
Содержание
Как избавиться от тополиного пуха – Наука – Коммерсантъ
Тополь — незаменимое для мегаполисов растение: он неприхотлив, легко размножается, быстро растет, эффективно очищает воздух и благодаря большой листовой поверхности выделяет кислорода в несколько раз больше, чем другие деревья, озеленяющие города средней полосы России. Однако у тополя есть серьезный недостаток: взрослые деревья образуют пух, а он пожароопасен, забивает водостоки и системы вентиляции, собирает и переносит пыльцу и пыль — аллергены.
Мужские (сверху) и женские (снизу) генеративные органы (сережки) тополя
Фото: РИА Новости
Мужские (сверху) и женские (снизу) генеративные органы (сережки) тополя
Фото: РИА Новости
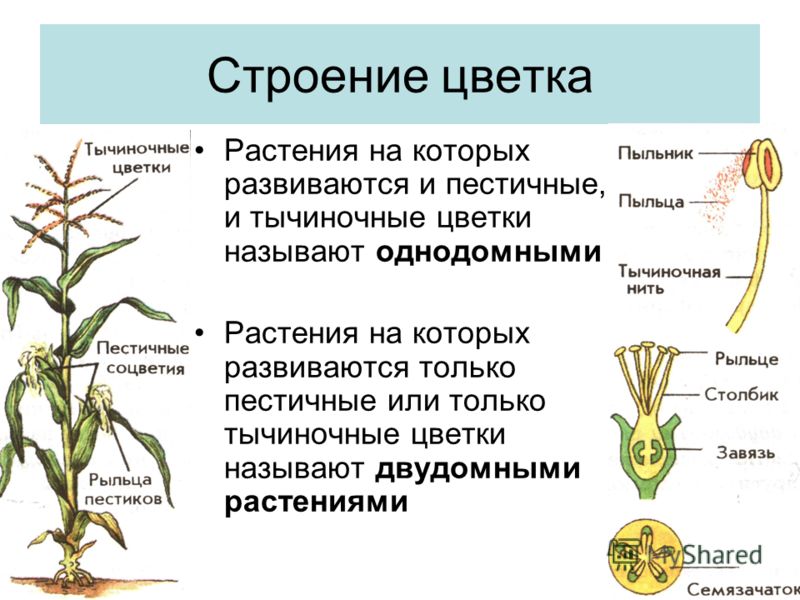
Тополь относится к двудомным растениям: на одних деревьях образуются только женские генеративные органы, а на других — только мужские. Если для животных такое разделение полов является привычным, то у растений оно встречается достаточно редко — только у 5–6% видов. То, что мы называем пухом, на самом деле является опушенными семенами, которые образуют только женские деревья, и именно они приносят вред городам и их жителям. Для эффективного использования тополя в озеленении необходимо выращивание исключительно мужских растений, причем выбирать их необходимо еще на начальных стадиях развития, а не через семь-десять лет, когда тополя начинают цвести и уже являются большими деревьями, а для этого требуются знания о механизмах определения пола и генетических отличиях мужских и женских растений.
То, что мы называем пухом, на самом деле является опушенными семенами, которые образуют только женские деревья, и именно они приносят вред городам и их жителям. Для эффективного использования тополя в озеленении необходимо выращивание исключительно мужских растений, причем выбирать их необходимо еще на начальных стадиях развития, а не через семь-десять лет, когда тополя начинают цвести и уже являются большими деревьями, а для этого требуются знания о механизмах определения пола и генетических отличиях мужских и женских растений.
Установление молекулярных механизмов детерминации пола тополя и оценка его генетического разнообразия являются одними из основных направлений исследований, проводимых нами в Институте молекулярной биологии им. В. А. Энгельгардта РАН. Ключевым используемым методом служит высокопроизводительное секвенирование, позволяющее быстро генерировать данные о нуклеотидных последовательностях геномов, транскриптомов или отдельных генов с высокой точностью. Нами получены высококачественные сборки геномов тополя, что позволило идентифицировать отличия мужских и женских растений на молекулярном уровне и дополнить наши знания о механизме детерминации пола.
Кроме того, на представительной выборке образцов определены нуклеотидные последовательности ассоциированных с полом участков генома тополя и обнаружены ДНК-полиморфизмы, встречающиеся только у мужских деревьев. На основе полученных данных, а также исследований, выполненных научными коллективами из других стран, нами разработана тест-система для определения пола тополя, которая основана на методе полимеразной цепной реакции (ПЦР). Наша разработка позволяет выбирать растения нужного пола даже среди однолетних саженцев, что делает возможным использование для озеленения города только не образующих пуха мужских деревьев.
Нами также показан высокий полиморфизм тополей, растущих на территории Москвы, и предложены генетические маркеры для создания коллекции высокоадаптивных к городским условиям деревьев, максимально охватывающей существующее разнообразие и пригодной для дальнейшего размножения и эффективного использования в озеленении. Таким образом, наши работы помимо фундаментального значения привели к разработке ДНК-маркеров, которые позволят отбирать саженцы тополя, наиболее подходящие для озеленения мегаполисов, и избавиться от пуха, максимально используя все преимущества этого растения для улучшения экологии городов.
Меры борьбы с тополиным пухом весьма неоднозначны. В основном используется кардинальная обрезка деревьев, однако этот метод при значительных финансовых затратах не приносит положительных результатов: пока у тополей обрезаны ветви, они образуют мало листьев и не способны эффективно очищать воздух и выделять кислород, а когда они восстанавливаются, то начинают цвести и женские растения вновь образуют пух. Про то, что для озеленения городов нужно использовать только мужские тополя, говорится давно, однако значимых шагов в этом направлении не предпринимается.
Существует мнение, что тополя могут менять пол и сажать мужские растения бесполезно, так как все равно через какое-то время они превратятся в женские. Однако крупномасштабные исследования последних лет, проведенные учеными в различных странах и опубликованные в том числе в журналах издательства Nature, показали, что пол тополя определяется генетически, поэтому его смена не может быть повсеместным явлением. Для большинства видов тополей характерна XY-система детерминации пола. Проведенные нами исследования на представительных выборках растений, в том числе подвергнутых кардинальной обрезке, из различных районов города Москвы продемонстрировали полное соответствие между генотипом и фенотипом: все деревья с Y-хромосомой образовывали мужские генеративные органы, а без нее — женские. Напрашивается вывод, что причина распространения в городах женских растений тополя, образующих пух, связана с использованием посадочного материала обоих полов.
Проведенные нами исследования на представительных выборках растений, в том числе подвергнутых кардинальной обрезке, из различных районов города Москвы продемонстрировали полное соответствие между генотипом и фенотипом: все деревья с Y-хромосомой образовывали мужские генеративные органы, а без нее — женские. Напрашивается вывод, что причина распространения в городах женских растений тополя, образующих пух, связана с использованием посадочного материала обоих полов.
Замену тополю для озеленения наиболее экологически неблагоприятных территорий средней полосы России найти сложно, а использование достижений современной науки может решить проблему пуха благодаря посадке только мужских саженцев. Реализация такого проекта улучшит экологию, снизит заболеваемость, повысит качество жизни населения и увеличит его социальную и трудовую активность, что будет иметь долгосрочный экономический эффект. Именно в ближайшие годы остро встанет проблема с естественным старением растущих сейчас тополей, связанная в том числе с повышением ломкости древесины деревьев старше 40 лет. Вопрос замены старых тополей на новые нельзя оставлять без внимания, так как если старые растения будут просто убраны, то город останется без своих «легких», что нанесет ущерб здоровью жителей. Посадку деревьев и их смену необходимо проводить в несколько этапов, чтобы обеспечить постоянный высокий уровень зеленых насаждений среднего возраста, а древесину заменяемых партий тополей можно использовать в промышленности — она высоко ценится при производстве фанеры и бумаги. В то же время для реализации такого масштабного проекта требуется поддержка на федеральном или региональном уровне, в первую очередь Москвы — крупнейшего мегаполиса.
Вопрос замены старых тополей на новые нельзя оставлять без внимания, так как если старые растения будут просто убраны, то город останется без своих «легких», что нанесет ущерб здоровью жителей. Посадку деревьев и их смену необходимо проводить в несколько этапов, чтобы обеспечить постоянный высокий уровень зеленых насаждений среднего возраста, а древесину заменяемых партий тополей можно использовать в промышленности — она высоко ценится при производстве фанеры и бумаги. В то же время для реализации такого масштабного проекта требуется поддержка на федеральном или региональном уровне, в первую очередь Москвы — крупнейшего мегаполиса.
Подписи к фотографиям
1. Мужские (сверху) и женские (снизу) генеративные органы (сережки) тополя
2. Опушенные семена на женском растении тополя
Наталья Мельникова, кандидат биологических наук, старший научный сотрудник лаборатории постгеномных исследований ИМБ РАН
от посадки до урожая. Часть 1.
Земляника — травянистое вечнозеленое многолетнее растение. Широкое применение получила потому, что отличается ранним плодоношением (на следующий год после посадки), высокой ежегодной урожайностью, сравнительно легким размножением и широкой приспособляемостью растений к различным климатическим условиям.
Широкое применение получила потому, что отличается ранним плодоношением (на следующий год после посадки), высокой ежегодной урожайностью, сравнительно легким размножением и широкой приспособляемостью растений к различным климатическим условиям.
Ягоды земляники и клубники отличаются высокими вкусовыми качествами, содержат много сахаров, кислот, минеральных солей и витамина С. Обладают лечебными свойствами, особенно при малокровии и болезнях желудка, очень полезны детям и пожилым людям.
На самом деле лечебными свойствами обладают не только ягоды, но и весь кустик! Вымойте в чистой воде листья, стебли, высушите их и сложите в бумажные мешки. В зимнее время будете иметь прекрасный чай, который рекомендуется пить 3 раза в день.
Большую ценность имеет и клубничный сок, особенно из темных ягод. Чем темнее ягода, тем в них больше витаминов.
Кстати с помощью клубники или земляники те, кто мечтает похудеть, могут устраивать разгрузочные дни. Правда, только в том случае, если нет аллергии. В течение одного дня можно съесть от 1 до 1,5кг ягод. Но больше – никаких продуктов! Запивать ягоды можно водой или чаем.
В течение одного дня можно съесть от 1 до 1,5кг ягод. Но больше – никаких продуктов! Запивать ягоды можно водой или чаем.
Часто садовую крупноплодную землянику называют клубникой. Это неверно. Эти растения относятся к различным ботаническим видам и отличаются друг от друга по ряду признаков:
Земляника относится к однодомным растениям ( тычинки и пестики расположены у нее на одном цветке), цветоносы ниже листьев; клубника же относится к двудомным растениям – мужские (тычиночные) и женские (пестичные) цветики расположены на различных растениях, кусты выше, чем у земляники, листья ребристые, цветоносы выше листьев.
Куст земляники состоит из укороченных надземных стеблей (рожков), листьев, корневища, удлиненных стеблей – усов и возникающих на них новых кустов (розетки).
Земляника главного корня не имеет. Вся корневая система, хорошо разветвленная и мочковатая, размещается в поверхностном слое почвы на глубине 10-30см, поэтому она очень влаголюбива и обработку почвы и внесение удобрений производить на небольшую глубину.
Один куст за лето может дать до 80 усов. Число укоренившихся розеток может быть 20-30, их используют как посадочный материал. Биологической особенностью земляники является усиленный рост листьев весной в течение 25-30 дней и в конце сбора урожая. Живут они 60-70 дней и отмирают. Сформированные после плодоношения листья перезимовывают и предохраняют почки и корни растения от вымерзания.
Рост стеблей начинается после открытия в начале мая, через 2-3 недели появляются цветоносы (конец мая – начало июня), цветение наступает в середине июня и длится 20-25 дней.
Соцветие земляники – полузонтик, первичный цветок наиболее развит, цветет первым, ягода его крупнее. Цветки крупные, обоеполые, самоплодные, однако перекрестное опыление повышает урожайность ягод. Плод земляники – ложная ягода, окраска сортовой признак от бледно- розовой до темно-красно-вишневой. Созревание их наступает через месяц после начала цветения, неодновременное, поэтому приходится делать 8-10 сборов.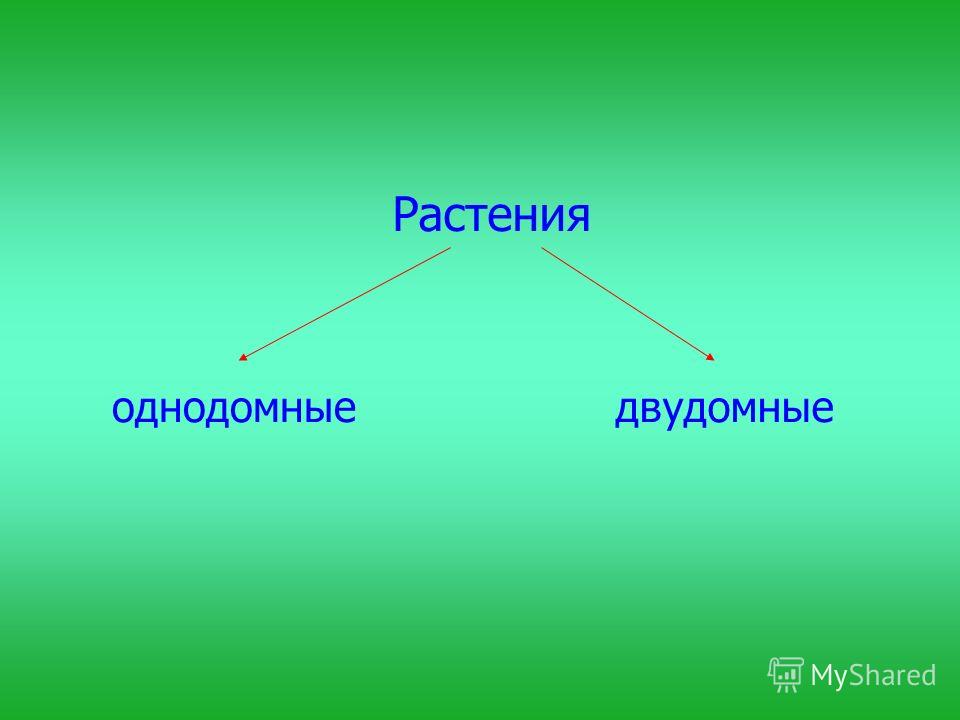
Земляника наименее зимостойка. При снижении температуры почвы до -10˚С вымерзает корневище, а затем надземная часть (-15 -18˚С). Лучшие условия зимовки для земляники складываются при высоте снега 50-60см.
Земляника – светолюбивое и теплолюбивое растение. Хорошо растет в условиях нашего короткого лета. Плохо переносит затемнение, дает мало усов и розеток, снижает урожай. Но в то же время не переносит солнцепека, на южных склонах без полива рост ее угнетен.
Земляник – влаголюбивое растение. При недостатке влаги рост замедляется, во время цветения плохо проходит опыление и завязывание ягод, они мельчают и урожай снижается. Переувлажнение почвы приводит к тому, что растение стабильно растет (жирует), но при этом урожай снижается. А от избытка влаги в местах скопления талых, дождевых, поливных и близкого расположения грунтовых вод земляника вымокает и гибнет.
Для земляники подходят практически любые почвы, но чтобы они были воздухопроницаемыми и плодородными . лучше растет на почвах со слабокислой реакцией (РН 5,5-6,5).
лучше растет на почвах со слабокислой реакцией (РН 5,5-6,5).
Семена высевают с января-февраля по апрель. Но зимой потребуется мощная подсветка, если ее нет, лучше не торопиться с посевом, провести лучше в конце марта-апреля, когда уже достаточно света.
Семена необходимо замочить в стимуляторах роста «Эпин-Экстра», «Гетероауксин», «Циркон», «Оберег», «Проросток», «Гуми».
Грунт лучше подходит универсальный для рассады , можно использовать торф, только сначала его или поливают крутым кипятком или поливают и ставят в микроволновку на 15минут. Остывший торф поливают любым биостимулятором для восстановления плодородия – «БИЭМ», «Гуми», «Гуми Кузнецова». Можно развести марганцовку и полить грунт.
Емкости использовать небольшие и высокие – 10-15см, можно выращивать в торфяных таблетках диаметром 24мм, 33мм, а можно в ящиках, горшках. Как удобнее. Есть несколько способов выращивания земляники из семян, поэтому вы можете выбрать тот способ, какой лучше подходит. Но в любом случае обязательна стратификация. Остановимся на 2 наиболее простых способах:
Но в любом случае обязательна стратификация. Остановимся на 2 наиболее простых способах:
1. Семена замачивают в стимуляторах роста (для лучшей всхожести и будущие ростки приобретают иммунитет к болезням). Когда они набухнут, раскладывают в емкости сверху на подготовленный грунт или торф (лучше положить снег, а уже на него – семена) закрыть пленкой. Получившийся парник ставится на холод на 14 дней. После этого перенести в теплое место (+23 +25˚С). Подкармливать можно после появления 2 пар настоящих листиков опрыскиванием стимуляторами (1-2 капли препарата на 1 литр воды) через каждые 10-12 дней или удобрениями.
2. Рассада высаживается в конце марта начале апреля, в зависимости от погодных условий. В посуду или цветочные горшки на дно насыпается керамзит или крупнозернистый песок для дренажа. Сверху насыпается слегка увлажненная почвенная смесь, которая уплотняется и на поверхности раскладываются семена. По верху горшка насыпается небольшой слой снега, закрывается стеклом или пленкой. Выбирается в саду самое затемненное место (где дольше всего не тает снег) и оставить там горшок на 1-2 недели. После этого их можно поместить в холодный парник или занести домой и ждать всходов. При таком способе семена лучше всходят и растения меньше болеют, а при пикировке большая часть остается жизнеспособной.
Выбирается в саду самое затемненное место (где дольше всего не тает снег) и оставить там горшок на 1-2 недели. После этого их можно поместить в холодный парник или занести домой и ждать всходов. При таком способе семена лучше всходят и растения меньше болеют, а при пикировке большая часть остается жизнеспособной.
Вообще посев на снег имеет несколько преимуществ: 1) посев не загущается, так как на белом хорошо видны темные семена, что исключает черную ножку; 2) когда снег тает, он тянет за собой семена, а поскольку грунт слегка утрамбован, они затягиваются не слишком глубоко, присыпать землей не надо.
Если укрывается пленкой, то делаются мелкие отверстия для проветривания и испарения конденсата.
Самое главное – не перелить при поливе и не пересушить. Излишек воды вреден для корней они могут загнить, поэтому поливать в зависимости от состояния почвы.
Пересушивание почвы также может повредить корням, так как они маленькие и пересыхание , даже на короткое время, отрицательно сказывается на растениях.
В первое время растения растут очень медленно. В фазе 1-2 настоящих листьев, примерно через 30-40 дней после посева сеянцы пикируются очень осторожно. После пикировки температура поддерживается в пределах +15 +24˚С, каждые 10-12 дней производится подкормка комплексными удобрениями для ягодных культур: «Ортон-ягодное», «Агровита Для клубники и земляники», «Мультифлор АКВА 250мл для ягодных культур» — водорастворимое удобрение.
Примерно за 2 недели до высадки рассаду закаливают, уменьшают полив, а непосредственно перед посадкой опрыскивают раствором «Эпин-Экстра» и после высадки для лучшей приживаемости.
Опасности изменения климата для двудомных видов растений
Хейлбут, Дж. К. Меньшее видовое богатство двудомных ветвей. утра. Нат . 156 , 221–241 (2000).
ПабМед
Google ученый
Маталлама Г.
 , Вендт Т., Араужо Д. С. Д. и Скарано Ф. Р. Большое количество двудомных растений в тропической прибрежной растительности. утра. Дж. Бот . 92 , 1513–1519 (2005).
, Вендт Т., Араужо Д. С. Д. и Скарано Ф. Р. Большое количество двудомных растений в тропической прибрежной растительности. утра. Дж. Бот . 92 , 1513–1519 (2005).Google ученый
Геринг, К.А. и Уитхэм, Т.Г. Уменьшение микоризы на Juniperus monosperma с омелой: влияние экологического стресса и пола дерева на растительного паразита и растительно-грибковый мутуализм. Oecologia 89 , 298–303 (1992).
ПабМед
Google ученый
Whitham, T. G. et al. Структура генетики сообществ и экосистем: от генов к экосистемам. Nature Rev. Gen . 7 , 510–523 (2006).
КАС
Google ученый
Холм, Дж. А., Миллер, С. Дж. и Кроппер-младший, В. П. Динамика популяции двудомной амазонской пальмы Mauritia flexuosa : анализ моделирования устойчивого сбора урожая.
Biotropica 40 , 550–558 (2008 г.).Google ученый
Куаракса, К., Эллиот, С. и Хоссарт-Макки, М. Фенология двудомных Ficus spp. Виды деревьев и их значение для лесовосстановления. Для. Экол. Управление . 265 , 82–93 (2012).
Google ученый
Петри, В. К. и др. Механизмы, лежащие в основе полового диморфизма растений в политрофных сообществах членистоногих. Экология 94 , 2055–2065 (2013).
ПабМед
Google ученый
Дарвин, К. Различные формы цветов на растениях одного и того же вида (Джон Мюррей, 1877).
Google ученый
Charlesworth, D. Определение пола растений и половые хромосомы. Наследственность 88 , 94–101 (2002).
ПабМед
Google ученый
Доусон, Т.Е. и Гебер, Массачусетс в Пол и диморфизм цветковых растений (под редакцией Гебер, Массачусетс, Доусон, Т.Е. и Делф, Л.Ф.) 175–215 (Springer, 1999).
Google ученый
Обесо, Дж. Р. Затраты на воспроизводство растений. Новый Фитол . 155 , 321–348 (2002).
Google ученый
Пармезан, К. Экологические и эволюционные реакции на недавнее изменение климата. Энн. Преподобный Экол. Эвол. Сис . 37 , 637–669 (2006).
Google ученый
Урбан, штат Мэриленд. Ускорение риска вымирания из-за изменения климата. Наука 348 , 571–573 (2015).
Артикул
КАСGoogle ученый
Bierzychudek, P.
& Eckhart, V. Пространственное разделение полов двудомных растений. утра. Нат . 132 , 34–43 (1988).Google ученый
Филд, Д. Л., Пикап, М. и Барретт, К. Х. Сравнительный анализ вариаций соотношения полов у двудомных цветковых растений. Эволюция 67 , 661–672 (2012).
ПабМед
Google ученый
Синклер, Дж. Р., Эмлен, Дж. И Фриман, Д. К. Предвзятое соотношение полов у растений: теория и тенденции. Бот. Версия . 78 , 63–86 (2012).
Google ученый
Барретт С.Ч., Якимовски С.Б., Филд Д.Л. и Пикап М. Экологическая генетика соотношения полов в популяциях растений. Фил. Транс. Р. Соц. Лонд. B 365 , 2549–2557 (2010).
Google ученый
Доусон, Т.
Э. и Элерингер, Дж. Р. Гендерная физиология, различение изотопов углерода и распределение среды обитания у бузины, 9 лет.0005 Клен черный . Экология 74 , 798–815 (1993).Google ученый
Делф, Л.Ф. в Пол и диморфизм цветковых растений (под редакцией Гебера, Массачусетса, Доусона, Т.Е., Делфа, Л.Ф.) 149–174 (Springer, 1999).
Google ученый
Фриман, Д. К., Кликофф, Л. Г. и Харпер, К. Т. Дифференциальное использование ресурсов полами раздельнополых растений. Наука 193 , 597–599 (1976).
КАС
пабмедGoogle ученый
Чарнов, Э. Т. Теория распределения полов (Princeton Univ. Press, 1982).
Google ученый
де Йонг, Т.
Дж., Ван Батенбург, Ф.Х.Д. и Ван Дейк, Дж. Соотношение полов в семенах двудомных растений зависит от относительного рассеивания пыльцы и семян: пример с использованием модели шахматной доски. Дж. Эвол. Биол . 15 , 373–379 (2002).Google ученый
Фриман, Д. К., Дауст, Дж. Л., Эль-Кеблави, А., Милья, К. Дж. и Макартур, Э. Д. Половой отбор и предотвращение инбридинга в эволюции двудомности. Бот. Версия . 63 , 65–92 (1997).
Google ученый
Булл, Дж. Дж. Эволюция соотношения полов при изменении приспособленности. Наследственность 46 , 9–26 (1981).
Google ученый
Булл, Дж. Дж. и Чарнов, Э. Л. Насколько фундаментальны соотношения полов по Фишеру? Оксфорд Сур. Эвол. Биол . 5 , 96–135 (1988).
Google ученый
Булл, Дж. Дж. и Чарнов, Э. Л. Загадочное соотношение полов рептилий. Эволюция 43 , 1561–1566 (1989).
КАС
пабмедGoogle ученый
Frank, S.A. & Swingland, I.R. Соотношение полов при условном выражении пола. Ж. Теор. Биол . 135 , 415–418 (1998).
Google ученый
Чарнов, Э. Л. и Булл, Дж. Дж. Соотношение полов, не связанное с рыбаками, со сменой пола и определением пола в окружающей среде. Природа 338 , 148–150 (1989а).
Google ученый
Чарнов Э. Л. и Булл Дж. Дж. Первичное соотношение полов при определении пола в окружающей среде. Ж. Теор. Биол . 139 , 431–436 (1989b).
КАС
пабмедGoogle ученый
Фридберг, С.
и Уэйд, М. Дж. Борьба мужчин поддерживает соотношение полов, ориентированное на женщин, при определении пола по окружающей среде. Аним. Поведение . 67 , 177–181 (2004).Google ученый
Шоу, Р. Ф. и Молер, Дж. Д. Селективное значение соотношения полов. утра. Нат . 87 , 337–342.
Уэйд, М. Дж., Шустер, С. М. и Демут, Дж. П. Половой отбор благоприятствует предвзятому соотношению полов: баланс между противоборствующими силами отбора по соотношению полов и полового отбора. утра. Нат . 162 , 403–414 (2003).
ПабМед
Google ученый
Филд, С. Б., Рандерсон, Дж. Л. и Мальмстрем, К. М. Глобальная чистая первичная продукция: сочетание экологии с дистанционным зондированием. Рем. Сенатор Окружающая среда . 51 , 74–88 (1995).
Google ученый
Гросс, К.
Л. и Соул, Дж. Д. Различия в распределении биомассы по репродуктивным и вегетативным структурам мужских и женских растений двудомного многолетнего растения, Silene alba (Миллер) Краузе. утра. Дж. Бот . 68 , 801–807 (1981).Google ученый
Хермс, Д. А. и Мэттсон, В. Дж. Дилемма растений: выращивать или защищать. кв. Преподобный Биол . 67 , 283–335 (1992).
Google ученый
Wardlaw, IF Tansley Review № 27. Контроль разделения углерода в растениях. Новый Фитол . 116 , 341–381 (1990).
КАС
Google ученый
Уотсон, М. А. и Каспер, Б. Б. Морфогенетические ограничения моделей распределения углерода в растениях. Энн. Преподобный Экол. Сис . 15 , 233–258 (1984).
Google ученый
Макдауэлл, С.
К.Л., Макдауэлл, Н.Г., Маршалл, Дж.Д. и Халтин, К. Распределение углерода и азота при размножении самцов и самок пихты Дугласа Скалистых гор ( Pseudotsuga menziesii вар. gluaca , Pinaceae). утра. Дж. Бот . 87 , 539–546 (2000).КАС
пабмедGoogle ученый
Доусон, Т. Е. и Блисс, Л. С. Модели использования воды и водные отношения в тканях двудомного кустарника, Salix arctica : физиологическая основа разделения среды обитания между полами. Экология 79 , 332–343 (1989).
КАС
пабмедGoogle ученый
Leigh, A. & Nicotra, A.B. Половой диморфизм, репродуктивное распределение и эффективность использования воды у Maireana pyramata (Chenopodiaceae), двудомного полузасушливого кустарника. Австрал. Дж. Бот . 51 , 509–514 (2003).
Google ученый
Грайм, Дж.
П. Стратегии растений, вегетационные процессы и свойства экосистем (Уайли, 2006).Google ученый
Халтин, К.Р., Буш, С.Е., Уэст, А.Г. и Элерингер, Дж.Р. Структура популяции, физиология и экогидрологическое воздействие раздельнополых видов прибрежных деревьев западной части Северной Америки. Экология 154 , 85–93 (2007).
КАС
пабмедGoogle ученый
Tognetti, R. Адаптация двудомных растений к изменению климата: имеет ли значение гендерный баланс? Дерево Физиол . 32 , 1321–1324 (2012).
ПабМед
Google ученый
Жювани, М. и Мунне-Бош, С. Половые различия в устойчивости к стрессу у двудомных растений: критическая оценка в физиологическом контексте. Дж. Экспл. Бот . 66 , 6083–6092 (2015).
КАС
пабмедGoogle ученый
Халтин, К. Р. и др. Гендерно-специфические модели надземного распределения, проводимости полога и использования воды доминирующими видами прибрежных деревьев. Физиол дерева . 28 , 1383–1394 (2008).
КАС
пабмедGoogle ученый
Walther, G. R. et al. Экологические реакции на недавнее изменение климата. Природа 416 , 389–395 (2002).
КАС
пабмедGoogle ученый
Jump, A. S. & Peñuelas, J. Бег, чтобы остановиться: адаптация и реакция растений на быстрое изменение климата. Экол. Пусть . 8 , 1010–1020 (2005 г.).
Google ученый
Уорд, Дж. К., Доусон, Т.
Е. и Элерингер, Дж. Р. Реакция полов Acer negundo на межгодовые различия в доступности воды, определенные на основе соотношения изотопов углерода в целлюлозе годичных колец. Дерево Физиол . 22 , 339–346 (2002).КАС
пабмедGoogle ученый
Доусон, Т. Е., Уорд, Дж. К. и Элерингер, Дж. Р. Временное масштабирование физиологических реакций от газообмена до годичных колец: гендерное исследование Acer negundo (Boxelder), растущего в разных условиях. Функц. Экол . 18 , 212–222 (2004).
Google ученый
Халтин, К.Р., Буш, С.Э., Уэст, А.Г. и Элерингер, Дж.Р. Влияние пола на транспирацию в масштабе сокодвижения у доминирующих прибрежных пород деревьев: бузины самшитовой ( Acer negundo ). Ж Геофиз. Рез. Биогеоск . 112 , G03S06 (2007).
Google ученый
Wallace, C.
S. & Rundel, P.W. Половой диморфизм и распределение ресурсов мужских и женских кустарников Simondsia chinensis . Oecologia 44 , 34–39 (1979).ПабМед
Google ученый
Дельф, Л. Ф. Модели распределения ресурсов в зависимости от пола в полудвудомных кустарниках Hebe subalpine . Экология 71 , 1342–1351 (1990).
Google ученый
Делф, Л. Ф., Лу, Ю. и Джейн, Л. Д. Модели распределения ресурсов в раздельнополом Осинка (Cyperaceae). утра. Дж. Бот . 80 , 607–615 (1993).
Google ученый
Барретт, С.Ч. и Хаф, Дж. Половой диморфизм у цветковых растений. Дж. Экспл. Бот. 64 , 67–82 (2013).
КАС
пабмедGoogle ученый
Вимп, Г.
М. и Уитэм, Т. Г. Последствия хищничества и гибридизации растений-хозяев для биоразнообразия на мутуализме тлей и муравьев. Экология 82 , 440–452 (2001).Google ученый
Allen, C.D. Глобальный обзор смертности деревьев, вызванной засухой и жарой, выявил возникающие риски изменения климата для лесов. Для. Экол. Управление . 259 , 660–684 (2010).
Google ученый
Williams, A. P. et al. Реакция лесов на усиление засушливости и тепла на юго-западе США. Проц. Натл акад. науч. США 107 , 21289–21294 (2010 г.).
КАС
пабмедGoogle ученый
Williams, A. P. et al. Температура как мощный фактор регионального стресса и гибели лесов от засухи. Природа Клим. Изменение 3 , 292–297 (2013).
Google ученый
McDowell, N.G. и др. Многомасштабные прогнозы массовой гибели хвойных деревьев из-за хронического повышения температуры. Природа Клим. Смена 6 , 295–300 (2016).
Google ученый
McDowell, N.G. и др. Механизмы выживания и гибели растений при засухе: почему одни растения выживают, а другие погибают от засухи? Новый Фитол . 178 , 719–739 (2008).
ПабМед
Google ученый
McDowell, N.G. et al. Взаимозависимость механизмов, лежащих в основе обусловленной климатом смертности растительности. Тренды Экол. Эвол. 26 , 523–532 (2011).
ПабМед
Google ученый
Xu, X. Половые реакции Populus cathayana на засуху и повышенные температуры.
Окружающая среда растительных клеток . 31 , 850–860 (2008).КАС
пабмедGoogle ученый
Бродрибб, Т. Дж. и Холбрук, Н. М. Снижение гидравлической эффективности по мере высыхания испаряющихся листьев: два типа реакции. Окружающая среда растительных клеток . 29 , 2205–2215 (2006).
КАС
пабмедGoogle ученый
Звинецки, М. А. и Холбрук, Н. М. Противостояние демону Максвелла: биофизика восстановления ксилемы. Trends Plant Sci . 14 , 530–534 (2009).
КАС
пабмедGoogle ученый
Вилагроса, А. и др. Скоординированы ли устойчивость симпласта к сильным засушливым условиям и уязвимость ксилемы к кавитации? Комплексный анализ фотосинтетических, гидравлических и листовых процессов у двух средиземноморских засухоустойчивых видов.
Окружающая среда. Эксп. Бот . 69 , 233–242 (2010).Google ученый
Буччи, С. Дж., Шольц, Ф. Г., Гольдштейн, Г., Мейнцер, Ф. К. и Штернберг, Л. Д. С. Л. Динамические изменения гидравлической проводимости в черешках двух видов деревьев саванны, факторы и механизмы, способствующие повторному наполнению эмболизированных сосудов. Окружающая среда растительных клеток . 26 , 1633–1645 (2003).
Google ученый
Саллео, С., Трифило, П., Эспозито, С., Нардини, А. и ЛоГулло, М. Превращение крахмала в сахар в паренхиме древесины в полеводстве Лавр благородный растений: компонент сигнального пути для восстановления эмболии. Функц. Растение Биол . 36 , 815–825 (2009).
Google ученый
Якобсен, А. Л., Эслер, К.
Дж., Пратт, Р. Б. и Эверс, Ф. В. Устойчивость кустарников к водному стрессу в регионах со средиземноморским климатом: конвергенция сообществ финбос и суккулентных кару с калифорнийскими кустарниковыми сообществами. утра. Дж. Бот . 96 , 1445–1453 (2009).ПабМед
Google ученый
Martinez-Vilalta, J., Piñol, J. & Beven, K. Гидравлическая модель для прогнозирования гибели древесных растений, вызванной засухой: приложение к изменению климата в Средиземноморье. Экол. Мод . 155 , 127–147 (2002).
Google ученый
Плаут, Дж. А. и др. Гидравлические пределы, предшествующие гибели в сосново-можжевеловом лесу в условиях экспериментальной засухи. Окружающая среда клеток растений . 35 , 1601–1617 (2012).
ПабМед
Google ученый
Плаут, Дж.
А. и др. Пониженная реакция транспирации на импульсы осадков предшествует гибели в сосново-можжевеловых лесах, подверженных продолжительной засухе. Новый Фитол . 200 , 375–387 (2013).ПабМед
Google ученый
Андерегг В.Р.Л., Андерегг Л.Д.Л., Берри Дж.А. и Филд С.Б. Потеря гидравлической проводимости всего дерева во время сильной засухи и многолетнего вымирания леса. Экология 175 , 11–23 (2014).
ПабМед
Google ученый
Севанто С., Макдауэлл Н. Г., Дикман Т. Л., Пэнгл Р. и Покман В. Т. Как умирают деревья? Проверка гипотезы гидравлического отказа и углеродного голодания. Окружающая среда растительных клеток . 37 , 153–161 (2014).
КАС
пабмедGoogle ученый
Фишер, Р. А.
Генетическая теория естественного отбора (Oxford Univ. Press, 1930).Google ученый
Лоари, С. Р. Скорость изменения климата. Природа 462 , 1052–1055 (2011).
Google ученый
Сандел Б. и др. Влияние скорости позднечетвертичного изменения климата на эндемизм видов. Наука 334 , 660–664 (2011).
КАС
пабмедGoogle ученый
Corlett, T. & Westcott, D. A. Будет ли перемещение растений поспевать за изменением климата. Тренды Экол. Эвол . 28 , 482–488 (2013).
ПабМед
Google ученый
Джонс, М. Х., Макдональд, С. Э. и Генри, Г. Х. Р. Реакция высокой арктической ивы на экспериментальное изменение климата в зависимости от пола и среды обитания.
Oikos 87 , 129–138 (1999).Google ученый
Wang, X. & Griffin, K.L. Половая физиологическая реакция и реакция роста на повышенное содержание CO2 в атмосфере у Silene latifolia Poiret. Глобальное изменение биол . 9 , 612–618 (2003).
Google ученый
Zhao, H., Xu, X., Zhang, Y., Korpelainen, H. & Li, C. Осаждение азота по-разному ограничивает фотосинтетическую реакцию на повышенное содержание CO2 у раздельнополых видов. Экология 165 , 41–54 (2011).
ПабМед
Google ученый
Wang, X. Размножение и потомство Silene latifolia (Caryophyllaceae) в зависимости от концентрации CO2 в атмосфере. амер. Дж. Бот . 92 , 826–832 (2005).
Google ученый
Монтесинос, Д.
, Де Луис, М., Верду, М., Равентос, Х. и Гарсия-Файос, П. Когда, как и сколько: гендерно-специфические стратегии использования ресурсов в раздельнополом дереве Можжевельник турифера . Энн. Бот . 98 , 885–889 (2006).КАС
пабмед
ПабМед ЦентральныйGoogle ученый
Montesinos, D., Villar-Salvador, P., García-Fayos, P. & Verdú, M. Полы в Juniperus thurifera по-разному реагируют на изменения в доступности питательных веществ. Новый Фитол . 193 , 705–712 (2011).
ПабМед
Google ученый
Руд, С. Б. и др. Гидрологические связи между колебаниями климата, речным стоком, растительностью и древесиной Δ 13 C мужских и женских тополей. Окружающая среда растительных клеток . 36 , 984–993 (2013).
КАС
пабмедGoogle ученый
Браун, Т.
Б. и др. Использование фенокамер для наблюдения за нашей изменяющейся Землей: к глобальной сети фенокамов. Фронт. Экол. Окружающая среда . 14 , 84–93 (2016).Google ученый
Кавеки, Т. Дж. и Эберт, Д. Концептуальные вопросы адаптации к местным условиям. Экол. Пусть . 7 , 1225–1241 (2004).
Google ученый
Grady, K.C. et al. Генетическая изменчивость продуктивности основных прибрежных видов на границе их распространения: последствия для восстановления и вспомогательной миграции в условиях потепления климата. Глобальное изменение биол . 17 , 3724–3735 (2011).
Google ученый
Грейди, К. С., Колб, Т. Э., Икеда, Д. Х. и Уитэм, Т. Г. Слишком далекий мост: холод и патогенные ограничения для генетически обусловленной миграции прибрежных лесов.
Рез. Экол . 23 , 811–820 (2015).Google ученый
Халтин, К.Р., Бертч, К.Г. и Элерингер, Дж.Р. Гендерные особенности поглощения углерода и использования воды у доминирующих прибрежных пород деревьев, подвергающихся воздействию потепления климата. Глобальное изменение биол . 19 , 3390–3405 (2013).
Google ученый
Эппли, С. М., Мерсер, К. А., Хаанинг, К. и Грейвс, К. Б. Половые различия во взаимодействии между Distichlis spicata (Poaceae) и микоризными грибами. утра. Дж. Бот . 96 , 1967–1973 (2009).
ПабМед
Google ученый
Varga, S. & Kytöviita, MM Гендерный диморфизм и микоризный симбиоз влияют на посетителей цветков и репродуктивную способность Geranium sylvaticum . Функц.
Экол . 24 , 750–758 (2010).Google ученый
Лаворель, С. и Гарнье, Э. Прогнозирование изменений в составе сообщества и функционировании экосистемы на основе признаков растений: новый взгляд на Святой Грааль. Функц. Экол . 16 , 545–556 (2002).
Google ученый
Харт, С. К., ДеЛука, Т. Х., Ньюман, Г. С., Маккензи, М. Д. и Бойл, С. И. Послепожарная вегетативная динамика как движущая сила структуры и функционирования микробного сообщества в лесных почвах. Для. Экол. Управление . 220 , 166–184 (2005).
Google ученый
Verdú, M. & García-Fayos, P. Плодоядные птицы опосредуют половую стимуляцию двудомного растения-кормилицы. Дж. Вег. наука . 14 , 35–42 (2003).
Google ученый
Графф, П.
, Розитано, Ф. и Агиар, М. Р. Изменения в соотношении полов двудомной травы с интенсивностью выпаса: взаимодействие между гендерными признаками, взаимодействием с соседями и пространственными паттернами. Дж. Экол . 101 , 1146–1157 (2013).Google ученый
Огрен, Дж. Данелл, К. и Элмквист, Т. в Пол и диморфизм цветковых растений (ред. Гебер, Массачусетс, Доусон, Т.Е., Делф, Л.Ф.) 217–246 (Springer, 1999).
Google ученый
Корнелиссен, Т. и Стайлинг, П. Травоядные с пристрастием к полу: метаанализ влияния пола на взаимодействие растений и травоядных. Oikos 111 , 488–500 (2005).
Google ученый
Эшман, Т. Л. Избирательность опылителей и ее значение для эволюции раздельнополого и полового диморфизма. Экология .
81 , 2577–2591 (2000).Google ученый
Вулф, Л. М. Дифференциальное травоядное растение и образование галлов у самцов и самок Neea психотроиды , двудомного дерева. Biotropica 29 , 169–174 (1997).
Google ученый
 , Вендт Т., Араужо Д. С. Д. и Скарано Ф. Р. Большое количество двудомных растений в тропической прибрежной растительности. утра. Дж. Бот . 92 , 1513–1519 (2005).
, Вендт Т., Араужо Д. С. Д. и Скарано Ф. Р. Большое количество двудомных растений в тропической прибрежной растительности. утра. Дж. Бот . 92 , 1513–1519 (2005).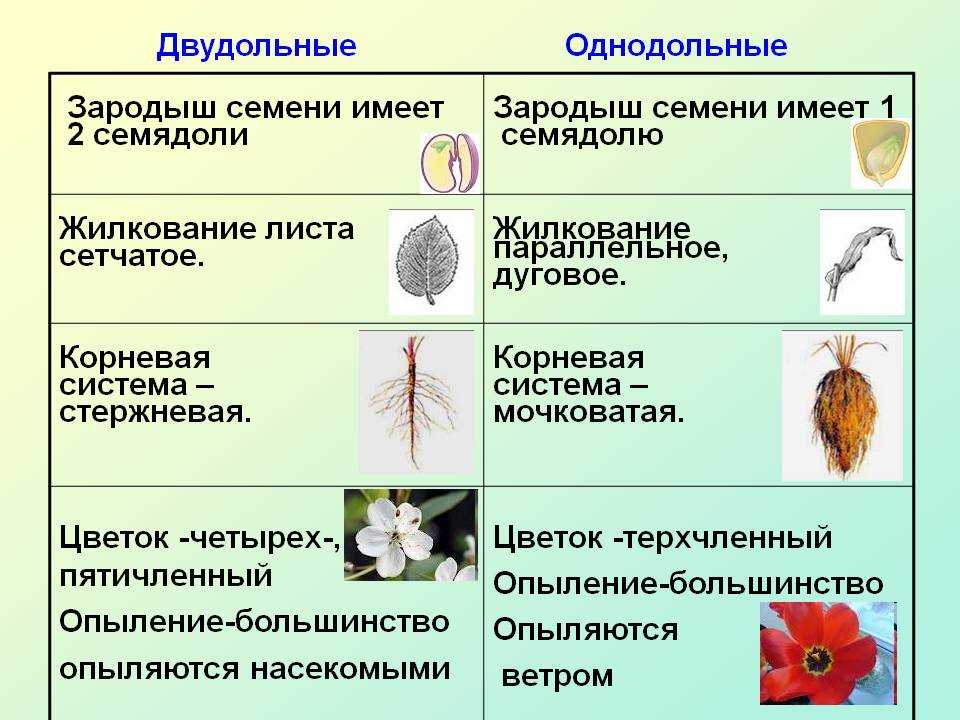 Biotropica 40 , 550–558 (2008 г.).
Biotropica 40 , 550–558 (2008 г.).
 & Eckhart, V. Пространственное разделение полов двудомных растений. утра. Нат . 132 , 34–43 (1988).
& Eckhart, V. Пространственное разделение полов двудомных растений. утра. Нат . 132 , 34–43 (1988). Э. и Элерингер, Дж. Р. Гендерная физиология, различение изотопов углерода и распределение среды обитания у бузины, 9 лет.0005 Клен черный . Экология 74 , 798–815 (1993).
Э. и Элерингер, Дж. Р. Гендерная физиология, различение изотопов углерода и распределение среды обитания у бузины, 9 лет.0005 Клен черный . Экология 74 , 798–815 (1993). Дж., Ван Батенбург, Ф.Х.Д. и Ван Дейк, Дж. Соотношение полов в семенах двудомных растений зависит от относительного рассеивания пыльцы и семян: пример с использованием модели шахматной доски. Дж. Эвол. Биол . 15 , 373–379 (2002).
Дж., Ван Батенбург, Ф.Х.Д. и Ван Дейк, Дж. Соотношение полов в семенах двудомных растений зависит от относительного рассеивания пыльцы и семян: пример с использованием модели шахматной доски. Дж. Эвол. Биол . 15 , 373–379 (2002).
 и Уэйд, М. Дж. Борьба мужчин поддерживает соотношение полов, ориентированное на женщин, при определении пола по окружающей среде. Аним. Поведение . 67 , 177–181 (2004).
и Уэйд, М. Дж. Борьба мужчин поддерживает соотношение полов, ориентированное на женщин, при определении пола по окружающей среде. Аним. Поведение . 67 , 177–181 (2004). Л. и Соул, Дж. Д. Различия в распределении биомассы по репродуктивным и вегетативным структурам мужских и женских растений двудомного многолетнего растения, Silene alba (Миллер) Краузе. утра. Дж. Бот . 68 , 801–807 (1981).
Л. и Соул, Дж. Д. Различия в распределении биомассы по репродуктивным и вегетативным структурам мужских и женских растений двудомного многолетнего растения, Silene alba (Миллер) Краузе. утра. Дж. Бот . 68 , 801–807 (1981).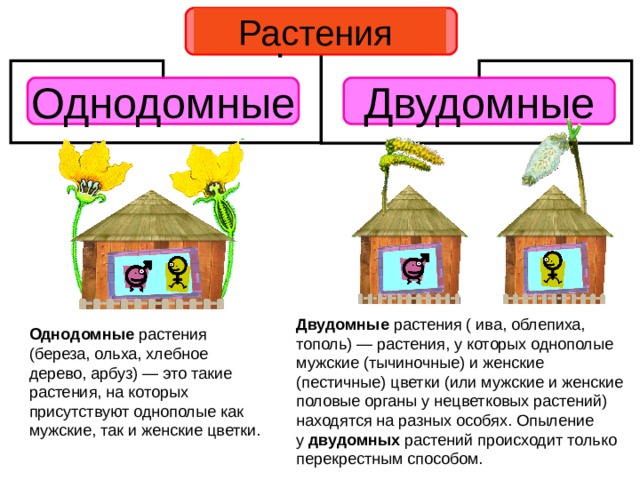 К.Л., Макдауэлл, Н.Г., Маршалл, Дж.Д. и Халтин, К. Распределение углерода и азота при размножении самцов и самок пихты Дугласа Скалистых гор ( Pseudotsuga menziesii вар. gluaca , Pinaceae). утра. Дж. Бот . 87 , 539–546 (2000).
К.Л., Макдауэлл, Н.Г., Маршалл, Дж.Д. и Халтин, К. Распределение углерода и азота при размножении самцов и самок пихты Дугласа Скалистых гор ( Pseudotsuga menziesii вар. gluaca , Pinaceae). утра. Дж. Бот . 87 , 539–546 (2000).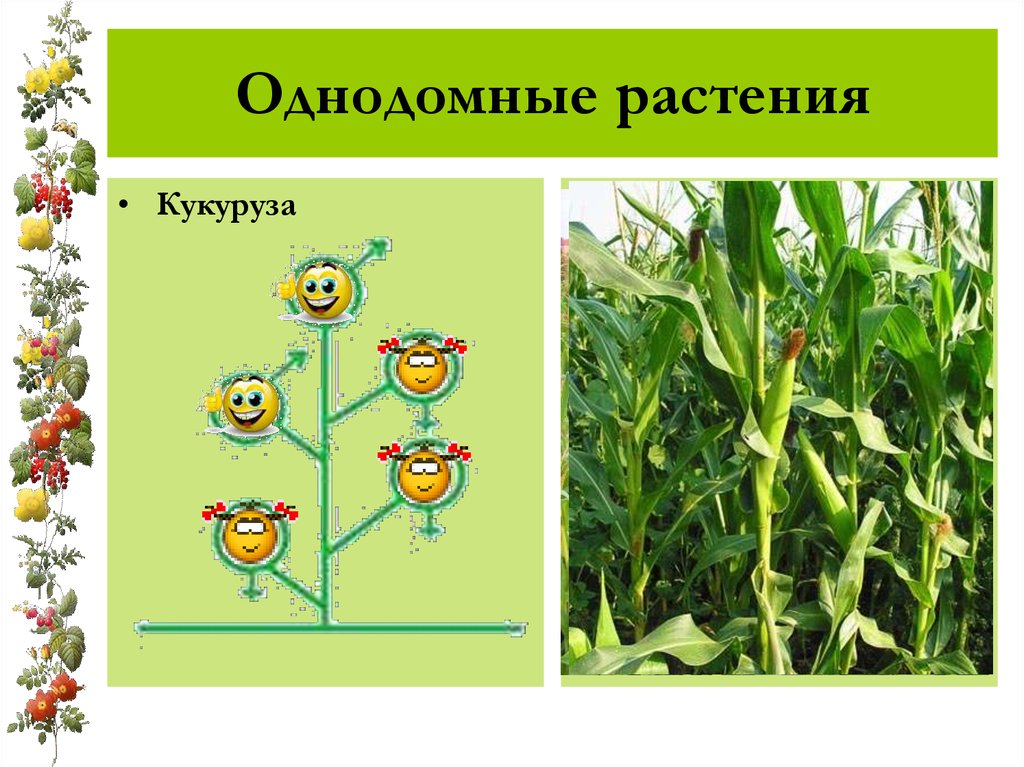 П. Стратегии растений, вегетационные процессы и свойства экосистем (Уайли, 2006).
П. Стратегии растений, вегетационные процессы и свойства экосистем (Уайли, 2006).
 Е. и Элерингер, Дж. Р. Реакция полов Acer negundo на межгодовые различия в доступности воды, определенные на основе соотношения изотопов углерода в целлюлозе годичных колец. Дерево Физиол . 22 , 339–346 (2002).
Е. и Элерингер, Дж. Р. Реакция полов Acer negundo на межгодовые различия в доступности воды, определенные на основе соотношения изотопов углерода в целлюлозе годичных колец. Дерево Физиол . 22 , 339–346 (2002). S. & Rundel, P.W. Половой диморфизм и распределение ресурсов мужских и женских кустарников Simondsia chinensis . Oecologia 44 , 34–39 (1979).
S. & Rundel, P.W. Половой диморфизм и распределение ресурсов мужских и женских кустарников Simondsia chinensis . Oecologia 44 , 34–39 (1979). М. и Уитэм, Т. Г. Последствия хищничества и гибридизации растений-хозяев для биоразнообразия на мутуализме тлей и муравьев. Экология 82 , 440–452 (2001).
М. и Уитэм, Т. Г. Последствия хищничества и гибридизации растений-хозяев для биоразнообразия на мутуализме тлей и муравьев. Экология 82 , 440–452 (2001).
 Окружающая среда растительных клеток . 31 , 850–860 (2008).
Окружающая среда растительных клеток . 31 , 850–860 (2008). Окружающая среда. Эксп. Бот . 69 , 233–242 (2010).
Окружающая среда. Эксп. Бот . 69 , 233–242 (2010). Дж., Пратт, Р. Б. и Эверс, Ф. В. Устойчивость кустарников к водному стрессу в регионах со средиземноморским климатом: конвергенция сообществ финбос и суккулентных кару с калифорнийскими кустарниковыми сообществами. утра. Дж. Бот . 96 , 1445–1453 (2009).
Дж., Пратт, Р. Б. и Эверс, Ф. В. Устойчивость кустарников к водному стрессу в регионах со средиземноморским климатом: конвергенция сообществ финбос и суккулентных кару с калифорнийскими кустарниковыми сообществами. утра. Дж. Бот . 96 , 1445–1453 (2009). А. и др. Пониженная реакция транспирации на импульсы осадков предшествует гибели в сосново-можжевеловых лесах, подверженных продолжительной засухе. Новый Фитол . 200 , 375–387 (2013).
А. и др. Пониженная реакция транспирации на импульсы осадков предшествует гибели в сосново-можжевеловых лесах, подверженных продолжительной засухе. Новый Фитол . 200 , 375–387 (2013).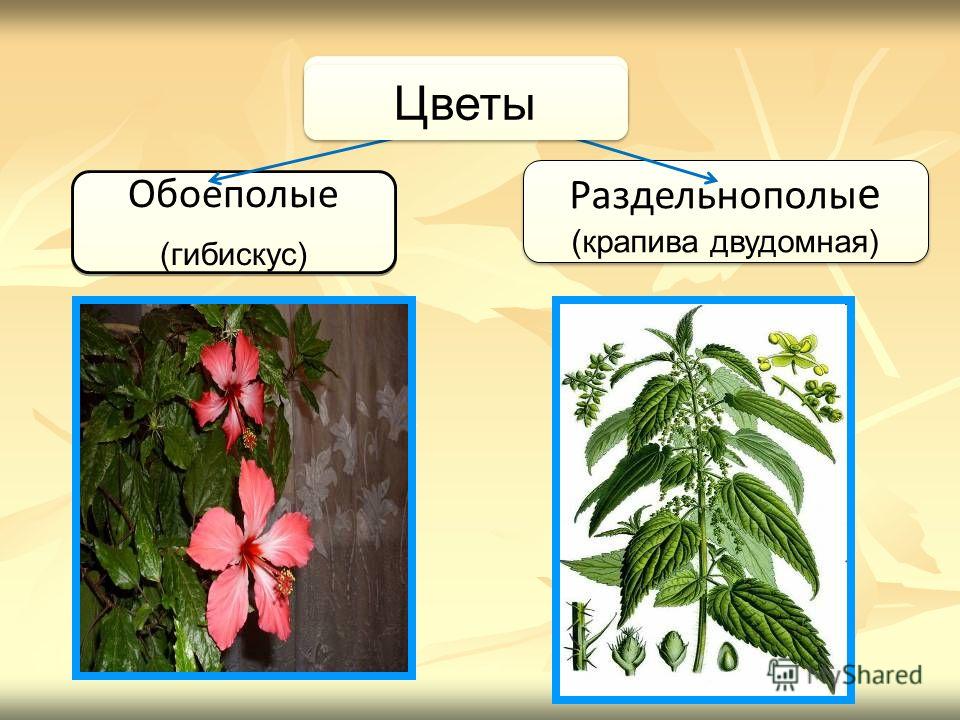 Генетическая теория естественного отбора (Oxford Univ. Press, 1930).
Генетическая теория естественного отбора (Oxford Univ. Press, 1930). Oikos 87 , 129–138 (1999).
Oikos 87 , 129–138 (1999). , Де Луис, М., Верду, М., Равентос, Х. и Гарсия-Файос, П. Когда, как и сколько: гендерно-специфические стратегии использования ресурсов в раздельнополом дереве Можжевельник турифера . Энн. Бот . 98 , 885–889 (2006).
, Де Луис, М., Верду, М., Равентос, Х. и Гарсия-Файос, П. Когда, как и сколько: гендерно-специфические стратегии использования ресурсов в раздельнополом дереве Можжевельник турифера . Энн. Бот . 98 , 885–889 (2006). Б. и др. Использование фенокамер для наблюдения за нашей изменяющейся Землей: к глобальной сети фенокамов. Фронт. Экол. Окружающая среда . 14 , 84–93 (2016).
Б. и др. Использование фенокамер для наблюдения за нашей изменяющейся Землей: к глобальной сети фенокамов. Фронт. Экол. Окружающая среда . 14 , 84–93 (2016). Рез. Экол . 23 , 811–820 (2015).
Рез. Экол . 23 , 811–820 (2015). Экол . 24 , 750–758 (2010).
Экол . 24 , 750–758 (2010).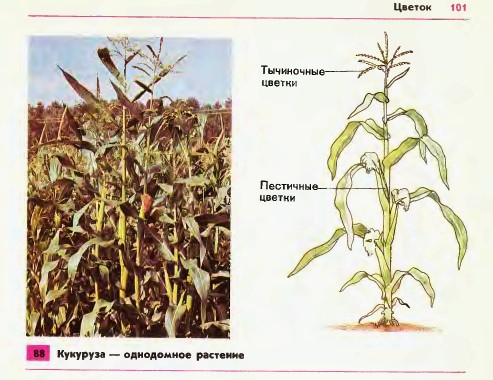 , Розитано, Ф. и Агиар, М. Р. Изменения в соотношении полов двудомной травы с интенсивностью выпаса: взаимодействие между гендерными признаками, взаимодействием с соседями и пространственными паттернами. Дж. Экол . 101 , 1146–1157 (2013).
, Розитано, Ф. и Агиар, М. Р. Изменения в соотношении полов двудомной травы с интенсивностью выпаса: взаимодействие между гендерными признаками, взаимодействием с соседями и пространственными паттернами. Дж. Экол . 101 , 1146–1157 (2013). 81 , 2577–2591 (2000).
81 , 2577–2591 (2000).Скачать ссылки
Двудомное растение | Энциклопедия.com
oxford
просмотров обновлено 11 июня 2018 г.
раздельнополые Обладающие мужскими и женскими цветками или другими репродуктивными органами на отдельных, однополых, отдельных растениях. См. также АНДРОЭЦИУМ; Сравните ОДНОДОМНЫЕ.
Словарь наук о растениях МАЙКЛ АЛЛАБИ
oxford
просмотров обновлено 08 июня 2018 г.
раздельнополые (бот.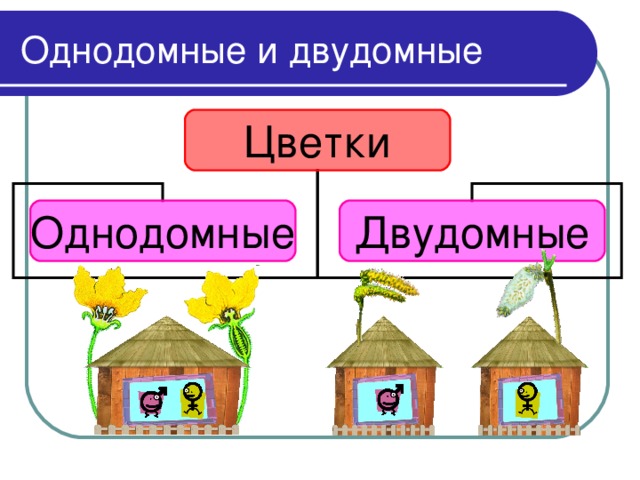 ), имеющие два пола у отдельных особей. XVIII. ф. ДИ- 2 + Гр. ойкос дом; см. -IOUS.
), имеющие два пола у отдельных особей. XVIII. ф. ДИ- 2 + Гр. ойкос дом; см. -IOUS.
Краткий Оксфордский словарь английской этимологии TF HOAD
oxford
просмотров обновлено 21 мая 2018 г.
двудомные растения Примерами двудомных растений являются ивы. Сравните однодомные.
Биологический словарь
Оксфорд
просмотров обновлен 14 мая 2018
двудомные Обладающие мужскими и женскими цветками или другими репродуктивными органами на отдельных однополых отдельных растениях. Сравните однодомные.
Экологический словарь МАЙКЛ АЛЛАБИ
oxford
просмотров обновлено 14 мая 2018
раздельнополые особи Обладающие отдельными половыми и женскими репродуктивными органами. Сравните ОДНОДОМНЫЕ.
Сравните ОДНОДОМНЫЕ.
Зоологический словарь МАЙКЛ АЛЛАБИ
Еще с encyclopedia.com
Однодомные , однодомные
1. Применяется к организму, в котором отдельные мужские и женские органы встречаются у одной и той же особи (например, к растению, имеющему мужские и женские… Androecium , Перейти к основному содержанию
андроцей
андроцей Совокупность тычинок, образующих мужские репродуктивные органы цветкового растения. Они могут принадлежать… Однополый цветок , однополый цветок Цветок, имеющий либо тычинки, либо плодолистики, но не то и другое вместе. Растение может быть однополым (двудомным), иметь только мужские цветки или… Archegonium , Перейти к основному содержанию
архегоний
archegonium (мн. archegonia) Многоклеточный женский половой орган в форме колбы мохообразных, плаунов, хвощей,… Бегонии, бегонии (род Begonia) — привлекательные многолетние травы с мягкими, сочными стеблями и белыми, розовыми, красными, оранжевыми или желтыми цветы. Бегонии мем… Семя , Семя
Семя – это часть цветкового растения, участвующая в размножении. Он состоит из трех основных частей: зародыша, эндосперма и тесты. Эмбрион я…
Он состоит из трех основных частей: зародыша, эндосперма и тесты. Эмбрион я…
Об этой статье
Все источники —
Обновлено август 24 2016 Облизительно Encyclopedia.com Содержание Тема печати
Вы также можете как
. °
Диодор Александрийский
Диодор Крон (р. IV век до н. э.)
Диодор Тарсийский
Diodontidae
Diodes Incorporated
diode-transistor logic
diode laser
Diodati, Giovanni
dioctyl sodium sulphosuccinate
Diocletian, Roman Emperor
Diocletian, Persecution of
Diocletian, Caius Valerianus°
Diocletian window
Dioclesian
diocesan
диок. син.
диок.
дим.
Дион Златоуст°
Дион Златоуст
Дион Кассий°
Dio Cassius
Dinwiddie, Emily (1879–1949)
dioecious plant
dioestrus
Diogenes Laertius (c.