
Содержание
Сравнение прокариотических и эукариотических клеток
Результаты обучения
- Определение признаков, общих для всех клеток
- Сравните состав и размер прокариотических и эукариотических клеток
Клетки делятся на две широкие категории: прокариотические и эукариотические. Одноклеточные организмы доменов Bacteria и Archaea классифицируются как прокариоты ( pro = до; karyon – = ядро). Клетки животных, растительные клетки, грибы и протисты являются эукариотами ( ес = правда).
Компоненты прокариотических клеток
Все клетки имеют четыре общих компонента: (1) плазматическая мембрана, внешнее покрытие, отделяющее внутреннюю часть клетки от окружающей среды; (2) цитоплазма, состоящая из желеобразной области внутри клетки, в которой находятся другие клеточные компоненты; (3) ДНК, генетический материал клетки; и (4) рибосомы, частицы, которые синтезируют белки. Однако прокариоты отличаются от эукариотических клеток по нескольким параметрам.
Рисунок 1. На этом рисунке показана обобщенная структура прокариотической клетки.
Прокариотическая клетка представляет собой простой одноклеточный (одноклеточный) организм, в котором отсутствует ядро или какая-либо другая мембраносвязанная органелла. Вскоре мы увидим, что у эукариот это существенно отличается. Прокариотическая ДНК находится в центральной части клетки: затемненная область, называемая нуклеоидом (рис. 1).
В отличие от архей и эукариот, бактерии имеют клеточную стенку из пептидогликана, состоящего из сахаров и аминокислот, и многие из них имеют полисахаридную капсулу (рис. 1). Клеточная стенка действует как дополнительный слой защиты, помогает клетке сохранять свою форму и предотвращает обезвоживание. Капсула позволяет клетке прикрепляться к поверхностям в окружающей среде. Некоторые прокариоты имеют жгутики, пили или фимбрии. Жгутики используются для передвижения, в то время как большинство пили используются для обмена генетическим материалом во время типа размножения, называемого конъюгацией.
Эукариотические клетки
В природе взаимосвязь между формой и функцией очевидна на всех уровнях, включая уровень клетки, и это станет ясно, когда мы будем исследовать эукариотические клетки. Принцип «форма следует за функцией» встречается во многих контекстах. Это означает, что в целом можно вывести функцию структуры, глядя на ее форму, потому что они совпадают. Например, птицы и рыбы имеют обтекаемые тела, которые позволяют им быстро перемещаться в среде, в которой они живут, будь то воздух или вода.
Эукариотическая клетка представляет собой клетку, имеющую мембраносвязанное ядро и другие мембраносвязанные компартменты или мешочки, называемые органеллами , которые выполняют специализированные функции. Слово «эукариотический» означает «истинное ядро» или «истинное ядро», намекая на наличие в этих клетках мембраносвязанного ядра. Слово «органелла» означает «маленький орган», и, как мы узнали ранее, у органелл есть специализированные клеточные функции, точно так же, как органы вашего тела имеют специализированные функции.
Размер клетки
При диаметре 0,1–5,0 мкм прокариотические клетки значительно меньше эукариотических клеток, диаметр которых колеблется в пределах 10–100 мкм (рис. 2). Небольшие размеры прокариот позволяют ионам и органическим молекулам, попавшим в них, быстро распространяться в другие части клетки. Точно так же любые отходы, образующиеся в прокариотической клетке, могут быстро выводиться наружу. Однако более крупные эукариотические клетки развили различные структурные приспособления для усиления клеточного транспорта. Действительно, большие размеры этих клеток были бы невозможны без этих приспособлений. Как правило, размер клетки ограничен, поскольку объем увеличивается гораздо быстрее, чем площадь поверхности клетки. По мере того, как клетка становится больше, ей становится все труднее и труднее получать достаточное количество материалов для поддержания процессов внутри клетки, потому что относительный размер площади поверхности, по которой должны транспортироваться материалы, уменьшается.
Рисунок 2. На этом рисунке показаны относительные размеры различных типов клеток и клеточных компонентов. Для сравнения показан взрослый человек.
Резюме: Сравнение прокариотических и эукариотических клеток
Прокариоты — это одноклеточные организмы доменов бактерий и архей. Все прокариоты имеют плазматические мембраны, цитоплазму, рибосомы, клеточную стенку, ДНК и лишены мембраносвязанных органелл. Многие также имеют полисахаридные капсулы. Прокариотические клетки имеют диаметр от 0,1 до 5,0 мкм.
Как и прокариотическая клетка, эукариотическая клетка имеет плазматическую мембрану, цитоплазму и рибосомы, но эукариотическая клетка, как правило, больше, чем прокариотическая клетка, имеет истинное ядро (это означает, что ее ДНК окружена мембраной) и другие мембраносвязанные органеллы, обеспечивающие компартментализацию функций. Эукариотические клетки имеют тенденцию быть в 10-100 раз больше прокариотических клеток.
Попробуйте
Внесите свой вклад!
У вас есть идеи по улучшению этого контента? Мы будем признательны за ваш вклад.
Улучшить эту страницуПодробнее
Клеточные стенки и внеклеточный матрикс — клетка
Хотя границы клеток определяются плазматической мембраной, многие клетки окружены нерастворимым массивом секретируемых макромолекул. Клетки бактерий, грибов, водорослей и высших растений окружены жесткими клеточными стенками, являющимися неотъемлемой частью клетки. Хотя животные клетки не заключены в клеточные стенки, они в тканях тесно связаны с внеклеточным матриксом, состоящим из белков и полисахаридов. Внеклеточный матрикс не только обеспечивает структурную поддержку клеток и тканей, но также играет важную роль в регуляции поведения клеток в многоклеточных организмах.
Бактериальные клеточные стенки
Жесткие клеточные стенки бактерий определяют форму клетки и предотвращают разрыв клетки в результате осмотического давления. Структура их клеточных стенок делит бактерии на два больших класса, которые можно различить с помощью процедуры окрашивания, известной как окраска по Граму, разработанной Кристианом Грамом в 1884 г. (). Как описано ранее в этой главе, грамотрицательные бактерии (такие как E . coli ) имеют двойную мембранную систему, в которой плазматическая мембрана окружена проницаемой внешней мембраной. Эти бактерии имеют тонкие клеточные стенки, расположенные между их внутренней и внешней мембранами. Напротив, грамположительные бактерии (такие как обычный человеческий патоген Staphylococcus aureus ) имеют только одну плазматическую мембрану, которая окружена гораздо более толстой клеточной стенкой.
(). Как описано ранее в этой главе, грамотрицательные бактерии (такие как E . coli ) имеют двойную мембранную систему, в которой плазматическая мембрана окружена проницаемой внешней мембраной. Эти бактерии имеют тонкие клеточные стенки, расположенные между их внутренней и внешней мембранами. Напротив, грамположительные бактерии (такие как обычный человеческий патоген Staphylococcus aureus ) имеют только одну плазматическую мембрану, которая окружена гораздо более толстой клеточной стенкой.
Рисунок 12.44
Стенки бактериальных клеток. Плазматическая мембрана грамотрицательных бактерий окружена тонкой клеточной стенкой под наружной мембраной. Грамположительные бактерии не имеют наружных мембран и имеют толстые клеточные стенки.
Несмотря на эти структурные различия, основным компонентом клеточных стенок как грамположительных, так и грамотрицательных бактерий является пептидогликан (), состоящий из линейных полисахаридных цепей, сшитых короткими пептидами. Из-за этой поперечно-сшитой структуры пептидогликан образует прочную ковалентную оболочку вокруг всей бактериальной клетки. Интересно, что уникальная структура их клеточных стенок также делает бактерии уязвимыми для некоторых антибиотиков. Пенициллин, например, ингибирует фермент, ответственный за образование поперечных связей между различными цепями пептидогликана, тем самым препятствуя синтезу клеточной стенки и блокируя рост бактерий.
Из-за этой поперечно-сшитой структуры пептидогликан образует прочную ковалентную оболочку вокруг всей бактериальной клетки. Интересно, что уникальная структура их клеточных стенок также делает бактерии уязвимыми для некоторых антибиотиков. Пенициллин, например, ингибирует фермент, ответственный за образование поперечных связей между различными цепями пептидогликана, тем самым препятствуя синтезу клеточной стенки и блокируя рост бактерий.
Рисунок 12.45
Пептидогликан E . кишечная палочка . Полисахаридные цепи состоят из чередующихся остатков N -ацетилглюкозамина (НАГ) и N -ацетилмурамовой кислоты (НАМ), соединенных β(1→4) гликозидными связями. Параллельные цепи сшиты тетрапептидами, присоединенными к (подробнее…)
Клеточные стенки растений
В отличие от бактерий, клеточные стенки эукариот (включая грибы, водоросли и высшие растения) состоят в основном из полисахаридов (). Основным структурным полисахаридом клеточных стенок грибов является хитин (полимер N -остатки ацетилглюкозамина), который также образует экзоскелет членистоногих (например, панцирей крабов). Клеточные стенки большинства водорослей и высших растений состоят в основном из целлюлозы, которая является самым распространенным полимером на Земле. Целлюлоза представляет собой линейный полимер остатков глюкозы, часто содержащий более 10 000 мономеров глюкозы. Остатки глюкозы соединены β(1→4) связями, что позволяет полисахариду образовывать длинные прямые цепи. Затем несколько десятков таких цепочек соединяются параллельно друг с другом, образуя микрофибриллы целлюлозы, длина которых может достигать многих микрометров.
Клеточные стенки большинства водорослей и высших растений состоят в основном из целлюлозы, которая является самым распространенным полимером на Земле. Целлюлоза представляет собой линейный полимер остатков глюкозы, часто содержащий более 10 000 мономеров глюкозы. Остатки глюкозы соединены β(1→4) связями, что позволяет полисахариду образовывать длинные прямые цепи. Затем несколько десятков таких цепочек соединяются параллельно друг с другом, образуя микрофибриллы целлюлозы, длина которых может достигать многих микрометров.
Рисунок 12.46
Полисахариды клеточных стенок грибов и растений. (A) Хитин (основной полисахарид клеточных стенок грибов) представляет собой линейный полимер остатков N -ацетилглюкозамина, тогда как целлюлоза представляет собой линейный полимер глюкозы. Углеводные мономеры соединены (подробнее…)
Внутри клеточной стенки микрофибриллы целлюлозы встроены в матрикс, состоящий из белков и двух других типов полисахаридов: гемицеллюлоз и пектинов (). Гемицеллюлозы представляют собой сильно разветвленные полисахариды, которые связаны водородными связями с поверхностью микрофибрилл целлюлозы. Это сшивает микрофибриллы целлюлозы в сеть прочных волокнистых молекул, которая отвечает за механическую прочность клеточных стенок растений. Пектины представляют собой разветвленные полисахариды, содержащие большое количество отрицательно заряженных остатков галактуроновой кислоты. Из-за этих множественных отрицательных зарядов пектины связывают положительно заряженные ионы (такие как Ca 2+ ) и улавливают молекулы воды с образованием гелей. Иллюстрацией их гелеобразующих свойств служит тот факт, что джемы и желе производятся путем добавления пектинов к фруктовому соку. В клеточной стенке пектины образуют гелеобразную сеть, которая сцепляется со сшитыми микрофибриллами целлюлозы. Кроме того, клеточные стенки содержат множество гликопротеинов, которые включены в матрикс и, как считается, обеспечивают дополнительную структурную поддержку.
Гемицеллюлозы представляют собой сильно разветвленные полисахариды, которые связаны водородными связями с поверхностью микрофибрилл целлюлозы. Это сшивает микрофибриллы целлюлозы в сеть прочных волокнистых молекул, которая отвечает за механическую прочность клеточных стенок растений. Пектины представляют собой разветвленные полисахариды, содержащие большое количество отрицательно заряженных остатков галактуроновой кислоты. Из-за этих множественных отрицательных зарядов пектины связывают положительно заряженные ионы (такие как Ca 2+ ) и улавливают молекулы воды с образованием гелей. Иллюстрацией их гелеобразующих свойств служит тот факт, что джемы и желе производятся путем добавления пектинов к фруктовому соку. В клеточной стенке пектины образуют гелеобразную сеть, которая сцепляется со сшитыми микрофибриллами целлюлозы. Кроме того, клеточные стенки содержат множество гликопротеинов, которые включены в матрикс и, как считается, обеспечивают дополнительную структурную поддержку.
Рисунок 12.47
Модель клеточной стенки растений. (A) Структуры репрезентативной гемицеллюлозы (ксилоглюкан) и пектина (рамногалактуронан). Ксилоглюкан состоит из остатков глюкозы (Glc) с боковыми цепями ксилозы (Xyl), галактозы (Gal) и фукозы (Fuc). (подробнее…)
Структура и функции клеточных стенок изменяются по мере развития растительных клеток. Стенки растущих растительных клеток (называемые первичными клеточными стенками ) относительно тонкие и гибкие, что позволяет клетке увеличиваться в размерах. Как только клетки перестают расти, они часто откладывают вторичные клеточные стенки между плазматической мембраной и первичной клеточной стенкой (). Такие вторичные клеточные стенки, более толстые и жесткие, чем первичные, особенно важны для типов клеток, отвечающих за проведение воды и обеспечение механической прочности растения.
Рисунок 12.48
Первичные и вторичные клеточные стенки. Вторичные клеточные стенки закладываются между первичной клеточной стенкой и плазматической мембраной.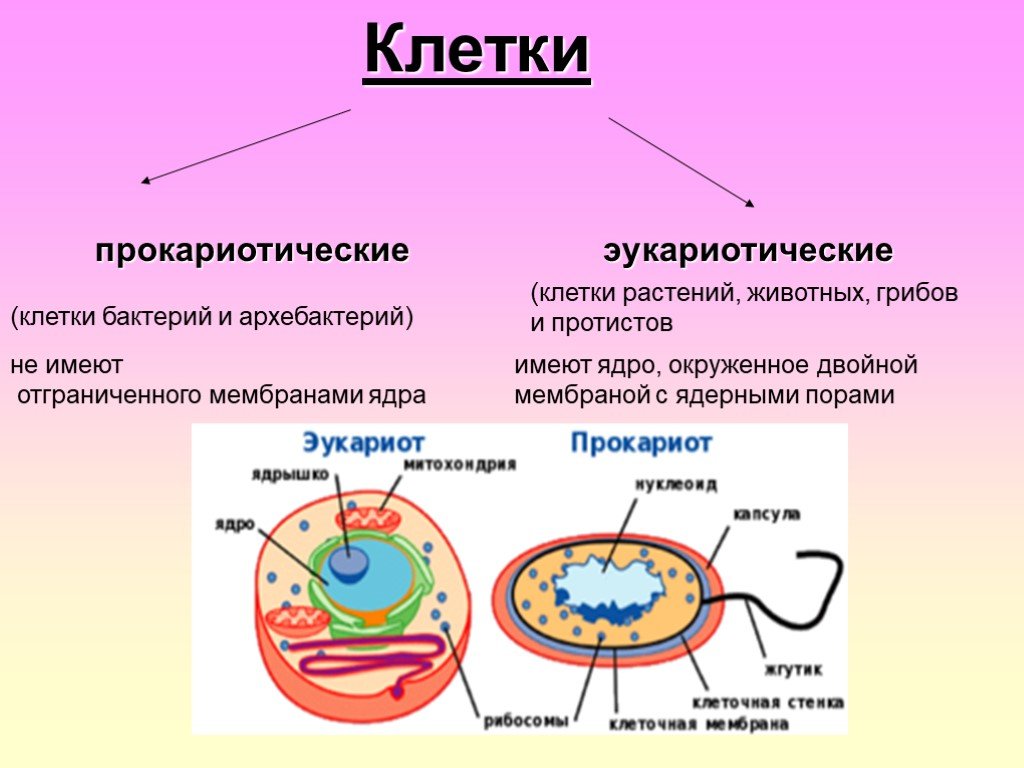 Вторичные стенки часто состоят из трех слоев, различающихся ориентацией микрофибрилл целлюлозы. Электронные микрофотографии (подробнее…)
Вторичные стенки часто состоят из трех слоев, различающихся ориентацией микрофибрилл целлюлозы. Электронные микрофотографии (подробнее…)
Первичные и вторичные клеточные стенки различаются по составу, а также по толщине. Первичные клеточные стенки содержат примерно равные количества целлюлозы, гемицеллюлозы и пектинов. Напротив, более жесткие вторичные стенки обычно не содержат пектина и содержат от 50 до 80% целлюлозы. Многие вторичные стены дополнительно укреплены лигнином, сложным полимером фенольных остатков, который в значительной степени отвечает за прочность и плотность древесины. Ориентация микрофибрилл целлюлозы также различается в первичных и вторичных клеточных стенках. Волокна целлюлозы первичных стенок, по-видимому, расположены беспорядочно, тогда как волокна вторичных стенок сильно упорядочены (см. рис. 1). Вторичные стенки часто укладываются слоями, в которых волокна целлюлозы различаются по ориентации, образуя слоистую структуру, которая значительно увеличивает прочность клеточных стенок.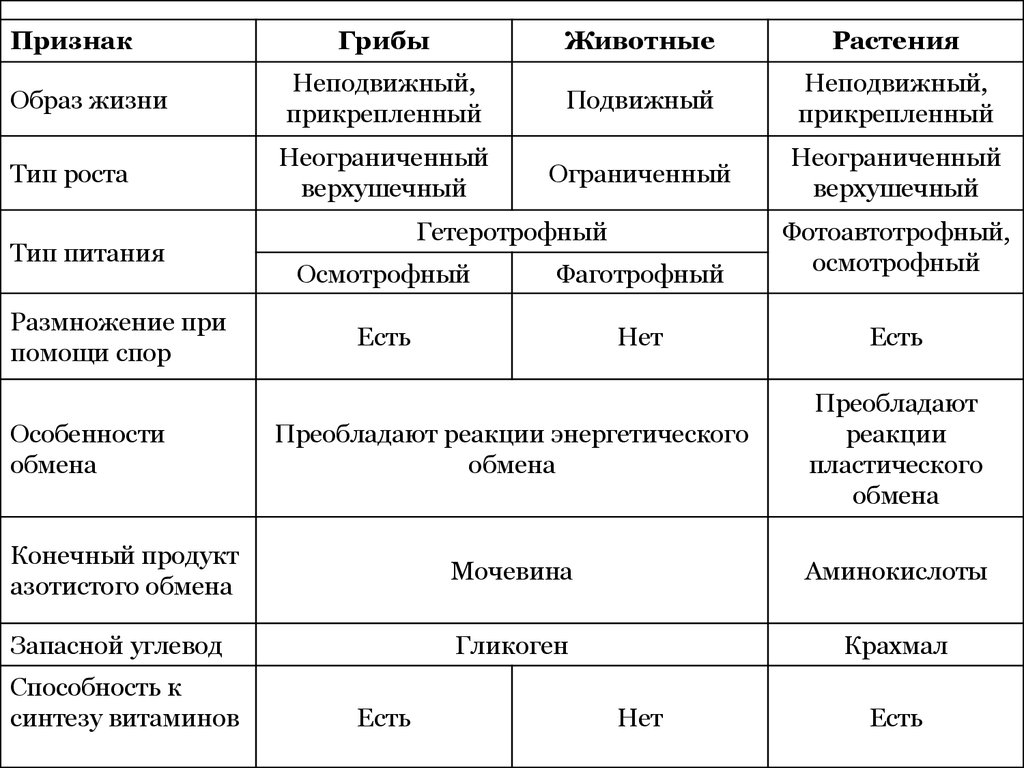
Одной из важнейших функций клеточных стенок растений является предотвращение набухания клеток в результате осмотического давления. В отличие от животных клеток, растительные клетки не поддерживают осмотического баланса между их цитозолем и внеклеточной жидкостью. Следовательно, осмотическое давление постоянно вызывает приток воды в клетку. Этот приток воды переносится растительными клетками, потому что их жесткие клеточные стенки предотвращают набухание и разрыв. Вместо этого внутри клетки создается внутреннее гидростатическое давление (называемое тургорным давлением), которое в конечном итоге выравнивает осмотическое давление и препятствует дальнейшему притоку воды.
Тургорное давление в значительной степени отвечает за жесткость растительных тканей, что легко видно при осмотре обезвоженного увядшего растения. Кроме того, тургорное давление обеспечивает основу для формы роста клеток, уникальной для растений. В частности, растительные клетки часто расширяются за счет поглощения воды без синтеза новых цитоплазматических компонентов (4). Расширение клеток по этому механизму сигнализируется растительными гормонами ( ауксины ), которые ослабляют область клеточной стенки, позволяя тургорному давлению управлять расширением клетки в этом направлении. При этом вода, поступающая в клетку, скапливается в большой центральной вакуоли, поэтому клетка расширяется, не увеличивая объем своего цитозоля. Такое расширение может привести к 10-100-кратному увеличению размеров растительных клеток в процессе развития.
Расширение клеток по этому механизму сигнализируется растительными гормонами ( ауксины ), которые ослабляют область клеточной стенки, позволяя тургорному давлению управлять расширением клетки в этом направлении. При этом вода, поступающая в клетку, скапливается в большой центральной вакуоли, поэтому клетка расширяется, не увеличивая объем своего цитозоля. Такое расширение может привести к 10-100-кратному увеличению размеров растительных клеток в процессе развития.
Рисунок 12.49
Расширение растительных клеток. Тургорное давление стимулирует расширение растительных клеток за счет поглощения воды, которая накапливается в большой центральной вакуоли.
По мере роста клеток новые компоненты клеточной стенки откладываются за пределами плазматической мембраны. Компоненты матрикса, включая гемицеллюлозы и пектины, синтезируются в аппарате Гольджи и секретируются. Однако целлюлоза синтезируется комплексом ферментов плазматической мембраны ( целлюлозсинтаза 9).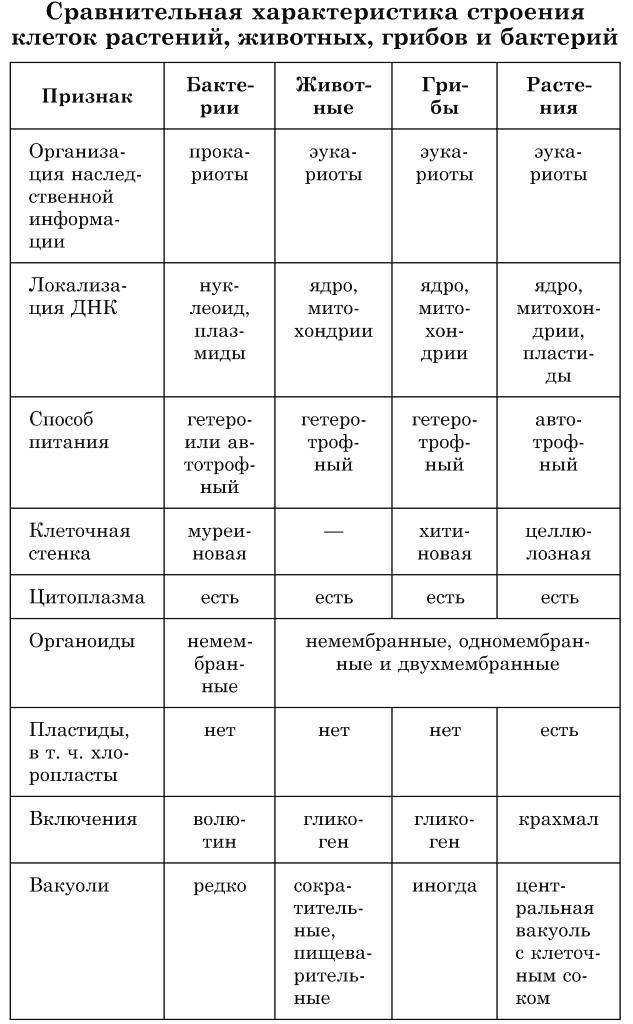 0098). В расширяющихся клетках вновь синтезированные микрофибриллы целлюлозы откладываются под прямым углом к направлению удлинения клетки — ориентация, которая, как считается, играет важную роль в определении направления дальнейшего расширения клетки (14). Интересно, что микрофибриллы целлюлозы в удлиняющихся клеточных стенках располагаются параллельно кортикальным микротрубочкам, лежащим в основе плазматической мембраны. Эти микротрубочки, по-видимому, определяют ориентацию вновь синтезированных микрофибрилл целлюлозы, возможно, путем определения направления движения комплексов целлюлозосинтазы в мембране. Таким образом, кортикальные микротрубочки определяют направление роста клеточной стенки, которое, в свою очередь, определяет направление расширения клеток и, в конечном счете, форму всего растения.
0098). В расширяющихся клетках вновь синтезированные микрофибриллы целлюлозы откладываются под прямым углом к направлению удлинения клетки — ориентация, которая, как считается, играет важную роль в определении направления дальнейшего расширения клетки (14). Интересно, что микрофибриллы целлюлозы в удлиняющихся клеточных стенках располагаются параллельно кортикальным микротрубочкам, лежащим в основе плазматической мембраны. Эти микротрубочки, по-видимому, определяют ориентацию вновь синтезированных микрофибрилл целлюлозы, возможно, путем определения направления движения комплексов целлюлозосинтазы в мембране. Таким образом, кортикальные микротрубочки определяют направление роста клеточной стенки, которое, в свою очередь, определяет направление расширения клеток и, в конечном счете, форму всего растения.
Рисунок 12.50
Синтез целлюлозы при удлинении клетки. Новые целлюлозные микрофибриллы, синтезируемые ферментным комплексом плазматической мембраны (целлюлозосинтазой), располагаются под прямым углом к направлению растяжения клетки.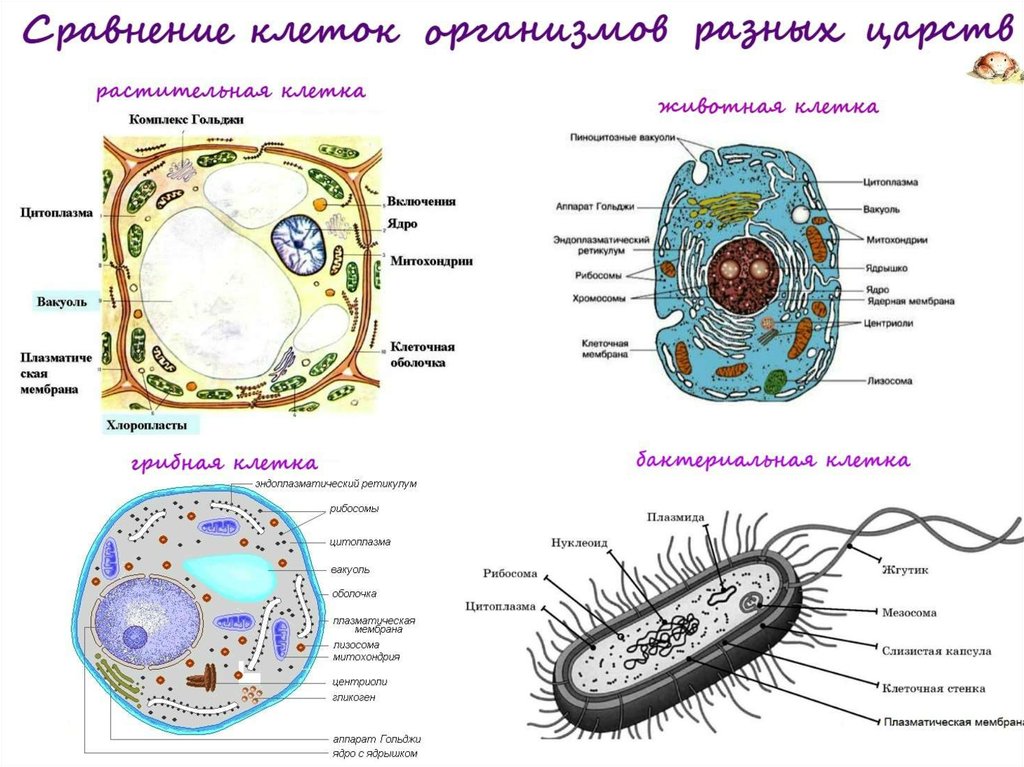 Направление синтеза целлюлозы параллельно (подробнее…)
Направление синтеза целлюлозы параллельно (подробнее…)
Внеклеточный матрикс
Хотя клетки животных не окружены клеточными стенками, многие клетки в тканях многоклеточных организмов погружены во внеклеточный матрикс, состоящий из секретируемых белков и полисахариды. Внеклеточный матрикс заполняет пространство между клетками и связывает клетки и ткани вместе. Примером одного типа внеклеточного матрикса являются тонкие пластинчатые базальные пластинки, или базальные мембраны, на которых лежат слои эпителиальных клеток (). Помимо поддерживающих слоев эпителиальных клеток, базальные пластинки окружают мышечные клетки, жировые клетки и периферические нервы. Однако внеклеточный матрикс наиболее распространен в соединительных тканях. Например, рыхлая соединительная ткань под слоями эпителиальных клеток состоит преимущественно из внеклеточного матрикса, в котором распределены фибробласты. Другие типы соединительной ткани, такие как кость, сухожилие и хрящ, также состоят в основном из внеклеточного матрикса, который в основном отвечает за их структуру и функцию.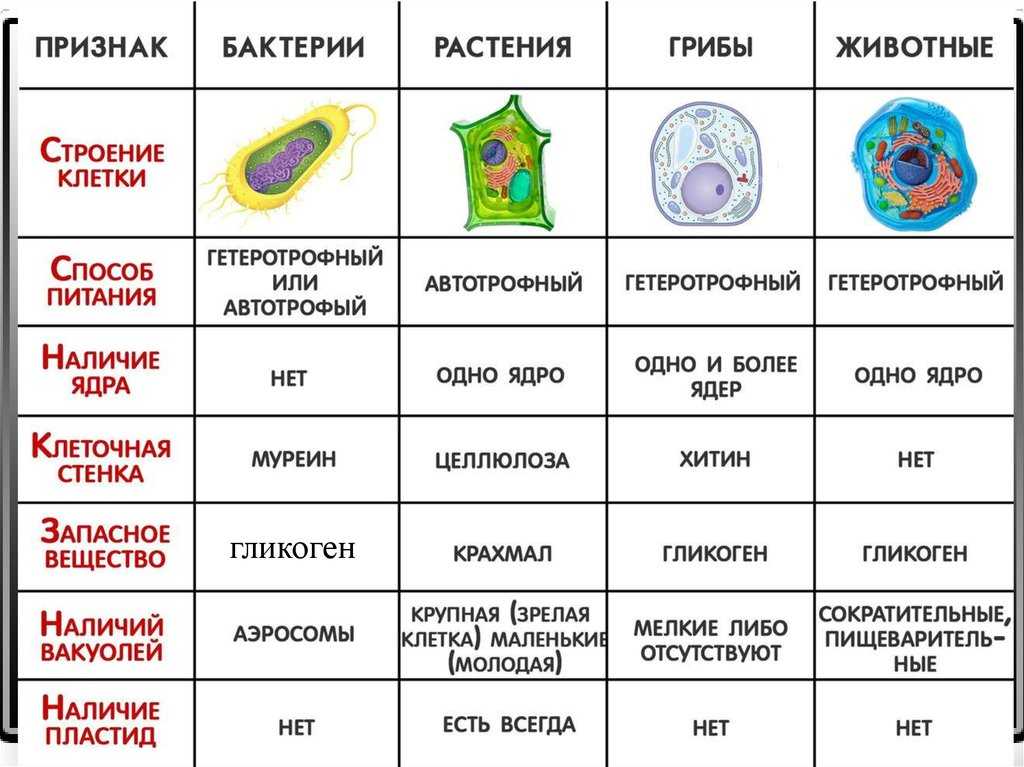
Рисунок 12.51
Примеры внеклеточного матрикса. Листы эпителиальных клеток покоятся на тонком слое внеклеточного матрикса, называемом базальной мембраной. Под базальной мембраной находится рыхлая соединительная ткань, состоящая в основном из внеклеточного матрикса, секретируемого фибробластами. (подробнее…)
Внеклеточный матрикс состоит из жестких волокнистых белков, встроенных в гелеобразное полисахаридное основное вещество, структура которого в основном аналогична структуре клеточных стенок растений. В дополнение к волокнистым структурным белкам и полисахаридам внеклеточный матрикс содержит белки адгезии, которые связывают компоненты матрикса как друг с другом, так и с прикрепленными клетками. Различия между различными типами внеклеточного матрикса являются результатом вариаций на эту общую тему. Например, сухожилия содержат большое количество волокнистых белков, тогда как хрящи содержат высокую концентрацию полисахаридов, которые образуют прочный гель, устойчивый к сжатию. В костях внеклеточный матрикс затвердевает за счет отложения кристаллов фосфата кальция. Пластинообразная структура базальных пластинок также является результатом использования компонентов матрикса, которые отличаются от тех, которые обнаруживаются в соединительных тканях.
В костях внеклеточный матрикс затвердевает за счет отложения кристаллов фосфата кальция. Пластинообразная структура базальных пластинок также является результатом использования компонентов матрикса, которые отличаются от тех, которые обнаруживаются в соединительных тканях.
Основным структурным белком внеклеточного матрикса является коллаген, который является наиболее распространенным белком в тканях животных. Коллагены представляют собой большое семейство белков, содержащее не менее 19 различных членов. Для них характерно образование тройных спиралей, в которых три полипептидные цепи плотно закручены друг вокруг друга в виде веревки. Домены тройной спирали коллагенов состоят из повторов аминокислотной последовательности Gly-X-Y. Глицин (наименьшая аминокислота, боковая цепь которой состоит только из водорода) требуется в каждом третьем положении для того, чтобы полипептидные цепи упаковывались достаточно близко друг к другу, образуя тройную спираль коллагена. Пролин часто находится в положении X, а гидроксипролин — в положении Y; благодаря своей кольцевой структуре эти аминокислоты стабилизируют спиральные конформации полипептидных цепей. Необычная аминокислота гидроксипролин образуется в эндоплазматическом ретикулуме путем модификации остатков пролина, которые уже были включены в полипептидные цепи коллагена (4). Остатки лизина в коллагене также часто превращаются в гидроксилизины. Считается, что гидроксильные группы этих модифицированных аминокислот стабилизируют тройную спираль коллагена, образуя водородные связи между полипептидными цепями. Эти аминокислоты редко встречаются в других белках, хотя гидроксипролин также часто встречается в некоторых гликопротеинах клеточных стенок растений.
Необычная аминокислота гидроксипролин образуется в эндоплазматическом ретикулуме путем модификации остатков пролина, которые уже были включены в полипептидные цепи коллагена (4). Остатки лизина в коллагене также часто превращаются в гидроксилизины. Считается, что гидроксильные группы этих модифицированных аминокислот стабилизируют тройную спираль коллагена, образуя водородные связи между полипептидными цепями. Эти аминокислоты редко встречаются в других белках, хотя гидроксипролин также часто встречается в некоторых гликопротеинах клеточных стенок растений.
Рисунок 12.52
Структура коллагена. (A) Три полипептидные цепи закручиваются друг вокруг друга в характерную структуру тройной спирали. (B) Аминокислотная последовательность домена тройной спирали коллагена состоит из повторов Gly-X-Y, в которых X часто представляет собой пролин, а Y представляет собой (подробнее…)
Рисунок 12.53
Образование гидроксипролина. Пролилгидроксилаза превращает остатки пролина в коллагене в гидроксипролин.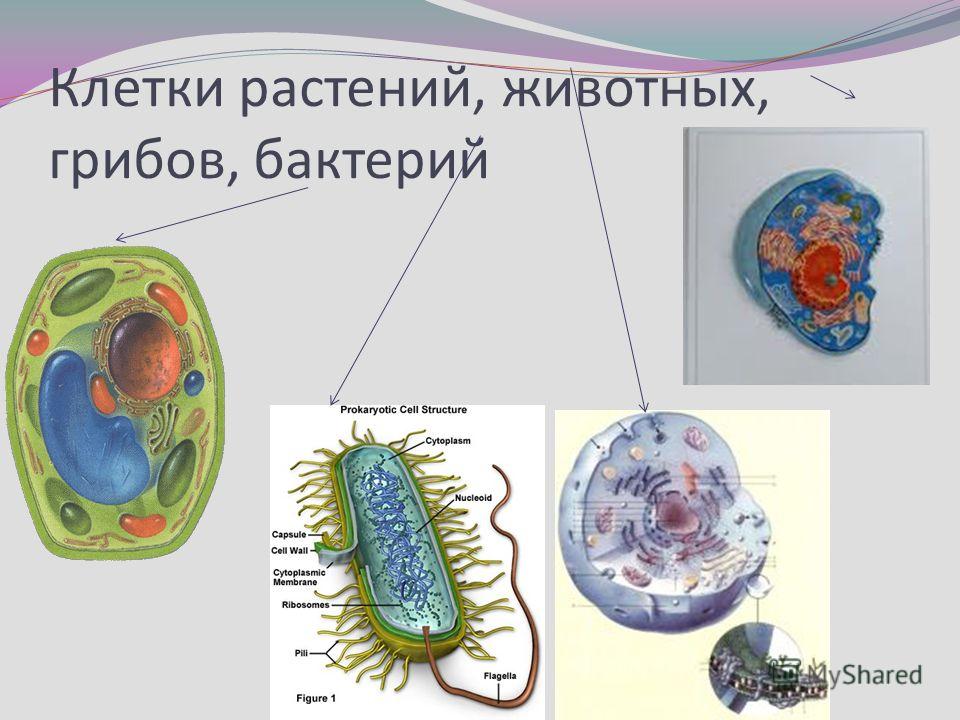
Наиболее распространенный тип коллагена (коллаген I типа) является одним из фибриллообразующих коллагенов, которые являются основными структурными компонентами соединительной ткани (). Полипептидные цепи этих коллагенов состоят примерно из тысячи аминокислот или 330 повторов Gly-X-Y. После выделения из клетки эти коллагены собираются в коллагеновые фибриллы , в которых молекулы тройной спирали связаны в правильные шахматные ряды (). Эти фибриллы не образуются внутри клетки, потому что образующие фибриллы коллагены синтезируются в виде растворимых предшественников (проколлагены ), которые содержат неспиральные сегменты на обоих концах полипептидной цепи. Проколлаген расщепляется до коллагена после его секреции, поэтому сборка коллагена в фибриллы происходит только вне клетки. Ассоциация молекул коллагена в фибриллах дополнительно усиливается за счет образования ковалентных поперечных связей между боковыми цепями остатков лизина и гидроксилизина. Часто фибриллы далее связываются друг с другом, образуя коллагеновые волокна, диаметр которых может достигать нескольких микрометров.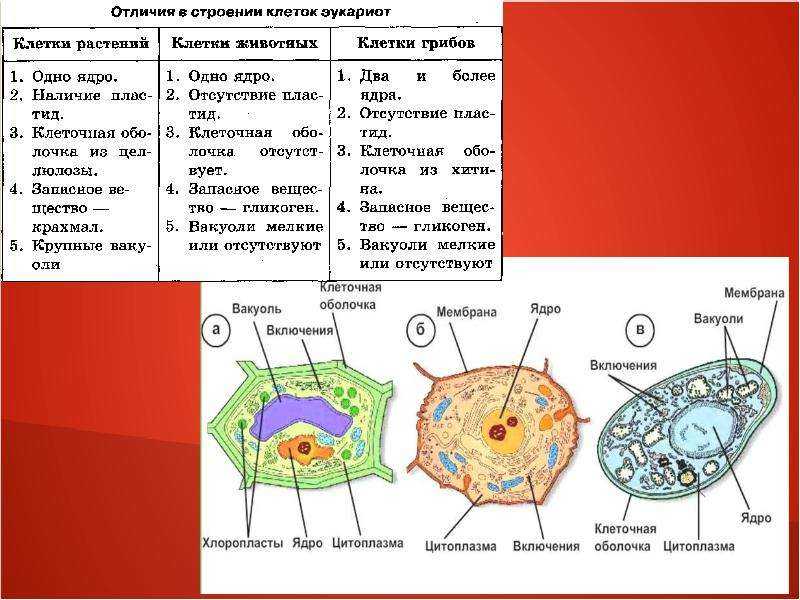
Таблица 12.2
Репрезентативные члены семейства коллагенов.
Рисунок 12.54
Коллагеновые фибриллы. (A) Молекулы коллагена собираются в правильном шахматном порядке, образуя фибриллы. Молекулы перекрываются на четверть своей длины, и между N-концом одной молекулы и С-концом другой имеется небольшой промежуток. Сборка (подробнее…)
Несколько других типов коллагена не образуют фибрилл, но играют разные роли в различных видах внеклеточного матрикса. Помимо фибриллообразующих коллагенов соединительные ткани содержат фибриллассоциированные коллагены, которые связываются с поверхностью коллагеновых фибрилл и связывают их как друг с другом, так и с другими компонентами матрикса. Базальные пластинки формируются из другого типа коллагена (коллагена IV типа), который представляет собой сеткообразующий коллаген. Повторы Gly-X-Y этих коллагенов часто прерываются короткими неспиральными последовательностями. Из-за этих прерываний коллагены, образующие сеть, более гибкие, чем коллагены, образующие фибриллы. Следовательно, они собираются в двумерные сшитые сети вместо фибрилл. Еще один тип коллагена образует якорные фибриллы, которые связывают некоторые базальные пластинки с нижележащими соединительными тканями.
Следовательно, они собираются в двумерные сшитые сети вместо фибрилл. Еще один тип коллагена образует якорные фибриллы, которые связывают некоторые базальные пластинки с нижележащими соединительными тканями.
Рисунок 12.55
Коллаген типа IV. (A) Структура повторов Gly-XY коллагена типа IV (желтый) прерывается несколькими неспиральными последовательностями (столбики). (B) Электронная микрофотография коллагеновой сети IV типа. (B, P.D. Yurchenco and J.C. Schittny, 1990. FASEB J. 4: 1577.) (подробнее…)
Соединительные ткани также содержат эластических волокон , которых особенно много в органах, которые регулярно растягиваются, а затем возвращаются к их первоначальной форме. Легкие, например, растягиваются при каждом вдохе и возвращаются к своей первоначальной форме при каждом выдохе. Эластические волокна состоят в основном из белка, называемого 9.0097 эластин , который сшит в сеть ковалентными связями, образованными между боковыми цепями остатков лизина (аналогично тем, которые обнаружены в коллагене). Эта сеть сшитых эластиновых цепей ведет себя как резиновая лента, растягиваясь при натяжении, а затем разрываясь, когда натяжение ослабевает.
Эта сеть сшитых эластиновых цепей ведет себя как резиновая лента, растягиваясь при натяжении, а затем разрываясь, когда натяжение ослабевает.
Волокнистые структурные белки внеклеточного матрикса встроены в гели, образованные из полисахаридов, называемых гликозаминогликанами, или ГАГ , которые состоят из повторяющихся звеньев дисахаридов (). Один сахар дисахарида равен N -ацетилглюкозамин или N -ацетилгалактозамин, а второй обычно кислый (либо глюкуроновая кислота, либо идуроновая кислота). За исключением гиалуроновой кислоты, эти сахара модифицированы путем добавления сульфатных групп. Следовательно, ГАГ сильно отрицательно заряжены. Подобно пектинам клеточных стенок растений, они связывают положительно заряженные ионы и улавливают молекулы воды, образуя гидратированные гели, тем самым обеспечивая механическую поддержку внеклеточного матрикса.
Рисунок 12.56
Основные типы гликозаминогликанов. Гликозаминогликаны состоят из повторяющихся дисахаридных звеньев. За исключением гиалуронана, сахара часто содержат сульфат. Гепарансульфат подобен гепарину, за исключением того, что он содержит меньше сульфатных групп. (подробнее…)
За исключением гиалуронана, сахара часто содержат сульфат. Гепарансульфат подобен гепарину, за исключением того, что он содержит меньше сульфатных групп. (подробнее…)
Гиалуронан является единственным ГАГ, который представляет собой единую длинную полисахаридную цепь. Все другие ГАГ связаны с белками с образованием протеогликанов, которые могут состоять до 95% полисахарида по весу. Протеогликаны могут содержать от одной до более сотни цепей ГАГ, присоединенных к сериновым остаткам основного белка. Было идентифицировано множество основных белков (в диапазоне от 10 до > 500 кДа), поэтому протеогликаны представляют собой разнообразную группу макромолекул. Помимо того, что они являются компонентами внеклеточного матрикса, некоторые протеогликаны представляют собой белки клеточной поверхности, которые участвуют в клеточной адгезии.
Ряд протеогликанов взаимодействует с гиалуроновой кислотой, образуя большие комплексы во внеклеточном матриксе. Хорошо охарактеризованным примером является аггрекан, основной протеогликан хряща (14).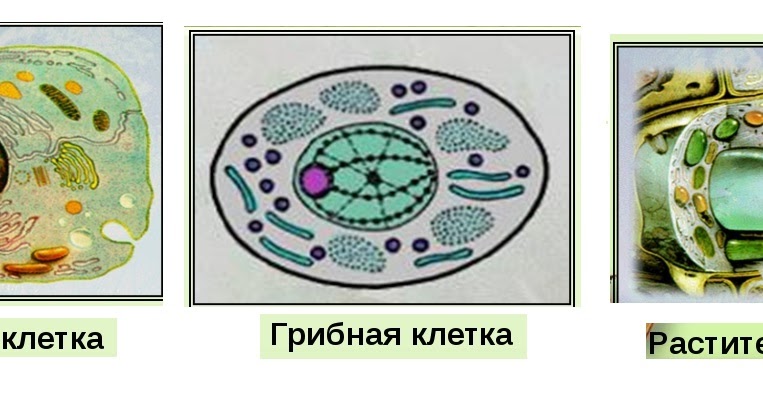 Более ста цепей хондроитинсульфата присоединены к коровому белку размером около 250 кДа, образуя протеогликан размером около 3000 кДа. Множественные молекулы аггрекана затем связываются с цепями гиалуроновой кислоты, образуя большие агрегаты (>100 000 кД), которые попадают в сеть коллагена. Протеогликаны также взаимодействуют как с коллагеном, так и с другими белками матрикса, образуя гелеобразные сети, в которые встроены волокнистые структурные белки внеклеточного матрикса. Например, перлекан (основной гепарансульфатный протеогликан базальных пластинок) связывается как с коллагеном типа IV, так и с адгезионным белком ламинином, который вскоре обсуждается.
Более ста цепей хондроитинсульфата присоединены к коровому белку размером около 250 кДа, образуя протеогликан размером около 3000 кДа. Множественные молекулы аггрекана затем связываются с цепями гиалуроновой кислоты, образуя большие агрегаты (>100 000 кД), которые попадают в сеть коллагена. Протеогликаны также взаимодействуют как с коллагеном, так и с другими белками матрикса, образуя гелеобразные сети, в которые встроены волокнистые структурные белки внеклеточного матрикса. Например, перлекан (основной гепарансульфатный протеогликан базальных пластинок) связывается как с коллагеном типа IV, так и с адгезионным белком ламинином, который вскоре обсуждается.
Рисунок 12.57
Комплексы аггрекана и гиалуроновой кислоты. Аггрекан представляет собой крупный протеогликан, состоящий из более чем 100 цепей хондроитинсульфата, соединенных с основным белком. Множественные молекулы аггрекана связываются с длинными цепями гиалуронана, образуя большие комплексы во внеклеточном (подробнее.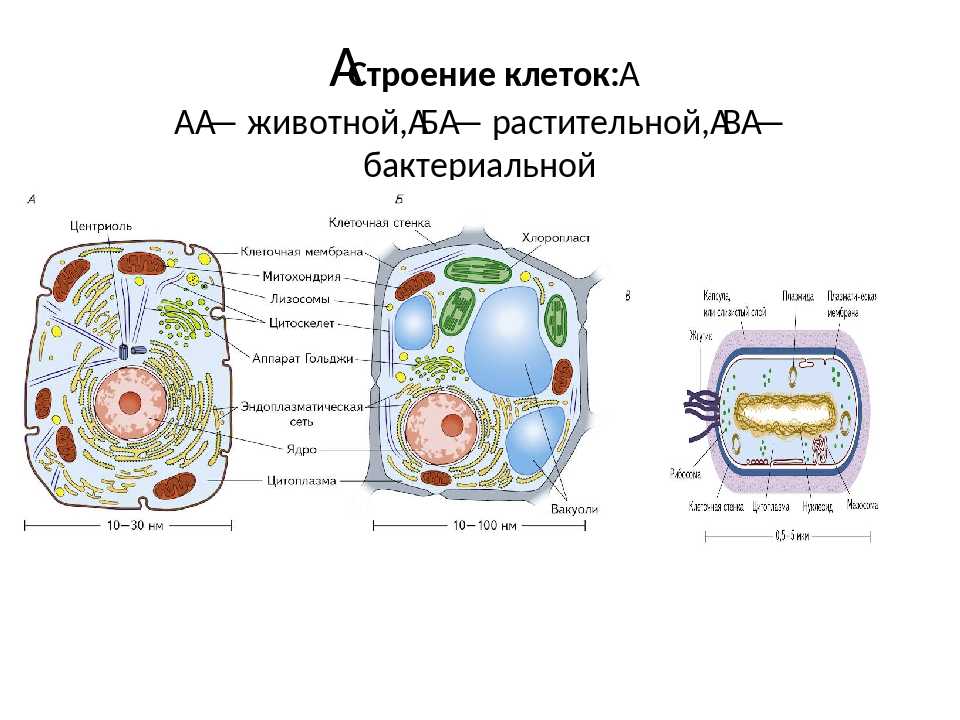 ..)
..)
Белки адгезии, третий класс компонентов внеклеточного матрикса, ответственны за связывание компонентов матрикса как друг с другом, так и с поверхности клеток. Прототипом этих молекул является фибронектин, основной белок адгезии соединительной ткани. Фибронектин представляет собой димерный гликопротеин, состоящий из двух полипептидных цепей, каждая из которых содержит около 2500 аминокислот (1). Во внеклеточном матриксе фибронектин дополнительно сшивается в фибриллы дисульфидными связями. Фибронектин имеет сайты связывания как для коллагена, так и для ГАГ, поэтому он сшивает эти компоненты матрикса. Отдельный участок молекулы фибронектина распознается рецепторами клеточной поверхности и, таким образом, отвечает за прикрепление клеток к внеклеточному матриксу.
Рисунок 12.58
Структура фибронектина. Фибронектин представляет собой димер сходных полипептидных цепей, соединенных дисульфидными связями вблизи С-конца. Указаны сайты для связывания с протеогликанами, клетками и коллагеном.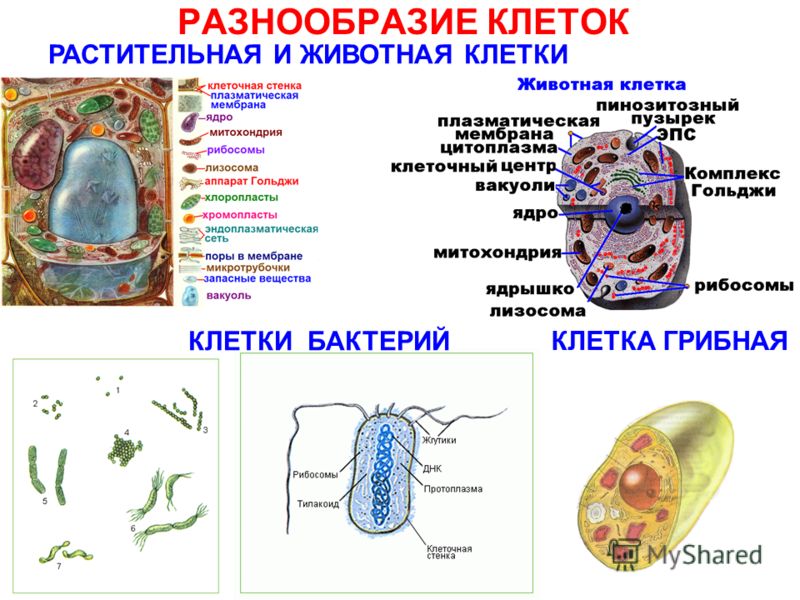 Молекула также содержит дополнительные сайты связывания, которые (подробнее…)
Молекула также содержит дополнительные сайты связывания, которые (подробнее…)
Базальные пластинки содержат отдельный белок адгезии, называемый ламинином (). Подобно коллагену типа IV, ламинины могут самостоятельно собираться в сетчатые полимеры. Такие ламининовые сети являются основными структурными компонентами базальных пластинок, синтезируемых у очень ранних эмбрионов, которые не содержат коллагена. Ламинины также имеют сайты связывания с рецепторами клеточной поверхности, коллагеном IV типа и перлеканом. Кроме того, ламинины тесно связаны с другим белком адгезии, называемым 9.0097 энтактин или нидоген , который также связывается с коллагеном IV типа. В результате этих множественных взаимодействий ламинин, энтактин, коллаген IV типа и перлекан образуют сшитые сети в базальной мембране.
Рисунок 12.59
Структура ламинина. Ламинин состоит из трех полипептидных цепей, обозначенных как A, B1 и B2. Указаны некоторые сайты связывания энтактина, коллагена IV типа, протеогликанов и рецепторов клеточной поверхности.
Основными рецепторами клеточной поверхности, ответственными за прикрепление клеток к внеклеточному матриксу, являются интегрины. Интегрины представляют собой семейство трансмембранных белков, состоящих из двух субъединиц, обозначенных как α и β (1). Было идентифицировано более 20 различных интегринов, образованных из комбинаций 18 известных α-субъединиц и 8 известных β-субъединиц. Интегрины связываются с короткими аминокислотными последовательностями, присутствующими во многих компонентах внеклеточного матрикса, включая коллаген, фибронектин и ламинин. Первым охарактеризованным таким сайтом связывания интегрина была последовательность Arg-Gly-Asp, которая распознается несколькими членами семейства интегринов. Другие интегрины, однако, связываются с различными пептидными последовательностями, включая распознаваемые последовательности в коллагенах и ламинине. Трансмембранные протеогликаны на поверхности различных клеток также связываются с компонентами внеклеточного матрикса и модулируют взаимодействия между клетками и матриксом.
Рисунок 12.60
Структура интегринов. Интегрины представляют собой гетеродимеры двух трансмембранных субъединиц, обозначенных α и β. Субъединица α связывает двухвалентные катионы (M 2+ ). Область связывания матрикса состоит из частей обеих субъединиц.
Помимо прикрепления клеток к внеклеточному матриксу, интегрины служат якорями для цитоскелета (). Возникающая в результате связь цитоскелета с внеклеточным матриксом отвечает за стабильность клеточно-матриксных соединений. Отличительные взаимодействия между интегринами и цитоскелетом обнаружены в двух типах клеточно-матриксных соединений, фокальных адгезиях и полудесмосомах, которые обсуждались в главе 11. Фокальные адгезии прикрепляют различные клетки, включая фибробласты, к внеклеточному матриксу. Цитоплазматические домены β-субъединиц интегринов в этих соединениях клетки с матриксом закрепляют актиновый цитоскелет, связываясь с пучками актиновых филаментов. Гемидесмосомы представляют собой специализированные места прикрепления эпителиальных клеток, к которым прикрепляется специфический интегрин (обозначается как α 9).