
РАСТЕНИЕВОДСТВО. Аминокислоты для растений. Аминокислоты для растений
Аминокислоты для растений
Часто растения нуждаются в тех же питательных веществах, что и человек. Они необходимы для полноценного функционирования растительного организма и его дальнейшего развития. Среди таких важных соединений — аминокислоты, использование которых в выращивании сельхозкультур может давать поразительные результаты.Активное изучение воздействия на растения подкормок аминокислотами началось в 70–80-е годы прошлого века. Многие ученые отмечали, что эти вещества активируют механизмы роста после соляного стресса и низких температур, повышают фертильность пыльцы и образование завязи плодов, увеличивают способность усвоения элементов питания и устойчивость к вредителям, болезням.
Растения и животные быстрее и лучше усваивают натуральные α-аминокислоты оптически активной L-конфигурации, из которых строятся белки. Такие модификации легко воспринимаются растительным организмом и быстро включаются в метаболизм как собственные. D-формы аминокислот встречаются в природе сравнительно редко, причем только как продукты обмена веществ низших организмов.
ПРАКТИЧЕСКИЙ ЭКЗАМЕН
Благодаря современным методам анализа эффект от проведения подкормок растений L-α-аминокислотами достаточно хорошо изучен. Первыми зарегистрированными в России агрохимикатами, содержащими эти вещества, был антистрессант «Мегафол», стимулятор гармоничного развития «Вива» и стимулятор корнеобразования «Радифарм». Они появились в нашей стране в 2004 году, который характеризовался затяжной холодной весной, поэтому результаты их применения на сельхозкультурах были сразу хорошо заметны. В 2005–2006 годах из-за резкого падения температуры до –35°С в середине января на Северном Кавказе отмечался большой процент вымерзания озимых культур, гибель виноградной лозы и плодовых почек косточковых культур. Семечковые сады встретили весну в состоянии глубокого ступора. Сложившиеся погодные условия дали возможность увидеть эффективность применения аминокислотных агрохимикатов — они способствовали сохранению озимых культур, а на семечковых садах весенняя антистрессовая программа позволила получить полноценный урожай плодов.
В 2006 году в ЗАО АФ «Солнечная» был заложен опыт на промышленных томатах в открытом грунте с использованием систем капельного полива. Для этого использовались два расположенных рядом земельных участка по три гектара. На контроле применялась разработанная в хозяйстве бюджетная схема питания простыми водорастворимыми удобрениями. На опытном наделе к ней добавлялись агрохимикаты «Радифарм», «Активейв» и «Вива», а на площади 0,5 га во время вегетации проводилась коррекция питания с помощью листовых подкормок. Антистрессант «Мегафол» в дозировке 1 л/га в сочетании с другими специальными агрохимикатами в этот период вносился шесть раз. При уборке с 7 августа по 18 сентября 2006 года сотрудники хозяйства констатировали высокую эффективность корневых и некорневых подкормок препаратами, содержащими в своем составе аминокислоты. Проведенный опыт и его результаты способствовали быстрому распространению по стране практики применения подобных удобрений на всех сельхозкультурах.
ЗНАКОМСТВО ПОБЛИЖЕ
Сегодня в России зарегистрировано множество агрохимикатов, содержащих аминокислоты, которые активно рекламируются в СМИ. Однако на практике аграрии не всегда могут правильно разобраться в некоторых определениях.
Термин «свободные аминокислоты» — САК — применим как к белковым, так и небелковым аминокислотам. Первые могут находиться в растительном организме в не связанной в белки или пептиды форме. Количество подобных веществ достаточно высоко в молодых растениях и с возрастом понижается, причем в вегетативных органах свободных аминокислот больше, чем в репродуктивных. Увеличение общего количества САК наблюдается при пониженном питании растений калием, фосфором, серой, кальцием и магнием. Такой же эффект возникает при недостатке цинка, меди, марганца, железа, что связано с ослаблением в этих условиях синтеза белков из аминокислот.
Второй тип этих веществ структурно аналогичен первому и также может содержаться в растениях. Небелковая разновидность может выполнять функцию азот-запасающего компонента, но в больших количествах является токсичной. Она принимает участие в биосинтезе рибосомального белка, что приводит к образованию дефектной формы этого вещества, вызывает у растений, микроорганизмов, насекомых уменьшение роста и в некоторых случаях гибель. Многие небелковые САК, образовавшиеся при метаболизме низших организмов, имеют свойства антибиотиков и являются конкурентными ингибиторами при обмене веществ, задерживая определенные ступени биосинтеза белковых аминокислот или способствуя образованию ложных последовательностей при создании белков. В связи с этим применение в описании агрохимикатов термина «свободные аминокислоты» как однозначно более эффективного материала требует определенной конкретизации.
Сегодня наукой описано около 300 аминокислот, найденных в природе. Однако из них только 20 входят в состав белков, то есть могут называться протеиногенными. Это основные составные части животных и растительных белков, при этом их встраивание в молекулу этого вещества регулируется информацией генетического кода. С 2015 года в рамках программы импортозамещения в России появились специальные высокотехнологичные агрохимикаты отечественного производства — «Аминофол» и «Максифол». Один из основных компонентов новых продуктов — свободные протеиногенные L-α-аминокислоты в концентрации более 10 процентов, которая обеспечивает их высокую эффективность.Текст: А. Хорошкин, канд. с.-х. наук, ведущий специалист ГК «АгроМастер»
www.agbz.ru
Агромастер - Аминокислоты в листовых подкормках
Хорошкин А.Б. кандидат с-х наук, ведущий специалист ГК «АгроМастер», г. Краснодар
Активное изучение действия подкормок аминокислотами на растения началось в 70-80-е годы прошлого века. Многие ученые отмечали, что аминокислоты активируют механизмы роста после соляного стресса и низких температур [1, 2, 3], повышают фертильность пыльцы и образование завязи плодов [4],повышают способность усвоения элементов питания[5] и устойчивость к вредителям и болезням [6] и т.д.
Было открыто, что растения и животные быстрее и лучше усваивают натуральные α-аминокислоты (из которых строятся белки) оптически активной L-конфигурации. L-α-аминокислоты легко усваиваются растениями и быстро включаются в метаболизм как собственные. «D-формы аминокислот встречаются в природе сравнительно редко»[7],«как продукты метаболизма низших организмов»[8].
В настоящее время эффект от проведения подкормок растений L-α-аминокислотами, благодаря современным методам анализа, достаточно хорошо изучен. Если свести воедино все известные данные, то получается следующая картина:
Действие свободных протеиногенных α-аминокислот на растения
|
L-Leucine (Лейцин) и L-Isoleucine (Изолейцин) Повышает устойчивость к засолению (солевому стрессу) Улучшает прорастание пыльцы L-Tyrosine (Тирозин) Улучшает прорастание пыльцы L-Aspartic Acid (Аспарагиновая кислота): Активизирует прорастание семян Участвует в метаболизме аминокислот Источник органического азота L-Glutamic Acid (Глютаминовая кислота): Хорошие свойства хелатора Стимулятор роста Активизирует прорастание семян Способствует открытию устьиц Улучшает опыляемость Предшественник хлорофилла Предшественник аминокислот Активатор механизмов устойчивости к патогенам L-Arginine (Аргинин): Повышает холодостойкость Стимулирует синтез гормонов связанных с цветением и плодоношением Усиливает развитие корней Предшественник полиаминов Повышает устойчивость к засолению L-Phenylaninine (Фенилаланин): Активизирует прорастание семян Предшественник лигнина α-Glycine (Глицин) (оптически неактивен): Хорошие свойства хелатора Способствует росту тканей Улучшает вкус плодов Предшественник пиррола (C4H5N) – ядро Пиррола составная часть хлорофилла, витамина В12, цитохромов и других биологически активных соединений. L-Histidine (Гистидин): Хорошие свойства хелатора Улучшает созревание плодов Регулирует открытие устьиц L-Alanine (Аланин): Повышает холодостойкость Стимулирует синтез хлорофилла Улучшает качество плодов Регулирует открытие устьиц Повышает устойчивость к суховеям и засухе |
L-Lysine (Лизин): Хорошие свойства хелатора Стимуляция синтеза хлорофилла Активизирует прорастание семян Улучшает процессы опыления и оплодотворения Повышает устойчивость к суховеям и засухе L-Methionine (Метионин): Активизирует прорастание семян Стимулирует производство этилена Улучшает процессы опыления и оплодотворения Предшественник факторов роста Усиливает рост корней Регулирует открытие устьиц L-Proline (Пролин): Антистрессовое действие Повышает сопротивляемость осмотическим стрессам, регулирует водный обмен в растении Способствует открытию устьиц Повышает содержание хлорофилла и фотосинтетическую способность Улучшает генеративное развитие растений Повышает фертильность пыльцы и завязывание плодов Улучшает вкус плодов Усиливает способность семян к прорастанию L-Serine (Серин): Предшественник ауксина Повышает сопротивляемость стрессовым воздействиям Улучшает опыление и оплодотворение Образование гумусовых составов L-Threonine (Треонин): Активизирует прорастание семян Регулирует механизм защиты во время стресса Усиливает процесс гумификации L-Tryptophan (Триптофан): Предшественник ауксина Стимулирует рост меристемных тканей L-Valine (Валин): Предшественник ауксина Улучшает качество плодов Повышает устойчивость к суховеям и засухе Улучшает формирование семян L-Cysteine (Цистеин): Хорошие свойства хелатора Антиокислительная активность Важный компонент баланса клеточных функций |
Первые, зарегистрированные в России агрохимикаты содержащие аминокислоты: антистрессант - Мегафол; стимулятор гармоничного развития – Вива и стимулятор корнеобразования - Радифарм (производства «Valagro») появились в России в 2004 году. Этот год характеризовался затяжной, холодной весной, поэтому результаты применения этих агрохимикатов на с/х культурах были заметны визуально.
В сезоне 2005-2006 г.г. в результате резкого падения температуры до минус 35°С в середине января, на Северном Кавказе фиксировали большой процент вымерзания озимых культур, гибель виноградной лозы и плодовых почек косточковых культур. Семечковые сады входили в весну в состоянии глубокого ступора. Было отмечено, что применение аминокислотных агрохимикатов для обработки семян способствовало сохранению озимых культур, а на семечковых садах весенняя антистрессовая программа позволила получить полноценный урожай плодов.
В 2006 году, в ЗАО АФ «Солнечная» (г. Краснодар) на промышленных томатах в открытом грунте с использованием систем капельного полива был заложен опыт на двух (расположенных рядом) участках по 3 га. На контроле применялась разработанная в хозяйстве бюджетная схема питания простыми водорастворимыми удобрениями. На опытном участке к этой базовой (фоновой) схеме, в «каплю» добавлялись Радифарм, Активейв и Вива, а на площади 0,5 га в период вегетации проводилась коррекция питания с помощью листовых подкормок. Мегафол в течение вегетации вносился 6 раз в сочетании с другими специальными агрохимикатами, в дозе 1,0 л/га. С 7.08.06 г. по 18.09.06 г. на опытном и контрольном участках производилась комиссионная учетная уборка томатов.
Урожайность томатов Перфект Пим за период с 07.08.06 г по 18.09.06 г.
на опытном и контрольном участках (ЗАО АФ «Солнечная», 2006 г.)
|
Участок |
Учетная площадь участка, га |
Вес, тн |
Урожайность, тн/га |
||
|
Контроль (схема фертигации хозяйства – Фон |
3,0 |
132,3 |
44,1 |
||
|
Опыт |
Фон+Радифарм, Активейв, Вива-фертигация |
3,0 |
2,5 |
155,9 |
62,4 |
|
Фон+Радифарм, Активейв, Вива-фертигация +листовые подкормки + Мегафол |
0,5 |
54,6 |
109,2 |
||
Проведенный опыт наглядно продемонстрировал эффективность корневых и некорневых подкормок специальными агрохимикатами, которые включают в состав аминокислоты. Это способствовало быстрому распространению по стране практики применения данных удобрений на всех с/х культурах.
В настоящее время в России зарегистрировано множество агрохимикатов содержащих аминокислоты. Их сопровождает большое количество рекламной информации, но некоторые термины могут иметь двоякое или широкое толкование, что требует дополнительных разъяснений.
Так, термин «свободные аминокислоты» (САК) применим как к белковым, так и небелковым аминокислотам. Белковые (протеиногенные) аминокислоты могут находиться в растительном организме в несвязанной в белки или пептиды форме. Количество белковых САК достаточно высоко в молодых растениях (или органах) и с возрастом понижается. В вегетативных органах растений свободных аминокислот больше, чем в репродуктивных. Увеличение общего количества САК наблюдается при пониженном питании растений калием, фосфором, серой, кальцием и магнием. Такое же действие происходит при недостатке ряда микроэлементов: цинка, меди, марганца, железа. Это связано с ослаблением синтеза белков из аминокислот в этих условиях.
По поводу непротеиногенных аминокислот доктор биологических наук Чекалин Н.М. в своей монографии «Генетические основы селекции зернобобовых культур на устойчивость к патогенам» (Полтава, 2003) писал: «Свободные (небелковые) аминокислоты (900 структур), структурно аналогичные белковым аминокислотам, в изобилии содержатся у некоторых бобовых (Vicieae, Phaseoleae, Mimosoideae), являются токсичными и выполняют функцию азот-запасающего компонента. Свободные аминокислоты (САК) могут: принимать участие в биосинтезе рибосомального белка, приводя к образованию дефектного белка; ингибировать синтез аминоацил-tРНК-синтетазы или другие пути биосинтеза белка. Свободные аминокислоты вызывают у растений, микроорганизмов и насекомых уменьшение роста и даже гибель; у позвоночных - эмбриональное уродство, нейротоксичные нарушения, паралич, цирроз печени, аритмию и др.» «Многие (небелковые САК – А.Х.) аминокислоты, образовавшиеся при обмене веществ низших организмов, имеют свойства антибиотиков. Они действуют как аминокислоты – антагонисты, т.е. являются конкурентными ингибиторами при обмене веществ, задерживая определенные ступени биосинтеза аминокислот или способствуя образованию ложных последовательностей при биосинтезе белков» [9].
Исходя из вышеизложенного, применение в описании агрохимикатов термина «свободные аминокислоты», как однозначно более эффективного материала, требует определенной конкретизации.
«В настоящее время описано около 300 аминокислот, найденных в природе. Однако только 20 аминокислот входят в состав белков» [10], т.е. называются протеиногенными. Они «являются основными составными частями животных и растительных белков, причем их встраивание в молекулу белка регулируется информацией генетического кода» [11].
С 2015 года, в рамках импортозамещения, в России появились специальные высокотехнологичные агрохимикаты собственного производства с торговыми марками «Аминофол» и «Максифол», одним из основных компонентов которых являются свободные протеиногенные L-α-аминокислоты в концентрации более 10%. Это важно, т.к. весь опыт ведущих мировых производителей показал, что более низкая концентрация аминокислот в агрохимикате существенно снижает их эффективность.
Линия отдельных мезо- и микроэлементов Аминофол (Mg, Fe, Mn, Zn, Cu и Mo) базируется на свойстве аминокислот образовывать комплексные соединения с металлами по типу хелатизации. Эти «аминокислотные комплексы металлов имеют октаэдрическое строение, причем два остатка аминокислоты связаны с центральным атомом металла амино- и карбоксильными группами, а свободные координационные связи заняты водой. Особой устойчивостью отличаются комплексы с аминокислотами, имеющими функциональные боковые цепи, как например, гистидин, азот имидазола в котором образует дополнительную (третью) связь с центральным атомом» [12].
В силу того, что в самом растении «в реакциях комплексообразования с микроэлементами принимают участие различные органические соединения, содержащиеся в них: аминокислоты, пептиды, белки» [13] и т.п., комплексы Аминофол не являются чужеродными и полностью усваиваются растением. Высокую степень усвоения элементов питания без риска фитотоксичности обеспечивают: Глютаминовая кислота; Цистеин; Глицин; Гистидин и Лизин, которые образуют хелатные соединения с микроэлементами, а Тирозин, Аргинин, Аланин, Пролин, Серин, Треонин и Валин стимулируют метаболизм, и способствуют лучшему усвоению питательных веществ даже в стрессовых ситуациях. Линия Аминофол обеспечивает эффективное лечение хлорозов при возникновении дефицита, а при своевременном применении отлично удовлетворяет индивидуальные потребности с/х культур в мезо- и микроэлементах.
Аминофол Плюс– специальный антистрессовый агрохимикат с высоким (59%) содержанием свободных протеиногенных аминокислот, применение которого помогает растениям преодолевать стрессовые ситуации, стимулирует метаболизм и усвоение питательных веществ, что существенно повышает урожайность и качество продукции даже в неблагоприятных условиях.
Аминофол NPK – специальный агрохимикат содержащий макроэлементы N-5, P-15, K-10 и 32% аминокислот. Применение Аминофол NPK помогает преодолевать не только стрессовые ситуации, стимулируя метаболизм, рост и развитие растений, но и повышает устойчивость ко многим заболеваниям, т.к. фосфор и калий в нем находятся в форме фосфита калия, который запускает функцию эндогенной защиты растения, стимулируя синтез фитоалексинов (внутренних растительных антибиотиков).
Агрохимикаты «Максифол Радифарм» - стимулятор корнеобразования и антистрессант «Максифол Динамикс», кроме специфических аминокислот содержат концентрированный экстракт морских водорослей Ascophyllum nodosum, который богат натуральными питательными элементами, антиоксидантами, альгиновой кислотой и фитогормонами: цитокинином, ауксином, гиббереллином и глицинбетаином. Эти активные компоненты усиливают устойчивость растений к стрессам различной этиологии, и способствуют повышению количественных и качественных параметров урожайности.
Библиографический список:
1. Mladenova Y.I., Rotcheva S., Vinarova K. 1989. Changes of growth and metabolism of maize seedlings under NaCl stress and interfering effect of Siapton leaf organic fertilizer on the stress responses. In: 20th Ann. ESNA Meeting, Lunteren (NL), Oct. (poster).
2. Soro R. 1985. Recuperacion de los citricos afectados por las heladas. Valencia-fruits, Febr.:9
3. Soro R. 1985. Estimulacion de naranjos. Agricola Vergel, 4: 166-169.
4. Marcucci M.C. 1984. The influence of storage and of organic nutrients on the germination of pollen and fruit set of apple and pear. Acta Hort., 149: 117-122
5. Stoyanov I. 1981. Restoration of maize plants after Magnesium starvation with the help of Magnesium and Siapton. Proc. 3rd Int. Symp. Plant Growth Regulators, Varna, Bulgaria, (B.A.S., Sofia, ed, publ. 1985) pp. 602-606
6. Kovacs A.I., Maini P., De Leonardis A. 1986. Effetto nematostatico del biostimolante Siapton. Atti Giornate Fitopatol., Riva del Garda, CLUEB Ed. BO, pp. 415-424
7. В.Л. Кретович, Биохимия растений. М.«Высшая школа», 1986, стр. 27
8. Х.-Д. Якубке, Х. Ешкайт, Аминокислоты Пептиды Белки. М. «Мир»,1985, стр.9
9. Х.-Д. Якубке, Х. Ешкайт, Аминокислоты Пептиды Белки. М. «Мир» 1985, стр.20
10. В.Л. Кретович, Биохимия растений. М. «Высшая школа», 1986, стр. 25
11. Х.-Д. Якубке, Х. Ешкайт, Аминокислоты Пептиды Белки. М. «Мир», 1985, стр.17
12. Х.-Д. Якубке, Х. Ешкайт, Аминокислоты Пептиды Белки. М. «Мир», 1985, стр.67
13. Н.П. Битюцкий, «Микроэлементы и растение», Изд. С-ПбУ, 1999, стр. 156

Группа компаний «АгроМастер»
Торгово-консультационный офис, склады – г. Тимашевск, ул. Промышленная, 2
тел. (861) 256-81-81, 256-83-83, 256-85-85, факс: 256-82-82; (861-30) 93-150, 93-170
www.agromaster.ru [email protected]
agromaster.ru
Аминокислоты для растений | AgroCounsel
Аминокислоты для растений
Так, например, температура 35 °С может вызвать повреждение гороха, но не приводит к повреждениям в тканях сои. Также очень важно время воздействия стрессовых факторов на растение и его интенсивность. Некоторые могут вызвать негативные последствия уже после нескольких минут воздействия - высокую температуру или избыток или дефицит питательных веществ (проявляется преимущественно через долгое время). Стрессовые факторы, в зависимости от генезиса (базис/причины) действия, можем разделить на абиотические и биотические.Стрессовые факторы■ Абиотические♦ Излучение♦ Температура (слишком высокая, слишком низкая)♦ Вода (засуха, чрезмерная влажность)♦ Газы (дефицит 02 наличие вулканических газов)♦ Минеральные компоненты (дефицит, избыток, окисления, тяжелые металлы, алкализация)♦ Механические (ветер, снежный покров, ледяная корка)■ Биотические♦ Растения (перенасыщение - конкуренция, аллелопатия, растения-паразиты)♦ Микроорганизмы (вирусы ,бактерии,грибы)♦ Животные (грызуны, паразиты, вытаптывание дикими животными)♦ Антропогенные (промышленное загрязнение, пестициды, пожара)
,бактерии,грибы)♦ Животные (грызуны, паразиты, вытаптывание дикими животными)♦ Антропогенные (промышленное загрязнение, пестициды, пожара)
Какие иммунные реакции в растении вызывает стрессовый фактор?Растения, «потревоженные» действием стрессового фактора, сразу активируют защитные механизмы и производят разнообразные субстанции, которые являются возбудителем для продуцирования аминокислот, а впоследствии - белков (отвечают за иммунные процессы). Среди большого количества аминокислот, которые выполняют в растениях различные функции, особого внимания заслуживают две - пролин и глицин. Пролин - способствует повышению иммунитета растений в стрессовых ситуациях и накоплению азота, является прекурсором вкуса, усиливает способность семян к прорастанию, улучшает эффективность фотосинтеза и увеличивает содержание хлорофилла. Его действие заключается также в улучшении генеративного развития растений и их плодородия, он влияет на завязывание плодов, регулирует водообмен в растении. Пролин накапливается в клетках растения, особенно в случае водного и осмотического стрессов. Глицин выполняет роль комплексообразующего вещества (хелата), которое является прекурсором хлорофилла, благодаря чему влияет на увеличение эффективности фотосинтеза, также прекурсором вкуса. Использование препаратов, в своем составе содержат вещества, естественным путем вырабатываются растениями (например, аминокислоты), способствует тому, что не тратится энергия на их выработку - они сразу включаются в метаболизм растения.
Накопление пролина в условиях водного стрессаО возникновении водного стресса говорят как в случае чрезмерной влажности, так и при недостатке соответствующего количества воды в среде роста растений. Это приводит к значительным нарушениям в развитии растений и даже к их отмиранию. Длительный дефицит или избыток воды вызывает нарушение азотного метаболизма, гормонального баланса растения, метаболизма восстановительных компонентов, происходит снижение содержания хлорофилла. В условиях водного стресса растения накапливают большое количество аминокислоты пролина (этому способствует также высокая температура, мороз, засоление, дефицит питательных веществ, заражение патогенами или газовое загрязнение воздуха). Накопление значительного количества пролина в условиях водного стресса способствует эффективному поглощению воды в условиях засухи и препятствует обезвоживанию растений (повышается их засухоустойчивость). Пролин является фактором, стабилизирующим структуру белков и их синтез. Стоит обратить внимание на тот факт, что во время засухи вещества, которые в обычных условиях направляются на рост растений, используется для синтеза пролина. Если эта аминокислота будет введена, например, в виде удобрения вместе с микроэлементами, растение не будет тратить энергию и питательные вещества на ее выработку, а назначит их на другие жизненные процессы.
Физиологическая роль глицинаГлицин является компонентом, так называемых структурных белков, которые высвобождаются в момент возникновения биотических стрессов. Эти белки укрепляют клеточные стенки и ограничивают проникновение патогенов в ткани растения. Глицин играет главную роль в защите клетки от последствий обезвоживания (или избытка соли). Эту аминокислоту используют также для комплексообразования микроэлементов в удобрениях. Благодаря тому, что молекула глицина гораздо меньше, чем молекула хелатирующего веществ, обычно используют (например, ЭДТА), удобрение, которое ее содержит, имеет большую концентрацию микроэлементов. Кроме этого, она является компонентом белков, и поэтому вместе с микроэлементами встраивается в скелет растения. При применение хелатных соединений растение «забирает» лишь элемент питания, а сам хелатор (как лишний балласт) попадает в почву.
Подытоживая, следует отметить, что применение аминокислот в внекорневых удобрениях (хотя и может вызвать немало вопросов и дискуссий), без сомнения, является одним из самых эффективных способов нивелирования влияния вредных условий окружающей среды на сельскохозяйственные растения.
www.agrocounsel.ru
АМИНОКИСЛОТЫ В БОРЬБЕ СО СТРЕССОМ У РАСТЕНИЙ
АМИНОКИСЛОТЫ В БОРЬБЕ СО СТРЕССОМ У РАСТЕНИЙ
Стресс – это реакция растительного организма на воздействие неблагоприятных факторов, которая сопровождается замедлением метаболических процессов, затратами энергии на преодоление негативных факторов внешней среды в ущерб формированию урожая. Основной принцип действия антистрессантов заключается в том, что специально подобранные органические биологически активные вещества запускают, поддерживают и стимулируют физиологические процессы растительного организма, улучшают усвоение питательных элементов и в кратчайшие сроки восстанавливают рост и развитие растений.
В практике сельскохозяйственного производства в условиях наступления ранней весны, когда идет активная вегетация озимых культур, на Северном Кавказе нередко наблюдается возврат холодов в апреле и мае. Это отрицательно сказывается на росте, развитии растений и закладке их потенциальной продуктивности. Низкие положительные температуры в дневное время (+6, +8 0С) и отрицательные в ночное (по прогнозу метеорологов в первой декаде апреля температура ночью будет опускаться до -2, -4 0С) не позволят провести в короткие агротехнические сроки обработку посевов гербицидами. Период борьбы с сорной растительностью может растянуться в крае до 20-25 дней.
Растения озимых культур будут в это время переходить к фазе трубкования и на данном этапе онтогенеза действие гербицидов особенно отрицательно скажется на процессах формирования элементов продуктивности колоса и обмене веществ, поскольку снизится интенсивность фотосинтеза, замедлится синтез важнейших аминокислот (структурных компонентов белков), приостановятся ростовые процессы растений. Стрессовый эффект в зависимости от роста и развития растений, характеру температурного режима в период до и после обработки посевов гербицидами может проявляться в течение 8-15-ти дней.
Применение аминокислот во внекорневых подкормках, хотя и может вызвать некоторые вопросы и дискуссии, без сомнения, в настоящее время является одним из самых эффективных способов нивелирования влияния вредных условий окружающей среды на сельскохозяйственные растения.
Стрессовые факторы в зависимости от генезиса действия, можно разделить на 2 группы:
- Абиотические
- Излучение
- Температура (повышенная, пониженная)
- Вода (засуха, чрезмерная влажность)
- Газы (дефицит СО2, О2)
- Минеральные компоненты (дефицит, избыток, тяжелые металлы)
- Механические (ветер, снежный покров, ледяная корка)
- Биотические
- Микроорганизмы (вирусы, бактерии, грибы)
- Животные (грызуны, паразиты, вытаптывание животными)
- Антропогенные (промышленное загрязнение, пестициды)
- Растения (перенасыщение, конкуренция, аллелопатия, растения-паразиты)
Таким образом, стрессы могут иметь различную природу и воздействовать на растения в коротком или длинном временном промежутке. Стрессовые факторы, обусловленные погодой (например, засуха, чрезмерная влажность, низкая и высокая температуры), могут длиться долгое время или иметь кратковременное действие, оказывая в большинстве случаев негативное влияние на величину и качество урожая.
ПУТИ МЕТАБОЛИЗМА АЗОТА В РАСТЕНИИ И СОХРАНЕНИЕ ЭНЕРГИИ
Как известно, растения синтезируют собственные аминокислоты из неорганического азота. Данный процесс включает превращение нитратов в нитриты и аммонийные соединения и их последующее усвоение в органические молекулы глутаминовой кислоты. Основываясь на этой аминокислоте, растения синтезируют остальные через процесс переаминирования.
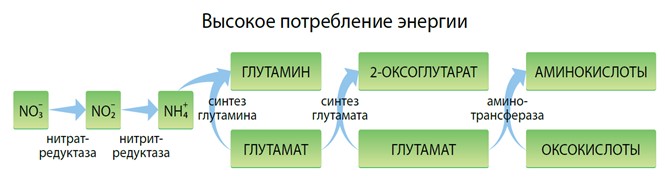
Вышеуказанные метаболические процессы поглощают много энергии, это и является причиной того, что при стрессовых ситуациях растения необходимо напрямую снабжать аминокислотами, высвобождая энергию для прочих физиологических реакций. Доказано, что аминокислоты легко усваиваются на листовом и корневом уровнях и могут напрямую использоваться растениями.
Подвергнутые стрессу растения, сразу активируют защитные механизмы и производят разнообразные субстанции, являющиеся активатором для продуцирования аминокислот, а впоследствии - белков. Использование препаратов, которые в своем составе содержат вещества, активизирующие синтез аминокислот и сами аминокислоты способствует тому, что не тратится энергия на их выработку - они сразу включаются в метаболизм растения.
 Аминокислоты (L-глутаминовая, глицин, лизин и др.), входящие в состав препарата Аминокат 10%, 30% представляют собой готовый строительный материал для биосинтеза молекул белка. Аминокислоты, попадая на лист, практически сразу включаются в обмен веществ без дополнительных затрат времени и энергии на преобразование неорганических веществ в органические. Поступая в растения через листовой аппарат, они в кратчайшие сроки - от нескольких часов – до 2-3-х дней - восстанавливают обмен веществ в растениях после воздействия любых стрессовых факторов (химических, физических, биологических) и активизируют функциональную деятельность всего растительного организма.
Аминокислоты (L-глутаминовая, глицин, лизин и др.), входящие в состав препарата Аминокат 10%, 30% представляют собой готовый строительный материал для биосинтеза молекул белка. Аминокислоты, попадая на лист, практически сразу включаются в обмен веществ без дополнительных затрат времени и энергии на преобразование неорганических веществ в органические. Поступая в растения через листовой аппарат, они в кратчайшие сроки - от нескольких часов – до 2-3-х дней - восстанавливают обмен веществ в растениях после воздействия любых стрессовых факторов (химических, физических, биологических) и активизируют функциональную деятельность всего растительного организма.
Среди большого количества аминокислот, которые выполняют в растениях различные функции, наряду с глутаминовой, особого внимания заслуживают ещё две – пролин и глицин. Пролин – способствует повышению иммунитета растений в стрессовых ситуациях и накоплению азота, усиливает способность семян к прорастанию, улучшает эффективность фотосинтеза и увеличивает содержание хлорофилла. Его действие заключается в улучшении генеративного развития растений и их плодоношения. Пролин влияет на завязывание плодов, регулирует водообмен в растении – он накапливается в клетках растений в случае водного и осмотического стресса. Глицин выполняет роль комплексообразующего вещества (хелата), которое является прекурсором хлорофилла, благодаря чему влияет на увеличение эффективности фотосинтеза.
МЕХАНИЗМ ДЕЙСТВЯ АНТИСТРЕССОВЫХ ПРЕПАРАТОВ НА ОСНОВЕ ДРУГИХ ОРГАНИЧЕСКИХ ВЕЩЕСТВ
В отличие от аминокислот, полимерные соединения различной органической природы - гуматы; Альбит и др. действуют на растения опосредованно и значительно медленнее, поскольку это крупные органические молекулы, которые из-за значительных размеров не способны проникать в клетки растений и принимать непосредственное участие в процессах биосинтеза.
Препараты на основе указанных соединений способны стимулировать обмен веществ у растений после достаточно длительного процесса микробиологического расщепления в течение 8-12 дней, а также за счет воздействия продуктов метаболизма микроорганизмов, оказывающих разлагающее воздействие на указанные вещества.
 Однако длительный процесс разложения сопряжен с более длительным периодом снижения у растений метаболической и фотосинтетической активности. Сложная и разнообразная химическая структура органических полимерных соединений различного происхождения, огромное разнообразие эпифитной микрофлоры, обитающей на поверхности растений, к сожалению, не позволяет точно прогнозировать состав и количество продуктов разложения, а, соответственно, и характер их влияния на растительный организм.
Однако длительный процесс разложения сопряжен с более длительным периодом снижения у растений метаболической и фотосинтетической активности. Сложная и разнообразная химическая структура органических полимерных соединений различного происхождения, огромное разнообразие эпифитной микрофлоры, обитающей на поверхности растений, к сожалению, не позволяет точно прогнозировать состав и количество продуктов разложения, а, соответственно, и характер их влияния на растительный организм.
Так, например, точные молекулярные формулы гуминовых кислот отсутствуют. Все предложенные учеными варианты имеют вид схем, в которых учтены лишь состав и некоторые свойства соединений. Расположение атомов, атомных групп в них определяется гипотетически. Отсутствие у гуминовых кислот формул объясняется их переменным составом и полидисперсностью даже, казалось бы, в однородных органических соединениях. Поэтому обычные методы определения формул биоорганических молекул для гуминовых кислот оказались недейственными.
Действующее вещество препарата Альбит – биополимер поли-бета-гидроксимасляная кислота. Это естественное запасное вещество полезных почвенных бактерий (подобно крахмалу у растений, жиру и гликогену у животных), которое служит субстратом для роста и развития микроорганизмов, но не способно использоваться растениями напрямую в биохимических процессах.

ПРОЦЕССЫ, АКТИВИЗИРУЮЩИЕСЯ В РАСТЕНИЯХ ПОД ДЕЙСТВИЕМ АМИНОКИСЛОТ
Осморегуляция
Свободные аминокислоты, накопленные в клетках во время стрессовых ситуаций, являются результатом синтеза или расщепления белков. Эти аминокислоты вместе с растворенными органическими веществами имеют высокую степень клеточной осморегуляции, которая увеличивает устойчивость клеток к неблагоприятным факторам. Примером является стресс под воздействием засоленности. Накопление в клеточной цитоплазме свободных аминокислот, особенно пролина, позволяет поддерживать низкий уровень ионов, которые могут быть токсичными, например Cl– и Na+.
Регулирование открытия устьиц и активности фотосинтеза
 Закрытие устьиц растений, провоцируемое стрессовыми условиями (высокой температурой, низкой относительной влажностью и т.п.), уменьшает активность фотосинтеза, замедляет все процессы обмена веществ. Экзогенное внесение аминокислот в растения при стрессовых условиях способствует открытию устьиц (рисунок 1, 2). Этот эффект позволяет увеличить удержание влаги в тканях, активизировать фотосинтез, скорректировать скорость испарения и отсрочить увядание.
Закрытие устьиц растений, провоцируемое стрессовыми условиями (высокой температурой, низкой относительной влажностью и т.п.), уменьшает активность фотосинтеза, замедляет все процессы обмена веществ. Экзогенное внесение аминокислот в растения при стрессовых условиях способствует открытию устьиц (рисунок 1, 2). Этот эффект позволяет увеличить удержание влаги в тканях, активизировать фотосинтез, скорректировать скорость испарения и отсрочить увядание.
Антиоксидантная активность
Во время стрессовых ситуаций растения реагируют накоплением веществ с большой оксидативной активностью (О2–, ОН, Н2О2 и О2), что ухудшает структуры клеток. При этих условиях растения используют, среди прочих, азотные соединения, которые действуют как естественные антиоксиданты: аминокислоты, амины, полиамины и определенные ферменты (пероксидазы, каталазы, пероксид дисмутазы). Среди аминокислот антиоксидантными свойствами выделяются аргинин, гистидин, цистеин, триптофан, лизин, метионин и треонин. Применение препарата АМИНОКАТ (10%, 30%) способствует увеличению содержания в растительных тканях антиоксидантов (SOD).
Стимуляция гормональной регуляции
Было замечено, что аминокислоты оказывают влияние на уровни гормонов в растениях. Растения испытывают гормональный дисбаланс из-за внешних факторов, особенно в критические фазы развития. Аминокислоты способны самостоятельно влиять на синтез гормонов, чтобы сбалансировать потребности растения. В то же время комбинированное применение аминокислотных препаратов с гиберрелинами и ауксинами проявляет важный эффект синергизма, когда аминокислоты и гормоны взаимно усиливают действие друг друга. Пуриновые и пиримидиновые основания, присутствующие в составе АМИНОКАТ (10%, 30%) способны стимулировать формирование цитокинов в растении, поскольку они являются прекурсорами этих гормонов. Они воздействуют на деление клеток и воспроизводство, что приводит к усилению роста.
Активизация и защита процессов опыления и плодоношения
Доступность аминокислот в органах цветения является основой успешного прорастания пыльцы и надлежащего и быстрого развития пыльцевой трубки. Развитие пыльцевой трубки происходит благодаря аминокислотам и сахарам, потребляемым пестиком цветка. Из-за этого растению важно иметь в своем распоряжении эти вещества. Особенно способствуют прорастанию пыльцы пролин и глутаминовая кислота.
Опыление и оплодотворение цветов — явление, существенно подвергающееся внутренним и внешним факторам, способным отрицательно повлиять на завязывание и формирование плодов. Было замечено, что внешняя обработка аминокислотами уменьшает время оплодотворения. Сокращение времени оплодотворения увеличивает возможность успешного завязывания плодов при неблагоприятных условиях, особенно экстремальных температурах, как высоких, так и низких. Кроме того, аминокислоты способствуют сбалансированному питанию растения, что увеличивает его способность к удержанию плодов и уменьшает сброс завязи.
Повышение эффективности средств защиты растений
Эффективность средств защиты растений зависит от интенсивности их поглощения через поверхностные структуры растения. Данный процесс, главным образом, идет через клеточные мембраны, а также через каналы и поры растения. Клеточные мембраны содержат высокий процент мембранных белков, способных к изменению проводимости в соответствии со структурой, в которой они находятся. Различные структуры определяются последовательностью аминокислот, с которыми они сочетаются. Обработка аминокислотными препаратами вместе со средствами защиты растений, регуляторами роста, листовыми подкормками, увеличивает проводимость мембраны, повышая скорость усвоения активных веществ.
РЕЗУЛЬТАТЫ ПРИМЕНЕНИЯ ПРЕПАРАТА АМИНОКАТ В ХОЗЯЙСТВАХ КРАЯ
На сегодняшний день одним из наиболее эффективных и востребованных с/х производством антистрессовым препаратом на основе аминокислот является препарат Аминокат 10%, 30%. Его применение в технологии возделывания с/х культур становится обязательным приемом, который прочно входит в практику выращивания различных культур на Ставрополье. Данный препарат является наиболее доступным и относительно недорогим средством управления стрессом у растений.
Высокую эффективность препарат демонстрирует в случае необходимости активизации роста и развития вегетативной массы, предупреждении стрессовых состояний растений (пестицидные обработки), а так же для ликвидации последствия различных повреждающих факторов - повреждение градобоем, избыточной дозой или последействием гербицидов. Даже при крайне угнетенном состояним посевов, обработка данным препаратом позволяет в короткие сроки восстановить обмен веществ растений, получить активно растущий листовой аппарат и достичь уровня продуктивности посева близкого к продуктивности неповрежденных растений. Урожайность сахарной свеклы серьезно поврежденной градобоем после применения аминокислотных препаратов составила 486 ц/га, при дегистии 16,1% (рисунок 4).
В одном из хозяйств Красногвардейского района обследование посевов кукурузы свидетельствовало о серьезном стрессовом состоянии растений, вызванным, по данным специалистов агрослужбы хозяйства, последействием гербицида Фенизан (состав: дикамба+хлорсульфорон). Данный гербицид обладает последействием в том случае, если между применением гербицидов и высевом последующей культуры выпало малое количество осадков. У растений отмечалась задержка роста, слабое развитие корневой системы, хотя по развитию они уже находились в фазе 6-7 листьев (рисунок 3). При проведении функциональной экспресс-диагностики была выявлена очень низкая активность хлоропластов (8,7 единиц) и существенный дефицит цинка и серы.
Применение препарата Аминокат 10% позволило вывести растения кукурузы из состояния стресса, при этом повторное проведение ФЭД через 16 дней после обработки указывало на то, что активность хлоропластов увеличилась до 26 единиц, наблюдался активный прирост биомассы, увеличилась площадь листовой поверхности, цвет растений стал более темно-зеленым, начала интенсивно развиваться корневая система (в том числе и воздушные корни). В результате на полях, где растения были подвержены стрессу, урожайность зерна составила 65 ц/га при средней урожайности кукурузы по хозяйству 80 ц/га.


agro-analiz.com
Аминокислоты для подкормки урожая - АгроПерспектива
В данной статье мы поговорим о том, как включить механизм защиты растений от неблагоприятных факторов.
В течении последних лет на рынке агрохимикатов повышенным интересом пользуются препараты, содержащие аминокислоты. Особенно эффективно применение таких препаратов путем внекорневой подкормки, а также для предпосевной обработки семян и в капельном орошении.
Попробуем рассмотреть роль аминокислот в жизнедеятельности растений и обобщить их функции в растениях. Эти знания помогут агрономам более эффективно использовать такие продукты.
Аминокислоты являются одними из самых активных участников метаболизма. Образуясь в процессе фотосинтеза или в результате синтетической деятельности корней, они в дальнейшем участвуют в самых разнообразных биохимических процессах, в том числе в синтезе белковых и ростовых веществ, от которых, в свою очередь, зависят ростовые процессы.
Аминокислоты являются строительным материалом для формирования (развития и роста) клеток и выполняют много других важных функций в растительных организмах. В условиях стресса и во время роста и развития самих растений аминокислоты играют важную роль. Образование аминокислот в клетках растений – сложный и энергозатратный процесс. Первичным строительным материалом для синтеза аминокислот являются углерод и кислород из воздуха, водород и кислород из воды и азот из почвы.
Аминокислоты, которые синтезируются в растениях, относятся к группам протеиногенных (α-аминокислоты, входящие в состав белков) и непротеиногенных. Протеиногенные аминокислоты – у растений это α-L-аминокислоты. Растения и животные быстрее и лучше усваивают натуральные α-аминокислоты оптически активной L-конфигурации, из которых строятся белки. Такие модификации легко воспринимаются растительным организмом и быстро включаются в метаболизм. D-формы α-аминокислот встречаются в природе сравнительно редко, причем только как продукты обмена веществ низших организмов.
Различия L-аминокислот и D-аминокислот не в химическом составе, а в стереохимическом строении молекул аминокислот (Рис.1.). Однако именно α-L-аминокислоты являются полезными для растений и пригодны для формирования белков.
 L-Аминокислота и D-Аминокислота
L-Аминокислота и D-АминокислотаВ растениях синтезируется 20 протеиногенных аминокислот. Из аминокислот синтезируются белки, которые, в свою очередь, образуют простые ферменты и ферментативные комплексы. Таким образом, они играют важную роль в жизнедеятельности растений. Именно от белков и их качественного и количественного состава в растениях зависит устойчивость растений к стрессовым условиямвегетации и преодоление их последствий. Некоторые аминокислоты важны для гормонального обмена растений, а также как строительный материал образования клеточных стенок у растений (табл. 1).
Таблица 1. Роль протеиногенных аминокислот в стойкостик стрессовым условиям роста, стимулировании обменных процессов и развития растений
| Аминокислота | Функции в растениях |
| Аргинин | Преодоление солевого стресса; развитие корне-вой системы |
| Аспарагиновая кислота | Стимуляция прорастания семян; как строительный материал для других аминокислот |
| Глутаминовая кислота | Синтез хлорофилла; прорастание семян; как строительный материал для других аминокислот |
| Аланин | Синтез хлорофилла; толерантность к засухе; регулирование работы листовых устьиц для оптимизации водного обмена |
| Глицин | Синтез хлорофилла; регулирование работы листовых устьиц, процесса опыления; хелатирование микроэлементов |
| Гистидин | Хелатирующий агент для улучшения поглощения элементов питания; регулирование работы листовых устьиц для оптимизации водного обмена |
| Треонин | Регулирование работы листовых устьиц во время жаркой погоды |
| Пролин | Осмотический протектант, толерантность к жаркой и засушливой погоде, солевому стрессу; регулирование работы листовых устьиц для оптимизации водного обмена; синтез хлорофилла |
| Тирозин | Солевой стресс; толерантность к жаркой погоде; прорастание пыльцы |
| Валин | Толерантность к жаркой и знойной погоде; прорастание семян; процесс опыления |
| Метионин | Стимулирование созревания; регулирование работы листовых устьиц для оптимизации водного обмена |
| Изолейцин | Осмотический протектант, толерантность к жаркой и засушливой погоде, солевому стрессу; прорастание пыльцы; опыление |
| Лейцин | Осмотический протектант, толерантность к жаркой и засушливой погоде, солевому стрессу; прорастание пыльцы |
| Фенилаланин | Синтез гуминовых кислот; процесс опыления; синтез лигнина для укрепления стенок клеток |
| Лизин | Толерантность к засухе; регулирование работы листовых устьиц; синтез хлорофилла; прорастание пыльцы |
| Триптофан | Материал для синтеза гормональных веществ ауксинового типа |
| Серин | Осмотический протектант, толерантность к жаркой и засушливой погоде, солевому стрессу |
| Таурин | Толерантность к засухе и солевому стрессу |
Аминокислоты, которые встречаются в растениях в свободном состоянии, известны как непротеиногенные, то есть, они не входят в состав белков. К непротеиногенным аминокислотам относится большая группа соединений. Их количество более 200, именно они определяют уникальные особенности аминокислотного обмена у растений.
Функции непротеиногенных аминокислот в растениях:
■ транспортная форма азота по растению;■ основная запасная форма азота в семени;■ быстрая и эффективная регуляция количества доступного Nh5+ для растения.
Непротеиногенные аминокислоты легко подвергаются метаболизму и при необходимости являются источником ионов аммония для синтеза белка.
Роль аминокислот в защите культур от стрессов
Вред, нанесенный стрессами культурам, может быть намного большим, чем тот, который они получают от разного рода патогенов (болезней) или вредителей. В то же время, производственный опыт показывает, что современные сорта и гибриды культур интенсивного типа более чувствительны к стрессовым факторам, чем «старые» сорта экстенсивного типа. Возможно это обусловлено тем, что при селекции высокоурожайных сортов культур, теряются гены, которые кодируют культуры на получение высокой урожайности при неблагоприятных «стресс-факторах». Приформировании высокой урожайности культурами активируются их обменные процессы, что затрудняет адаптацию к стрессам.
Главными причинами, которые вызывают физиологическую депрессию растений являются температурный, водный, световой, солевой (почвенный) и пестицидный стрессы, иногда их еще называют «фитострессантами».
Именно ненормированное их влияние на культуры порождает обострение развития болезней, вредителей, проявление некрозов и хлорозов, ухудшение качества, товарности продукции, общее снижение урожайности, а при чрезмерном воздействии приводит к повреждению, а то и к гибели.
Внутреннее проявление стресса «тревоги» на культурах сопровождается снижением метаболических (обменных) процессов, преобладанием распада над синтезом, деградацией белково-синтетического аппарата, старением культур и огромными затратами их энергии на восстановление обмена, что, в конце концов, приводит к снижению продуктивности.
В зависимости от того, в какой фазе роста и развития культуры подвержены влиянию фитостреcсантов, снижение урожайности может составить от 5 до 70%. Однако, научными исследованиями доказано, что обработка культур аминокислотами – пролином, аргинином, аспарагиновой, глутаминовой, янтарной кислотами, а также олигосахаридами, полиаминами, моно- и дисахаридами и другими биологически активными веществами усиливает защиту от фитострессов.
Так, в частности:
Глутаминовая кислота:
хорошо активирует обменные процессы, синтез новых аминокислот, восстанавливает водный баланс культур, улучшает их опыление, является источником синтеза хлорофилла и эффективным комплексоном (хелатирующим агентом). Она также выступает источником энергии и обезвреживает аммиак в растительном организме, отнимая атомы азота в процессе образования другой аминокислоты.
Глицин и лизин:
принимают активное участие в биосинтезе хлорофилла. Глицин также входит в состав листовой ткани, участвует в опылении и завязывании плодов и имеет высокую активность как комплексон, а лизин является предшественником полиаминов, источником азота и улучшает устойчивость культур при стрессах.
Пролин:
улучшает засухоустойчивость и противодействие водному стрессу. Исследования показали, что подкормка пролином может задержать увядание, когда растение подвергается осмотическому стрессу. По результатам исследований, гистидин, пролин и другие аминокислоты являются частью механизма открытия и закрытия устьиц, способствуют удержанию воды и обмену газов.
Все аминокислоты, включая вышеприведенные, являются предшественниками или активаторами фитогормонов. Именно гормоны определяют, какие органы растительного организма нужно формировать: корни, стебли, листья, цветки или плоды. При благоприятных условиях выращивания культур, производство продуктов их фотосинтеза проходит нормально, что обеспечивается фитогормональным балансом.
При фитострессах в организме растений (происходит гидролиз белков в аммоний, который становится токсичным и заставляет растение производить гормон старения – этилен) повышается концентрация гормонов стресса – абсцизовой кислоты и этилена. Как результат – тормозятся ростовые процессы, ухудшается поглощение и утилизация элементов минерального питания, прироста биомассы (растение, не завершив вегетативного развития, переходит к репродуктивной фазе). Продукты фотосинтеза и метаболических реакций перенаправляются на формирование плодов. Гормональный дисбаланс, возникший в результате стресса, приводит к блокированию поступления продуктов фотосинтеза в корневую систему, торможению ее развития и отмиранию. Это приводит к снижению урожайности культур. Контролируя стресс, мы имеем возможность контролировать проблему снижения урожайности.
Важность аминокислот для растений убедительна и несомненна. Благодаря свойствам аминокислот легко растворяться в воде и способности к быстрому проникновению через листовую поверхность в клетки растений, отработаны технологии листовой подкормки растений аминокислотами. Результаты подкормок растений аминокислотными препаратами свидетельствуют о высокой эффективности этих мер для повышения стойкости к жаркой и засушливой погоде, преодоления солевого стресса, улучшения процессов опыления и плодообразования, интенсивности процесса фотосинтеза, поддержания гормонального баланса, улучшения азотного обмена в растениях и прочее.
agroperspectiva.com.ua
Рост растений и аминокислотный обмен
Аминокислоты являются основными исходными веществами, обеспечивающими синтез белка растущих органов и частей растений.
Биосинтез белка является одним из важнейших внутренних условий роста, хотя, по-видимому, будет правильным утверждение, что ни один из физиологических процессов нельзя назвать основным или более важным, ибо все они внутренне неразделимы и могут быть правильно поняты только при рассмотрении их в неразрывном взаимодействии.
Физиология растений ныне опирается на изучение химического, физико-химического и физического аспектов жизнедеятельности организма; возникли самостоятельные науки — биохимия, биофизика, биофизическая химия, посвященные этим аспектам. Однако они являются базисом физиологии, ибо последняя может преуспевать в познании явлений жизнедеятельности лишь при вскрытии закономерностей взаимосвязи химических и физических процессов. Поскольку «организм есть, несомненно, высшее единство, связывающее в себе в одно целое механику, физику и химию» изучение физико-химических процессов в организме приближает пас к пониманию специфических закономерностей организма, являющихся результатом необычайной сложности и замечательной организованности всей совокупности реакций, составляющих процессы жизнедеятельности.
Как нам кажется, при рассмотрении основного интегрального процесса жизнедеятельности организма — роста — необходимо ближе подходить к характеристике отдельных сторон обмена веществ растущих органов с тем, чтобы когда-нибудь возникли реальные возможности для создания полной картины единой сложной сети реакций, в которой переплелись сотни химических реакций со строго упорядоченной их последовательностью в пространстве и времени.
Исследуя растворимые аминокислоты и амиды в органах растений в процессе их роста, мы, конечно, учитывали исключительно изменчивый характер их содержания. На концентрацию и качественный состав аминокислот влияют как внутренние процессы, определяющие течение азотного обмена в широком смысле слова (обмен белков, нуклеиновых кислот, пигментов, ростовых веществ и т. д.), так и внешние факторы. Теперь известно более 100 аминокислот, из которых только 22 встречаются в растениях в составе белков. Многие из аминокислот обнаруживаются в низких концентрациях, а другие, по-видимому, существующими методами пока не удается уловить. Хотя не все аминокислоты участвуют в синтезе белка и потому, казалось бы, непосредственно не влияют на рост растений, следует сказать, что свободные аминокислоты, не входящие в состав белков, необходимы для жизнедеятельности организмов и играют важную роль во многих обменных процессах, в том числе в процессах обмена с белковыми аминокислотами, с органическими кислотами, с синтезом физиологически активных веществ. Иначе говоря, непосредственно или опосредствованно все свободные аминокислоты участвуют в метаболизме ростовых процессов.
По составу и количеству аминокислот не всегда представляется возможным судить об их роли в отдельных процессах жизнедеятельности растений. Состав аминокислот, конечно, тесно связан с процессами роста и развития, поскольку последние вызывают изменения в оттоке аминокислот из листа и их использовании в синтезе белка. Однако эти связи аминокислотного обмена с ростом растений нередко оказываются мало заметными или их трудно установить, так как во время вегетации по самым разнообразным причинам меняется состав веществ, в том числе и аминокислот. Изменения в составе свободных и белковых аминокислот могут быть связаны с процессом фотосинтеза, но могут явиться и результатом передвижения по растению, когда, например, аминокислоты поступают в листья из корней, где они образуются в значительных количествах и в большом разнообразии.
При оценке синтетической деятельности корней по веществам, содержащимся в пасоке растений, следует иметь в виду то обстоятельство, что в корневой системе, изолированной от надземных органов, тотчас после удаления последних изменяется направленность биохимических процессов. Так, Э. С. Авунджян в пасоке гречихи, собранной за первые сутки после декапитации растений, обнаружил лишь две лабильно связанные аминокислоты (глицин и α-аланин), что указывает, очевидно, на наличие соответствующего дипептида. Однако уже начиная со вторых суток картина совершенно меняется: появляются новые аминокислоты пептидной фракции, набор которых достигает максимума на четвертые сутки, затем уменьшается, а на шестые сутки опять возрастает. Интересно отметить, что количество глютаминовой кислоты составляет 26—40%, аргинина— 7—23% общей суммы всех аминокислот, находящихся в пептидной связи. Аминокислотный состав трудно гидролизуемой белковой фракции пасоки более устойчив. На этом основании можно предполагать, что в составе пасоки корни транспортируют к надземным органам белковые вещества в основном определенного аминокислотного состава. При изучении аминокислот с применением С14 необходимо учитывать, что они могут сразу же после образования включаться в новые соединения и потому не обнаруживаться среди свободных аминокислот на хроматограммах.
Аминокислоты являются одними из самых активных участников метаболизма. Образуясь в процессе фотосинтеза или в результате синтетической деятельности корней, они в дальнейшем участвуют в самых разнообразных биохимических процессах, в том числе в синтезе белковых и ростовых веществ, от которых, в свою очередь, зависят ростовые процессы. Установлено, что аланин и серии у Chlorella pyrenoidosa образуются в результате дальнейших превращений первого стабильного продукта фотосинтеза — фоофоглицериновой кислоты. Ранее Кальвин установил, что лист ячменя при экспозиции с С14 на свету за 60 сек образует аланин, глицин, серии, аспарагиновую кислоту; лист герани образует только аланин, серии и глицин.
Содержание суммы и отдельных аминокислот может быть показателем интенсивности ростовых процессов. В молодых листьях фасоли и бобов, например, количественно преобладают амиды и аспарагиновая кислота: они могут составлять 50—60% всего аминного азота. Однако по мере роста листьев исчезает аспарагин, начинают преобладать аланин, серии, глютаминовая кислота. В зрелых листьях накапливается γ-аминомасляная кислота, что, возможно, связано с замедлением и прекращением ростовых процессов через торможение новообразования белка.
О. И. Романовская проследила сезонные изменения аминокислот в тканях одиночных побегов яблони и сливы в связи с их ростовой активностью. Оказалось, что в начале роста (май) в коре яблони имеется 9, а сливы —13 аминокислот.
В клеточном соке коры и древесины много аспарагина. В период наиболее интенсивных ростовых процессов и синтеза белков (июнь) содержание свободных аминокислот снижается. Исчезают или обнаруживаются в меньших количествах аргинин, пролин, цистин, гистидин и др. Содержание аспарагина остается высоким. Период глубокого покоя (октябрь—декабрь) характеризуется резким уменьшением количества или полным исчезновением аспарагина, накоплением аргинина, треонина, пролина и других аминокислот. При переходе к «вынужденному» покою резко увеличивается количество всех свободных аминокислот.
По-видимому, в этот период происходят сложные превращения белковых веществ, высвобождение азота в виде свободных аминокислот и подготовка к началу ростовых процессов.
Аминокислоты, поступая к меристем этическим клеткам почек яблони, стимулируют формирование качественно новых органов — цветков. Аминокислотный состав листьев плодоносных и неплодоносных кольчаток изменяется как в различные часы суток, так и в зависимости от состояния кольчаток, от которых были взяты листья для анализа.
Если учесть факты образования ростовых веществ типа гетероауксина и фенилпропионовой кислоты при дезаминировании триптофана, образования β-аланина при декарбоксилировании аспарагиновой кислоты, а также действие, оказываемое этими веществами на растительные клетки, то станет ясным, что аминокислоты и продукты их окислительно-восстановительных превращений не только играют роль питательных веществ в растении, но и являются физиологически активными веществами, регулирующими жизнедеятельность протоплазмы и рост растительных тканей.
Имеются указания на то, что гиббереллин оказывает влияние на аминокислотный обмен в ходе прорастания семян. Уже в набухших семенах конопли, обработанных гибберелловой кислотой, увеличивается количество аланина, причем это увеличение происходит параллельно с концентрацией ГК, что может рассматриваться как показатель наличия близкой связи между гиббереллином и синтезом аланина. Обнаружены качественные и количественные изменения аминокислот в проростках (длина зародышей — до 0,5 см), вызванные влиянием ГК.
Сравнение меристемы верхушек побегов люпина, сирени и папоротника с тканями прилегающих, более зрелых частей этих растений показывает, что основные аминокислоты — аргинин и лизин — более заметны в области меристемы. Глютамин, глютаминовая и γ-аминомасляная кислоты также находятся в меристеме в больших количествах, чем в других тканях.
При попытке по количественному содержанию того или иного вещества судить о его участии в физиолого-биохимических процессах необходимо учитывать то весьма важное обстоятельство, что это содержание чрезвычайно изменчиво и зависит не только от скорости синтеза, но и от скорости оттока веществ, который происходит в растении избирательно. Избирательный характер передвижения веществ из органа в орган показан, в частности, исследованиями по изучению поступления ассимилятов в проводящие пути листьев ревеня. Сахара и аминокислоты, образовавшиеся в мезофилле листа ревеня в процессе фотосинтеза, протекавшего в присутствии C14O2, уже в первые минуты поступают в проводящую систему. Скорость поступления этих веществ значительно превосходит таковую органических кислот и других соединений.
Однако не все аминокислоты поступают в проводящие пути с одинаковой скоростью. Легче других проникает треонин, хотя при фотосинтезе в наибольших количествах образуются аланин и серин. Эти последние все же относительно быстро поступают в русло движения веществ, тогда как аспарагиновая кислота, пролин, а иногда и γ-аминомасляная кислота участвуют в оттоке довольно слабо.
Таким образом, отдельные вещества оттекают из одних органов в другие с различной скоростью, непропорциональной их концентрации. Это указывает на метаболический характер саморегулирующегося механизма передвижения веществ по растению. Только скорость протекания биохимических превращений, в свою очередь зависящая от множества внешних и внутренних факторов, определяет интенсивность поступления и оттока веществ в различных органах растений. А это означает, что все наши заключения относительно закономерностей, характеризующих особенности протекания метаболических процессов по содержанию отдельных веществ в определенный момент, в значительной степени являются условными, так как при современном состоянии знаний об отдельных звеньях обмена веществ мы не всегда имеем возможность опереться на достаточно обширные сведения, объясняющие причинно-следственные зависимости между количеством вещества, скоростью его притока—оттока и превращения и другими сторонами процесса, одним из участников которого является данное вещество.
По-видимому, все аминокислоты прямо или опосредствованно связаны с биохимическими процессами, лежащими в основе роста растений.
Мы имеем в виду не только те аминокислоты, которые участвуют в создании белковой молекулы, но и те, которые обнаруживаются лишь в свободном состоянии и количество которых в три раза выше первых.
Из давно открытых аминокислот при нормальных условиях развития во всех растениях широко распространены и содержатся в значительных количествах γ-аминомасляная кислота, пипеколиновая кислота, β-аланин, α-аминоадипиновая кислота. Остальные новые аминокислоты содержатся во многих растениях либо в незначительных количествах, либо вообще отсутствуют.
Ассимиляция аммиака растениями и включение его в различные органические азотистые соединения осуществляется путем синтеза глютаминовой кислоты из α-кетоглютарата и аммония под действием глютамикодегидразы и через образование глютамина из глютаминовой кислоты и аммония при участии глютаминсинтетазы.
Говоря о превращении тех или иных веществ в растении, необходимо помнить, что одно и то же вещество иногда одновременно, иногда последовательно участвует в разных звеньях обмена веществ. В. Л. Кретович и В. И. Яковлева отмечали, в частности, что α-кетоглютаровая кислота, вводимая в живую растительную ткань, подвергается превращениям в цикле Кребса, аминируется аммонием и вступает в реакции переаминирования с аминокислотами и амидами, в первую очередь с глютамином.
Глютаминовая кислота амидируется аммонием, декарбоксилируется с образованием γ-аминомасляной кислоты и принимает участие в реакциях переаминирования. Глютамин используется в реакциях переаминирования с кетокислотами и может вместе с тем дезаминироваться под действием глютаминазы.
Путь превращения метаболита, его содержание в растении зависят от соотношения скоростей реакций и условий выращивания растений. Учет всех этих обстоятельств крайне важен при оценке роли тех или иных веществ, в данном случае аминокислот, в ростовых процессах и их отдельных звеньях.
Тесные взаимосвязи между ростовыми процессами и аминокислотным составом растений устанавливаются уже в период начального роста, когда метаболизм поступающих из эндосперма азотистых соединений глубоко воздействует на рост проростка, но сам метаболизм подвергается действию со стороны роста и развития системы. Изменяется качественный состав аминокислот в последовательных отрезках корня, и в расчете на клетку содержание аминокислот к концу роста клеток увеличивается в 10 раз.
Количественная бумажная хроматография позволила установить значительные качественные и количественные изменения метаболизма азотистых соединений в верхушках побега некоторых растений (люпина, сирени и папоротника) в зависимости от зон роста. Обнаружено, что у люпина аспарагин составляет 68% азотнорастворимой фракции верхушечной меристемы. Аргинин и лизин более заметны в областях меристемы по сравнению с тканями прилежащих и более зрелых частей.
Из других аминокислот наибольшее внимание многих исследователей привлек триптофан, который считается предшественником гетероауксина в растении.
Предполагают, что биосинтез этой важной аминокислоты осуществляется у разных организмов по-разному. О. Л. Поляновский подтвердил возможность образования триптофана из индолпировиноградной кислоты и связующую роль последней между триптофаном и индол ил уксусной кислотой. Он показал, что синтез триптофана из индолпировиноградной кислоты идет в растении двумя путями: переаминированием аминокислот и другим путем — с участием АТФ и Nh4, что скорее всего связано с образованием глютамина, который затем переаминируется с иидолпировиноградной кислотой (ИП).
При исследовании синтеза триптофана из ИП в дельных проростках и колосьях пшеницы было показано, что синтез триптофана в течение 10 час идет линейно во времени. Это указывает на то, что синтезировавшийся триптофан медленно расходуется растительным организмом на синтез белка. По способности к образованию триптофана в проростках пшеницы исследованные его предшественники могут быть расположены в следующий ряд: антраниловая кислота < индол < β-3-индолпировиноградная кислота. Синтез триптофана зависит от освещения растения. В этиолированных проростках и корнях гороха синтез триптофана из индола и серина шел слабо, в зеленых проростках — сильнее, а в зеленых верхушках проростков — наиболее интенсивно. Синтез триптофана из антраниловой кислоты, индола и ИП на свету шел значительно интенсивнее, чем в темноте. При насасывании в проростки на свету ИП и триптофана наблюдается образование ряда аминокислот: лейцина, валина, треонина и γ-аминомасляной кислоты.
Существенное значение для понимания роли свободных аминокислот в ростовых процессах имеют исследования по изучению их качественного и количественного состава в молодых растущих растениях на разных этапах роста. Мы считали, что показательность таких исследований возрастет, если удастся получить данные, устанавливающие закономерности между интенсивностью линейного прироста растений и содержанием аминокислот. В связи с этим в опыты были взяты высокорослый (Капитал) и низкорослый (Комсомолец) сорта гороха. Начиная с 9-дневного возраста количественно определяли аспарагиновую и глютаминовую кислоты, α-аланин, аргинин, метионин, лизин, аспарагин, цистеин, лейцин. Качественно были также определены валин, триптофан, гистидин, пролин, серии, содержание которых было слишком ничтожным (кроме пролина), чтобы определить их фотоэлектроколориметрически.
Количественное определение свободных аминокислот проводилось методом хроматографии на бумаге. Четкое разделение пятен достигалось применением одномерной хроматограммы, но при многократном пропускании одной и той же смеси растворителя (н-бутанол : уксусная кислота : вода в соотношении 4:1:1) с подсушиванием хроматограмм на воздухе. Элюцию окрашенных нингидрином пятен и измерение интенсивности окраски полученных растворов проводили по методу, описанному Т. Ф. Андреевой и О. П. Осиповой. Оптическую плотность окрашенных растворов определяли на ФЭК-М, используя зеленый светофильтр против элюата из бумаги контрольного участка хроматограммы (фона).
Затем вычисляли среднее арифметическое значение оптической плотности для каждой аминокислоты стандартной и опытной смеси. Расчет концентрации аминокислот производили по отношению среднего значения оптической плотности соответствующей аминокислоты известной концентрации в стандартной смеси к среднему значению оптической плотности той же аминокислоты в исследуемой смеси.
В наиболее значительных количествах в растениях гороха в первый месяц их роста обнаруживаются аспарагиновая и глютаминовая кислоты. В меньшем количестве, но превышающем содержание остальных аминокислот (по крайней мере, начиная с 16-дневного возраста), найден аланин. Эти три аминокислоты наиболее быстро обрадуются в растениях. Высокая начальная активность этих соединений предшествует появлению меченого углерода в других аминокислотах, в связи с чем предполагается, что аланин, глютаминовая и аспарагиновая кислоты являются предшественниками других аминокислот. Переаминирование как общая реакция способствует образованию других аминокислот путем переноса аминогрупп от дикарбоновых аминокислот, которые, по-видимому, играют регулирующую роль в синтезе белка.
Большинство других аминокислот (аргинин, цистсин, серии, метионин) остаются, по нашим данным, на одном уровне в течение первых недель роста. Имеет место постепенное нарастание количества триптофана.
Указанные закономерности в количественном и качественном содержаний аминокислот одинаково характерны для растений как высокорослого, так и низкорослого сортов. Вместе с тем между ними обнаружены также и определенные различия по содержанию аминокислот. Высокорослый сорт отличается более высоким содержанием аспарагиновой и глютаминовой кислот и аланина. Особенно следует отметить, что содержание триптофана, который, безусловно, связан с биосинтезом никотиновой, идолилуксусной и других биоактивных производных индола, достигает максимального значения у растений высокорослого сорта. Все эти факты, на наш взгляд, убедительно свидетельствуют о наличии прямой зависимости между содержанием отдельных аминокислот и интенсивностью роста растений. Эта зависимость, надо полагать, осуществляется через участие аминокислот в регуляции синтеза белка как необходимого условия ростовой активности растения.
Важные закономерности удается установить при исследовании количественных показателей образования некоторых аминокислот в отдельных междоузлиях и растущих листьях растения. В этой связи следует упомянуть работу Л. Д. Прусаковой, посвященную особенностям роста листа и некоторым физиолого-биохимическим аспектам ростовых процессов в нем. Автор подчеркивает необходимость детального изучения биосинтетической способности листа при обязательном учете состояния его клеток. Преодолев определенные методические трудности выделения клеток листа, находящихся в той или иной фазе роста, она смогла установить, что начиная с момента заложения листа яровой пшеницы до того времени, когда он достигнет 7 мм длины, все его клетки находятся в состоянии деления, о чем свидетельствуют митозы и амитозы. К этому времени появляется первый проводящий сосуд, лист длиной свыше 7 мм и проявляет признаки дифференциации (в зоне 1 мм начиная с верхушки деление клеток прекращается), из клеток образуются устьица, волоски, к ним прокладывают путь проводящие сосуды. Одновременно в большей части листа происходит интенсивное деление клеток во всех направлениях до тех пор, пока лист не достигнет 10 мм. Хотя от момента заложения до 10 мм длины клетки листа успевают пройти в основном фазу деления, все же одновременно происходят процессы растяжения и дифференциации клеток. В эту раннюю фазу развития листа заметных отличий в растениях как при оптимальном водоснабжении, так и недостаточном нет, за исключением большего размера клеток у первых.
В период удлинения листа от 10 до 50 мм интенсивным делением охвачены не все клетки и не во всех направлениях — намечаются делящиеся ряды клеток вдоль листа, т. е. дифференцируется сама меристем этическая ткань. Эта фаза роста листа наряду с интенсивным делением клеток включает их растяжение, а также и дифференциацию. В условиях недостаточного водоснабжения уменьшается интенсивность деления клеток и величина растяжения; наряду с этим быстрее идет дифференциация клеток на ткани.
Дифференциация включает в себя не только образование выполняющих определенную функцию тканей в листе (проводящую, ассимиляционную, устьица, эпидермис и др.), но и такие процессы, как утолщение первичных стенок, лигнификация их и др. При задержке роста листа, вызванной недостатком воды, образующиеся в листе ассимиляты, не используемые на ростовые процессы, вовлекаются в процессы дифференциации и тем самым стимулируют последние. При оптимальных условиях водного режима этот процесс идет медленнее, сопутствуя более длительному делению и растяжению клеток листа.
Лист, находящийся в фазе деления и растяжения клеток, содержал при оптимальном водоснабжении значительно больше триптофана, чем выросший. Кроме того, в растущем листе обнаруживалось большее содержание тирозина. Недостаточное водоснабжение листа в период его интенсивного роста сильно снижало в нем содержание триптофана. Тирозин, по-видимому, тоже принимает непосредственное участие в росте. Пролин (возможно, оксипролин) появляется тогда, когда заканчиваются процессы роста, причем его количество при недостаточном водоснабжении в несколько раз больше по сравнению с количеством при оптимальном водоснабжении. В растущем листе обнаруживается и большее количество аланина.
Опыты по синтезу триптофана из его возможных предшественников (индола и серина) показали, что растущий лист за 2 час инкубации синтезировал больше триптофана, чем лист, закончивший рост.
Результаты наших исследований также показали, что содержание аминокислот заметно коррелирует с состоянием ростовой активности отдельных органов и частей растений. Верхние междоузлия (четвертое и пятое) десятидневных проростков гороха содержат большие количества аспарагиновой, глютаминовой кислот и аргинина у сорта Капитал по сравнению с теми же междоузлиями низкорослого сорта Комсомолец. Первые две из названных аминокислот имеют максимальное значение во втором и третьем листках, находящихся на начальных фазах развития. Более высокое содержание аминокислот в верхушке растений сорта Комсомолец по сравнению с таковой у сорта Капитал следует объяснить тем, что вместе, с верхушкой для анализа брали зачатки третьего листа, которые, по-видимому, содержат много аминокислот, по крайней мере дикарбоновых, как это имеет место и в молодых листьях растений сорта Капитал.
Физиологически активные вещества (гиббереллин и гетероауксин) влияют на ростовые процессы растений через изменение обмена веществ, в том числе аминокислотного обмена. Эти изменения сводятся в основном к уменьшению количества свободных аминокислот без заметных качественных изменений аминокислотного состава. Так, обработка гиббереллином несколько снижает содержание серина, глютамина, тирозина и приводит ‘К исчезновению гистидина в листьях кукурузы. В корнях это уменьшение содержания аминокислот распространяется на аспарагиновую и глютаминовую кислоты, серии, треонин, α-аланин, тирозин, триптофан, валин, лейцин. Гистидин в корнях обработанных гиббереллином растений не обнаруживается совсем. Количество валина, лейцина и лизина возрастает в листьях, а последний появляется и в корнях.
Гетероауксин снижает содержание серина, глютаминовой кислоты, гистидина и тирозина в листьях и гистидина, аспарагина, глютаминовой кислоты, треонина, тирозина, триптофана, валина и лейцина — в корнях. В листьях совсем исчезает аспарагин и увеличивается количество лизина, триптофана и валина. Больше становится серина и α-аланина в корнях.
При обработке гиббереллином раздельно или совместно уменьшается количество таких аминокислот, как глютаминовая и триптофан, играющих важнейшую роль в ростовых процессах, однако при двойной норме азотного питания гиббереллин повышает уровень не только глютаминовой кислоты, но и аспарагиновой, а также α-аланина. Мы уже отмечали, что патологические отклонения в росте растений, вызванные гиббереллином, могут быть результатом нарушения условий азотного питания.
Изучение влияния гиббереллина на обмен аминокислот в набухших семенах и проростках конопли показало, что в этих условиях имеют место изменения в качестве и количестве аминокислот. Особого внимания заслуживает увеличение содержания аланина, наблюдаемое в набухших семенах после внесения гибберелловой кислоты. Возрастание количества аланина параллельно с концентрацией гиббереллина указывает на коррелятивные отношения между гиббереллином и синтезом аланина.
В наших опытах изменения в содержании аминокислот в растущих растениях гороха в зависимости от влияния гиббереллина носили настолько скачкообразный характер, что трудно установить какую-либо определенную закономерность, кроме, возможно, той, что образование, приток и передвижение, взаимопревращения и использование аминокислот в биосинтезе белков носят неравномерный характер и потому не всегда удается обнаружить прямую зависимость между воздействием стимулятора роста и количественным содержанием аминокислот. Имеют значение также не только видовые, но и сортовые отличия.
Аминокислоты участвуют в важнейших для роста растений биохимических реакциях в тесной взаимосвязи с другими соединениями, от содержания которых в значительной степени зависит качественный и количественный состав аминокислот и их превращение, в частности, в белковом обмене. Особенно тесно связан аминокислотный обмен с углеводным обменом. От содержания углеводов зависит в первую очередь превращение свободного аммиака, который может образовываться в растениях при дезаминировании аминокислот. При наличии достаточного запаса углеводов аммиак вступает в реакцию прямого аминирования с кетокислотами, образующимися из углеводов. Однако и при недостатке углеводов или избыточном поступлении аммонийных солей в растении не может накапливаться аммиак. В этом случае последний обезвреживается путем превращения в амиды — аспарагин и глютамин.
Амидообразование непосредственно связано с ростом растений. Эта связь осуществляется двумя путями. Во-первых, накопившиеся в неблагоприятных для азотно-углеводного обмена условиях амиды при нормализации условий используются для синтеза белков в молодых тканях растений, как это имеет место при освещении этиолированных проростков бобовых, когда образующиеся в процессе фотосинтеза углеводы создают возможность для превращения аспарагина и участия его продуктов в белковом обмене. Кроме того, аспарагин и глютамин играют не менее важную роль в качестве резерва дикарбоновых кислот, необходимых для осуществления реакций ферментативного переаминирования. Поскольку последние, а также дикарбоновые аминокислоты имеют чрезвычайно важное значение для синтеза и превращения аминокислот в растительном организме и, в конечном счете, для анаболических процессов, становится еще более ясной роль амидов в росте растений.
Для роста растений важное значение имеют процессы декарбоксилирования аминокислот. Образующиеся при этом промежуточные соединения участвуют вновь в синтезе аминокислот и стимуляторов роста. Так, триптофан может синтезироваться из индола и серина. Образовавшийся при декарбоксилировании из аспарагиновой кислоты α-аланин в дальнейшем может быть использован при синтезе белка.
Белковый и аминокислотный обмены весьма тесно взаимодействуют с обменом витаминов и ростовых веществ. Фосфорилированное производное витамина B6 — пиридоксальфосфат — входит в состав активных групп аминотрансфераз и декарбоксилаз аминокислот. Имеются предположения, что никотиновая кислота образуется в результате превращений триптофана. Уже отмечалось, что β-индолилуксусная кислота своим наиболее ранним предшественником имеет триптофан.
Таким образом, можно считать установленной прямую или опосредствованную взаимосвязь аминокислотного обмена с ростовыми процессами. Однако экспериментальные данные, характеризующие эту связь, весьма скудны. Необходимы углубленные исследования количественного и качественного состава аминокислот в зонах роста, в меристемах, а также в связи с интенсивностью роста и особенностями белкового и нуклеинового обменов. Успехи биохимии в изучении аминокислотного обмена позволяют в ближайшие годы решить эти вопросы.
Если вы нашли ошибку, пожалуйста, выделите фрагмент текста и нажмите Ctrl+Enter.
www.activestudy.info
РАСТЕНИЕВОДСТВО. Аминокислоты для растений |
АМИНОКИСЛОТЫ ДЛЯ РАСТЕНИЙ
ВОЗДЕЙСТВИЕ СТРЕССОВЫХ ФАКТОРОВ (ТЕМПЕРАТУРА И ВРЕМЯ) НА РАСТЕНИЕ И ЕГО ИНТЕНСИВНОСТЬ ОЧЕНЬ ВАЖНО. НЕКОТОРЫЕ МОГУТ ВЫЗВАТЬ НЕГАТИВНЫЕ ПОСЛЕДСТВИЯ УЖЕ ПОСЛЕ НЕСКОЛЬКИХ МИНУТ ВОЗДЕЙСТВИЯ — ВЫСОКУЮ ТЕМПЕРАТУРУ ИЛИ ИЗБЫТОК ИЛИ ДЕФИЦИТ ПИТАТЕЛЬНЫХ ВЕЩЕСТВ (ПРОЯВЛЯЕТСЯ ПРЕИМУЩЕСТВЕННО ЧЕРЕЗ ДОЛГОЕ ВРЕМЯ).
Стрессовые факторы, в зависимости от генезиса (базис/причины) действия, можем разделить на абиотические и биотические.
Стрессовые факторы.
Абиотические
- Излучение
- Температура (слишком высокая, слишком низкая)
- Вода (засуха, чрезмерная влажность)
- Газы (дефицит 02 наличие вулканических газов)
- Минеральные компоненты (дефицит, избыток, окисления, тяжелые металлы, алкализация)
Механические (ветер, снежный покров, ледяная корка)
Биотические
- Растения (перенасыщение — конкуренция, аллелопатия, растения-паразиты)
- Микроорганизмы (вирусы, бактерии, грибы)
- Животные (грызуны, паразиты, вытаптывание дикими животными)
- Антропогенные (промышленное загрязнение, пестициды, пожара)
Итак, как видим, стрессовые факторы, в зависимости от «виновника», могут иметь разную причину и воздействовать на растения в коротком или длинном временном промежутке. Стрессовые факторы, обусловленные погодой (например, засуха, чрезмерная влажность, низкая и высокая температуры), могут длиться долгое время или иметь кратковременное действие, оказывая в большинстве случаев негативное влияние на величину и качество урожая.
Следующими «преступниками», которые вызывают стрессы, являются болезни, вредители, сорняки и т.д. Любое легкомыслие фермера в борьбе с вышеупомянутыми факторами может привести к существенному уменьшению урожая.
Какие иммунные реакции в растении вызывает стрессовый фактор?
Растения, «потревоженные» действием стрессового фактора, сразу активируют защитные механизмы и производят разнообразные субстанции, которые являются возбудителем для продуцирования аминокислот, а впоследствии — белков (отвечают за иммунные процессы). Среди большого количества аминокислот, которые выполняют в растениях различные функции, особого внимания заслуживают две — пролин и глицин.
Пролин — способствует повышению иммунитета растений в стрессовых ситуациях и накоплению азота, является прекурсором вкуса, усиливает способность семян к прорастанию, улучшает эффективность фотосинтеза и увеличивает содержание хлорофилла. Его действие заключается также в улучшении генеративного развития растений и их плодородия, он влияет на завязывание плодов, регулирует водообмен в растении.
Пролин накапливается в клетках растения, особенно в случае водного и осмотического стрессов.
Глицин выполняет роль комплексообразующего вещества (хелата), которое является прекурсором хлорофилла, благодаря чему влияет на увеличение эффективности фотосинтеза, также прекурсором вкуса. Использование препаратов, в своем составе содержат вещества, естественным путем вырабатываются растениями (например, аминокислоты), способствует тому, что не тратится энергия на их выработку — они сразу включаются в метаболизм растения.
Накопление пролина в условиях водного стресса.
О возникновении водного стресса говорят как в случае чрезмерной влажности, так и при недостатке соответствующего количества воды в среде роста растений. Это приводит к значительным нарушениям в развитии растений и даже к их отмиранию. Длительный дефицит или избыток воды вызывает нарушение азотного метаболизма, гормонального баланса растения, метаболизма восстановительных компонентов, происходит снижение содержания хлорофилла.
В условиях водного стресса растения накапливают большое количество аминокислоты пролина (этому способствует также высокая температура, мороз, засоление, дефицит питательных веществ, заражение патогенами или газовое загрязнение воздуха). Накопление значительного количества пролина в условиях водного стресса способствует эффективному поглощению воды в условиях засухи и препятствует обезвоживанию растений (повышается их засухоустойчивость).
Пролин является фактором, стабилизирующим структуру белков и их синтез. Стоит обратить внимание на тот факт, что во время засухи вещества, которые в обычных условиях направляются на рост растений, используется для синтеза пролина. Если эта аминокислота будет введена, например, в виде удобрения вместе с микроэлементами, растение не будет тратить энергию и питательные вещества на ее выработку, а назначит их на другие жизненные процессы.
Физиологическая роль глицина.
Глицин является компонентом, так называемых структурных белков, которые высвобождаются в момент возникновения биотических стрессов. Эти белки укрепляют клеточные стенки и ограничивают проникновение патогенов в ткани растения. Глицин играет главную роль в защите клетки от последствий обезвоживания (или избытка соли).
Эту аминокислоту используют также для комплексообразования микроэлементов в удобрениях. Благодаря тому, что молекула глицина гораздо меньше, чем молекула хелатирующего веществ, обычно используют (например, ЭДТА), удобрение, которое ее содержит, имеет большую концентрацию микроэлементов.
Кроме этого, она является компонентом белков, и поэтому вместе с микроэлементами встраивается в скелет растения. При применение хелатных соединений растение «забирает» лишь элемент питания, а сам хелатор (как лишний балласт) попадает в почву.
Подытоживая, следует отметить, что применение аминокислот в внекорневых удобрениях (хотя и может вызвать немало вопросов и дискуссий), без сомнения, является одним из самых эффективных способов нивелирования влияния вредных условий окружающей среды на сельскохозяйственные растения.
www.agrocounsel.ru
yariks.info










