
Содержание
2. Проводящие ткани. Проводящие пучки. Проводящие элементы ксилемы и флоэмы.
Проводящие
ткани. Значение и разнообразие проводящих
тканей
Проводящие
ткани являются важнейшей составной
частью большинства высших растений.
Они являются обязательным структурным
компонентом вегетативных и репродуктивных
органов споровых и семенных растений.
Проводящие ткани в совокупности с
клеточными стенками и межклетниками,
некоторыми клетками основной паренхимы
и специализированными передаточными
клетками образуют проводящую систему,
которая обеспечивает дальний и радиальный
транспорт веществ. Благодаря особой
конструкции клеток и их расположению
в теле растений проводящая система
выполняет многочисленные, но взаимосвязанные
функции:
1)
передвижение воды и минеральных веществ,
поглощенных корнями из почвы, а также
органических веществ, образуемых в
корнях, в стебель, листья, репродуктивные
органы;
2)
передвижение продуктов фотосинтеза из
зелёных частей растения в места их
использования и запасания: в корни,
стебли, плоды и семена;
3)
передвижение фитогормонов по растению,
что создает определённый их баланс,
который определяет темпы роста и развития
вегетативных и репродуктивных органов
растений;
4)
радиальный транспорт веществ из
проводящих тканей в рядом расположенные
живые клетки других тканей, например,
в ассимилирующие клетки мезофилла листа
и делящиеся клетки меристем. В нем могут
В нем могут
также принимать участие паренхимные
клетки сердцевинных лучей древесины и
коры. Большое значение в радиальном
транспорте имеют передаточные клетки
с многочисленными выпячиваниями
клеточной оболочки, находящиеся между
проводящими и паренхимными тканями;
5)
проводящие ткани повышают устойчивость
органов растений к деформирующим
нагрузкам;
6)
проводящие ткани образуют непрерывную
разветвленную систему, связывающую
органы растений в единое целое;
В
зависимости от выполняемых функций
проводящие ткани подразделяются на
ткани восходящего тока и ткани нисходящего
тока. Основным назначением тканей
восходящего тока является транспорт
воды и растворенных в ней минеральных
веществ от корня к выше расположенным
надземным органам. Кроме того, по ним
перемещаются органические вещества,
образуемые в корне и стебле, например,
органические кислоты, углеводы и
фитогормоны. Однако термин «восходящий
ток» не следует воспринимать однозначно
как передвижение снизу — вверх. Ткани
восходящего тока обеспечивают поток
веществ по направлению от зоны всасывания
к апексу побега. При этом транспортируемые
При этом транспортируемые
вещества используются как самим корнем,
так и стеблем, ветвями, листьями,
репродуктивными органами, независимо
от того, выше или ниже уровня корней они
находятся. Например, у картофеля вода
и элементы минерального питания поступают
по тканям восходящего тока в столоны и
клубни, образуемые в почве, а также в
надземные органы.
Ткани
нисходящего тока обеспечивают отток
продуктов фотосинтеза в растущие части
растений и в запасающие органы.
Проводящие
ткани восходящего тока
К
тканям восходящего тока относятся
трахеиды и сосуды (трахеи), которые
располагаются в древесинной (ксилемной)
части органов растений. В этих тканях
передвижение воды и растворенных в ней
веществ происходит пассивно под действием
корневого давления и испарением воды
с поверхности растения.
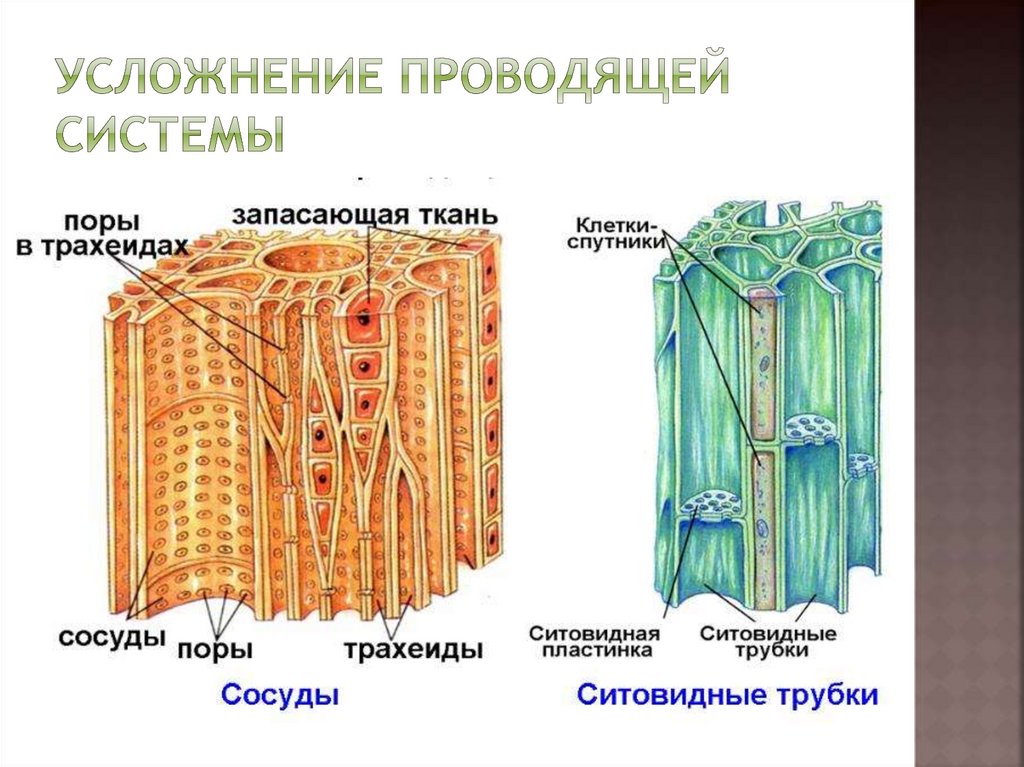
Трахеиды
имеют более древнее происхождение. Они
встречаются у высших споровых растений,
голосеменных и реже — у покрытосеменных.
У покрытосеменных они типичны для
мельчайших разветвлений жилок листа.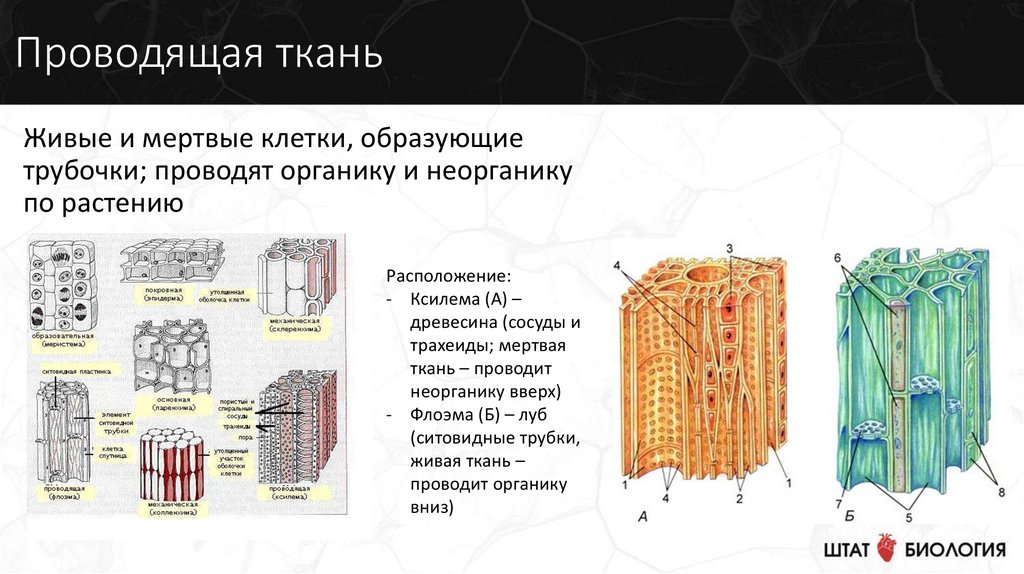
Клетки трахеид мертвые. Они имеют
вытянутую, часто веретеновидную форму.
Их длина составляет 1 — 4 мм. Однако у
голосеменных, например у араукарии, она
достигает 10 мм. Стенки клеток толстые,
целлюлозные, часто пропитываются
лигнином. В клеточных оболочках имеются
многочисленные окаймленные поры.
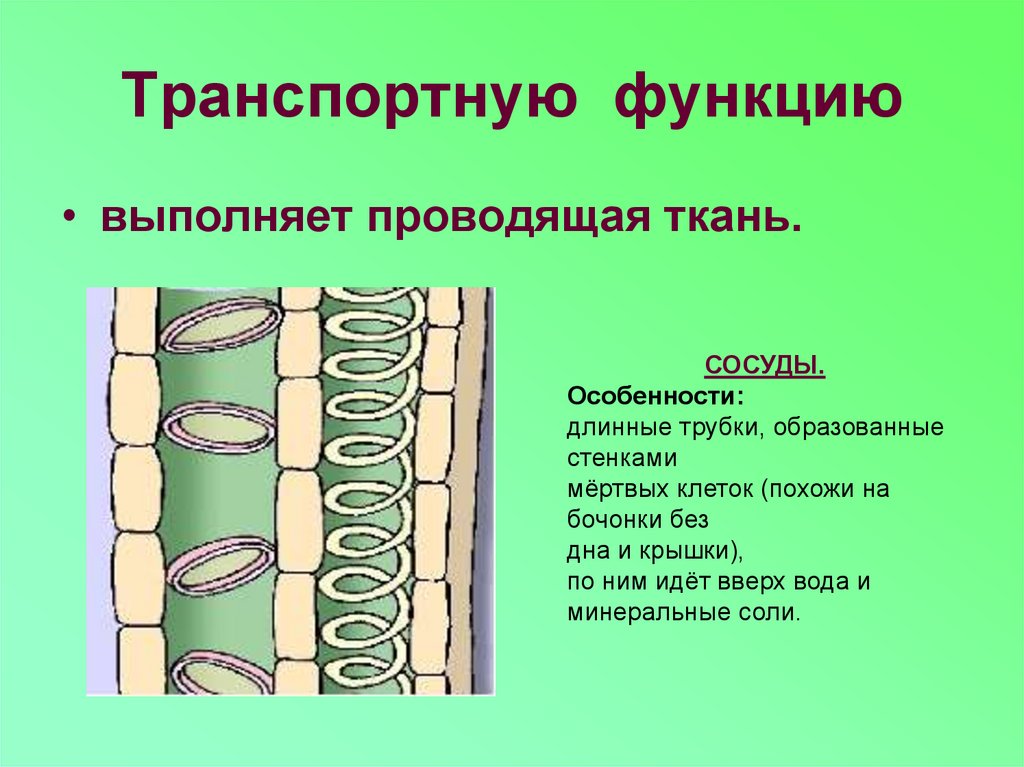
Сосуды
сформировались на более поздних этапах
эволюции. Они характерны для покрытосеменных,
хотя встречаются и у некоторых современных
представителей отделов Плауны (род
Селлагинелла), Хвощи, Папоротники и
Голосеменные (род Гнетум).
Сосуды
состоят из удлиненных мертвых клеток,
расположенных одна над другой и называемых
члениками сосуда. В торцевых стенках
члеников сосуда имеются крупные сквозные
отверстия — перфорации, через которые
осуществляется дальний транспорт
веществ. Перфорации возникли в ходе
эволюции из окаймленных пор трахеид. В
составе сосудов они бывают лестничными
и простыми. Многочисленные лестничные
перфорации образуются на торцевых
стенках члеников сосуда при их косом
заложении.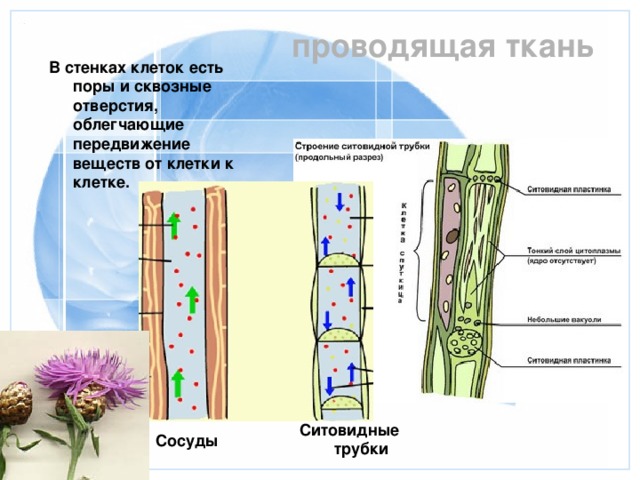
Боковые
стенки имеют неравномерные целлюлозные
утолщения, которые предохраняют сосуды
от избыточного давления, создаваемого
рядом расположенными живыми клетками
других тканей. В боковых стенках могут
находиться многочисленные поры,
обеспечивающие выход воды за пределы
сосуда.
Сосуды
имеют ограниченный срок деятельности.
Они могут разрушаться в результате
закупорки тиллами — выростами соседних
паренхимных клеток, а также под действием
центростремительных сил давления новых
клеток древесины, образуемых камбием.
В ходе эволюции сосуды подвергаются
изменениям. Членики сосудов становятся
короче и толще, косые поперечные
перегородки сменяются прямыми, а
лестничные перфорации — простыми.
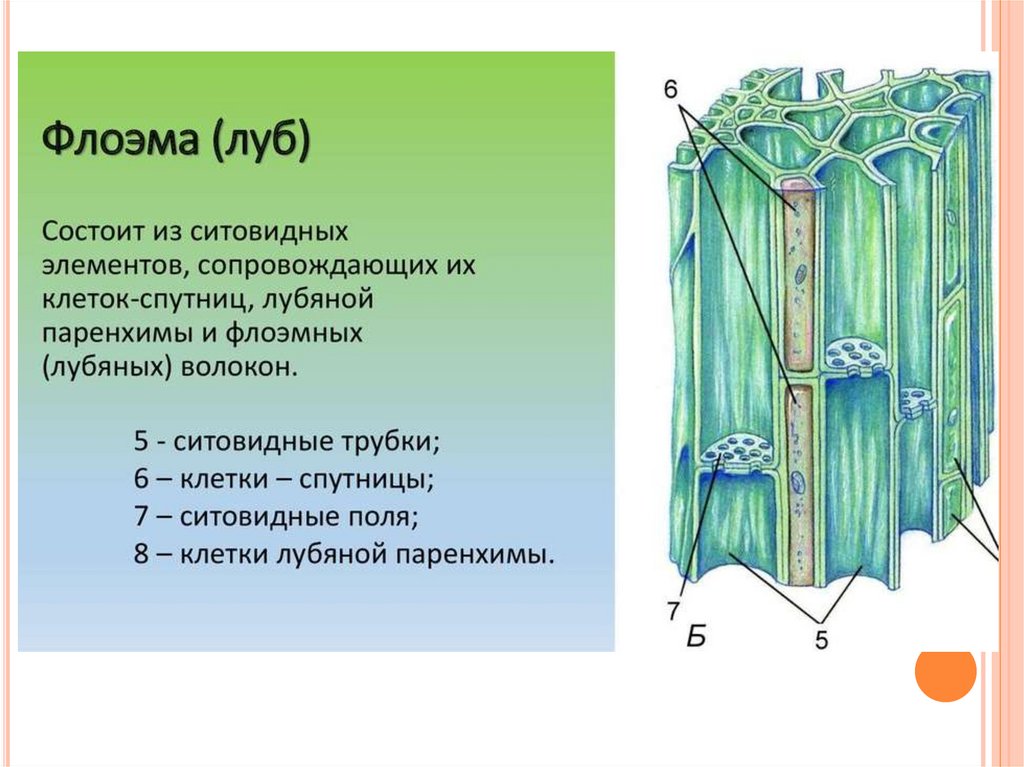
Проводящие
ткани нисходящего тока
К
тканям нисходящего тока относятся
ситовидные клетки и ситовидные трубки
с клетками-спутницами. Ситовидные клетки
имеют более древнее происхождение. Они
встречаются у высших споровых растений
и голосеменных. Это живые, удлиненные
клетки с заостренными концами.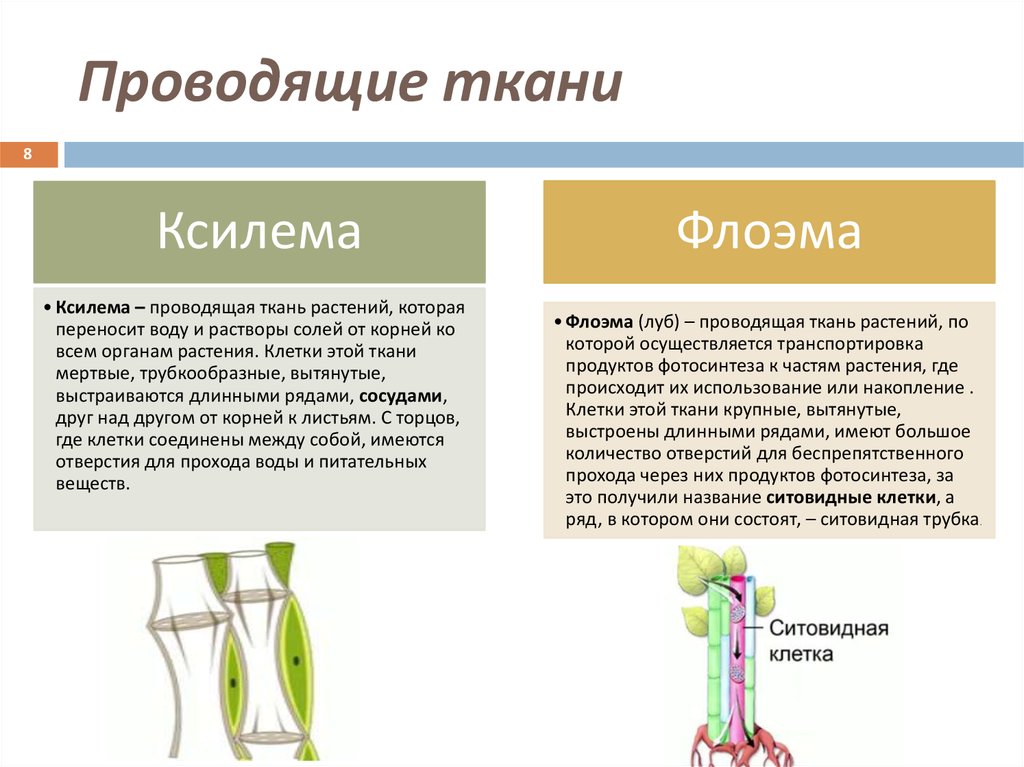 В зрелом
В зрелом
состоянии они содержат ядра в составе
протопласта. В их боковых стенках, в
местах соприкосновения смежных клеток,
имеются мелкие сквозные перфорации,
которые собраны группами и образуют
ситовидные поля, через которые
осуществляется передвижение веществ.
Ситовидные
трубки состоят из вертикального ряда
удлиненных клеток, разделенных между
собой поперечными стенками и называемыми
ситовидными пластинками, в которых
расположены ситовидные поля. Если
ситовидная пластинка обладает одним
ситовидным полем, она считается простой,
а если несколькими — то сложной. Ситовидные
поля образуются многочисленными
сквозными отверстиями — ситовидными
перфорациями небольшого диаметра. Через
эти перфорации из одной клетки в другую
проходят плазмодесмы. На стенках
перфораций размещается полисахарид
каллоза, которая уменьшает просвет
перфораций. По мере старения ситовидной
трубки каллоза полностью закупоривает
перфорации и трубка прекращает работу.
При
формировании ситовидной трубки в
образующих их клетках синтезируется
специальный флоэмный белок (Ф-белок) и
развивается крупная вакуоль. Она
Она
оттесняет цитоплазму и ядро к стенке
клетки. Затем мембрана вакуоли разрушается
и внутреннее пространство клетки
заполняется смесью цитоплазмы и
клеточного сока. Тельца Ф-белка теряют
отчетливые очертания, сливаются, образуя
тяжи около ситовидных пластинок. Их
фибриллы проходят через перфорации из
одного членика ситовидной трубки в
другой. К членикам ситовидной трубки
плотно прилегают одна или две
клетки-спутницы, которые имеют удлиненную
форму, тонкие стенки и живую цитоплазму
с ядром и многочисленными митохондриями.
Проводящие
пучки
Проводящие
ткани располагаются в органах растений
в виде продольных тяжей, образуя
проводящие пучки. Различают четыре типа
проводящих пучков: простые, общие,
сложные и сосудисто-волокнистые.
Простые
пучки состоят из одного типа проводящих
тканей. Например, в краевых частях
листовых пластинок многих растений
встречаются небольшие по диаметру пучки
из сосудов и трахеид, а в цветоносных
побегах у лилейных — из одних лишь
ситовидных трубок.
Общие
пучки образуются трахеидами, сосудами
и ситовидными трубками. Иногда этот
термин используется для обозначения
пучков метамера, которые проходят в
междоузлии и являются листовыми следами.
В состав сложных пучков входят проводящие
и паренхимные ткани. Наиболее совершенными,
многообразными по строению и местоположению
являются сосудисто-волокнистые пучки.
В
таких пучках выделяются функционально
разные части — флоэма и ксилема. Флоэма
обеспечивает отток ассимилятов из листа
и передвижение их в места использования
или запасания. По ксилеме вода и
растворенные в ней вещества передвигаются
из корневой системы в лист и другие
органы. Объем ксилемной части в несколько
раз превосходит объем флоэмной, поскольку
объем поступающей в растение воды
превышает объем образуемых ассимилятов,
так как значительная часть воды испаряется
растением.
Разнообразие
сосудисто-волокнистых пучков определяется
их происхождением, гистологическим
составом и местонахождением в растении.
Если пучки образуются из прокамбия и
завершают своё развитие по мере
использования запаса клеток образовательной
ткани, как у однодольных, они называются
закрытыми для роста.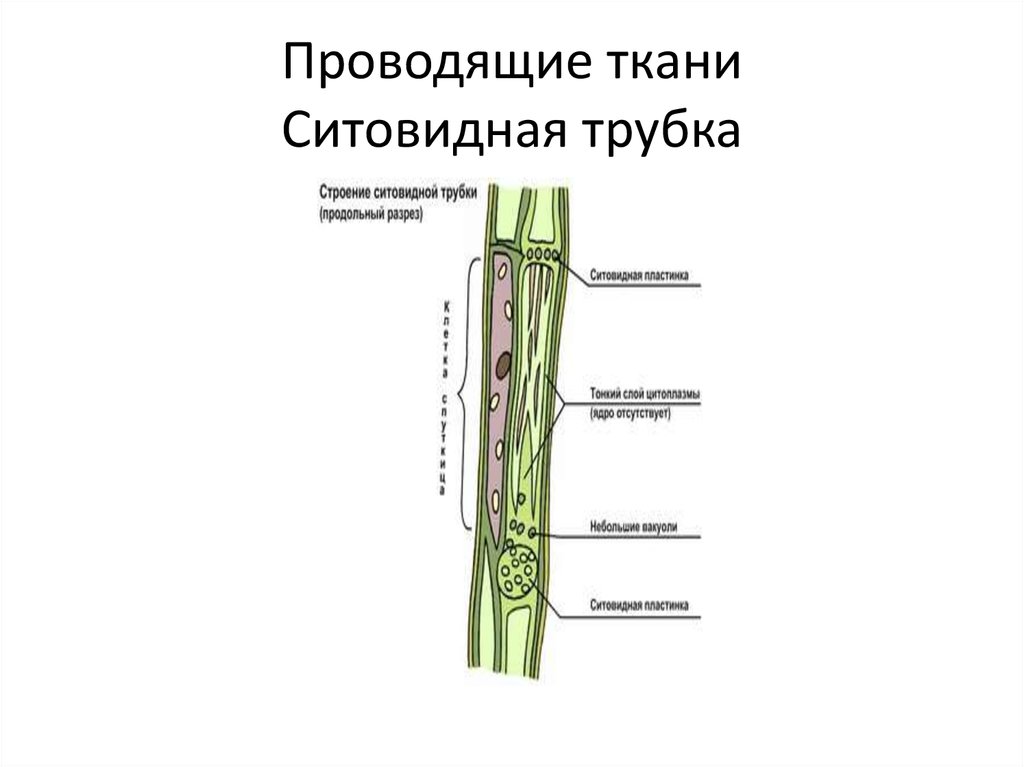 В отличие от них,
В отличие от них,
у двудольных открытые пучки не ограничены
в росте, поскольку они формируются
камбием и увеличиваются в диаметре на
протяжении всей жизни растения. В состав
сосудисто-волокнистых пучков кроме
проводящих могут входить основные и
механические ткани. Например, у двудольных
флоэма образуется ситовидными трубками
(проводящая ткань восходящего тока),
лубяной паренхимой (основная ткань) и
лубяными волокнами (механическая ткань).
В состав ксилемы входят сосуды и трахеиды
(проводящая ткань нисходящего тока),
древесинная паренхима (основная ткань)
и древесинные волокна (механическая
ткань). Гистологический состав ксилемы
и флоэмы генетически детерминирован и
может быть использован в систематике
растений для диагностики разных таксонов.
Кроме того, степень развития составных
частей пучков может изменяться под
влиянием условий произрастания растений.
Экзаменационные вопросы по курсу общей ботаники (высшие растения) ЭКОЛОГИ (2016/2017 уч. год)
Экзаменационные вопросы по курсу общей ботаники (высшие растения)
по направлению подготовки “ЭКОЛОГИЯ И ПРИРОДОПОЛЬЗОВАНИЕ”
cкачать файл. docx
docx
Модуль 1. Растительные клетки и ткани
- Отличительные особенности растительной клетки. Пластиды, их типы, ультрамикроскопическое строение, выполняемые функции. Взгляды на происхождение пластид. Тургесцентность растительной клетки.
- Клеточная оболочка. Порядок формирования и особенности строения первичной и вторичной оболочки. Строение плазмодесмы. Строение поры, поровой мембраны, простые и окаймленные поры.
- Формирование клеточной стенки при цитокинезе. Плазмодесмы, первичные поровые поля, поры. Химические изменения оболочки: лигнификация, суберинизация, кутинизация, кутикуляризация, минерализация.
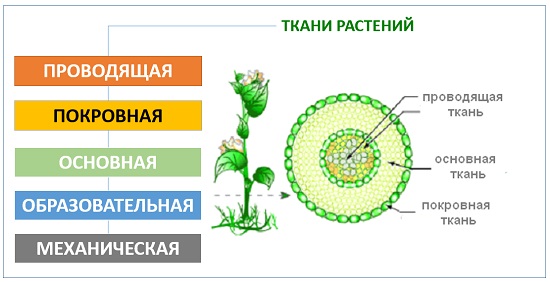
- Понятие ткани, типы растительных тканей. Образовательные и постоянные ткани. Понятие о первичных и вторичных, простых и сложных тканях. Идиобласты.
- Меристемы и полумеристемы: роль в жизни растений, принципы организации, локализация в теле растения; классификация меристем. Цитологические особенности меристематических клеток.
 Зона интеркалярного роста.
Зона интеркалярного роста. - Покровные ткани. Эпидерма, её функции, типы слагающих её клеток. Общее представление о строении и функционировании устьиц, разнообразие устьичных аппаратов. Кутикула, её роль в жизнедеятельности растения. Трихомы и эмергенцы. Особенности ризодермы и экзодермы корня. Перидерма. Строение и функционирование чечевички.
- Механические ткани. Колленхима, типы. Склеренхима, особенности волокон и склереид. Сходство и различия в принципах функционирования и строении клеток колленхимы и склеренхимы. Закономерности расположения механических тканей в различных органах растения. Механическая функция флоэмы и ксилемы и гистологические элементы, ее выполняющие.
- Разнообразие постоянных тканей: система выделения веществ, фотосинтезирующие и запасающие ткани. Абсорбционные ткани, ткани “проветривания”.
- Запасающие ткани, их типы и локализация в теле растения. Форма и место отложения различных типов запасных питательных веществ в растительной клетке.
- Организация газообмена с внешней средой. Ближний и дальний транспорт газов в растении.
- Абсорбционные ткани. Веламен. Абсорбционная функция ризодермы (эпиблемы).
- Аэренхима, её функциональная нагрузка и локализация в теле растения. Особенности анатомического строения пневматофоров.
- Понятие об эргастических веществах растительной клетки. Различные формы отложения оксалата кальция.
- Проблемы секреции у растений. Экзогенные и эндогенные секреторные структуры. Железистые трихомы, нектарники, гидатоды, солевыводящие структуры (желёзки и волоски). Схизогенные и лизигенные секреторные каналы, млечники.
- Ксилема и флоэма: типы проводящих элементов, их отличия, развитие и строение. Первичные и вторичные проводящие ткани и меристемы, их образующие. Полифункциональность проводящих тканей, их паренхимная и механическая составляющая. Особенности взаимного расположение проводящих тканей в органах растения: типы проводящих пучков.
- Ксилема: типы проводящих элементов, их отличия и строение у разных таксонов высших растений. Окаймленные поры и перфорационные пластинки, типы перфорационных пластинок. Дифференциация прото- и метаксилемы. Первичная и вторичная ксилема, происхождение.
- Флоэма: типы проводящих элементов, их отличия и строение у разных таксонов высших растений. Ситовидные поля и ситовидные пластинки, типы ситовидных пластинок. Особенности функционирования флоэмы: клетки Страсбургера и сопровождающие клетки. Дифференциация прото- и метафлоэмы. Первичная и вторичная флоэма, происхождение.
- Разнообразие перфорационных пластинок члеников сосудов высших растений. Эволюционные преобразования перфорационных пластинок. Возрастные изменения в проводящих элементах ксилемы и флоэмы – затилловывание и образование мозолистых тел.
- Трансфузионнная ткань как разновидность проводящей ткани, её строение, функции, локализация.
- Понятие о метаморфозе у растений. Основные метаморфозы побега, корня и листа с примерами. Аналогия и гомология, критерии гомологии. Примеры аналогичных и гомологичных органов.
 Зона интеркалярного роста.
Зона интеркалярного роста.
 Основные метаморфозы побега, корня и листа с примерами. Аналогия и гомология, критерии гомологии. Примеры аналогичных и гомологичных органов.
Основные метаморфозы побега, корня и листа с примерами. Аналогия и гомология, критерии гомологии. Примеры аналогичных и гомологичных органов.
Модуль 2. Вегетативные органы высших растений
- Выход растений на сушу. Первые наземные растения на примере представителей риниофитов. Их отличие от зеленых водорослей. Проблемы, связанные с существованием в новой среде обитания и реализованные пути их решения.
- Корень, разнообразие его функций. Алло- и гоморизия; типы корневых систем. Ветвление корня. Морфофункциональные зоны молодого корня, функции корневого чехлика. Строение, местонахождение и функции ризодермы. Строение и функции корневого чехлика.
- Анатомо-топографические зоны корня. Первичное строение корня, особенности строения и функции экзодермы и эндодермы. Строение стелы корня. Особенности строения стелы корня двудольных растений при первичном строении в сравнении со стелой однодольных.
- Радиальный транспорт веществ в корне. Понятие апопласта и симпласта. Экзодерма и эндодерма, их роль в “загрузке” проводящих элементов ксилемы. Строение и функции клеток эндодермы зоне поглощения (корневых волосков). Возрастные изменения в эндодерме и первичной коре у однодольных и двудольных покрытосеменных растений.
- Функции перицикла в корне двудольных растений. Вторичное утолщение корня, роль камбия и феллогена. Строение корня двудольного растения после заложения и развития вторичных меристем.
- Современные взгляды на происхождение корня. Корнеподобные образования у современных и ископаемых высших споровых растений. Метаморфозы корня, их функциональное значение.
- Морфологическое расчленение побега, метамерия. Побеги удлиненные и укороченные. Почки, их строение и расположение. Различные подходы к классификации почек. Ветвление и нарастание побега, их типы и биологическое значение.
- Строение апекса побега. Различия в строении апекса побега у споровых и семенных. Сравнение апекса побега с апексом корня. Апикальный и интеркалярный рост.
- Молодой стебель, многообразие вариантов анатомического строения. Анатомо-топографические зоны стебля. Эндодерма (крахмалоносное влагалище) и перицикл. Принципы классификации стел и основные факторы их эволюции. Разнообразие устройства стелы у современных высших растений.
- Роль листьев и аксиллярных почек в формировании структурных особенностей центрального цилиндра (стелы). Листовые и веточные следы, листовые прорывы и прорывы ветвления. Строения стеблей двудольных и однодольных покрытосеменных растений в зоне узлов.
- Вторичное утолщение в стебле древесных двудольных и голосеменных растений. Морфологическое строение многолетних стеблей древесных растений. Работа камбия и феллогена и обусловленные ею изменения в анатомическом строении древесного растения. Особенности вторичного утолщения стебля древесных однодольных.
- Структурные элементы вторичной флоэмы (луба) и вторичной ксилемы (древесины). Понятия твёрдого, мягкого луба, годичных колец в древесине, первичных и вторичных лубо-древесинных (сердцевинных) лучей. Рассеяннососудистая и кольцесосудистая древесина.
- Строение и функциональная нагрузка первичных и вторичных лубо-древесинных (сердцевинных) лучей у хвойных и покрытосеменных растений. Понятие гетероцеллюлярных и гомоцеллюлярных лучей. Тяжевая древесинная паренхима, её типы.
- Возрастные изменения вторичных проводящих тканей: ядровая древесина, тиллообразование. Дилатация коры, её значение. Ритидом.
- Метаморфозы побега, их функциональное значение.
- Современные взгляды на происхождение листьев. Листья энационные и кладодификационные, их особенности и встречаемость у представителей разных таксонов высших растений. Макрофильная линия эволюции высших растений: роль листьев и аксиллярных почек в формировании структурных особенностей стелы.
- Лист, определение, основные функции. Структурные компоненты листа, порядок их развития. Особенности расчленения пластинки листа. Простые и сложные листья. Филлотаксис и его закономерности, листовая мозаика. Ярусные категории листьев. Гетерофиллия.
- Развитие листа как результат работы различных меристем. Анатомическое строение листовой пластинки, его разнообразие у семенных растений: листья голосеменных, листья цветковых. Бифациальные, унифациальные и эквифациальные листья. Жилкование листовой пластинки, его типы. Механические ткани в листе. Особенности строения проводящих пучков и их окончаний.
- Зависимость анатомического строения пластинки листа от типа фотосинтеза, анатомические особенности листьев растений с С-4 типом фотосинтеза. Особенности клеток листьев растений с CAM-метаболизмом.
- Понятие о продолжительности жизни листьев и их сменяемости. Листопад, его механизм и биологическое значение. Метаморфозы листьев.
 Понятие апопласта и симпласта. Экзодерма и эндодерма, их роль в “загрузке” проводящих элементов ксилемы. Строение и функции клеток эндодермы зоне поглощения (корневых волосков). Возрастные изменения в эндодерме и первичной коре у однодольных и двудольных покрытосеменных растений.
Понятие апопласта и симпласта. Экзодерма и эндодерма, их роль в “загрузке” проводящих элементов ксилемы. Строение и функции клеток эндодермы зоне поглощения (корневых волосков). Возрастные изменения в эндодерме и первичной коре у однодольных и двудольных покрытосеменных растений.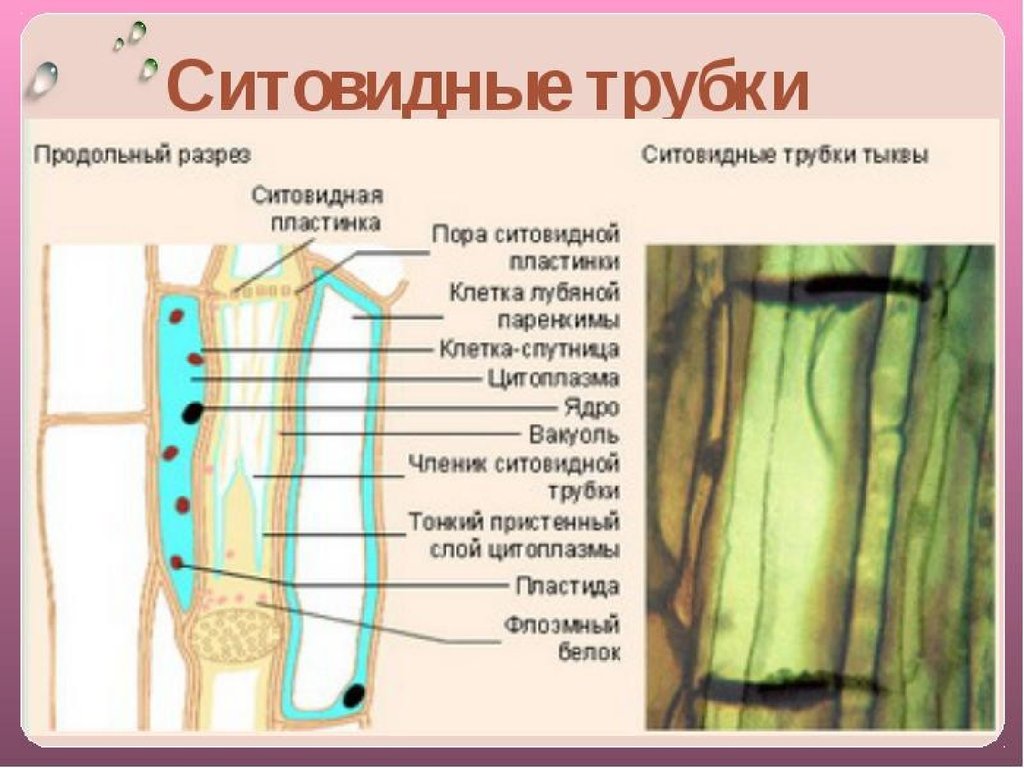 Сравнение апекса побега с апексом корня. Апикальный и интеркалярный рост.
Сравнение апекса побега с апексом корня. Апикальный и интеркалярный рост.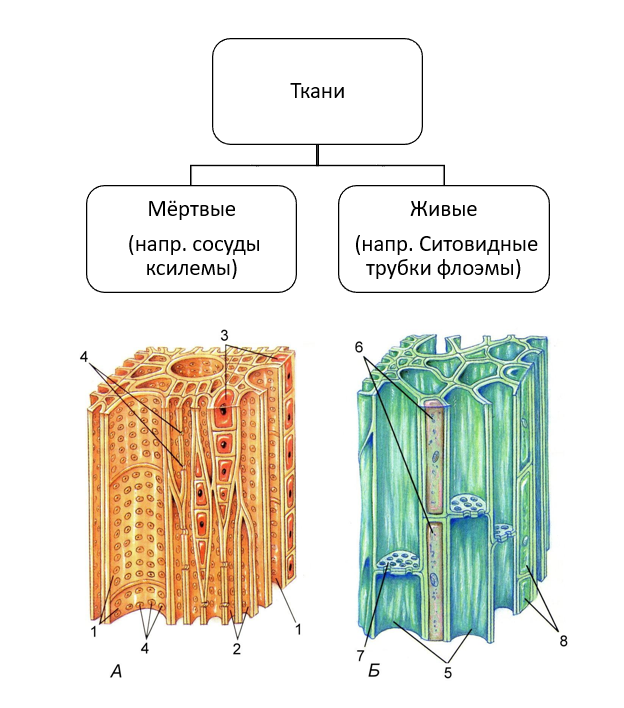 Понятия твёрдого, мягкого луба, годичных колец в древесине, первичных и вторичных лубо-древесинных (сердцевинных) лучей. Рассеяннососудистая и кольцесосудистая древесина.
Понятия твёрдого, мягкого луба, годичных колец в древесине, первичных и вторичных лубо-древесинных (сердцевинных) лучей. Рассеяннососудистая и кольцесосудистая древесина.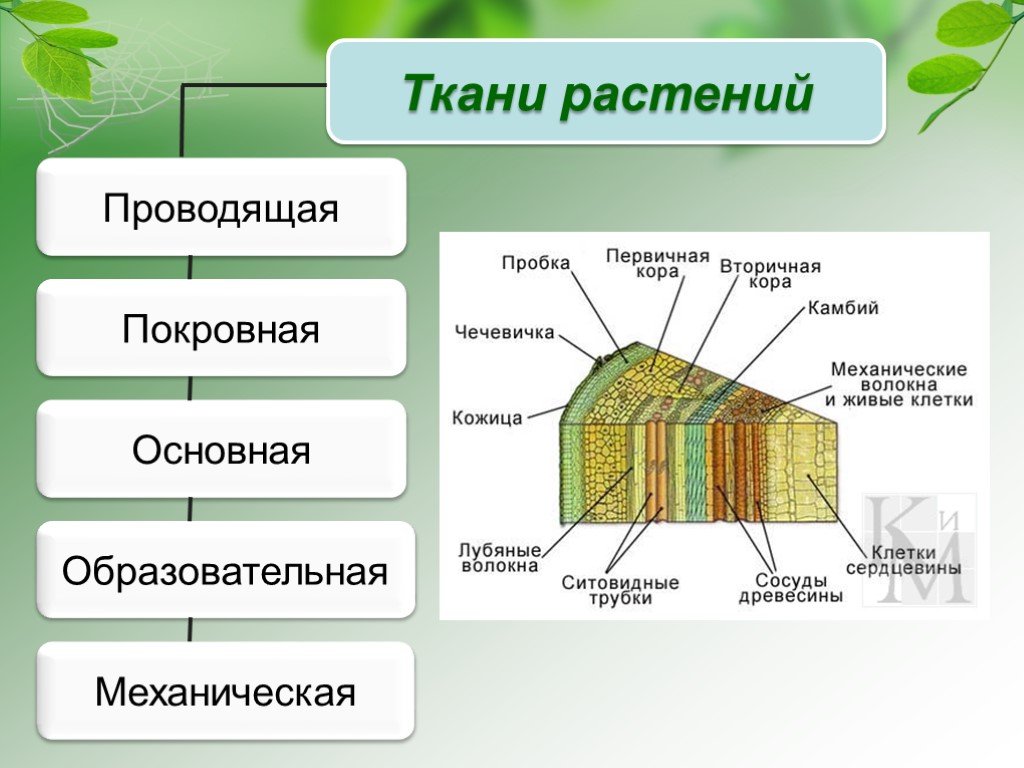 Особенности расчленения пластинки листа. Простые и сложные листья. Филлотаксис и его закономерности, листовая мозаика. Ярусные категории листьев. Гетерофиллия.
Особенности расчленения пластинки листа. Простые и сложные листья. Филлотаксис и его закономерности, листовая мозаика. Ярусные категории листьев. Гетерофиллия.
Модуль 3. Размножение растений
- Цикл развития высших растений, чередование гаметофита и спорофита. Бесполое размножение и половое воспроизведение высших растений. Типы спорангиев. Типы гаметангиев. Зоидо- и сифоногамия. Пространственное разделение гаметангиев у плауновидных и папоротниковидных. Редукционные явления в строении гаметофитов и гаметангиев в процессе эволюции таксонов высших растений.
- Общие представления об особенностях полового процесса у семенных растений, о строении и происхождении семязачатка. Семенные папоротники как уровень организации первых семенных растений. Особенности строения женских и мужских фруктификаций у вымершего порядка тригонокарповые (медуллозовые). Взгляды на происхождение интегумента.
- Тенденции к возникновению покрытосемянности у ископаемых растений на примере арбериевых (глоссопртерисовых), кейтониевых и беннеттитовых. Основные положения эуантовой (стробильной) теории происхождения цветка. Представления её авторов о типе цветка первых покрытосеменных. Значение теории для современного понимания проблем морфогенеза цветка. Многообразие взглядов на происхождение покрытосеменных растений.
- Оболочкосеменные. Особенности строения мега- и микростробилов оболочкосеменных и основные положения псевдантовой теории происхождения цветка. Представления её автора – P. Веттштейна – о типе цветка первых покрытосеменных. Значение теории для современного понимания проблем морфогенеза цветка. Многообразие взглядов на происхождение покрытосеменных растений.
- Основные характеристики покрытосеменных как таксона, отличия от голосеменных. Многообразие взглядов на происхождение покрытосеменных растений. Время и место происхождения покрытосеменных: проблемы понимания. Внешний облик древнейших ископаемых покрытосеменных на примере Archaefructus (Archaefructaceae) и Archaeanthus (Liriodendraceae).
- Цветок – общее представление о природе цветка, разнообразие цветков. Различные взгляды исследователей на происхождение цветка, его особенности и присутствие у первых покрытосеменных растений. Строение цветка – околоцветник, андроцей, гинецей, расположение на цветоложе. Закон чередования кругов. Филлотаксис частей цветка. Симметрия цветка. Диаграммы и формулы цветков.
- Околоцветник, строение, функции. Ахламидные и апохламидные цветки. Простой и двойной околоцветник, происхождение двойного околоцветника. Чашечка, её особенности у представителей разных таксонов покрытосеменных, роль чашечки при плоде. Особенности лепестков: отгиб и ноготок, шпорец, привенчик. Конгенитальное и постгенитальное срастание органов цветка и их частей. Нектарники в цветке.
- Андроцей, разнообразие вариантов строения. Морфологическая природа и строение тычинки, тычиночная нить, пыльники (микроспорангии), ориентация микроспорангиев относительно центра цветка. Число кругов тычинок, нарушение закона чередования кругов. Процессы срастания в андроцее. Типы вскрывания пыльников. Стаминодии.
- Андроцей, разнообразие вариантов строения. Строение микроспорофилла и стенки микроспорангия. Микроспорогенез. Развитие и строение микрогаметофита. Апертуры, их названия, расположение. Особенности оболочек микрогаметофита. Поллинии.
- Гинецей, разнообразие вариантов строения. Морфологическая природа плодолистика. Понятие плацентации. Кондупликатный плодолистик и его эволюция. Асцидиатный плодолистик. Зональность плодолистика: сочетание в одном гинецее пликатных (синпликатных) и асцидиатных (синасцидиатных) зон. Изменения в расположении воспринимающей пыльцу (рыльцевой) ткани в процессе специализации плодолистика. Понятие компитума.
- Гинецей, срастание плодолистиков между собой в системе понятий кондупликатного плодолистика. Типы гинецея и плацентации. Биологическое значение завязи, типы завязи в цветке, понятие гипогинии и эпигинии. Гипантий. Гиностемий и гиностегий. Гинофор и андрогинофор.
- Гинецей, строение семязачатка, типы семязачатков. Происходящие внутри семязачатка процессы. Разнообразие вариантов формирования и строения зрелых мегагаметофитов покрытосеменных. Рост пыльцевой трубки через ткани семязачатка (халазо- и порогамия). Процесс двойного оплодотворения, развитие зародыша и эндосперма. Вариативность процесса. Первые этапы развития зародыша: суспензор и проэмбрио. Апомиксис.
- Цветение и опыление. Авто- и аллогамия. Клейстогамные и хазмогамные цветки. Способы предотвращения самоопыления. Абиотическое и биотическое опыление, разнообразие вариантов. Адаптации, связанные с различными типами опыления. Связь с агентами опыления, примеры.
- Соцветия, определение и биологическое значение. Разнообразие соцветий, принципы их классификации. Основные морфологические типы соцветий. Антодии.
- Плод как результат преобразования цветка. Разнообразие плодов и сложности их классификации. Важность анатомии для реконструкции морфогенеза плодов. Перикарпий, консистенция перикарпия. Типы вскрывания или распадения плодов. Дробные и членистые плоды. Ариллусы и ариллоиды.
- Плод, определение. Плод как результат преобразования цветка. Критерии классификации плодов. Основные типы плодов из апокарпного гинецея.
- Плод, определение. Плод как результат преобразования цветка. Критерии классификации плодов. Основные типы плодов из синкарпного гинецея.
- Плод, определение. Плод как результат преобразования цветка. Критерии классификации плодов. Основные типы плодов из пара-, фрагмо- и лизикарпногоо гинецеев.
- Плод, типы распространения плодов. Понятие диаспоры. Биологическое значение партенокарпии и гетерокарпии. Варианты гетерокарпии. Соплодия, примеры соплодий.
- Семя, определение. Биологическое значение возникновения семян. Основные отличия в строении семязачатков и семян голосеменных и покрытосеменных растений. Разные типы семян покрытосеменных по особенностям питательных тканей. Типы зародышей.
 Бесполое размножение и половое воспроизведение высших растений. Типы спорангиев. Типы гаметангиев. Зоидо- и сифоногамия. Пространственное разделение гаметангиев у плауновидных и папоротниковидных. Редукционные явления в строении гаметофитов и гаметангиев в процессе эволюции таксонов высших растений.
Бесполое размножение и половое воспроизведение высших растений. Типы спорангиев. Типы гаметангиев. Зоидо- и сифоногамия. Пространственное разделение гаметангиев у плауновидных и папоротниковидных. Редукционные явления в строении гаметофитов и гаметангиев в процессе эволюции таксонов высших растений. Многообразие взглядов на происхождение покрытосеменных растений.
Многообразие взглядов на происхождение покрытосеменных растений.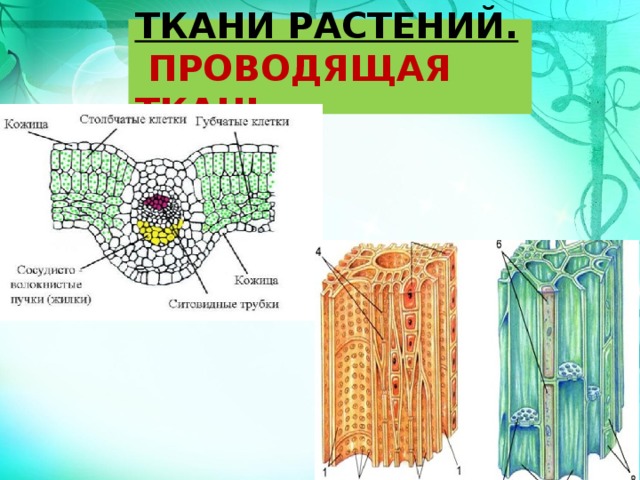 Строение цветка – околоцветник, андроцей, гинецей, расположение на цветоложе. Закон чередования кругов. Филлотаксис частей цветка. Симметрия цветка. Диаграммы и формулы цветков.
Строение цветка – околоцветник, андроцей, гинецей, расположение на цветоложе. Закон чередования кругов. Филлотаксис частей цветка. Симметрия цветка. Диаграммы и формулы цветков. Микроспорогенез. Развитие и строение микрогаметофита. Апертуры, их названия, расположение. Особенности оболочек микрогаметофита. Поллинии.
Микроспорогенез. Развитие и строение микрогаметофита. Апертуры, их названия, расположение. Особенности оболочек микрогаметофита. Поллинии. Рост пыльцевой трубки через ткани семязачатка (халазо- и порогамия). Процесс двойного оплодотворения, развитие зародыша и эндосперма. Вариативность процесса. Первые этапы развития зародыша: суспензор и проэмбрио. Апомиксис.
Рост пыльцевой трубки через ткани семязачатка (халазо- и порогамия). Процесс двойного оплодотворения, развитие зародыша и эндосперма. Вариативность процесса. Первые этапы развития зародыша: суспензор и проэмбрио. Апомиксис.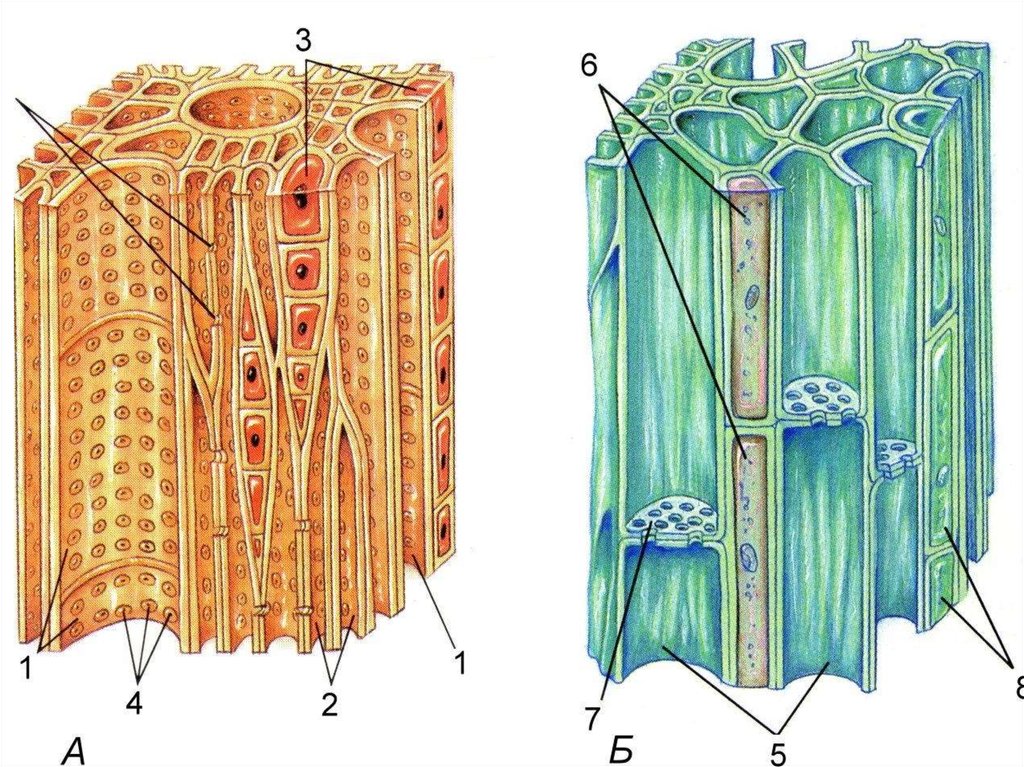 Критерии классификации плодов. Основные типы плодов из апокарпного гинецея.
Критерии классификации плодов. Основные типы плодов из апокарпного гинецея.
Основная литература:
- Ботаника в четырёх томах. Том 3. Высшие растения: учебник для студ. высш. учеб. заведений / А. К. Тимонин. –M.: Издательский центр “Академия”, 2007. –
- 352 .
- Ботаника в четырёх томах. Том 4. Книга 2. Систематика высших растений: учебник для студ. высш. учеб. заведений / А.К. Тимонин, Д.Д.Соколов, А.Б.Шипунов –M.: Издательский центр “Академия”, 2009. –
- 352 .
- Морфология и анатомия высших растений / Лотова Л.И. – M.: Эдиториал УРСС, 2000. – 528 с.
 К. Тимонин. –M.: Издательский центр “Академия”, 2007. –
К. Тимонин. –M.: Издательский центр “Академия”, 2007. –
Дополнительная литература:
- Ботаника. Учебник для вузов в 4 т. / П.Зитте, Э.В. Вайлер, Й.В. Кадерайт, А.Брезински, К.Кёрнер; на основе учебника Э.Страсбургера [и др.]; пер. с нем. –М.: Издательский центр “Академия”, 2007. – 368 с.
- Т. 1 Клеточная биология. Анатомия. Морфология.
Ботаника, высшие растения
Почему ксилема и флоэма называются сложными тканями?
Анатомия цветковых растений включает организацию клеток и тканей в телах цветковых растений. Ткани образованы группой клеток для выполнения определенной функции. Ткани растений подразделяются на два типа в зависимости от их способности делиться. Они являются меризматичными и постоянными тканями.
Ткани образованы группой клеток для выполнения определенной функции. Ткани растений подразделяются на два типа в зависимости от их способности делиться. Они являются меризматичными и постоянными тканями.
Меризматическая ткань
Это простая ткань, обладающая способностью к непрерывному делению клеток. В этом типе ткани рост происходит в корнях и кончиках побегов растения. Основываясь на области меристематической ткани в теле растения, мы можем разделить меристемы на три типа. Их:
- Апикальные меристемы. Эти меристемы расположены на кончиках корней и побегов.
- Вставочные меристемы – Вставочные меристемы расположены в междоузлиях или в основании листьев.
- Боковые меристемы – имеются на боковой стороне стебля и основания растения.
Постоянная ткань
Эти ткани происходят из меризматических тканей и утратили способность делиться. Этот тип ткани можно найти у взрослых растений. Они подразделяются на два типа: простые и сложные ткани 9. 0003
0003
Простая ткань: Это также называется гомогенной тканью, состоящей из клеток одного типа. Простые ткани бывают трех типов: паренхима, колленхима и склеренхима.
- Паренхима : Эти ткани находятся в нежных частях растений, таких как корни, стебли, листья и цветы. Клетки этой ткани содержат между собой огромные межклеточные пространства. Каждая клетка имеет вакуоль посередине. Функции тканей паренхимы – фотосинтез, запасание и помощь растению в дрейфе по воде.
- Колленхима : Колленхима похожа на клетки паренхимы, которые содержат овальные, круглые или многоугольные клетки с толстыми стенками. Толщина стенок обусловлена пектином, целлюлозой и гемицеллюлозой. Они предназначены для механической поддержки структуры растения в таких частях, как черешок листа.
- Склеренхима — Клетки этой ткани мертвы. Этот тип клеток лишен ядра и цитоплазмы. Их клеточные стенки утолщены лигнином и целлюлозой. Их основная способность заключается в придании механической прочности частям зрелого растения.
 Их основная способность заключается в придании механической прочности частям зрелого растения.
Их основная способность заключается в придании механической прочности частям зрелого растения.Сложная ткань
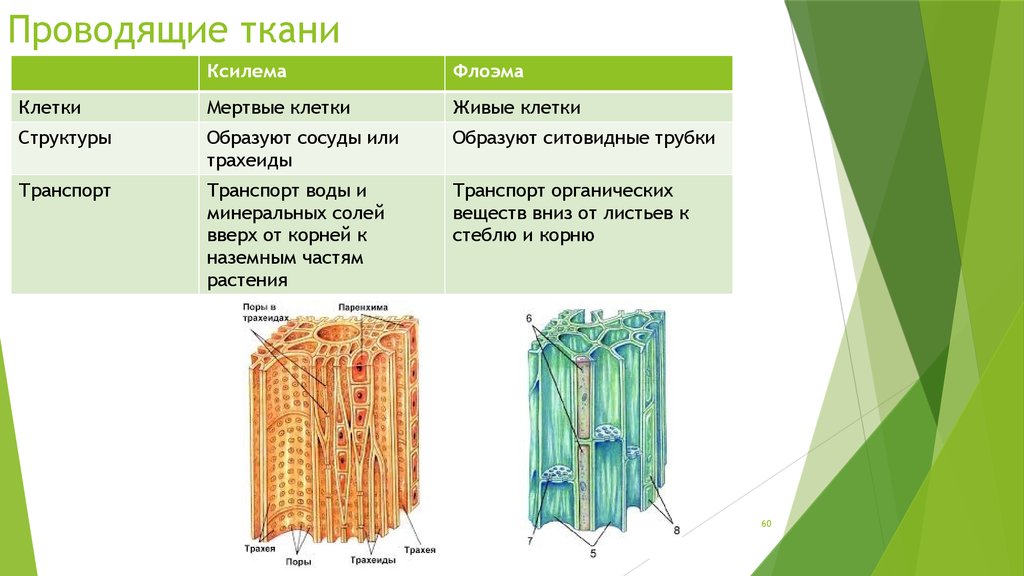
Этот тип ткани также известен как гетерогенная ткань, которая состоит из различных типов клеток, и эти дифференцированные типы клеток координируются для выполнения определенной функции. Сложные ткани бывают двух типов: ксилема и флоэма.
Ксилема (водопроводящая ткань)
Ксилема образована трансцендентно трахеальными компонентами, такими как трахеиды и сосуды. Эссенциальная ксилема начинается с прокамбия во время основного развития, в то время как факультативная ксилема начинается в сосудистом камбии во время вспомогательного развития. Он находится в глубоком слое растений, который состоит из трахеи, волокон ксилемы, трахеид и паренхимы ксилемы. Трахеиды и сосуды имеют цилиндрическую форму. Это позволяет им транспортировать воду и минералы в восходящем направлении (вертикально). Основной функцией ксилемы является перемещение воды от корней к стеблям и листьям, но она также содержит добавки.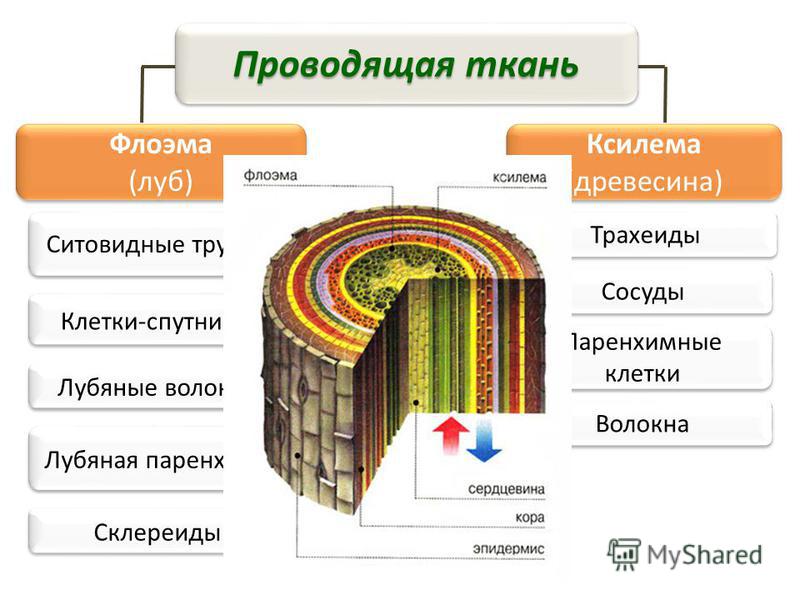 Он также обеспечивает механическую прочность растений. Ткани ксилемы представляют собой трубчатые структуры без поперечных стенок. Эта ткань напоминает форму звезды. Волокна ксилемы мельче и располагаются в центре сосудистых пучков. Волокна ксилемы присутствуют в корнях, стеблях и листьях. Он отвечает за возмещение общего количества потерянных молекул воды в результате транспирации и фотосинтеза.
Он также обеспечивает механическую прочность растений. Ткани ксилемы представляют собой трубчатые структуры без поперечных стенок. Эта ткань напоминает форму звезды. Волокна ксилемы мельче и располагаются в центре сосудистых пучков. Волокна ксилемы присутствуют в корнях, стеблях и листьях. Он отвечает за возмещение общего количества потерянных молекул воды в результате транспирации и фотосинтеза.
Флоэма (Пищепроводящая ткань)
Флоэма начинается с меристематических клеток в сосудистом камбии – основная флоэма из апикальной меристемы и необязательная флоэма из сосудистого камбия. Он находится во внешних слоях растения, состоящих из клеток-компаньонов, паренхимы флоэмы, волокон флоэмы, ситовидных клеток и ситовидных трубок. В отличие от ксилемы флоэма движется в обоих направлениях. Он отвечает за транспортировку пищи из листьев в другие части растения. Он не обеспечивает механической прочности растений. Волокна флоэмы крупнее и располагаются с внешней стороны сосудистых пучков. Они присутствуют в стеблях и листьях, которые позже транспортируются и растут в корнях, плодах и семенах. Он отвечает за транспортировку белков и мРНК по всему растению.
Они присутствуют в стеблях и листьях, которые позже транспортируются и растут в корнях, плодах и семенах. Он отвечает за транспортировку белков и мРНК по всему растению.
Характеристика ксилемы и флоэмы
Флоэма и ксилема представляют собой сложные ткани, осуществляющие транспортировку пищи и воды к частям растений. Это сосудистые ткани растения.
Характеристики | Xylem | Phloem |
| . внутри сосудов ксилемы. Существует два типа: протоксилема и метаксилема, в зависимости от структуры лигнина. | Удлиненной трубчатой формы с тонкостенными ситовыми трубками. Ситовидные трубки имеют поры на каждом конце в поперечных стенках и микротрубочки, которые проходят между ситовыми элементами, обеспечивая продольный поток материала. | |
| Элементы | Ксилема включает сосуды ксилемы, волокна и трахеиды. | Ксилема включает волокна ксилемы флоэмы, ситовидные трубки, ситовидные клетки, паренхиму флоэмы и сопутствующие клетки. |
| Форма | Ксилема имеет форму звезды. | Флоэма не звездчатая. |
| Расположение | Расположен в центре сосудистого пучка, глубоко в растении. | Располагается с наружной стороны сосудистого пучка. |
| Движение | Движение однонаправленное. | Двунаправленное движение. |
| Функции | Обеспечивает механическую прочность и отвечает за транспортировку воды и других питательных веществ к растениям. | Не обеспечивает механической прочности, а флоэма представляет собой живую ткань, отвечающую за транспортировку пищи и других органических материалов. |
| Природа ткани | Ксилема состоит из мертвых клеток (паренхима – это единственные живые клетки, присутствующие в ксилеме). Ксилема составляет основную часть тела растения.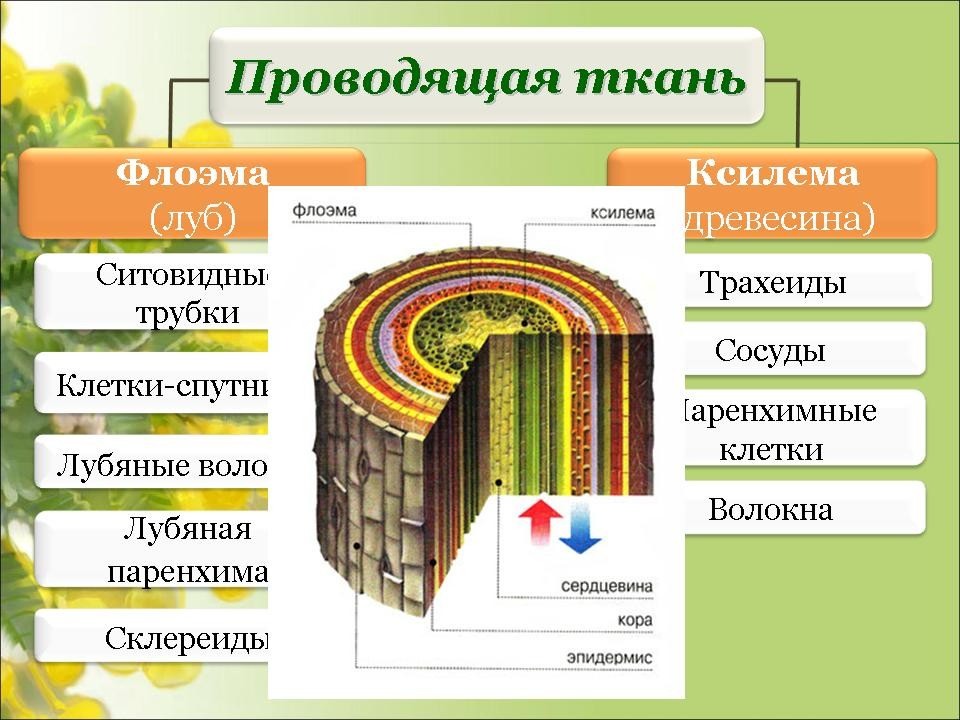 | Флоэма содержит живые клетки (волокна — единственные мертвые клетки во флоэме). Он составляет небольшую часть тела растения. |
Часто задаваемые вопросы
Вопрос 1: Почему ксилема известна как водопроводящая ткань?
Ответ:
Ксилему называют водопроводящей тканью, так как она переносит молекулы воды от корней к частям растений.
Вопрос 2: Почему флоэма известна как проводящая пищу ткань?
Ответ:
Флоэма называется пищевой тканью, так как она переносит пищевые вещества и питательные вещества из листьев в другие части растения.
Вопрос 3: Какие существуют сложные ткани и типы сложных тканей?
Ответ:
Сложная ткань представляет собой гетерогенную ткань, состоящую из более чем одного типа клеток, и эти различные типы клеток объединены для выполнения определенной функции.
 Поэтому их называют сосудистыми тканями. Они переносят воду и пищу к частям растений. Сложная ткань классифицируется как ксилема и флоэма.
Поэтому их называют сосудистыми тканями. Они переносят воду и пищу к частям растений. Сложная ткань классифицируется как ксилема и флоэма.Вопрос 4: Каковы характеристики ксилемы и флоэмы?
Ответ:
- Ксилема: Основной функцией ксилемы является перемещение воды от корней к стеблям и листьям, однако она дополнительно доставляет питательные вещества. Он также обеспечивает механическую прочность растений. Ткани ксилемы представляют собой трубчатые структуры без поперечных стенок. Эта ткань напоминает форму звезды. Волокна ксилемы мельче и располагаются в центре сосудистых пучков. Волокна ксилемы присутствуют в корнях, стеблях и листьях. Он отвечает за возмещение общего количества потерянных молекул воды в результате транспирации и фотосинтеза.
- Флоэма: Отвечает за транспортировку пищи из листьев в другие части растения. Он не обеспечивает механической прочности растений.
 Волокна флоэмы крупнее и располагаются на внешней стороне сосудистых пучков. Они присутствуют в стеблях и листьях, которые позже транспортируются и растут в корнях, плодах и семенах. Он отвечает за транспортировку белков и мРНК по всему растению.
Волокна флоэмы крупнее и располагаются на внешней стороне сосудистых пучков. Они присутствуют в стеблях и листьях, которые позже транспортируются и растут в корнях, плодах и семенах. Он отвечает за транспортировку белков и мРНК по всему растению.Вопрос 5: Каковы функции ксилемы и флоэмы?
Ответ:
Ксилема и флоэма участвуют в транспортировке воды, минералов и пищи по всему растению. Ксилема переносит воду и минеральные вещества от корней к листьям. Тем не менее, флоэма передает пищу, организованную проходами, к различным частям растения.
Документ без названия
Документ без названия
Botany Primer
Добро пожаловать в
учебник по ботанике! Этот краткий обзор вторичного роста стеблей предназначен для
предоставить только основы этого увлекательного процесса завода. Мы рекомендуем проконсультироваться
базовая книга по ботанике, такая как « Биология растений » П. Х. Рэйвен и др. (1999) для
Х. Рэйвен и др. (1999) для
Дополнительная информация. В этом учебнике мы познакомим вас с основами ботаники.
термины, обсудить вторичный рост, наметить изменения в теле растения во время вторичного
рост, и посмотрите на структуру древесины. Создавая этот обзор, мы держали вас,
дендрохронолог, в виду.
Введение
Основной
Тело растения состоит из корней, листьев и стеблей. Как дендрохронологи,
нас больше всего интересуют стебли (хотя некоторые очень интересные дендрохронологические
работа выполнена с корнями). Стержни выполняют несколько функций, таких как
в качестве поддержки и проводки. Когда мы смотрим на годичные кольца деревьев, мы смотрим на некоторые
проводящих и поддерживающих структур, образующихся в стебле.
Для перемещения материалов
по всему телу растения (сосудистые растения) используют особую проводящую
ткани или сосудистые ткани, известные как ксилема и флоэма . Ксилем транспортирует
вода и растворенные вещества от корней вверх. Флоэма транспортирует материалы
Флоэма транспортирует материалы
вниз с листьев.
Рост
Первичный рост
приводит к увеличению длины основного тела растения. Первичный рост
происходит в апикальной меристеме (меристема, расположенная на кончике стебля).
Меристема представляет собой область недифференцированных клеток, из которой образуются новые клетки.
возникают в результате клеточного деления. Вторичный рост – увеличение толщины
или обхват тела растения за счет активности боковых меристем. Боковой
меристемы расположены латерально вокруг стебля (не на концах, как апикальные
меристемы). Есть два типа латеральных меристем или камбий, сосудистых
камбий и пробковый камбий . Мы будем говорить только о росте
сосудистый камбий (хотя некоторые исследователи изучали временные вариации
в производстве пробки).
Сосудистый камбий
состоит из двух типов инициалов: лучевые инициалы и веретенообразные инициалы
( инициалы — это клетки, которые остаются в меристематических областях после клеточного
деление и продолжать делать больше клеток).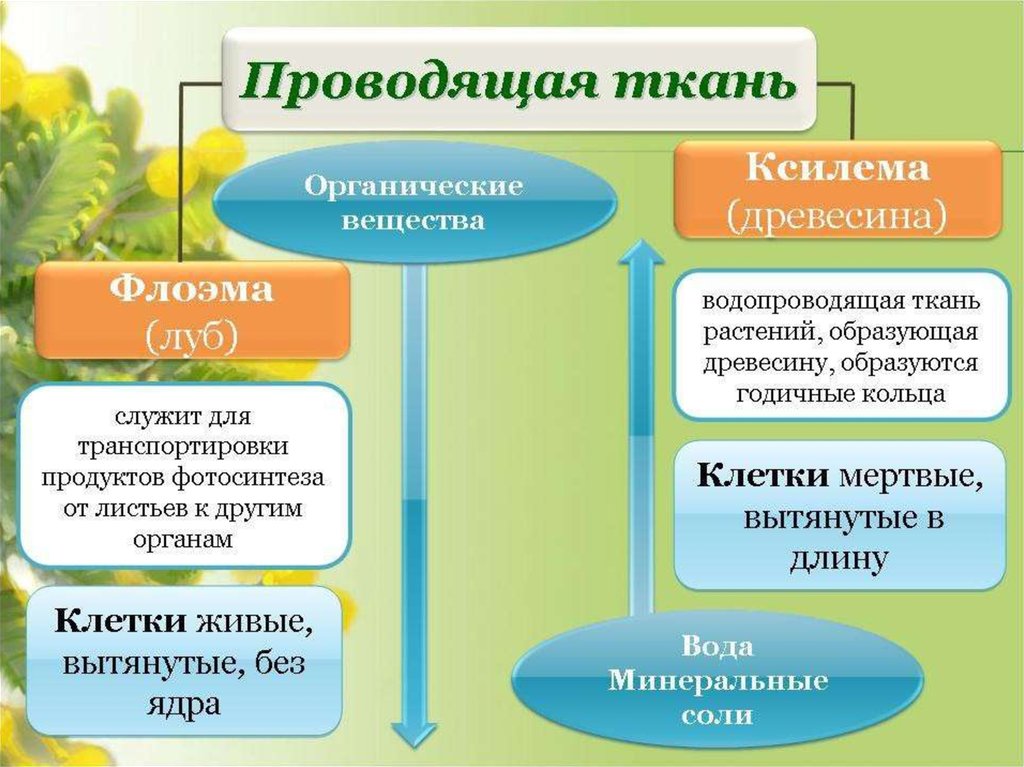 Инициалы лучей производят лучи. Лучи
Инициалы лучей производят лучи. Лучи
горизонтально ориентированы в стебле и выполняют несколько функций: транспортировку пищи
и вода радиально и запасающая (крахмал, белок, липид).
Как дендрохронологи,
нас больше интересуют веретенообразные инициалы. Веретенообразные инициалы производят
вторичная ксилема и вторичная флоэма (ксилема и флоэма образуются
во время вторичного роста). Когда мы изучаем древесину или годичные кольца, мы смотрим на
вторичная ксилема. По мере деления веретенообразных инициалей образуется вторичная ксилема.
внутрь клетки, а вторичная флоэма наружу. Следовательно,
сосудистый камбий (где мы находим веретенообразные инициали) считается
быть двусторонним.
Цифры в
клетки указывают порядок, в котором они были образованы. По мере роста,
клетки будут оттеснены от сосудистого камбия.
Изменения в
корпус растения
Во время среднего
роста в организме растения происходят большие изменения.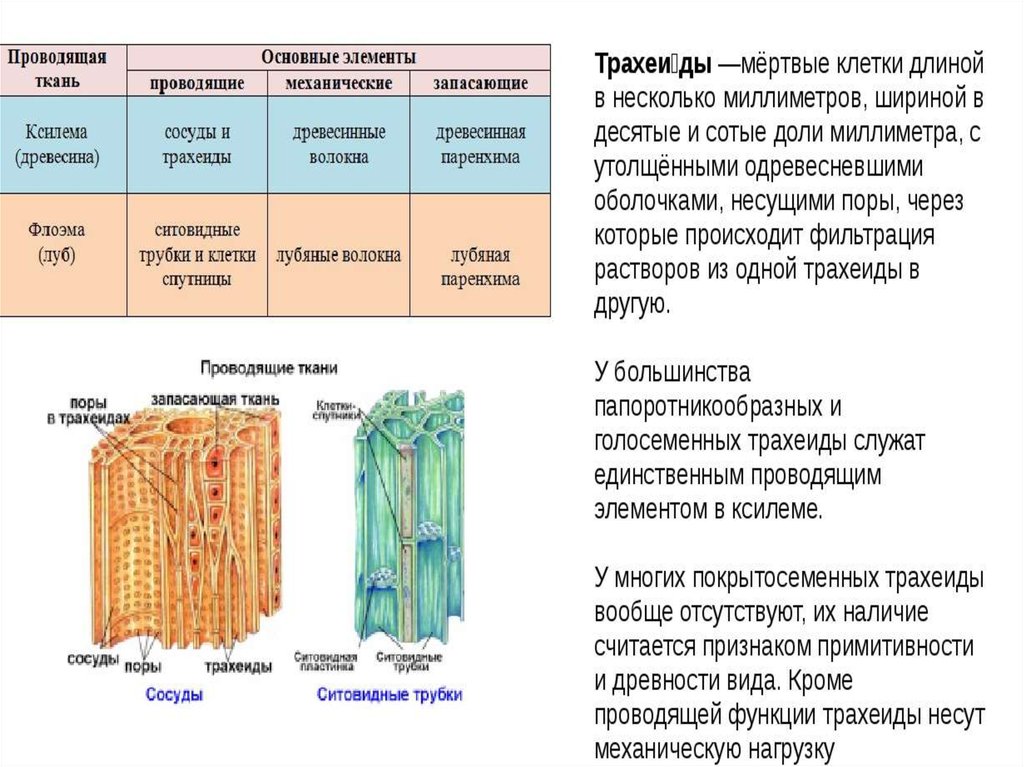 Ниже приведена схематическая интерпретация
Ниже приведена схематическая интерпретация
как меняется стебель в первый год роста древесного растения. Примечание,
если вы не уверены в значении какого-либо из приведенных ниже терминов, обязательно
найти их в книге ботаники! Вы должны понимать каждого из игроков
если вы хотите знать, как играется игра!
В регионах с умеренным климатом
мира, камбий образует вторичную ксилему и вторичную флоэму.
в течение вегетационного периода и будет оставаться в состоянии покоя при благоприятных условиях роста
закончились за год. Когда камбий «реактивируется»,
в следующем году вокруг стебля образуется еще один слой ксилемы и флоэмы.
с ксилемой внутрь и флоэмой наружу.
Этот ствол четыре
лет. Цифры обозначают год, в котором каждое из колец ксилемы
(Примечание: на этой диаграмме показана только ксилема).
Основные изменения
в кожной системе (системе тканей, служащих наружным покровом
стебель) также возникают во время вторичного роста.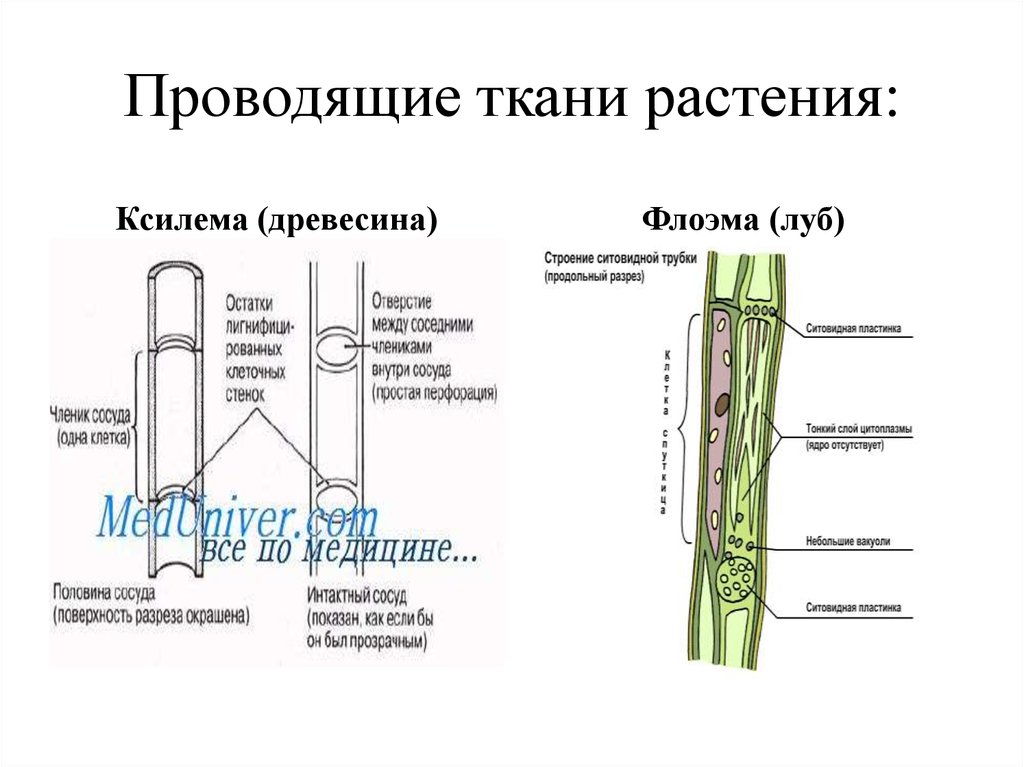 По мере расширения стебля во время вторичного
По мере расширения стебля во время вторичного
рост, вторичная флоэма и ткани снаружи сосудистого камбия
растягиваются и в конечном итоге отслаиваются от стебля. Мы не будем обсуждать эти
изменения, поскольку нас больше всего интересует производство древесины или вторичной ксилемы.
Проверьте базовую книгу по ботанике, чтобы узнать подробности, связанные с этим интересным
явление.
Кольца роста
Как дендрохронологи,
нас интересуют годичные кольца (также известные как кольца деревьев или годовые приросты).
Так как же определить индивидуальное кольцо роста? Годичные кольца в регионах с умеренным климатом
в мире обычно состоят из ранней древесины и поздней древесины .
Вы можете отличить раннюю древесину от поздней, ища различия.
по плотности: ранняя древесина менее плотная и состоит из более крупных ячеек с тонкими стенками
в то время как поздняя древесина более плотная и состоит из более мелких клеток с более толстыми стенками.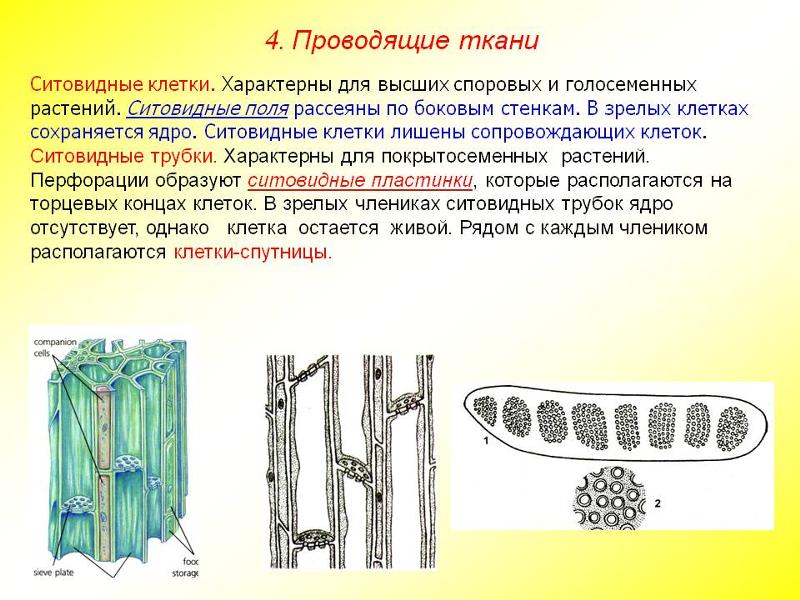
Принимая во внимание
узор ранней древесины и поздней древесины (иногда называемой весенней древесиной и летней
древесина; эта терминология не совсем корректна!) мы можем выделить отдельные
кольца роста. Годовое кольцо состоит из ранней и поздней древесины.
На ширину годичного кольца деревьев в данный год влияет множество факторов (прим.
изменение ширины в годичных кольцах деревьев показано ниже). Щелкните эту ссылку для
Дополнительная информация. Внимание: в данные годы (из-за различных стрессов или климатических
аномалии) могут возникать ложные кольца или отсутствующие кольца. Поэтому нам нужно перекреститься
годы, которые мы присваиваем отдельным кольцам.
Древесина и дерево
Кольца
Как видите
из приведенных выше фотографий, есть некоторые существенные различия в анатомии древесины и
структура среди различных пород деревьев. Дуб, показанный выше, представляет собой твердую древесину, и
сосна относится к хвойным породам. Лиственные породы и Мягкие породы происходят из покрытосеменных растений.
Лиственные породы и Мягкие породы происходят из покрытосеменных растений.
(цветковые растения) и хвойные растения соответственно. Будь осторожен! Есть
твердые хвойные и мягкие лиственные породы! Два вида древесины различаются по многим свойствам.
включая наличие или отсутствие элементов сосуда, долю специализированных
типы тканей, сложность и состав лучевой системы. Лиственные породы часто
подразделяются на разные типы в зависимости от анатомических особенностей. Для начала
дендрохронологи, мы рекомендуем использовать кольцевидно-пористую древесину, так как ранняя древесина
и поздняя древесина настолько разительно отличаются друг от друга, что делают индивидуальную идентификацию колец
Полегче.
За отличное
обзор структуры древесины, идентификация и многое другое, мы настоятельно рекомендуем R.B.
Книга Ходли Идентификация древесины: точные результаты с помощью простых инструментов (1990).
Вы найдете отличные фотографии и описания многих видов древесины.
Вы также можете
найти разнообразие свойств древесины в одном стволе. При вырубке деревьев вы можете
обратите внимание, что древесина ближе к сердцевине часто темнее, чем в регионах
ближе к коре. Эти разноцветные зоны — сердцевина
и заболонь . Сердцевина является непроводящей древесиной, в то время как заболонь проводит
материал через ствол. Превращение сердцевины в заболонь часто
включает инфильтрацию, осаждение и накопление химических соединений, которые
часто делают сердцевину темнее. Помните, что древесина — это вторичная ксилема, а древесина
мертвая ткань (единственная живая часть ствола дерева — камбий и прилегающие
клетки).
Juglans nigra
л.
Быстрая викторина!
Если верхняя часть
этот забор в настоящее время находится на высоте 2 м над землей, а дерево растет на 0,5 м каждые
год, какой высоты будет вершина забора через 3 года?
Ответ :
Верх забора будет на высоте 2 м от земли! Когда начинается вторичный рост
на участке стебля прекращается первичный рост! Простой способ визуализировать это
заключается в использовании «аналогии укладочного конуса».