Содержание
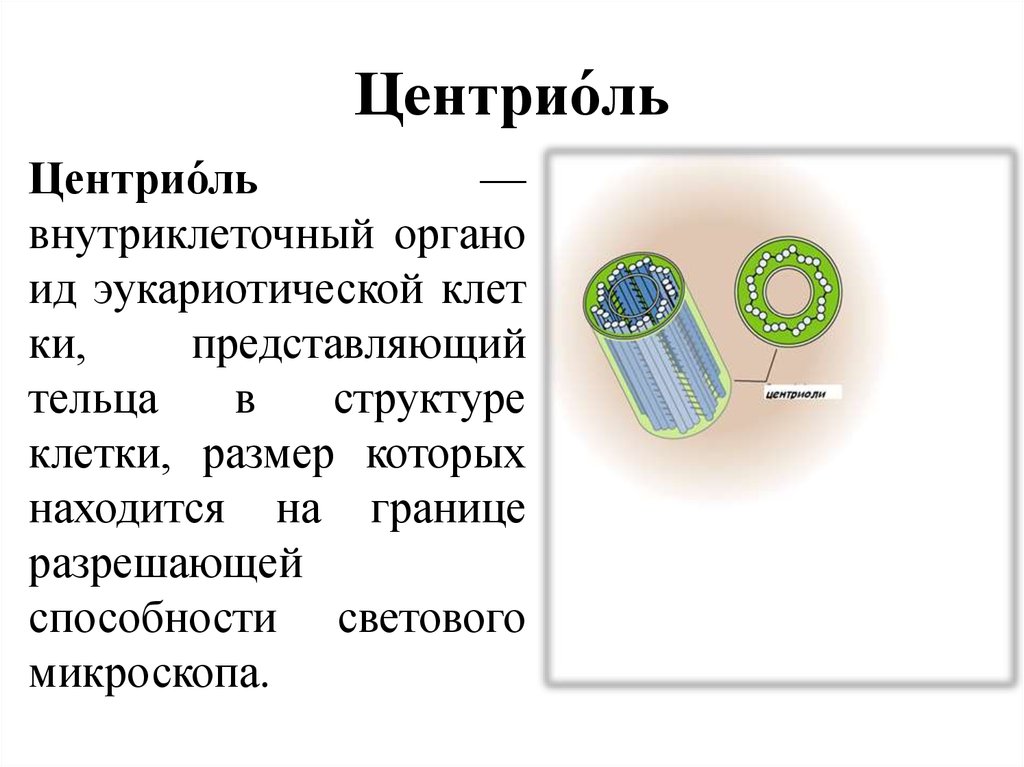
Описан механизм неканонической сборки центриолей у мхов — PCR News
Подготовила
Елизавета Минина
У животных при делении клеток центриоли удваиваются по хорошо изученному каноническому механизму. Однако у многих эукариот сборка центриолей происходит de novo. В статье, опубликованной в Current Biology, описан механизм неканонической сборки центриолей с нуля у мха Physcomitrium patens.
Подготовила
Елизавета Минина
Новое исследование показывает. что механизм сборки центриолей менее консервативен, чем считалось. Центриоли — структуры, состоящие из микротрубочек: обычно это девять триплетов, которые располагаются по кругу, образуя цилиндр. Пара центриолей, окруженная плотным перицентриолярным материалом, называется центросомой. Центросома участвует в организации митоза и поддерживают структуру микротрубочек в цитоплазме. Однако центриоли также могут выступать в роли базальных телец — закрепляться на клеточной мембране и становиться матрицей для роста ресничек, которые обеспечивают подвижность клеток или передачу сигналов. Это происходит не только у жгутиковых и инфузорий, но и в клетках млекопитающих. Центриоли сперматозоидов участвуют в формировании жгутиков, а также необходимы для развития эмбриона после оплодотворения. Неспособность центриолей преобразовываться в базальные тельца может быть причиной генетических заболеваний и пороков развития.
Однако центриоли также могут выступать в роли базальных телец — закрепляться на клеточной мембране и становиться матрицей для роста ресничек, которые обеспечивают подвижность клеток или передачу сигналов. Это происходит не только у жгутиковых и инфузорий, но и в клетках млекопитающих. Центриоли сперматозоидов участвуют в формировании жгутиков, а также необходимы для развития эмбриона после оплодотворения. Неспособность центриолей преобразовываться в базальные тельца может быть причиной генетических заболеваний и пороков развития.
Деление клеток эукариот сопровождается удвоением их центриолей, которое может происходить по разным механизмам. Чаще всего сборка дочерней центриоли начинается вблизи материнской центриоли перпендикулярно ей. Однако так происходит далеко не у всех эукариот. У цветковых и голосеменных растений центриолей нет, а у мхов, папоротников, гингко и саговников единственные клетки, обладающие центриолями — это спермии, мужские половые клетки, которые формируются в специальных структурах — антеридиях. В новой работе детально описан процесс неканонического формирования центриолей на примере мха Physcomitrium patens. Авторы исследования изучили сперматиды P. patens с помощью электронной и световой микроскопии, а также молекулярных методов, чтобы понять, как центриоли возникают de novo.
В новой работе детально описан процесс неканонического формирования центриолей на примере мха Physcomitrium patens. Авторы исследования изучили сперматиды P. patens с помощью электронной и световой микроскопии, а также молекулярных методов, чтобы понять, как центриоли возникают de novo.
В начале сперматогенеза, как и ожидалось, центриоли найти не удалось, однако вблизи ядра сперматид присутствовало электронно-плотное облако, из которого выходили микротрубочки. Эту электронно-плотную структуру авторы назвали «концентратором». Затем вблизи концентратора появлялись две центриоли (бицентриоли), расположенные друг за другом, соединенные перемычкой и окруженные единым облаком микротрубочек. Далее центриоли обособлялись друг от друга, и одновременно с этим под ними начинала формироваться особая многослойная структура. В зрелом спермии многослойная структура и лежащие над ней центриоли располагаются вблизи поверхности клетки, причем каждая центриоль выступает в роли базального тела, от которого отходит ресничка стандартного состава — 9 дублетов тубулиновых микротрубочек и две микротрубочки в центре (9+2).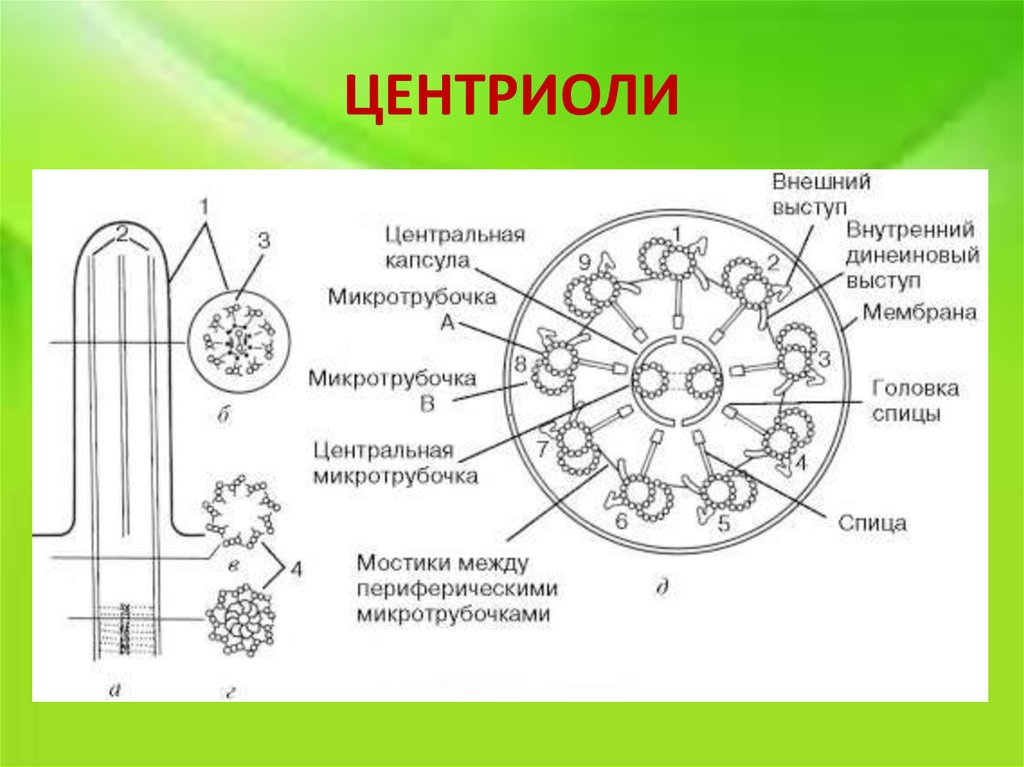 Перемычка, соединявшая две центриоли в составе бицентриоли, на стадии зрелого спермия отсутствует.
Перемычка, соединявшая две центриоли в составе бицентриоли, на стадии зрелого спермия отсутствует.
Детальное изучение структуры новособранных центриолей показало, что на стадии бицентриолей они «закручены» в разные стороны: в одной центриоли ряды микротрубочек ориентированы по часовой стрелке, а во второй — против. Тот факт, что от бицентриолей берут начало микротрубочки, подтверждает предположение о роли бицентриоли как центра организации этой составляющей цитоскелета. После разделения центриолей и появления многослойной структуры центриоли становятся практически параллельны друг другу.
Несмотря на кажущуюся одновременность развития, на молекулярном уровне сестринские центриоли развиваются асинхронно. В частности, об этом свидетельствует различная локализация двух эволюционно консервативных центриолярных белков, SAS6 и POC1, в созревающих центриолях. Авторы работы показали, что SAS6 необходим для сборки перемычки, а POC1, наряду с консервативным белком центриолей Bld10, — для элонгации перемычки и формирования стенок центриолей.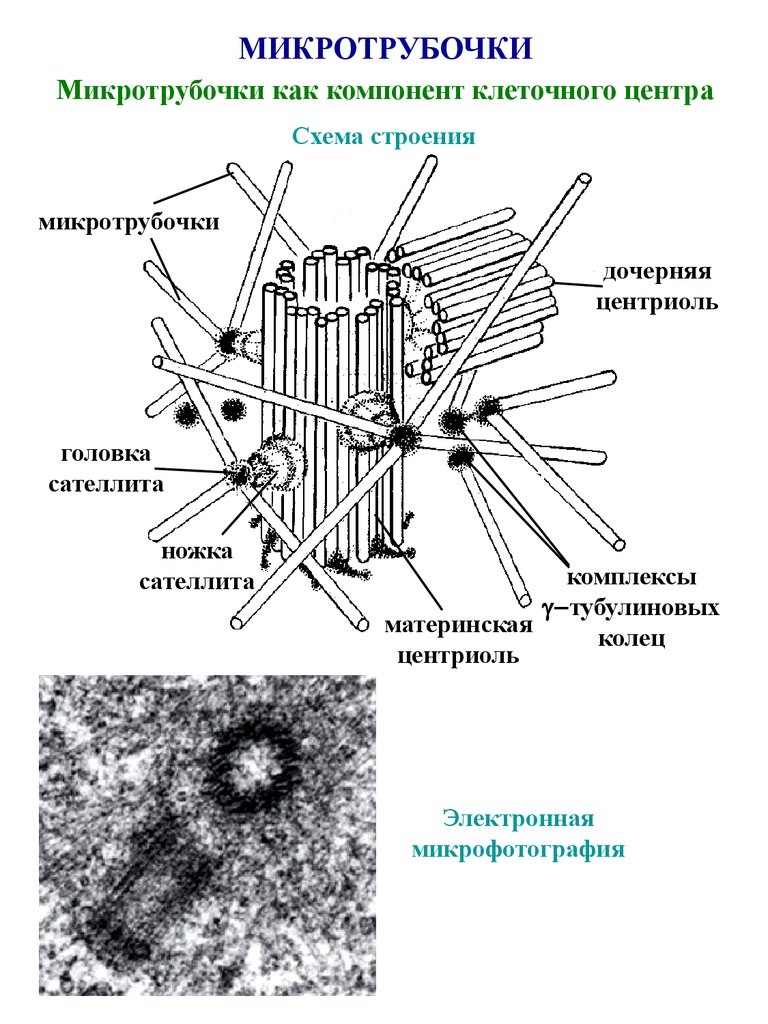 Будущие исследования должны определить дальнейшие детали регуляции процесса сборки центриолей de novo, уточнить, как возникает асимметрия сестринских центриолей.
Будущие исследования должны определить дальнейшие детали регуляции процесса сборки центриолей de novo, уточнить, как возникает асимметрия сестринских центриолей.
Реснички необходимы сперматозоидам P. patens для того, чтобы плыть к яйцеклетке, и авторы предполагают в дальнейшем исследовать, не влияет ли асимметрия базальных тел на те или иные свойства ресничек, влияющие на подвижность.
Источник
Sónia Gomes Pereira, et al. The 3D architecture and molecular foundations of de novo centriole assembly via bicentrioles // Current Biology, Published August 24, 2021, DOI:
10.1016/j.cub.2021.07.063.
Et сetera Клеточная биология
10 класс. Биология. Митоз. Амитоз — Митоз. Амитоз
Комментарии преподавателя
Митоз – процесс непрямого деления соматических клеток эукариот, в результате которого генетический материал материнской клетки (после удвоения в синтетическом периоде интерфазы) равномерно распределяется между дочерними клетками.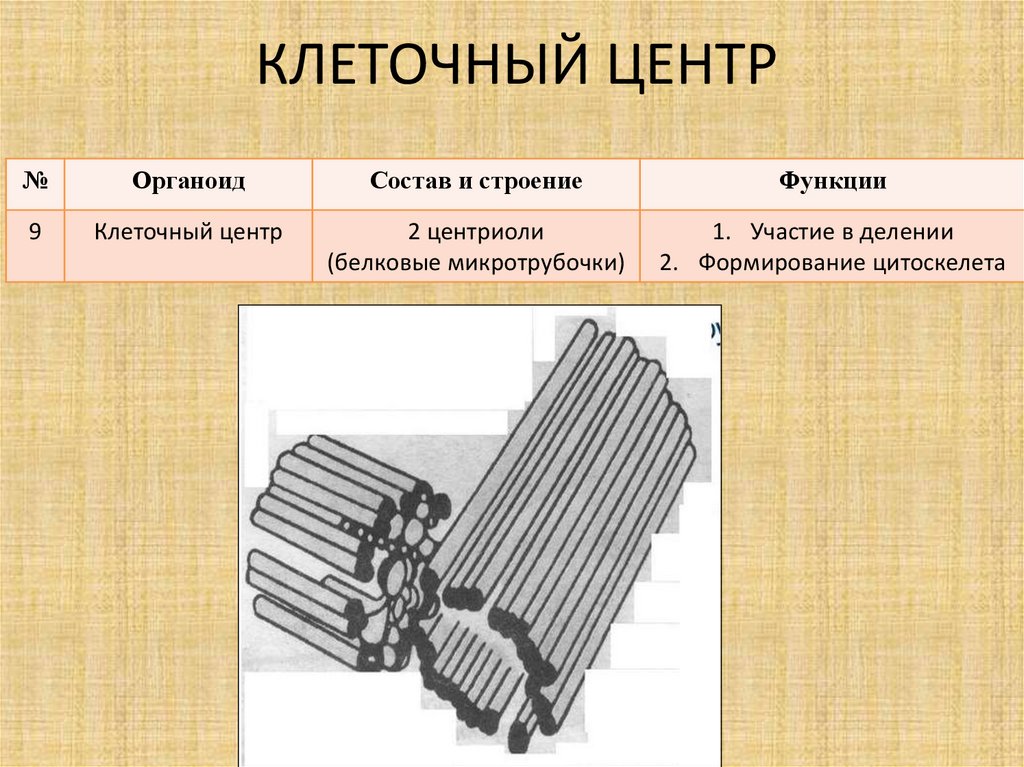 Он является основным способом деления клеток эукариот.
Он является основным способом деления клеток эукариот.
Продолжительность митоза клеток животных составляет от 30 до 60 минут, а клеток растительных организмов – от 2-х до 3-х часов. Митоз включает в себя 2 этапа, а именно: деление ядра, или кариокинез, и деление цитоплазмы или цитокинез.
Центриоли и образование веретена деления
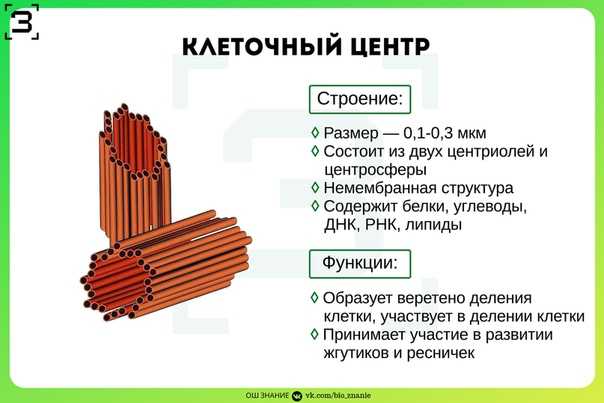
Центриоли – органеллы, которые располагаются в цитоплазме около ядерной оболочки в клетках животных и низших растений. Две центриоли, образующие пару, лежат перпендикулярно друг другу. Каждая центриоль имеет примерно 50 нм в длину и 200 нм в диаметре. Она состоит из девяти групп микротрубочек по три в каждой группе (рис. 1).
Рис. 1. Строение центриолей
Полагают, что соседние тройки микротрубочек соединены между собой фибриллами. Нити веретена деления имеет трубчатую форму и в диаметре около 25 нм.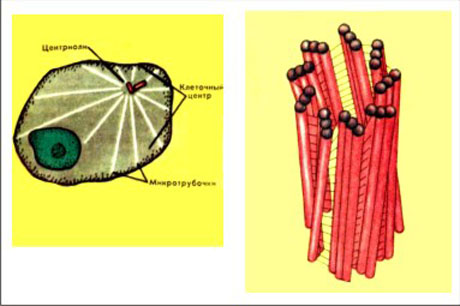 Они образуются во время митоза или мейоза и состоят из микротрубочек, которые, в свою очередь, состоят из белка тубулина.
Они образуются во время митоза или мейоза и состоят из микротрубочек, которые, в свою очередь, состоят из белка тубулина.
Раньше считали, что центриоли играют главную роль в организации нитей веретена деления, но потом от этого мнения пришлось отказаться, потому что у высших растений нити веретена деления образуются, а центриоли – нет.
Нити веретена деления бывают двух типов. Первый тип протягивается от одного полюса к другому, а второй тип образует пучки, которые присоединяются к центромерам хромосом. Перемещение хромосом во время анафазы связано с движением этих нитей относительно друг друга.
Как показывает результат электронных микроскопических исследований, между этими нитями имеются поперечные мостики.
Деление цитоплазмы, или цитокинез
Деление цитоплазмы называют цитокинезом, оно обычно следует за телофазой и различается у животных и растительных клеток. У животных клеток плазматическая мембрана во время телофазы начинает впячиваться внутрь на том уровне, где прежде располагался экватор веретена. Полагают, этот процесс происходит под действием микрофиламентов (рис. 2).
У животных клеток плазматическая мембрана во время телофазы начинает впячиваться внутрь на том уровне, где прежде располагался экватор веретена. Полагают, этот процесс происходит под действием микрофиламентов (рис. 2).
Рис. 2. Телофаза и цитокинез у животных
В результате этого процесса образуется непрерывная борозда, опоясывающая клетку по экватору (рис. 2).
В конце концов, клеточные мембраны в области борозды смыкаются, полностью разделяя обе клетки.
В растительных клетках нити веретена деления во время телофазы начинают исчезать, сохраняясь лишь в области экваториальной пластинки.
Рис. 3. Механизм цитокинеза у растений
Здесь они сдвигаются к периферии клетки, число их увеличивается, и они образуют бочковидное тельце, которое носит название фрагмопласт (рис. 3).
В эту область перемещаются микротрубочки, рибосомы, митохондрии, эндоплазматическая сеть (ЭПС) и аппарат Гольджи (АГ). ЭПС и АГ образуют множество мелких пузырьков с жидкостью.
ЭПС и АГ образуют множество мелких пузырьков с жидкостью.
Пузырьки появляются в центре клетки, а затем, направляемые микротрубочками, сливаются друг с другом, образуя клеточную пластинку. Клеточная пластинка располагается в экваториальной плоскости.
Пластинка разрастается и, в конце концов, сливается с клеточной стенкой родительской клетки. Образуется так называемая первичная клеточная стенка. А вторичная клеточная стенка образуется путем отложения на первичной клеточной стенке целлюлозы и лигнина.
Митоз в животных и растительных клетках
Самое важное событие, которое происходит в митозе, это равномерное распределение генетического материала. Митоз в животных и растительных клетках почти одинаков, но имеется ряд различий, которые указаны в нашей таблице (рис. 4). В растительной клетке центриолей нет, а в животной клетке центриоли имеются, в растительной клетке образуется клеточная пластинка, у животной клетки не образуется.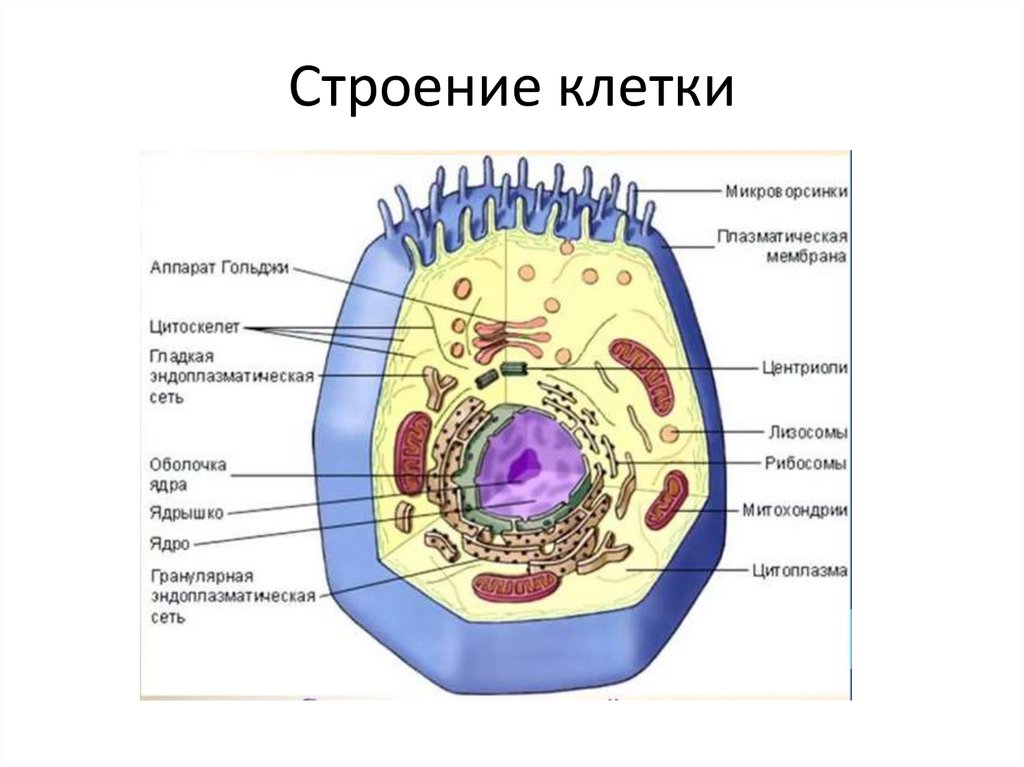
Рис. 4. Сравнение особенностей митоза в клетках животных и растений
В растительных клетках при цитокинезе не образуется перетяжка, а в животных клетка образуется. Митозы в растительных клетках происходят, в основном, в меристемах, а в животных клетках митозы происходят в различных тканях и участках организма.
Митоз подразделяют на четыре последовательные фазы: профазу, метафазу, анафазу и телофазу (рис. 5). Интерфаза – основная стадия жизненного цикла клетки (см. предыдущий урок), является подготовкой к делению или предшествует гибели клетки, поэтому фазой митоза не является.
Рис. 5. Интерфаза и следующие за ней фазы митоза: профаза, метафаза, анафаза и телофаза
В профазе происходит спирализация ДНК в ядре и, рассматривая клетку через микроскоп, можно увидеть туго скрученные хромосомы (рис.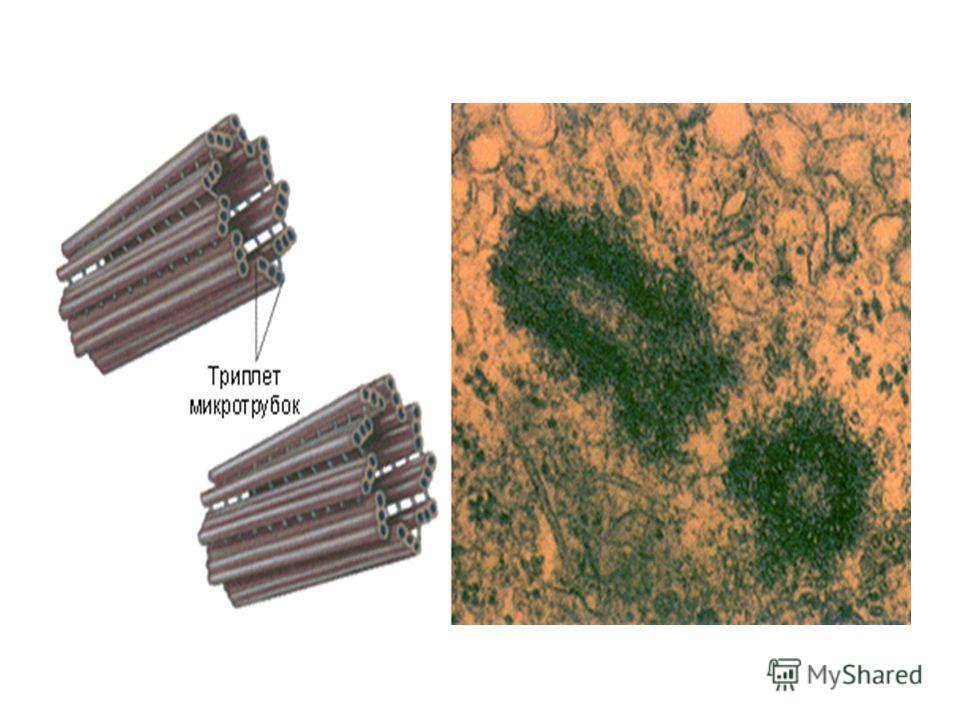 6).
6).
Рис. 6. Профаза митоза
Обычно видно, что каждая хромосома состоит из двух хроматид и объединяющих областей – центромер. Ядрышки на этом этапе исчезают. У животных клеток и у низших растений центриоли расходятся к полюсам клетки.
От каждой центриоли в виде лучей отходят короткие микротрубочки. Они образуют структуру, по форме напоминающую звезду.
Рис. 7. Профаза митоза в клетках животных и растений
К концу профазы (рис. 7) ядерная оболочка распадается или растворяется и микротрубочки начинают образовывать веретено деления (рис. 8).
Рис. 8. Завершение профазы и переход к метафазе
Следующая фаза – метафаза. Хромосомы располагаются таким образом, что их центромеры находятся на плоскости экватора клетки (рис. 9).
Рис. 9. Метафаза: веретено деления. На экваторе – метафазная пластинка.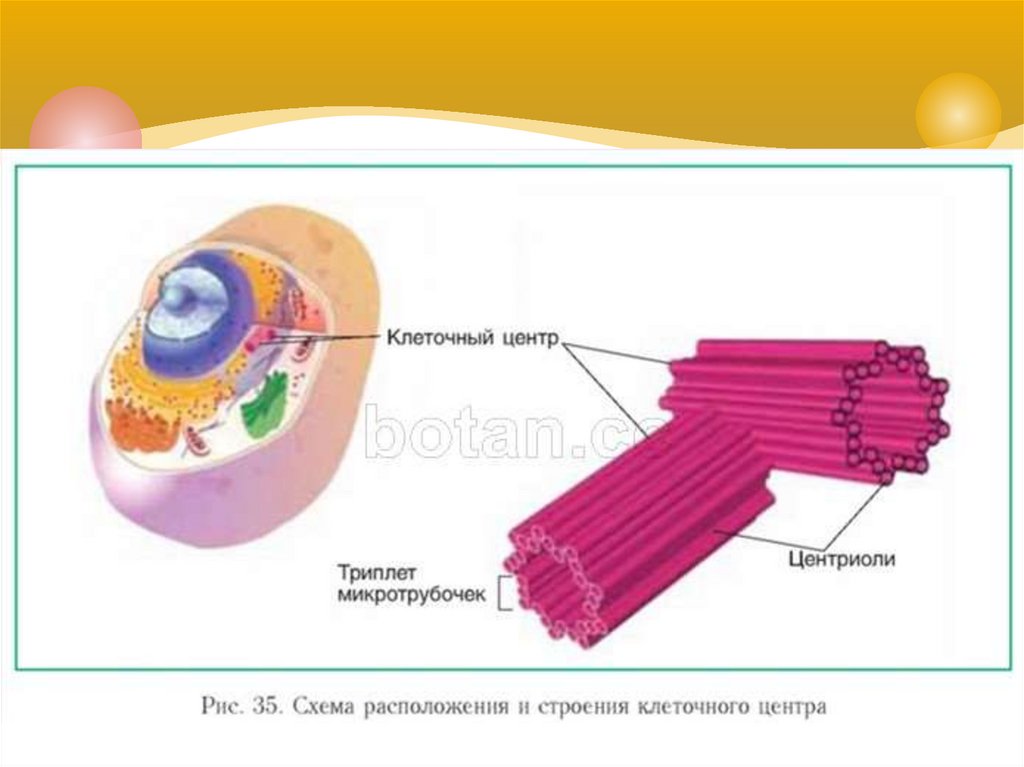
Образуется так называемая метафазная пластинка (рис. 10), которая состоит из хромосом. Нити веретена деления прикрепляются к центромерам каждой хромосомы.
Рис. 10. Метафаза. Окрашенный препарат. Веретено деления образованно центромерами (голубые), микрофибриллами (фиолетовые) и хромосомами метафазной пластинки – желтые.
Анафаза – очень короткая фаза (рис. 11). Каждая хромосома продольно расщепляется на две идентичные хроматиды, которые расходятся к противоположным полюсам клетки, теперь их называют дочерними хромосомами (или хроматидами).
Рис. 11. Анафаза митоза
За счет идентичности дочерних хромосом у двух полюсов клетки оказывается одинаковый генетический материал. Тот же, что был в клетке до начала митоза. Стоит отметить, что при этом возле каждого полюса носителей информации – молекул ДНК, компактно упакованных в хромосомы – в два раза меньше, чем в исходной клетке.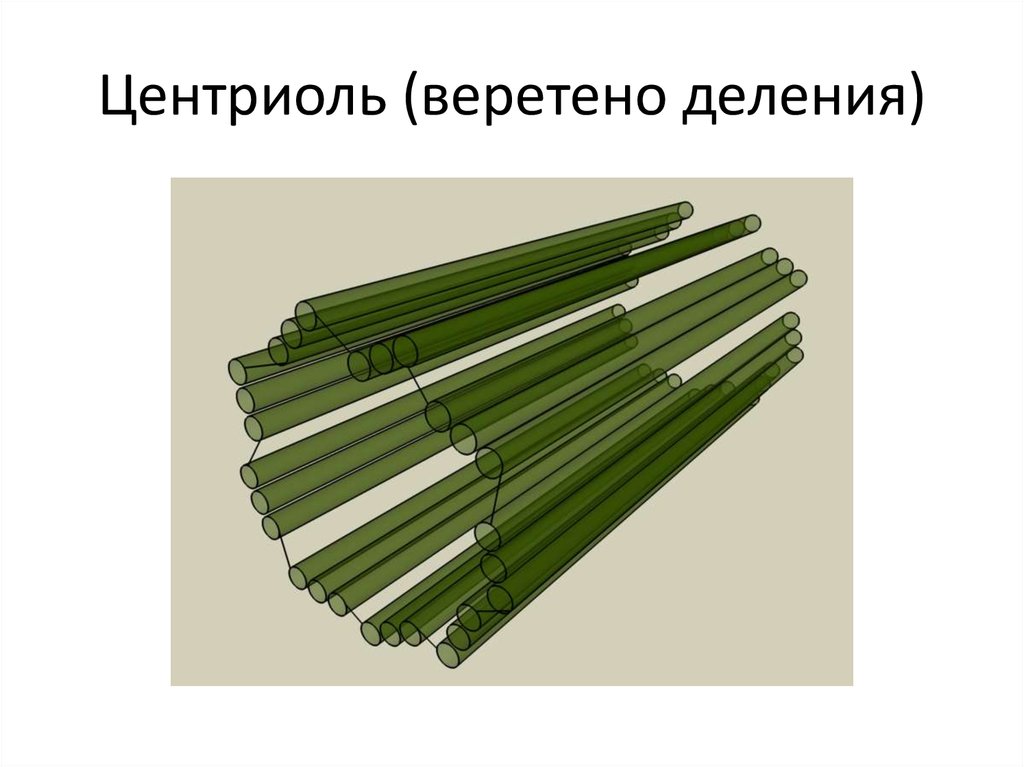
Телофаза – последняя фаза, дочерние хромосомы деспирализируются у полюсов клетки и становятся доступными для транскрипции, начинается синтез белков, формируются ядерные оболочки и ядрышки (рис. 12).
Рис. 12. Телофаза митоза в клетках животных и растений
Нити веретена деления распадаются. На этом кариокинез заканчивается, и начинается цитокинез (рис. 13), при этом у животных клеток в экваториальной плоскости возникает перетяжка. Она углубляется до тех пор, пока не происходит разделение двух дочерних клеток.
Рис. 13. Цитокинез
В образовании перетяжки важную роль играют структуры цитоскелета. Цитокинез у растительных клеток происходит иначе, поскольку растения имеют жесткую клеточную стенку, и они не делятся с образованием перетяжки, а образуют внутриклеточную перегородку.
Митоз, в первую очередь, дает генетическую стабильность. В результате митоза образуются два ядра, которые содержат столько же хромосом, сколько и было их в материнской или родительской клетках.
В результате митоза образуются два ядра, которые содержат столько же хромосом, сколько и было их в материнской или родительской клетках.
Эти хромосомы образуются путем точной репликации молекулы ДНК родительских хромосом, в результате чего гены их содержат совершенно одинаковую наследственную информацию.
Таким образом, дочерние клетки генетически идентичны родительской клетке, поскольку митоз не может внести никаких изменений в наследственную информацию. Клеточные популяции, полученные путем митоза от родительских клеток, обладают генетической стабильностью.
Митоз необходим для нормального роста и развития многоклеточных организмов, поскольку в результате митоза количество клеток увеличивается.
Митоз является одним из главных механизмов роста многоклеточных эукариот.
Митоз лежит в основе бесполого размножения многих животных и растений, обеспечивает регенерацию утраченных частей (например, конечностей ракообразных), а также замещение клеток, происходящее в многоклеточном организме.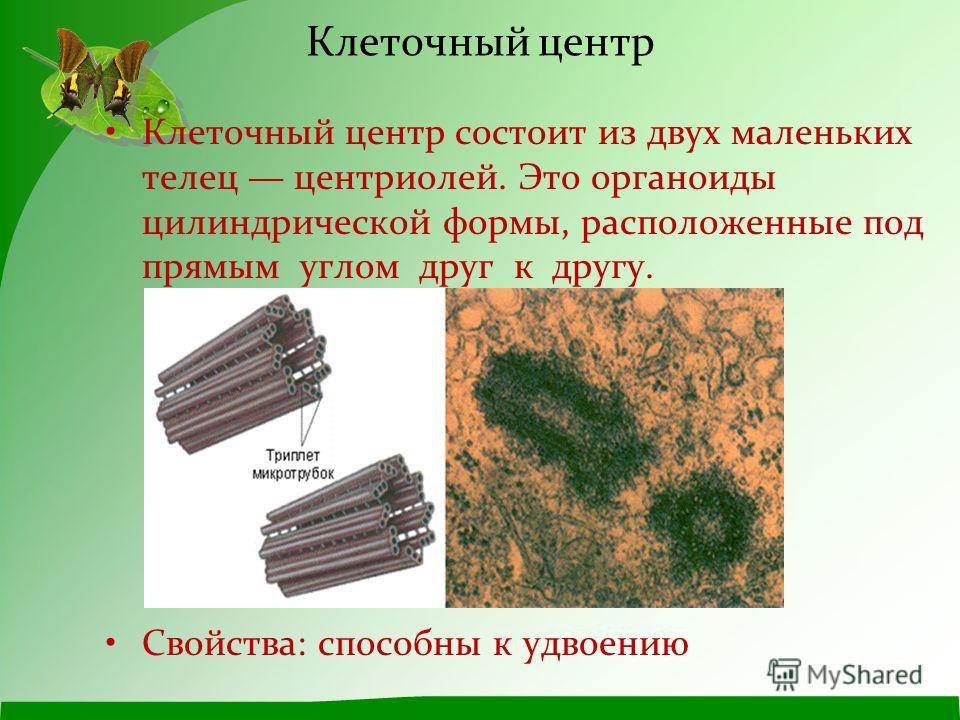
Рис. 14. Амитоз – прямое деление
Амитоз – прямое деление клеток. Амитоз встречается у эукариот достаточно редко. При амитозе ядро начинает делиться без видимых предварительных изменений. При этом не обеспечивается равномерное распределение генетического материала между дочерними клетками. Иногда при амитозе не происходит цитокинеза, то есть деления цитоплазмы, и тогда образуется двухъядерная клетка (рис. 14).
Если же все-таки произошло деление цитоплазмы, то велика вероятность того, что обе дочерние клетки будут неполноценными. Амитоз чаще встречается в опухолевых или отмеряющих тканях.
источник конспекта — http://interneturok.ru/ru/school/biology/10-klass/undefined/mitoz-amitoz
источник видео — http://www.youtube.com/watch?v=peeUBmOgx8k
источник видео — http://www.youtube.com/watch?v=Nz0j4oKIDUo
источник презентации — http://prezentacii.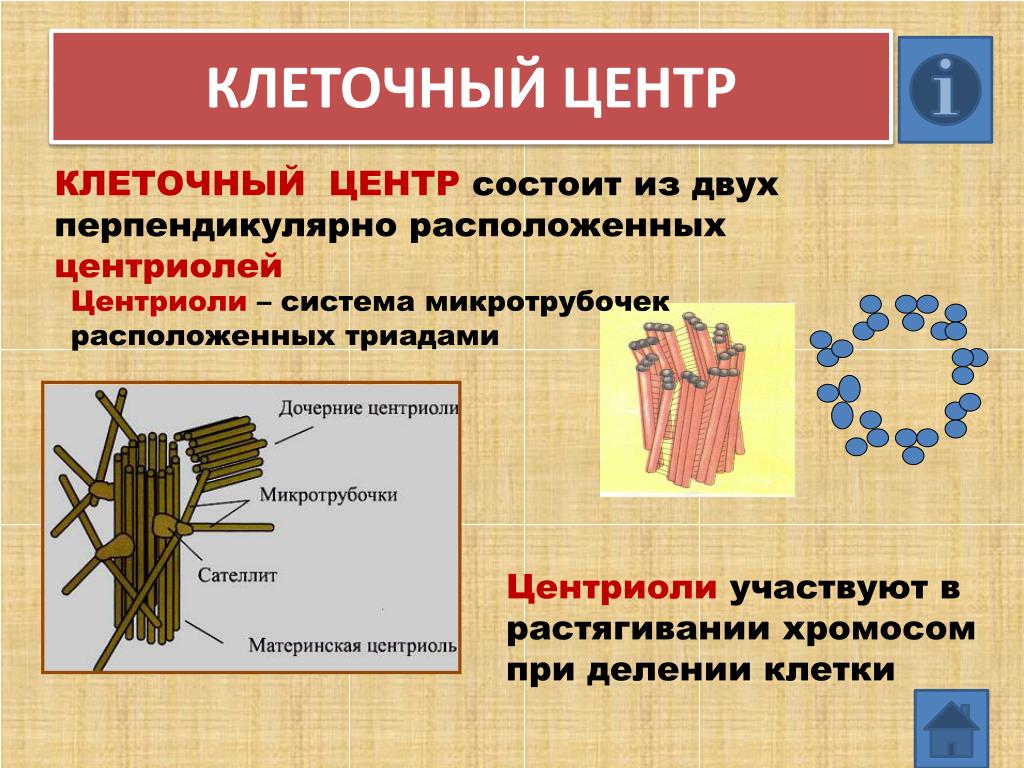 com/biologiya/14211-zhiznennyy-cikl-kletki-mitoz-amitoz-10-klass.html
com/biologiya/14211-zhiznennyy-cikl-kletki-mitoz-amitoz-10-klass.html
Британское общество клеточной биологии
Краткий обзор: Обнаруженная только в клетках животных и некоторых низших растений, центриоль состоит из коротких отрезков микротрубочек, расположенных параллельно друг другу и расположенных вокруг центральной полости в форме цилиндра.
В клетках животных центриоли расположены в центросоме и образуют ее часть, где они представляют собой парные структуры, расположенные под прямым углом друг к другу. В этом контексте они, возможно, участвуют в сборке веретена во время митоза. Центросома расположена в цитоплазме вне ядра, но часто рядом с ним.
На базальных концах ресничек и жгутиков также можно обнаружить одну центриоль. В этом контексте оно называется «базальным тельцем» и связано с ростом и работой микротрубочек в ресничке или жгутике.
Чтобы просмотреть изображение центриолей в сперматозоидах дрозофилы, интерпретированное с использованием технологии CIMR GridPoint, НАЖМИТЕ ЗДЕСЬ
Центриоли представляют собой нечто загадочное
Центриоли присутствуют в (1) клетках животных и (2) базальных область ресничек и жгутиков у животных и низших растений (например, у хламидомонады). В ресничках и жгутиках центриоли называются «базальными тельцами», но их можно считать взаимопревращаемыми.
В ресничках и жгутиках центриоли называются «базальными тельцами», но их можно считать взаимопревращаемыми.
Центриоли отсутствуют в клетках высших растений.
Когда клетки животных подвергаются митозу, некоторые считают, что они получают пользу от присутствия центриолей, которые, по-видимому, контролируют образование веретенообразных волокон и которые впоследствии влияют на разделение хромосом. Однако исследования показали, что митоз может происходить в клетках животных после разрушения центриолей. Иногда кажется, что это происходит за счет аномалий развития веретена и последующих проблем с разделением хромосом. Недавние исследования также показывают, что эмбрионы Drosophila арестовывают очень рано, если репликация центриолей не может происходить.
У высших растений митоз вполне удовлетворительно протекает с микротрубочками, образующими веретенообразные волокна, но без помощи центриолей. Таким образом, функция центриолей остается загадкой.
Структура
Центриоль состоит из коротких отрезков микротрубочек, расположенных в форме открытого цилиндра длиной около 500 нм и диаметром 200 нм.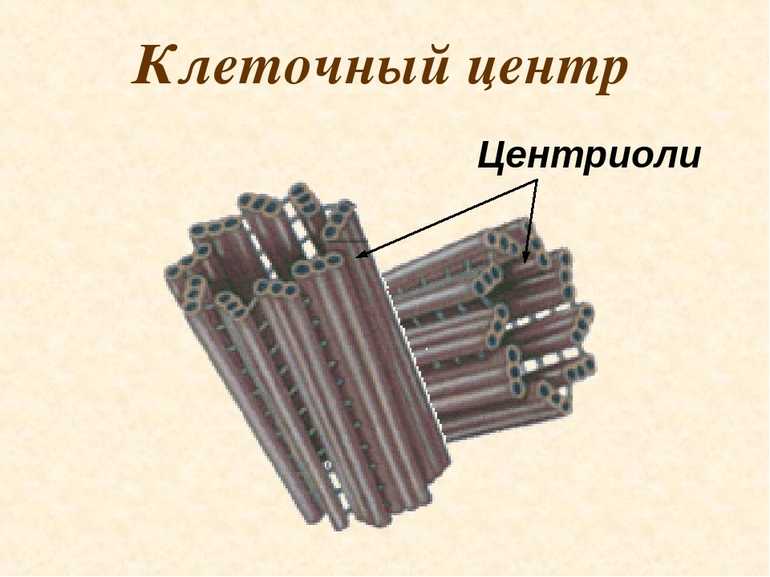 Микротрубочки, образующие стенку цилиндра, сгруппированы в девять наборов пучков по три микротрубочки в каждом.
Микротрубочки, образующие стенку цилиндра, сгруппированы в девять наборов пучков по три микротрубочки в каждом.
В ресничках и жгутиках, где центриоли лежат в основе структуры и называются базальными тельцами, архитектура стенок и полостей немного отличается. Помимо стенок цилиндра, состоящих из девяти наборов пучков из трех микротрубочек, имеются стенки из девяти наборов из двух пучков. В обоих типах имеется центральная матрица, от которой расходятся спицы, как в тележном колесе.
В клетках животных центриоли обычно расположены парами, причем цилиндрические центриоли расположены под прямым углом друг к другу.
Центриоли организуют вокруг себя «облако» белкового материала; это перицентриолярный материал (ПКМ). Вместе они составляют всю важную центросому.
Функция
Центриоли функционируют как пара в большинстве клеток животных, но как одна центриоль или базальное тельце в ресничках и жгутиках.
Центриоли в парах
Клетки, вступающие в митоз, имеют центросому, содержащую две пары центриолей и связанный с ними перицентриолярный материал (ПКМ). Во время профазы центросома делится на две части, и пара центриолей мигрирует к каждому концу или полюсу на внешней стороне ядерной мембраны или оболочки. В этот момент микротрубочки образуются на внешнем крае перицентриолярного материала и растут в радиальной форме. Пара центриолей и ПКМ называется звездой. Микротрубочки от звездочки на одном полюсе растут к звездочке на противоположном полюсе. Эти микротрубочки называются волокнами веретена. Некоторые из них прикрепляются центромерами к хромосомам, выстроенным на «экваторе» делящейся клетки. Другие, хотя и не прикрепленные к хроматидам/хромосомам центромерами, будут способствовать раздвижению двух частей делящейся клетки.
Во время профазы центросома делится на две части, и пара центриолей мигрирует к каждому концу или полюсу на внешней стороне ядерной мембраны или оболочки. В этот момент микротрубочки образуются на внешнем крае перицентриолярного материала и растут в радиальной форме. Пара центриолей и ПКМ называется звездой. Микротрубочки от звездочки на одном полюсе растут к звездочке на противоположном полюсе. Эти микротрубочки называются волокнами веретена. Некоторые из них прикрепляются центромерами к хромосомам, выстроенным на «экваторе» делящейся клетки. Другие, хотя и не прикрепленные к хроматидам/хромосомам центромерами, будут способствовать раздвижению двух частей делящейся клетки.
Одиночное центриоль или базальное тельце.
В основании каждой реснички или жгутика находится одна центриоль. Эта структура и связанный с ней перицентриолярный материал образуют микротрубочки в линейном направлении. Эти микротрубочки формируют большую часть внутренней части ресничек и жгутиков и, используя белковые моторы, в значительной степени ответственны за механические аспекты их движения.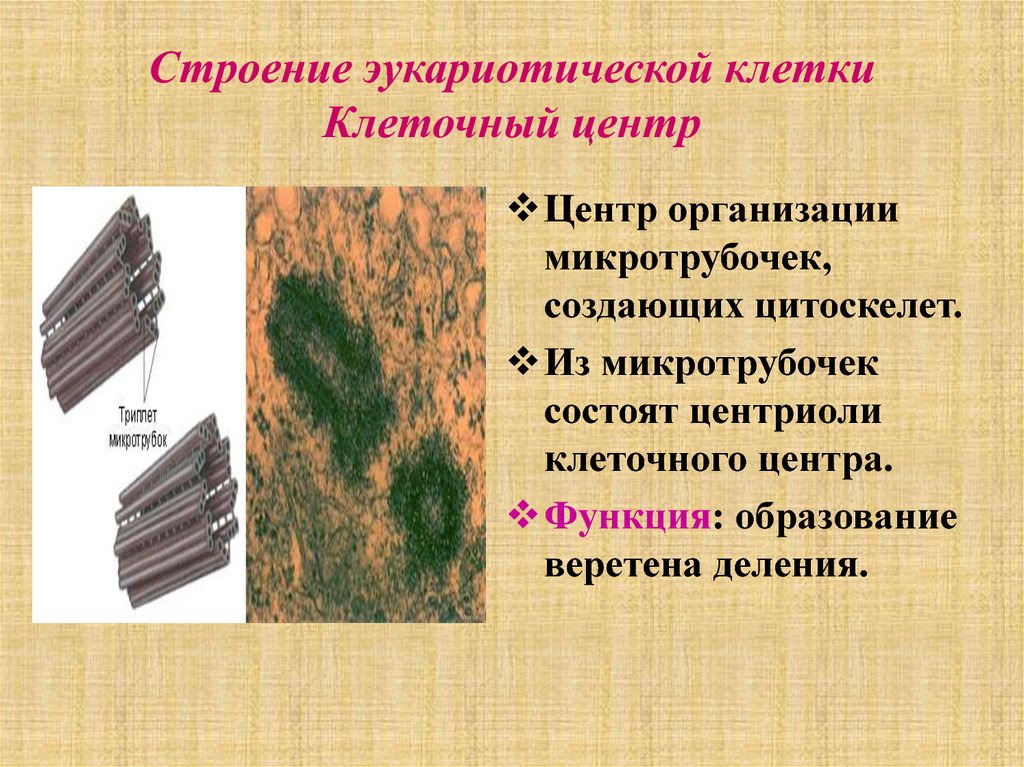 Центриоли в основании каждого из них также, по-видимому, в некоторой степени направляют и контролируют движение ресничек и жгутиков.
Центриоли в основании каждого из них также, по-видимому, в некоторой степени направляют и контролируют движение ресничек и жгутиков.
Репликация
В клетках, где центриоли присутствуют в виде пары, репликация происходит в течение всего клеточного цикла. В фазе G1 два центриолевых цилиндра очень незначительно расходятся друг от друга. Во время фазы S новые цилиндры микротрубочек формируются рядом и под прямым углом к двум «материнским» цилиндрам. Две пары центриолей держатся очень близко друг к другу до профазной стадии митоза. В этот момент они разделяются, и обе пары центриолей движутся по внешней поверхности ядерной оболочки к противоположным концам или «полюсам» клетки, образуя астральные полюса делящейся клетки.
Резюме
- Центриоли встречаются в виде парных цилиндрических органелл вместе с перицентриолярным материалом (ПКМ) в центросоме животной клетки.
- Центриоли обнаружены в виде одиночных структур в ресничках и жгутиках в клетках животных и некоторых низших растительных клетках.

- Центриоли построены из микротрубочек.
- В клетках животных центриоли организуют перицентриолярный материал с образованием микротрубочек, включая волокна митотического веретена.
- Центриоли представляют собой нечто вроде загадки; они, по-видимому, влияют на исход митоза в клетках животных. При их наличии происходит удовлетворительное деление, но без них митоз все же имеет место, но иногда с неудовлетворительным исходом. Центриоли отсутствуют в клетках высших растений, но имеет место нормальный митоз с удовлетворительными результатами.

ВАЖНОЕ ЗАМЕЧАНИЕ
Иногда новая информация бросает вызов текущему мышлению только для того, чтобы быть замененной другими идеями по мере развития знаний. Так работает наука. Британское общество клеточной биологии (BSCB) стремится предоставлять достоверную и актуальную информацию, но не может нести ответственность за любую потерю оценок в результате использования информации на этом веб-сайте, которая может противоречить общепринятым взглядам проверяющих органов.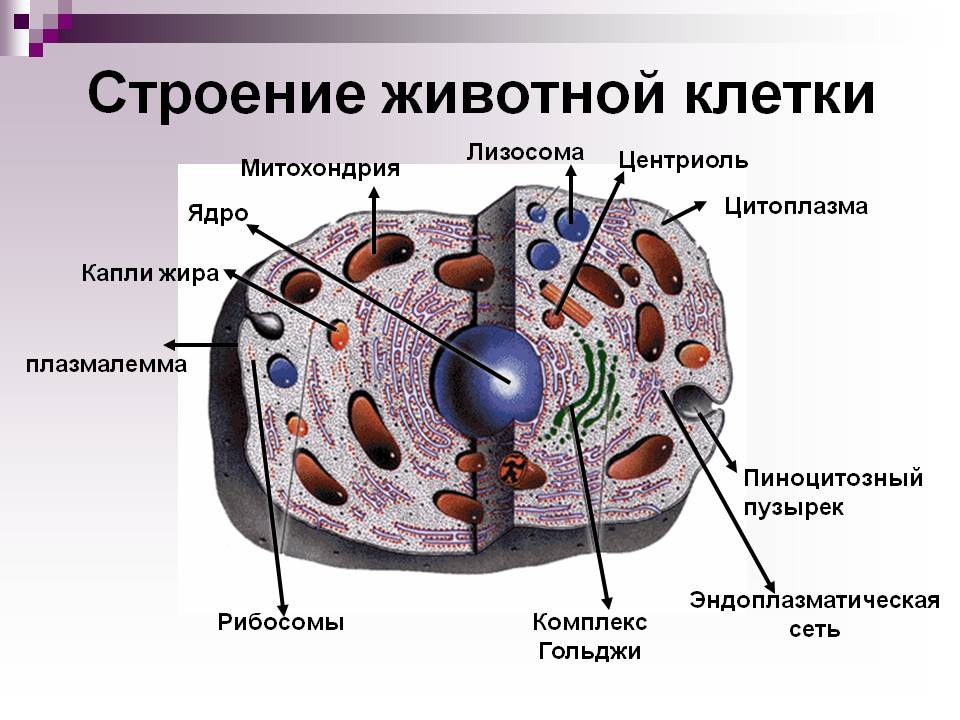 .
.
| Центриоли Обнаруженные только в клетках животных, эти парные органеллы обычно располагаются вместе вблизи ядра в центросоме , зернистой массе, которая служит организующим центром для микротрубочек. Центриоли играют заметную роль в клеточном делении. Во время интерфазы животной клетки центриоли и другие компоненты центросомы дублируются, хотя ученые еще не уверены, как это происходит. Сначала две пары центриолей остаются в непосредственной близости друг от друга, но по мере инициации митоза исходная центросома делится, и пары разделяются так, что один набор центриолей располагается в каждом из новых центров организации микротрубочек. Эти новые центры излучают микротрубочки в звездчатых скоплениях, известных как астры . По мере того, как звезды перемещаются к противоположным полюсам клеток, микротрубочки с помощью центриолей организуются в веретенообразные образования, охватывающие клетку (см. рис. 2). Эти волокна веретена действуют как направляющие для выравнивания хромосом, поскольку они разделяются позже в процессе клеточного деления. Хотя центриоли играют роль в митозе животных клеток, растительные клетки способны размножаться без них. Поэтому исследователи были очень заинтересованы в точном определении того, насколько важны эти органеллы на самом деле. Исследования показали, что некоторые клетки животных, особенно женские гаметы (ооциты), могут успешно делиться даже при разрушении их центриолей. Однако некоторые исследователи обнаружили также, что отсутствие центриолей в клетках животных связано с повышенным числом ошибок деления и существенной задержкой митотического процесса, особенно перед сегрегацией хромосом. Следовательно, было высказано предположение, что центриоли развились как усовершенствование клетки, что сделало митоз гораздо более эффективным и менее подверженным ошибкам процессом. В клетках, которые имеют реснички или жгутики, присутствуют базальных телец , которые демонстрируют ту же структурную форму, что и центриоли. Эти сборки расположены, однако, вблизи клеточной поверхности у основания каждой реснички или жгутика, а не в центросоме вблизи ядра. НАЗАД К СТРУКТУРЕ ЖИВОТНЫХ КЛЕТОК Вопросы или комментарии? Отправить нам письмо. © 1995-2022 автор | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
 gif»>
gif»> Внутри центросомы центриоли расположены под прямым углом друг к другу, как показано на рис. 1. Каждая центриоль состоит из девяти пучков микротрубочек (по три на пучок), расположенных кольцом.
Внутри центросомы центриоли расположены под прямым углом друг к другу, как показано на рис. 1. Каждая центриоль состоит из девяти пучков микротрубочек (по три на пучок), расположенных кольцом.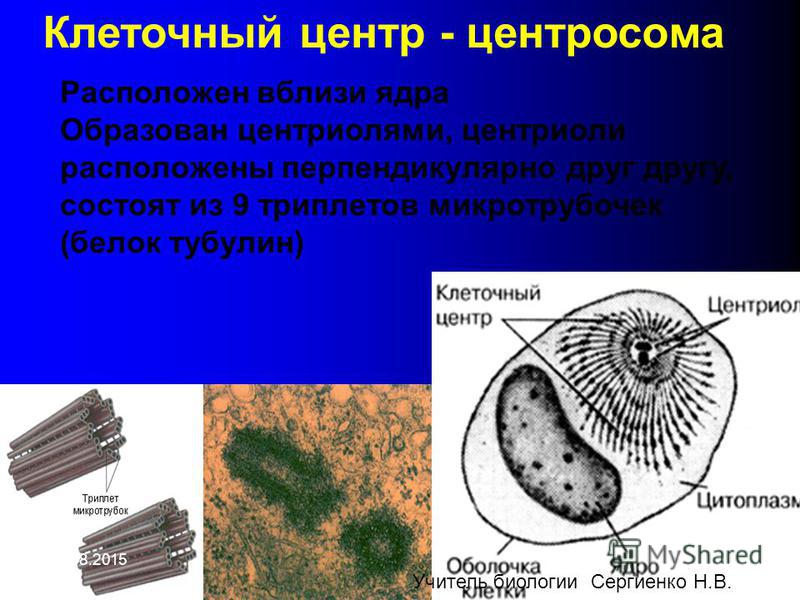
 Базальные тельца закрепляются в своих цитоплазматических положениях с помощью так называемой системы корешков в клетке. У некоторых организмов, таких как одноклеточные Chlamydomonas базальные тельца меняют свое расположение и функционально превращаются в центриоли до митотического процесса.
Базальные тельца закрепляются в своих цитоплазматических положениях с помощью так называемой системы корешков в клетке. У некоторых организмов, таких как одноклеточные Chlamydomonas базальные тельца меняют свое расположение и функционально превращаются в центриоли до митотического процесса.