Содержание
Корень. Строение и функции — презентация онлайн
Похожие презентации:
Эндокринная система
Анатомо — физиологические особенности сердечно — сосудистой системы детей
Хронический панкреатит
Топографическая анатомия верхних конечностей
Анатомия и физиология сердца
Мышцы головы и шеи
Эхинококкоз человека
Черепно-мозговые нервы
Анатомия и физиология печени
Топографическая анатомия и оперативная хирургия таза и промежности
кроссворд :
?
1.
?
2
?
3
4
?
?
5
6
?
1. Молодое растеньице, появившееся при прорастании семени.
2. Состоит из стебелька, почечки, корешка, семядолей.
3. Они обуславливают зеленый цвет растений.
4. Орган размножения и расселения растений.
5. Промежуток между клетками
6. Группа клеток, сходных по строению и выполняемым функциям.
п
р
м
о
е
р
ж
о
с
т
о
к
з
а
р
о
д
ы
ш
х
л
о
р
о
п
л
с
е
м
я
и
к
к
л
е
т
н
т
к
а
н
ь
а
с
т
ы
1. Молодое растеньице, появившееся при прорастании семени.
Молодое растеньице, появившееся при прорастании семени.
2. Состоит из стебелька, почечки, корешка, семядолей.
3. Они обуславливают зеленый цвет растений.
4. Орган размножения и расселения растений.
5. Промежуток между клетками
6. Группа клеток, сходных по строению и выполняемым функциям.
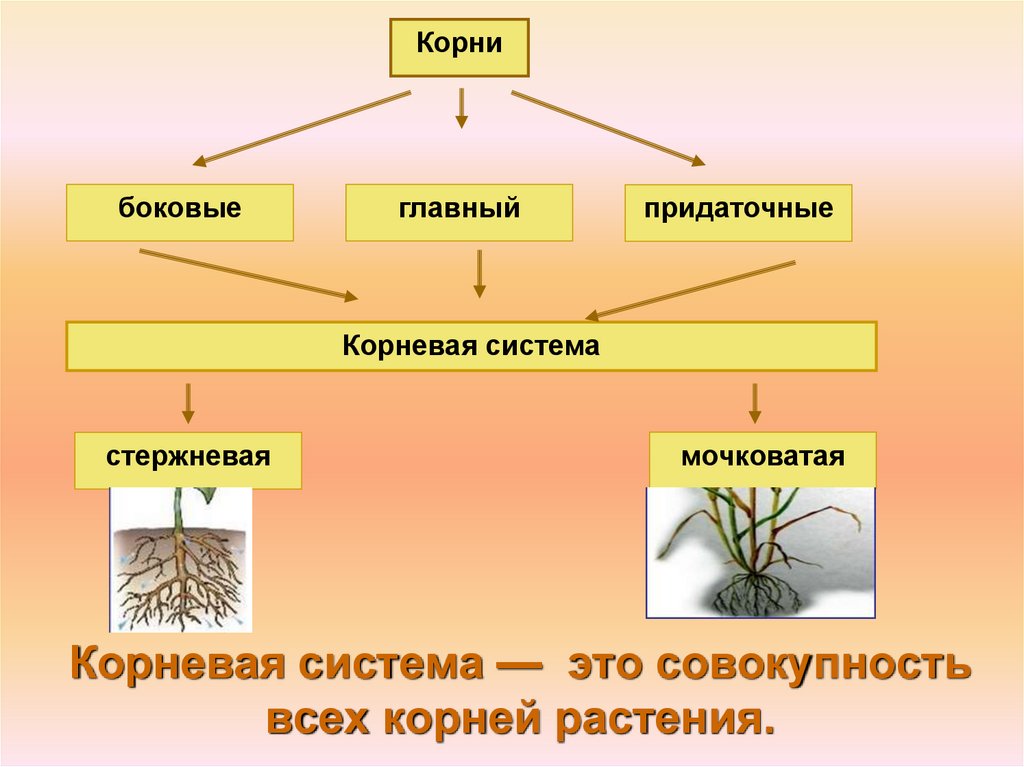
3. Корень. Строение и функции
Корень – вегетативный орган
растения, выполняющий функцию
почвенного питания
Придаточные
— развиваются
на побеге
Главный — развивается из
зародышевого
корешка семени
Боковые — развиваются
на главном
корне
Все корни одного растения образуют
корневую систему
5. Корневая система — это совокупность всех корней растения.
Корни
боковые
главный
придаточные
Корневая система
стержневая
мочковатая
Корневая система — это совокупность
всех корней растения.
6. Типы корневых систем
Стержневая
корневая система
Мочковатая
корневая система
1- придаточные корни
2 – главный корень
3- боковые корни
Рассмотрите рисунок.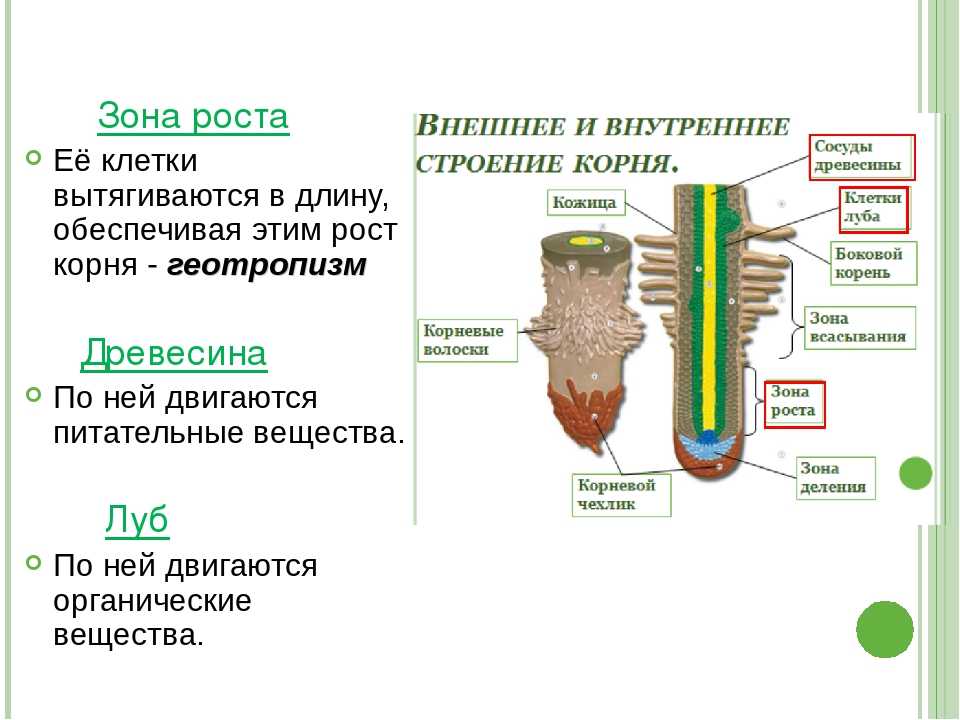
Какие корневые системы изображены под буквами
а — д растений?
8. По выполняемым функциям корень делится на зоны:
9. Корневой чехлик
• Точка роста корня, не относится к зонам
корня
• Верхняя часть корня, образует конус
нарастания
10. Зона деления
• Состоит из образовательной ткани
• Клетки интенсивно делятся,
обеспечивая рост корня в длину
11. Зона роста
• Вновь образованные клетки растут и
увеличиваются в размерах
• Происходит постепенное
формирование проводящей ткани
12. Зона всасывания
• Покрыта большим количеством
волосков, обеспечивающих всасывание
веществ
13. Зона проведения
• Обеспечивает проведение веществ ко
всем частям растения
• Имеется много боковых корней,
образуются восходящие (древесина) и
нисходящие (луб) пути проведения
веществ
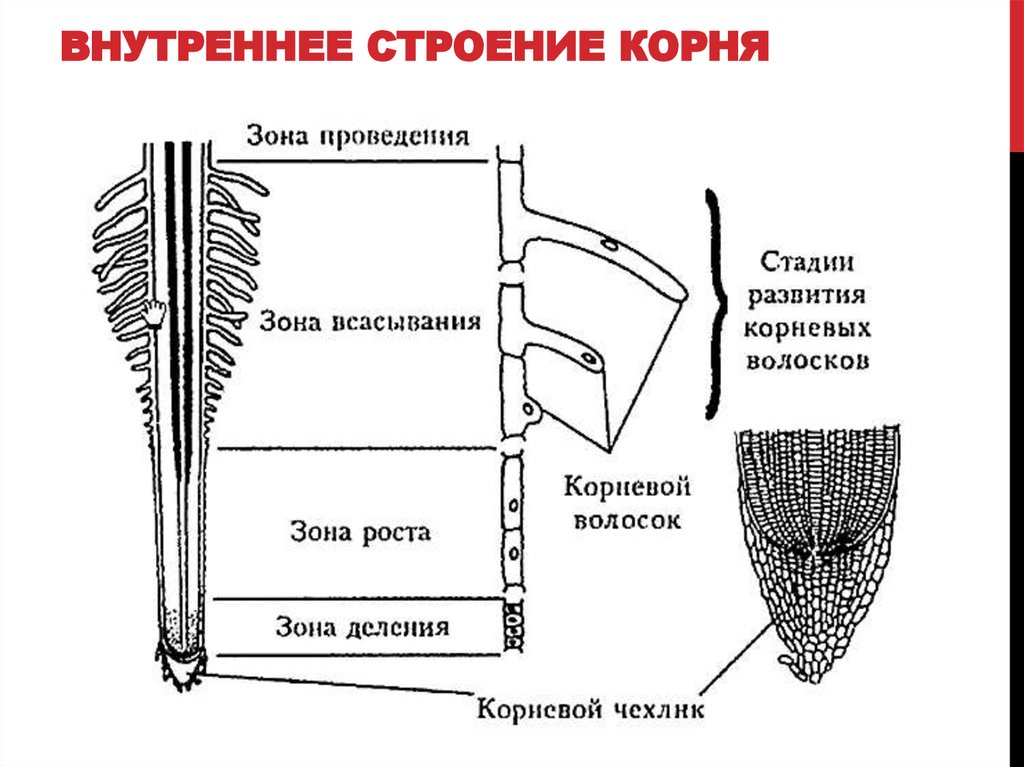
14. Внутреннее строение корня
зона корня
функция
тип ткани
Корневой
чехлик
защита зоны деления
покровная ткань
Зона
деления
деление клеток
первичная
образовательная ткань
зона роста
клетки быстро растут в длину
вторичная
образовательная ткань
Зона
всасывания
поглощение воды и мин. веществ покровная ткань
веществ покровная ткань
Зона
проведения
передвижение веществ
Вывод:
проводящая ткань
Тесное взаимодействие всех зон корня обеспечивает его
бесперебойную, нормальную работу, важную для
жизнедеятельности
Почвенное питание
-поглощение воды
с питательными
веществами
Укрепление
в почве
Запасание
питательных
веществ
Вегетативное
размножение
17. Видоизменения корней
корнеплоды
Корневые шишки (корневые клубни)
клубеньки на корнях
КОРНЕПЛОД – утолщенный
главный корень, в котором
откладываются запасные
питательные вещества
КОРНЕВЫЕ ШИШКИ –
утолщенные боковые и
придаточные корни в
мочковатой корневой системе
Дыхательные корни мангровых деревьев
Воздушные корни орхидеи
Корни присоски плюща
Корни подпорки баньяна
19. Вывод:
Разнообразие корней
– результат
видоизменений
главного, боковых и
придаточных корней.
20. Лабораторная работа №2 «Строение корня проростка»
21.
 Рост проростка
Рост проростка
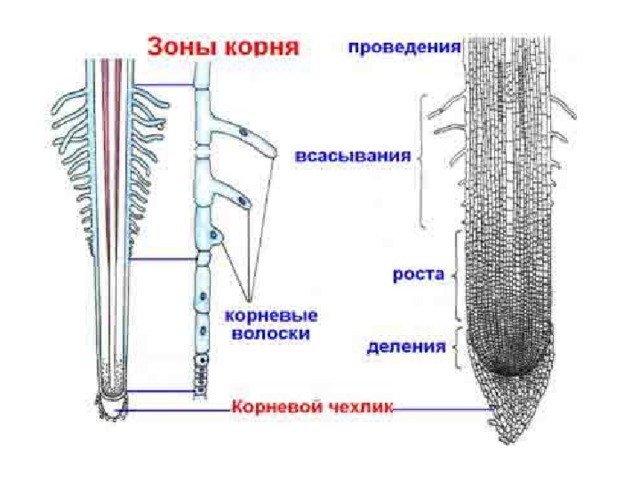
22. Зоны корня
зона проведения
зона всасывания
зона роста
корневой чехлик
English
Русский
Правила
Корень. Строение и функции — презентация онлайн
Царство Растения. Морфология и анатомия
растений.
Тема:
«Корень. Строение и
функции.»
Задачи:
o Изучить внешнее и внутреннее строение
o Дать характеристику основным функциям
o Рассмотреть видоизменения корня
https://vk.com/exam_biology
Внешнее строение корня
Корень — осевой орган, обладающий способностью к
неограниченному росту и свойством положительного геотропизма.
Функции корня:
1. Укрепление растения в почве и удержание надземной части растения;
2. Поглощение воды и минеральных веществ;
3. Проведение веществ;
4. Может служить местом накопления питательных веществ;
5. Служит органом вегетативного размножения.
Внешнее строение корня
По происхождению корни делят на главный, боковые и придаточные.
Главный корень — корень, развивающийся из зародышевого корешка.
Обладает наиболее активной верхушечной меристемой.
Придаточные корни — корни, развивающиеся от стеблей, листьев и даже
от старых корней. Появляются за счет деятельности вторичных меристем.
Боковые корни — корни, развивающиеся на другом корне любого
происхождения и являющиеся образованиями второго и последующих
порядков ветвления.
Внешнее строение корня
Корневая система — это совокупность всех корней растения.
Стержневая корневая система — корневая система с хорошо
выраженным главным корнем. Характерна для двудольных растений.
У высших споровых растений – плаунов, хвощей, папоротников
формируется мочковатая корневая система.
Мочковатая корневая система — корневая система, образованная
боковыми и придаточными корнями. Главный корень растет слабо и рано
прекращает свой рост. Типична для однодольных растений.
Олимпиадникам: вторичногоморизная корневая система? (Ответ в буфере).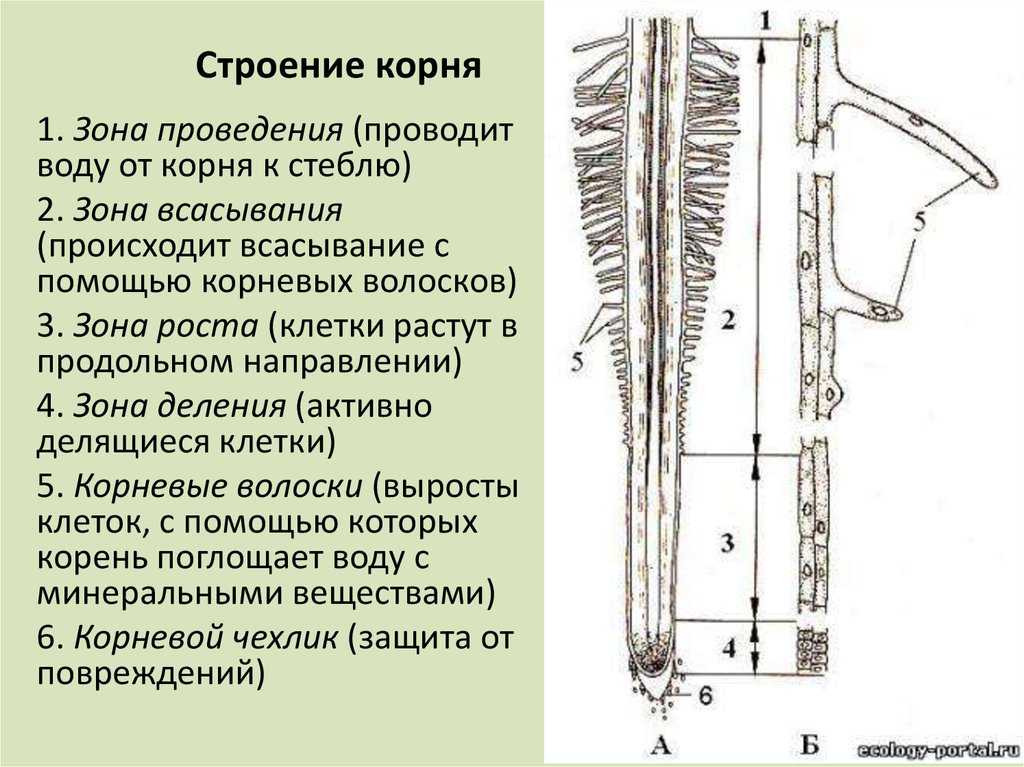
Внешнее строение корня
Зона деления. Самое окончание корня длиной
1-2 мм. Апикальная меристема корня защищена
корневым чехликом.
Зона роста, или растяжения. Протяженность
зоны — несколько миллиметров.
Зона поглощения, всасывания, или корневых
волосков. Протяженность зоны – несколько
сантиметров. Здесь уже различают наружный
слой эпиблемы (ризодермы) с корневыми
волосками, слой первичной коры и центральный
цилиндр. Корневой волосок представляет собой
боковой вырост клетки эпиблемы (ризодермы).
Почти всю клетку занимает вакуоль, окруженная
тонким слоем цитоплазмы. Вакуоль создает
высокое осмотическое давление, за счет которого
вода с растворенными солями поглощается
клеткой. Длина корневых волосков до 8 мм. В
среднем на 1 мм2 поверхности корня образуется
от 100 до 300 корневых волосков.
Внешнее строение корня
В результате суммарная
площадь зоны всасывания
больше площади поверхности
надземных органов (у растения
озимой пшеницы в 130 раз,
например).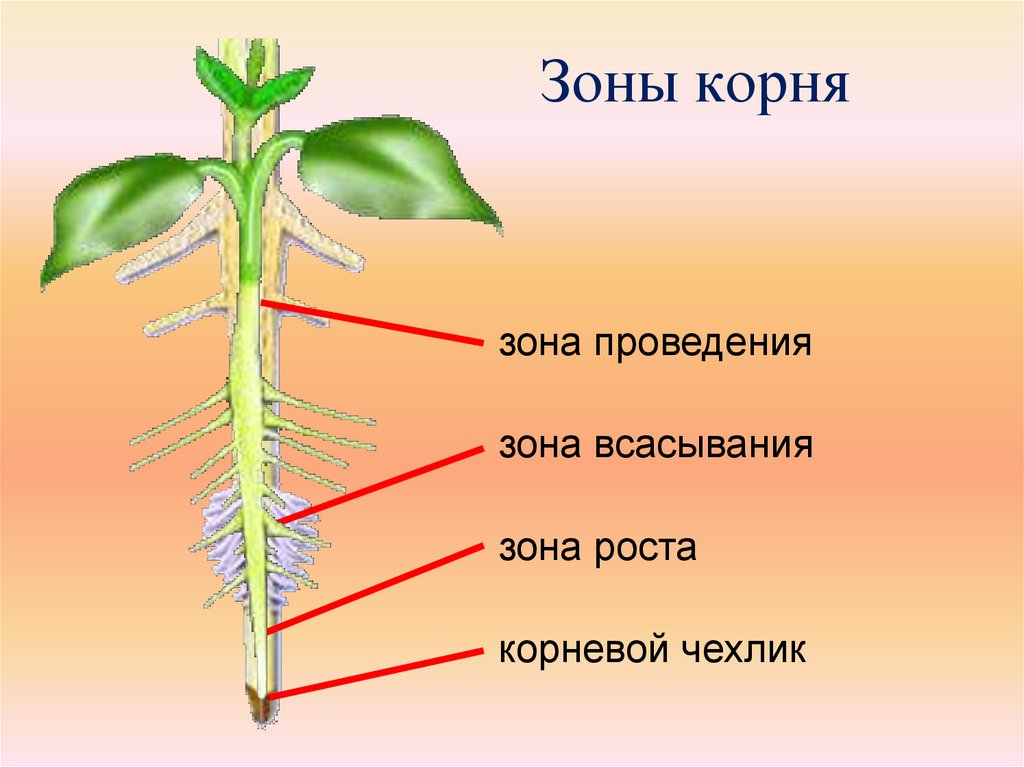 Поверхность
Поверхность
корневых волосков ослизняется
и склеивается с частицами
почвы, что облегчает
поступление воды и
минеральных веществ в
растение.
Поглощению способствует и выделение корневыми волосками кислот,
растворяющих минеральные соли.
Корневые волоски недолговечны, отмирают через 10-20 дней. На смену
отмерших (в верхней части зоны) приходят новые (в нижней части зоны).
Зона проведения.. Вода и минеральные соли передвигаются от вверх к
стеблю и листьям. Здесь же за счет образования боковых корней происходит
ветвление корня.
Внутреннее строение корня
Первичное строение корня
формируется за счет первичных
меристем, характерно для
молодых корней всех групп
растений. На поперечном срезе
корня в зоне всасывания можно
различить три части: эпиблему,
первичную кору и центральный
осевой цилиндр (стелу). У
плаунов, хвощей, папоротников и
однодольных растений
сохраняется в течение всей
жизни.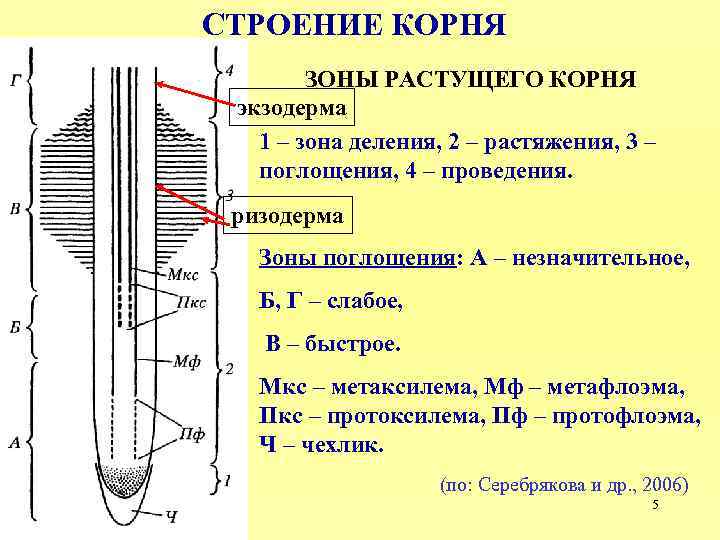
Эпиблема, или кожица – первичная покровная ткань корня. Состоит из
одного ряда плотно сомкнутых клеток, в зоне всасывания имеющих выросты –
корневые волоски, которые в верхней части зоны отмирают.
Первичная кора представлена экзодермой, мезодермой и эндодермой.
Экзодерма располагается под эпиблемой, это наружная часть первичной коры.
По мере отмирания эпиблемы оказывается на поверхности корня и в этом
случае выполняет роль покровной ткани.
Внутреннее строение корня
Под экзодермой располагается мезодерма. Здесь происходит
передвижение воды в осевой цилиндр корня, накапливаются питательные
вещества.
Внутренний слой первичной коры – эндодерма, образованная одним слоем
клеток. У двудольных растений клетки эндодермы имеют утолщения на
радиальных стенках (пояски Каспари), пропитанные непроницаемым для
воды жироподобным веществом – суберином.
Внутреннее строение корня
У однодольных растений в клетках эндодермы образуются
подковообразные утолщения клеточных стенок. Среди них встречаются
Среди них встречаются
живые тонкостенные клетки – пропускные клетки, также имеющие пояски
Каспари. Клетки эндодермы с помощью живого протопласта контролируют
поступление воды и растворенных в ней минеральных веществ из коры в
центральный цилиндр и обратно органических веществ.
Внутреннее строение корня
Центральный цилиндр, осевой цилиндр, или стела. Наружный слой стелы,
примыкающий к эндодерме, называется перицикл. Его клетки долго сохраняют
способность к делению. Здесь происходит заложение боковых корешков.
В центральной части осевого цилиндра находится сосудисто-волокнистый
пучок. Ксилема образует звезду, а между ее лучами располагается флоэма.
Перицикл – подробнее в буфере.
Внутреннее строение корня
Количество лучей ксилемы различно – от двух нескольких десятков. У
двудольных до пяти, у однодольных – пять и более пяти. В самом центре
цилиндра могут находиться элементы ксилемы или тонкостенная паренхима.
Вторичное строение корня.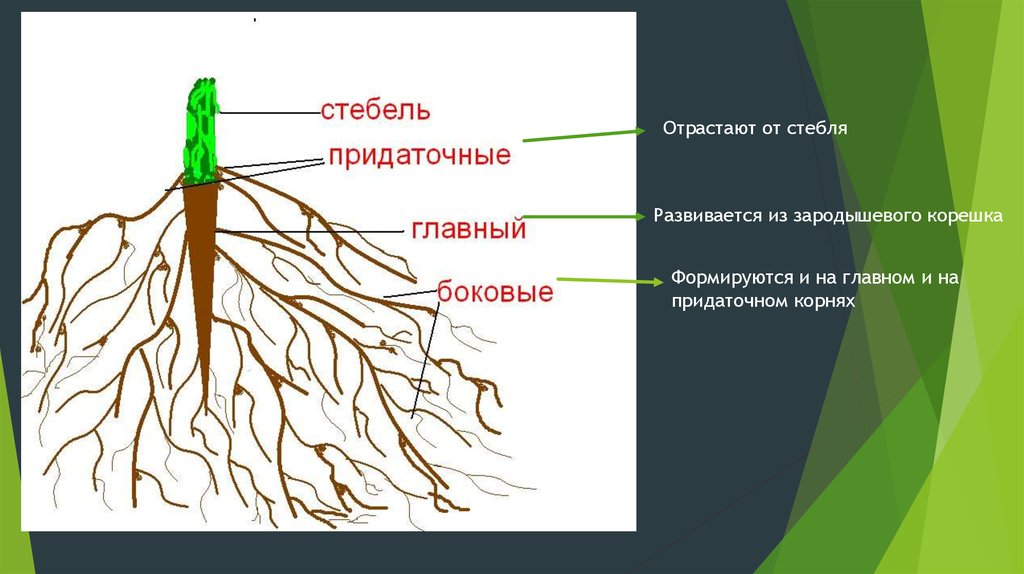 У двудольных и голосеменных растений
У двудольных и голосеменных растений
первичное строение корня сохраняется недолго. В результате деятельности
вторичных меристем формируется вторичное строение корня.
Внутреннее строение корня
Процесс вторичных изменений
начинается с появления прослоек
камбия между флоэмой и
ксилемой. Внутрь он откладывает
элементы вторичной ксилемы
(древесины), наружу элементы
вторичной флоэмы (луба). Сначала
прослойки камбия разобщены,
затем смыкаются, образуя
сплошной слой. При делении
клеток камбия исчезает
радиальная симметрия,
характерная для первичного
строения корня.
1 – эпиблема.
2 – первичная кора.
3 – перицикл.
4 – флоэма.
5 – ксилема.
6 – камбий.
Внутреннее строение корня
Внутреннее строение корня
Первичная кора слущивается. Перицикл образует феллоген, за счет
которого образуется перидерма.
Подведем итоги:
1 – эпиблема, корневые волоски
2 – первичная кора
3 – стела, осевой цилиндр
4 – пропускная клетка
5 – клетки эндодермы
6 – перицикл
7 – лучи ксилемы
8 – флоэма
9 – корневой чехлик
10 – зона деления
11 – зона роста
12 – зона всасывания
13 – зона проведения
Подведем итоги:
Микропрепараты корней каких растений на рисунках?
1
2
3
Однодольного
Двудольного
Однодольного
4
5
Двудольного
Однодольного
Подведем итоги:
Какой корень называется главным?
Корень, развивающийся из зародышевого корешка.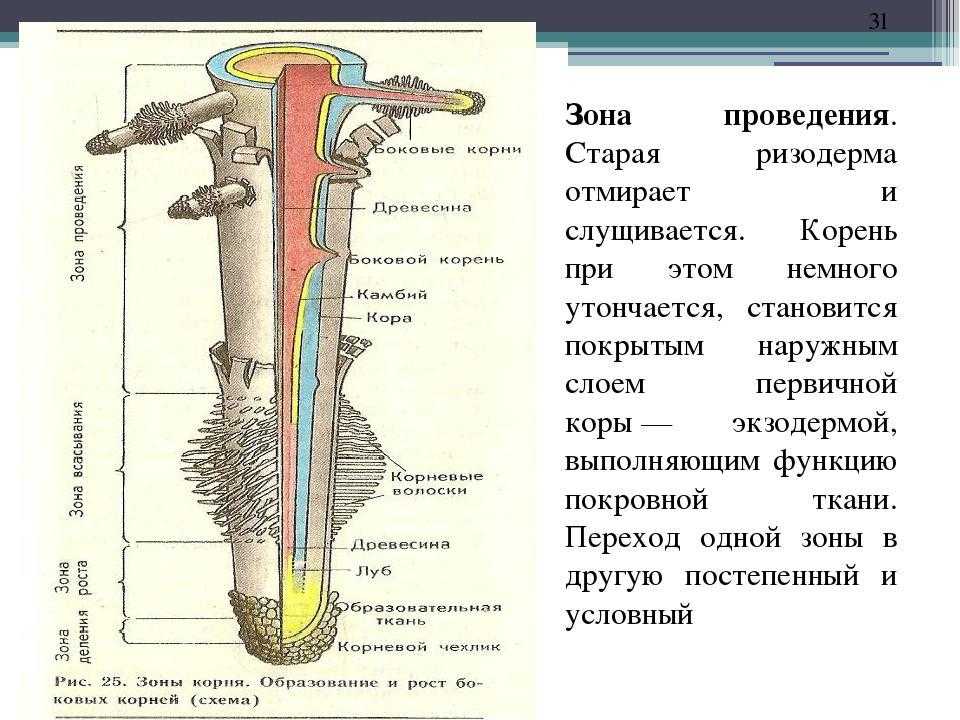
Какие корни называются придаточными?
Корни, развивающиеся от стебля или листьев.
Какие корни называются боковыми?
Корни, развивающиеся на главном или придаточных корнях.
Что такое корневая система?
Все корни одного растения.
Какие зоны различают во внешнем строении корня?
Корневой чехлик, зона деления, зона роста, зона всасывания, зона проведения.
Какие три слоя различают в первичной коре корня?
Экзодерму, мезодерму, эндодерму.
Что такое перицикл, где он находится?
Под эндодермой.
Сколько лучей ксилемы у однодольных и двудольных растений?
У однодольных 5 и более 5, у двудольных – до 5.
Чем отличается эндодерма однодольных и двудольных растений?
У двудольных в эндодерме пояски Каспари, у однодольных – клетки эндодермы
с подковообразными утолщениями и с пропускными клетками.
Подведем итоги:
1 – эпиблема.
2 – первичная кора.
3 – перицикл.
4 – флоэма.
5 – ксилема.
6 – камбий.
Олимпиадникам:
Олимпиадникам:
Олимпиадникам:
1 – перицикл;
2 – метаксилема;
3 – флоэма;
4 – эндодерма;
5 – пропускная клетка;
6 – протоксилема.
У цветковых и голосеменных первые элементы протоксилемы и протофлоэмы
закладываются экзархно, центростремительно, ближе к центру – метаксилема и
метафлоэма. В стебле будет наоборот.
Олимпиадникам:
Всероссийская олимпиада.
Экзархная протоксилема формируется непосредственно под:
а) флоэмой;
б) перициклом;
в) эндодермой;
г) пропускными клетками.
Физиология корня:
Корень обладает неограниченным ростом.
Растет он верхушкой, на которой располагается
апикальная меристема. Возьмем 3-4 дневные
проростки семян фасоли, нанесем на
развивающийся корень тушью тонкие метки на
расстоянии 1 мм друг от друга и поместим их во
влажную камеру. Через несколько дней можно
обнаружить, что расстояние между метками на
кончике корня увеличилось, в то время как в
более высоко расположенных участках корня
оно не меняется. Этот опыт доказывает
верхушечный рост корня.
Корень растет верхушкой, на которой
располагается апикальная меристема.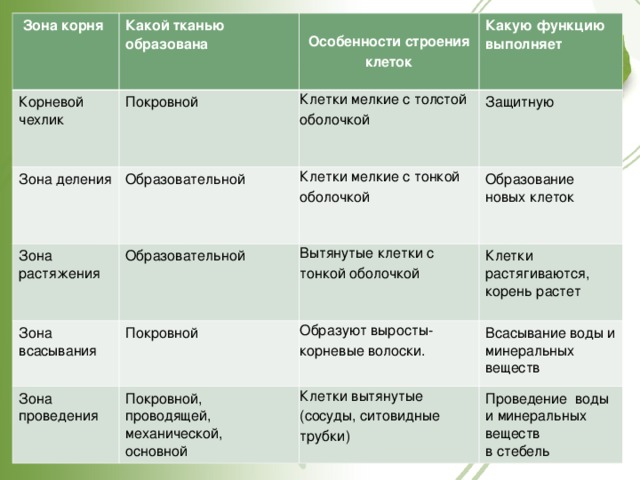
При пересадке рассады культурных растений
проводят пикировку — удаление верхушки
корня.
Для развития придаточных и боковых корней
проводят окучивание.
Физиология корня:
Поглощение из почвы и передвижение к наземным органам воды и
минеральных веществ – одна из важнейших функций корня. Эта функция
возникла у растений в связи с выходом на сушу. Вода попадает в тело
растения благодаря наличию корневых волосков. В этой зоне в стеле корня
формируется проводящая система корня – ксилема, необходимая для
обеспечения восходящего тока воды и минеральных веществ.
Вода поступает в растение в основном по закону осмоса. Корневые волоски
имеют огромную вакуоль, обладающую большим осмотическим потенциалом,
который обеспечивает поступление воды из почвенного раствора в корневой
волосок.
Физиология корня:
В корне горизонтальное движение
воды и минеральных веществ
осуществляется в следующем порядке:
корневой волосок, клетки первичной
коры, клетки стелы – перицикл,
паренхима осевого цилиндра, сосуды
корня.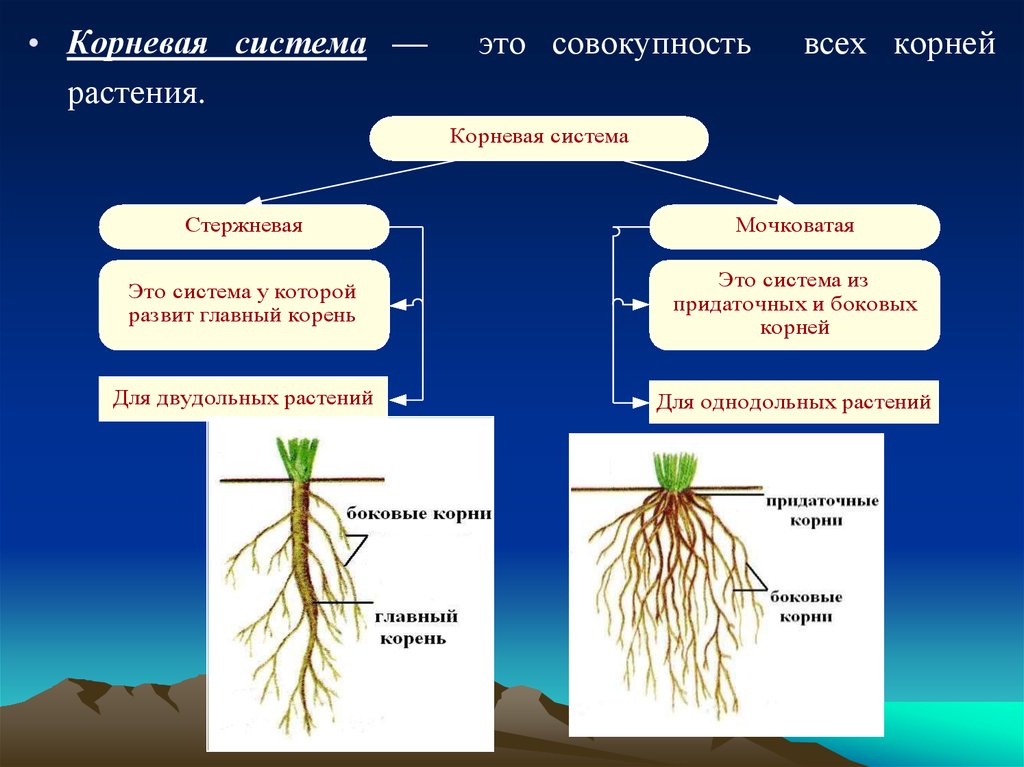
Горизонтальный транспорт воды и
минеральных веществ происходит по
трем путям: путь через апопластный,
симпластный и вакуолярный.
Апопластный путь включает в себя
все межклеточные пространства и
клеточные стенки. Данный путь
является основным для транспорта
воды и ионов неорганических веществ.
Путь через симпласт – систему
протопластов клеток, соединенных
посредством плазмодесм. Служит для
транспортировки минеральных и
органических веществ.
Физиология корня:
В зависимости от уровня
организации процесса различают
три типа транспорта веществ в
растении: внутриклеточный,
ближний (внутри органа) и дальний
(между органами).
В корне вода передвигается по
апопласту до эндодермы. Здесь ее
дальнейшему продвижению
мешают водонепроницаемые
клеточные стенки, пропитанные
суберином (пояски Каспари).
Поэтому вода попадает в стелу по
симпласту через пропускные
клетки (вода проходит через
плазматическую мембрану под
контролем цитоплазмы пропускных
клеток эндодермы).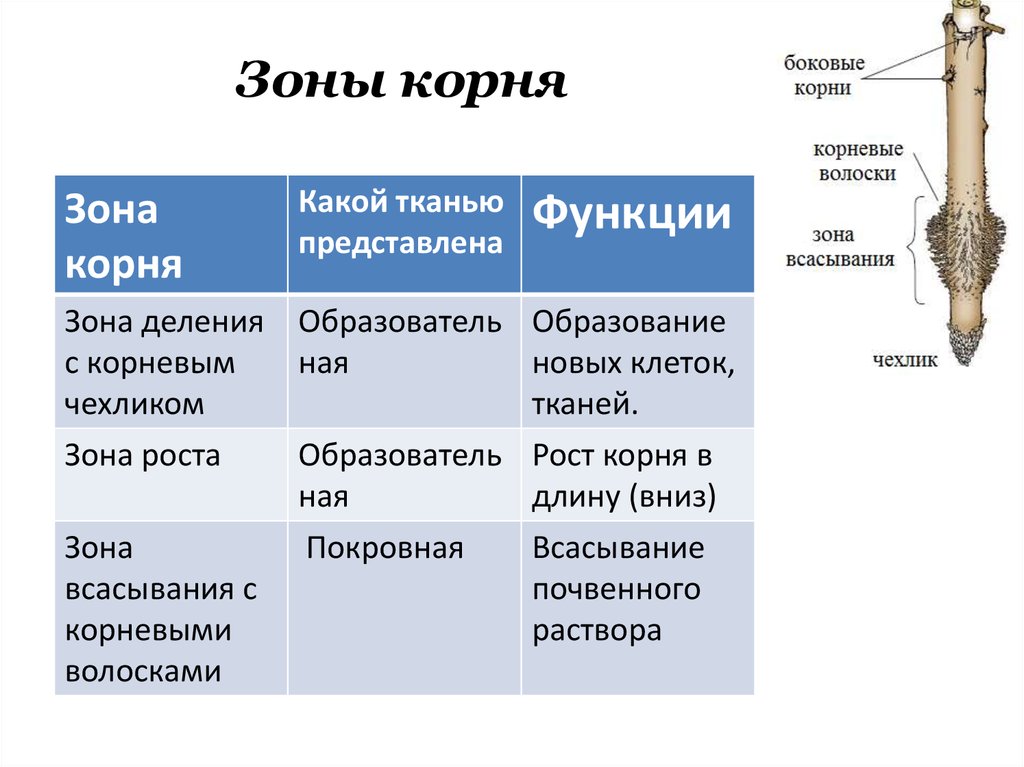 Благодаря
Благодаря
этому происходит регуляция
движения воды и минеральных
веществ из почвы в ксилему.
Физиология корня:
Корни не только поглощают
воду и минеральные вещества из
почвы, но и подают их к
надземным органам.
Вертикальное перемещение воды
происходит по мертвым клеткам,
которые не способны толкать воду
к листьям. Вертикальный
транспорт воды и растворенных
веществ обеспечивается
деятельностью самого корня и
листьев. Корень представляет
собой нижний концевой
двигатель, подающий воду в
сосуды стебля под давлением,
называемым корневым.
Физиология корня:
Корневое давление возникает главным образом
в результате повышения осмотического давления в
сосудах корня над осмотическим давлением
почвенного раствора. Оно является следствием
активного выделения клетками корня минеральных
и органических веществ в сосуды. Величина
корневого давления обычно – 1-3 атм.
Доказательство наличия корневого давления
служит выделение пасоки.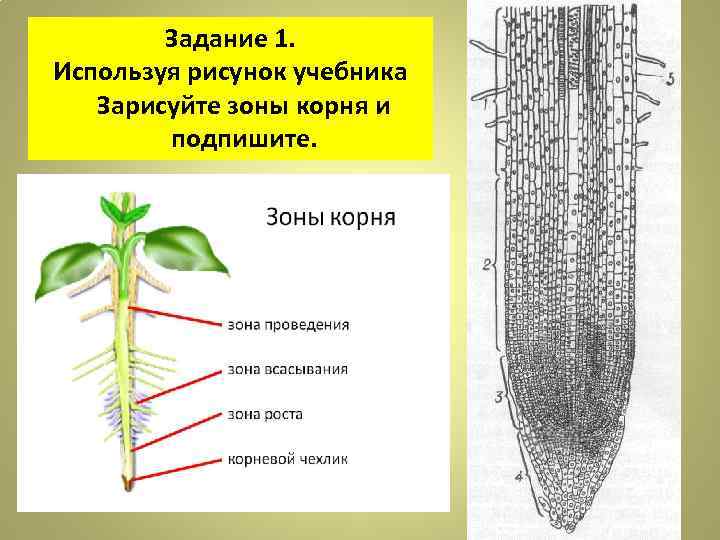
Пасока – это жидкость, которая выделяется из
перерезанного стебля.
Верхний концевой двигатель, обеспечивающий
вертикальный транспорт воды – присасывающая
сила листьев. Она возникает в результате
транспирации – испарения воды с поверхности
листьев. При непрерывном испарении воды
создается возможность для нового притока воды к
листьям. Сосущая сила листьев у деревьев может
достигать 15-20 атм. В сосудах ксилемы вода
движется в виде непрерывных водяных нитей.
Физиология корня:
При движении вверх молекулы воды сцепляются друг с другом (когезия),
что заставляет их двигаться друг за другом. Кроме того, молекулы воды
способны прилипать к стенкам сосудов (адгезия). Таким образом, поднятие
воды по растению осуществляется благодаря верхнему и нижнему двигателям
водного тока и силам сцепления молекул воды в сосудах. Основной движущей
силой является транспирация.
Видоизменения корня:
Георгин
Часто корни выполняют и другие функции, при этом возникают различные
видоизменения корней.
Запасающие корни. Среди запасающих корней различают корневые клубни и
корнеплоды. Корневые клубни образуются в результате видоизменения
боковых или придаточных корней (чистяк, ятрышник, любка, батат, георгин,
лилейник). Корнеплод образуется, в основном, в результате утолщения
главного корня, но его образовании принимает участие и стебель (цикорий,
одуванчик, женьшень, хрен, морковь, пастернак, петрушка, сельдерей, репа,
редька, редис). При этом запасающая ткань может развиваться как в ксилеме,
так и в флоэме. В утолщении главного корня может принимать участие и
перицикл, формируя добавочные камбиальные кольца (у свеклы).
Видоизменения корня:
1. Веламен
2. Первичная
кора
3. Стела
Растения, растущие на болотах, часто образуют корни, растущие вверх –
дыхательные корни, пневматофоры. В таких корнях хорошо развита
воздухоносная паренхима. Таким образом, корни болотных растений получают
достаточное количество кислорода.
Растения-эпифиты, произрастающие на других растениях высоко над
землей (но не паразитирующие на них, например, многие виды орхидей)
образуют воздушные корни, которые полностью находятся в воздухе.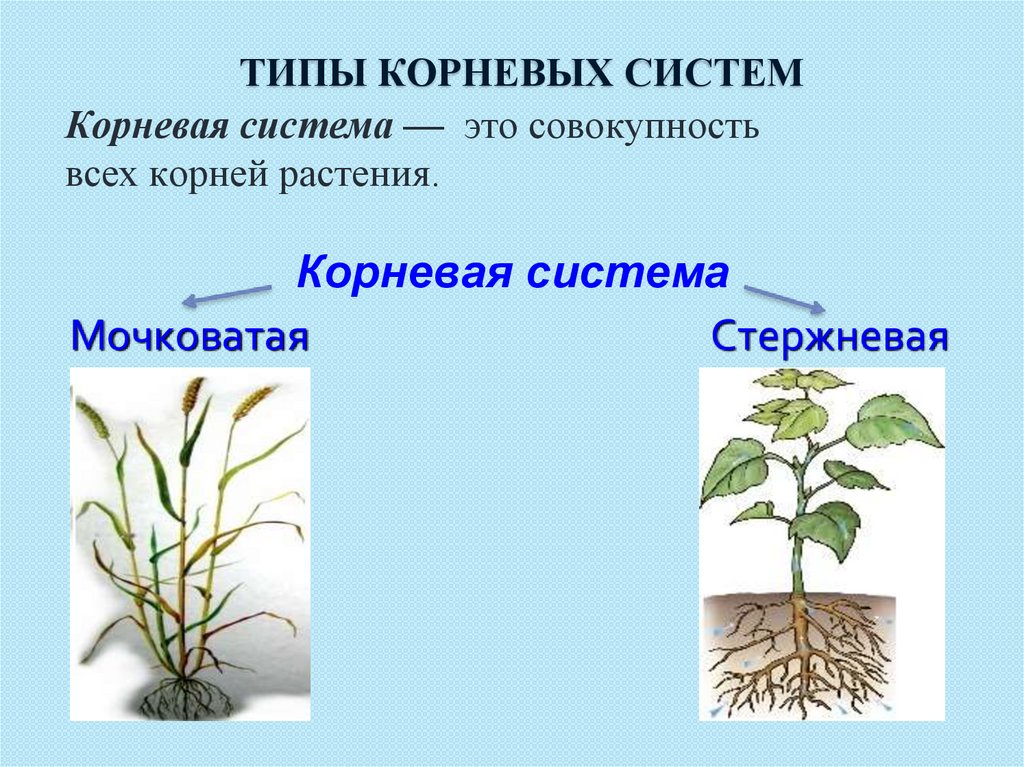 Такие
Такие
воздушные корни образуют на поверхности веламен – слой мертвых,
опробковевших клеток, образующих губчатую гигроскопическую ткань,
поглощающую влагу, находящейся в воздухе. Веламен становится емкостью,
из которой растение получает влагу в периоды между дождями.
Видоизменения корня:
У индийского дерева баньян корни, которые образуются на ветвях,
достигают земли и служат опорой ветвям, такие корни называют корнямиподпорками. У мангровых деревьев в связи с приливами и отливами
сформировались ходульные корни. Интересны досковидные корни,
выполняющие функцию опоры, корни-прицепки у плюща, с помощью которых
это растение может подниматься по вертикальной стене. Корни-присоски
растений паразитов и полупаразитов врастают в корни растения-хозяина. У
многих луковичных растений корни способны сокращаться на 10-70% от
первоначальной длины и осенью втягивать луковицу глубже в почву. Такие
втягивающие корни спасают луковицу от промерзания в зимний период.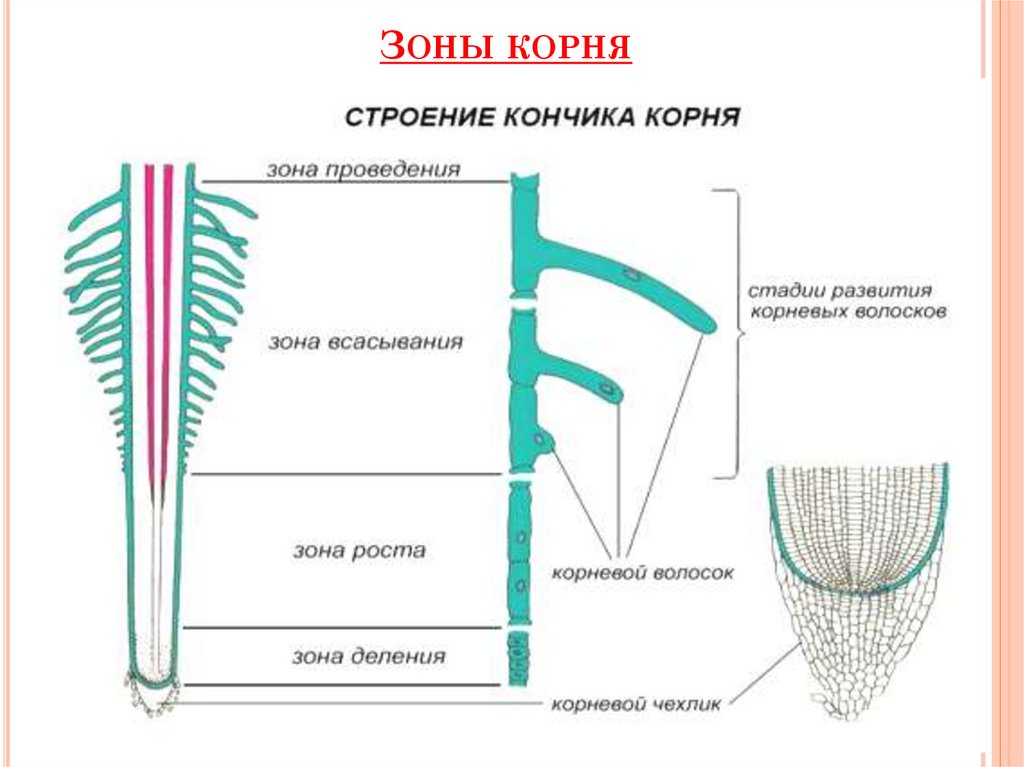
Видоизменения корня:
Видоизменения корня:
В корнях многих растений (бобовых, березовых, лоховых и др.) могут
поселяться клубеньковые бактерии, которые вызывают разрастание
клеток паренхимы и образование клубеньков. Эти бактерии – активные
азотфиксаторы, они поглощают из воздуха атмосферный азот, который
становится доступен растениям. В воздухе около 79% азота, но растения не
способны его использовать для синтеза аминокислот, азотистых основания и
поглощают азот из почвы. Растения, живущие в симбиозе с клубеньковыми
бактериями не испытывают недостатка в азоте, содержат много белка и при
отмирании обогащают почву азотом. Клевер или люцерна, например,
накапливают в клубеньках до 300 кг/га азота в год.
Удобрения:
К органическим удобрениям относят навоз, торф, птичий помет, фекалии,
компосты. Они соединяют в себе и минеральные соли и органические
вещества, постепенно образующие при разложении минеральные соединения.
К минеральным удобрениям относятся азотные, фосфорные, калийные и
другие промышленные удобрения, а из местных удобрений – зола.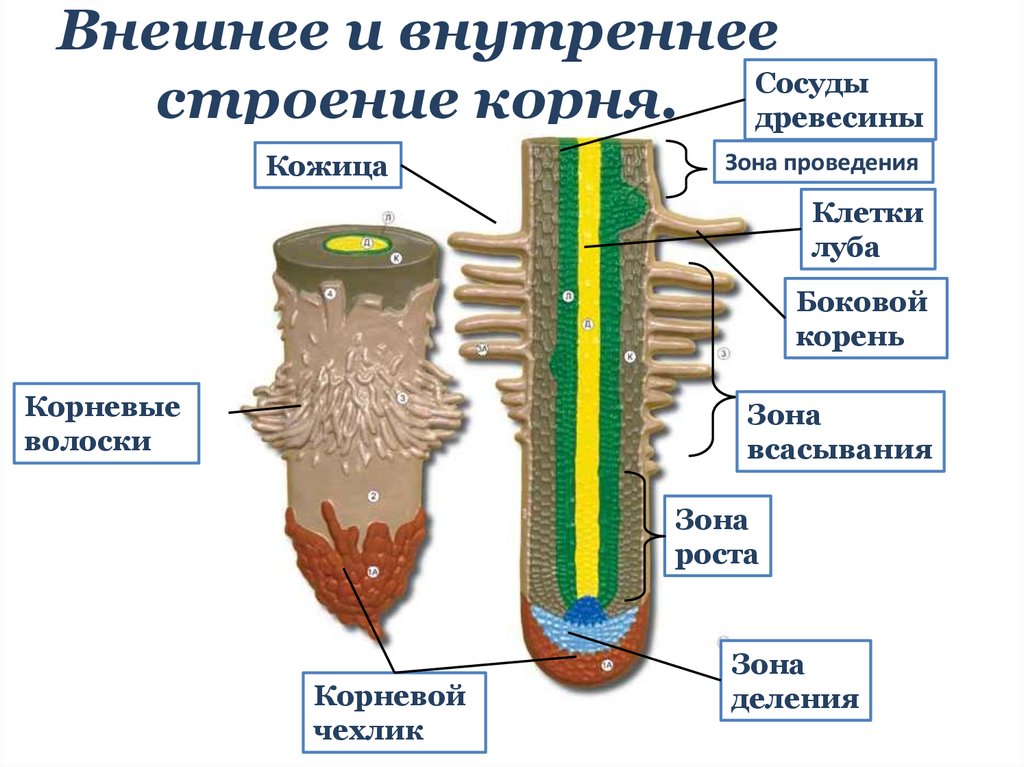
o Азотные удобрения усиливают рост стеблей и листьев.
o Фосфорные удобрения продлевают цветение, ускоряют созревание плодов.
o Калийные удобрения усиливают рост подземных органов растений корней,
луковиц, клубней.
o Фосфорные и калийные удобрения повышают холодоустойчивость
растений.
Такие элементы как бор, марганец, медь, молибден, цинк и другие,
требуются в незначительных количествах и получили название
микроэлементов, а удобрения, их содержащие – микроудобрениями.
Подведем итоги:
Для чего проводят пикировку?
Пикировка – удаление кончика корня. При этом происходит развитие
боковых корней, которые развиваются в верхнем, более плодородном слое.
Для чего проводят окучивание?
Для развития дополнительных придаточных корней.
Что такое апопластный путь?
Апопластный путь включает в себя все межклеточные пространства и
клеточные стенки. Данный путь является основным для транспорта воды и
ионов неорганических веществ.
Что такое симпластный путь? Для чего он служит?
Путь через симпласт – систему протопластов клеток, соединенных
посредством плазмодесм.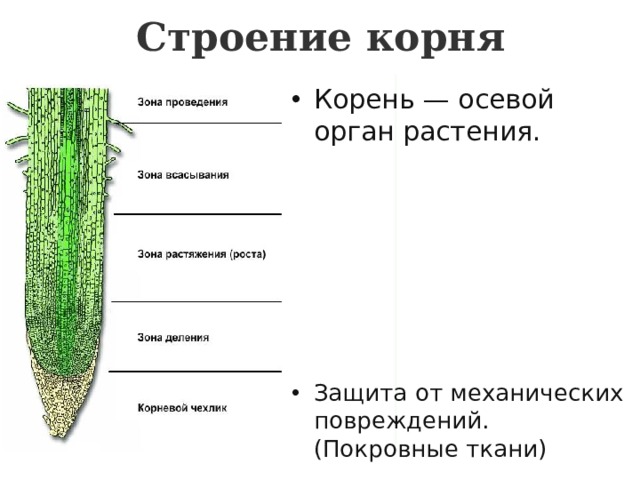 Служит для транспортировки минеральных и
Служит для транспортировки минеральных и
органических веществ.
Почему возникает корневое давление, нижний концевой двигатель?
Корневое давление возникает главным образом в результате повышения
осмотического давления в сосудах, что является следствием активного
выделения клетками корня минеральных и органических веществ в сосуды.
Величина корневого давления обычно – 1-3 атм.
Что такое верхний концевой двигатель?
Присасывающая сила листьев. Она возникает в результате транспирации
– испарения воды с поверхности листьев.
Подведем итоги:
Чем корневые клубни отличаются от корнеплодов?
Корневые клубни образуются в результате видоизменения боковых или
придаточных корней (чистяк, ятрышник, георгин). Корнеплод образуется, в
основном, в результате утолщения главного корня, но его образовании
принимает участие и стебель (морковь, редис, свекла).
Какие еще видоизменения корней известны?
Дыхательные корни болотных растений, воздушные корни с веламеном,
ходульные корни, корни-подпорки, досковидные корни, корни паразитов и
полупаразитов, корни с клубеньками.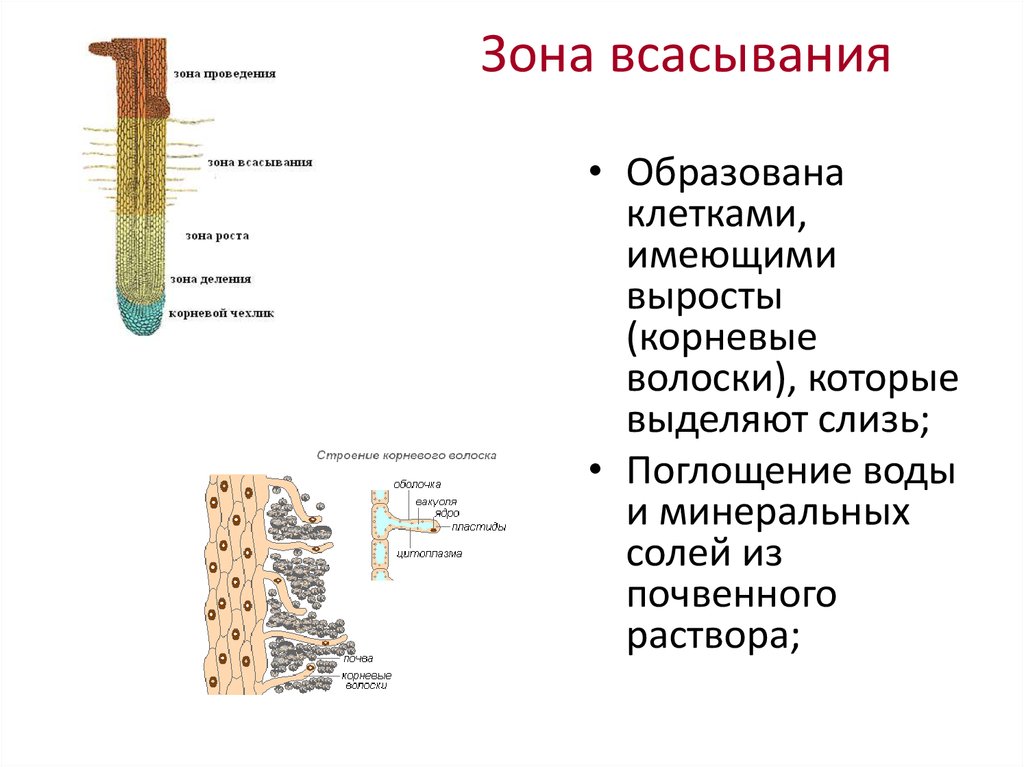
Какие удобрения усиливают рост стеблей и листьев, цветение и созревание
плодов, рост подземных органов?
Азотные удобрения усиливают рост стеблей и листьев. Фосфорные
удобрения продлевают цветение, ускоряют созревание плодов. Калийные
удобрения усиливают рост подземных органов растений корней, луковиц,
клубней.
Какие удобрения повышают холодоустойчивость растений?
Калийные и фосфорные удобрения.
Олимпиадникам:
Теория гистогенов
выдвинута немецким ботаником
Иоганном
Ганштейном (1868) и применима
только к корню. В апикальной
меристеме корня можно
выделить 3 слоя
меристематических клеток,
названных гистогенами,
расположены они ярусами друг
над другом:
Всероссийская олимпиада. Кора корня, согласно теории
Ганштейна, развивается из:
а) дерматогена;
б) плеромы;
в) периблемы;
г) всех перечисленных гистогенов.
1. Гистоген – дерматоген, при
делении инициалей которого
формируется покровная
ткань – ризодерма;
2. Гистоген – периблема,
Гистоген – периблема,
формирующая клетки
первичной коры;
3. Гистоген – плерома, дает
начало центральному
цилиндру корня.
границ | Биология корневых систем
Корни растений, которые необходимы для закрепления в почве, получения минеральных питательных веществ и воды, а также для синтеза множества метаболитов, представляют собой прекрасную модель для изучения физиологических, эволюционных и метаболических процессов на системном уровне. Сложность понимания таких процессов сравнима с расшифровкой принципа радио с помощью редукционистского подхода, то есть путем случайного удаления частей из серии идентичных радио и наблюдения за «фенотипами», получающимися в результате этой процедуры (Лазебник, 2002). Несомненно, понимание (корневой) биологии в целом представляет собой гораздо более сложную задачу, но постоянная разработка новых инструментов и алгоритмов, а также технический прогресс в технологиях омики способствуют быстрому продвижению к более интегративной, целостной картине корневой биологии. 13 статей в этой электронной книге освещают последние результаты, подходы и ресурсы в области биологии корневых систем.
13 статей в этой электронной книге освещают последние результаты, подходы и ресурсы в области биологии корневых систем.
Одной из проблем при изучении корней является их многоклеточная сложность. Qiao and Libault (2013) описывают метод, в котором ультразвуковая аэропонная система используется для создания большого количества клеток корневых волосков, что позволяет проводить однородную и долгосрочную обработку одного типа клеток различными биотическими и абиотическими стимулами для последующих функциональных геномные приложения. На развитие корневых волосков влияют факторы почвенной среды, которые максимизируют поглощающую способность и, в конечном счете, приспособленность растения. Lee и Cho (2013) резюмируют роль ауксина как ключевого игрока и организующего узла для экологической/гормональной модуляции роста корневых волосков. Ауксин также играет ключевую роль в формировании боковых корней, которые постэмбрионально инициируются из первичного корня в ответ на стимулы развития и окружающей среды, обеспечивая высокий уровень пластичности архитектуры корневой системы.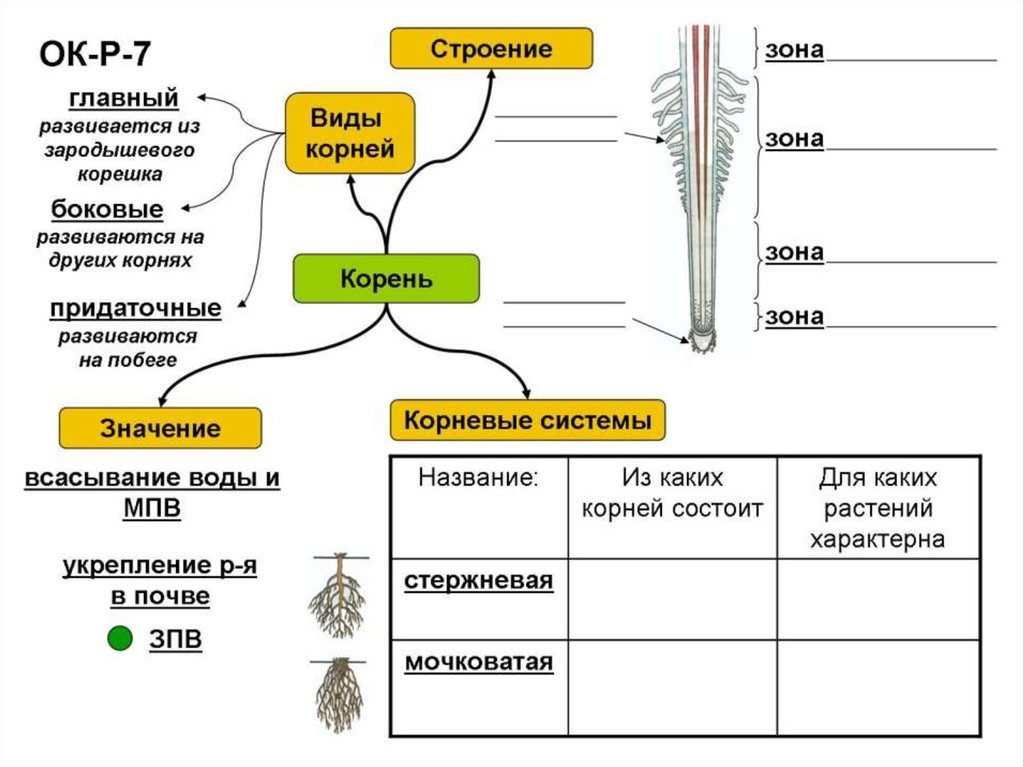 Методы визуализации нового поколения и подходы с высокой пропускной способностью, часто используемые в сочетании с компьютерным моделированием, вызвали возрождение исследований развития корней. В своей обзорной статье Cuesta et al. (2013) описывают традиционные и новые инструменты и оценивают их потенциал для решения давних вопросов об органогенезе боковых корней на качественно новом уровне.
Методы визуализации нового поколения и подходы с высокой пропускной способностью, часто используемые в сочетании с компьютерным моделированием, вызвали возрождение исследований развития корней. В своей обзорной статье Cuesta et al. (2013) описывают традиционные и новые инструменты и оценивают их потенциал для решения давних вопросов об органогенезе боковых корней на качественно новом уровне.
Структура корней тесно взаимосвязана и определяется наличием питательных веществ, в частности нитратов и фосфатов. Стратегии по расширению использования ресурсов сельскохозяйственных культур приобретают все большее значение для обеспечения устойчивого производства продуктов питания. В последнее время такие стратегии были сосредоточены на корневых признаках с целью более эффективного использования почвенных ресурсов, что облегчило бы переход от высокопроизводительного монокультурного сельского хозяйства к продуктивным, устойчивым агроэкосистемам с низкими затратами. Тиан и Дёрнер (2013) оценивают важность поиска корневых ресурсов и возможность использования естественных вариантов местных сортов или диких родственников сельскохозяйственных культур для программ селекции с целью получения культур с корневыми характеристиками, которые обеспечивают более устойчивые характеристики при воздействии экологических стрессов.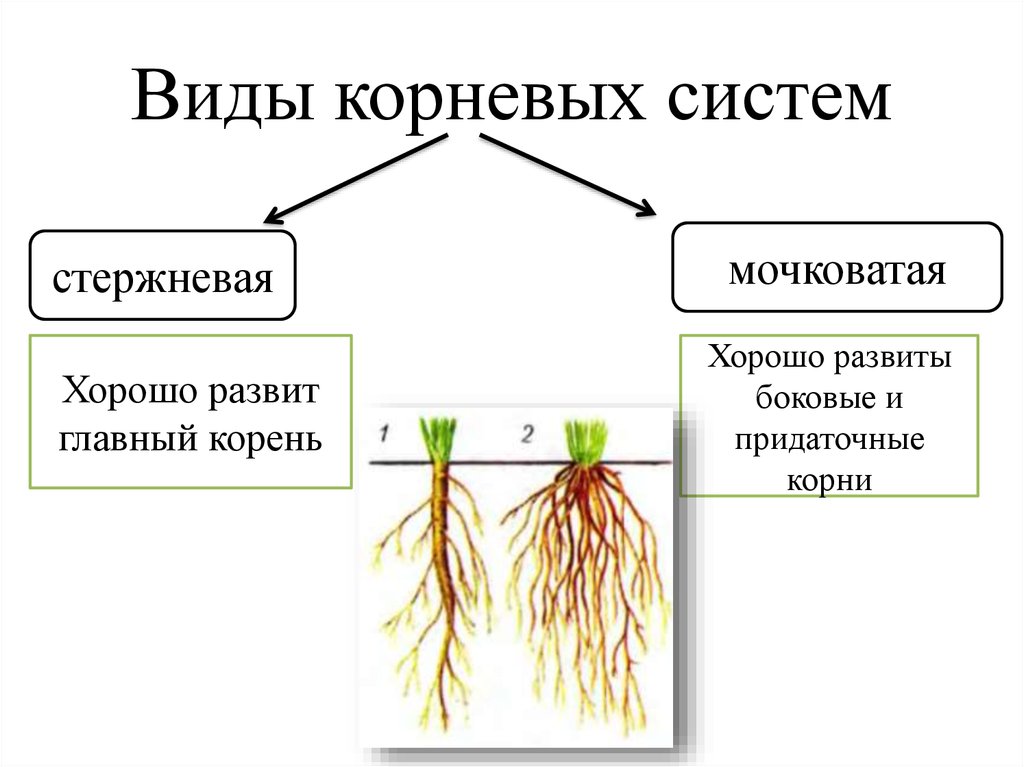 например дефицит фосфатов. Азот, в основном потребляемый в виде нитратов, является еще одним важным питательным веществом, которое сильно влияет на структуру корней и имеет решающее значение для продуктивности растений. Модулирование развития корней доступностью азота имеет большое сельскохозяйственное значение, и его понимание обеспечивает основу для создания зародышевой плазмы с улучшенной корневой архитектурой. Мохд-Радзман и др. (2013) обновляют современные знания о сигнальных компонентах, участвующих в N-опосредованной корневой архитектуре, уделяя особое внимание корневой системе бобовых. Дефицит нитратов приводит к экспрессии примерно 2000 генов, из которых только меньшинство функционально охарактеризовано. Объединив общедоступные данные микрочипов из 27 независимых наборов экспериментальных данных, связанных с нитратами, Canales et al. (2014) сгенерировали несколько кластеров генов с высокой степенью коэкспрессии с надежными функциями в транспорте нитратов, передаче сигналов и метаболизме в Корни арабидопсиса .
например дефицит фосфатов. Азот, в основном потребляемый в виде нитратов, является еще одним важным питательным веществом, которое сильно влияет на структуру корней и имеет решающее значение для продуктивности растений. Модулирование развития корней доступностью азота имеет большое сельскохозяйственное значение, и его понимание обеспечивает основу для создания зародышевой плазмы с улучшенной корневой архитектурой. Мохд-Радзман и др. (2013) обновляют современные знания о сигнальных компонентах, участвующих в N-опосредованной корневой архитектуре, уделяя особое внимание корневой системе бобовых. Дефицит нитратов приводит к экспрессии примерно 2000 генов, из которых только меньшинство функционально охарактеризовано. Объединив общедоступные данные микрочипов из 27 независимых наборов экспериментальных данных, связанных с нитратами, Canales et al. (2014) сгенерировали несколько кластеров генов с высокой степенью коэкспрессии с надежными функциями в транспорте нитратов, передаче сигналов и метаболизме в Корни арабидопсиса . В дополнение к определению приоритетов потенциально важных генов для дальнейшей функциональной характеристики, метаанализ выявил несколько предполагаемых ключевых регуляторных факторов, которые контролируют эти модули генной сети, и выявил новые процессы развития, контролируемые нитратами, такие как образование корневых волосков.
В дополнение к определению приоритетов потенциально важных генов для дальнейшей функциональной характеристики, метаанализ выявил несколько предполагаемых ключевых регуляторных факторов, которые контролируют эти модули генной сети, и выявил новые процессы развития, контролируемые нитратами, такие как образование корневых волосков.
Переходная зона корня соединяет высокочувствительную верхушку корня с зоной удлинения, в которой осуществляются ответы на стимулы окружающей среды, приводящие к изменениям в судьбе клеток и изменениям архитектуры корня. Baluška and Mancuso (2013) обсуждают специфические особенности переходной зоны и предполагают, что она действует как командная зона, которая объединяет информацию об окружающей среде, полученную от апекса, для регулирования ответов клеток в зоне растяжения. Абиотические стрессы, такие как засуха, засоление, наводнения и холод, отрицательно сказываются на росте растений и снижают урожайность сельскохозяйственных культур. Сигнатуры стрессор-специфических белков, которые диктуют адаптивные механизмы, описаны с точки зрения протеомики Ghosh and Xu (2014). Достижения в области масс-спектрометрии и фрагментации пептидов значительно улучшают охват протеомных профилей и открывают новые перспективы для анализа молекулярных механизмов, лежащих в основе адаптивных реакций на абиотические стрессы.
Достижения в области масс-спектрометрии и фрагментации пептидов значительно улучшают охват протеомных профилей и открывают новые перспективы для анализа молекулярных механизмов, лежащих в основе адаптивных реакций на абиотические стрессы.
Внутренне неупорядоченные белки не принимают складчатую структуру в своей функциональной форме, но выполняют критически важные функции в сигнальных каскадах и сетях факторов транскрипции. Благодаря присущей им конформационной гибкости неупорядоченные белки могут связываться с несколькими партнерами с высокой специфичностью и низкой аффинностью, тем самым усложняя интерактомы. Неупорядоченные белки далеко не редкие или анекдотические, они являются одними из наиболее важных белков в данном протеоме, очевидно, противореча классическим отношениям структура-функция. Пазос и др. (2013) постулируют, что нарушение белков особенно важно для сидячего образа жизни растений, предоставляя им быстрый механизм для получения сложных, взаимосвязанных и универсальных молекулярных сетей для взаимодействия с окружающей средой.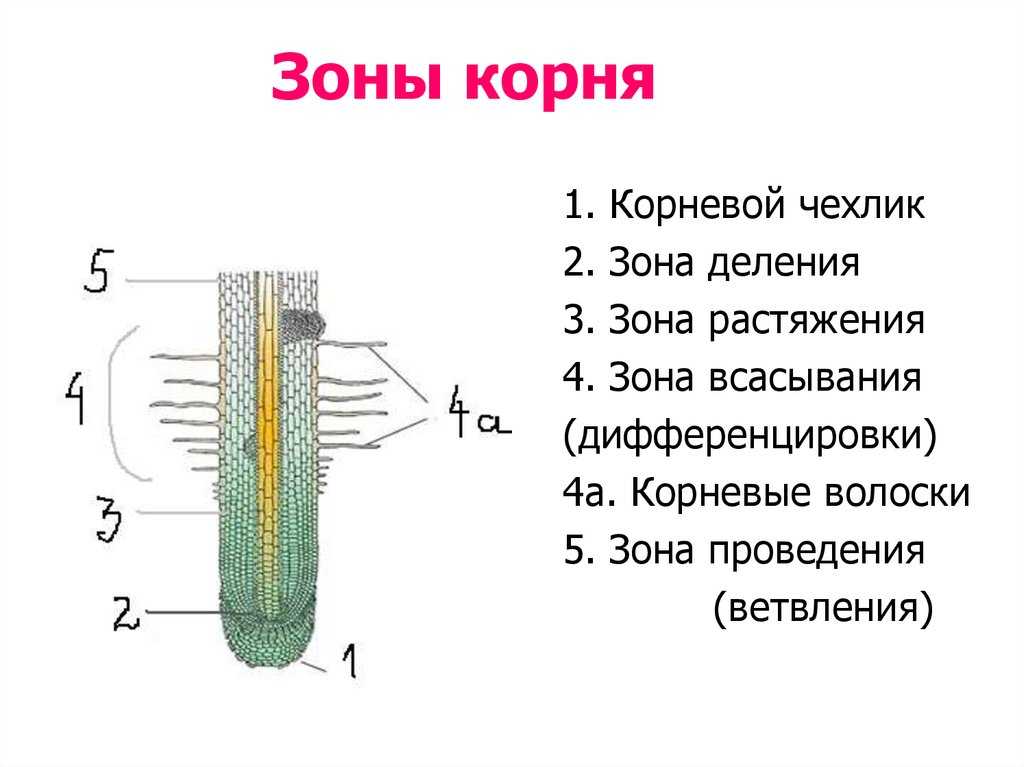
Более 7000 небольших неаннотированных открытых рамок считывания, многие из которых могут кодировать регуляторные пептиды, существуют в геноме Arabidopsis (Hanada et al., 2013). Малые сигнальные пептиды представляют собой растущий класс регуляторных молекул, которые являются частью множества сигнальных сетей, контролирующих развитие корней растений. Делэй и др. (2013) рассматривают участие регуляторных пептидов в нескольких аспектах развития корней растений, включая, помимо прочего, поддержание меристемы, гравитропную реакцию, развитие боковых корней и формирование сосудов, подчеркивая недавний скачок в нашем понимании их роли в регуляции развивающие программы.
Генетические регуляторные сети (GRN) являются отличным инструментом для интеграции и анализа сложных биомолекулярных систем на структурном и динамическом уровне. Однако большинство моделей GRN неполны, потому что в них, вероятно, отсутствуют компоненты или взаимодействия из-за отрывочных экспериментальных данных и вычислительных ограничений.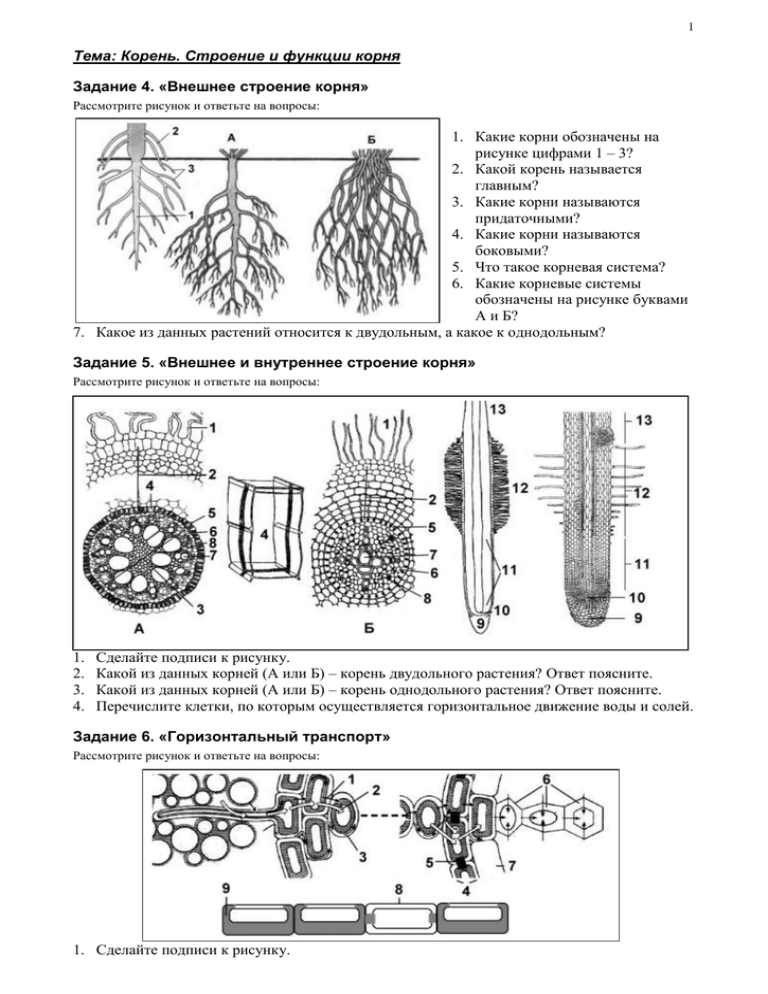 Аспейтиа и др. (2013) предлагают набор процедур для обнаружения и прогнозирования отсутствующих взаимодействий в булевых сетях и оценивают их применимость для прогнозирования предполагаемых отсутствующих взаимодействий с использованием ранее опубликованного 9Например, сеть корневых стволовых клеток 0007 Arabidopsis (Azpeitia and Alvarez-Buylla, 2012).
Аспейтиа и др. (2013) предлагают набор процедур для обнаружения и прогнозирования отсутствующих взаимодействий в булевых сетях и оценивают их применимость для прогнозирования предполагаемых отсутствующих взаимодействий с использованием ранее опубликованного 9Например, сеть корневых стволовых клеток 0007 Arabidopsis (Azpeitia and Alvarez-Buylla, 2012).
Исследования в области биологии корней значительно выиграли от конструирования растений для экспрессии многокомпонентных конструкций ДНК, таких как слияния промоторных/репортерных генов. Эмами и др. (2013) представили оптимизированный протокол для быстрого и недорогого создания многокомпонентных трансгенов на основе стратегии клонирования «Золотые ворота». Одновременный мониторинг изменений мембранного потенциала в популяциях клеток может дать количественную характеристику для оценки вместе с глобальными изменениями активности генов и уровнями метаболитов в исследованиях системной биологии. Matzke and Matzke (2013) описывают получение трансгенных растений, сконструированных для экспрессии различных версий генетически кодируемых чувствительных к напряжению флуоресцентных белков, нацеленных на плазматическую мембрану и внутренние мембраны растительных клеток. В их статье «Гипотеза и теория» описывается прогресс в адаптации технологии, первоначально использовавшейся на нервных клетках животных, для записи электрических паттернов, которые выходят за пределы отдельных клеток и систем с одной мембраной в ответ на различные стимулы в живых растениях.
В их статье «Гипотеза и теория» описывается прогресс в адаптации технологии, первоначально использовавшейся на нервных клетках животных, для записи электрических паттернов, которые выходят за пределы отдельных клеток и систем с одной мембраной в ответ на различные стимулы в живых растениях.
Заявление о конфликте интересов
Автор заявляет, что исследование проводилось в отсутствие каких-либо коммерческих или финансовых отношений, которые могли бы быть истолкованы как потенциальный конфликт интересов.
Ссылки
Аспейтиа Э. и Альварес-Буйла Э. Р. (2012). Комплексный системный подход к механизмам развития ниш корневых стволовых клеток арабидопсиса: от молекул к сетям и к морфогенезу. Завод Мол. Биол . 80, 351–363. дои: 10.1007/s11103-012-9954-6
Опубликовано Резюме | Опубликован полный текст | Полный текст CrossRef
Аспейтиа, Э., Вайнштейн, Н., Бенитес, М., Мендоса, Л., и Альварес-Буйла, Э. Р. (2013). Обнаружение отсутствующих взаимодействий регуляторной сети генов ниш корневых стволовых клеток Arabidopsis thaliana . Фронт. Завод Наука . 4:110. doi: 10.3389/fpls.2013.00110
Фронт. Завод Наука . 4:110. doi: 10.3389/fpls.2013.00110
Pubmed Abstract | Опубликован полный текст | Полный текст CrossRef
Балушка, Ф., и Манкузо, С. (2013). Переходная зона верхушки корня как колебательная зона. Фронт. Завод Наука . 4:354. doi: 10.3389/fpls.2013.00354
Pubmed Abstract | Опубликован полный текст | Полный текст CrossRef
Каналес, Дж., Мояно, Т.С., Вильярроэль, Э., и Гутьеррес, Р.А. (2014). Системный анализ данных транскриптома дает новые гипотезы о реакции корней Arabidopsis на обработку нитратами. Фронт. Завод Наука . 5:22. doi: 10.3389/fpls.2014.00022
Pubmed Abstract | Опубликован полный текст | Полный текст CrossRef
Куэста, К., Вабник, К., и Бенкова, Э. (2013). Системные подходы к изучению динамики корневой архитектуры. Фронт. Завод Наука . 4:537. doi: 10.3389/fpls.2013.00537
Pubmed Abstract | Опубликован полный текст | Полный текст CrossRef
Делей, К., Имин, Н.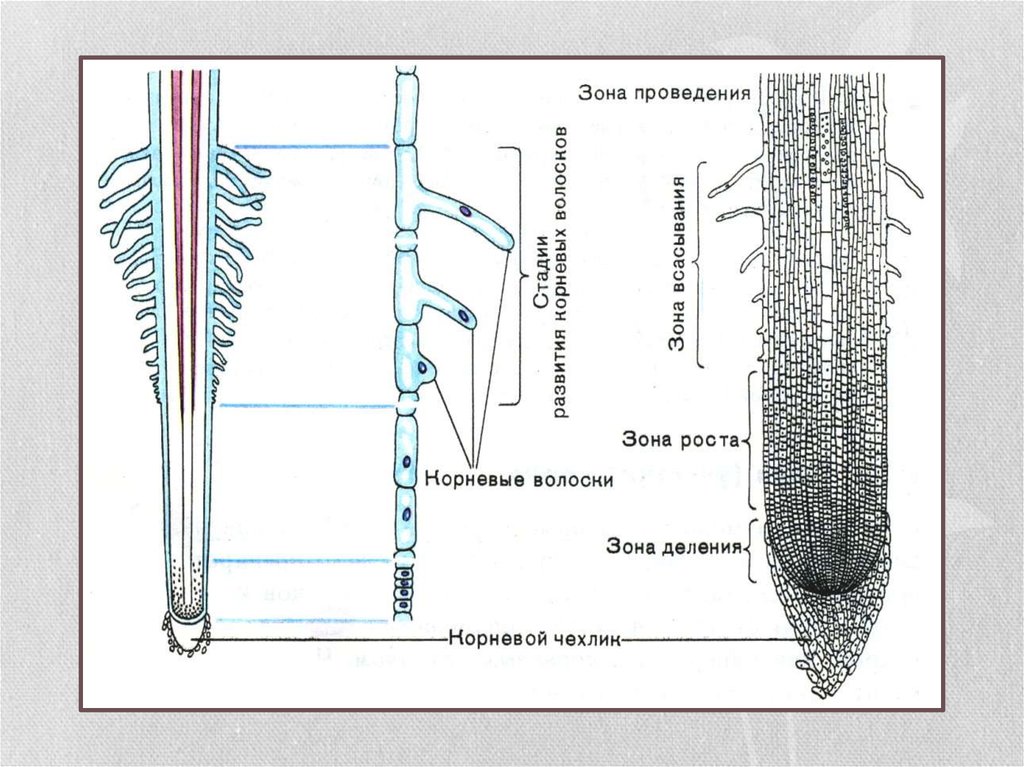 , и Джорджевич, М. А. (2013). Регуляция развития корней малыми сигнальными пептидами. Фронт. Завод Наука . 4:352. doi: 10.3389/fpls.2013.00352
, и Джорджевич, М. А. (2013). Регуляция развития корней малыми сигнальными пептидами. Фронт. Завод Наука . 4:352. doi: 10.3389/fpls.2013.00352
Pubmed Abstract | Опубликован полный текст | Полный текст CrossRef
Эмами, С., Йи, М. К., и Диннени, Дж. Р. (2013). Надежное семейство Golden Gate Agrobacterium векторов для синтетической биологии растений. Фронт. Завод Наука . 4:339. doi: 10.3389/fpls.2013.00339
Pubmed Abstract | Опубликован полный текст | Полный текст CrossRef
Гош, Д., и Сюй, Дж. (2014). Реакции на абиотический стресс в корнях растений: перспектива протеомики. Фронт. Завод Наука . 5:6. doi: 10.3389/fpls.2014.00006
Pubmed Abstract | Опубликован полный текст | CrossRef Полный текст
Ханада К., Хигучи-Такеучи М., Окамото М., Йошизуми Т., Симидзу М., Накаминами К. и др. (2013). Небольшие открытые рамки считывания, связанные с морфогенезом, скрыты в геномах растений. Проц. Натл. акад. науч. США .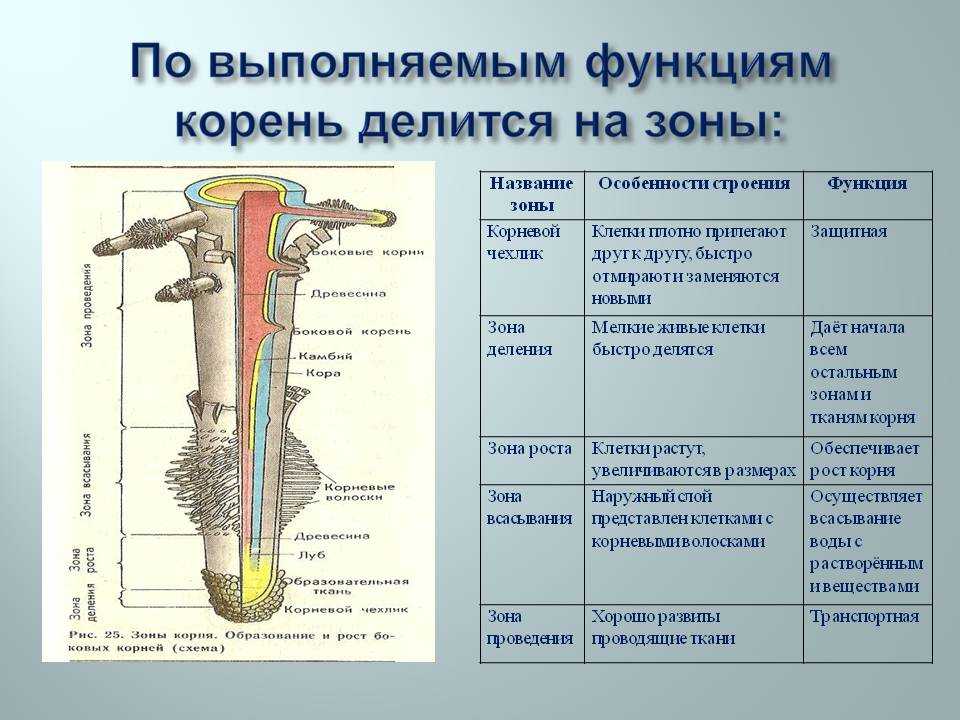 110, 2395–2400. doi: 10.1073/pnas.1213958110
110, 2395–2400. doi: 10.1073/pnas.1213958110
Pubmed Abstract | Опубликован полный текст | Полный текст CrossRef
Лазебник, Ю. (2002). Может ли биолог починить радио? Или то, что я узнал, изучая апоптоз. Раковая ячейка 2, 179–182. doi: 10.1016/S1535-6108(02)00133-2
Опубликовано Аннотация | Опубликован полный текст | Полный текст CrossRef
Lee, RD, and Cho, HT (2013). Ауксин, организатор гормональных/средовых сигналов для роста корневых волосков. Фронт. Завод Наука . 4:448. doi: 10.3389/fpls.2013.00448
Pubmed Abstract | Опубликован полный текст | Полный текст CrossRef
Мацке, А.Дж.М., и Мацке, М. (2013). Мембранная «потенциал-омика»: к визуализации напряжения на уровне клеточной популяции в корнях живых растений. Фронт. Завод Наука . 4:311. doi: 10.3389/fpls.2013.00311
Pubmed Abstract | Опубликован полный текст | Полный текст CrossRef
Мохд-Радзман, Н. А., Джорджевич, М. А., и Имин, Н. (2013). Азотная модуляция сигнальных путей корневой архитектуры бобовых включает фитогормоны и небольшие регуляторные молекулы.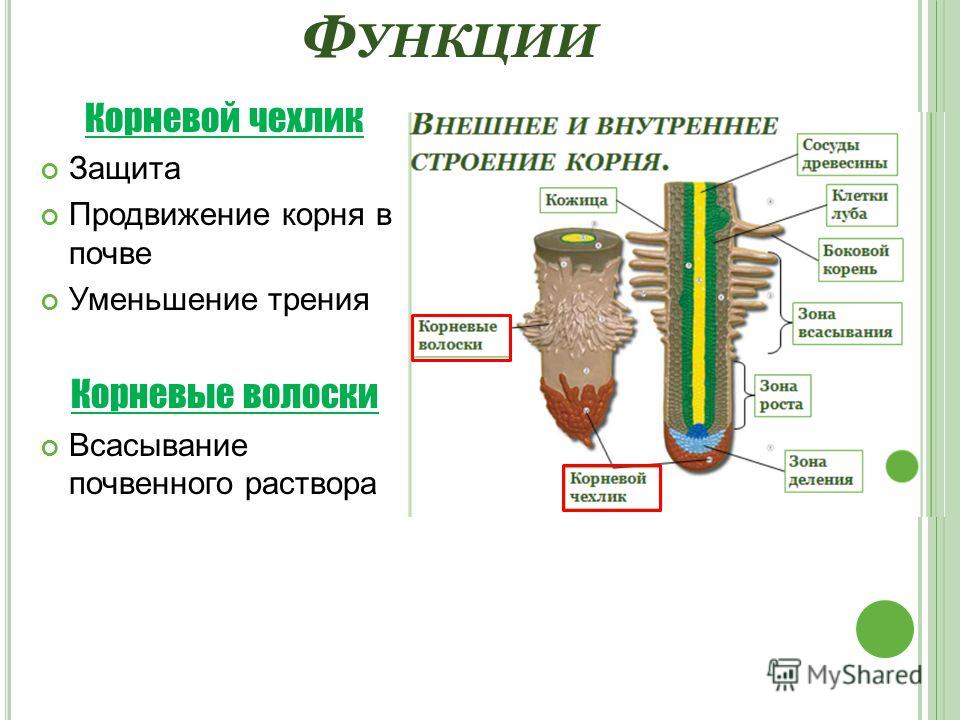 Фронт. Завод Наука . 4:385. doi: 10.3389/fpls.2013.00385
Фронт. Завод Наука . 4:385. doi: 10.3389/fpls.2013.00385
Pubmed Abstract | Опубликован полный текст | Полный текст CrossRef
Пазос, Ф., Пьеросемоли, Н., Гарсия-Мартин, Дж. А., и Солано, Р. (2013). Внутреннее нарушение белков у растений. Фронт. Завод Наука . 4:363. doi: 10.3389/fpls.2013.00363
Pubmed Abstract | Опубликован полный текст | Полный текст CrossRef
Цяо, З., и Либо, М. (2013). Раскрытие потенциала клеток корневых волосков как модели единственного типа растительных клеток в биологии корневых систем. Фронт. Завод Наука . 4:484. doi: 10.3389/fpls.2013.00484
Pubmed Abstract | Опубликован полный текст | Полный текст CrossRef
Тиан, X., и Доернер, П. (2013). Добыча корневых ресурсов: имеет ли это значение? Фронт. Завод Наука . 4:303. doi: 10.3389/fpls.2013.00303
Pubmed Abstract | Опубликован полный текст | CrossRef Full Text
Биология, структура и функция растений, форма и физиология растений, корни
Обзор
К концу этого раздела вы сможете:
- Определить два типа корневых систем
- Описать три зоны кончика корня и обобщить роль каждой зоны в росте корня
- Описать структуру корень
- Перечислите и опишите примеры модифицированных корней
Корни семенных растений выполняют три основные функции: закрепляют растение в почве, поглощают воду и минералы и транспортируют их вверх, а также сохраняют продукты фотосинтеза.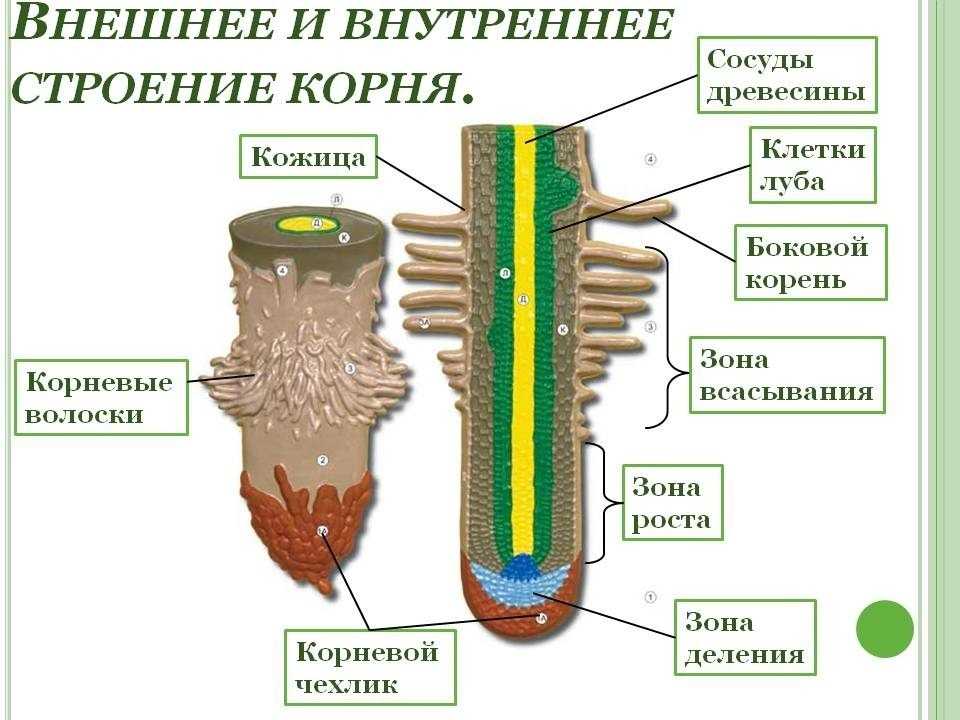 Некоторые корни модифицированы для поглощения влаги и газообмена. Большинство корней находятся под землей. Некоторые растения, однако, также имеют придаточные корни , которые выходят над землей из побега.
Некоторые корни модифицированы для поглощения влаги и газообмена. Большинство корней находятся под землей. Некоторые растения, однако, также имеют придаточные корни , которые выходят над землей из побега.
Корневые системы в основном бывают двух типов (рисунок). У двудольных корневая система стержневая, а у однодольных корневая система мочковатая. Стержневая корневая система имеет главный корень, который растет вертикально вниз и от которого отходит множество более мелких боковых корней. Одуванчики — хороший пример; их стержневые корни обычно обрываются при попытке вырвать эти сорняки, и они могут отрастить новый побег из оставшегося корня). Стержневая корневая система глубоко проникает в почву. Напротив, мочковатая корневая система расположена ближе к поверхности почвы и образует густую сеть корней, которая также помогает предотвратить эрозию почвы (хорошим примером являются газонные травы, а также пшеница, рис и кукуруза).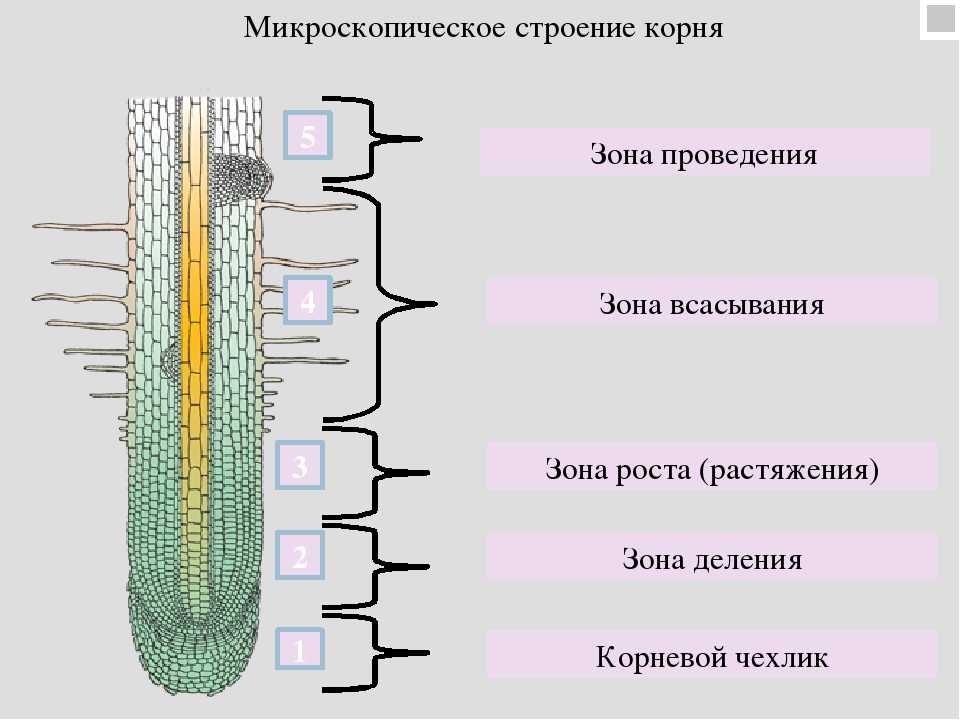 Некоторые растения имеют сочетание стержневых и мочковатых корней. Растения, произрастающие в засушливых районах, часто имеют глубокую корневую систему, в то время как растения, растущие в районах с большим количеством воды, скорее всего, имеют более мелкую корневую систему.
Некоторые растения имеют сочетание стержневых и мочковатых корней. Растения, произрастающие в засушливых районах, часто имеют глубокую корневую систему, в то время как растения, растущие в районах с большим количеством воды, скорее всего, имеют более мелкую корневую систему.
(а) стержневые корневые системы имеют основной корень, растущий вниз, а (б) мочковатые корневые системы состоят из множества мелких корней. (кредит b: модификация работы Austen Squarepants/Flickr)
Рост корней начинается с прорастания семян. Когда зародыш растения выходит из семени, корешок зародыша образует корневую систему. Кончик корня защищен корневым чехликом , структурой, эксклюзивной для корней и не похожей ни на одну другую структуру растений. Корневой чехлик постоянно заменяется, потому что он легко повреждается, когда корень проталкивается через почву. Кончик корня можно разделить на три зоны: зону клеточного деления, зону удлинения и зону созревания и дифференцировки (рис.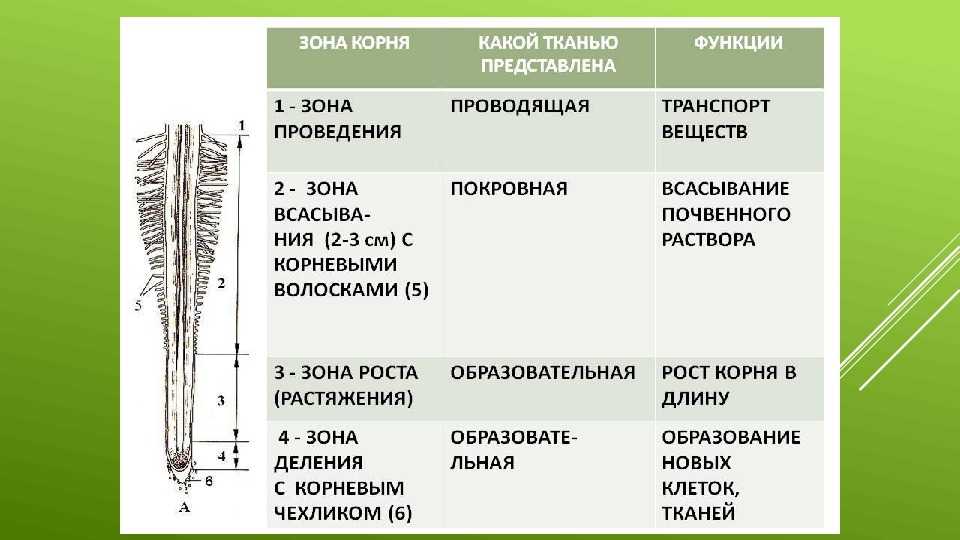 ). Зона деления клеток находится ближе всего к кончику корня; она состоит из активно делящихся клеток меристемы корня. Зона удлинения — это место, где новообразованные клетки увеличиваются в длину, тем самым удлиняя корень. Начиная с первого корневого волоска, начинается зона созревания клеток, где клетки корня начинают дифференцироваться в особые типы клеток. Все три зоны находятся примерно в первом сантиметре от кончика корня.
). Зона деления клеток находится ближе всего к кончику корня; она состоит из активно делящихся клеток меристемы корня. Зона удлинения — это место, где новообразованные клетки увеличиваются в длину, тем самым удлиняя корень. Начиная с первого корневого волоска, начинается зона созревания клеток, где клетки корня начинают дифференцироваться в особые типы клеток. Все три зоны находятся примерно в первом сантиметре от кончика корня.
На продольном срезе корня видны зоны клеточного деления, удлинения и созревания. Деление клеток происходит в апикальной меристеме.
Корень имеет внешний слой клеток, называемый эпидермисом, который окружает участки основной ткани и сосудистой ткани. Эпидермис обеспечивает защиту и помогает впитыванию. Корневые волоски , являющиеся отростками эпидермальных клеток корня, увеличивают площадь поверхности корня, в значительной степени способствуя поглощению воды и минералов.
Внутри корня основная ткань образует две области: кору и сердцевину (рисунок).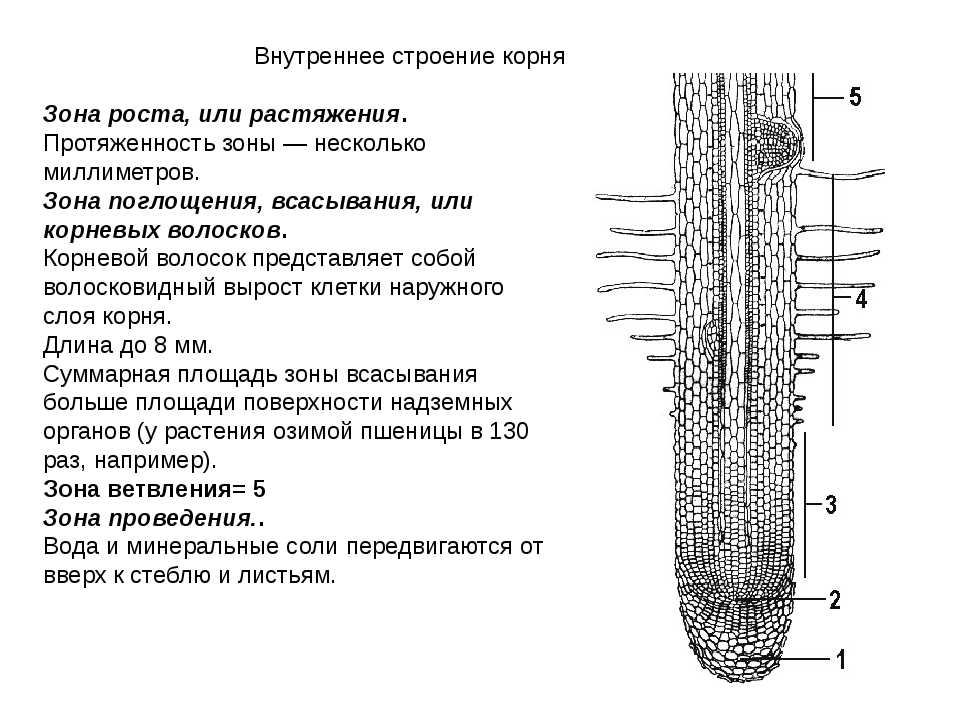 По сравнению со стеблями, корни имеют много коры и мало сердцевины. Обе области включают клетки, хранящие продукты фотосинтеза. Кора находится между эпидермисом и сосудистой тканью, тогда как сердцевина лежит между сосудистой тканью и центром корня. Окрашивание
По сравнению со стеблями, корни имеют много коры и мало сердцевины. Обе области включают клетки, хранящие продукты фотосинтеза. Кора находится между эпидермисом и сосудистой тканью, тогда как сердцевина лежит между сосудистой тканью и центром корня. Окрашивание
показывает различные типы клеток на этой световой микрофотографии поперечного среза корня пшеницы ( Triticum ). Клетки склеренхимы экзодермы и клетки ксилемы окрашиваются в красный цвет, а клетки флоэмы — в синий. Другие типы клеток окрашиваются в черный цвет. Стела, или сосудистая ткань, представляет собой область внутри эндодермы (обозначена зеленым кольцом). Корневые волоски видны вне эпидермиса. (кредит: данные масштабной линейки от Мэтта Рассела)
Сосудистая ткань в корне расположена во внутренней части корня, которая называется стелой (рисунок). Слой клеток, известный как эндодерма , отделяет стелу от основной ткани во внешней части корня.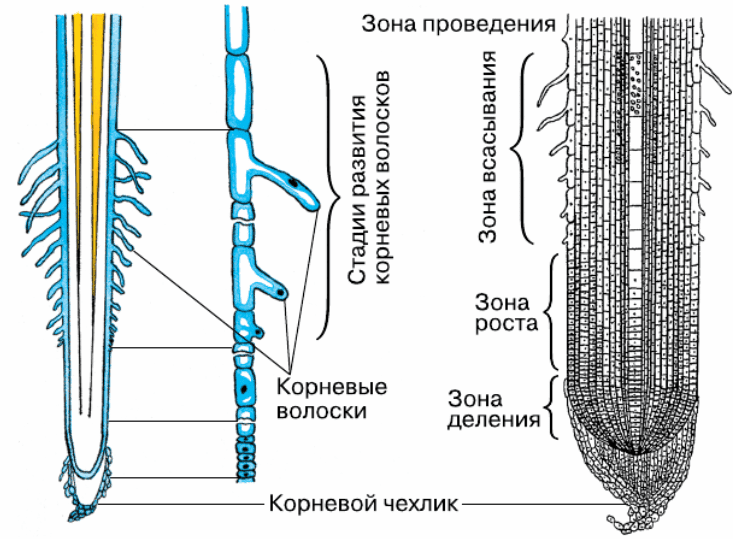 Энтодерма присуща только корням и служит контрольным пунктом для материалов, поступающих в сосудистую систему корня. На стенках энтодермальных клеток присутствует воскообразное вещество, называемое суберином. Эта восковидная область, известная как полоса Каспариана , заставляет воду и растворенные вещества пересекать плазматические мембраны энтодермальных клеток вместо того, чтобы скользить между клетками. Это гарантирует, что через эндодерму проходят только материалы, необходимые для корня, в то время как токсичные вещества и патогены, как правило, исключаются. Самый наружный клеточный слой сосудистой ткани корня — это перицикл , область, которая может дать начало боковым корням. У корней двудольных ксилема и флоэма стелы расположены попеременно в форме буквы X, тогда как у корней однодольных проводящая ткань располагается кольцом вокруг сердцевины.
Энтодерма присуща только корням и служит контрольным пунктом для материалов, поступающих в сосудистую систему корня. На стенках энтодермальных клеток присутствует воскообразное вещество, называемое суберином. Эта восковидная область, известная как полоса Каспариана , заставляет воду и растворенные вещества пересекать плазматические мембраны энтодермальных клеток вместо того, чтобы скользить между клетками. Это гарантирует, что через эндодерму проходят только материалы, необходимые для корня, в то время как токсичные вещества и патогены, как правило, исключаются. Самый наружный клеточный слой сосудистой ткани корня — это перицикл , область, которая может дать начало боковым корням. У корней двудольных ксилема и флоэма стелы расположены попеременно в форме буквы X, тогда как у корней однодольных проводящая ткань располагается кольцом вокруг сердцевины.
У типичных двудольных (слева) сосудистая ткань образует Х-образную форму в центре корня. У типичных однодольных (справа) клетки флоэмы и более крупные клетки ксилемы образуют характерное кольцо вокруг центральной сердцевины.
Корневые структуры могут быть изменены для определенных целей. Например, некоторые корни луковичные и хранят крахмал. Воздушные корни и опорные корни — это две формы надземных корней, которые обеспечивают дополнительную поддержку для закрепления растения. Стержневые корнеплоды, такие как морковь, репа и свекла, являются примерами корнеплодов, модифицированных для хранения пищевых продуктов (рис.).
Многие овощи представляют собой модифицированные корни.
Эпифитные корни позволяют растению расти на другом растении. Например, эпифитные корни орхидей развивают губчатую ткань для поглощения влаги. Баньян ( Ficus sp.) начинается как эпифит, прорастая в ветвях дерева-хозяина; воздушные корни развиваются из ветвей и со временем достигают земли, обеспечивая дополнительную опору (рис.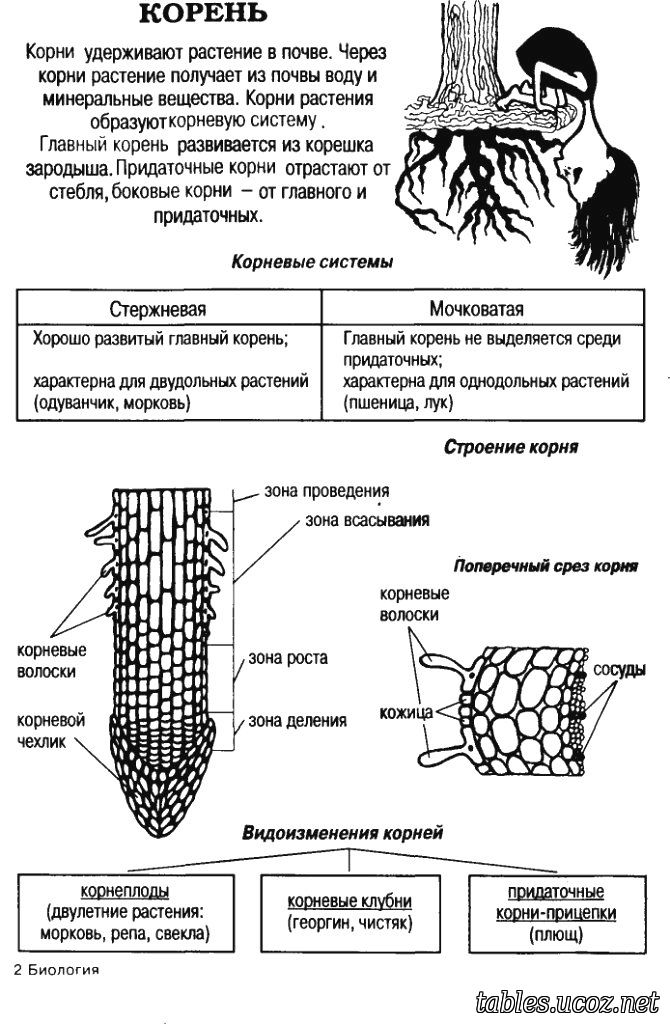 ). У сосны ( Pandanus sp.), похожего на пальму дерева, растущего на песчаных тропических почвах, из узлов развиваются надземные опорные корни, обеспечивающие дополнительную поддержку.
). У сосны ( Pandanus sp.), похожего на пальму дерева, растущего на песчаных тропических почвах, из узлов развиваются надземные опорные корни, обеспечивающие дополнительную поддержку.
(а) Баньяновое дерево, также известное как инжир-душитель, начинает жизнь как эпифит в дереве-хозяине. Воздушные корни простираются до земли и поддерживают растущее растение, которое в конечном итоге душит дерево-хозяин. У сосны (b) развиваются надземные корни, которые помогают поддерживать растение на песчаных почвах. (кредит а: модификация работы «psyberartist»/Flickr; кредит б: модификация работы Дэвида Эйхоффа)
Корни помогают закрепить растение, поглощают воду и минералы, а также служат местом хранения пищи. Стержневые и мочковатые корни являются двумя основными типами корневых систем. В стержневой корневой системе главный корень растет вертикально вниз с несколькими боковыми корнями. Мочковатые корневые системы возникают у основания стебля, где скопление корней образует густую сеть, более мелкую, чем стержневой корень.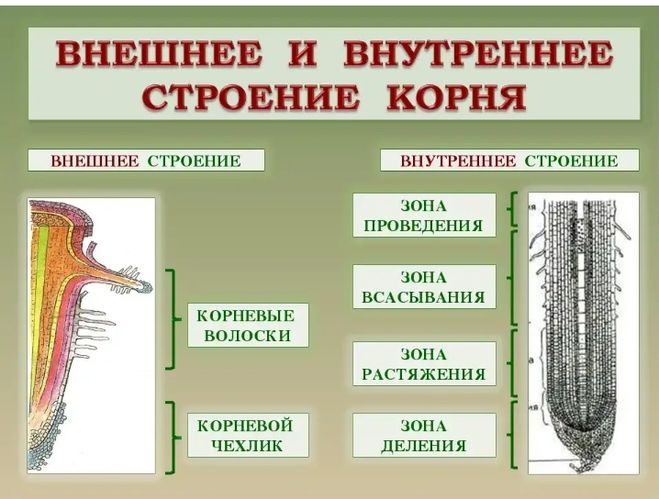 Растущий кончик корня защищен корневым чехликом. Кончик корня имеет три основные зоны: зону клеточного деления (клетки активно делятся), зону удлинения (клетки увеличиваются в длину) и зону созревания (клетки дифференцируются, образуя разные типы клеток). Корневая сосудистая ткань проводит воду, минеральные вещества и сахара. В некоторых местообитаниях корни некоторых растений могут видоизменяться, образуя воздушные или эпифитные корни.
Растущий кончик корня защищен корневым чехликом. Кончик корня имеет три основные зоны: зону клеточного деления (клетки активно делятся), зону удлинения (клетки увеличиваются в длину) и зону созревания (клетки дифференцируются, образуя разные типы клеток). Корневая сосудистая ткань проводит воду, минеральные вещества и сахара. В некоторых местообитаниях корни некоторых растений могут видоизменяться, образуя воздушные или эпифитные корни.
Корни, которые позволяют растению расти на другом растении, называются ________.
- эпифитные корни
- опорные корни
- придаточные корни
- воздушные корни
Подсказка:
А
________ вызывает избирательное поглощение минералов корнями.
- перицикл
- эпидермис
- эндодерма
- корневой чехол
Подсказка:
C
Новообразованные клетки корня начинают формировать различные типы клеток в ________.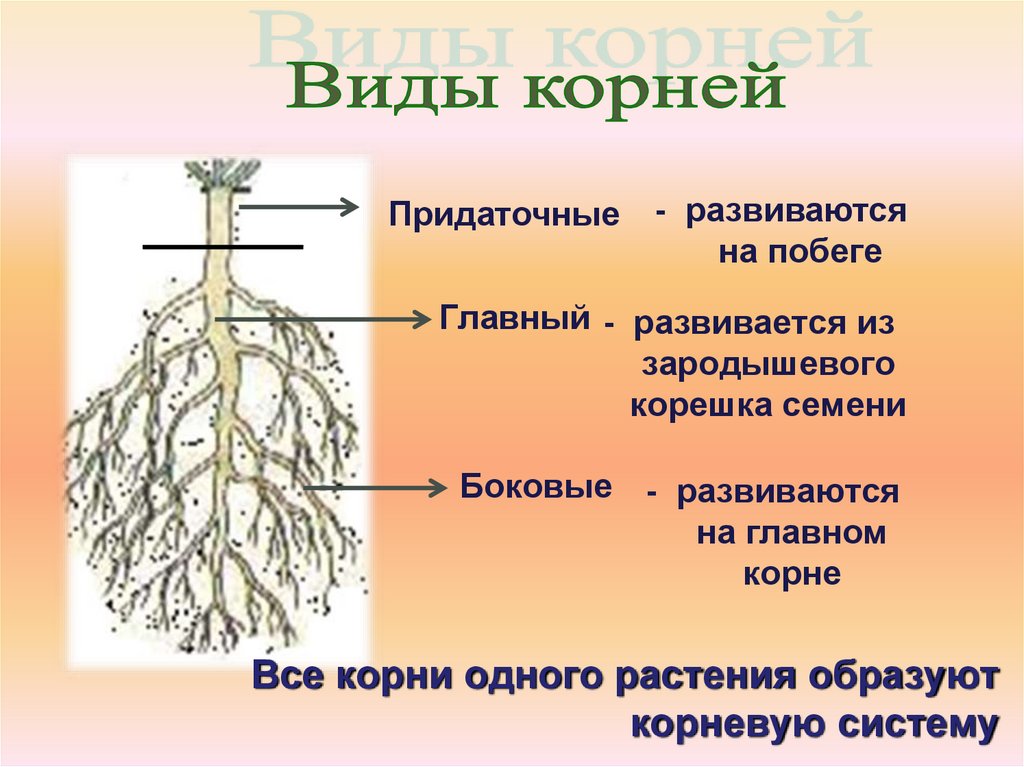
- зона удлинения
- зона созревания
- корневая меристема
- зона клеточного деления
Подсказка:
B
Сравните стержневую корневую систему с мочковатой корневой системой. Для каждого типа назовите растение, которое обеспечивает пищу в рационе человека. Какой тип корневой системы характерен для однодольных растений? Какой тип корневой системы характерен для двудольных растений?
Подсказка:
Стержневая корневая система имеет один главный корень, который растет вниз. Мочковатая корневая система образует густую сеть корней, расположенную ближе к поверхности почвы. Примером стержневой корневой системы является морковь. Травы, такие как пшеница, рис и кукуруза, являются примерами мочковатой корневой системы. Мочковатые корневые системы встречаются у однодольных; стержневые корневые системы встречаются у двудольных.
Что может произойти с корнем, если перицикл исчезнет?
Подсказка:
Корень не сможет дать боковые корни.