Содержание
Корни боковые
Боковые корни распространяются в сторону на 60— 70 см под острым углом от главного стержневого корня.[ …]
Корни ели чаще всего развиваются в самых поверхностных горизонтах почвы, стержневой корень у нее не развивается. Лишь на рыхлых, достаточно легких и хорошо дренированных почвах ель образует глубоко расположенную корневую систему с массой якорных корней, вертикально ответвляющихся от боковых. Обычно на таких местоположениях ель оказывается вполне ветроустойчивой, в большинстве же случаев она является очень ветровальной породой. Эта последняя особенность ели создает ряд затруднений при ведении хозяйства в ельниках на тяжелых почвах. На болотистых и на умеренно влажных почвах ель, так же как и лиственница, способна образовывать придаточные корни, начиная с 3—4-летнего возраста.[ …]
Корни начинают формироваться уже в зародыше растения. При прорастании семени из зародышевого корешка образуется главный корень. Через некоторое время от него отрастают многочисленные боковые корни. У ряда растений от стеблей и листьев образуются придаточные корни.[ …]
У ряда растений от стеблей и листьев образуются придаточные корни.[ …]
В корнях сосудистая система отделена от коры специализированными тканями — перициклом и эндодермой. Перицикл опоясывает сосуды. Он состоит из паренхимати-ческой ткани и дает начало боковым корням и стеблям, возникающим на корнях. Эндодерма обычно состоит из одного ряда клеток, отделяющих сосуды от коры, и до сих пор не выяснено, является ли она частью сосудистой системы или коры. По-видимому, она выполняет защитные функции. В стебле перицикл и эндодерма обычно отсутствуют.[ …]
У основания боковой почки образуется зачаток корня. Изучение развития почки показывает, что почка и корень происходят от одной материнской клетки, при делении которой возникает более или менее шаровидная масса клеток. В верхней части клеточной массы дифференцируется побег, а в нижней — корень. Зачатки корней у надземных побегов обычно не прорастают, но если побег присыпать землей или погрузить в воду, на его узлах из зачатков корней вырастут корни, хотя некоторые почки останутся спящими. [ …]
[ …]
Горизонтальные, или боковые, корни. Уступают по толщине вертикальным и в почве расположены примерно параллельно ее поверхности. Они содержат много запасных веществ и более обильно усажены адвентивными почками, чем вертикальные. В вегетативном возобновлении боковые корни часто играют большую роль, чем вертикальные, так как благодаря боковым корням от главного корня отводятся на различные расстояния дочерние растения.[ …]
Большую часть стелы в корне занимает сложный проводящий пучок с двумя-тремя и более радиально вытянутыми ксилемными участками, чередующимися с участками флоэмы. В месте прикрепления главного корня к стеблю обычно имеется до восьми протоксилемных тяжей. Ближе к концу (верхушке) корни становятся диархными, т. е. имеющими два ксилемных участка (рис. 167).[ …]
Плодовые тела образуются на корнях дерева или у основания ствола, чаще на нижней стороне боковых корней ели, приподнятых над землей. Заражение происходит базидиоспорами и конидиями, распространяемыми грызунами, насекомыми и ветром. Грибница проникает в древесину через поранения на корнях, реже через поранения корневой шейки и вызывает корневую гниль, переходящую в напенную и иногда стволовую. Очаговое заражение происходит при соприкосновении больных корней со здоровыми. Зараженная древесина становится красноватой или темно-фиолетовой; на ней появляются белые пятна с черными точками или штрихами в центре. В последней стадии гниения древесина делается ячеистой.[ …]
Грибница проникает в древесину через поранения на корнях, реже через поранения корневой шейки и вызывает корневую гниль, переходящую в напенную и иногда стволовую. Очаговое заражение происходит при соприкосновении больных корней со здоровыми. Зараженная древесина становится красноватой или темно-фиолетовой; на ней появляются белые пятна с черными точками или штрихами в центре. В последней стадии гниения древесина делается ячеистой.[ …]
Наибольшая глубина проникновения корней в почву и лучший рост боковых корней наблюдается на площадках, расположенных между волоками и на огнищах. Так, минимальная глубина проникновения корней на огнищах на 36% больше, чем у сеянцев на трелевочном волоке. Объяснить это можно тем, что на волоках уплотненная в резульгате трелевки древесины почва препятствует проникновению корней вглубь.. Основная масса корней здесь располагается в верхнем, разрыхленном слое почвы.[ …]
Верхушечная почка побега тормозит рост боковых почек, а верхушка корня подавляет образование зачатков боковых корней. Оба процесса регулируются гормонами. Транспортируемые вниз от верхушечной почки ауксины, вероятно, оказывают тормозящее действие на рост боковых почек, в то время как поднимающиеся вверх из корня цитойшины стимулируют их рост. Напротив, регуляция образования боковых корней, очевидно, связана со стимуляцией этого процесса ауксином из верхушки корня или из побега и с его торможением образующимися в корневой верхушке цитокининами. Вопрос об участии других гормонов остается спорным.[ …]
Оба процесса регулируются гормонами. Транспортируемые вниз от верхушечной почки ауксины, вероятно, оказывают тормозящее действие на рост боковых почек, в то время как поднимающиеся вверх из корня цитойшины стимулируют их рост. Напротив, регуляция образования боковых корней, очевидно, связана со стимуляцией этого процесса ауксином из верхушки корня или из побега и с его торможением образующимися в корневой верхушке цитокининами. Вопрос об участии других гормонов остается спорным.[ …]
Со стимуляцией образования придаточных корней в принципе сравнима и стимуляция ауксином образования боковых корней, хотя этот вариант не имеет существенного практического значения. В этом случае в глубоко-лежащей ткани возникают меристемные комплексы, из которых потом дифференцируются боковые корни. Образование боковых корней — это тоже коррелятивно управляемый процесс, и контролируется он главным образом кончиком корня, хотя гормональные факторы, видимо, действуют здесь совсем иначе, нежели при развитии боковых почек под влиянием верхушки побега (см.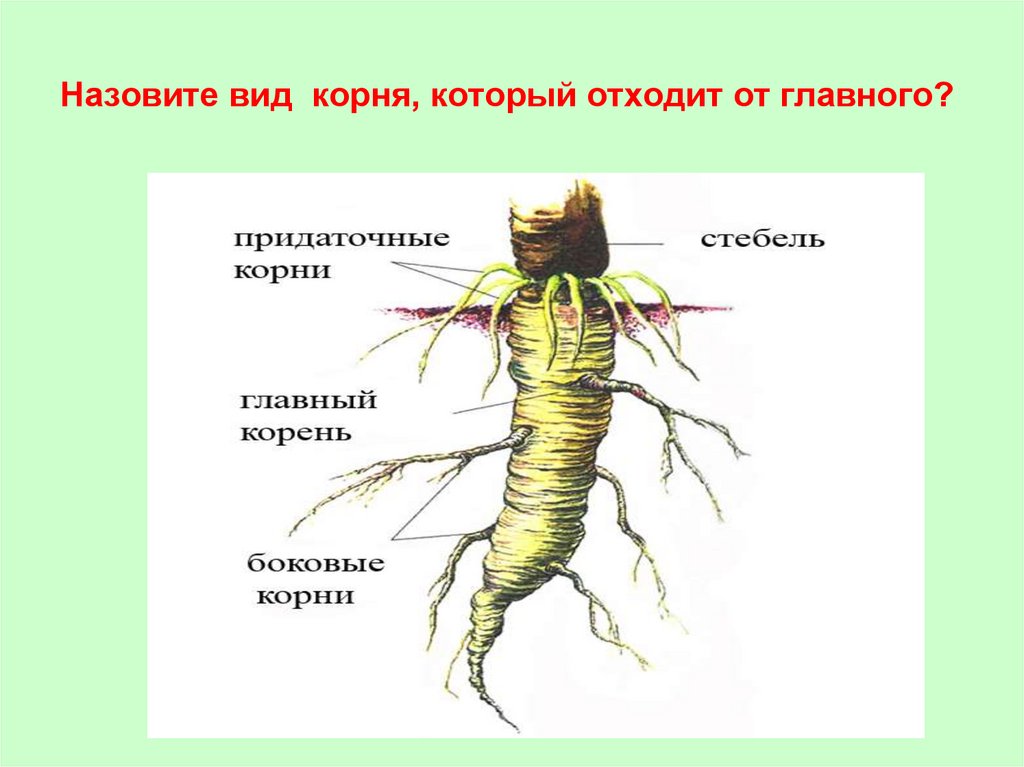 также разд. 14.2).[ …]
также разд. 14.2).[ …]
Корневая система у вьюнка состоит из главного корня, уходящего в почву на глубину до 2,5 м, который дает боковые ответвления и новые побеги.[ …]
Одновременно с отгниванием заболони отгнивают и корни. Через 10—15 лет остаются только стержневой и крупные боковые корни, в результате чего пень сравнительно легко извлекается из .почвы.[ …]
Различие в характере возникновения клубеньков и боковых корней особенно четко наблюдается у сераделлы, поскольку коровая ткань главного корня этого растения — место возникновения первых клубеньков — состоит из относительно небольшого слоя клеток и клубеньки становятся видимыми очень быстро после инфицирования корня бактериями. Они образуют сначала выступы уплощенной формы на корне, что позволяет отличить их от конических выступов боковых корней. Клубеньки отличаются от боковых корней и рядом анатомических признаков: отсутствием центрального цилиндра, корневых чехликов и эпидермиса, наличием значительного слоя коры, покрывающей клубенек. [ …]
[ …]
Корневая система состоит из мощного стержневого и боковых корней, разветвляющихся в верхних слоях почв.[ …]
Цитокинины представляют собой производные адс-нина с боковой цепью различного строения. Они образуются в кончике корня и транспортируются по ксилеме. Цитокинины индуцируют клеточное деление, а в листьях— и растяжение клеток. Еще одним свойством цито-кининов является их способность задерживать старение путем стимуляции процессов биосинтеза. Наряду с процессами распада происходит также образование конъюгатов с сахарами и аминокислотами. Известны такие механизмы действия цитокининов, как активация и подавление ферментов, влияние на свойства мембран, а также на синтез РНК и белка.[ …]
В благоприятных почвенных условиях развивает стержневой и боковые корни, что придает деревьям ветроустойчивость; на вечной мерзлоте или близком уровне грунтовых вод образует поверхностную корневую систему и страдает от ветровала. На моховых болотах лучше, чем лиственница сибирская, способна образовывать придаточные корни по мере роста торфяника и мерзлоты вверх, благодаря чему небольшие и искривленные деревья могут расти до 300—400 лет.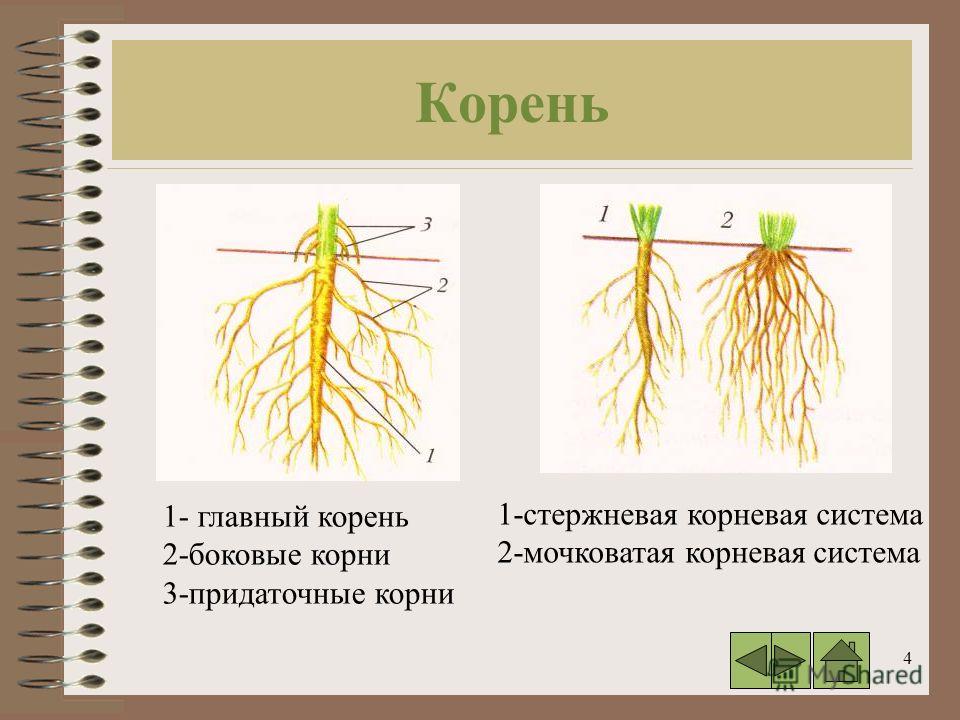 [ …]
[ …]
Корневая система картофеля мочковатая. При посадке клубнями корни образуются в узлах подземной части стеблей. При высеве семенами из зачаточного корешка семени развивается главный корень с многочисленными боковыми.[ …]
Кальций необходим для нормального роста надземных органов и корней растений. Потребность в нем проявляется еще в фазе прорастания. При недостатке кальция и резком преобладании в питательном растворе одновалентных катионов (Н’, Na К ) или катионов Mg нарушается физиологическая уравновешенность раствора и прежде всего страдает корневая система растений. Рост и развитие корней приостанавливаются, они становятся утолщенными, не образуют боковых корешков и корневых волосков, ослизняются и темнеют. Наружные клетки корня, непосредственно соприкасающиеся с таким раствором, разрушаются, клеточные стенки их ослизняются, так как пропитывающие их пектиновые вещества и липоиды в отсутствии кальция растворяются, и содержимое клеток вытекает, ткань превращается в слизистую бесструктурную массу.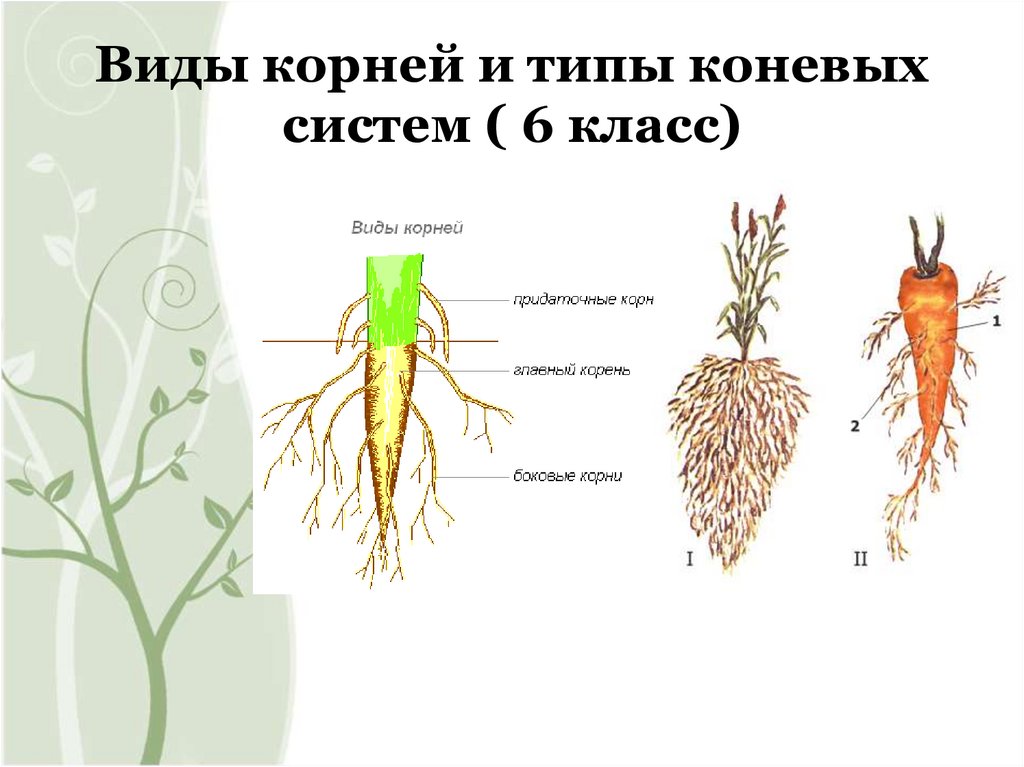 В результате нарушается поглощение растениями питательных веществ.[ …]
В результате нарушается поглощение растениями питательных веществ.[ …]
Приросты образуются как из одиночной верхушечной почки, так и из боковых, расположенных в пазухах листьев. Кроме того, почки могут формироваться на междоузлиях стебля, листьях или корнях часто в результате повреждения. Такие почки называются придаточными.[ …]
Через каждые 5 дней измеряют длину веточки, отмечают образование корней и боковых веточек и измеряют их длину, а у ряски образование лопастей. Дополнительно наблюдают состояние клеток и изменение общего вида растений (побурение, отрыв листочков, разрушение веточки). После 25 дней испытания получаемые результаты по росту веточек и длины корней выражают графически (на оси абсцисс — время в днях, на оси ординат — величины). Снижение роста на 50% по сравнению с контролем отмечаем как условную максимальную безвредную концентрацию. Дополнительные показатели иногда дают весьма (важные сведения о состоянии организма (побурение или исчезновение хлорофилла, начинающееся разрушение растения) и могут несколько коррегировать окончательное суждение о токсичности для высших растений.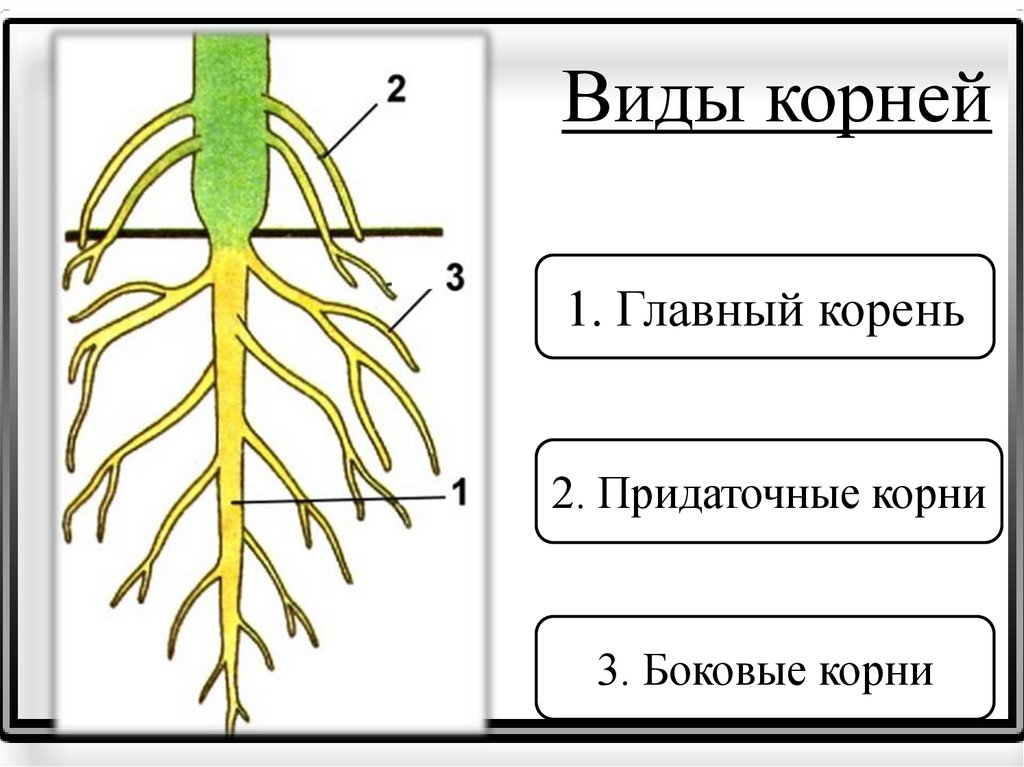 [ …]
[ …]
Мегатроф — требователен к богатству почв зольными веществами. Это является одной из причин его слабого продвижения на север, куда он заходит лишь по современным заливным поймам рек.[ …]
Корневая гниль земляники. Гриб, кроме земляники, поражает овощные культуры. Корни вначале чернеют, на них появляются черные окольцованные пятна. Главный и боковые корни обесцвечиваются и отмирают. Растение легко вынимается из почвы. Гриб распространяется в почве и с посадочным материалом. Зимует возбудитель болезни в почве.[ …]
Корень имеет также несколько пониженную смолистость. В свежих пнях смолистость корней низкая; в спелых пнях она выше. Смолистыми являются не только стержневой, но и крупные боковые корни.[ …]
Корневая система обнаружила еще более резкое различие: общая длина горизонтального и боковых корней у сосны при сильном затенении уменьшилась в 71/2 раз, у ели — почти в 2 раза.[ …]
Общим признаком воздействия динитроанилинов является опухолевое перерождение кончиков корней.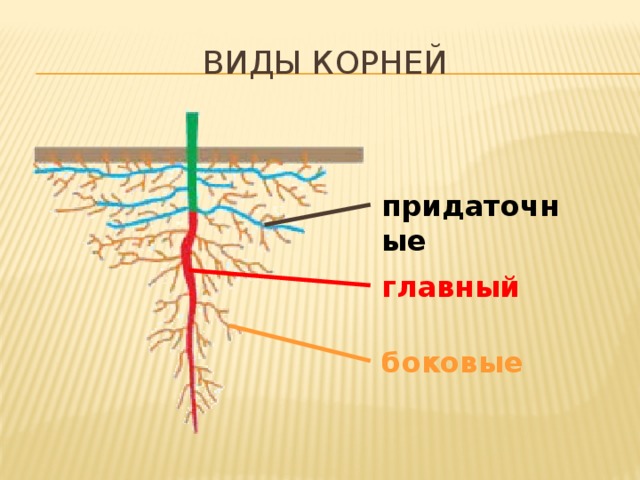 Клетки многоядерные, небольшого размера, в паренхиме коры гипертрофированы, имеют тонкие стенки. Процессы дифференцировки неупорядочены, ксилема чрезмерно утолщается. Динитроанилины подавляют митоз, действуя в тех фазах деления, в которых должны образоваться и функционировать микротрубочки (метафаза, анафаза, телофаза). Волокна веретена состоят из микротрубочек. При нормальном делении микротрубочки перемещают хромосомы, упорядочивая их в метафазе определенным образом, и именно на стадии метафазы динитроанилины нарушают этот процесс. По своему действию они напоминают колхицин, поскольку также препятствуют полимеризации тубулина в микротрубочкн. Однако по точке приложения действия они отличаются от колхицина. Микротрубочки играют определенную роль в переносе веществ, необходимых для строительства клеточной стенки, в размещении ее скелетных элементов.[ …]
Клетки многоядерные, небольшого размера, в паренхиме коры гипертрофированы, имеют тонкие стенки. Процессы дифференцировки неупорядочены, ксилема чрезмерно утолщается. Динитроанилины подавляют митоз, действуя в тех фазах деления, в которых должны образоваться и функционировать микротрубочки (метафаза, анафаза, телофаза). Волокна веретена состоят из микротрубочек. При нормальном делении микротрубочки перемещают хромосомы, упорядочивая их в метафазе определенным образом, и именно на стадии метафазы динитроанилины нарушают этот процесс. По своему действию они напоминают колхицин, поскольку также препятствуют полимеризации тубулина в микротрубочкн. Однако по точке приложения действия они отличаются от колхицина. Микротрубочки играют определенную роль в переносе веществ, необходимых для строительства клеточной стенки, в размещении ее скелетных элементов.[ …]
Главной частью оси над землей является стебель, или ствол; он постепенно делится на сучья, ветви и боковые побеги (ветки), расположение которых часто является характерным для данной породы. Часть вертикальной оси находится в земле; подобно наземной части она разветвляется и образует корневую систему. Ствол обеспечивает механическую опору для кроны, служит проводящей системой и иногда может накопить довольно значительный запас питательных веществ. Корни закрепляют дерево в почве и поддерживают его в вертикальном положении, они всасывают воду и минеральные вещества из почвы посредством корневых волосков, или микоризы, а также транспортируют и накопляют эти вещества. Как корни, так и ствол имеют радиальную симметрию, т. е. различные части расположены вокруг общего центра.[ …]
Часть вертикальной оси находится в земле; подобно наземной части она разветвляется и образует корневую систему. Ствол обеспечивает механическую опору для кроны, служит проводящей системой и иногда может накопить довольно значительный запас питательных веществ. Корни закрепляют дерево в почве и поддерживают его в вертикальном положении, они всасывают воду и минеральные вещества из почвы посредством корневых волосков, или микоризы, а также транспортируют и накопляют эти вещества. Как корни, так и ствол имеют радиальную симметрию, т. е. различные части расположены вокруг общего центра.[ …]
К 15 июня (т. е. через 30 дней) пригнутые прошлогодние лозы в местах отрастания от отведенных боковых побегов дали корни длиной от /2 вершка до 2 вершков, толстые, белые, еще не ветвистые, № 44 цветет, но без пестиков. Дожди продолжаются ежедневно, очень тепло.[ …]
Растения этой группы имеют укороченный главный корень или вообще не имеют стержневого корня и развивают мочку боковых корней. К ним относятся лютик едкий, подорожник большой.[ …]
К ним относятся лютик едкий, подорожник большой.[ …]
Размножается семенами и вегетативно. Образовавшееся из семени растение в первый год образует розетку листьев и корни с придаточными почками. На следующий год из почек развиваются многочисленные плодоносящие побеги. В вегетативном возобновлении решающее значение принадлежит боковым утолщенным корням с придаточными почками. При обработке почвы они легко дробятся на многочисленные кусочки, которые даже при длине 1—3 см быстро приживаются. Образовавшиеся из отрезков растения уже с фазы 4—5 листьев формируют способную к вегетативному возобновлению новую корневую систему. К осени такое растение плодоносит, общая длина корней размножения увеличивается до 4—5 м, от них отрастает множество дочерних розеток.[ …]
Примечательной особенностью всех саговниковых являются растущие вверх над землей и дихотомически ветвящиеся коралловидные корни — кораллоиды. Они возникают как разветвления боковых корней эндогенно из многорядного перицикла напротив лучей первичной ксилемы.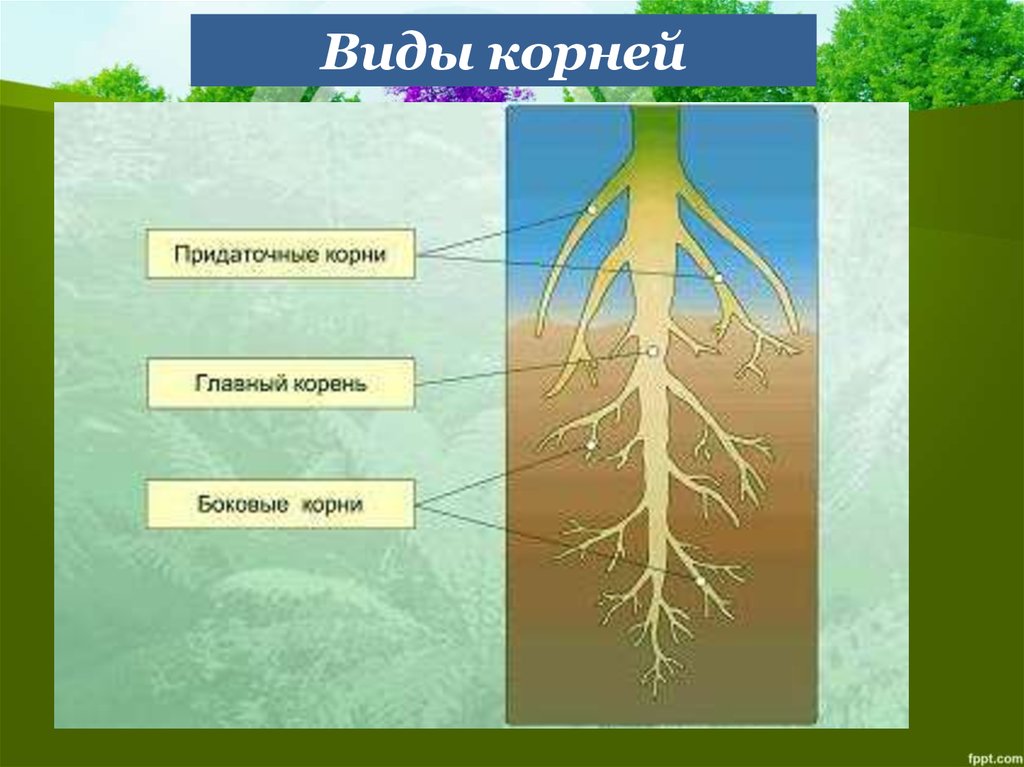 Благодаря интенсивному дихотомическому ветвлению коротких и тонких боковых корней образуются целые грозди клубеньков, окружающие ствол у его основания и напоминающие внешне кораллы. Сначала считали, что это происходит под влиянием бактерий, проникающих в клетки коры корней. В последнее время высказывается предположение, что бактерии, как и сине-зеленые водоросли, являются вторичными поселенцами в сформировавшихся уже клубеньках, а сам клубенек представляет разрастание несущего его корня, вызванное эндофитным грибом, мицелий которого обильно заполняет межклетники в коровой паренхиме этого корня.[ …]
Благодаря интенсивному дихотомическому ветвлению коротких и тонких боковых корней образуются целые грозди клубеньков, окружающие ствол у его основания и напоминающие внешне кораллы. Сначала считали, что это происходит под влиянием бактерий, проникающих в клетки коры корней. В последнее время высказывается предположение, что бактерии, как и сине-зеленые водоросли, являются вторичными поселенцами в сформировавшихся уже клубеньках, а сам клубенек представляет разрастание несущего его корня, вызванное эндофитным грибом, мицелий которого обильно заполняет межклетники в коровой паренхиме этого корня.[ …]
Макрофиты — -водные фотосинтезирующие растения, плавающие на поверхности воды или погруженные в ее толщу. Плавающие растения не имеют корней и держатся на поверхности воды. К наиболее распространенным плавающим растениям относится ряска, маленькое растение с тремя листьями, имеющее диаметр 5 мм. Другое распространенное растение данного типа — водяной гиацинт. Все или большинство лиственных погруженных в толщу воды растений растут под поверхностью воды.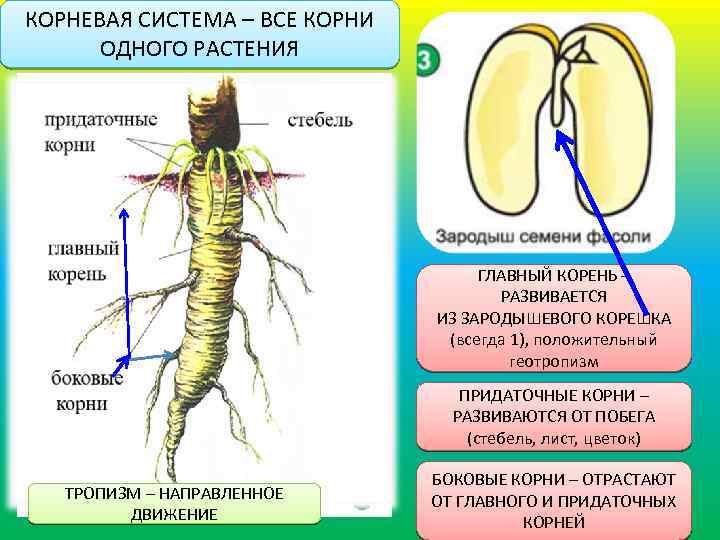 Они могут в зависимости от чистоты воды иметь корни на глубине более 3 м. Погруженные растения закрепляются корнями в донном иле, а их лиственная часть располагается над поверхностью воды. Озера с каменистым и гравийным дном и небольшим количеством питательных веществ в воде не являются благоприятными для роста водных растений, тогда как в эвтрофицированных озерах, в мелких заводях и вдоль береговых линий они растут в изобилии. Слив сточных вод в озера и водоемы может стимулировать рост растений при других благоприятных условиях, таких, как достаточно высокая температура и наличие солнечного света. В биологических прудах сдерживают рост водных растений, устраивая достаточно крутые боковые стенки и сохраняя глубину воды не менее 1 м, чтобы предотвратить проникание солнечных лучей на дно.[ …]
Они могут в зависимости от чистоты воды иметь корни на глубине более 3 м. Погруженные растения закрепляются корнями в донном иле, а их лиственная часть располагается над поверхностью воды. Озера с каменистым и гравийным дном и небольшим количеством питательных веществ в воде не являются благоприятными для роста водных растений, тогда как в эвтрофицированных озерах, в мелких заводях и вдоль береговых линий они растут в изобилии. Слив сточных вод в озера и водоемы может стимулировать рост растений при других благоприятных условиях, таких, как достаточно высокая температура и наличие солнечного света. В биологических прудах сдерживают рост водных растений, устраивая достаточно крутые боковые стенки и сохраняя глубину воды не менее 1 м, чтобы предотвратить проникание солнечных лучей на дно.[ …]
Так, средняя высота 3-летних сеянцев сосны на 4-летней вырубке колеблется от 10 до 12,5 см, а на 7-летней — от 8 до 10,8 см. На 7-летней вырубке меньшую длину стержневого корня имеет и корневая система.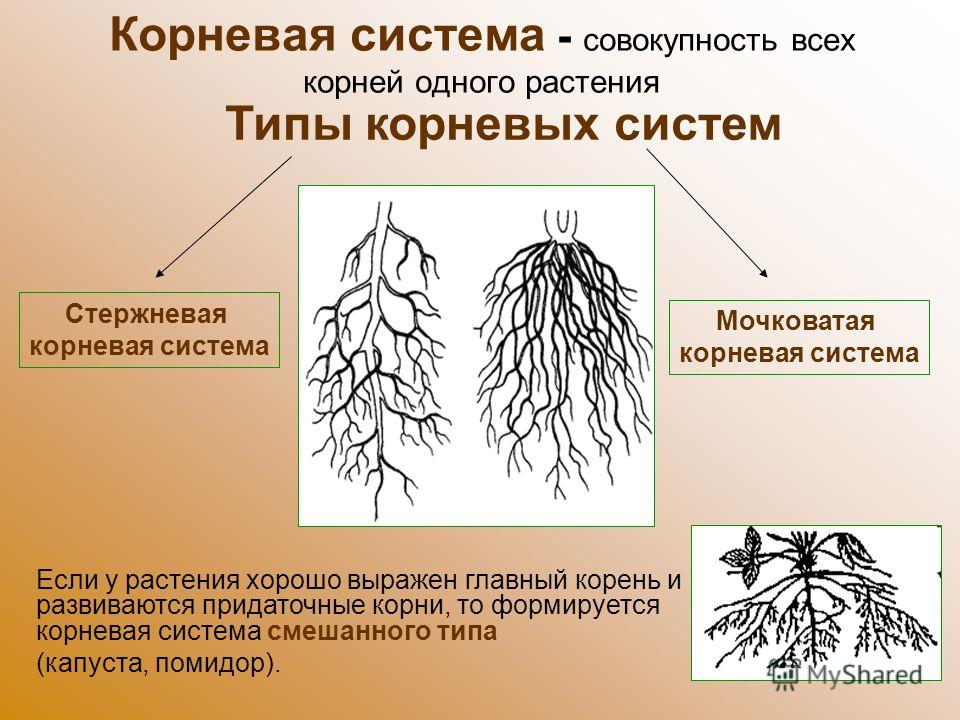 Если на 4-летней вырубке глубина проникновения корней сеянцев сосны колеблется от 8 до 13 см, то на 7-летней — от 7 до 11,5 см. Примерно такое же отличие наблюдается и в длине боковых корней.[ …]
Если на 4-летней вырубке глубина проникновения корней сеянцев сосны колеблется от 8 до 13 см, то на 7-летней — от 7 до 11,5 см. Примерно такое же отличие наблюдается и в длине боковых корней.[ …]
Это дерево высотой до 20 м при диаметре ствола до 80 см, с характерной поверхностной корневой системой. На длинных боковых корнях возникают почки, из которых развиваются корневые отпрыски (поодиночке или пучками). В дальнейшем они могут вырастать в нормальные деревья. Древесина у дакридиума Коленсо крепкая, эластичная, стойкая, желтовато-белого цвета (торговое название «серебряная сосна»). Используется как строительный и поделочный материал.[ …]
Корневая система довольно сильно варьирует в зависимости от свойств субстрата. На глубоких и дренированных отложениях развивает мощную корневую систему с сильно развитым стержневым корнем и боковыми корнями, на каменистых мерзлотных и избыточно увлажненных почвах корневая система поверхностная. В таких условиях лиственница сибирская подвержена ветровалу. На моховых болотах может образовывать придаточные корни выше корневой шейки, благодаря чему способна существовать до 200-300 лет в виде искривленного небольшого деревца в условиях роста торфяника и поднимающейся за ним вечной мерзлоты.[ …]
На моховых болотах может образовывать придаточные корни выше корневой шейки, благодаря чему способна существовать до 200-300 лет в виде искривленного небольшого деревца в условиях роста торфяника и поднимающейся за ним вечной мерзлоты.[ …]
Особенности питания хлопчатника. Корневая система хлопчатника довольно быстро развивается сразу же после всходов и через 5—6 дней достигает в длину 12—15 см. К этому времени обычно начинается образование боковых корешков; через две недели после появления всходов хлопчатника корни достигают глубины 40—50 см. В период цветения и плодообразования боковые корни хлопчатника в верхнем 10-сантиметровом слое почвы отмирают вследствие сухости этого горизонта. Зона наиболее активного поглощения у корневой системы к началу цветения хлопчатника на сероземных почвах, расположенных на повышенных элементах рельефа, находится довольно глубоко, и поэтому корнями усваиваются только питательные вещества удобрений, заделанных на глубину не мельче 18—20 см.[ …]
Сосна имеет пластичную и мощную корневую систему и приспосабливается к жизни при различных режимах водности.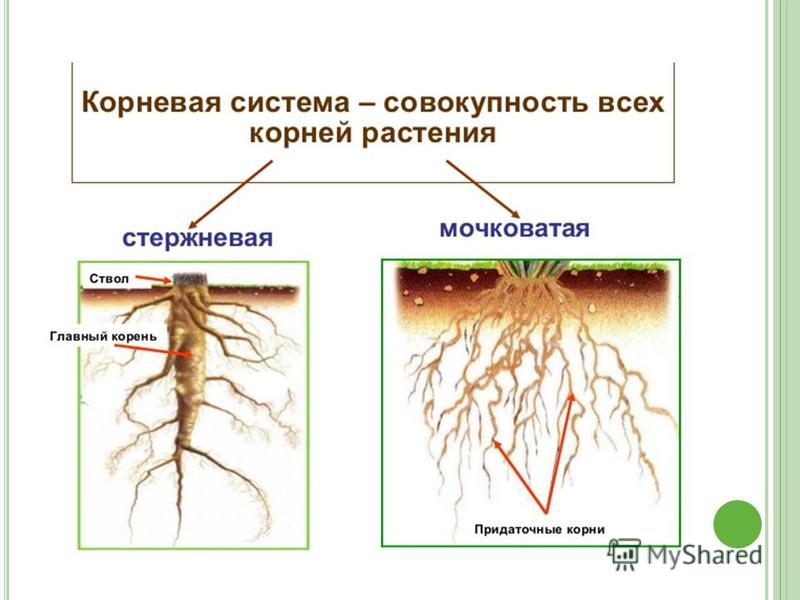 На глубоких сухих песках вершин дюн (гигротоп I) корневая система поверхностная с коротким стержневым корнем и сильно разветвленными боковыми корнями, приспособленными к перехватыванию атмосферных осадков. На свежих дренированных почвах развивается мощная корневая система с глубоким стержневым корнем — редькой и боковыми корнями, которые достигают зеркала грунтовых вод.[ …]
На глубоких сухих песках вершин дюн (гигротоп I) корневая система поверхностная с коротким стержневым корнем и сильно разветвленными боковыми корнями, приспособленными к перехватыванию атмосферных осадков. На свежих дренированных почвах развивается мощная корневая система с глубоким стержневым корнем — редькой и боковыми корнями, которые достигают зеркала грунтовых вод.[ …]
Ювенильные деревья ф обычно уже не имеют семядолей, но обладают детскими (инфантильными) структурами. Первичный побег (стволик) неветвящийся; листья или хвоя ювенильной формы; корневая система состоит из первичного корня и небольшого числа боковых корней. Проростки и ювенильные особи входят в состав травяно-кустарничкового яруса и характеризуются высокой теневыносливостью.[ …]
В работе [53] были проведены расчеты параметров тела, гравитационный эффект от которого позволил бы наилучшим образом согласовать наблюденные и расчетные гравитационные аномалии в осевой зоне хребта. Для моделирования “габброидного корня” было использовано клинообразное, сужающееся книзу тело, наибольший дефицит плотности в котором приурочен к оси, а к периферии тела плотность увеличивается по линейному закону, приближаясь на границе к плотности окружающих пород.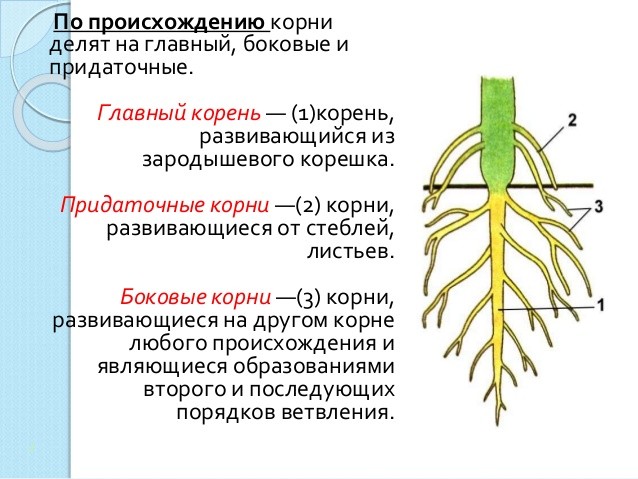 На рис. 7.5, а, где приведены наблюденные гравитационные аномалии, точками показаны рассчитанные значения Ag с учетом термического строения, серпентинизации и эффекта “габброидного корня”. При расчете использовались следующие параметры тела: глубина верхней границы 8 км, ширина верхнего сечения 106 км, глубина распространения 40 км, плотность на оси -0,09 г/см3. Сравнение расчетных аномалий с наблюденными показывает хорошее совпадение в осевой зоне вплоть до боковых максимумов.[ …]
На рис. 7.5, а, где приведены наблюденные гравитационные аномалии, точками показаны рассчитанные значения Ag с учетом термического строения, серпентинизации и эффекта “габброидного корня”. При расчете использовались следующие параметры тела: глубина верхней границы 8 км, ширина верхнего сечения 106 км, глубина распространения 40 км, плотность на оси -0,09 г/см3. Сравнение расчетных аномалий с наблюденными показывает хорошее совпадение в осевой зоне вплоть до боковых максимумов.[ …]
На 7-летней вырубке на волоках при посеве вразброс высота 3-летних сеянцев сосны на 2—2,5 см меньше, тем на огнищах н минерализованных площадках. У 3-летних сеянцев сосны, расположенных на волоках, менее развита и корневая система. Так, при луночном посеве у сеянцев сосны корни на огнищах и минерализованных площадках проникают в почву на 3,5—4 см глубже, чем на волоках. Тоже можно сказать » о распространении боковых корней.[ …]
Молочай лозный — многолетний сорняк семейства молочайных. Распространен почти повсеместно и засоряет все культуры, паровые поля, луга и т..jpg) д. В основном размножается вегетативно корневой порослью. Главный вертикальный корень уходит в почву до 4 м, от него на глубине 5—25 см образуются боковые корни, на которых закладываются почки. Почки на боковых корнях и на корневой шейке главного корня могут сохранять жизнеспособность до 8 лет. Небольшой отрезок корня способен быстро отрастать и давать новое растение.[ …]
д. В основном размножается вегетативно корневой порослью. Главный вертикальный корень уходит в почву до 4 м, от него на глубине 5—25 см образуются боковые корни, на которых закладываются почки. Почки на боковых корнях и на корневой шейке главного корня могут сохранять жизнеспособность до 8 лет. Небольшой отрезок корня способен быстро отрастать и давать новое растение.[ …]
По сравнению с ортогеотропической реакцией другие геотропические реакции изучены значительно меньше. Сюда относится диагеотропизм, т. е. тенденция к горизонтальному росту (например, у корневищ), и плагиогео-тропизм, т. е. стремление расти косо вверх или вниз, как это, например, можно наблюдать у боковых корней и боковых ветвей. Участие гормонов в таких формах движе-, ния весьма вероятно, но этот вопрос еще недостаточно исследован.[ …]
От R. quercina страдают молодые дубы. Гриб поражает их корпи, почему и называется «дубовым корнегубителем». Болезнь эта наиболее распространяется на северо-западе ФРГ в сырые, дождливые годы. Молодые (однолетние и трехлетние) дубки увядают, сохнут и гибнут. Отмирание начинается с верхушки стеблей. На корнях увядающего растения можно разглядеть черные шарики величиной с булавочную головку — склероции. Кроме того, местами на корне заметны еще нежные, нитевидные, ветвящиеся тяжи, состоящие из слабо сросшихся гиф. Они протягиваются в почве от одного корня к другому и служат для распространения гриба. Если дубки со склероциями посажены летом в сырую почву, то склероции дают начало паутинистому мицелию, который распространяется по поверхности почвы. Гифы, достигнув корней, обвивают их, проникают внутрь до самой сердцевины и довольно быстро убивают дерево. В живых паренхимных клетках коры гифы разрастаются, заполняя их полость плотным сплетением. Старые части корня защищены от проникновения мицелия пробковой тканью, но в случае повреждения (особенно при отрывании боковых корешков) они могут также поражаться. Летом на внешнем мицелии, образовавшемся из склероция, развиваются конидиеносцы с конидиями.
Молодые (однолетние и трехлетние) дубки увядают, сохнут и гибнут. Отмирание начинается с верхушки стеблей. На корнях увядающего растения можно разглядеть черные шарики величиной с булавочную головку — склероции. Кроме того, местами на корне заметны еще нежные, нитевидные, ветвящиеся тяжи, состоящие из слабо сросшихся гиф. Они протягиваются в почве от одного корня к другому и служат для распространения гриба. Если дубки со склероциями посажены летом в сырую почву, то склероции дают начало паутинистому мицелию, который распространяется по поверхности почвы. Гифы, достигнув корней, обвивают их, проникают внутрь до самой сердцевины и довольно быстро убивают дерево. В живых паренхимных клетках коры гифы разрастаются, заполняя их полость плотным сплетением. Старые части корня защищены от проникновения мицелия пробковой тканью, но в случае повреждения (особенно при отрывании боковых корешков) они могут также поражаться. Летом на внешнем мицелии, образовавшемся из склероция, развиваются конидиеносцы с конидиями. На таком же мицелии развиваются перитеции в виде черных точек. Аскоспоры прорастают только на следующий год, тогда как конидии прорастают тем же летом и обычно разносятся мышами.[ …]
На таком же мицелии развиваются перитеции в виде черных точек. Аскоспоры прорастают только на следующий год, тогда как конидии прорастают тем же летом и обычно разносятся мышами.[ …]
В. Schischk. et Serg. — развивается как полупаразит-ное растение (рис. 75). Стебель слабоволосистый, часто с буро-фиолетовыми черточками, в верхней части иногда ветвящийся, высотой 15—40 см. Листья яйцевидно-ланцетные, городчато-зубчатые, супротивные. Венчик желтый, с изогнутой трубкой. Зубцы верхней губы венчика фиолетовые. На боковых корнях имеются сосочкообразные выросты (гаустории), которыми присасывается к корням ближайшего растения-хозяина. Цветки крупные, собраны в колос, которым заканчивается стебель, обычно простой, с 6—9 междоузлиями.[ …]
Осот полевой — злостный сорняк семейства сложноцветных. Засоряет все культуры. Распространяется как семянками-летучками (одно растение образует до 20 тыс. семянок), так и корневой порослью. В отличие от бодяка полевого и других корнеотпрысковых сорняков, у осота основной стержневой корень не углубляется в почву более чем на 50 см.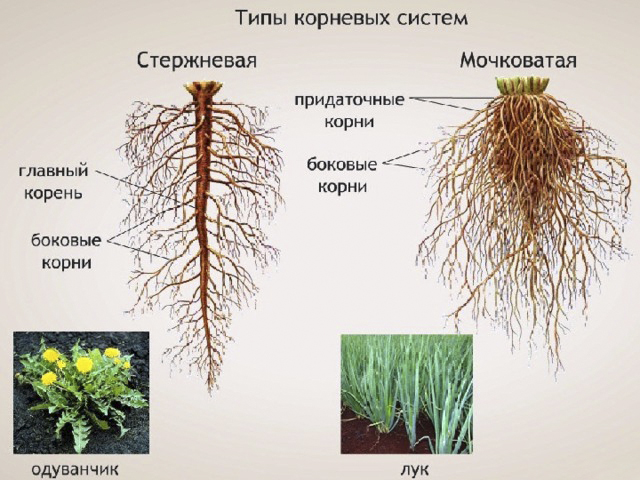 На глубине 6—18 см от вертикального корня образуются боковые корни размножения, которые достигают длины 1,5 м, и на них образуется много почек. Корни осота полевого хрупки, содержат до 40—60% углеводов, обломки корней очень хорошо приживаются.[ …]
На глубине 6—18 см от вертикального корня образуются боковые корни размножения, которые достигают длины 1,5 м, и на них образуется много почек. Корни осота полевого хрупки, содержат до 40—60% углеводов, обломки корней очень хорошо приживаются.[ …]
Рост и развитие растений подвержены регуляции со стороны фитогормонов (регуляторов роста растений), которые представляют собой сигнальные молекулы и которыми являются ауксины, гиббереллины, цитокинины, абсцизовая кислота и этилен. Перечисленные соединения либо синтезируются в клетках, либо транспортируются к клеткам-мишеням. Наибольший эффект этих соединений проявляется при их сочетанном действии. Например, ауксин, индолилуксусная кислота, стимулирует образование корней, но в сочетании с гиббереллином содействует росту корней в длину, а в сочетании с цитокинином — стимулирует закладку и рост боковых почек.[ …]
А. многолетняя — Ambrosia psilostachya D. С.— карантинный корнеотпрысковый сорняк (рис. 71). Растение сизовато-зеленое от густого опушения.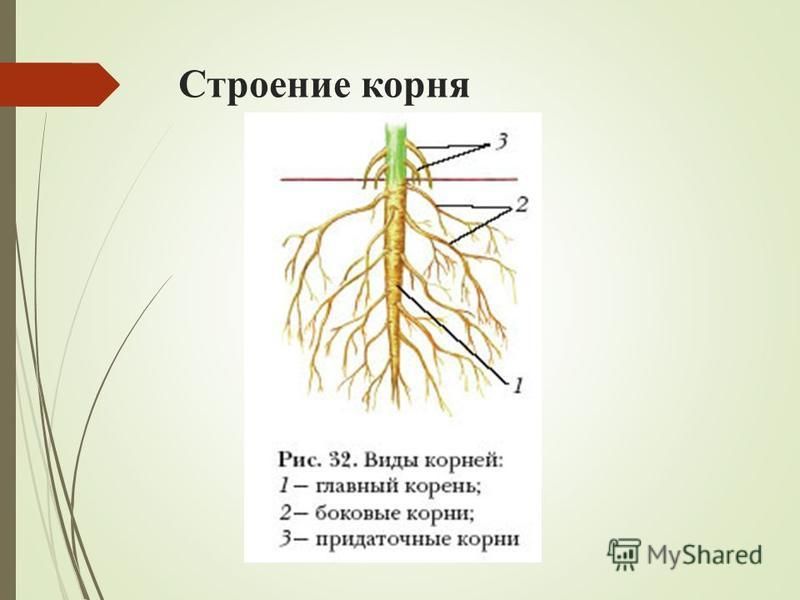 Стебель прямой, бороздчатый, ветвистый, высотой 50—150 см. Листья черешковые, перистонадрезанные. Однодомное растение. Цветки в корзинках, собранных в рыхлые кисти. Почти все цветки мужские, а одиночные женские расположены у основания соцветия. Морфологически сходно с амброзией полыннолистной. Корневая система состоит из главного вертикально углубляющегося в почву корня и боковых корней, расположенных на глубине до 15— 20 см. Горизонтальные корни, усаженные выводковыми почками, расходятся в стороны от вертикального корня и сильно ветвятся.[ …]
Стебель прямой, бороздчатый, ветвистый, высотой 50—150 см. Листья черешковые, перистонадрезанные. Однодомное растение. Цветки в корзинках, собранных в рыхлые кисти. Почти все цветки мужские, а одиночные женские расположены у основания соцветия. Морфологически сходно с амброзией полыннолистной. Корневая система состоит из главного вертикально углубляющегося в почву корня и боковых корней, расположенных на глубине до 15— 20 см. Горизонтальные корни, усаженные выводковыми почками, расходятся в стороны от вертикального корня и сильно ветвятся.[ …]
Виргинильные деревья (х) имеют почти полностью сформированные черты взрослого дерева, но еще не приступили к семяношению. У них хорошо развиты ствол и крона, а прирост в высоту максимальный за весь онтогенез. Величины годичного прироста ствола по длине значительно превышают таковые у крупных ветвей, что определяет удлиненную форму кроны с заостренной вершиной. Диаметр ствола превышает диаметр скелетных ветвей в 3 раза и более. Побеговая система состоит из ветвей 4—7 (8-го) порядков. Корневая система включает главный корень (или его основание), боковые корни разных порядков и придаточные корни. Ствол покрыт перидермой (корка обычно еще не начала развиваться). В начале своего развития виргинильные деревья находятся в ярусе кустарников, в конце — входят в древесный полог. В этом онтогенетическом состоянии у всех деревьев потребности в свете максимальные.[ …]
Корневая система включает главный корень (или его основание), боковые корни разных порядков и придаточные корни. Ствол покрыт перидермой (корка обычно еще не начала развиваться). В начале своего развития виргинильные деревья находятся в ярусе кустарников, в конце — входят в древесный полог. В этом онтогенетическом состоянии у всех деревьев потребности в свете максимальные.[ …]
Корень
Историческое развитие корня
Филогенетически корень возник позже стебля и листа — в связи с переходом растений к жизни на суше и вероятно, произошёл от корнеподобных подземных веточек. У корня нет ни листьев, ни в определённом порядке расположенных почек. Для него характерен верхушечный рост в длину, боковые разветвления его возникают из внутренних тканей, точка роста покрыта корневым чехликом. Корневая система формируется на протяжении всей жизни растительного организма. Иногда корень может служить местом отложения в запас питательных веществ. В таком случае он видоизменяется.
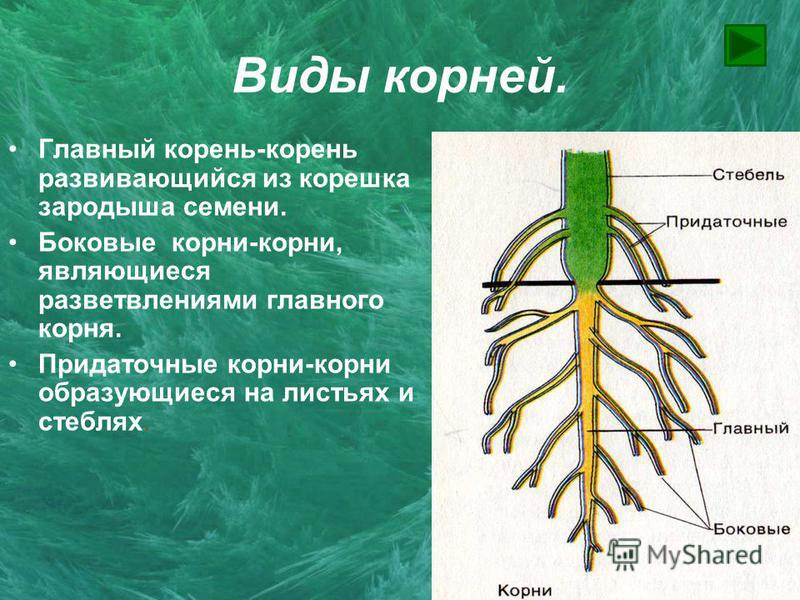
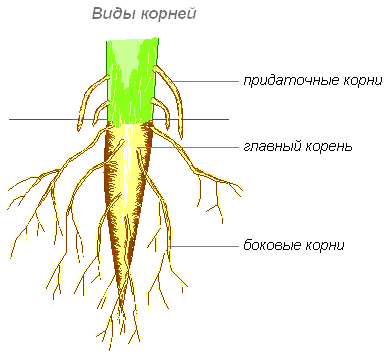
Виды корней
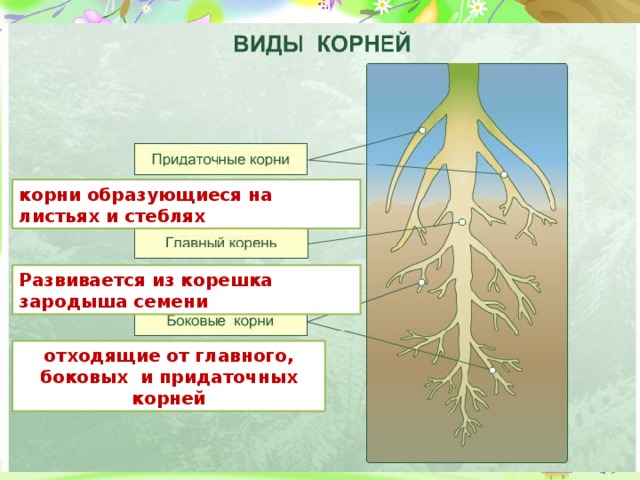
Главный корень образуется из зародышевого корешка при прорастании семени.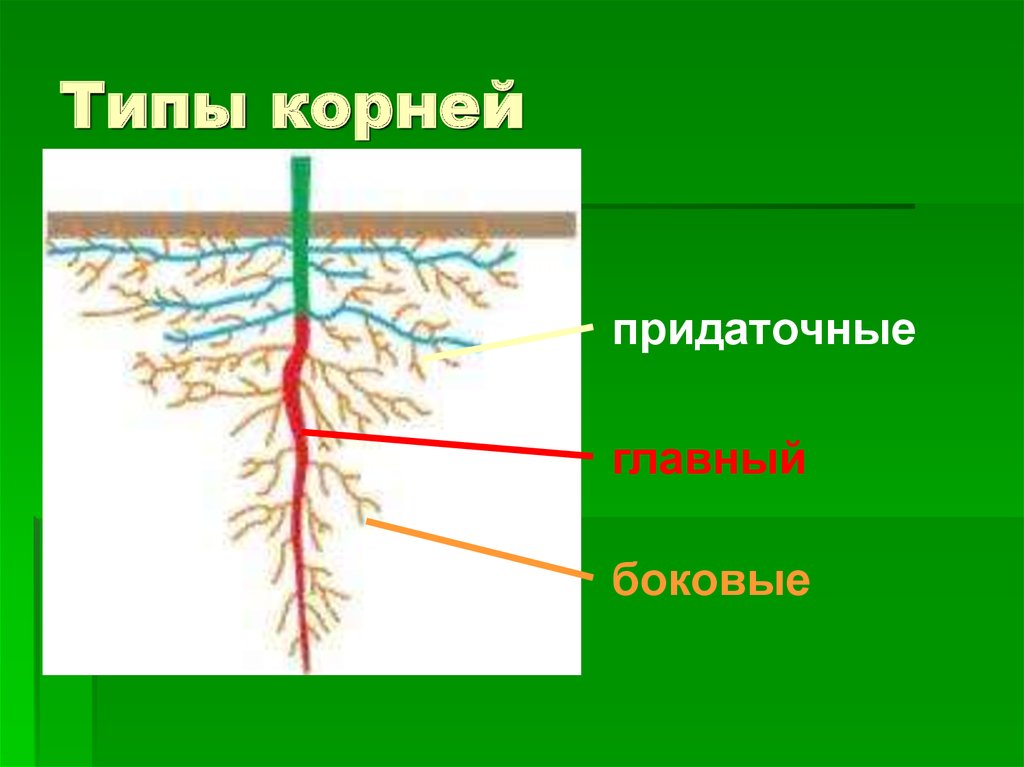 От него отходят боковые корни.
От него отходят боковые корни.
Придаточные корни развиваются на стеблях и листьях.
Боковые корни представляют собой ответвления любых корней.
Каждый корень (главный, боковые, придаточные) обладает способностью к ветвлению, что значительно увеличивает поверхность корневой системы, а это способствует лучшему укреплению растения в почве и улучшению его питания.
Типы корневых систем
Различают два основных типа корневых систем: стержневая, имеющая хорошо развитый главный корень, и мочковатая. Мочковатая корневая система состоит из большого числа придаточных корней, одинаковых по величине. Вся масса корней состоит из боковых или придаточных корешков и имеет вид мочки.
Сильно разветвлённая корневая система образует огромную поглощающую поверхность. Например,
- общая длина корней озимой ржи достигает 600 км;
- длина корневых волосков — 10 000 км;
- общая поверхность корней — 200 м2.
Это во много раз превышает площадь надземной массы.
Если у растения хорошо выражен главный корень и развиваются придаточные корни, то формируется корневая система смешанного типа (капуста, помидор).
Внешнее строение корня. Внутреннее строение корня
Зоны корня
Корневой чехлик
Корень растёт в длину своей верхушкой, где находятся молодые клетки образовательной ткани. Растущая часть покрыта корневым чехликом, защищающим кончик корня от повреждений, и облегчает продвижение корня в почве во время роста. Последняя функция осуществляется благодаря свойству внешних стенок корневого чехлика покрываться слизью, что уменьшает трение между корнем и частичками почвы. Могут даже раздвигать частички почвы. Клетки корневого чехлика живые, часто содержат зёрна крахмала. Клетки чехлика постоянно обновляются за счёт деления. Участвует в положительных геотропических реакциях (направление роста корня к центру Земли).
Клетки зоны деления активно делятся, протяженность этой зоны у разных видов и у разных корней одного и того же растения неодинакова.
За зоной деления расположена зона растяжения (зона роста). Протяжённость этой зоны не превышает нескольких миллиметров.
По мере завершения линейного роста наступает третий этап формирования корня — его дифференциация, образуется зона дифференциации и специализации клеток (или зона корневых волосков и всасывания). В этой зоне уже различают наружный слой эпиблемы (ризодермы) с корневыми волосками, слой первичной коры и центральный цилиндр.
Строение корневого волоска
Корневые волоски — это сильно удлинённые выросты наружных клеток, покрывающих корень. Количество корневых волосков очень велико (на 1 мм2 от 200 до 300 волосков). Их длина достигает 10 мм. Формируются волоски очень быстро (у молодых сеянцев яблони за 30-40 часов). Корневые волоски недолговечны. Они отмирают через 10-20 дней, а на молодой части корня отрастают новые. Это обеспечивает освоение корнем новых почвенных горизонтов. Корень непрерывно растёт, образуя всё новые и новые участки корневых волосков. Волоски могут не только поглощать готовые растворы веществ, но и способствовать растворению некоторых веществ почвы, а затем всасывать их. Участок корня, где корневые волоски отмерли, некоторое время способен всасывать воду, но затем покрывается пробкой и теряет эту способность.
Волоски могут не только поглощать готовые растворы веществ, но и способствовать растворению некоторых веществ почвы, а затем всасывать их. Участок корня, где корневые волоски отмерли, некоторое время способен всасывать воду, но затем покрывается пробкой и теряет эту способность.
Оболочка волоска очень тонкая, что облегчает поглощение питательных веществ. Почти всю клетку волоска занимает вакуоль, окружённая тонким слоем цитоплазмы. Ядро находится в верхней части клетки. Вокруг клетки образуется слизистый чехол, который содействует склеиванию корневых волосков с частицами почвы, что улучшает их контакт и повышает гидрофильность системы. Поглощению способствует выделение корневыми волосками кислот (угольной, яблочной, лимонной), которые растворяют минеральные соли.
Корневые волоски играют и механическую роль — они служат опорой верхушке корня, которая проходит между частичками почвы.
Под микроскопом на поперечном срезе корня в зоне всасывания видно его строение на клеточном и тканевом уровнях.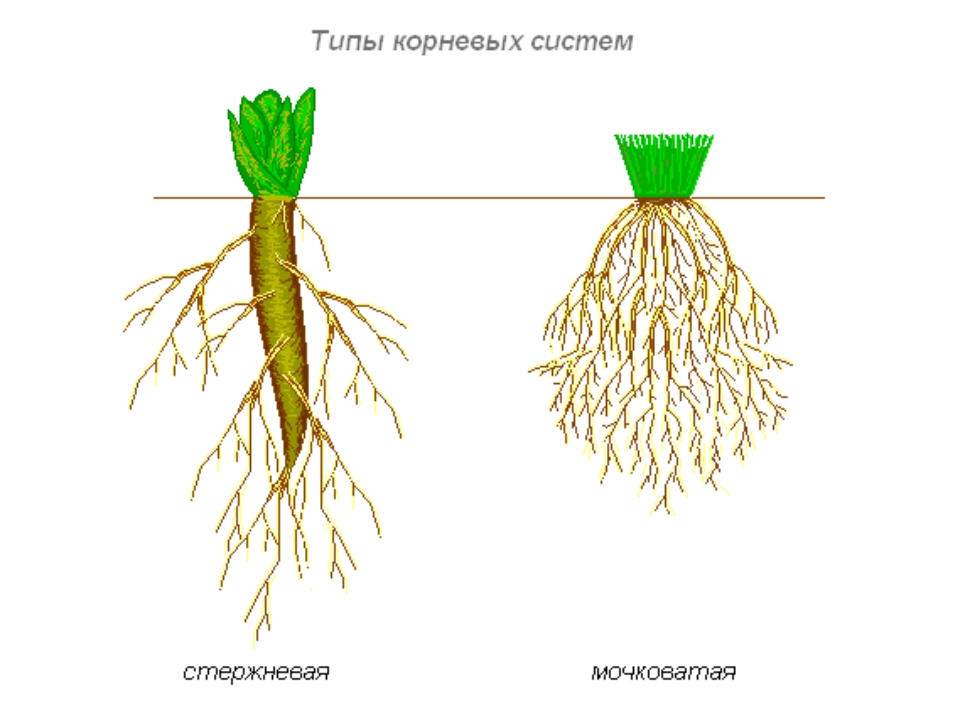 На поверхности корня — ризодерма, под ней — кора. Наружный слой коры — экзодерма, вовнутрь от неё — основная паренхима. Её тонкостенные живые клетки выполняют запасающую функцию, проводят растворы питательных веществ в радиальном направлении — от всасывающей ткани к сосудам древесины. В них же происходит синтез ряда жизненно важных для растения органических веществ. Внутренний слой коры — эндодерма. Растворы питательных веществ, поступающие из коры в центральный цилиндр через клетки эндодермы, проходят только через протопласт клеток.
На поверхности корня — ризодерма, под ней — кора. Наружный слой коры — экзодерма, вовнутрь от неё — основная паренхима. Её тонкостенные живые клетки выполняют запасающую функцию, проводят растворы питательных веществ в радиальном направлении — от всасывающей ткани к сосудам древесины. В них же происходит синтез ряда жизненно важных для растения органических веществ. Внутренний слой коры — эндодерма. Растворы питательных веществ, поступающие из коры в центральный цилиндр через клетки эндодермы, проходят только через протопласт клеток.
Кора окружает центральный цилиндр корня. Она граничит со слоем клеток, долго сохраняющих способность к делению. Это перицикл. Клетки перицикла дают начало боковым корням, придаточным почкам и вторичным образовательным тканям. Вовнутрь от перицикла, в центре корня, находятся проводящие ткани: луб и древесина. Вместе они образуют радиальный проводящий пучок.
Проводящая система корня проводит воду и минеральные вещества из корня в стебель (восходящий ток) и органические вещества из стебля в корень (нисходящий ток).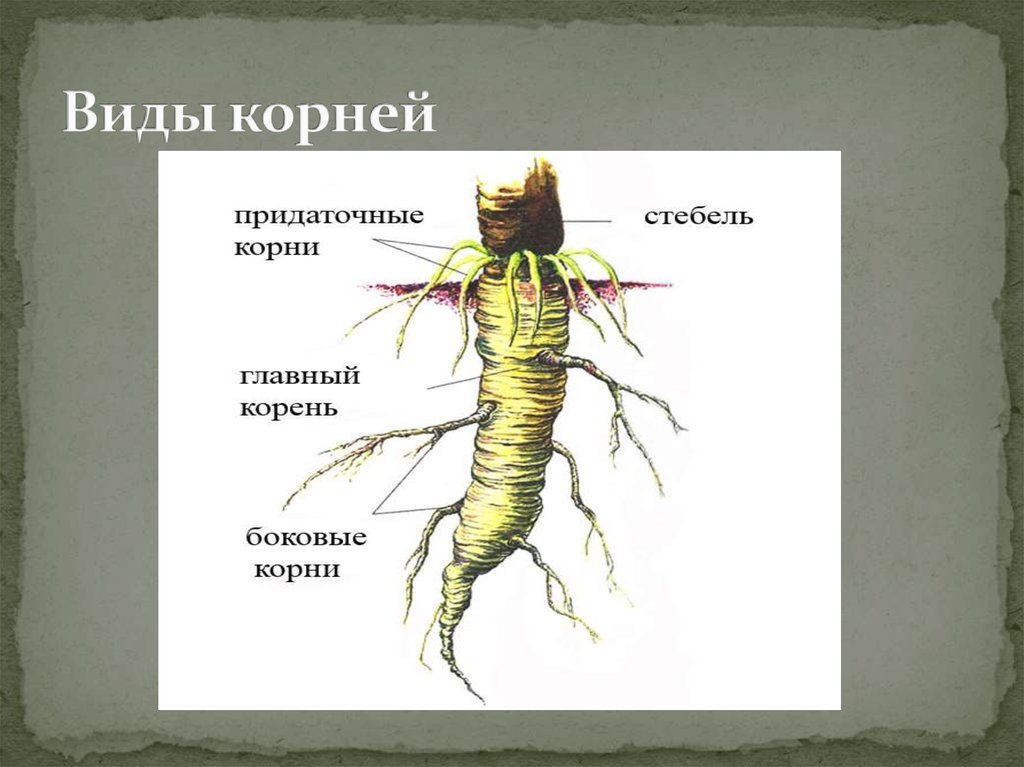 Состоит она из сосудисто-волокнистых пучков. Основными слагаемыми частями пучка являются участки флоэмы (по ним вещества передвигаются к корню) и ксилемы (по которым вещества передвигаются от корня). Основные проводящие элементы флоэмы — ситовидные трубки, ксилемы — трахеи (сосуды) и трахеиды.
Состоит она из сосудисто-волокнистых пучков. Основными слагаемыми частями пучка являются участки флоэмы (по ним вещества передвигаются к корню) и ксилемы (по которым вещества передвигаются от корня). Основные проводящие элементы флоэмы — ситовидные трубки, ксилемы — трахеи (сосуды) и трахеиды.
Процессы жизнедеятельности корня
Транспорт воды в корне
Всасывание воды корневыми волосками из почвенного питательного раствора и проведение её в радиальном направлении по клеткам первичной коры через пропускные клетки в эндодерме к ксилеме радиального проводящего пучка. Интенсивность поглощения воды корневыми волосками называется сосущей силой (S), она равна разнице между осмотическим (P) и тургорным (T) давлением: S=P-T.
Когда осмотическое давление равно тургорному (P=T), то S=0, вода перестаёт поступать в клетку корневого волоска. Если концентрация веществ почвенного питательного раствора будет выше, чем внутри клетки, то вода будет выходить из клеток и наступит плазмолиз — растения завянут. Такое явление наблюдается в условиях сухости почвы, а также при неумеренном внесении минеральных удобрений. Внутри клеток корня сосущая сила корня возрастает от ризодермы по направлению к центральному цилиндру, поэтому вода движется по градиенту концентрации (т. е. из места с большей её концентрацией в место с меньшей концентрацией) и создаёт корневое давление, которое поднимает столбик воды по сосудам ксилемы, образуя восходящий ток. Это можно обнаружить на весенних безлистных стволах, когда собирают «сок», или на срезанных пнях. Истекание воды из древесины, свежих пней, листьев, называется «плачем» растений. Когда распускаются листья, то они тоже создают сосущую силу и притягивают воду к себе — образуется непрерывный столбик воды в каждом сосуде — капиллярное натяжение. Корневое давление является нижним двигателем водного тока, а сосущая сила листьев — верхним. Подтвердить это можно с помощью несложных опытов.
Такое явление наблюдается в условиях сухости почвы, а также при неумеренном внесении минеральных удобрений. Внутри клеток корня сосущая сила корня возрастает от ризодермы по направлению к центральному цилиндру, поэтому вода движется по градиенту концентрации (т. е. из места с большей её концентрацией в место с меньшей концентрацией) и создаёт корневое давление, которое поднимает столбик воды по сосудам ксилемы, образуя восходящий ток. Это можно обнаружить на весенних безлистных стволах, когда собирают «сок», или на срезанных пнях. Истекание воды из древесины, свежих пней, листьев, называется «плачем» растений. Когда распускаются листья, то они тоже создают сосущую силу и притягивают воду к себе — образуется непрерывный столбик воды в каждом сосуде — капиллярное натяжение. Корневое давление является нижним двигателем водного тока, а сосущая сила листьев — верхним. Подтвердить это можно с помощью несложных опытов.
Всасывание воды корнями
Цель: выяснить основную функцию корня.
Что делаем: растение, выращенное на влажных опилках, отряхнём его корневую систему и опустим в стакан с водой его корни. Поверх воды для защиты её от испарения нальём тонкий слой растительного масла и отметим уровень.
Что наблюдаем: через день-два вода в ёмкости опустилась ниже отметки.
Результат: следовательно, корни всосали воду и подали её наверх к листьям.
Можно ещё проделать один опыт, доказывающий всасывание питательных веществ корнем.
Что делаем: срежем у растения стебель оставив пенёк высотой 2-3 см. На пенёк наденем резиновую трубку длиной 3 см, а на верхний конец наденем изогнутую стеклянную трубку высотой 20-25 см.
Что наблюдаем: вода в стеклянной трубке поднимается, и вытекает наружу.
Результат: это доказывает, что воду из почвы корень всасывает в стебель.
А влияет ли температура воды на интенсивность всасывания корнем воды?
Цель: выяснить, как температура влияет на работу корня.
Что делаем: один стакан должен быть с тёплой водой (+17-18ºС), а другой с холодной (+1-2ºС).
Что наблюдаем: в первом случае вода выделяется обильно, во втором — мало, или совсем приостанавливается.
Результат: это является доказательством того, что температура сильно влияет на работу корня.
Тёплая вода активно поглощается корнями. Корневое давление повышается.
Холодная вода плохо поглощается корнями. В этом случае корневое давление падает.
Минеральное питание
Физиологическая роль минеральных веществ очень велика. Они являются основой для синтеза органических соединений, а также факторами, которые изменяют физическое состояние коллоидов, т.е. непосредственно влияют на обмен веществ и строение протопласта; выполняют функцию катализаторов биохимических реакций; воздействуют на тургор клетки и проницаемость протоплазмы; являются центрами электрических и радиоактивных явлений в растительных организмах.
Установлено, что нормальное развитие растений возможно только при наличии в питательном растворе трёх неметаллов — азота, фосфора и серы и — и четырёх металлов — калия, магния, кальция и железа.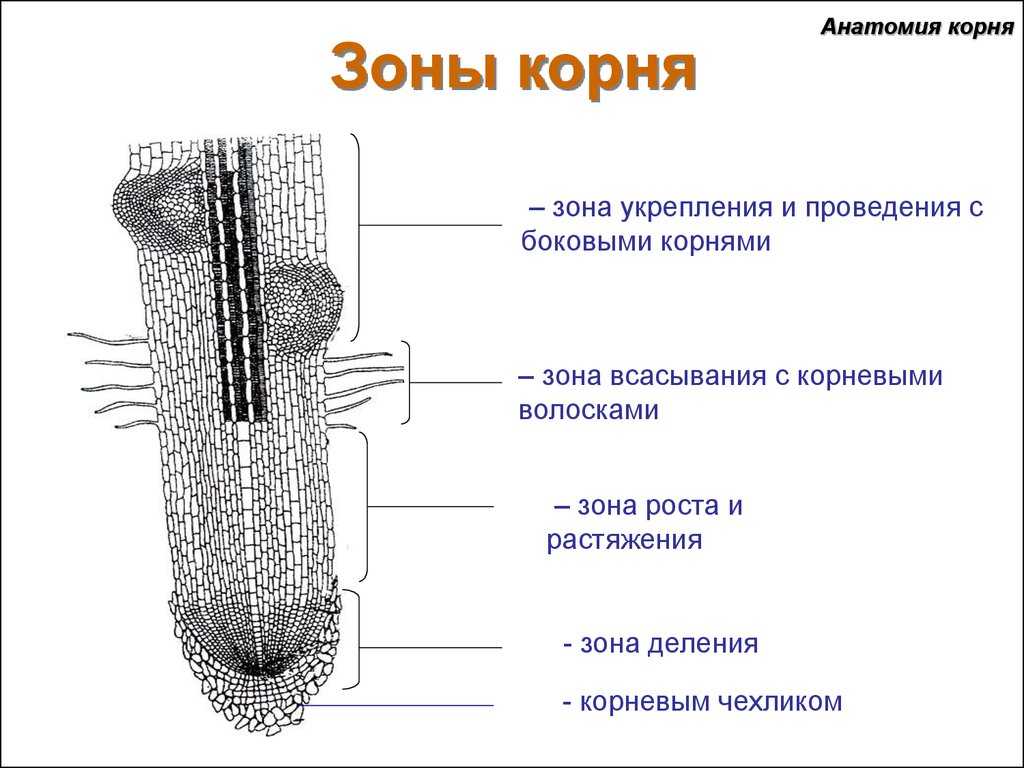 Каждый из этих элементов имеет индивидуальное значение и не может быть заменён другим. Это макроэлементы, их концентрация в растении составляет 10-2–10%. Для нормального развития растений нужны микроэлементы, концентрация которых в клетке составляет 10-5–10-3%. Это бор, кобальт, медь, цинк, марганец, молибден др. Все эти элементы есть в почве, но иногда в недостаточном количестве. Поэтому в почву вносят минеральные и органические удобрения.
Каждый из этих элементов имеет индивидуальное значение и не может быть заменён другим. Это макроэлементы, их концентрация в растении составляет 10-2–10%. Для нормального развития растений нужны микроэлементы, концентрация которых в клетке составляет 10-5–10-3%. Это бор, кобальт, медь, цинк, марганец, молибден др. Все эти элементы есть в почве, но иногда в недостаточном количестве. Поэтому в почву вносят минеральные и органические удобрения.
Растение нормально растёт и развивается в том случае, если в окружающей корни среде будут содержаться все необходимые питательные вещества. Такой средой для большинства растений является почва.
Дыхание корней
Для нормального роста и развития растения необходимо чтобы к корню поступал свежий воздух. Проверим, так ли это?
Цель: нужен ли воздух корню?
Что делаем: возьмём два одинаковых сосуда с водой. В каждый сосуд поместим развивающие проростки. Воду в одном из сосудов каждый день насыщаем воздухом с помощью пульверизатора. На поверхность воды во втором сосуде нальём тонкий слой растительного масла, так как оно задерживает поступление воздуха в воду.
На поверхность воды во втором сосуде нальём тонкий слой растительного масла, так как оно задерживает поступление воздуха в воду.
Что наблюдаем: через некоторое время растение во втором сосуде перестанет расти, зачахнет, и в конце концов погибнет.
Результат: гибель растения наступает из-за недостатка воздуха, необходимого для дыхания корня.
Видоизменения корней
У некоторых растений в корнях откладываются запасные питательные вещества. В них накапливаются углеводы, минеральные соли, витамины и другие вещества. Такие корни сильно разрастаются в толщину и приобретают необычный внешний вид. В формировании корнеплодов участвуют и корень, и стебель.
Корнеплоды
Если запасные вещества накапливаются в главном корне и в основании стебля главного побега, образуются корнеплоды (морковь). Растения, образующие корнеплоды, в основном двулетники. В первый год жизни они не цветут и накапливают в корнеплодах много питательных веществ. На второй — они быстро зацветают, используя накопленные питательные вещества и образуют плоды и семена.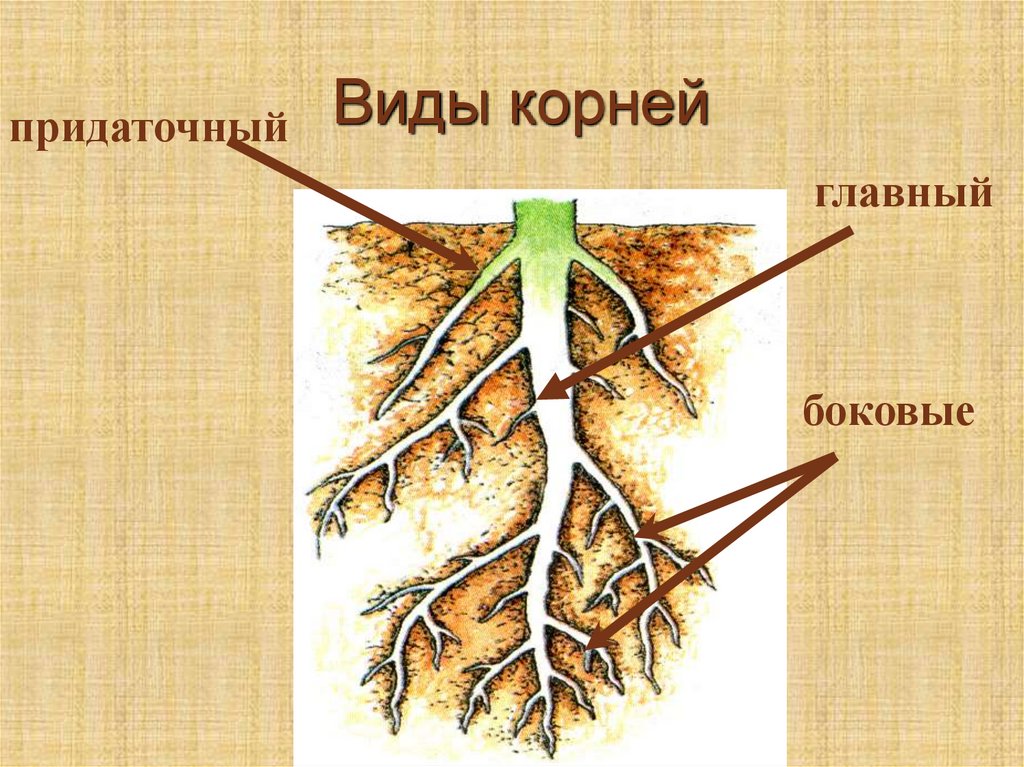
Корневые клубни
У георгина запасные вещества накапливаются в придаточных корнях, образуя корневые клубни.
Бактериальные клубеньки
Своеобразно изменены боковые корни у клевера, люпина, люцерны. В молодых боковых корешках поселяются бактерии, что способствует усвоению газообразного азота почвенного воздуха. Такие корни приобретают вид клубеньков. Благодаря этим бактериям эти растения способны жить на бедных азотом почвах и делать их более плодородными.
Ходульные
У пандуса, произрастающего в приливно-отливной зоне, развиваются ходульные корни. Они высоко над водой удерживают на зыбком илистом грунте крупные облиственные побеги.
Воздушные
У тропических растений, живущих на ветвях деревьев, развиваются воздушные корни. Они часто встречаются у орхидей, бромелиевых, у некоторых папоротников. Воздушные корни свободно висят в воздухе, не достигая земли и поглощая попадающую на них влагу от дождя или росы.
Втягивающие
У луковичных и клубнелуковичных растений, например у крокусов, среди многочисленных нитевидных корней имеется несколько более толстых, так называемых втягивающих, корней.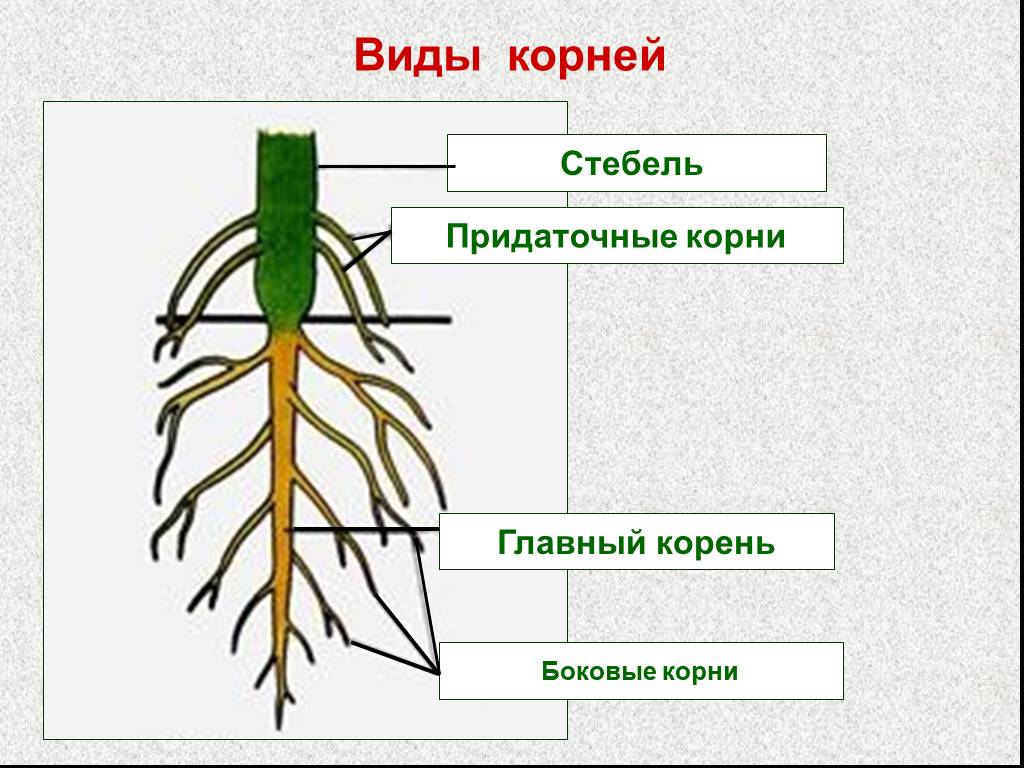 Сокращаясь, такие корни втягивают клубнелуковицу глубже в почву.
Сокращаясь, такие корни втягивают клубнелуковицу глубже в почву.
Столбовидные
У фикуса развиваются столбовидные надземные корни, или корни-подпорки.
Почва как среда обитания корней
Почва для растений является средой, из которой оно получает воду и элементы питания. Количество минеральных веществ в почве зависит от специфических особенностей материнской горной породы, деятельности организмов, от жизнедеятельности самих растений, от типа почвы.
Почвенные частицы конкурируют с корнями за влагу, удерживая её своей поверхностью. Это так называемая связанная вода, которая подразделяется на гигроскопическую и плёночную. Удерживается она силами молекулярного притяжения. Доступная растению влага представлена капиллярной водой, которая сосредоточена в мелких порах почвы.
Между влагой и воздушной фазой почвы складываются антагонистические отношения. Чем больше в почве крупных пор, тем лучше газовый режим этих почв, тем меньше влаги удерживает почва. Наиболее благоприятный водно-воздушный режим поддерживается в структурных почвах, где вода и воздух находятся одновременно и не мешают друг другу — вода заполняет капилляры внутри структурных агрегатов, а воздух — крупные поры между ними.
Характер взаимодействия растения и почвы в значительной степени связан с поглотительной способностью почвы — способностью удерживать или связывать химические соединения.
Микрофлора почвы разлагает органические вещества до более простых соединений, участвует в формировании структуры почвы. Характер этих процессов зависит от типа почвы, химического состава растительных остатков, физиологических свойств микроорганизмов и других факторов. В формировании структуры почвы принимают участие почвенные животные: кольчатые черви, личинки насекомых и др.
В результате совокупности биологических и химических процессов в почве образуется сложный комплекс органических веществ, который объединяют термином «гумус».
Метод водных культур
В каких солях нуждается растение, и какое влияние оказывают они на рост и развитие его, было установлено на опыте с водными культурами. Метод водных культур — это выращивание растений не в почве, а в водном растворе минеральных солей. В зависимости от поставленной цели в опыте можно исключить отдельную соль из раствора, уменьшить или увеличить ее содержание. Было выяснено, что удобрения, содержащие азот, способствуют росту растений, содержащие фосфор — скорейшему созреванию плодов, а содержащие калий — быстрейшему оттоку органических веществ от листьев к корням. В связи с этим содержащие азот удобрения рекомендуется вносить перед посевом или в первой половине лета, содержащие фосфор и калий — во второй половине лета.
Было выяснено, что удобрения, содержащие азот, способствуют росту растений, содержащие фосфор — скорейшему созреванию плодов, а содержащие калий — быстрейшему оттоку органических веществ от листьев к корням. В связи с этим содержащие азот удобрения рекомендуется вносить перед посевом или в первой половине лета, содержащие фосфор и калий — во второй половине лета.
С помощью метода водных культур удалось установить не только потребность растения в макроэлементах, но и выяснить роль различных микроэлементов.
В настоящее время известны случаи, когда выращивают растения методами гидропоники и аэропоники.
Гидропоника — выращивание растений в сосудах, заполненных гравием. Питательный раствор, содержащий необходимые элементы, подаётся в сосуды снизу.
Аэропоника — это воздушная культура растений. При этом способе корневая система находится в воздухе и автоматически (несколько раз в течение часа) опрыскивается слабым раствором питательных солей.
* * *
Ранняя пластичность боковых корней в ответ на асимметричную водообеспеченность
- Краткое сообщение
- Опубликовано:
- Даниэль фон Вангенхайм
ORCID: orcid.org/0000-0002-6862-1247 1,2 , - Джейсон Банда 1 ,
- Александр Шмитц 2 ,
5 Йенс Боланд0012 3 ,
ORCID: orcid.org/0000-0003-1545-0736 2 и
ORCID: orcid.org/0000-0003-0475-390X 1
Природные растения
том 6 , страницы 73–77 (2020)Процитировать эту статью
3597 доступов
18 цитирований
327 Альтметрический
Сведения о показателях
Субъекты
- Трехмерная реконструкция
- Морфогенез растений
Abstract
Ветвление корней зависит от почвенной среды и демонстрирует высокий уровень пластичности.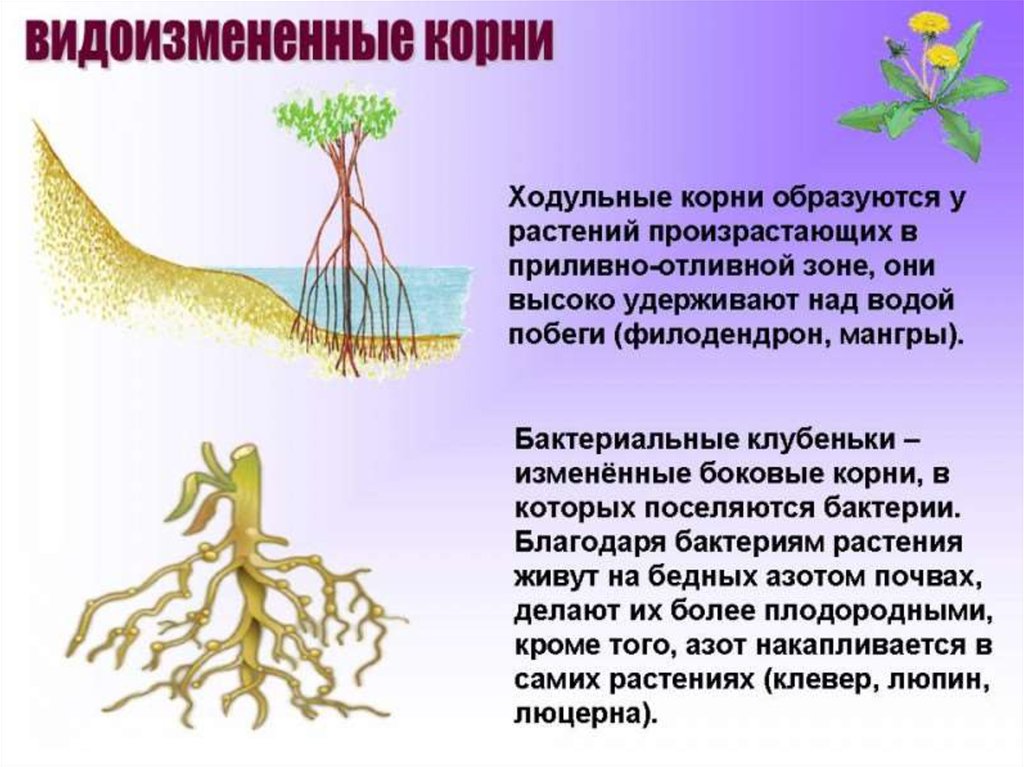 Мы сообщаем, что на радиальное расположение появляющихся боковых корней влияет их гидрологическая среда на ранних стадиях развития. Новые зачатки боковых корней имеют как высокую степень гибкости с точки зрения закладки, так и угла развития по отношению к доступной воде. Наши наблюдения показывают, как внешняя гидрологическая среда регулирует морфогенез боковых корней.
Мы сообщаем, что на радиальное расположение появляющихся боковых корней влияет их гидрологическая среда на ранних стадиях развития. Новые зачатки боковых корней имеют как высокую степень гибкости с точки зрения закладки, так и угла развития по отношению к доступной воде. Наши наблюдения показывают, как внешняя гидрологическая среда регулирует морфогенез боковых корней.
Это предварительный просмотр содержимого подписки, доступ через ваше учреждение
Варианты доступа
Подписаться на журнал
Получить полный доступ к журналу на 1 год
118,99 €
всего 9,92 € за выпуск
Подписаться
Расчет налогов будет завершен во время оформления заказа.
Купить статью
Получите ограниченный по времени или полный доступ к статье на ReadCube.
32,00 $
Купить
Все цены указаны без учета стоимости.
Рис. 1: Ветвление Arabidopsis зависит от положения корня в агаре. Рис. 2: Развитие боковых корней является гибким на всех стадиях развития.
Рис. 2: Развитие боковых корней является гибким на всех стадиях развития.
Доступность данных
Данные для рис. 1f показаны в дополнительном видео 1. Другие наборы данных могут быть предоставлены по разумному запросу.
Ссылки
Morris, E.C. et al. Формирование трехмерной архитектуры корневой системы. Курс. биол. 27 , R919–R930 (2017).
Артикул
КАСGoogle ученый
Дрю, М. С. Сравнение эффектов локализованного поступления фосфатов, нитратов, аммония и калия на рост семенной корневой системы и побегов ячменя. Н. Фитол. 75 , 479–490 (1975).
Артикул
КАСGoogle ученый
Орман-Лигеза, Б. и др. Реакция ксероветвления подавляет образование боковых корней, когда корни не контактируют с водой.
 Курс. биол. 28 , 3165–3173 (2018).
Курс. биол. 28 , 3165–3173 (2018).Артикул
КАСGoogle ученый
Бао Ю. и др. Корни растений используют механизм формирования паттерна для размещения боковых корневых ветвей по направлению к доступной воде. Проц. Натл акад. науч. США 111 , 9319–9324 (2014).
Артикул
КАСGoogle ученый
Ороса-Пуэнте, Б. и др. Ветвление корня в сторону воды связано с посттрансляционной модификацией фактора транскрипции ARF7. Наука 362 , 1407–1410 (2018).
Артикул
КАСGoogle ученый
фон Гуттенберг, Х. в Handbuch der Pflanzenanatomie (изд. Линсбауэр, К.) Band 8 (Gebrüder Bornträger, 1940).
Casero, P.J., Casimiro, I. & Lloret, P.G. Закладка боковых корней путем асимметричных поперечных делений клеток перицикла у четырех видов растений: Raphanus sativus , Helianthus annuus , Zea mays и Daucus carota .
Protoplasma 188 , 49–58 (1995).Артикул
Google ученый
Ласковски М.Дж., Уильямс М.Е., Нусбаум Х.К. и Сассекс И.М. Формирование меристем боковых корней представляет собой двухстадийный процесс. Разработка 121 , 3303–3310 (1995).
КАС
пабмедGoogle ученый
Касимиро, И. и др. Транспорт ауксина способствует инициации боковых корней Arabidopsis . Plant Cell 13 , 843–852 (2001).
Артикул
КАСGoogle ученый
Дубровский, Дж. Г., Рост, Т. Л., Колон-Кармона, А. и Доернер, П. Ранний морфогенез примордиев во время инициации боковых корней у Arabidopsis thaliana . Planta 214 , 30–36 (2001).
Артикул
КАСGoogle ученый
фон Вангенхайм, Д. и др. Правила и самоорганизующие свойства постэмбриональных моделей деления клеток органов растений. Курс. биол. 26 , 439–449 (2016).
Артикул
КАСGoogle ученый
Касимиро И. и др. Препарирование Arabidopsis развития боковых корней. Trends Plant Sci. 8 , 165–171 (2003).
Артикул
КАСGoogle ученый
Го Т., Джой С., Мимура Т. и Фукаки Х. Установление асимметрии в Клетки-основатели боковых корней Arabidopsis регулируются LBD16/ASL18 и родственными белками LBD/ASL.
Развитие 139 , 883–893 (2012).Артикул
КАСGoogle ученый
Stelzer, EHK. Световая флуоресцентная микроскопия для количественной биологии. Нац. Методы 12 , 23–26 (2015).
Артикул
КАСGoogle ученый
фон Вангенхайм, Д., Хаушильд, Р. и Фримл, Дж. Световая флуоресцентная микроскопия корней растений, растущих на поверхности геля. Дж. Вис. Эксп. 2017 , e55044 (2017).
Google ученый
Schindelin, J. et al. Фиджи: платформа с открытым исходным кодом для анализа биологических изображений. Нац. Методы 9 , 676–682 (2012).
Артикул
КАСGoogle ученый
Preibisch, S., Saalfeld, S.
, Schindelin, J. & Tomancak, P. Программное обеспечение для регистрации данных микроскопии с селективным плоским освещением на основе шариков. Нац. Методы 7 , 418–419 (2010).Артикул
КАСGoogle ученый
Preibisch, S. et al. Эффективная байесовская многоракурсная деконволюция. Нац. Методы 11 , 645–648 (2014).
Артикул
КАСGoogle ученый
 Курс. биол. 28 , 3165–3173 (2018).
Курс. биол. 28 , 3165–3173 (2018).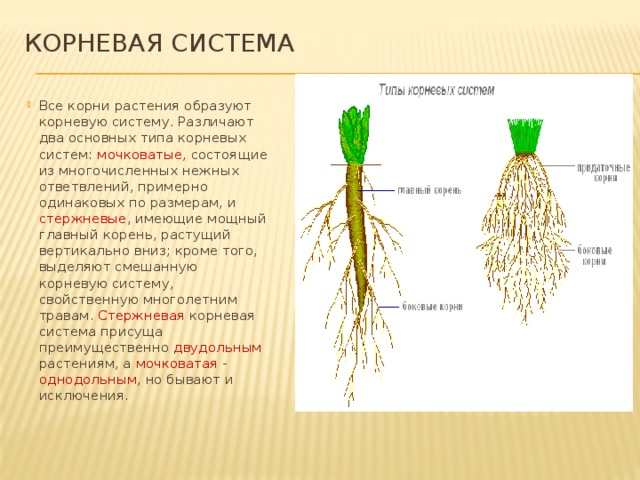 Protoplasma 188 , 49–58 (1995).
Protoplasma 188 , 49–58 (1995).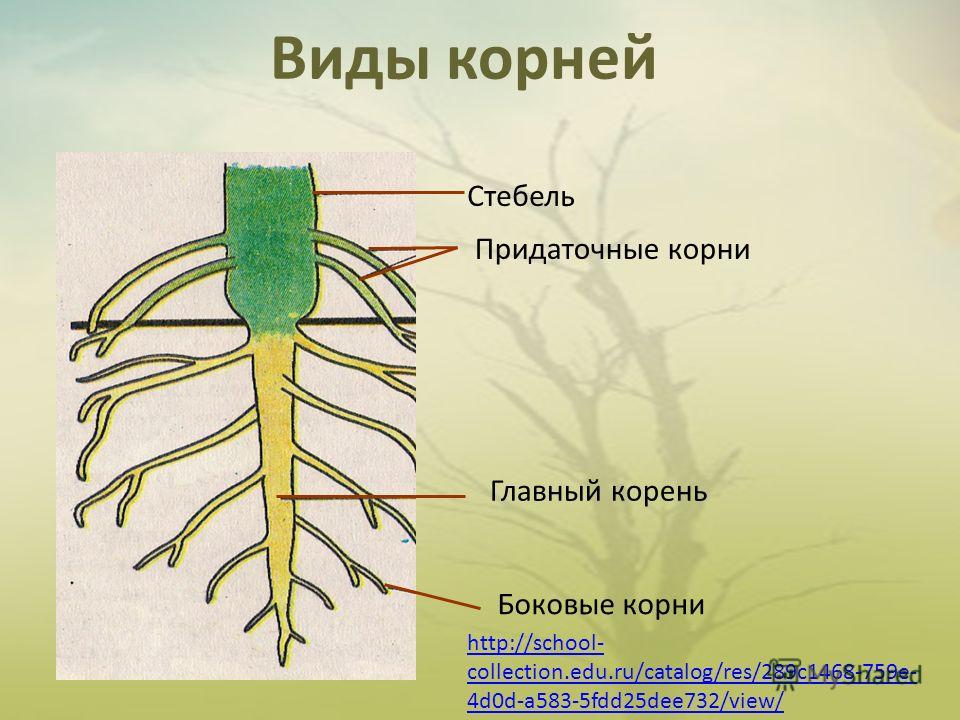 «>
«>Лукас, М. и др. Морфогенез боковых корней зависит от механических свойств покрывающих тканей. Проц. Натл акад. науч. США 110 , 5229–5234 (2013).
Артикул
КАС
Google ученый
.jpg) Развитие 139 , 883–893 (2012).
Развитие 139 , 883–893 (2012). , Schindelin, J. & Tomancak, P. Программное обеспечение для регистрации данных микроскопии с селективным плоским освещением на основе шариков. Нац. Методы 7 , 418–419 (2010).
, Schindelin, J. & Tomancak, P. Программное обеспечение для регистрации данных микроскопии с селективным плоским освещением на основе шариков. Нац. Методы 7 , 418–419 (2010).Ссылки на скачивание
Благодарности
Эта работа была поддержана наградами Совета по исследованиям в области биотехнологии и биологических наук (гранты №№ BB/M012212, BB/G023972/1, BB/R013748/1, BB/L026848/1 , BB/M018431/1, BB/PO16855/1 и BB/M001806/1), Европейского исследовательского совета FUTUREROOTS Advanced Investigator (грант № 294729) и Leverhulme Trust (грант № RPG-2016-409). Э.Х.К.С. финансируется Deutsche Forschungsgemeinschaft (CEF-MC I/II, DFG Exc 115). Исследования в лаборатории Майзеля поддерживаются DFG FOR2581, Землей Баден-Вюртемберг, фондом Chica und Heinz Schaller Stiftung, передовым кластером CellNetworks и фондом Boehringer Ingelheim Foundation.
Исследования в лаборатории Майзеля поддерживаются DFG FOR2581, Землей Баден-Вюртемберг, фондом Chica und Heinz Schaller Stiftung, передовым кластером CellNetworks и фондом Boehringer Ingelheim Foundation.
Информация о авторе
Авторы и принадлежности
Растительные и сельскохозяйственные науки, Школа биологических наук, Университет Ноттингема, Ноттингем, Великобритания
Daniel Von Wangenheim, Jason Banda, Anthony Bishopp & Malcolmolm. Bnenett
. Молекулярные науки о жизни, Гете-Университет Франкфурта-на-Майне, Франкфурт-на-Майне, Германия
Даниэль фон Вангенхайм, Александр Шмитц и Эрнст Х. К. Штельцер
Центр изучения организмов, Гейдельбергский университет, Гейдельберг, Германия
Jens Boland & Alexis Maizel
Авторы
- Daniel von Wangenheim
Просмотреть публикации этого автора Вы также можете искать этого автора
PubMed Google Scholar
- Jason Banda
Просмотр публикаций автора
Вы также можете искать этого автора в
PubMed Google Scholar - Александр Шмитц
Посмотреть публикации автора
Вы также можете искать этого автора в
PubMed Google Scholar - Jens Boland
Просмотр публикаций автора
Вы также можете искать этого автора в
PubMed Google Scholar - Anthony Bishopp
Просмотр публикаций автора
Вы также можете искать этого автора в
PubMed Google Scholar - Алексис Майзел
Просмотр публикаций автора
Вы также можете искать этого автора в
PubMed Google Scholar - Ernst H. K. Stelzer
Просмотр публикаций автора
Вы также можете искать этого автора в
PubMed Google Scholar - Malcolm Bennett
Просмотр публикаций автора
Вы также можете искать этого автора в
PubMed Google Scholar
 K. Stelzer
K. StelzerВклады
Д.в.В., Дж. Банда, А.Б., А.М., Э.Х.К.С. и М.Б. проектировал эксперименты. Д.в.В. и J. Banda выполнили эксперименты, а D.v.W., J. Banda, A.S., J. Boland, A.B., A.M., E.H.K.S. и М.Б. проанализировал данные. Д.в.В., Дж. Банда и М.Б. написал рукопись с участием всех других авторов.
Авторы переписки
Переписка с
Даниэль фон Вангенхайм или Малкольм Беннетт.
Декларации этики
Конкурирующие интересы
Авторы заявляют об отсутствии конкурирующих интересов.
Дополнительная информация
Примечание издателя Springer Nature остается нейтральной в отношении юрисдикционных претензий в опубликованных картах и институциональной принадлежности.
Дополнительная информация
Дополнительная информация
Дополнительная информация Рис. 1–4 с легендами.
Сводка отчетов
Дополнительное видео 1
Набор данных бокового корневого угла.
Дополнительное видео 2
3D-вид рис. 2d–g.
Дополнительные данные 1
Исходные статистические данные.
Права и разрешения
Перепечатка и разрешения
Об этой статье
Эта статья цитируется
Многомасштабные сети предприятий: построение графиков соединений предприятий с помощью методов многоуровневого анализа и визуализации
- Си Чжан
- Йи Ман
- Цзиньсин Линь
Наука Китая Life Sciences (2021)
Корневая архитектура и гидравлика объединяются для адаптации к изменяющейся доступности воды
- Кристоф Морель
- Филипп Накри
Природные растения (2020)
Раннее развитие боковых корней обусловлено жаждой
Корни необходимы растениям для поиска воды и питательных веществ.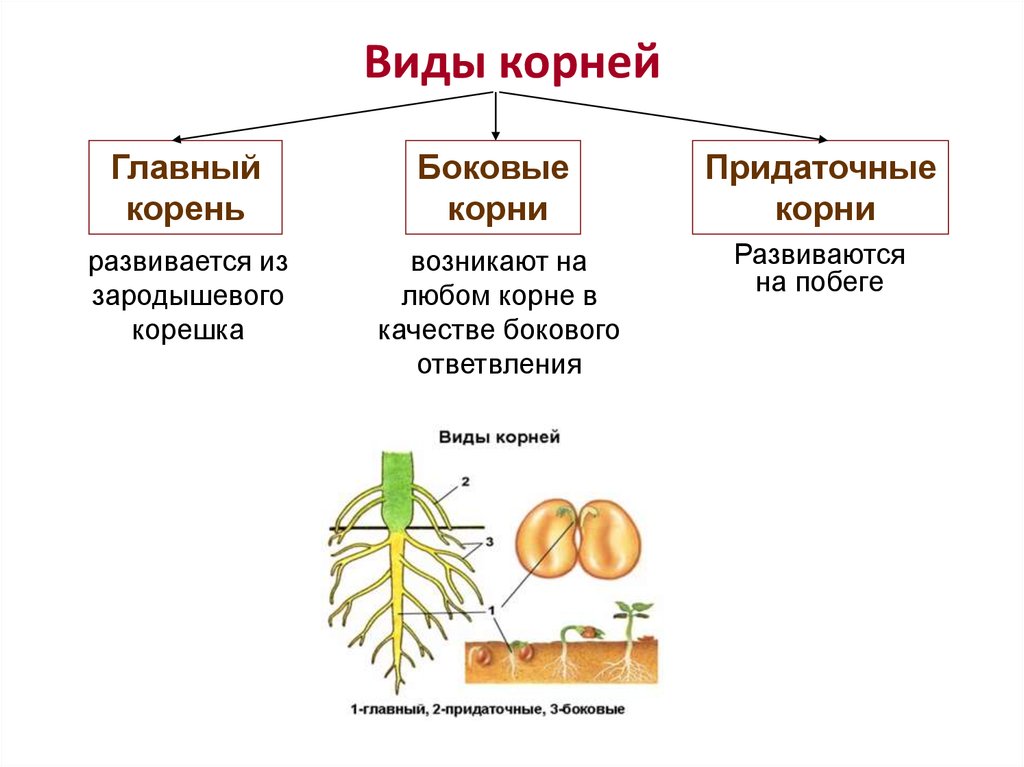 Они также могут быть дорогими в выращивании, поэтому растение должно направить их в правильном направлении. Как они это делают? В новой статье Даниэля фон Вангенхайма и его коллег из Nature Plants исследуется, как боковые корни растут по направлению к воде на самых ранних стадиях их формирования.
Они также могут быть дорогими в выращивании, поэтому растение должно направить их в правильном направлении. Как они это делают? В новой статье Даниэля фон Вангенхайма и его коллег из Nature Plants исследуется, как боковые корни растут по направлению к воде на самых ранних стадиях их формирования.
Внутренняя часть молодого корня представляет собой почти ряд концентрических слоев. Снаружи вы проходите через эпидермис, кору и эндодерму, пока не достигнете перицикла. Перицикл представляет собой слой клеток, окружающий стелу, сердцевину корня, содержащую сосудистую ткань. Сосудистая ткань — это ксилема, которая несет воду и минеральные вещества от корней, и флоэма, которая переносит сахара от листьев. Если бы вы разрезали корень в поперечном сечении, вы бы увидели, как ксилема тянется от одной стороны стелы к другой, как стержень, как если бы у нее было два конца или «полюса».
Полюса ксилемы в корне Arabidopsis . Ive De Smet 2011.
Де Смет написал мини-обзор, в котором подытожил то, что было известно в 2011 году. То, что боковые корни растут из перицикла, не новость. Неудивительно и то, что корни, по-видимому, растут из полюсов ксилемы. Боковые корни будут дополнением к внутренней сантехнике растения, поэтому «трубы» должны соединяться. Следовательно, должен быть путь к ксилеме в главном корне.
То, что боковые корни растут из перицикла, не новость. Неудивительно и то, что корни, по-видимому, растут из полюсов ксилемы. Боковые корни будут дополнением к внутренней сантехнике растения, поэтому «трубы» должны соединяться. Следовательно, должен быть путь к ксилеме в главном корне.
Особенность, обнаруженная фон Вангенхаймом и его коллегами, заключается в том, что если есть тенденция к тому, чтобы вода находилась на одной стороне корня, боковые корни растут в эту сторону. Это тоже не должно вызывать удивления. Наращивание корней требует энергии и материалов, растение захочет делать это там, где оно получает наибольшую отдачу. Но есть сюрприз, с какой стороны от перицикла начинают расти боковые корни. Если есть два полюса ксилемы, то вы с одинаковой вероятностью найдете боковые корни, начиная с сухой стороны, как и с влажной стороны.
Но когда команда фон Вангенхайма вырастила корни на агаровом геле, чтобы посмотреть, как развиваются первичные боковые корни, они не обнаружили, что корни с двух сторон ведут себя по-разному.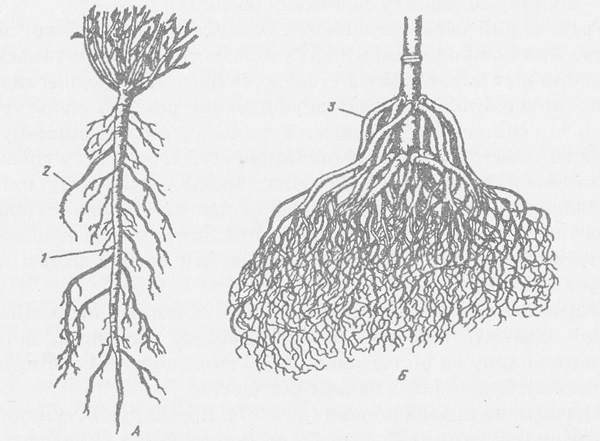 «Зачатки, появляющиеся на воздушной стороне, ориентированы в основном на поверхность геля. Напротив, — пишут они, — зачатки, зарождающиеся на контактной стороне, ориентированы в основном параллельно поверхности геля. Эти результаты показывают, что угол бокового отростка корня очень пластичен и направляет развитие органов преимущественно в сторону доступных извне источников воды».
«Зачатки, появляющиеся на воздушной стороне, ориентированы в основном на поверхность геля. Напротив, — пишут они, — зачатки, зарождающиеся на контактной стороне, ориентированы в основном параллельно поверхности геля. Эти результаты показывают, что угол бокового отростка корня очень пластичен и направляет развитие органов преимущественно в сторону доступных извне источников воды».
Команда решила выяснить, как рано боковой корень начинает расти под углом. Ранее считалось, что именно клетки над полюсом ксилемы наиболее важны для развития нового корня. Однако из-за асимметричного роста клетки, окружающие центральный клеточный массив, также направляют корень.
Глядя на предыдущую работу по этому вопросу для написания этого поста, я нашел много молекулярных вкладов в рост боковых корней, но мало на клеточном уровне. Казалось бы, дело не только в моих плохих поисковых способностях. Фон Вангенхайм и его коллеги пишут: «В отличие от этих более ранних исследований, которые были сосредоточены на основных молекулярных механизмах, контролирующих это поведение, это исследование было сосредоточено на механизмах в масштабе клетки-органа, которые способствуют росту LRP в ответ на воду. доступность. Наше исследование показывает, что, в отличие от оси полюса ксилемы, выбор файлов клеток перицикла, которые инициируют новый зачаток бокового корня, связан с внешним гидрологическим ландшафтом».
доступность. Наше исследование показывает, что, в отличие от оси полюса ксилемы, выбор файлов клеток перицикла, которые инициируют новый зачаток бокового корня, связан с внешним гидрологическим ландшафтом».
Несоответствие между двумя элементами особенно интересно. Направление корня от перицикла показывает, что наличие воды имеет решающее значение для направления корня. Напротив, инициация от любого полюса ксилемы показывает, что где-то сигнал к началу процесса не зависит от этого гидрологического смысла. Я задаюсь вопросом, есть ли что-то очевидное, что я упускаю из виду, что объясняет, почему рост не всегда начинается на полюсе ксилемы на самой влажной стороне.
Благодаря очень умной и искусной работе команде фон Вангенхайма удалось показать, что есть проблема в соединении развития клеток и органов с молекулярным механизмом, который управляет ростом боковых корней. В газете также есть несколько ярких изображений и видео, иллюстрирующих их работу. Когда общая ссылка станет доступной, я вернусь, чтобы отредактировать это и направить вас к этому.