтому размеры клеток невелики, форма паренхимная – кубическая, многогранная до почти округлой. Клеточные стенки тонкие, без вторичных целлюлозных утолщений. Цитоплазма густая, вязкая, оптически более плотная, чем у других тканей. Ядро крупное, в период роста растений интерфаза – автосинтетическая, т.е. связана с подготовкой клетки к очередному делению. Хорошо развиты органоиды, которые обеспечивают синтез белка. Комплекс Гольджи и вакуоли выражены слабо. Между массой ядра и цитоплазмы поддерживается постоянное соотношение. Увеличение массы ядра является условием начала клеточного деления. Меристемы, связанные с образованием вегетативных частей растения, делятся путем митоза. Поэтому в их клетках набор хромосом и генов поддерживается постоянным. В спорогенных меристемах проходит мейоз, что объясняет возникновение генетически разнокачественных гаплоидных спор.
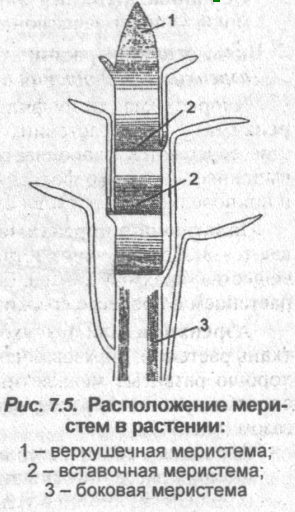
По месту расположения в растительном организме меристемы бывают апикальными, интеркалярными, латеральными, раневыми и спорогенными.
Апикальные меристемы
Апикальные меристемы находятся на верхушке побега в составе тканей конуса нарастания, а также в кончике корня. Поэтому их называют также верхушечными меристемами. В конусе нарастания выделяют: клетки туники, из которых развивается эпидермис; центральную меристематическую зону, которая является резервом образовательных клеток для других зон; серединную меристему, порождающую клетки сердцевины; периферическую зону, из клеток которой формируется прокамбий; и основную меристему, обеспечивающую образование первичной коры и паренхимы центрального цилиндра.
В кончике корня апикальная меристема расположена в подзоне деления. Из инициальных клеток этой меристемы образуется несколько групп клеток: калиптроген, характерный для однодольных, клетки которого порождают корневой чехлик; дерматоген, дифференцирующийся в эпиблему – первичную покровную ткань корня; периблему, из которой развивается первичная кора корня; плерому, используемую для построения центрального цилиндра.
Интеркалярные меристемы
Интеркалярные, или вставочные, меристемы являются первичными по происхождению. Они представляют собой остатки апикальных меристем и локализованы в базальной части междоузлий и в основаниях листьев. Их клетки длительное время могут находиться в эмбриональном состоянии и значительно отставать в своем развитии от рядом расположенных клеток постоянных тканей. Активное деление клеток интеркалярных меристем начинается по мере замедления и прекращения апикального роста побега. Например, у пшеницы и других мятликовых при длине побега около 1 см конус нарастания дифференцируется и вместо метамеров вегетативной части побега продуцирует метамеры сложного колоса или другого соцветия. К этому времени на побеге образуется 4 – 6 надземных междоузлий, у которых последующий рост в длину обеспечивается делением клеток интеркалярной меристемы и растяжением их производных. Благодаря интеркалярным меристемам лист у покрытосеменных нарастает своим основанием после выхода из почки.
Латеральные меристемы
Латеральные, или боковые, меристемы располагаются в стебле и корне параллельно их поверхности и обеспечивают прирост растений в толщину.
К первичным латеральным меристемам относятся прокамбий и перицикл. Из прокамбия образуются ткани проводящих пучков стебля. Если прокамбий закладывается в конусе нарастания сплошным кольцом, то при последующем развитии формируется стебель непучкового типа строения, как у льна. При заложении прокамбия отдельными тяжами по кругу, в стебле развиваются обособленные проводящие пучки, как, например, у пшеницы и других мятликовых.
Клетки перицикла функционально более многообразны. Из них в стебле образуется механическая ткань склеренхима, в корне – может возникнуть камбий и феллоген (пробковый камбий). Кроме того, клетки перицикла участвуют в образовании боковых корней.
Вторичными латеральными меристемами являются камбий и феллоген, которые образуются либо из перицикла, либо из клеток основной паренхимы; камбий может также возникнуть из прокамбия. Они характерны для осевых вегетативных органов двудольных покрытосеменных и голосеменных. Камбий обеспечивает образование вторичных элементов проводящих тканей и пучков, луба и древесины. Феллоген порождает клетки, из которых образуется пробка и феллодерма. Заложение повторных слоев феллогена в лубе приводит у древесных пород к образованию корки.
Раневые меристемы
При травмировании растений клетки основной паренхимы, расположенные рядом с поврежденным участком, дедифференцируются, т.е. приобретают способность к делению и порождают особую ткань – раневую, или травматическую, меристему, из клеток которой сначала образуется каллюс. Таким образом, эта меристема является вторичной по происхождению. При пропитывании клеточных оболочек каллюса суберином формируется пробка. Каллюс и пробка защищают травмированный участок от поражения патогенами.
Спорогенные ткани
Назначение спорогенных тканей заключается в образовании спор. У цветковых растений они развиваются при формировании цветка в теках пыльников и семязачатках пестиков. В развитых пыльниках спорогенная ткань является многоклеточной, а в семязачатке представлена лишь несколькими клетками. Клетки крупные, тонкостенные, одноядерные, с диплоидным набором хромосом, большим запасом питательных веществ в цитоплазме.
В пыльниках из каждой спорогенной клетки в результате мейоза образуется четыре гаплоидные микроспоры. При последующем митотическом делении из микроспоры образуется пыльца. В семязачатке у большинства покрытосеменных мейотическое деление спорогенной клетки приводит к образованию четырех неравноценных клеток. Из них одна развивается в гаплоидную макроспору, остальные – отмирают. Последующее митотическое деления макроспоры является основой образования зародышевого мешка.
biofile.ru
Образовательные ткани - Википедия
Меристе́мы (с др.-греч. — «μεριστός» — делимый), или образовательные ткани, или меристематические ткани, — обобщающее название для тканей растений, состоящих из интенсивно делящихся и сохраняющих физиологическую активность на протяжении всей жизни клеток, обеспечивающих непрерывное нарастание массы растения и предоставляющих материал для образования различных специализированных тканей (проводящих, механических и т. п.)[1].
Особенности клеток меристем[ | ]
Эумеристемы, дающие начало всем тканям растения, кроме проводящих, состоят из изодиаметрических, в очертании многоугольных клеток. Каждая клетка эумеристемы имеет крупное ядро, занимающее около половины объёма клетки, а ядерная оболочка имеет много пор. В цитоплазме много диффузно расположенных рибосом. Имеются пропластиды, митохондрии и . Вакуоли немногочисленные и мелкие. Соседние клетки соединены более или менее диффузно расположенными плазмодесмами[2].
Клетки меристем, образующих проводящие ткани (камбий и прокамбий), прозенхимные, снабжены крупными вакуолями[2].
Клеткам меристем свойственно неравное (дифференцированное) деление. Клетка делится митозом на 2: одна остаётся клеткой меристемы, а другая, поделившись один или несколько раз, приступает к дифференцировке[3].
Митотическая активность клеток меристемы различна. Наиболее активные в этом отношении клетки — инициальные клетки, дающие начало производным клеткам. Производные могут отличаться от инициальных формой, размерами, количеством вакуолей.
Увеличение общего объёма меристемы может быть следствием только инициальных клеток (так, например, нарастают талломы некоторых бурых водорослей), либо чередованием делений инициальной клетки и производных, причём большее количество делений приходится на производные клетки. Такая периодичность свойственна конусам нарастания большинства высших споровых. У семенных же инициальная клетка и производные морфологически неразличимы[4].
Классификация меристем и их производных[ | ]
Морфологическая классификация[ | ]
Морфологическая классификация основана на ориентации перегородок в делящихся клетках. Выделяется три морфологических типа меристем:
- Пластинчатые меристемы однослойные, клетки, таблитчатые на поперечном срезе, делятся антиклинально (перпендикулярно плоскости органа). Участвует в образовании эпидермы.
- Колончатые, или стержневые меристемы состоят из кубических или призматических клеток, располагающихся продольными рядами. У некоторых растений такие меристемы образуют сердцевину стебля.
- Массивные меристемы состоят из многоугольных в очертании клеток, делящихся в разных направлениях и обеспечивающих более или менее равномерное увеличение объёма ткани. Из массивных меристем развивается, например, спорогенная ткань спорангия[4].
Онтогенетическая классификация[ | ]
Все меристемы, функционирующие на протяжении всей жизни растения, можно подразделить на общие и специальные. К общим меристемам относится меристема зародыша, из которой развиваются апикальные (верхушечные) меристемы, а они, в свою очередь, дают начало третьей группе общих меристем — интеркалярным.
От апикальных меристем берут начало:
Клетки этих трёх тканей более вакуолизированы, чем апикальные меристемы, и обладают ограниченной способностью к делению, поэтому часто их называют полумеристемами.
Часть клеток прокамбия даёт начало первичным проводящим тканям — первичным ксилеме и флоэме; из других его клеток развивается камбий, который, в свою очередь, образует вторичные проводящие ткани — вторичные ксилему и флоэму[5]. Вторичные проводящие ткани образует и т. н. добавочный камбий, развивающийся не из прокамбия, а из постоянных тканей, живые клетки которых при этом дедифференцируются, то есть возвращаются из специализированного состояния к состоянию, близкому к эмбриональному, и вновь приобретают способность к делению[6].
Таким же образом в растении возникают и раневые меристемы, восстанавливающие повреждённые участки растения. На возможности новообразования меристем основано вегетативное размножение растений[3].
Кроме добавочного камбия, постоянные ткани также участвуют в образовании феллогена (пробкового камбия). Делясь периклинально (то есть параллельно поверхности органа), клетки феллогена отделяют наружу будущие клетки пробки (феллемы), а внутрь — феллодерму[6].
Топографическая классификация[ | ]
В соответствии с расположением, помимо апикальных меристем, локализованных на кончиках осевых органов — стебля и корня, выделяют меристемы:
- латеральные (боковые) меристемы расположены внутри осевых органов и осуществляют их утолщение. К латеральным меристемам относят камбий, добавочный камбий и феллоген.
- интеркалярные меристемы осуществляют вставочный рост. За счёт их деятельности удлиняются междоузлия на ранних этапах развития побега, развиваются черешки листьев. Принципиально они отличаются от прочих меристем тем, что, помимо недифференцированных клеток, в них существуют и клетки на разных стадиях дифференциации. Поэтому корректнее здесь говорить не об интеркалярных меристемах, а о зонах интеркалярного (вставочного) роста[6].
Все эти меристемы, кроме феллогена, являются производными апикальных меристем.
Генетическая классификация[ | ]
Согласно генетической классификации выделяют первичные и вторичные меристемы. Первичные меристемы связаны с меристемами зародыша и апикальными меристемами, а вторичные развиваются позднее, когда начинается утолщение осевых органов и заканчивается верхушечный рост. Однако такое разделение условно, так как, например, камбий закладывается до окончания верхушечного роста[6].
Апикальная меристема[ | ]
 Модель апикальной меристемы «туника-корпус». Эпидермальный (L1) и субэпидермальный (L2) слои формируют наружный слой, называемый туникой. Внутренний слой (L3) называется корпусом. Клетки слоёв L1 и L2 делятся в боковом направлении, благодаря чему эти слои не сливаются, в то время как клетки слоя L3 делятся в разных направлениях
Модель апикальной меристемы «туника-корпус». Эпидермальный (L1) и субэпидермальный (L2) слои формируют наружный слой, называемый туникой. Внутренний слой (L3) называется корпусом. Клетки слоёв L1 и L2 делятся в боковом направлении, благодаря чему эти слои не сливаются, в то время как клетки слоя L3 делятся в разных направлениях Развивающийся зародыш любого растения состоит из эумеристемы. По мере формирования зачатков вегетативных органов у зародыша эумеристема остаётся только в двух местах: в верхушке зачаточного побега (будущий конус нарастания). Эти меристемы и называют апикальными меристемами (от лат. apex — вершина)[4]. Они осуществляют рост побегов и корней в длину и дают начало большинству других меристем.
Когда растение вступает в фазу цветения, верхушечная меристема стебля преобразуется в меристему соцветия. От неё берёт начало флоральная меристема, образующая структуры цветка: лепестки, чашелистики, плодолистики и тычинки.
Примечания[ | ]
- ↑ Меристемы — статья из Большой советской энциклопедии. И. С. Михайловская.
- ↑ 1 2 Лотова, 2010, с. 46.
- ↑ 1 2 Лотова, 2010, с. 47.
- ↑ 1 2 3 Лотова, 2010, с. 48.
- ↑ Лотова, 2010, с. 50.
- ↑ 1 2 3 4 Лотова, 2010, с. 49.
Литература[ | ]
- Лотова Л. И. Ботаника: Морфология и анатомия высших растений. — Изд. 4-е, доп.. — М.: Книжный дом «ЛИБРОКОМ», 2010. — 512 с. — ISBN 978-5-397-01047-4.
- Эсау К. Анатомия растений, пер. с англ.. — М., 1969.
- Lehrbuch der Botanik für Hochschulen, 30 Aufl.. — Jena, 1971.
encyclopaedia.bid
Ткани растений и их виды
У первых организмов Земли все клетки были почти одинаковыми. Позднее появились водоросли, мхи, хвощи, папоротники. У этих растений клетки имеют специфическое строение. Поэтому можно достаточно точно определить, растениям какой группы они принадлежат. Однако общее строение клетки у всех растений примерно одинаково.
У высших растений клетки в результате деления образуют целые группы клеток — ткани. В них находятся клетки, сходные по своему строению и значению в жизни растения. В некоторых тканях клетки лежат очень плотно друг к другу, в других — рыхло. Промежутки, образующиеся между клетками, называют межклеточным пространством (или межклетниками). Не только клетки, но и межклетники входят в состав ткани. Рассмотрим особенности различных типов тканей растений.
Образовательная ткань состоит из клеток, которые способны делиться в течение всей жизни растения. Клетки здесь лежат очень быстро друг к другу. Благодаря делению они образуют множество новых клеток, обеспечивая тем самым рост растения в длину и толщину. Появившиеся в ходе деления образовательных тканей клетки затем преобразуются в клетки других тканей растения.
Основная ткань выполняет такие функции в организме растения, как создание и накопление веществ. Например, в основной ткани находится пигмент хлорофилл, а значит, создается органическое вещество и запасается энергия солнечного излучения. Ткань, в которой образуются (синтезируются) органические вещества, преимущественно находится в мякоти листа.
Основные ткани, специализирующиеся на запасании питательных веществ, широко представлены в семенах, а также в клубнях (картофеля), в луковицах и в других органах растений. У некоторых растений (нимфея, кубышка желтая) в основной ткани развиваются очень большие межклетники, заполненные воздухом. Такие ткани выполняют в организме функцию вентиляции, что очень важно для растений, особенно в водной среде.
Покровная ткань защищает снаружи все органы растения. Клетки покровной ткани могут быть плотно сомкнутыми между собой. Например, в кожице, которая покрывает листья и молодые побеги, эти клетки с очень тонкой прозрачной клеточной стенкой легко пропускают солнечный свет в глубь растения. В корнях и стеблях оболочки клеток покровной ткани другого вида — пробки — могут опробковевать. Покровная ткань защищает растение от высыхания, перегрева и от механических повреждений.
Проводящая ткань осуществляет передвижение растворенных питательных веществ по растению. У многих высших растений она представлена проводящими элементами (сосудами, трахеидами и ситовидными трубками). В стенках проводящих элементов есть поры и сквозные отверстия, облегчающие передвижение веществ от клетки к клетке.
Проводящая ткань образует в теле растения непрерывную разветвленную сеть, соединяющую все его органы в единую систему — от тончайших корешков до молодых побегов, почек и кончиков листа.Ученые считают, что возникновение тканей связано в истории Земли с выходом растений на сушу. Когда часть растения оказалась в воздушной среде, а другая часть (корневая) — в почве, появилась необходимость доставки воды и минеральных солей от корней к листьям, а органических веществ — от листьев к корням. Так в ходе эволюции растительного мира возникло два типа проводящих тканей — древесина и луб. По древесине (по трахеидам и сосудам) вода с растворенными минеральными веществами поднимается от корней к листьям — это водопроводящий, или восходящий, ток. По лубу (по ситовидным трубкам) образовавшиеся в зеленых листьях органические вещества поступают к корням и другим органам растения — это нисходящий ток.
Механическая ткань образована клетками с очень прочными клеточными стенками. Благодаря ей растения могут противостоять большим механическим нагрузкам, например переносить раскачивание ствола порывами ветра.
Механическая ткань обеспечивает большую твердость некоторым органам растения. Например, скорлупа грецкого ореха столь прочная, что ее в размолотом виде используют как шлифовальный порошок для очистки металлических и каменных поверхностей. В стеблях некоторых растений (лен, крапива) механическая ткань состоит из длинных частично одревесневших клеток. Они крепче стали. Сталь при многократных изгибах ломается, а стебли растений способны тысячекратно в течение жизни раскачиваться, сгибаться, каждый раз выпрямляться и не ломаться.
Таким образом, ткани растений выполняют различные функции, они тесно взаимодействуют друг с другом, обеспечивая существование и развитие организма.
Разнообразие тканей обусловлено их различными функциями и особенностями клеток, входящих в них.
Клетки образуют ткани, которые формируют тело растения, придают ему упругость, формируют защитные внешние покровы, обеспечивают поступление в организм питательных веществ, их передвижение и хранение. Клетки благодаря способности образовывать ткани объединены в одно целое — организм. Жизнедеятельность организма полностью зависит от жизнедеятельности его клеток.
blgy.ru










