Содержание
доклад для 2 класса на урок окружающего мира
Окружающий мир
12.11.21
12 мин.
Растения — «лёгкие» планеты, ведь именно благодаря процессу фотосинтеза, происходящему в них, вырабатывается кислород. Наука, исследующая их, называется ботаникой, в ней, конечно же, присутствует определённая классификация изучаемых организмов. Какие бывают растения, их группы и виды (окружающий мир, 2 класс) — одна из самых интересных тем.
Оглавление:
- Научная классификация
- Меньшие таксоны
- Какие есть группы растений
- Другое деление представителей флоры
Растения — «лёгкие» планеты, ведь именно благодаря процессу фотосинтеза, происходящему в них, вырабатывается кислород. Наука, исследующая их, называется ботаникой, в ней, конечно же, присутствует определённая классификация изучаемых организмов. Какие бывают растения, их группы и виды (окружающий мир, 2 класс) — одна из самых интересных тем.
Научная классификация
Она считается наиболее точной из всех, поскольку её составляли и согласовывали в течение долгого времени учёные.
Самый большой «элемент» (или по-научному таксон) классификации — это царство — группа организмов, похожих по определённым признакам. Как известно, их существует 5:
- Животных.
- Грибов.
- Вирусов.
- Бактерий.
Пятое царство — растений (или по-латыни Regnum Plantae) — включает в себя абсолютно все их известные виды. Вот по каким признакам они были выделены в отдельную большую группу:
- наличие хлоропластов;
- все они являются автотрофами и осуществляют фотосинтез;
- наличие плотной оболочки на клетках, состоящей большей частью из целлюлозы;
- практически неподвижная жизнь;
- особенная, свойственная только им, структура клеток.
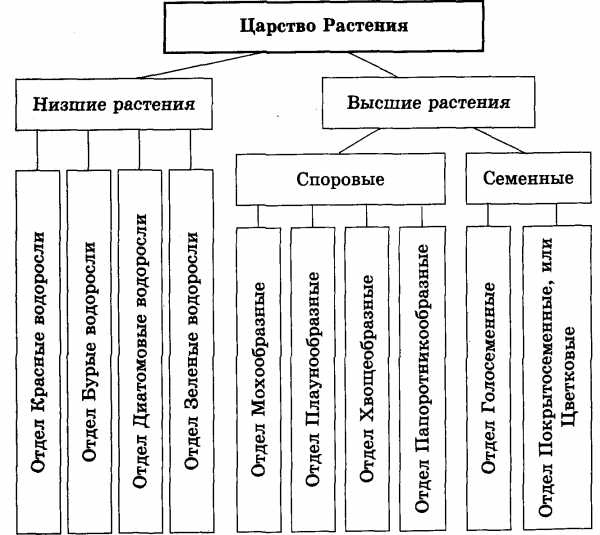
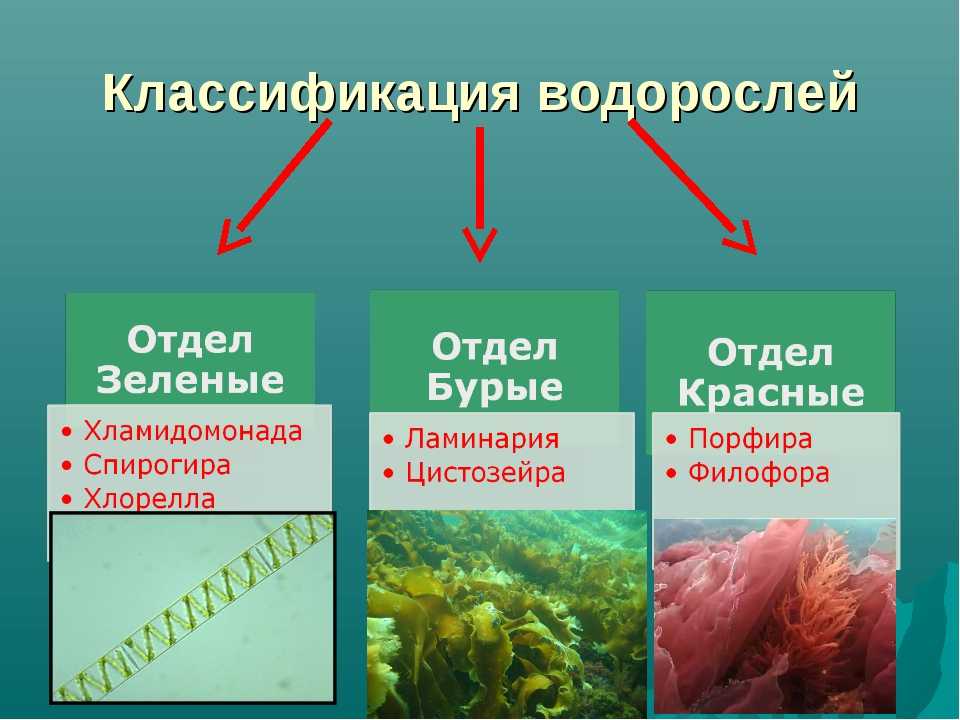
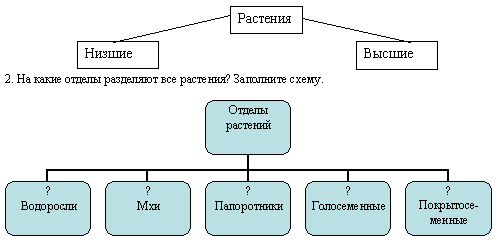
Далее следуют подцарства. Всего их 3: багрянки или красные водоросли, высшие растения и настоящие водоросли.
Раньше выделяли ещё и низшие растения, но теперь такой группы не существует, а её представители были присоединены к другим группам.
 Раньше выделяли ещё и низшие растения, но теперь такой группы не существует, а её представители были присоединены к другим группам.
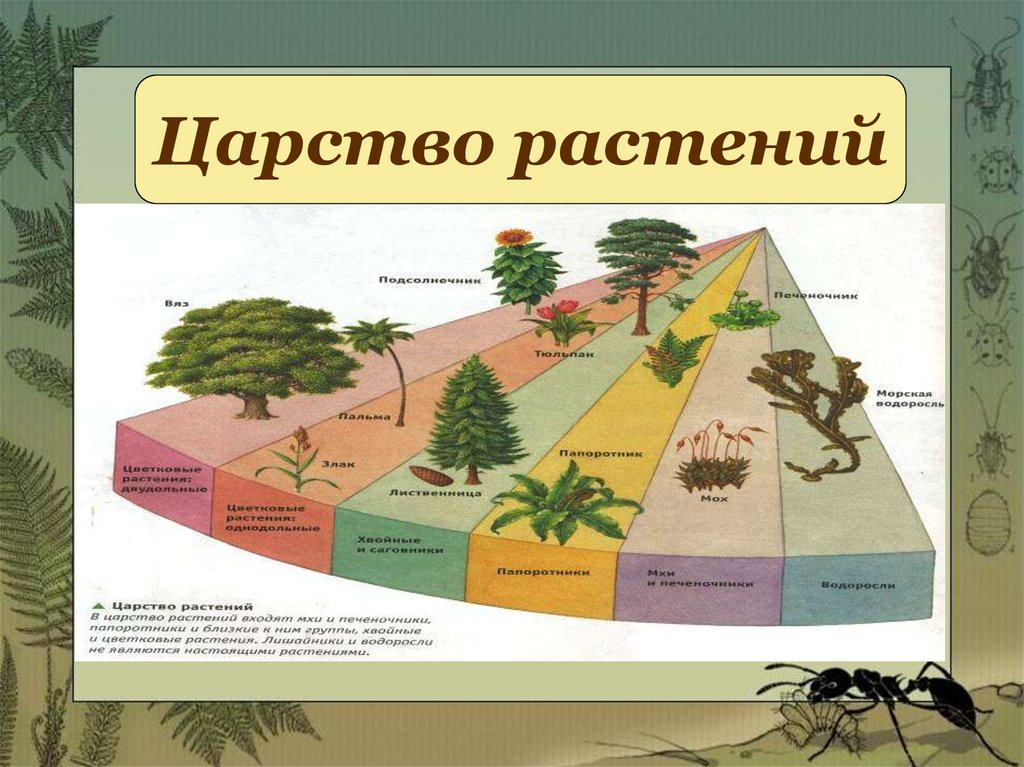
Раньше выделяли ещё и низшие растения, но теперь такой группы не существует, а её представители были присоединены к другим группам.Следующими выделяют отделы. Их очень много. Например, высшие растения:
- Риниофиты — уже исчезнувшие виды, которые находят лишь как ископаемые.
- Псилофиты — небольшая группка из 3 видов примитивнейших и очень древних представителей флоры.
- Мохообразные — здесь можно найти все виды мха, например, сфагнум, кукушкин лён и другие.
- Ликоподиофиты — вымерли лишь частично, более развиты, чем мхи. Примеры: плаун, сигиллярия.
- Эквизетофиты — травы-многолетники с развитыми корнями. По-другому их называют хвощами.
- Полиподиофиты — включают все виды папоротников и папоротникообразных.
- Пинофиты или голосеменные — деревья. Их название говорит само за себя: семена таких растений не заключены в плод и не образуются из завязи. К ним можно отнести ель, сосну, кедр.
- Магнолиофиты или покрытосеменные — эти виды образуют из завязи плод, который покрывает своей мякотью семена. Примеры: яблоня, груша, арбуз, вишня и т. п.
 Примеры: яблоня, груша, арбуз, вишня и т. п.
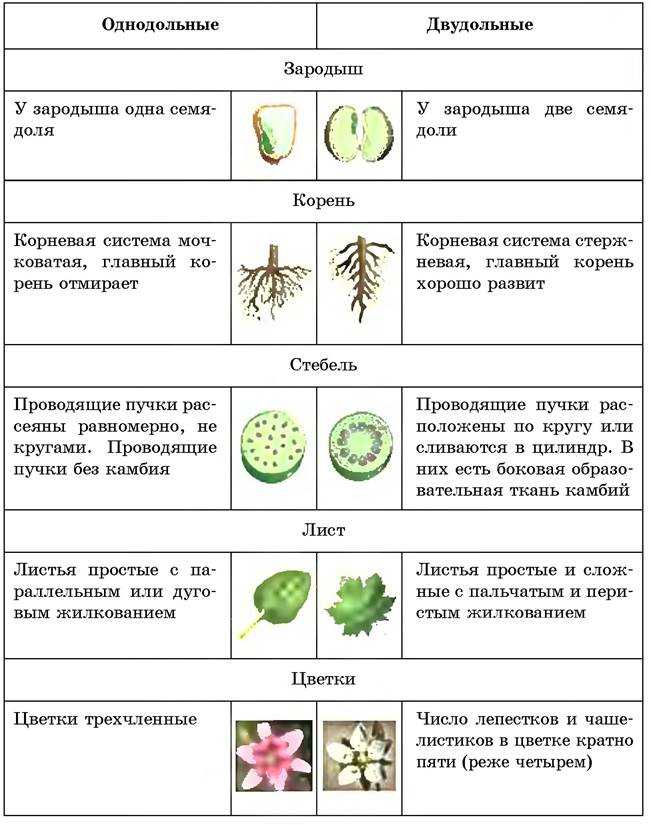
Примеры: яблоня, груша, арбуз, вишня и т. п.Отделы в свою очередь разбивают на классы. Например, покрытосеменные растения делятся на двудольные и однодольные. Первые названы так, поскольку их семечко имеет только одну так называемую семядолю (зародыш будущего листика), а у вторых их уже 2.
Меньшие таксоны
Класс, отдел и царство — это большие группы. Далее же следуют средние и маленькие начиная с подклассов. У двудольных растений выделяются:
- Магнолииды. Считаются самыми примитивными и древними среди всех подклассов двудольных. Некоторых из них природа обделила сосудистой системой, что для подобных растений — большая редкость. Подкласс представляют в большинстве своём древесные формы.
- Ранункулиды — похожи на магнолиидов, представлены в основном травами.
- Гамамелидиды — в эту группу включены такие известные виды, как дуб, бук, каштан, орех.
- Кариофиллиды. Они особенны тем, что у их цветков нет лепестков.
- Диленииды. Именно из представителя этого подкласса — тыквы — делают знаменитые украшения на Хэллоуин.
- Розиды. Судя по названию, именно здесь можно найти королеву цветов розу.
- Астериды. Цветы этих растений особенные: их лепестки срастаются, образуя из нескольких сегментов один большой.
После этого идёт деление на порядки. Например, ранункулиды подразделяются на лютикоцветных и саррациниевых. В свою очередь, в порядки объединяются семейства, в семейства — роды, а в роды — виды. Вид — самая маленькая единица, которая определяет только одного представителя живого мира.
Каждый вид можно охарактеризовать по классификации. Для примера пшеница мягкая:
- Название на латыни — Triticum aestivum.
- Род: Triticum.
- Семейство: злаки.
- Порядок — злакоцветные.
- Класс — однодольные.
- Отдел — цветковые.
- Царство — растения.
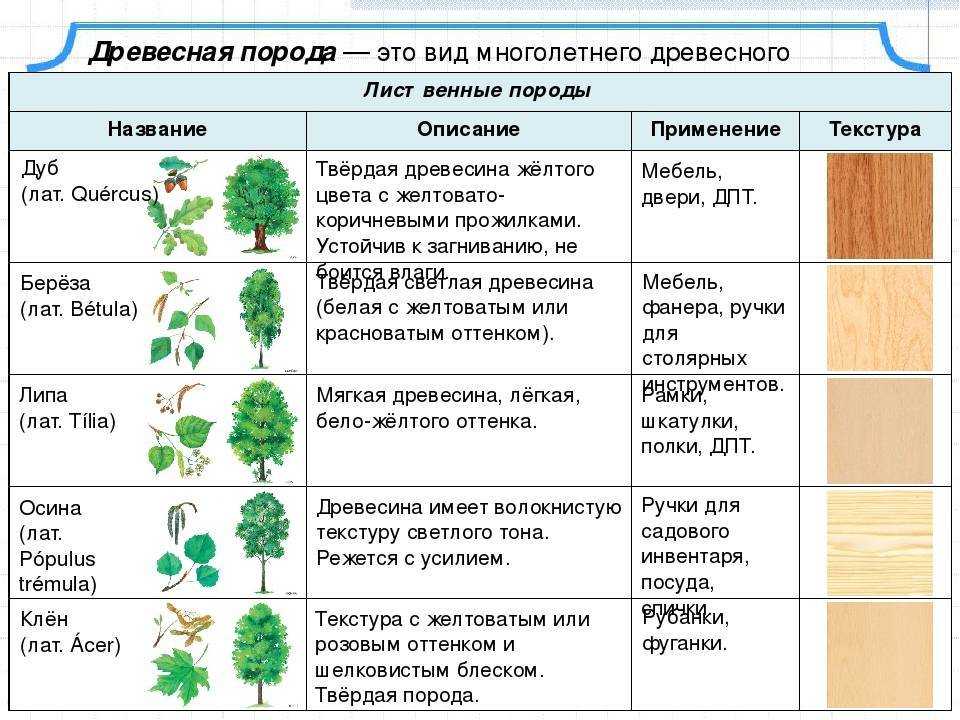
Какие есть группы растений
Помимо научной классификации, есть и более упрощённая, которая не предполагает такой точности, но позволяет определить основные отличительные черты.
Деревья — самые большие из всех растений. Их очень легко распознать по твёрдому стволу, разветвлённости, наличию кроны и сильных корней, уходящих глубоко в землю. По типу покрытия кроны они делятся на лиственных и хвойных. Ещё деревья могут жить очень долго. Именно среди них учёные нашли самого старого живого представителя флоры на планете — дуб Палмера возрастом целых 13 000 лет!
Среди деревьев есть рекордсмены и по росту. В этом смысле никто ещё не превзошёл вечнозелёную секвойю. Одна из них, которую люди называют Гиперионом, выросла до 116 метров и является самым высоким деревом Земли.
Деревья бывают самых разных типов. Есть плодоносящие, с которых собирают фрукты, декоративные, выращиваемые для красоты, экзотические, растущие в тропических и субтропических широтах.

Вторая группа — кустарники. Они тоже имеют прочные стволы и ветви, но по размерам уступают большинству деревьев. Кроме того, в отличие от больших собратьев, у кустов несколько стволов вместо одного главного.
Кустарники, как и деревья, могут быть разных типов. Самые красивые используются как декоративные растения. Например, из туи получаются отличные живые изгороди не хуже настоящих каменных заборов. Есть и плодоносные, например, барбарис, голубика, малина. Дикие виды образуют нижний ярус растительности в лесу или густые чапыжники.
В третью группу можно отнести травы. У таких растений мягкий стебель, рост не более 1,5−2 метров, тонкие корешки. Листья могут быть разного размера, это зависит от природной зоны: например, у степных трав листья тонкие, так как им хватает солнца, у лесных — большей площади, так как света не очень много.
Именно травянистое растение человек приручил первым, это была рожь. И сейчас травы преобладают в сельском хозяйстве: пшеница, рис, зелень, греча, подсолнечник считаются именно ими.
Существуют и лекарственные виды, их люди использовали ещё в те далёкие времена, когда вели кочевой образ жизни. На основе их сегодня создают многие лекарства: настойки валерианы, зверобоя, пустырника, натуральные мази, горчичники и многое другое. А самые простые дикие травы являются основной пищей для травоядных животных.
Другое деление представителей флоры
Все они без исключения делятся на комнатные, дикорастущие и культурные.
Комнатные — цветы или деревца, которые люди используют как декоративный элемент интерьера, поэтому они чаще всего очень красивы внешне, имеют приятный запах. Некоторые могут приносить пользу: отпугивать насекомых, поглощать вредные вещества из воздуха и даже увлажнять его, выделять успокаивающие запахи, благодаря которым легче заснуть.
Дикорастущие — те, что обитают в нетронутых цивилизацией лесах, степях, пустынях и горах.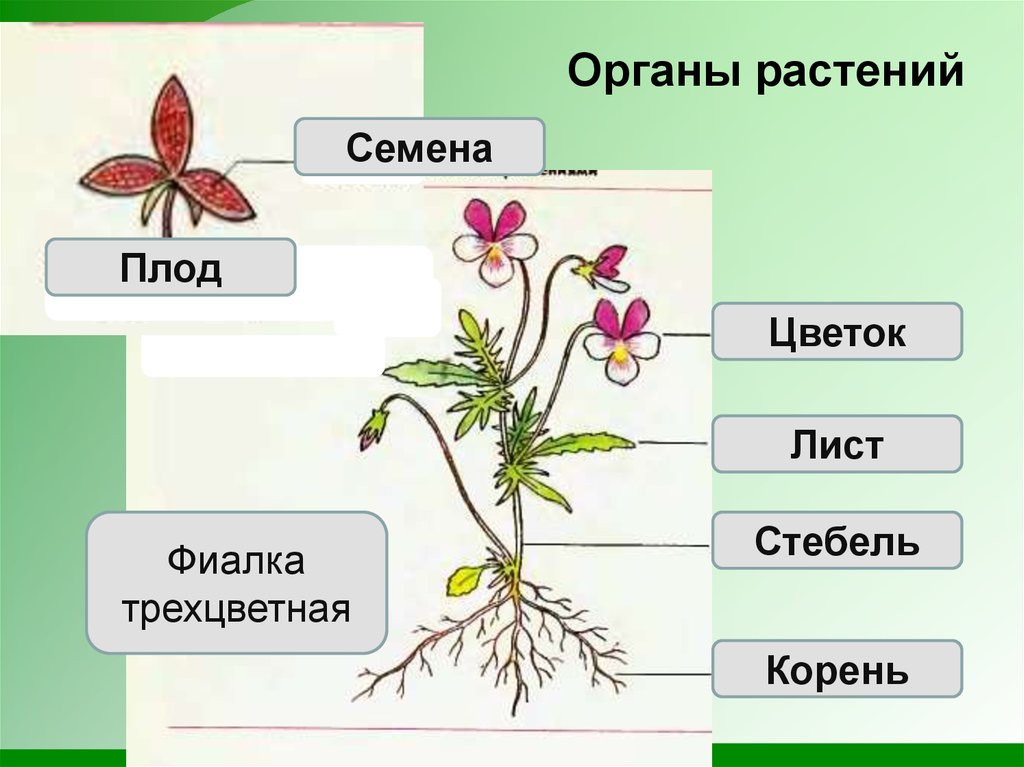 Среди них встречаются как совсем невзрачные, так и очень красивые представители, не уступающие во внешности многим комнатным цветам. Однако и такие растения приносят пользу: из них получают древесину, используют как лекарственные, заготавливают на корм животным.
Среди них встречаются как совсем невзрачные, так и очень красивые представители, не уступающие во внешности многим комнатным цветам. Однако и такие растения приносят пользу: из них получают древесину, используют как лекарственные, заготавливают на корм животным.
Культурные растения появились сравнительно недавно — около 10 000 лет назад, когда человек начал заниматься земледелием. С тех пор было выведено немало сортов самых разных трав, деревьев, корнеплодов, кустарников. Благодаря им люди могут полноценно питаться и обеспечивать пищей всё население планеты. Именно появление земледелия и растительных культур дало толчок к переходу к оседлой жизни и началу технического прогресса.
Какие бывают отделы и классы растений ответьте
Биология, 26.04.2021 19:40, 1903vita
Посмотреть ответы
Другие вопросы по: Биология
Частота колебаний маятника 10 гц. чему равен период колебаний маятника? какой путь он пройдёт за период колебаний, если амплитуда колебаний 4 см?. ..
..
Опубликовано: 27.02.2019 02:20
Ответов: 1
Берестейская уния была а-1596 б-1569 в-1591…
Опубликовано: 27.02.2019 13:30
Ответов: 3
Написати усмішку(гумористичний твір)…
Опубликовано: 28.02.2019 04:20
Ответов: 1
.(Квартира состоит из трех комнат, общей площадью 42 кв. метра. первая комната в два раза меньше второй, а вторая на три кв. метра больше третьей. какова площадь каждой комнаты в э…
Опубликовано: 28.02.2019 13:50
Ответов: 3
Найдите частное а)1.989.680: 187; б)572.163: 709; в)9.018009: 1001 г)533.368.000: 83.600…
Опубликовано: 01.03.2019 11:40
Ответов: 1
Магазин 2,4 т груш и 3,6 т яблок. во сколько раз больше ривезли яблок, чем груш? какую часть фруктов составляли груши? сколько процентов от всех фруктов составляют яблоки? p. s…
s…
Опубликовано: 02.03.2019 20:00
Ответов: 1
Знаешь правильный ответ?
Какие бывают отделы и классы растений ответьте …
Популярные вопросы
Найдите 2 числа если их разность равна 6,а семь двеннадцатых одного числа равны 70% второго?…
Опубликовано: 27.02.2019 19:30
Ответов: 3
Решение по битва на курской дуге началась 5июля1943года а закончилась 23августа того же года и вчесть победы был дан первый салют сколько дней длилась битва на курской дуге? ско…
Опубликовано: 01.03.2019 22:00
Ответов: 3
She now had her own opinions, and at supper discussed with sasha’s parents, saying how difficult the studies had become for the children at the school. but after all, she felt a cl…
Опубликовано: 02.03.2019 09:30
Ответов: 3
Диагонали параллелограмма равны 12 см и 20 см, а угол между ними равен 60 градусам. найдите стороны параллелограмма…
найдите стороны параллелограмма…
Опубликовано: 03.03.2019 12:10
Ответов: 1
Твір «врятували»з поданих слів: прекрасний ранок, ранкова прохолода, легенький морозець, друзі побігли до села. потрібно на сьогодні…
Опубликовано: 03.03.2019 13:10
Ответов: 1
:к источнику тока эдс е=9в и внутренним сопротивлением r=10м подключили параллельно соединенные с сопротивлением r=80 и плоский конденсатор, расстояние между пластинами которого d…
Опубликовано: 06.03.2019 23:40
Ответов: 1
Четырнадцать целых двадцать двадцать девятых минус пять целых двадцать двадцать девятых минус пять целых шесть седьмых…
Опубликовано: 07.03.2019 14:20
Ответов: 3
Составить предложения с выражениями: 1)that`s an idea! 2) what a good idea! 3) to be full of ideas 4)to have an idea of smth 5)to give an idea of smth 6)to book a ticket 7)to fi. ..
..
Опубликовано: 07.03.2019 18:00
Ответов: 3
Чтоб поддерживать скворечню иль антенну, я гожусь….
Опубликовано: 07.03.2019 21:00
Ответов: 1
Со следующими словосочетаниями составьте предложения ; искреннею просьбой…
Опубликовано: 08.03.2019 00:50
Ответов: 1
Случайные вопросы
Рост растений | Биология для специальностей II
Определение ключевых элементов и процессов роста растений
Большинство растений продолжают расти на протяжении всей своей жизни. Как и другие многоклеточные организмы, растения растут за счет сочетания клеточного роста и клеточного деления. Рост клеток увеличивает размер клеток, а деление клеток (митоз) увеличивает количество клеток.
Цели обучения
- Описать, как растут растения
- Различают первичный рост и вторичный рост стеблей
- Понять, как гормоны влияют на рост и развитие растений
Как растут растения
Рисунок 1.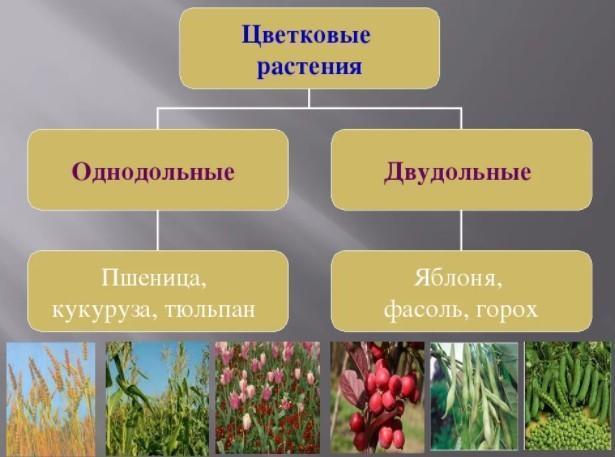 Должна быть область роста, похожая на то, как удлиняются кости пальцев, рук и ног. Есть, и она называется апикальной меристемой, которая здесь показана.
Должна быть область роста, похожая на то, как удлиняются кости пальцев, рук и ног. Есть, и она называется апикальной меристемой, которая здесь показана.
Большинство растений продолжают расти на протяжении всей своей жизни. Как и другие многоклеточные организмы, растения растут за счет сочетания клеточного роста и клеточного деления. Рост клеток увеличивает размер клеток, а деление клеток (митоз) увеличивает количество клеток. По мере роста растительных клеток они также становятся специализированными в различные типы клеток посредством клеточной дифференциации. Как только клетки дифференцируются, они больше не могут делиться. Как после этого растения растут или заменяют поврежденные клетки?
Ключом к непрерывному росту и восстановлению растительных клеток является меристема . Меристема — это тип растительной ткани, состоящий из недифференцированных клеток, которые могут продолжать делиться и дифференцироваться.
Апикальные меристемы находятся на верхушке или кончике корней и почек, позволяя корням и стеблям расти в длину, а листьям и цветкам дифференцироваться. Корни и стебли растут в длину, потому что меристема добавляет ткань «за собой», постоянно продвигаясь дальше в землю (для корней) или воздух (для стеблей). Часто апикальная меристема одной ветви становится доминирующей, подавляя рост меристем на других ветвях и приводя к развитию одного ствола. У трав меристемы у основания листовых пластинок позволяют отрастать после выпаса травоядными животными или скашивания газонокосилками.
Корни и стебли растут в длину, потому что меристема добавляет ткань «за собой», постоянно продвигаясь дальше в землю (для корней) или воздух (для стеблей). Часто апикальная меристема одной ветви становится доминирующей, подавляя рост меристем на других ветвях и приводя к развитию одного ствола. У трав меристемы у основания листовых пластинок позволяют отрастать после выпаса травоядными животными или скашивания газонокосилками.
Апикальные меристемы дифференцируются в три основных типа меристемной ткани, которые соответствуют трем типам ткани: протодерма образует новый эпидермис, основная меристема образует основную ткань, а прокамбий образует новую ксилему и флоэму. Эти три типа меристемы считаются первичной меристемой , поскольку они обеспечивают рост в длину или высоту, известный как первичный рост .
Рисунок 2. Микрофотография кончика корня фасоли показывает быстро делящуюся ткань апикальной меристемы сразу за корневым чехликом. Можно наблюдать многочисленные клетки на разных стадиях митоза.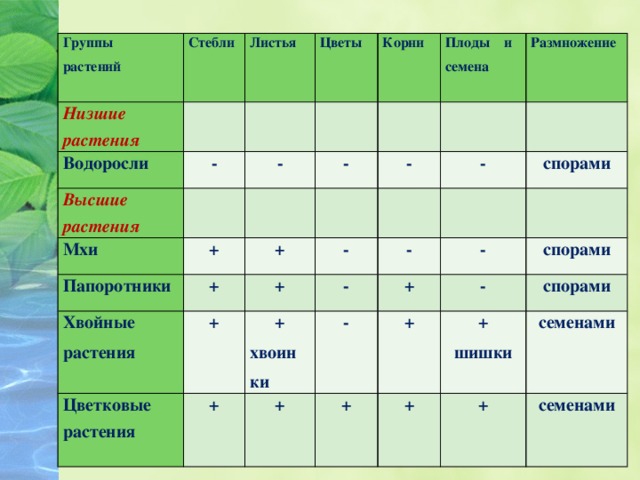
Вторичные меристемы обеспечивают рост в диаметре ( вторичный рост ) у древесных растений. Травянистые растения не имеют вторичного роста. Два типа вторичной меристемы оба называются камбием , что означает «обмен» или «изменение». Сосудистый камбий образует вторичную ксилему (по направлению к центру стебля или корня) и флоэму (по направлению к внешней стороне стебля или корня), увеличивая диаметр растения. Этот процесс производит древесину и строит крепкие стволы деревьев. Пробковый камбий лежит между эпидермисом и флоэмой и замещает эпидермис корней и стеблей корой, один слой которой представляет собой пробку.
Рисунок 3. Первичный и вторичный рост
Древесные растения растут двумя способами. Первичный рост добавляет длину или высоту, опосредованную тканью апикальной меристемы на кончиках корней и побегов, что трудно четко показать на диаграммах поперечного сечения.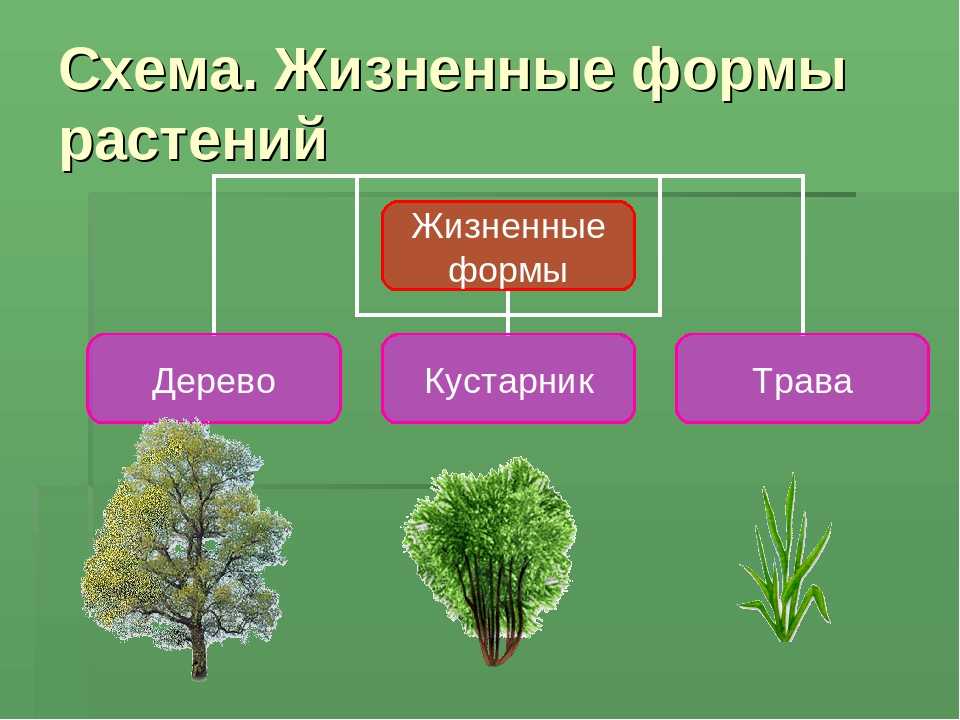 Вторичный рост добавляется к диаметру стебля или корня; сосудистый камбий добавляет ксилему (внутрь) и флоэму (наружу), а пробковый камбий заменяет эпидермис корой.
Вторичный рост добавляется к диаметру стебля или корня; сосудистый камбий добавляет ксилему (внутрь) и флоэму (наружу), а пробковый камбий заменяет эпидермис корой.
Посмотрите это покадровое видео о росте растений. Обратите внимание, что в видео нет повествования.
Кратко: Как растут растения
Большинство растений продолжают расти, пока живут. Они растут за счет сочетания клеточного роста и клеточного деления (митоза). Ключом к росту растений является меристема, тип растительной ткани, состоящий из недифференцированных клеток, которые могут продолжать делиться и дифференцироваться. Меристема позволяет стеблям и корням растений становиться длиннее (первичный рост) и шире (вторичный рост).
Рост стебля
Рост растений происходит по мере удлинения стебля и корня. Некоторые растения, особенно древесные, также увеличиваются в толщине в течение жизни. Увеличение длины побега и корня называется первичным ростом и является результатом деления клеток в апикальной меристеме побега. Вторичный рост характеризуется увеличением толщины или обхвата растения и вызывается делением клеток боковой меристемы. На рисунке 4 показаны области первичного и вторичного роста растения. Травянистые растения в основном подвергаются первичному росту, практически без вторичного роста или увеличения толщины. У древесных растений заметен вторичный рост или «древесина»; это встречается у некоторых двудольных, но очень редко у однодольных.
Увеличение длины побега и корня называется первичным ростом и является результатом деления клеток в апикальной меристеме побега. Вторичный рост характеризуется увеличением толщины или обхвата растения и вызывается делением клеток боковой меристемы. На рисунке 4 показаны области первичного и вторичного роста растения. Травянистые растения в основном подвергаются первичному росту, практически без вторичного роста или увеличения толщины. У древесных растений заметен вторичный рост или «древесина»; это встречается у некоторых двудольных, но очень редко у однодольных.
Рисунок 4. У древесных растений за первичным ростом следует вторичный рост, который позволяет стеблю растения увеличиваться в толщине или в обхвате. Вторичная сосудистая ткань добавляется по мере роста растения, а также пробковый слой. Кора дерева простирается от сосудистого камбия до эпидермиса.
Некоторые части растений, такие как стебли и корни, продолжают расти на протяжении всей жизни растения: явление, называемое неопределенным ростом.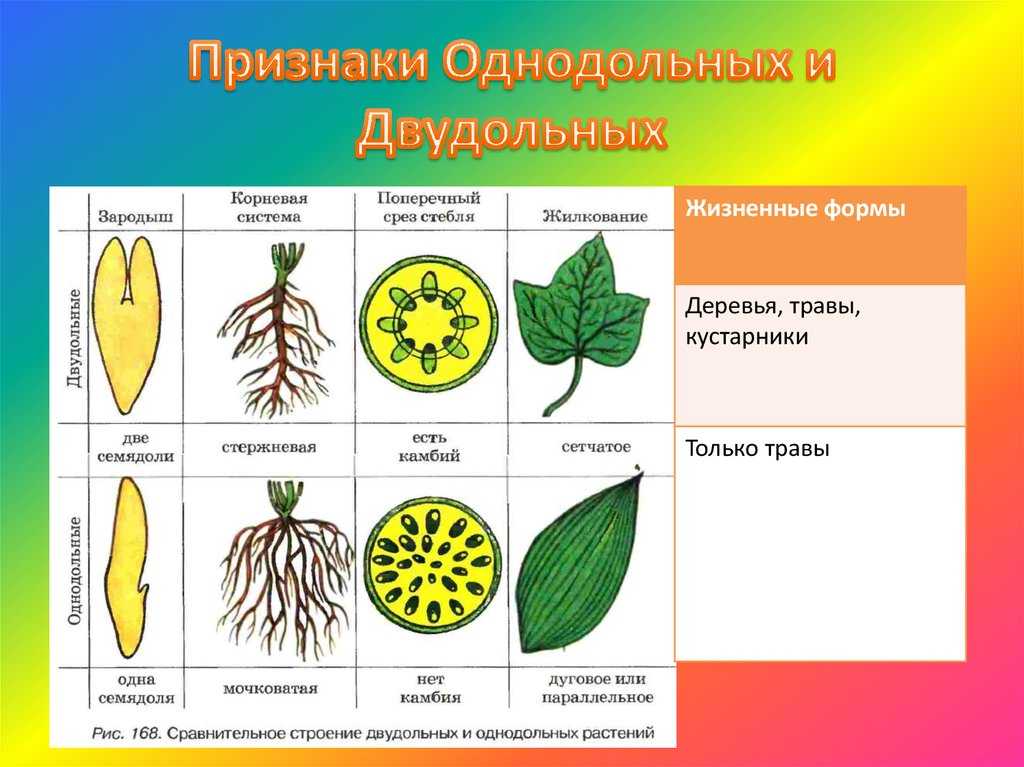 Другие части растения, такие как листья и цветы, демонстрируют детерминированный рост, который прекращается, когда часть растения достигает определенного размера.
Другие части растения, такие как листья и цветы, демонстрируют детерминированный рост, который прекращается, когда часть растения достигает определенного размера.
Первичный рост
Основной первичный рост происходит на верхушках или кончиках стеблей и корней. Первичный рост является результатом быстрого деления клеток в апикальных меристемах на кончике побега и кончике корня. Последующее удлинение клеток также способствует первичному росту. Рост побегов и корней во время первичного роста позволяет растениям постоянно искать воду (корни) или солнечный свет (побеги).
Влияние верхушечной почки на общий рост растений известно как верхушечное доминирование, которое уменьшает рост пазушных почек, образующихся по бокам ветвей и стеблей. Большинство хвойных деревьев демонстрируют сильное преобладание вершины, что приводит к типичной конической форме рождественской елки. Если верхушечную почку удалить, то пазушные почки начнут формировать боковые ответвления. Садоводы используют этот факт, когда обрезают растения, срезая верхушки ветвей, тем самым стимулируя рост пазушных почек, придавая растению кустистую форму.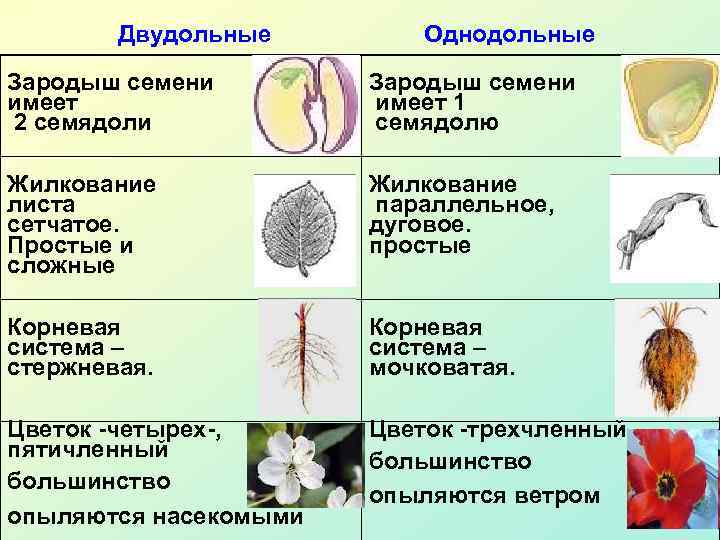
Посмотрите это видео BBC Nature, показывающее, как интервальная съемка позволяет запечатлеть рост растений на высокой скорости.
Вторичный рост
Увеличение толщины стебля в результате вторичного роста происходит из-за активности боковых меристем, которых нет у травянистых растений. Боковые меристемы включают сосудистый камбий, а у древесных растений — пробковый камбий (см. Рисунок 4).
Рисунок 5. Чечевицы на коре вишневого дерева позволяют древесному стволу обмениваться газами с окружающей атмосферой. (кредит: Роджер Гриффит)
Сосудистый камбий расположен сразу за пределами первичной ксилемы и внутри первичной флоэмы. Клетки сосудистого камбия делятся и образуют вторичную ксилему (трахеиды и элементы сосудов) внутрь и вторичную флоэму (ситовидные элементы и клетки-спутницы) наружу. Утолщение стебля, которое происходит при вторичном росте, связано с образованием вторичной флоэмы и вторичной ксилемы сосудистым камбием, а также действием пробкового камбия, который формирует самый прочный наружный слой стебля.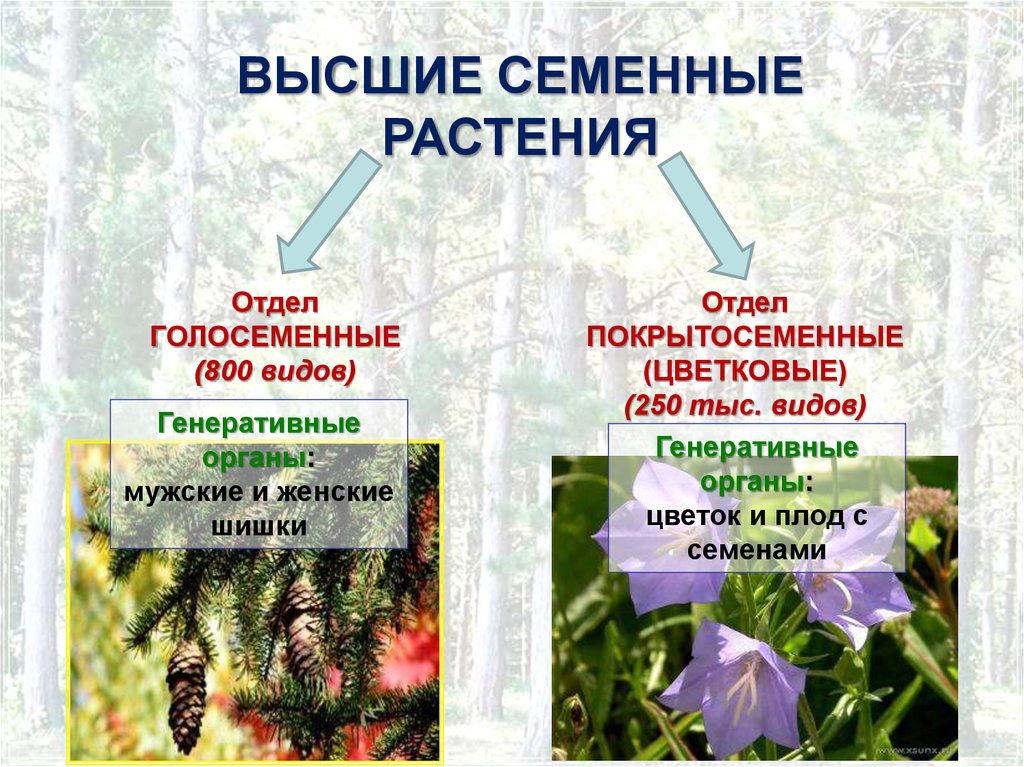 Клетки вторичной ксилемы содержат лигнин, обеспечивающий зимостойкость и прочность.
Клетки вторичной ксилемы содержат лигнин, обеспечивающий зимостойкость и прочность.
У древесных растений пробковый камбий представляет собой наружную латеральную меристему. Он производит пробковые клетки (кору), содержащие воскообразное вещество, известное как суберин, которое может отталкивать воду. Кора защищает растение от физического повреждения и помогает уменьшить потерю воды. Пробковый камбий также образует слой клеток, известный как феллодерма, который растет внутрь камбия. Пробковый камбий, пробковые клетки и феллодерма вместе называются перидермой . Перидерма заменяет эпидермис у взрослых растений. У некоторых растений перидерма имеет много отверстий, известных как 9.0079 чечевичек , которые позволяют внутренним клеткам обмениваться газами с внешней атмосферой (рис. 5). Это снабжает кислородом живые и метаболически активные клетки коры, ксилемы и флоэмы.
Годовые кольца
Рисунок 6. Скорость роста древесины увеличивается летом и уменьшается зимой, образуя характерное кольцо для каждого года роста. Сезонные изменения погодных условий также могут влиять на скорость роста — обратите внимание на то, как кольца различаются по толщине. (кредит: Адриан Пингстоун)
Сезонные изменения погодных условий также могут влиять на скорость роста — обратите внимание на то, как кольца различаются по толщине. (кредит: Адриан Пингстоун)
Деятельность сосудистого камбия дает образование годичных колец роста. В весенний вегетационный период клетки вторичной ксилемы имеют большой внутренний диаметр, а их первичные клеточные стенки не сильно утолщены. Это известно как ранняя древесина или весенняя древесина. В осенний сезон вторичная ксилема развивает утолщенные клеточные стенки, образуя позднюю или осеннюю древесину, которая более плотная, чем ранняя древесина. Такое чередование ранней и поздней древесины обусловлено в основном сезонным уменьшением числа сосудистых элементов и сезонным увеличением числа трахеид. Это приводит к образованию годового кольца, которое можно увидеть в виде круглого кольца на поперечном сечении стебля (рис. 6). Изучение количества годичных колец и их характера (например, их размера и толщины клеточной стенки) может выявить возраст дерева и преобладающие климатические условия в течение каждого сезона.
Реакция роста
Сенсорная реакция растения на внешние раздражители зависит от химических мессенджеров (гормонов). Гормоны растений влияют на все аспекты жизни растений, от цветения до завязывания и созревания плодов, от фототропизма до листопада. Потенциально каждая клетка растения может производить растительные гормоны. Они могут действовать в исходной клетке или переноситься в другие части тела растения, при этом многие реакции растений включают синергетическое или антагонистическое взаимодействие двух или более гормонов. Напротив, животные гормоны вырабатываются в определенных железах и транспортируются в отдаленные места для действия, и они действуют в одиночку.
Гормоны растений представляют собой группу неродственных химических веществ, влияющих на морфогенез растений. Традиционно описывают пять основных фитогормонов: ауксины (особенно ИУК), цитокинины, гиббереллины, этилен и абсцизовую кислоту. Кроме того, другие питательные вещества и условия окружающей среды могут быть охарактеризованы как факторы роста.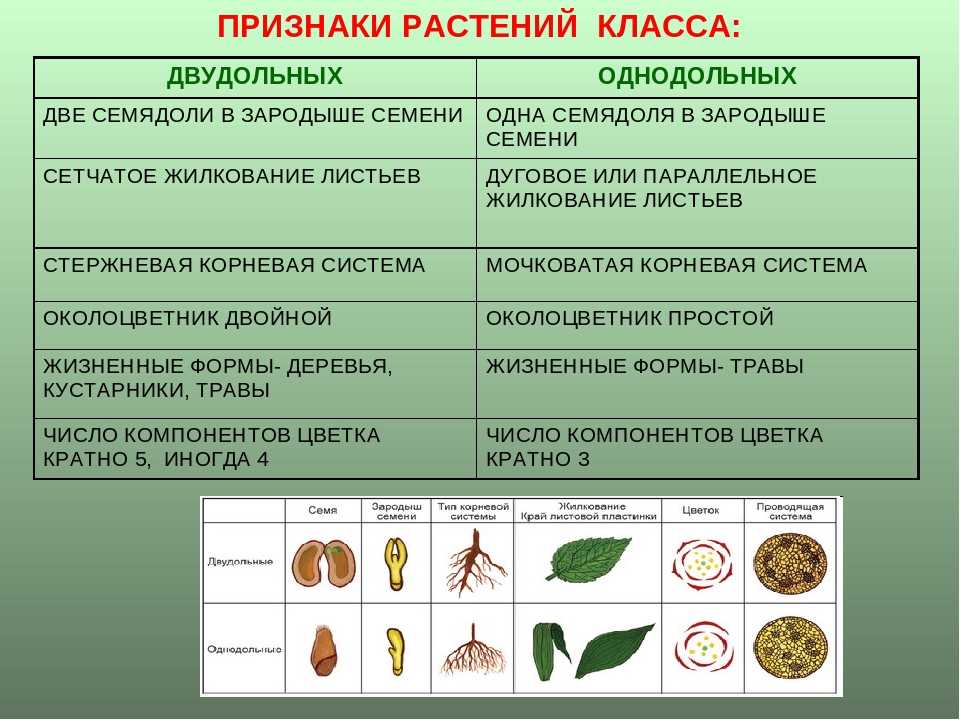
Ауксины
Термин ауксин происходит от греческого слова ауксеин , что означает «расти». Ауксины являются основными гормонами, отвечающими за удлинение клеток при фототропизме и гравитропизме. Они также контролируют дифференцировку меристемы в сосудистую ткань и способствуют развитию и расположению листьев. Хотя многие синтетические ауксины используются в качестве гербицидов, ИУК является единственным встречающимся в природе ауксином, проявляющим физиологическую активность. Апикальное доминирование — ингибирование образования боковых почек — запускается ауксинами, продуцируемыми в апикальной меристеме. Цветение, завязывание и созревание плодов, ингибирование опадение (листопад) — это другие реакции растений, находящиеся под прямым или косвенным контролем ауксинов. Ауксины также действуют как реле эффектов синего света и реакций красного/дальнего красного.
Коммерческое использование ауксинов широко распространено в питомниках растений и растениеводстве.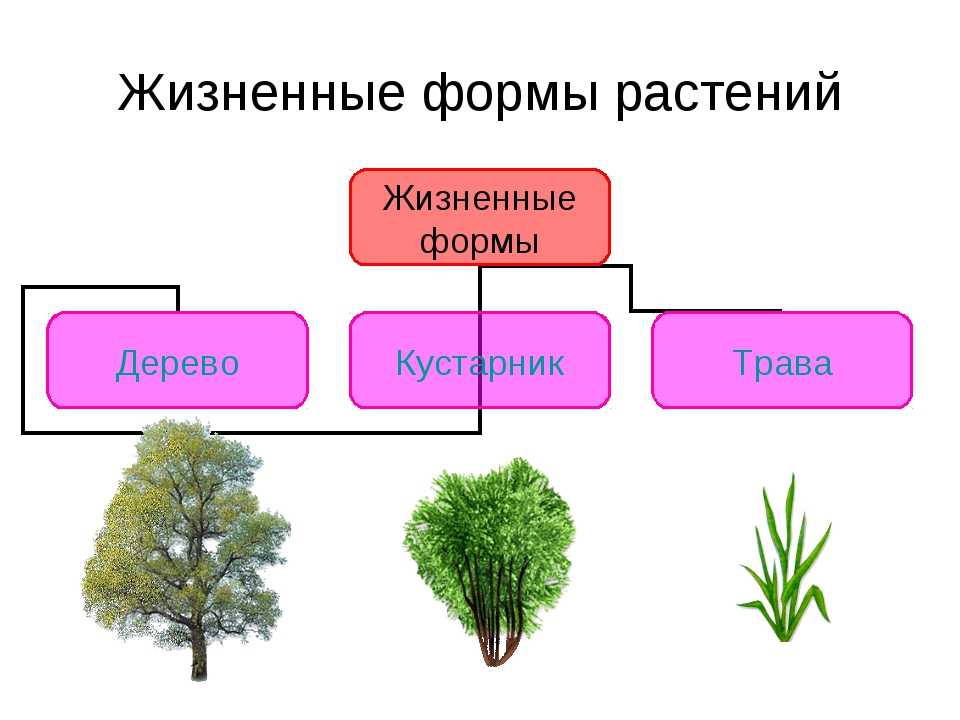 ИУК используется в качестве гормона укоренения для стимулирования роста придаточных корней на черенках и отдельных листьях. Применение синтетических ауксинов к растениям томатов в теплицах способствует нормальному развитию плодов. Внесение ауксина на открытом воздухе способствует синхронизации завязывания и опадения плодов, чтобы координировать сезон сбора урожая. Плоды, такие как огурцы без косточек, можно заставить завязывать плоды, обрабатывая неоплодотворенные цветки растений ауксинами.
ИУК используется в качестве гормона укоренения для стимулирования роста придаточных корней на черенках и отдельных листьях. Применение синтетических ауксинов к растениям томатов в теплицах способствует нормальному развитию плодов. Внесение ауксина на открытом воздухе способствует синхронизации завязывания и опадения плодов, чтобы координировать сезон сбора урожая. Плоды, такие как огурцы без косточек, можно заставить завязывать плоды, обрабатывая неоплодотворенные цветки растений ауксинами.
Цитокинины
О влиянии цитокининов впервые сообщили, когда было обнаружено, что добавление жидкого эндосперма кокосовых орехов к развивающимся зародышам растений в культуре стимулирует их рост. Было обнаружено, что стимулирующим фактором роста является цитокинин , гормон, который способствует цитокинезу (делению клеток). На сегодняшний день известно почти 200 встречающихся в природе или синтетических цитокининов. Цитокинины наиболее распространены в растущих тканях, таких как корни, зародыши и плоды, где происходит деление клеток.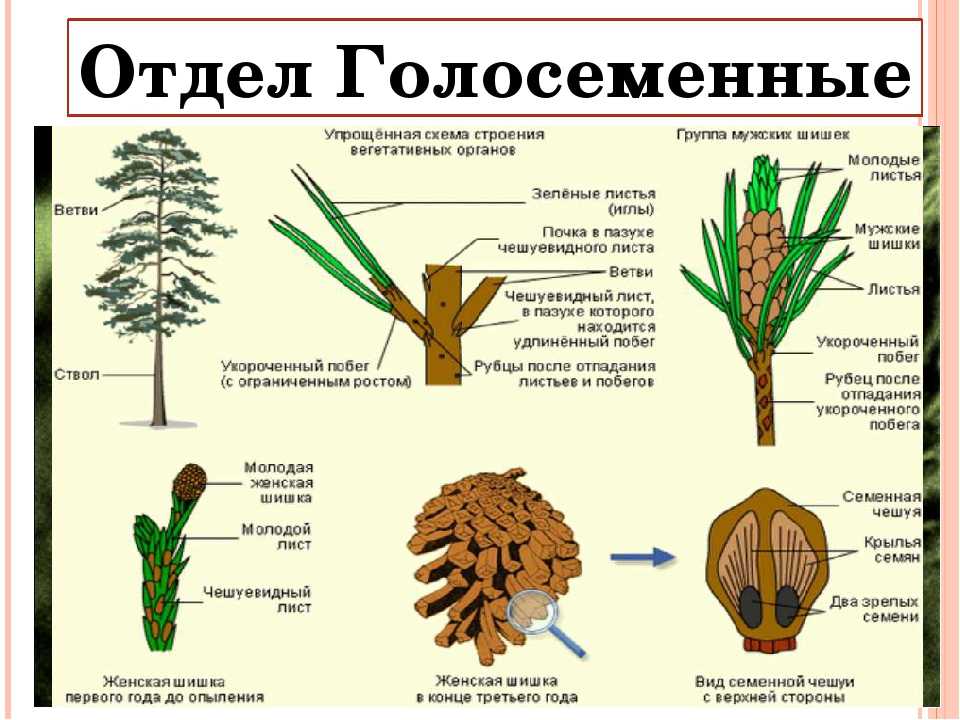 Известно, что цитокинины задерживают старение в тканях листа, способствуют митозу и стимулируют дифференцировку меристемы побегов и корней. Многие эффекты на развитие растений находятся под влиянием цитокининов либо в сочетании с ауксином, либо с другим гормоном. Например, апикальное доминирование, по-видимому, является результатом баланса между ауксинами, которые ингибируют боковые почки, и цитокининами, которые способствуют более густому росту.
Известно, что цитокинины задерживают старение в тканях листа, способствуют митозу и стимулируют дифференцировку меристемы побегов и корней. Многие эффекты на развитие растений находятся под влиянием цитокининов либо в сочетании с ауксином, либо с другим гормоном. Например, апикальное доминирование, по-видимому, является результатом баланса между ауксинами, которые ингибируют боковые почки, и цитокининами, которые способствуют более густому росту.
Гиббереллины
Рисунок 7. Внесение гибберелловой кислоты в виноград увеличивает размер плодов и ослабляет гроздья. (Фото: Bob Nichols, USDA)
Гиббереллины (GA) представляют собой группу из примерно 125 близкородственных растительных гормонов, которые стимулируют удлинение побегов, прорастание семян и созревание плодов и цветов. ГК синтезируются в апикальных меристемах корней и стеблей, молодых листьях и зародышах семян. В городских районах антагонисты ГА иногда наносят на деревья под линиями электропередач, чтобы контролировать рост и уменьшить частоту обрезки.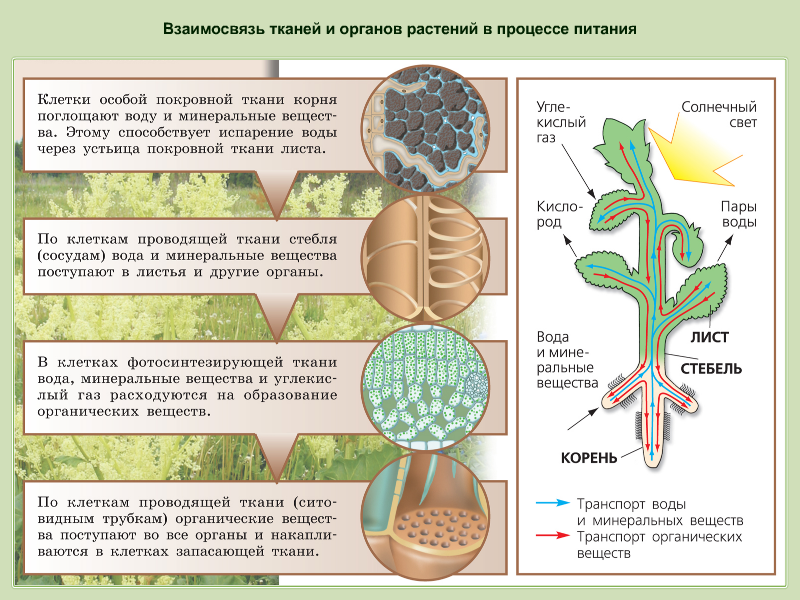
ГА нарушают покой (состояние задержки роста и развития) в семенах растений, которым для прорастания требуется воздействие холода или света. Абсцизовая кислота является сильным антагонистом действия ГК. Другие эффекты ГА включают выражение пола, развитие бессемянных плодов и задержку старения листьев и плодов. Виноград без косточек получают стандартными методами селекции и содержат незаметные семена, которые не развиваются. Поскольку ГА вырабатываются семенами, а развитие плодов и удлинение стебля находятся под контролем ГА, эти сорта винограда обычно дают маленькие плоды в компактных гроздьях. Созревающий виноград обычно обрабатывают ГА, чтобы увеличить размер плодов, а также получить более рыхлые грозди (более длинные стебли), что снижает вероятность заражения милдью (рис. 7).
Абсцизовая кислота
Растительный гормон абсцизовая кислота (АБК) была впервые обнаружена как агент, вызывающий опадение или опадение коробочек хлопка. Однако более поздние исследования показывают, что АБК играет лишь незначительную роль в процессе абсциссии. АБК накапливается в ответ на стрессовые условия окружающей среды, такие как обезвоживание, низкие температуры или укороченный световой день. Его активность противостоит многим стимулирующим рост эффектам ГК и ауксинов. АБК ингибирует удлинение стебля и вызывает состояние покоя боковых почек.
АБК накапливается в ответ на стрессовые условия окружающей среды, такие как обезвоживание, низкие температуры или укороченный световой день. Его активность противостоит многим стимулирующим рост эффектам ГК и ауксинов. АБК ингибирует удлинение стебля и вызывает состояние покоя боковых почек.
АБК вызывает состояние покоя семян, блокируя прорастание и стимулируя синтез запасных белков. Растениям, адаптированным к умеренному климату, требуется длительный период низких температур, прежде чем семена прорастут. Этот механизм предохраняет молодые растения от слишком раннего прорастания в не по сезону теплой зимней погоде. По мере того, как гормон постепенно разрушается в течение зимы, семя выходит из состояния покоя и прорастает весной, когда условия благоприятны. Еще одним эффектом АБК является содействие развитию зимних почек; он опосредует превращение апикальной меристемы в спящую почку. Низкая влажность почвы вызывает увеличение АБК, что приводит к закрытию устьиц, уменьшая потерю воды зимними почками.
Этилен
Этилен связан с созреванием плодов, увяданием цветов и листопадом. Этилен необычен, потому что это летучий газ (C 2 H 4 ). Сотни лет назад, когда на городских улицах устанавливали газовые уличные фонари, деревья, росшие близко к фонарным столбам, развивали искривленные, утолщенные стволы и сбрасывали листву раньше, чем предполагалось. Эти эффекты были вызваны испарением этилена из ламп.
Стареющие ткани (особенно стареющие листья) и узлы стеблей производят этилен. Однако наиболее известным эффектом гормона является стимуляция созревания плодов. Этилен стимулирует превращение крахмала и кислот в сахара. Некоторые люди хранят незрелые фрукты, такие как авокадо, в запечатанных бумажных пакетах, чтобы ускорить их созревание; газ, выделяемый первым созревающим плодом, ускорит созревание остальных плодов. Этилен также вызывает опадение листьев и плодов, увядание и опадение цветков, способствует прорастанию некоторых злаков и прорастанию луковиц и картофеля.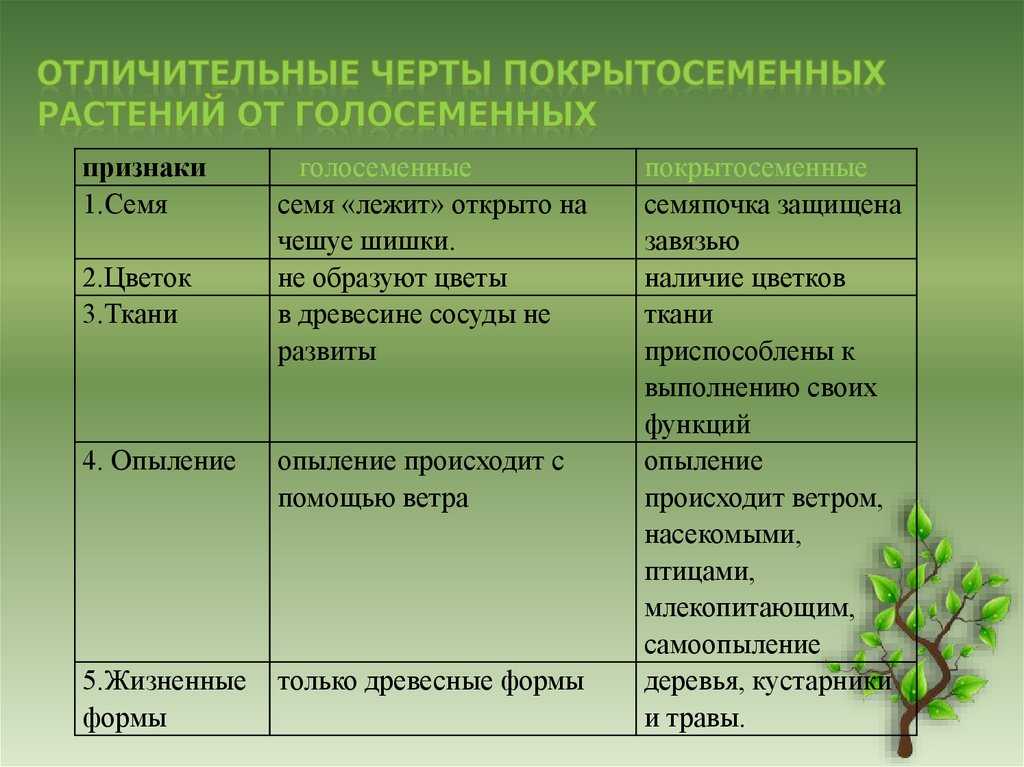
Этилен широко используется в сельском хозяйстве. Коммерческие садоводы контролируют сроки созревания фруктов с помощью газа. Садоводы подавляют опадение листьев у декоративных растений, удаляя этилен из теплиц с помощью вентиляторов и вентиляции.
Нетрадиционные гормоны
Недавние исследования обнаружили ряд соединений, которые также влияют на развитие растений. Их роль менее понятна, чем эффекты основных гормонов, описанных до сих пор.
Жасмонаты играют важную роль в защитных реакциях на травоядных. Их уровни увеличиваются, когда растение ранено хищником, что приводит к увеличению токсичных вторичных метаболитов. Они способствуют выработке летучих соединений, привлекающих естественных врагов хищников. Например, пережевывание гусеницами растений томата приводит к повышению уровня жасмоновой кислоты, что, в свою очередь, вызывает выделение летучих соединений, привлекающих хищников вредителя.
Олигосахарины также играют роль в защите растений от бактериальных и грибковых инфекций.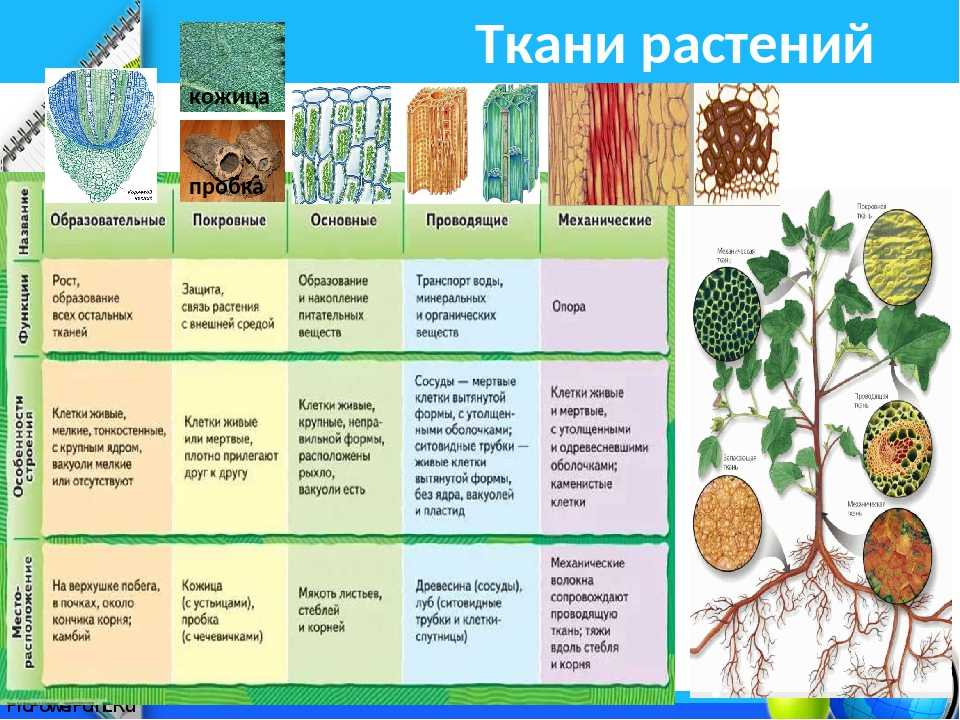 Они действуют локально в месте повреждения, а также могут транспортироваться в другие ткани. Стриголактоны способствуют прорастанию семян у некоторых видов и подавляют боковое верхушечное развитие в отсутствие ауксинов. Стриголактоны также играют роль в образовании микоризы, мутуалистической ассоциации корней растений и грибов. Брассиностероиды важны для многих процессов развития и физиологических процессов. Сигналы между этими соединениями и другими гормонами, особенно ауксином и ГК, усиливают их физиологический эффект. Гормоны положительно влияют на верхушечное доминирование, прорастание семян, гравитропизм и устойчивость к замораживанию. Рост корней и опадение плодов подавляются стероидами.
Они действуют локально в месте повреждения, а также могут транспортироваться в другие ткани. Стриголактоны способствуют прорастанию семян у некоторых видов и подавляют боковое верхушечное развитие в отсутствие ауксинов. Стриголактоны также играют роль в образовании микоризы, мутуалистической ассоциации корней растений и грибов. Брассиностероиды важны для многих процессов развития и физиологических процессов. Сигналы между этими соединениями и другими гормонами, особенно ауксином и ГК, усиливают их физиологический эффект. Гормоны положительно влияют на верхушечное доминирование, прорастание семян, гравитропизм и устойчивость к замораживанию. Рост корней и опадение плодов подавляются стероидами.
Проверьте свое понимание
Ответьте на вопросы ниже, чтобы узнать, насколько хорошо вы понимаете темы, затронутые в предыдущем разделе. В этом коротком тесте , а не учитываются при подсчете вашей оценки в классе, и вы можете пересдавать его неограниченное количество раз.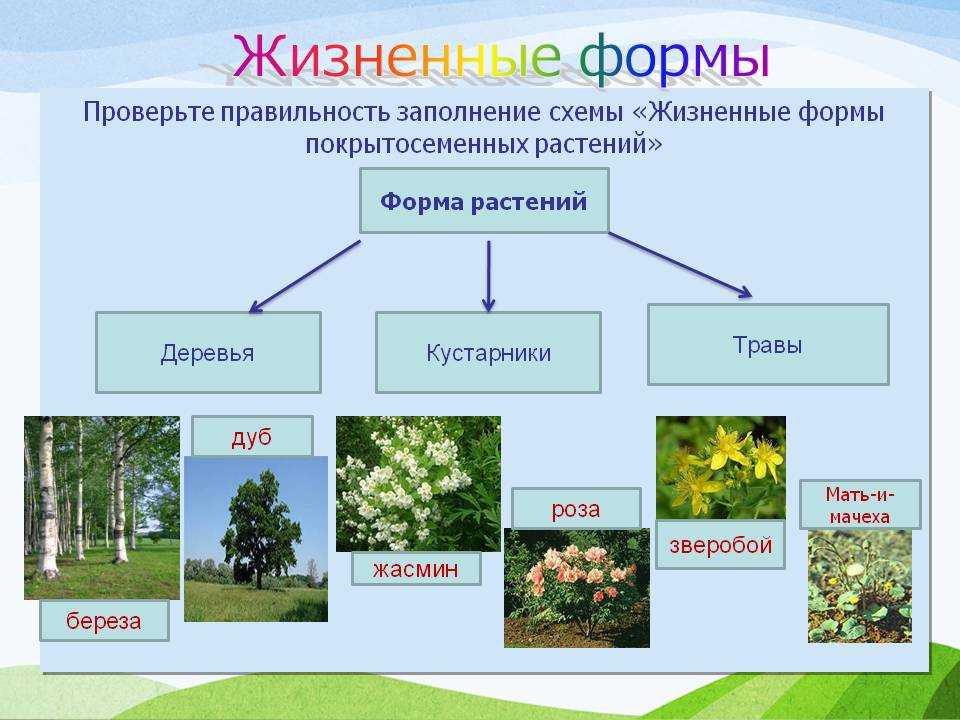
Используйте этот тест, чтобы проверить свое понимание и решить, следует ли (1) изучить предыдущий раздел дальше или (2) перейти к следующему разделу.
Деление растительной клетки: варианты кратчайшего симметричного пути
1. Hofmeister W. (1863) Zusatze und Berichtigungen zu den 1851 veroffentlichen Untersuchungen der Entwicklung hoherer Kryptogamen. Ярб Висс Бот
3, 259–293 [Google Scholar]
2. Sachs J. (1878) Ueber die anordnung der zellen in jüngsten pflanzentheilen. Арбайтен д. Бот. Инст
2, 46–104. Анналы бельжийского общества микроскопии
103, 822–824 [Google Scholar]
4. Фландерс Д. Дж., Роулинз Д. Дж., Шоу П. Дж. и Ллойд К. В. (1990) Связанные с ядром микротрубочки помогают определить плоскость деления эпидермальных клеток растений: предотвращение четырехсторонних соединений и роль геометрии клеток. Дж. Клеточная биология.
110, 1111–1122 гг.
10.1083/jcb.110.4.1111 [бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
5. Бессон С. и Дюме Дж. (2011) Универсальное правило симметричного деления клеток растений. проц. Натл. акад. науч. США.
и Дюме Дж. (2011) Универсальное правило симметричного деления клеток растений. проц. Натл. акад. науч. США.
108, 6294–6299
10.1073/pnas.1011866108 [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
6. Картер Р., Санчес-Корралес Ю.Э., Хартли М., Гринейзен В.А. и Мари А.Ф.М. (2017) Ячейки дорожного покрытия и загадка топологии. Разработка
144, 4386–4397
10.1242/dev.157073 [бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
7. Gibson W.T., Veldhuis J.H., Rubinstein B., Cartwright H.N., Perrimon N., Brodland G.W. и другие. (2011)Контроль плоскости митотического расщепления с помощью локальной топологии эпителия. Клетка
144, 427–438
10.1016/j.cell.2010.12.035 [бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
8. Jackson M.D.B., Duran-Nebreda S., Kierzkowski D., Strauss S., Xu H., Landrein B. et al. (2019) Глобальный топологический порядок возникает благодаря локальному механическому контролю клеточных делений в апикальной меристеме побега арабидопсиса. Сотовая система
Сотовая система
8, 53–65.e3
10.1016/j.cels.2018.12.009 [бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
9. Синнотт Э. У. и Блох Р. (1940) Цитоплазматическое поведение во время деления вакуолятных растительных клеток. проц. Натл. акад. науч. США.
26, 223–227
10.1073/pnas.26.4.223 [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
10. Venverloo C.J., Hovenkamp P.H., Weeda A.J. и Либбенга К.Р. (1980) Деление клеток в эксплантах наутилокаликса I. Фрагмосома, препрофазная полоса и плоскость деления. Z. Pflanzenphysiol.
100, 161–174
10.1016/S0044-328X(80)80209-1 [CrossRef] [Google Scholar]
11. Traas J.A., Doonan J.H., Rawlins D.J., Shaw P.J., Watts J. and Lloyd C.W. (1987). цитоплазма на протяжении всего клеточного цикла клеток моркови и связана с делящимся ядром. Дж. Клеточная биология.
105, 387–395
10.1083/jcb.105.1.387 [бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
12. Stoppin V., Vantard M., Schmit A.C. и Lambert A.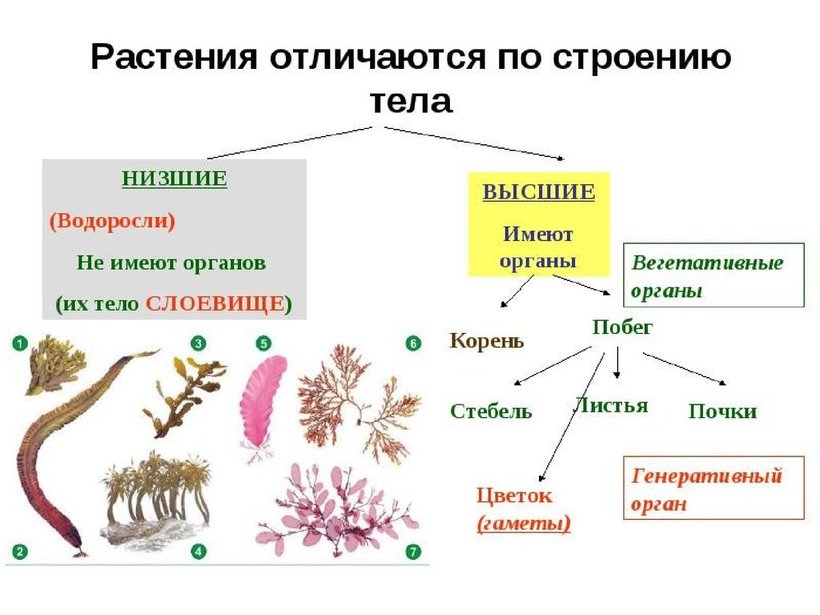 M. (1994) Сборка микротрубочек в изолированных ядрах растений: ядерная поверхность высших растений обладает активностью, подобной центросоме. Растительная клетка
M. (1994) Сборка микротрубочек в изолированных ядрах растений: ядерная поверхность высших растений обладает активностью, подобной центросоме. Растительная клетка
6, 1099–1106 гг.
10.2307/3869888 [бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
13. Пикетт-Хипс Дж. Д. и Норткот Д. Х. (1966) Организация микротрубочек и эндоплазматического ретикулума во время митоза и цитокинеза в меристемах пшеницы. Дж. Клеточные науки.
1, 109–120 PMID: [PubMed] [Google Scholar]
14. Caillaud M.-C. (2019) Анионные липиды: трубопровод, соединяющий ключевых участников деления растительных клеток. Фронт. Завод. науч.
10, 419
10.3389/fpls.2019.00419 [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
15. Gunning B.E.S. и Sammut M. (1990). Перестройки микротрубочек, участвующие в установлении плоскостей клеточного деления, начинаются сразу после синтеза ДНК и завершаются непосредственно перед митозом. Растительная клетка
2, 1273–1282 гг.
10.2307/3869345 [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
16. Miart F., Desprez T., Biot E., Morin H., Belcram K., Höfte H. et al. (2014)Пространственно-временной анализ синтеза целлюлозы во время формирования клеточной пластинки у арабидопсиса. Плант Дж.
Miart F., Desprez T., Biot E., Morin H., Belcram K., Höfte H. et al. (2014)Пространственно-временной анализ синтеза целлюлозы во время формирования клеточной пластинки у арабидопсиса. Плант Дж.
77, 71–84
10.1111/tpj.12362 [PubMed] [CrossRef] [Google Scholar]
17. Strasburger E. (1882) Ueber den theilungsvorgang der zellkerne und das verhältniss der kerntheilung zur zelltheilung. Архив микроскопической анатомии
21, 476–590
10.1007/BF02952628 [CrossRef] [Google Scholar]
18. Bajer A. (1965) Киномикрографический анализ образования клеточных пластинок в эндосперме. Эксп. Сотовый рез.
37, 376–398
10.1016/0014-4827(65)
-2 [PubMed] [CrossRef] [Google Scholar]
19. Смертенко А., Хьюитт С.Л., Жак С.Н., Кацпршик Р., Лю Ю., Марсец М.Дж. (2018) Динамика микротрубочек фрагмопластов — игра зон. Дж. Клеточные науки.
131, jcs203331
10.1242/jcs.203331 [бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
20. Лукас Дж.Р. и Сак Ф.Д. (2012)Полярное развитие полос препрофазы и клеточных пластин в эпидермисе листьев арабидопсиса. Плант Дж.
Плант Дж.
69, 501–509
10.1111/j.1365-313X.2011.04809.x [PubMed] [CrossRef] [Google Scholar]
21. Guérin A., Gravelle S. and Dumais J. (2016) Силы, лежащие в основе деления клеток растений. проц. Натл. акад. науч. США.
113, 8891–8893
10.1073/pnas.1609309113 [бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
22. Hamant O., Heisler M.G., Jönsson H., Krupinski P., Uyttewaal M., Bokov P. et al. (2008) Паттерн развития с помощью механических сигналов у арабидопсиса. Наука
322, 1650–1655 гг.
10.1126/наука.1165594 [PubMed] [CrossRef] [Google Scholar]
23. Hamant O., Inoue D., Bouchez D., Dumais J. and Mjolsness E. (2019) Являются ли микротрубочки датчиками натяжения?
Нац. коммун.
10, 2360
10.1038/s41467-019-10207-y [бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
24. Robinson S. and Kuhlemeier C. (2018) Глобальное сжатие переориентирует кортикальные микротрубочки в эпидермисе гипокотиля арабидопсиса и способствует росту . Курс. биол.
28, 1794–1802. e2
e2
10.1016/j.cub.2018.04.028 [PubMed] [CrossRef] [Google Scholar]
25. Verger S., Long Y., Boudaoud A. and Hamant O. (2018) Петля обратной связи между натяжением и адгезией в эпидермисе растений. электронная жизнь
7, е34460
10.7554/eLife.34460 [бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
26. Lloyd C.W. (1991) Лазерная микрохирургия демонстрирует, что цитоплазматические нити, закрепляющие ядро поперек вакуоли премитотических растительных клеток, находятся под напряжением. Последствия для выравнивания плоскости деления. Разработка
113, 931–939 [Google Scholar]
27. Ашкин А., Дзедзич Ю.М. (1989) Внутренние манипуляции с клетками с использованием инфракрасных лазерных ловушек. проц. Натл. акад. науч. США.
86, 7914–7918
10.1073/pnas.86.20.7914 [бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
28. Martinez P., Allsman L.A., Brakke K.A., Hoyt C., Hayes J., Liang H. et al. (2018) Прогнозирование плоскостей деления трехмерных клеток путем минимизации мыльной пленки.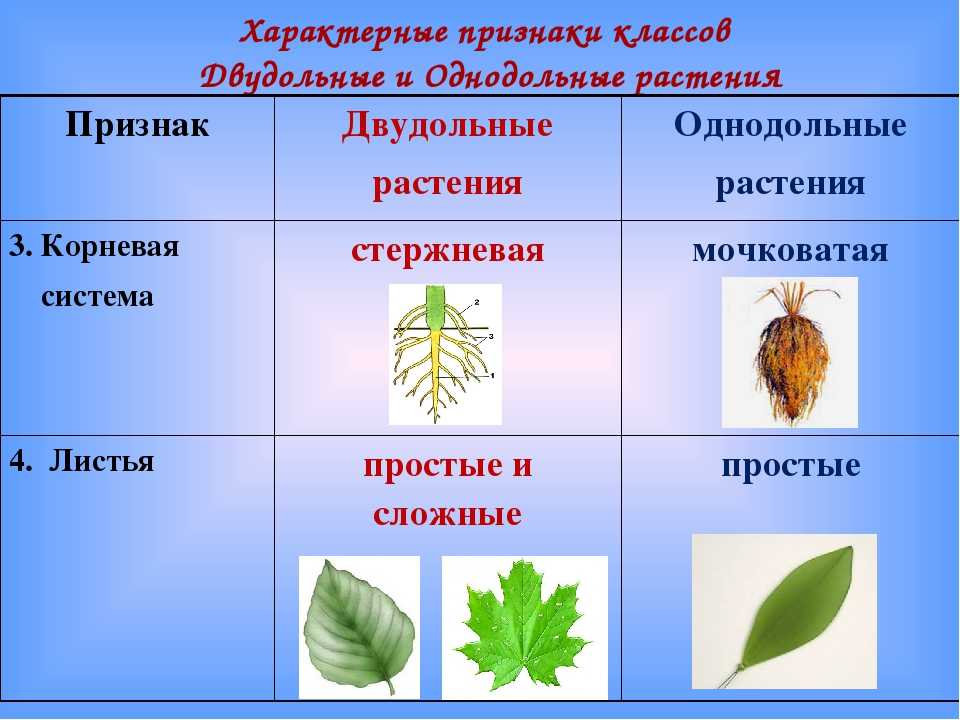 Растительная клетка
Растительная клетка
30, 2255–2266 гг.
10.1105/tpc.18.00401 [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
29. Jin Q. and Hasenstein K.H (2009) Изменения цитоскелета во время сперматогенеза у chara antheridia In The Plant Cytoskeleton: A Key Tool for Agro-Biotechnology (Blume Y.B., Baird W.V., Yemets A.I., Breviario D., eds), стр. 129–142, Springer, Дордрехт, Нидерланды [ Google Scholar]
30. Yoshida S., de Reuille PB., Lane B., Basssel G.W., Prusinkiewicz P., Smith R.S. и другие. (2014) Генетический контроль развития растений путем преодоления правила геометрического деления. Дев. Клетка
29, 75–87
10.1016/j.devcel.2014.02.002 [PubMed] [CrossRef] [Google Scholar]
31. Азимзаде Дж., Накри П., Христодулиду А., Древенсек С., Камиллери К., Амиур Н. и соавт. (2008) Белки TONNEAU1 Arabidopsis необходимы для формирования препрофазных полос и взаимодействуют с centrin. Растительная клетка
20, 2146–2159 гг.
10.1105/tpc.107.056812 [бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
32. Schaefer E., Belcram K., Uyttewaal M., Duroc Y., Goussot M., Legland D. et al. (2017) Препрофазная полоса микротрубочек контролирует надежность ориентации деления у растений. Наука
Schaefer E., Belcram K., Uyttewaal M., Duroc Y., Goussot M., Legland D. et al. (2017) Препрофазная полоса микротрубочек контролирует надежность ориентации деления у растений. Наука
356, 186–18910.1126/science.aal3016 [PubMed] [CrossRef] [Google Scholar]
33. Chan J., Calder G., Fox S. and Lloyd C. (2005) Локализация белка EB1, связывающего концы микротрубочек, выявляет альтернативные пути веретена развитие в суспензионных клетках арабидопсиса. Растительная клетка
17, 1737–1748 гг.
10.1105/tpc.105.032615 [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
34. Lloyd C. and Chan J. (2006) Не так разделены: общая основа деления клеток растений и животных. Нац. Преподобный Мол. Клеточная биол.
7, 147–152
10.1038/nrm1831 [PubMed] [CrossRef] [Google Scholar]
35. Ота Т. (1961) Роль цитоплазмы в цитокинезе растительных клеток. Цитология (Токио)
26, 428–447
10.1508/cytologia.26.428 [CrossRef] [Google Scholar]
36. Gunning B.E. и Вик С.М. (1985)Полосы препрофазы, фрагмопласты и пространственный контроль цитокинеза. Дж. Клеточные науки. Доп.
Дж. Клеточные науки. Доп.
2, 157–179
10.1242/jcs.1985.Supplement_2.9 [PubMed] [CrossRef] [Google Scholar]
37. Arima K., Tamaoki D., Mineyuki Y., Yasuhara H., Nakai T., Shimmen T. et al. (2018)Смещение митотических аппаратов центрифугированием выявляет кортикальную организацию актина во время цитокинеза в культивируемых клетках табака BY-2. Дж. Плант Рез.
131, 803–815
10.1007/s10265-018-1047-4 [PubMed] [CrossRef] [Google Scholar]
38. Палевиц Б.А. и Хеплер П.К. (1974) Контроль плоскости деления во время устьичной дифференциации у Allium. Хромосома
46, 297–326
10.1007/BF00284884 [CrossRef] [Google Scholar]
39. Ливанос П. и Мюллер С. (2019) Создание плоскости деления и цитокинез. Анну. Преподобный завод биол.
70, 239–267
10.1146/annurev-arplant-050718-100444 [PubMed] [CrossRef] [Google Scholar]
40. Мюллер С., Хан С. и Смит Л.Г. (2006) Два кинезина участвуют в пространственном контроле цитокинеза в Arabidopsis thaliana . Курс. биол.
16, 888–894
10. 1016/j.cub.2006.03.034 [PubMed] [CrossRef] [Google Scholar]
1016/j.cub.2006.03.034 [PubMed] [CrossRef] [Google Scholar]
41. Stöckle D., Herrmann A., Lipka E., Lauster T., Gavidia R., Zimmermann S. et al. (2016)Предполагаемые RopGAP влияют на выбор плоскости деления и взаимодействуют с кинезин-12 POK1. Нац. Растения
2, 16120
10.1038/nplants.2016.120 [PubMed] [CrossRef] [Google Scholar]
42. Lipka E., Gadeyne A., Stöckle D., Zimmermann S., De Jaeger G., Ehrhardt D.W. и другие. (2014) Белки класса кинезинов-12, ориентированные на фрагмопласт, переводят информацию о положении препрофазной полосы, чтобы установить зону коркового деления в Arabidopsis thaliana . Растительная клетка
26, 2617–2632
10.1105/tpc.114.124933 [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
43. Martinez P., Dixit R., Balkunde R.S., Zhang A., O’Leary S.E., Brakke K.A. и другие. (2020) TANGLED1 опосредует взаимодействия микротрубочек, которые могут способствовать позиционированию плоскости деления у кукурузы. Дж. Клеточная биология.
219, е201907184
10.1083/jcb.201907184 [бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
44. Herrmann A., Livanos P., Lipka E., Gadeyne A., Hauser M.-T., Van Damme D. . и другие. (2018) Двойной локализованный кинезин-12 POK2 играет несколько ролей во время клеточного деления и взаимодействует с MAP65-3. Представитель ЕМБО
19, е46085
10.15252/embr.201846085 [бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
45. Panteris E., Adamakis I.-D.S., Voulgari G. and Papadopoulou G. (2011) Роль катанина в растениях клеточное деление: организация микротрубочек в делящихся клетках корня мутантов fra2 и lue1 Arabidopsis thaliana . Цитоскелет (Хобокен)
68, 401–413
10.1002/cm.20522 [PubMed] [CrossRef] [Google Scholar]
46. Комис Г., Луптовчак И., Овечка М., Самаковли Д., Шамаёва О. и Шамай Ю. (2017) Влияние катанина на динамику корковые микротрубочки и митотические массивы в Arabidopsis thaliana , обнаруженный с помощью расширенной визуализации живых клеток.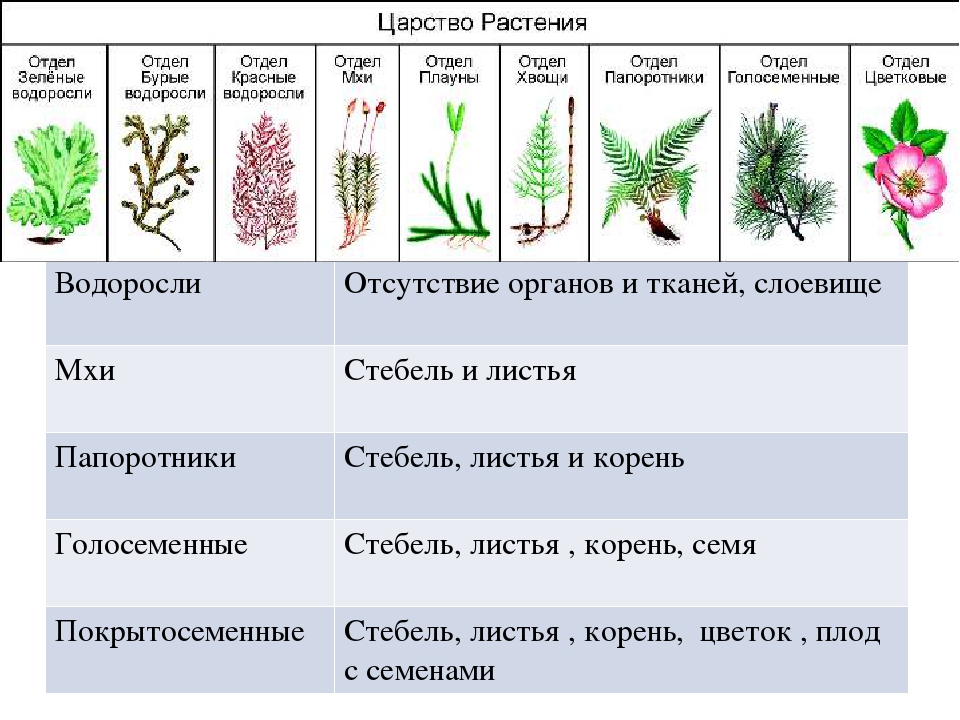 Фронт. Растениевод.
Фронт. Растениевод.
8, 866
10.3389/fpls.2017.00866 [бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
47. Sasaki T., Tsutsumi M., Otomo K., Murata T., Yagi N., Nakamura M. et al. (2019) Новый механизм связывания катанина ускоряет цитокинез. Курс. биол.
29, 4060–4070.e3
10.1016/j.cub.2019.09.049 [PubMed] [CrossRef] [Google Scholar]
48. Смит Л.Г., Хейк С. и Сильвестр А.В. (1996) Мутация tangled-1 изменяет ориентацию клеточных делений на всем протяжении развития листьев кукурузы без изменения формы листьев. Разработка
122, 481–489PMID: [PubMed] [Google Scholar]
49. Louveaux M., Julien J.-D., Mirabet V., Boudaoud A. and Hamant O. (2016) Ориентация плоскости клеточного деления на основе напряжения растяжения в Arabidopsis thaliana . проц. Натл. акад. науч. США.
113, Е4294–Е4303
10.1073/pnas.1600677113 [Статья бесплатно PMC] [PubMed] [CrossRef] [Google Scholar]
50. Квятковска Д. (2006) Формирование зачатка цветка на верхушке побега арабидопсиса: количественный анализ геометрии поверхности и роста.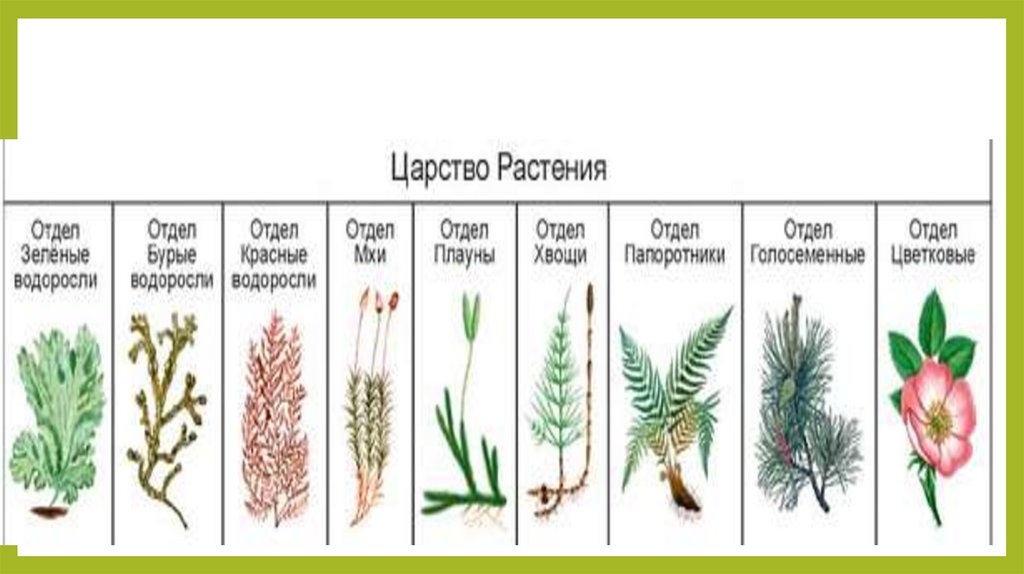 Дж. Эксп. Бот.
Дж. Эксп. Бот.
57, 571–580
10.1093/jxb/erj042 [PubMed] [CrossRef] [Google Scholar]
51. Квятковска Д. (2008) Цветение и динамика роста апикальной меристемы. Дж. Эксп. Бот.
59, 187–201
10.1093/jxb/erm290 [PubMed] [CrossRef] [Google Scholar]
52. Квятковска Д. и Рутье-Кежковска А.-Л. (2009) Морфогенез на верхушке побега соцветия Anagallis arvensis : геометрия поверхности и рост по сравнению с вегетативным побегом. Дж. Эксп. Бот.
60, 3407–3418
10.1093/jxb/erp176 [бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
53. Kwiatkowska D. и Dumais J. (2003) Рост и морфогенез на верхушке вегетативного побега Anagallis arvensis L. J. Exp. Бот.
54, 1585–1595 гг.
10.1093/jxb/erg166 [PubMed] [CrossRef] [Google Scholar]
54. Lintilhac P.M. и Весецкий Т.Б. (1984)Стресс-индуцированное выравнивание плоскости деления в растительных тканях, выращенных in vitro. Природа
307, 363–364
10.1038/307363a0 [CrossRef] [Google Scholar]
55. Гудбоди К.С. и Lloyd C.W. (1990) Актиновые филаменты выстраиваются вдоль эпидермальных клеток Tradescantia, предвосхищая индуцированные ранами плоскости деления. протоплазма
Гудбоди К.С. и Lloyd C.W. (1990) Актиновые филаменты выстраиваются вдоль эпидермальных клеток Tradescantia, предвосхищая индуцированные ранами плоскости деления. протоплазма
157, 92–101
10.1007/BF01322641 [CrossRef] [Google Scholar]
56. Venverloo C.J. (1990) Регуляция плоскости клеточного деления в вакуолизированных клетках. протоплазма
155, 85–94
10.1007/BF01322618 [CrossRef] [Google Scholar]
57. Landré P. (1972) Origine et développement des épidermes cotylédonaires et foliaires de la Moutarde ( Sinapis alba L.). Ультраструктурная дифференциация устьиц. Анна. науч. Нац. Бот. биол. Вег.
12, 247–322 [Google Scholar]
58. Ян М. и Сак Ф.Д. (1995) Мутации слишком много ртов и четыре губы влияют на образование устьиц у арабидопсиса. Растительная клетка
7, 2227–2239 [бесплатная статья PMC] [PubMed] [Google Scholar]
59. Nadeau J.A. и Сак Ф.Д. (2002) Развитие устьиц у арабидопсиса. Книга арабидопсиса
1, e0066
10.1199/tab.0066 [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
60. Стеббинс Г.Л. и Шах С.С. (1960) Исследования развития дифференцировки клеток в эпидермисе однодольных растений. Дев. биол.
Стеббинс Г.Л. и Шах С.С. (1960) Исследования развития дифференцировки клеток в эпидермисе однодольных растений. Дев. биол.
2, 477–500
10.1016/0012-1606(60)-6 [CrossRef] [Google Scholar]
61. Casimiro I., Marchant A., Bhalerao R.P., Beeckman T., Dhooge S., Swarup R. et al. (2001) Транспорт ауксина способствует инициации боковых корней арабидопсиса. Растительная клетка
13, 843–852
10.1105/tpc.13.4.843 [бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
62. Лукас М., Кеноби К., фон Вангенхайм Д., Вобет У., Сваруп К., Де Смет И. . и другие. (2013) Морфогенез боковых корней зависит от механических свойств покрывающих тканей. проц. Натл. акад. науч. США.
110, 5229–5234
10.1073/pnas.1210807110 [бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
63. von Wangenheim D., Fangerau J., Schmitz A., Smith R.S., Leitte H., Stelzer E.H.K. и другие. (2016) Правила и самоорганизующие свойства моделей постэмбрионального деления клеток органов растений.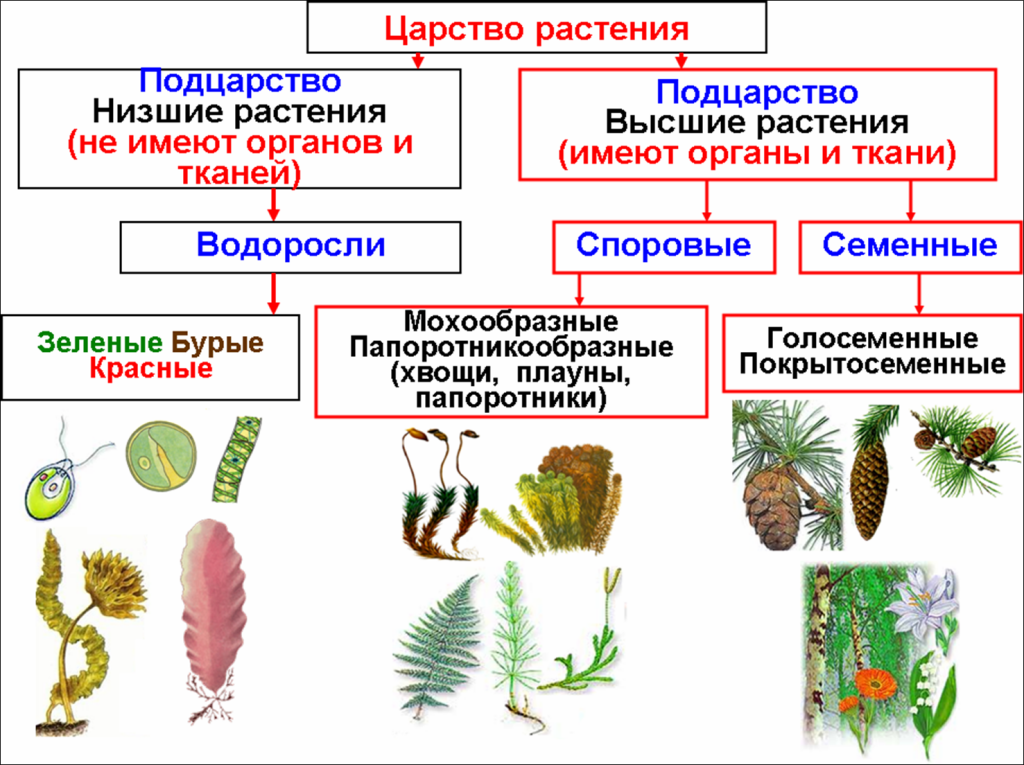 Курс. биол.
Курс. биол.
26, 439–449
10.1016/j.cub.2015.12.047 [PubMed] [CrossRef] [Google Scholar]
64. Мэнсфилд С.Г. и Бриарти Л.Г. (1991) Ранний эмбриогенез у Arabidopsis thaliana . II. Развивающийся эмбрион. Можно. Дж. Бот.
69, 461–476
10.1139/b91-063 [CrossRef] [Google Scholar]
65. Capron A., Chatfield S., Provart N. and Berleth T. (2009) Эмбриогенез: формирование структуры из одной клетки. Книга арабидопсиса
7, e0126
10.1199/tab.0126 [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
66. Мухтар Дж., Трубуил А., Белкрам К., Легланд Д., Хадир З., Урбейн А. и др. (2019)Геометрия клеток определяет симметричный и асимметричный выбор плоскости деления у ранних эмбрионов арабидопсиса. PLoS-компьютер. биол.
15, е1006771
10.1371/journal.pcbi.1006771 [бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
67. Chakrabortty B., Willemsen V., de Zeeuw T., Liao C.-Y., Weijers D., Mulder B. et al. (2018)Вероятный основанный на микротрубочках механизм ориентации клеточного деления в эмбриогенезе растений. Курс. биол.
Курс. биол.
28, 3031–3043.e2
10.1016/j.cub.2018.07.025 [PubMed] [CrossRef] [Google Scholar]
68. Робинсон С., де Рей П. Б., Чан Дж., Бергманн Д., Прусинкевич П. и Коэн Э. (2011 ) Генерация пространственных паттернов посредством переключения полярности клеток. Наука
333, 1436–1440 гг.
10.1126/science.1202185 [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
69. Мурояма А., Гонг Ю. и Бергманн Д.К. (2020)Противоположные ядерные миграции, обусловленные полярностью, лежат в основе асимметричных делений, образующих устьица арабидопсиса. Курс. биол.
30, 4467–4475.e4
10.1016/j.cub.2020.08.100 [бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
70. Роу М.Х., Донг Дж., Веймер А.К. и Бергманн Д.К. (2019) Модуль полярности, специфичный для растений, устанавливает асимметрию клеточных судеб в устьичной линии арабидопсиса. БиоРксив
10.1101/614636 [CrossRef] [Академия Google]
71. Донг Дж., Макалистер К.А. и Bergmann D.C. (2009) BASL контролирует асимметричное деление клеток у арабидопсиса. Клетка
Клетка
137, 1320–1330 гг.
10.1016/j.cell.2009.04.018 [бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
72. Cartwright H.N., Humphries J.A. и Смит Л.Г. (2009) PAN1: рецептороподобный белок, который способствует поляризации асимметричного деления клеток у кукурузы. Наука
323, 649–651
10.1126/science.1161686 [PubMed] [CrossRef] [Google Scholar]
73. Facette M.R., Park Y., Sutimantanapi D., Luo A., Cartwright H.N., Yang B. et al. (2015)Комплекс SCAR/WAVE поляризует рецепторы PAN и способствует асимметрии делений у кукурузы. Нац. Растения
1, 14024
10.1038/nplants.2014.24 [PubMed] [CrossRef] [Google Scholar]
74. Йошида С., ван дер Шурен А., ван Доп М., ван Гален Л., Сайга С., Адиби М. и соавт. (2019) Система координат на основе SOSEKI интерпретирует сигналы глобальной полярности у арабидопсиса. Нац. Растения
5, 160–166
10.1038/s41477-019-0363-6 [бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
75. Ван Доп М., Фидлер М., Мутте С., де Кейзер Дж. , Олиджслагер Л., Альбрехт С. и др. (2020) Полимеризация домена DIX стимулирует сборку комплексов полярности растительных клеток. Клетка
, Олиджслагер Л., Альбрехт С. и др. (2020) Полимеризация домена DIX стимулирует сборку комплексов полярности растительных клеток. Клетка
180, 427–439.e12
10.1016/j.cell.2020.01.011 [бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
76. Ле Дж., Лю С.-Г., Ян К.-З., Чен С.-Л., Цзоу Дж.-Дж., Ван Х.-З. и другие. (2014) Транспорт и активность ауксина регулируют формирование и развитие устьиц. Нац. коммун.
5, 3090
10.1038/ncomms4090 [PubMed] [CrossRef] [Google Scholar]
77. Малами Дж. Э. и Бенфи П. Н. (1997) Организация и дифференцировка клеток в боковых корнях Arabidopsis thaliana . Разработка
124, 33–44 PMID: [PubMed] [Google Scholar]
78. Дубровский Дж.Г., Дёрнер П.В., Колон-Кармона А. и Рост Т.Л. (2000)Пролиферация клеток перицикла и инициация боковых корней у арабидопсиса. Завод Физиол.
124, 1648–1657 гг.
10.1104/pp.124.4.1648 [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
79. Дубровский Ю.Г., Зауэр М., Напсучиали-Мендивиль С. , Иванченко М.Г., Фримл Ю., Шишкова С. и соавт. (2008) Auxin действует как локальный морфогенетический триггер для определения клеток-основателей боковых корней. проц. Натл. акад. наук. США.
, Иванченко М.Г., Фримл Ю., Шишкова С. и соавт. (2008) Auxin действует как локальный морфогенетический триггер для определения клеток-основателей боковых корней. проц. Натл. акад. наук. США.
105, 8790–8794
10.1073/pnas.0712307105 [бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
80. Marhavy P., Montesinos J.C., Abuzeineh A., Van Damme D., Vermeer J.E.M., Duclercq J. et al. (2016) Направленная элиминация клеток выявляет управляемый ауксином двухфазный способ инициации боковых корней. Гены Дев.
30, 471–483
10.1101/гад.276964.115 [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
81. Möller B.K., Ten Hove C.A., Xiang D., Williams N., López L.G., Yoshida S. et al. (2017) Клетка ответа на ауксин автономно контролирует инициацию основной ткани у раннего эмбриона арабидопсиса. проц. Натл. акад. науч. США.
114, Е2533–Е2539
10.1073/pnas.1616493114 [PMC free article] [PubMed] [CrossRef] [Google Scholar]
82. Овечка М., Луптовчак И., Комис Г., Шамаёва О. , Самаковли Д. и Шамай Ю. (2020) Пространственно-временной паттерн эктопических клеточных делений способствует неправильной форме фенотипа первичных и боковых корней мутанта katanin1. Фронт. Растениевод.
, Самаковли Д. и Шамай Ю. (2020) Пространственно-временной паттерн эктопических клеточных делений способствует неправильной форме фенотипа первичных и боковых корней мутанта katanin1. Фронт. Растениевод.
11 734
10.3389/fpls.2020.00734 [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
83. Bringmann M. and Bergmann D.C. (2017) Механические силы во всей ткани влияют на полярность устьичных стволовых клеток у арабидопсиса. Курс. биол.
27, 877–883
10.1016/j.cub.2017.01.059 [PubMed] [CrossRef] [Google Scholar]
84. Nakayama N., Smith R.S., Mandel T., Robinson S., Kimura S., Boudaoud A. et al. (2012) Механическая регуляция роста, опосредованного ауксином. Курс. биол.
22, 1468–1476 гг.
10.1016/j.cub.2012.06.050 [PubMed] [CrossRef] [Google Scholar]
85. Heisler M.G., Hamant O., Krupinski P., Uyttewaal M., Ohno C., Jönsson H. et al. (2010) Выравнивание между полярностью PIN1 и ориентацией микротрубочек в апикальной меристеме побега выявляет тесную связь между морфогенезом и транспортом ауксина.