Химический состав и молекулярная организация клеточной оболочки. Основное скелетное вещество оболочки растений
Химический состав и молекулярная организация клеточной оболочки.
КЛЕТОЧНАЯ ОБОЛОЧКА
Наличие клеточной целлюлозной оболочки хорошо отличает растительные клетки от животных. Клеточная оболочка ограничивает размер протопласта, защищая его от разрыва, и определяет форму клетки.
Функции:
1) Защитная. Оболочка обеспечивает прежде всего механическую прочность.
2) Обеспечивает специализацию клеток, так как оболочка определяет форму клеток, что связано с их функциями.
Физические свойства: оболочка, как правило, бесцветна и прозрачна, легко пропускает солнечный свет. Она довольно жесткая, и в то же время эластичная, поэтому хорошо сохраняет форму. По оболочке (и сквозь неё) могут передвигаться растворы и газы.
Химический состав и молекулярная организация клеточной оболочки.
Компоненты клеточной оболочки:
1) скелетное вещество – образует каркас, остов оболочки. Это целлюлоза (клетчатка).
2) матрикс– основное вещество клеточной оболочки, заполняет пространство между молекулами целлюлозы. Состоит из пектинов и гемицеллюлозы.
Скелетное вещество
Целлюлоза – (С6Н10О5)n – в значительной степени определяет архитектуру и свойства оболочки. Молекулы целлюлозы состоят из молекул глюкозы, соединенных в неразветвленную цепочку, поэтому нити целлюлозы располагаются параллельно друг другу. И образуют волокна – фибриллы. Сначала молекулы целлюлозы объединяются в микрофибриллы, толщиной 10-25 нм.
Благодаря упорядоченному расположению молекул на отдельных участках микрофибрилл – мицеллах, целлюлоза обладает кристаллическими свойствами.
Микрофибриллы перевиваются, образуя тонкие нити, обматываются одна вокруг другой и объединяются в более толстые нити – макрофибриллы,
толщиной 0,5 мкм
и длиной до 4 мкм.
Макрофибриллы очень эластичны и прочны (также как равная им по толщине стальная проволока использование целлюлозы человеком (хлопок, лен, бумага, древесина и др.)), они видны в микроскоп. Целлюлоза нерастворима в воде, кислотах, щелочах, органических растворителях, не набухает. Поэтому целлюлоза определяет такие характерные для оболочки свойства, как прочность, эластичность и химическую инертность.
Т.о., целлюлозный каркас образован макро- и микрофибриллами.
Матрикс.
Пространство между макро- и микрофибриллами заполнено молекулами матрикса. Он имеет аморфную гелеобразную консистенцию. По составу он представляет собой сложную смесь полимеров, среди которых преобладают полисахариды. Цепочки их более короткие и разветвленные. Это: а) пектины – в состав которых входят различные моносахариды. Они сильно набухают в воде и некоторые растворимы, легко разрушаются под действием щелочей и кислот.
б) гемицеллюлозы – близки к пектинам, но более химически устойчивы, меньше набухают.
Также в состав матрикса входят: белки-гликопротеиды, у некоторых клеток полимеры лигнин, суберин, кутин, различные воска,минеральные соли, витамины, которые придают оболочкам дополнительные свойства (например, лигнин увеличивает жёсткость оболочки и характерен для клеток, выполняющих опорную функцию, кутин, суберин и воска – жироподобные вещества, снижают проницаемость клеточной оболтчки для воды).
Молекулы матрикса располагаются упорядоченно, переплетаются с фибриллами целлюлозного каркаса и образуют ковалентные связи, повышая прочность оболочки.
Матрикс определяет такие свойства клеточной оболочки как сильная набухаемость, проницаемость для воды и растворенных в ней мелких молекул и ионов, катионообменные свойства.
На определенных участках – порах, оболочка пронизана многочисленными плазмодесмами, обеспечивающими связь протопластов соседних клеток.
Т.о., оболочки растительных клеток сочетают в себе свойства живых и неживых структур.
Формирование, рост и структура клеточной оболочки.
Клеточная оболочка синтезируется протопластом в результате его жизнедеятельности. Клеточная оболочка отделена от протопласта плазмалеммой.
Плазмалемма и аппарат Гольджи играют основную роль в образовании оболочки.
В момент своего возникновения из материнской клетки дочерние клетки уже имеют 2/3 оболочки, являющейся частью материнской клетки. Между дочерними клетками начинает формироваться перегородка - срединная пластинка из пектиновых веществ, которая растёт радиально, от центра к периферии.
Одновременно с ростом срединной пластинки каждая дочерняя клетка со своей стороны начинает формировать недостающую часть оболочки.
От аппарата Гольджи к плазмалемме направляются пузырьки 2 типов: со светлым и тёмным содержимым.
Светлые пузырьки содержат ферменты и вещества-предшественники целлюлозы.
Тёмные пузырьки содержат полисахариды матрикса.
Подойдя к плазмалемме, пузырьки Гольджи встраиваются в неё и изливают своё содержимое наружу за пределы плазмалеммы.
Синтез и кристаллизация целлюлозы, образование и ориентация микрофибрилл ведёт плазмалемма при помощи своих белков-ферментов. Ориентировать микрофибриллы помогают микротрубочки, которые в больших количествах скапливаются вдоль плазмалеммы, располагаясь параллельно образующимся микрофибриллам, а затем и макрофибриллам целлюлозы.
Т.о., роль аппарата Гольджи – поставка строительного материала, а роль плазмалеммы – синтез целлюлозы, ориентация микро- и макрофибрилл и окончательное построение оболочки.
Рост молодой первичной оболочки происходит за счет внедрения одних макрофибрилл между другими. Происходит растяжение клеточной оболочки. Рост путём внедрения носит название интосусцепция.
Рост за счёт растяжения обусловлен тем, что вакуоль активно поглощает воду и увеличивает свой объём. Тургорное давление растягивает оболочку, но она не рвётся, а растягивается, так как наряду с аморфными полисахаридами матрикса в оболочку откладываются новые молекулы целлюлозы, которые формируют с молекулами матрикса ковалентные связи.
Т.о., между молодыми клетками формируется клеточная стенка, которая состоит: из срединной пластинки и 2 соседних оболочек клеток.
То есть, клеточная стенка – это совокупность оболочек соседних клеток с расположенной между ними срединной пластинкой.
На определённых участках клеточная стенка пронизана плазмодесмами.
Толщина оболочек клеток сильно варьирует, обычно оболочка многослойна и состоит из нескольких основных слоёв.
Каждая клетка имеет первичную оболочку, часть которой молодая клетка получает от материнской клетки, а часть надстраивает сама. Кроме того, многие клетки, специализируясь и теряя способность к делению, откладывают дополнительные слои – вторичную клеточную оболочку.
Первичная клеточная оболочка – начинает откладываться сразу после деления клетки, до начала или во время роста клетки. Первичные оболочки прозрачные, содержат 60-90% воды и 10% сухого вещества (матрикс (пектины и гемицеллюлозы) – 60-70%, целлюлоза - не более 30%, структурные белки гликопротеины– до 10%).
Т.о, в первичных оболочках клеток компоненты матрикса преобладают над скелетным веществом. Пектиновый компонент придаёт первичной оболочке эластичность, она хорошо растягивается по мере роста клетки и роста молодых органов. Первичные оболочки имеют молодые и активно делящиеся клетки, а также зрелые клетки активно участвующие в обмене веществ (фотосинтез, дыхание, секреция).
Первичные клеточные оболочки из-за преобладания пектинового компонента легко набухают и хорошо проницаемы для воды и газов. Клетка в этом случае может поглощать воду и газы всей своей поверхностью.
Когда клетка достигнет своих предельных размеров в некоторых случаях протопласт начинает откладывать вторичную оболочку.
Вторичная клеточная оболочка – откладывается не во всех клетках, а только в сильно специализированных (клетки механических, покровных и водопроводящих тканей). Протопласт откладывает вторичную оболочку изнутри, на внутреннюю поверхность первичной оболочки, наслаивая новые молекулы целлюлозы и матрикса. Оболочка не растягивается, а утолщается. Такой рост путём наложения называется аппозиция. Внутренний объём клетки при этом уменьшается.
Вторичная оболочка содержит гораздо меньше воды, преобладает сухое вещество, а в нем – целлюлоза – 40-50% (до 90% у волокон льна), матрикса содержится всего 20-30%, и в нём преобладают гемицеллюлозы, пектинов очень мало, гликопротеинов нет совсем. Зато в составе матрикса часто присутствует лигнин – 25-30%, а в клетках покровных тканей – суберин.
Как правило, во вторичной оболочке выделяется 3 слоя - наружный, средний и внутренний. Они отличаются по ориентации микрофибрилл целлюлозы (строго упорядочено).
Вторичная оболочка, благодаря высокому содержанию целлюлозы, а также лигнина приобретает жёсткость, твёрдость и плохо растягивается. Теряется проницаемость оболочки для воды и газов. Поэтому вторичная оболочка не откладывается сплошь по всей поверхности клетки. На отдельных участках, где сосредоточены скопления плазмодесм, вторичная оболочка не откладывается. Здесь остаётся только первичная оболочка и образуются характерные углубления - поры.
Поры – это неутолщённые участки в клеточной оболочке, через которые проходят плазмодесмы. Поры образуются с 2 сторон у соседних клеток.
Клеточная стенка (состоящая из срединной пластинки и двух соседних первичных оболочек) образует замыкающую плёнку поры. Через замыкающую плёнку свободно проходит вода и газы.
В живых клетках поры пронизаны плазмодесмами, через которые идёт транспорт крупных молекул и передаются раздражения.
Поры бывают: а) простые – поровый канал имеет одинаковую толщину – характерны для клеток паренхимы, волокон. Диаметр порового канала различен у разных клеток.
б) окаймлённые – поровый канал у замыкающей плёнки расширяется и вторичная оболочка нависает над поровым каналом в виде валика, образуя камеру поры (со стороны клетки поровый канал кажется очень узким). Такие поры характерны для рано отмирающих водопроводящих клеток древесины покрытосеменных.
в) у хвойных растений замыкающая плёнка окаймлённых пор несёт дискообразное утолщение – торус. Он лигнифицируется и непроницаем для воды, а вода проходит через окружающую торус замыкающую плёнку. Торус может смещаться в разные стороны при различном давлении воды и блокировать прохождение веществ. (торус=клапан)
Т.о., поры – это не сквозные отверстия, сквозные отверстия в оболочках клеток называются перфорации и характерны для клеток проводящих тканей.
Видоизменения клеточной оболочки.
Видоизменения клеточной оболочки химического характера и зависят от специализации клеток.
1) Одревеснение (лигнификация) – (самое распространённое) отложение лигнинаво вторичной оболочке клетки. Внешне такие клетки мало чем отличаются от обычных, но оболочка прекращает рост, теряет эластичность, зато приобретает прочность. Протопласт при этом обычно отмирает. (Иногда одревеснение обратимо, например, каменистые клетки плода груши). Одревеснению подвергаются клетки механических и водопроводящих тканей.
Лигнин играет большую роль в круговороте веществ в природе. В почве лигнин разлагается до гуминовых кислот, которые входят в состав гумуса. Отложения каменного угля образовались за счёт лигнина.
2) Опробковение (суберинизация) - отложение суберина во вторичной клеточной оболочке. Характерно для вторичных покровных тканей.
При суберинизации протопласт отмирает, так как суберин – жироподобное вещество, не пропускающее воду и газы.
Опробковевшие оболочки хорошо защищают от резких перепадов температур и испарения.
3) Кутинизация– отложение кутина на поверхности клеточных оболочек, тем самым снижается потеря воды, так как кутин - жироподобное вещество+ воск (кутикула– плёнка на поверхности клеток - защита от испарения). При кутинизации протопласт остаётся живым, так как кутикула не препятствует газообмену (кислорода и углекислого газа).
4) Минерализация– в оболочке откладываются соли Са, Мg, Si (у хвощей, бамбука, осоки, некоторые злаки). Оболочки становятся жёсткими, но хрупкими. Минерализованные оболочки хорошо сохраняются во времени (Баевское окаменелое дерево).
5) Ослизнение – никаких новых веществ не образуется, а целлюлоза разлагается до слизей (кончик молодого корня, клетки водорослей) – защита от высыхания, облегчает передвижение в почве.
Дата добавления: 2015-12-08; просмотров: 542 | Нарушение авторских прав
mybiblioteka.su - 2015-2018 год. (0.019 сек.)mybiblioteka.su
1 основным скелетным веществом первичной оболочки является
1. Основным скелетным веществом первичной оболочки является:
а) целлюлоза, в) крахмал,
б) глюкоза, г) гемицеллюлоза.
2. Структурной единицей первичной оболочки растительной клетки является:
а) микрофибрилла, в) макрофибрилла,
б) мицелла, г) молекула целлюлозы.
3. Срединная пластинка между двумя соседними клетками составлена:
а) гемицеллюлозами, в) пектиновыми веществами,
б) целлюлозой, г) галактозой.
4. Матрикс первичной оболочки растительной клетки состоит из:
а) гемицеллюлоз и рибоз, в) фруктозы и галактозы,
б) галактозы и рибоз, г) пектиновых веществ и гемицеллюлоз.
5. Наружное положение в клетке занимает:
а) вторичная оболочка, в) первичная оболочка,
б) плазмалемма, г) срединная пластинка.
6. Наиболее тонкие участки в первичной оболочке называют:
а) порами, в) устьицами,
б) поровыми полями, г) плазмодесмами.
7. Утолщения первичной оболочки в окаймленных порах называют:
а) торусом, в) замыкающей пленкой пор,
б) апертурой, г) поровым каналом.
8. Связь между двумя растительными клетками осуществляется через
а) синапсы, в) десмосомы,
б) плазмодесмы, г) замыкающие контакты.
9. Первичная оболочка сохраняется:
а) в течение всей жизни клетки,
б) на время роста клетки,
в) до вступления клетки в функциональную фазу,
г) только в эмбриональной фазе.
10. Формирование первичной оболочки между двумя дочерними клетками по окончании митоза называют:
а) интеркинезом, в) цитокинезом,
б) интерфазой, г) симбиогенезом.
11. Образование первичной оболочки между двумя дочерними клетками у растений происходит с участием:
а) фрагмопласта, в) хлоропласта,
б) лейкопласта, г) хромопласта.
12. Рост первичной оболочки растительных клеток происходит путем:
а) сжатия, в) встраивания,
б) растяжения, г) наложения.
13. Вторичная оболочка откладывается:
а) во время деления клетки, в) во время формирования органов,
б) после прекращения роста клетки, г) в ходе цитокинеза.
14. Пропитывание вторичной оболочки суберином называют:
а) опробковение, в) одревеснение,
б) ослизнение, г) минерализация.
Тест - билет по теме «Оболочка растительной клетки»Вариант 2
1. Вторичная оболочка откладывается:
а) одновременно с первичной, в) после отмирания клетки,
б) после прекращения роста клетки, г) до начала образования первичной оболочки.
2. Вторичная оболочка составляет:
а) сплошной слой на первичной оболочке,
б) сплошной слой под первичной оболочкой,
в) инкрустирована в первичную оболочку,
г) прерывистый слой на первичной оболочке.
3. Вторичная оболочка состоит:
а) из трех слоев микрофибрилл, в) из одного слоя микрофибрилл,
б) из двух слоев микрофибрилл, г) из четырех слоев микрофибрилл
4.К видоизменениям вторичной оболочки не относится:
а) минерализация, в) одревеснение,
б) ослизнение, г) хитинизация.
5. Отложение во вторичной оболочке кремнезема и углекислого кальция называется:
а) кутинизация, в) одревеснение,
б) опробковение, г) минерализация.
6. Кутикула:
а) газо- и водонепроницаема,
б) придает прочность стенкам клеток,
в) предохраняет от воздействия высоких температур,
г) придает жесткость стенкам клеток.
7. На стенках сосудов ксилемы вторичная оболочка может образовывать утолщения:
а) дуговое, в) спиральное,
б) ребристое, г) зигзагообразное.
8. Прочность стенкам сосудов придает:
а) минерализация, в) кутинизация,
б) опробковение, г) одревеснение.
9. Перерывы во вторичной оболочке обеспечивают существование:
а) устьиц, в) пор,
б) трихомов, г) эмергенцев.
10. Для трахеид хвойных характерен следующий тип пор:
а) окаймленные, в) полуокаймленные,
б) простые, г) сложные.
11. Основная функция пор:
а) запасающая, в) механическая,
б) защитная, г) ионообменная.
12. Мицелла – это
а) группа молекул целлюлозы, в) цепочка молекул глюкозы,
б) объединение молекул пектина, г) молекулы гемицеллюлоз.
13. Молекулы целлюлозы в первичной оболочке растительной клетки:
а) расположены хаотично, в) формируют сеть,
б) не соприкасаются друг с другом, г) образуют пространственную решетку.
14. Связь между двумя растительными клетками осуществляется через:
а) синапсы, в) десмосомы,
б) плазмодесмы, г) замыкающие контакты.
Вопросы коллоквиума по теме «Образовательные ткани»
1. Особенности клеток меристем.
2. Классификация меристем по положению в теле растения.
3. Строение апекса побега.
4. Строение апекса корня двудольных.
5. Строение апекса корня однодольных.
6. Сравнение строения апексов корней стебля и корня.
7. Особенности латеральных меристем.
8. Интеркалярные меристемы.
Вопросы контрольной работы по теме «Ткани»
1 Вариант
1. Перечислите основные группы элементов проводящих тканей.
2. В чем сходство и различие проводящих сосудов и ситовидных трубок? Ответ оформите в виде таблицы:
Признаки | Ситовидные трубки | Сосуды |
1. Функции | ||
2. Строение | ||
3. Происхождение | ||
4. В состав какой ткани входят |
3. Изобразите онтогенетический и эволюционный ряд сосудов древесных покрытосеменных растений.
2 Вариант
1. Перечертите и заполните таблицу:
Строение | Функции | Особенности каждого из элементов | Из какой меристемы образуется | Время жизни | |
Эпидерма | 1) 2) 3) |
2. По какому признаку покровные ткани делят на первичные, вторичные, и третичные?
3. Приведите примеры, свидетельствующие о различиях и строениях эпидермы у разных растений.
3 Вариант
1. Перечертите и заполните таблицу
Ткани | Функции | Расположение в стебле | Типы и их особенности | Из какой меристемы возникают |
Колленхима | ||||
Склеренхима |
2. В чем заключаются различия между древесными и лубяными волокнами?
3. Одинаково ли соотношение склеренхимы и колленхимы в стеблях травянистых и древесных растений?
4 Вариант
1. Схема строения апекса побега. Укажите, какие ткани формируются из каждого слоя конуса нарастания.
2. Объясните, почему растения называют «открытыми системами роста», в отличие от животных. Благодаря какой ткани существует это свойство.
3. Классификация меристем по местоположению в теле растения, их функции (в виде схемы).
5 Вариант
1. Перечертите и заполните таблицу:
Ткани | Состав | Строение проводящих элементов (зарисовать) | Функции | Расположение в органах растений | Из какой меристемы возникают |
Флоэма | |||||
Ксилема | |||||
2. Классификация проводящих пучков (в виде схемы)
3. Какие пучки называют простыми, сложными, сосудисто-волокнистыми?
6 Вариант
1. Зарисуйте схему строения апекса корня двудольных. Укажите, какие ткани формируются из каждого слоя.
2. В чем состоят различия в строении апексов корней двудольных и однодольных растений?
3. Основная функция апикальных меристем.
7 Вариант
1. Перечертите и заполните таблицу:
Ткани | Строение (назвать основные элементы) | Функции | Время возникновения | Место образования |
Перидерма | ||||
Корка |
2. Что такое чечевички и какова их функция?
3. Почему перидерму называют вторичной покровной тканью, а корку третичной?
Тест-билет по теме "Семена и проростки"
1. Семя - орган размножения и расселения:
а) плауновидных в) папоротниковидных
б) моховидных г) покрытосеменных
2. Составными частями семени являются:
а) корешок, гипокотиль, эндосперм, семенная кожура
б) семенная кожура, запас питательных веществ, зародыш
в) спермодерма, эпикотиль, эндосперм
г) перисперм, зародыш, колеоптиль
3. Происхождение зародыша однодольных объясняет "гипотеза недоразвития":
а) Гегельмайера в) Гёте
б) Шмидта г) Ганштейна
4. Зрелый зародыш двудольных состоит из:
а) зародышевого корешка и побега в) апекса и побега
б) почечки и корешка г) корешка и гипокотиля
5. Последовательные фазы формирования зародыша двудольных:
а) проэмбрио, октант, глобулярная фаза, сердцевидная фаза,
торпедо
б) проэмбрио, сердцевидная фаза, октант, глобулярная фаза,
торпедо
в) прэмбрио, глобулярная фаза, октант, сердцевидная фаза,
торпедо
г) октант, проэмбрио, глобулярная фаза, сердцевидная фаза,
торпедо
6. Зародыш двудольных становится зелёным на стадии:
а) торпедо в) глобулярной
б) сердцевидной г) октанта
7. Какую часть зародыша двудольных формирует проэмбрио:
а) апекс побега в) семядоли
б) корневой чехлик г) гипокотиль
8. Какие части зародыша двудольных образованы нижним октантом клеток:
а) семядоли и апекс в) апекс и гипокотиль
б) гипокотиль и корешок г) гипокотиль
9. Составные части зародыша злаков:
а) колеориза, корешок, гипокотиль, почечка, колеотиль, щиток, эпибласт
б) колеотиль, почечка, гипокотиль, корешок, колеориза, щиток
в) колеоптиль, почечка, гипокотиль, корешок
г) колеориза, корешок, гипокотиль, почечка, колеоптиль, щиток
10. Функция колеотиля:
а) защита почечки
б) раздвигание частиц почвы при прорастании семени
в) защита корешка
г) доставка первых порций воды при прорастании семени
11. Колеориза - это:
а) "колпачок" корня в) одна из семядолей
б) "колпачок" почки г) запасная ткань семени
12. Надземным называют прорастание семян при котором семядоли:
а) остаются в семени в) остаются под землёй
б) выносятся на поверхность г) не образуются
13. Процессу прорастания семян предшествует:
а) высыхание в) разрушение
б) набухание г) деление
14. Спермодерма - это:
а) часть зародыша в) семенная кожура
б) запасная часть семени г) запас питательных веществ
15. Запасные вещества семени могут откладываться в:
а) эндосперме в) эпикотиле
б) гипокотиле г) колеоптиле
16. Эндосперм и перисперм различаются:
а) временем образования в) типом запасных веществ
б) способом формирования г) функциями
textarchive.ru
Клеточная оболочка.
Общее понятие о клеточной оболочке.
У высших растений лишь немногие клетки лишены клеточной оболочки, например, генеративная клетка и спермин в пыльцевом зерне. Все прочие клетки непосредственно после их образования покрываются оболочкой. Полисахаридная клеточная оболочка с плазмодесмами - неотъемлемый специфический компонент растительной клетки.
Она играет большую роль в жизни клетки:
1 - обеспечивает прочность и защищает протопласт от повреждений; своим противодавлением поддерживает клетку в тургорном (напряженном) состоянии. Благодаря наличию оболочки, растяжение протопласта осмотически активной вакуолью ограничено, а размер и форма клетки, достигшей зрелости, перестают изменяться.
2 - тип клеточных оболочек определяет анатомические особенности тканей. В периферических тканях органов клеточные оболочки содержат вещества, которые защищают лежащие глубже клетки от высыхания. Оболочки обеспечивают такие важные стороны деятельности растительных тканей, как поглощение воды и ионов солей, а также некрупных молекул, транспирацию, транспорт и секрецию веществ.
3 - совокупность всех клеточных оболочек представляет собой механический остов, который придает растению, не имеющему внутреннего скелета, прочность.
4 - система оболочек и межклетников между клетками обеспечивает передвижение веществ от клетки к клетке и называется апопластом (греч. аро — из, от, без и plastos — оформленный).
Клеточные оболочки соседних клеток как бы сцементированы между собой межклеточной, или срединной, пластинкой.
Она образуется из клеточной пластинки, которая со временем пропитывается особыми веществами, например, лигнином. Срединная пластинка очень тонкая и неразличима под световым микроскопом.
Протопласты соседних клеток разделены между собой клеточной стенкой, состоящей из двух клеточных оболочек и срединной пластинкой между ними.
Оболочка, как правило, бесцветна, прозрачна, пропускает солнечный свет. По химическому составу и строению оболочки можно судить о происхождении и функциях клеток разных типов и определить, к каким тканям данные клетки относятся. Именно поэтому внутреннее строение ископаемых растений изучают, в основном, путем исследования оболочек их клеток.
Клеточная оболочка строится непосредственно протопластом и растет, только находясь в контакте с ним. Очень часто клеточная оболочка сохраняется дольше протопласта.
Однако гибель протопласта не прекращает существования клетки как структурной единицы организма, поскольку форма клетки благодаря прочной оболочке сохраняется. Поэтому некоторые растительные клетки и после отмирания протопласта выполняют свои функции, особенно проведения растворов и механической опоры. Волокна, трахеиды, членики сосудов, клетки пробки во взрослом состоянии представляют собой одни клеточные оболочки. В основном из оболочек отмерших клеток состоит древесина.
Химический состав и молекулярная организация оболочки.
Клеточная оболочка построена в основном из полисахаридов. По своей роли и нахождению в составе оболочки они подразделяются на скелетные вещества и вещества матрикса.
Скелетным веществом оболочки высших растений является целлюлоза (клетчатка) — Р-1,4-О-глюкан.
Это наиболее распространенное вещество биосферы (более половины органического углерода планеты сосредоточено в целлюлозе). Молекулы целлюлозы, представляющие полимерные цепи из многочисленных мономеров: глюкозы, ксилозы, маннозы, могут достигать в длину несколько микрометров.
Целлюлоза нерастворима в воде и органических растворителях, не набухает, не растворяется в разбавленных кислотах и даже в концентрированных щелочах.
Обычно молекулы целлюлозы особыми связями собраны по несколько десятков в группы. В результате этого образуются тончайшие (1,5-4 нм), видимые только в электронный микроскоп волоконца неопределенной длины — микрофибриллы.
В определенных частях микрофибриллы молекулы целлюлозы располагаются упорядоченно на близком расстоянии параллельно друг другу, образуя мицеллы.
Между мицеллами находятся менее упорядоченно расположенные цепи из мономеров, образующие паракристаллические образования. Поэтому целлюлоза обладает свойствами кристаллов.
Микрофибриллы создают структурный каркас оболочки, ее «арматуру» и обеспечивают прочность. Они объединяются в более сложные структуры, называемые макрофибриллами, которые различимы уже в световом микроскопе.
Микрофибриллы эластичны и очень прочны на разрыв (сходны со сталью). На основании этих свойств целлюлоза и ее продукты широко используются для производства хлопкового волокна, ацетатного шелка, бумаги, вискозы, бездымного пороха.
Макрофибриллы целлюлозы погружены в обладающий свойствами коллоида матрикс.
По физическим свойствам матрикс представляет собой насыщенный водой пластичный гель. Он является сложной смесью полимеров, среди которых также преобладают полисахариды, но с более короткими, чем у целлюлозы, и часто разветвленными молекулами.
Цепи матричных полисахаридов заполняют пространство между макрофибриллами целлюлозы и образуют как с ними, так и между собой, многочисленные поперечные (ковалентные и водородные) связи.
Это настолько увеличивает прочность клеточной оболочки, что ее иногда сопоставляют с железобетоном или стекловолокном.
Матричные полисахариды гидрофильны (греч. гидро — влага и филю - люблю), способны сильно набухать и поглощать много воды. Они обладают высокой проницаемостью растворенных в воде молекул и ионов. По клеточным оболочкам может происходить передвижение веществ от клетки к клетке путем диффузии по градиенту концентрации (пассивный транспорт).
mykonspekts.ru
ЛЕКЦИЯ 5 - Стр 2
Состав, концентрация и вязкость клеточного сока у разных видов растений различны и неодинаковы даже в тканях, органах и клетках одного растения. Далеко не все перечисленные вещества одновременно присутствуют в клеточном соке. Некоторые из них (алкалоиды, гликозиды) характерны только для определенных групп растений, тогда как другие распространены более широко. Нередко в специализированных клетках происходит накопление только какого-либо одного вещества. На состав и свойства клеточного сока большое влияние оказывает возраст клетки и окружающие условия.
Вещества клеточного сока могут быть разносторонне использованы, и поэтому он является ценнейшим комплексным сырьем для промышленности. Особенно большое практическое значение имеет млечный сок как источник получения каучука, гуттаперчи, опиума, кодеина и других веществ. Большое количество каучука содержится в млечном соке бразильской гевеи, а также травянистых каучуконосов — кок-сагыза, крым-сагыза и тау-сагыза, произрастающих в РФ.
Оболочка растительной клетки. Одной из характерных особенностей растительной клетки является плотная оболочка, которую образует на поверхности клетки протопласт в процессе своей жизнедеятельности. Наличие или отсутствие оболочки служит надежным признаком, который позволяет отличить растительную клетку от животной. Оболочка защищает протопласт от внешних воздействий и придает клетке форму и прочность. Изнутри клеточная оболочка выстлана плазмалеммой. Некоторые клетки растений оболочки не имеют (половые клетки, клетки слизевиков).
Клеточные оболочки значительно изменяются в зависимости от возраста и типа клетки. Обычно молодые клетки имеют оболочку более тонкую, чем клетки, полностью сформировавшиеся. Клеточной оболочке свойственна пластичность, т. е. способность принимать и сохранять в дальнейшем новую форму и размеры, а также эластичность, благодаря которой оболочка может восстанавливать прежнюю форму и размеры после деформации. Клеточная оболочка обладает значительной прочностью на растяжение. Строение обо-' лочки тесно связано с функцией клетки.
Химический состав и структура оболочки. В состав оболочки чаще всего входят - целлюлоза (клетчатка), гемицеллюлоза (полуклетчатка) и пектиновые вещества. Наибольшее значение и распространение имеет целлюлоза, нередко составляющая до 90% вещества оболочки. Она представляет собой углевод (полисахарид), близкородственный крахмалу, и имеет такую же эмпирическую формулу — (С6Н10О5) п ,— но с другим значением коэффициента п и с более сложным молекулярным строением. Молекулы целлюлозы имеют нитчатую структуру и, располагаясь параллельно, группируются в пучки — мицеллы. Мицеллы в свою очередь образуют более крупные структурные элементы — фибриллы, промежутки между которыми заполнены основным веществом оболочки (матриксом), состоящим из пектиновых веществ и гемицеллюлозы. Целлюлоза осахаривается в крепких кислотах, а растворяется только в реактиве Швейцера (аммиачный раствор окиси меди). Гемицеллюлоза также является очень стойким веществом, но поддается разложению несколько легче, чем целлюлоза.
Петкиновые вещества в отличие, от целлюлозы и гемицеллюлозы состоят не из нитчатых, а из сильно разветвленных молекул, вследствие чего они обычно аморфны. Особенностью пектиновых веществ является их способность набухать в йоде. Кроме того, пектиновые вещества обладают значительно меньшей прочностью и сравнительно легко разрушаются. У некоторых низших растений оболочка клеток полностью состоит из пектиновых веществ.
За л ож е н u е и рост оболочки. В клетке различают первичную и вторичную оболочки. Каждая вновь образовавшаяся клетка сразу окружается очень тонкой прозрачной оболочкой. Эта оболочка является первичной, и в ней преобладают гемицеллюлоза и пектиновые вещества, а также содержится большое количество воды. Формирование первичной оболочки заканчивается, когда клетка достигает своего окончательного размера и перестает расти. Некоторые клетки до конца жизни остаются покрытыми первичной оболочкой. Однако в большинстве случаев после прекращения роста клетки протопласт ее начинает формировать вторичную оболочку, вещество которой откладывается на внутреннюю поверхность первичной оболочки. В состав вторичной оболочки входит главным образом целлюлоза. В ней обычно хорошо заметны слоистость и щтриховатость, обусловленные ее субмикроскопической структурой. Преобладание целлюлозы определяет высокие механические качества вторичной оболочки, особенно ее прочность на растяжение и эластичность. Иногда в клетках различают третичную оболочку в виде тонкого внутреннего слоя, в состав которого входит особое вещество — ксилан.
Между первичными оболочками соседних клеток находится прослойка межклеточного пектинового вещества, которая называется срединной пластинкой. Совокупность первичных оболочек двух соседних клеток и заключенной между ними тонкой прослойки межклеточного вещества образует клеточную стенку. Некоторые авторы отождествляют клеточную оболочку с клеточной стенкой, что, по-видимому, не совсем правильно. Разрушение срединной пластинки приводит к разъединению клеток — мацерации. Обособленные, мацерированные, клетки обычно приобретают шаровидную форму, тогда как будучи соединены одна с другой и испытывая взаимное давление, они имеют форму многогранников. Рост клеточной оболочки может осуществляться двумя способами: наложением (обычно изнутри) новых слоев оболочки на старые (аппозиция) и внедрением частиц вещества оболочки между старыми (интуссусцепция). При аппозиции происходит утолщение клеточной оболочки, при интуссусцепции — растяжение и увеличение ее поверхности. Оболочки имеют различную толщину, что обусловлено функцией клетки. Так, у опорных клеток толщина оболочки может достигать 10 мкм. Нередко оболочка настолько утолщается, что занимает всю полость клетки, вследствие чего происходит отмирание протопласта. Иногда наблюдается местное утолщение оболочки — отдельными участками в виде колец, спиралей и т. и.
П о р ы и п л а з м о д е с м ы. При формировании первичной оболочки в ней возникают участки, на которых отложение вещества оболочки происходит менее интенсивно. В результате в первичной оболочке появляются многочисленные углубления, получившие название первичных поровых полей. Во вторичной оболочке также имеются участки, на которых вещество оболочки не откладывается, вследствие чего в ней возникают прорывы, достигающие первичной оболочки и называемые порами.
Поры двух смежных клеток, как правило, совпадают. Между ними имеется участок тонкой первичной оболочки, называемый замыкающей пленкой поры. Следовательно, полость поры с внутренней стороны непосредственно соединяется с полостью клетки, а с наружной, там, где она соприкасается с соседней клеткой, прикрыта замыкающей пленкой.
В клетках с мощно развитой вторичной оболочкой поры превращаются в поровые каналы, идущие от полости клетки до первичной оболочки. Обычно поры образуются непосредственно над первичными поровыми полями, но могут возникать и над другими участками первичной оболочки.
Различают 2 типа пор — простые и окаймленные. Упростых пор диаметр порового канала приблизительно одинаков на всем протяжении. У окаймленных пор он резко суживается по мере отложения вторичной оболочки, вследствие чего внутреннее отверстие поры, ведущее в полость клетки, гораздо уже, чем наружное, граничащее с первичной оболочкой. При этом вторичная оболочка в виде валика нависает над расширенной частью канала.
Замыкающие пленки пор пронизаны мельчайшими отверстиями в виде канальцев, через которые из одной клетки в другую проходят нити цитоплазмы — плазмодесмы. Ввиду того, что плазмодесмы являются очень тонкими и нежными, увидеть их в световой микроскоп удается не всегда. Однако применение электронного микроскопа позволило обнаружить плазмодесмы почти у всех растений и во всех тканях. Количество плазмодесм в клетке очень велико и у некоторых растений (омела) достигает 6...24 тыс.
Плазмодесмы имеют большое биологическое значение. Они связаны с эндоплазматической сетью, а также соединяют протопласты отдельных клеток, обеспечивая непрерывность эндоплазматической сети и всей цитоплазмы организма. С помощью плазмодесм осуществляются проведение различных веществ, передача раздражений из одной клетки в другую и регуляция всех жизненных процессов, протекающих в организме. Плазмодесмы были впервые описаны в 1877 г. русским ученым И. Н. Горожанкиным, а затем Э. Руссоным, Э. Танглем, Э. Страсбургером и другими исследователями.
В последнее время нити цитоплазмы были обнаружены также в оболочках клеток, граничащих с внешней средой. Они получили название эк т о д е с м. По-видимому, они играют роль в выделении наружу в поглощении клеткой из внешней среды воды и растворенных в ней веществ.
В и д о и з м с н с. н и я клеточной оболочк и. Многие клетки сохраняют целлюлозные оболочки до конца своей жизни. Однако часто в процессе жизнедеятельности протопласта клеточная оболочка подвергается различным изменениям и приобретает новые химические и физические свойства. К числу таких изменений относятся одревеснение, опробковение, кутинизация, минерализация и ослизнение.
Одревеснение клеточной оболочки происходит в результате накопления в ней особого вещества — лигнина, который откладывается в промежутках между фибриллами целлюлозы, не вступая с ней в химическое соединение. Лигнин отличается от углеводов более высоким содержанием углерода, его эмпирическая формула С57Нв0О10. Однако химическая природа лигнина окончательно еще не выяснена. Ультраструктура одревесневших оболочек напоминает структуру железобетона, причем микрофибриллы можно сравнить с арматурой, а лигнин играет роль основного вещества. Одревесневшая оболочка теряет эластичность, становится более твердой, хрупкой и приобретает большую прочность на сжатие. Особенно сильное одревеснение клеточных оболочек наблюдается у кустарников и деревьев. При этом клетки могут сохранять живое содержимое, и в них не прекращается обмен веществ. Однако чаще такие клетки отмирают. Лигнин предохраняет клетки высших растений от разрушительного действия бактерий и грибов. В некоторых случаях происходит раздревеснение клеточных оболочек — они теряют лигнин и снова становятся мягкими. Подобное явление можно наблюдать, например, при созревании плодов груши или айвы, сопровождающемся раздревеснением оболочек каменистых клеток.
Опробковение заключается в пропитывании оболочки жироподобным веществом — суберином, который делает ее непроницаемой для воды и газов. Суберин не образует скелетные структуры, как это наблюдается при пропитывании оболочки лигнином. Он обычно накладывается изнутри па первичную оболочку в виде тонкого слоя. Вскоре после образования суберинового слоя клетка, будучи изолирована от внешней среды, отмирает и наполняется воздухом, как у пробкового дуба, или в ней накапливаются различные вещества.
Кутинизация — это пропитывание клеточных оболочек жироподобным веществом - к у т и и о м, который по своей химической природе близок к суберину. Как правило, кутин пропитывает только ту часть клеточной оболочки, которая непосредственно соприкасается с атмосферой. Часто кутин образует на поверхности клеток непрерывный застывший слой –кутикулу - в виде очень тонкой блестящей пленки. Кутинизация является защитным приспособлением против- слишком интенсивного испарения. Кроме того, кутикула отражает солнечные лучи, что предохраняет растение от перегрева, а иногда защищает листья от ультрафиолетового излучения, поглощая ультрафиолетовые лучи.
Ослизнение клеточных оболочек заключается в превращении клетчатки или пектиновых веществ в более высокомолекулярные углеводы — слизи и камеди, способные к сильному набуханию при соприкосновении с водой. Чаще всего ослизнению подвергаются оболочки клеток семенной кожуры у семян льна, тыквы, арбуза и листьев некоторых засухоустойчивых растений. Ослизнение ускоряет прорастание семян, а также предохраняет растения от перегрева. Иногда ослизнение клеточных оболочек и содержимого клеток наблюдается при поранениях. При этом происходит камедетечение (гуммоз), характерное для вишни, сливы и других растений.
Минерализация представляет собой отложение минеральных солей (кремнезема, углекислого кальция и др.) в клеточных оболочках стеблей и листьев многих растений — осок, злаков, хвощей. Минерализация повышает прочность оболочки и придает ей особую твердость, защищая растение от поедания животными.
Оболочка растительных клеток имеет большое практическое значение и используется в качестве сырья для получения клетчатки, древесины и других веществ, из которых вырабатываются бумага, искусственный шелк, кинопленка, целлофан и др.
studfiles.net
Оболочка
Категория: Анатомия растений
Оболочка
Клетки растений в большинстве случаев имеют хорошо выраженную оболочку, или стенку, которая защищает протопласт, определяет форму клеток, участвует в проведении веществ, а также играет механическую роль.
Образованию оболочки предшествует деление ядра. В телофазе митоза (см. с. 39) во фрагмопласте закладывается тонкая межклеточная пектиновая мембрана. На нее в результате деятельности протопласта с обеих сторон откладывается тонкая эластичная первичная оболочка, состоящая из пектиновых веществ, гемицеллюлоз и целлюлозы. Такую оболочку имеют клетки меристематических (например, камбия), фотосинтезирующих и некоторых других тканей. У большинства клеток после окончания их роста на первичную оболочку откладывается более плотная вторичная, преимущественно целлюлозная оболочка. В некоторых местах вторичной оболочки не образуется. Эти места называются порами.
Пора одной клетки расположена напротив поры соседней клетки. Такая пара пор, находящаяся в смежной стенке между двумя клетками, разделена замыкающей пленкой, или мембраной поры, состоящей из двух первичных оболочек соседних клеток и разделяющего их межклеточного вещества. В живой клетке полость поры заполнена цитоплазмой. В замыкающей пленке через тончайшие субмикроскопические отверстия проходят тяжи цитоплазмы — плазмодесмы, осуществляющие связь между протопластами соседних клеток.
Толщина оболочки зависит от выполняемой клеткой функции. Клетки, обусловливающие механическую прочность и твердость органов растения, имеют толстые оболочки; клетки, несущие запасающую, выделительную и некоторые другие функции, снабжены тонкой оболочкой.
Строение оболочек можно изучать на срезах того или иного органа, а также на мацерированном материале, получение которого основано на искусственном разъединении клеток вследствие растворения межклеточного вещества.
Тонкие оболочки клеток сердцевины бузины красной (Sambucus racemosa L.)
Сердцевина бузины служит одним из лучших объектов для начинающих работать бритвой. Ее получают из окончивших рост толстых однолетних побегов, отходящих от основания куста (так называемые жировые побеги). Междоузлия режут на куски длиной около 10 см и коротким шомполом выбивают из них сердцевину. Диаметры шомпола и сердцевины должны быть одинаковыми. Если сердцевина очень влажная и ее не удается выбить, кусочки стебля подсушивают в сухом помещении в течение двух-трех дней. Сухую или выдержанную в спирте сердцевину резать очень легко. Площадь среза и направление его по отношению к оси стебля для изучения клеток, слагающих сердцевину, значения не имеют.
Тонкий срез сухой сердцевины рассматривают в воде. Желательно, чтобы толщина среза была меньше диаметров клеток, иначе наложение одних клеток на другие помешает рассмотреть объект.
Чтобы удалить пузырьки воздуха из разрезанных клеток, по поверхности покровного стекла следует слегка постучать препаровальной иглой. Тогда полости этих клеток заполнятся водой. Там, где толщина среза больше диаметра клетки и они оказались неповрежденными, их полости останутся заполненными воздухом, который под микроскопом имеет вид пузырьков, окруженных черной каймой.
При малом увеличении надо найти наиболее тонкое место среза. Оно должно быть прозрачным и без пузырьков воздуха.
Сердцевина состоит из почти округлых тонкостенных, лишенных содержимого клеток различных размеров. Клетки имеют приблизительно одинаковые контуры и размеры в трех взаимно перпендикулярных направлениях. Такие клетки называют парен-х и м н ы м и. Клетки расположены на разных уровнях, поэтому на срезе они обычно лежат одна на другой. Между клетками имеются межклетники, очертания которых зависят от числа окружающих их клеток.
При большом увеличении микроскопа можно видеть, что в некоторых местах оболочка клетки очень тонка. Эти тонкие участки оболочки представляют собой поры. Они выглядят по-разному в зависимости от положения: в плане они округлые или несколько продолговатые, в разрезанных стенках между двумя клетками поры имеют вид коротких каналов, пересеченных тонкой замыкающей пленкой. Поры, канал которых на всем протяжении одинаковой ширины, называют простыми. Если пора находится в стенке, обращенной к межклетнику, то ее канал замкнут снаружи только одной первичной оболочкой. Эти поры называют слепыми.
Задание. При большом увеличении микроскопа зарисовать несколько клеток сердцевины, отметить форму клеток, межклетники, поры в плане и в разрезе.
Строение стенок паренхимных клеток с простыми порами можно рассмотреть и з сердцевине многих древесных растений, а также в основной ткани стеблей кукурузы или сорго.
Оболочки клеток эпидермиса листа аспидистры (Aspidistra elatior Blume)
Простые поры в тонкостенных клетках хорошо видны в эпидермисе аспидистры — обычного комнатного растения.
Из живого или фиксированного спиртом листа вырезают тонкую полоску, которую обвертывают вокруг указательного пальца левой руки, и, придерживая лист с двух сторон большим и средним пальцами, делают возможно более тонкий небольшой поверхностный срез. Его кладут в воду и накрывают покровным стеклом. Лучше всего рассматривать края среза, где клетки чаще всего расположены в один ряд. Средняя часть среза обычно получается более толстой, так как кроме клеток эпидермиса здесь могут быть и глубже лежащие клетки листа с многочисленными хлоропластами.
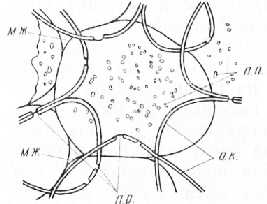
Рис. 1. Поперечный срез сердцевины бузины: мж — межклетники, п. п. — поры в плане, п. р. — поры в разрезе, о. к. — оболочка клетки
Эпидермис состоит из клеток, вытянутых вдоль листа. Клетки живые, с цитоплазмой и ядром. В стенках смежных клеток хорошо видны в разрезе поров ые каналы с тонкой замыкающей пленкой (рис. 2).
Задание. При большом увеличении зарисовать несколько клеток эпидермиса, отметить в них содержимое, на боковых стенках показать поры в разрезе и тонкие мембраны, пересекающие каналы пор.
Строение оболочек волокон в стебле льна-долгунца (Linum usitatissimum L.)
Клетки, длина которых во много раз превышает ширину, называются прозенхимными. Обычно они заострены на концах и имеют вид волокон.
Для изучения оболочек волокон пригодны стебли многих прядильных растений, в том числе льна-долгун-ца. Стебли льна фиксируют спиртом через 2—3 недели после начала цветения. Тонкий поперечный срез междоузлия рассматривают в воде. Срез может быть неполным.
При малом увеличении микроскопа видно, что в середине среза стебля находятся тонкостенные рыхлые клетки сердцевины, вокруг нее плотным широким кольцом располагается древесина. В периферической, коровой, части стебля хорошо заметны группы клеток со светлыми блестящими, сильно утолщенными оболочками, толщина которых зависит от сорта льна, сроков сбора материала и агротехники выращивания растения. При большом увеличении следует рассмотреть группу этих клеток.
Рис. 2. Клетки эпидермиса листа аспидистры: п. о. — первичная оболочка, в. о. — вторичная оболочка, п. р. — пора в разрезе, я — ядро, м. п. — мембрана поры
Если клетки расположены рыхло, то они имеют округлые или овальные контуры; плотно сомкнутые клетки в очертании многоугольные. В полостях молодых клеток заметны сгустки свернувшегося при фиксации содержимого. В старых клетках, в которых закончились рост и утолщение оболочек, протопласты отмирают.
В оболочке хорошо видны слои, параллельные поверхности клетки. Поровые каналы встречаются редко.
Чтобы рассмотреть общий вид этих клеток, следует выделить их из стебля, приготовив тотальный препарат. Для этого с кусочка стебля длиной 3—5 см снимают коровую часть и кладут ее на предметное стекло в каплю воды эпидермисом кверху. Скальпелем или препаровальной иглой удаляют периферические ткани, пока на стекле не останется пучок параллельно лежащих волокон. Накрыв их покровным стеклом, при малом увеличении надо найти свободно лежащие волокна. Длина волокна в сотни раз превышает его ширину. Концы волокна узкие, заостренные. В средней части волокна иногда встречаются местные расширения. Поскольку толщина волокна значительно превышает глубину поля зрения микроскопа, при большом увеличении, пользуясь микрометренным винтом, можно увидеть волокно с поверхности и в оптическом разрезе.
При рассмотрении волокна в оптическом разрезе видна полость клетки с остатками содержимого, состоящего из мелкозернистых сгустков. В оболочке заметна слоистость, параллельная продольной оси клетки (рис. 3).
Рассматривая волокно с поверхности, можно заметить тонкую косую исчерченность оболочки. Это так называемая полосатость, или штриховатость. На разной глубине, т. е. в разных слоях вторичной оболочки, штрихи могут быть направлены в противоположные стороны. Это объясняется тем, что каждый слой вторичной оболочки состоит из параллельных, винтообразно закрученных вокруг продольной оси клетки тонких волоконец целлюлозы — фибрилл. Такая структура оболочки значительно увеличивает прочность волокна, подобно тому, как достигается значительная крепость веревки, скрученной из волокон какого-либо прядильного растения.
Угол наклона фибрилл имеет диагностическое значение. У многих прядильных растений фибриллы расположены под определенным углом к продольной оси клетки.
Фибриллярным строением объясняются высокие технические свойства волокон льна, их прочность, эластичность и упругость.
В качестве реактива на целлюлозу используют хлор-цинк-иод, вызывающий ее посинение1. Он представляет собой раствор иода в воднод растворе хлористого цинка. Действие реактива основано на том, что насыщенный раствор хлористого цинка, входящего в его состав, реагируя с молекулой клетчатки, гидролизу-ет ее, доводя до состояния амилоида, близкого по свойствам к крахмальному клейстеру. Амилоидное соединение, взаимодействуя с иодом, входящим в состав реактива, дает уже известное синее окрашивание. Препарат смотрят в реактиве. Следует иметь в виду, что при действии хлор-цинк-иода оболочка сильно набухает.
Задание. 1. При большом увеличении микроскопа зарисовать волокно льна в поперечном разрезе, отметить очертание клетки, утолщенную слоистую оболочку, полость клетки с остатками содержимого.2. На тотальном препарате при большом увеличении микроскопа зарисовать участок волокна в оптическом разрезе и с поверхности, отметив слоистость и полоса-тость оболочки, а также полость клетки с остатками содержимого.
Рис. 3. Волокно льна. А — окончание волокна; Б— оптическое сечение участка волокна; В — участок волокна с поверхности: п. к. — полость клетки, о. к. — оболочка клетки, с. к. — содержимое клетки, сл — слоистость, пс — полосатость
Подобное строение оболочек волокон можно наблюдать в стеблях барвинка, конопли, крапивы и некоторых других волокнистых растений.
Оболочки каменистых клеток околоплодника груши (Pyrus communis L.)
С возрастом в оболочке происходят химические изменения, обусловленные появлением некоторых веществ, придающих ей новые свойства. Очень распространена инкрустация оболочки лигнином, вызывающим ее одревеснение.
Паренхимные клетки с очень толстой одревесневшей оболочкой называют каменистыми. С их строением можно ознакомиться на срезах мякоти плодов дикой груши или незрелых плодов культурной груши плохих сортов.
С фиксированного спиртом или свежего околоплодника делают тонкий небольшой срез, который кладут в каплю воды на предметное стекло. Срезы лучше делать с более твердых участков плода, под кожицей или ближе к середине. Направление разреза значения не имеет.
При малом увеличении видно, что среди тонкостенных, богатых клеточным соком клеток мякоти расположены более мелкие толстостенные клетки. Нужно выбрать участок, где эти клетки лежат поодиночке или небольшими группами. Значительная толщина оболочек каменистых клеток объясняется мощным развитием вторичной оболочки, которая откладывается слоями на первичную. Полость клетки постепенно уменьшается, а ее размеры и форма не меняются. После окончания роста в толщину оболочка одревесневает. При этом она теряет эластичность и упругость, но приобретает твердость и прочность. Эти свойства важны для клеток, несущих механическую и опорную функции. Одревеснение каменистых клеток сопровождается отмиранием их протопласта.
Для распознавания одревесневших оболочек существуют специальные цветные реакции, которыми широко пользуются в анатомии растений. Одревесневшую оболочку можно выявить реакцией со спиртовым раствором флороглюцина и соляной кислотой.
Реакцию проводят следующим образом. Сняв покровное стекло и оттянув воду фильтровальной бумагой, на срез наносят кайлю 1—0,5%-ного раствора флоро-глюцина в 50%-ном спирте. Через несколько секунд, оттянув раствор фильтровальной бумагой, срез обрабатывают концентрированной соляной кислотой. Флоро-глюцин необходимо удалить со стекла потому, что реакция идет только в присутствии концентрированной кислоты и нельзя допускать ее разбавления. Одревесневшие оболочки приобретают вишнево-красный цвет, слабоодревесневшие выглядят более светлыми (розовыми), а неодревесневшие окраску не изменяют. При действии крепкой соляной кислоты срез просветляется, что значительно облегчает изучение объекта. После проведения реакции кислоту удаляют фильтровальной бумагой и заменяют ее каплей глицерина, после чего срез накрывают покровным стеклом и переходят к рассмотрению объекта (вместо глицерина употреблять воду не рекомендуется, так как в воде довольно быстро обесцвечиваются одревесневшие оболочки).
При работе с кислотой необходимо соблюдать ряд предосторожностей, так как ее пары вредно действуют на металлические части микроскопа, линзы объективов, а также на одежду и организм работающего. В связи с этим посуду с кислотой необходимо держать закрытой, а реакцию проводить на столе, отодвинув предметное стекло от микроскопа.
Цветную реакцию на одревеснение можно также получить, действуя на препараты водным раствором сернокислого анилина (см. с. 220). При этом одревесневшие оболочки приобретают лимонно-желтый цвет.
Если срез очень толстый и каменистые клетки лежат в несколько слоев, то, сняв покровное стекло и отыскав невооруженным глазом группы окрашенных каменистых клеток, можно раздавить их скальпелем.
В зависимости от установки трубы микроскопа каменистые клетки можно видеть либо с поверхности, либо в оптическом разрезе. Если клетку рассматривать в разрезе, то в толще ее оболочки иногда заметна слоистость, параллельная поверхности клетки. Оболочка пронизана ветвящимися поров ы ми каналами: канал, начинающийся в полости клетки и проходящий через толщу вторичной оболочки, нередко разделяется на 2—3 канала, доходящих до первичной оболочки. Такое «ветвление» на самом деле представляет собой результат слияния отдельных поровых каналов, происходящего в процессе утолщения оболочки.
Если рассматривать поры в смежных стенках клеток, то можно видеть, что поровый канал одной клетки служит продолжением такого же порового канала соседней клетки. Их разделяет тонкая мембрана.
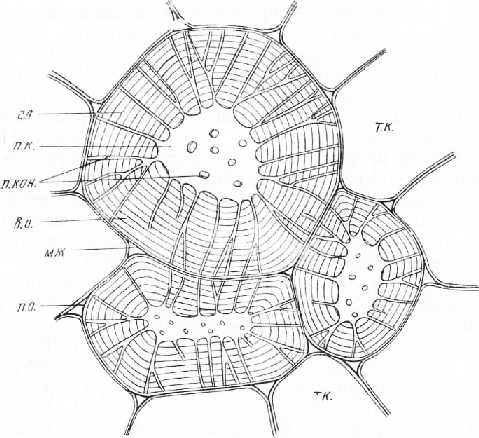
Рис. 2. Каменистые клетки из околоплодника груши: п. о. — первичная оболочка, в. о. — вторичная оболочка, п. кан. — поровые каналы в плане и в разрезе, мж — межклетники, т. к. — тонкостенные клетки, п. к. — полость клетки, сл — слои вторичной оболочки
На препарате часто бывают видны каналы, как бы обрывающиеся в толще оболочки. Это объясняется тем, что поры проходят не строго по радиусу, а изгибаются и оказываются вне плоскости оптического сечения. Продолжение этих поровых каналов можно увидеть, опуская или поднимая трубу микроскопа микрометрен-ным винтом. Поровые каналы в плане выглядят как небольшие овальные или округлые отверстия, открывающиеся в полость клетки. Диаметр отверстий соответствует ширине канала.
Если клетку рассматривать с поверхности, то полость клетки и слоистость оболочки незаметны, а поровые каналы, пронизывающие всю верхнюю (или нижнюю) стенку, видны в плане.
Задание. При большом увеличении микроскопа зарисовать: а) две-три каменистые клетки в оптическом разрезе и с поверхности, отметив в них первичную оболочку, слоистую вторичную оболочку, поры в плане и в разрезе; б) тонкостенные клетки мякоти плода, окружающие группы каменистых клеток.
Начать рисунок удобнее с контуров каменистых клеток, наметив мембрану, состоящую из двух первичных оболочек и межклеточного вещества, затем показать очертания полостей клеток, после чего можно перейти к зарисовке пор в плане и в разрезе. Поры в разрезе изображают в виде тонких каналов, наружные концы которых упираются в первичную оболочку, а внутренние открываются в полость клетки. Слоистость, если она видна, можно показать тонкими линиями.
Оболочки каменистых клеток скорлупы ореха лещины обыкновенной (Corylus avellana L.)
Околоплодник (скорлупа) ореха лещины состоит из каменистых клеток с сильно утолщенными одревесневшими оболочками, в которых хорошо видна слоистость. Так как сделать тонкий срез такого околоплодника практически невозможно, пользуются мацерированным материалом.
Небольшие кусочки скорлупы осторожно кипятят в пробирке с азотной кислотой в течение нескольких минут. В результате сильного окислительного процесса разрушается склеивающее клетки межклеточное вещество, происходит мацерация. Реакция протекает очень бурно, с выделением окислов азота в виде бурых паров. Пробирку периодически снимают с огня и встряхивают, следя за тем, чтобы пенящаяся жидкость не выплеснулась. Работать следует в вытяжном шкафу, так как выделяющиеся окислы азота ядовиты. Для ускорения реакции в азотную кислоту можно добавить несколько кристалликов бертолетовой соли (КС103).
После мацерации кислоту сливают, материал несколько раз промывают водой и хранят в спирте.
Каплю взвеси, состоящую из отдельных мацерированных клеток или их небольших групп, наносят на предметное стекло и добавляют воду. Клетки рассматривают сначала при малом, а затем при большом увеличениях микроскопа.
Клетки могут быть округлыми, продолговатыми, ромбическими, несколько разветвленными и т. д. Толщина оболочки у разных клеток неодинакова. В клетках с сильно утолщенными оболочками полости очень малы.
В некоторых клетках хорошо видна слоистость вторичной оболочки, пронизанной многочисленными узкими поровыми каналами. Их настолько много, что оболочка кажется исчерченной в радиальном направлении. Так выглядит клетка в оптическом разрезе. Если клетку рассматривать с поверхности, то поры видны в плане; они имеют округлые или щелевидные очертания.
Задание. При большом увеличении зарисовать две-три клетки, отметить слоистость оболочки и показать вид поровых каналов при разной установке трубы микроскопа. На рисунке число поровых каналов можно сократить.
Объектами для изучения строения оболочки каменистых клеток могут служить также клетки мякоти плодов айвы, рябины, мацерированной скорлупы (внутриплодника) грецких орехов, сливы, вишни, абрикоса и т. п.
Анатомия растений - Оболочкаgardenweb.ru
Клеточная стенка (оболочка) растений
Если выделить любую клетку из организма животного и поместить ее в воду, то через короткое время клетка после набухания лопнет, лизируется. Это происходит из-за того, что через плазматическую мембрану вода будет поступать в цитоплазму, в зону с более высокой концентрацией солей и органических молекул. При этом будет увеличиваться внутренний объем клетки до тех пор, пока не разорвется плазматическая мембрана. В составе организма животных этого не происходит, потому что клетки низших и высших животных существуют в окружении жидкостей внутренней среды, концентрация солей и веществ в которой близка к таковой в цитоплазме. Свободноживущие в пресной воде одноклеточные простейшие организмы не лизируются (при отсутствии клеточной стенки) из-за того, что у них постоянно работает клеточный насос, откачивающий воду из цитоплазмы, - сократительная вакуоль.
Если же мы в воду поместим клетки бактерий или растений, то они не будут лизироваться до тех пор, пока цела их клеточная стенка. Воздействием набора различных ферментов эти стенки можно растворить. В этом случае моментально происходит набухание и разрыв, лизис, клеток. Следовательно, в естественных условиях клеточная стенка предотвращает этот гибельный для клетки процесс. Более того, наличие клеточных стенок является одним из главных факторов, регулирующих поступление воды в клетку. Клетки бактерий и растений обитают чаще всего в гипотонической водной среде, они не имеют сократительных (выделительных) вакуолей, чтобы откачать воду, но зато прочная клеточная стенка предохраняет их от чрезвычайного набухания. По мере поступления воды в клетке возникает внутреннее давление, тургор, которое препятствует дальнейшему поступлению воды.
Интересно, что у многих низших растений, например у зеленых водорослей, клетки имеют хорошо сформированную клеточную оболочку, но при половом размножении, когда образуются подвижные зооспоры, последние теряют клеточную оболочку и у них появляются пульсирующие вакуоли.
Клеточная стенка растений формируется при участии плазматической мембраны и является экстраклеточным (внеклеточным) многослойным образованием, защищающим поверхность клетки, служащим как бы наружным скелетом растительной клетки (рис. 158). Клеточная стенка растений состоит из двух компонентов: аморфного пластичного гелеобразного матрикса (основы) с высоким содержанием воды и опорной фибриллярной системы. Часто для придания свойств жесткости, несмачиваемости и др. в состав оболочек входят дополнительные полимерные вещества и соли.
В химическом отношении главные компоненты оболочек растений относятся к структурным полисахаридам.
В состав матрикса оболочек растений входят гетерогенные группы полисахаридов, растворяющиеся в концентрированных щелочах, гемицеллюлозы и пектиновые вещества. Гемицеллюлозы представляют собой ветвящиеся полимерные цепи, состоящие из различных гексоз (глюкоза, манноза, галактоза и др.), пентоз (ксилоза, арабиноза) и уроновых кислот (глюкуроновая и галактуроновая кислоты). Эти компоненты гемицеллюлоз сочетаются между собой в разных количественных отношениях и образуют разнообразные комбинации. Цепи гемицеллюлозных молекул не кристаллизуются и не образуют элементарных фибрилл. Из-за наличия полярных групп уроновых кислот они сильно гидратированы.
Пектиновые вещества - гетерогенная группа, в которую входят разветвленные сильно гидратированные полимеры, несущие отрицательные заряды из-за множества остатков галактуроновой кислоты. Благодаря свойствам своих компонентов матрикс представляет собой мягкую пластическую массу, укрепленную фибриллами.
Волокнистые компоненты клеточных оболочек растений состоят обычно из целлюлозы, линейного, неветвящегося полимера глюкозы. Молекулярный вес целлюлозы варьирует от 5 х 104 до 5 х 105, что соответствует 300-3000 остаткам глюкозы. Такие линейные молекулы целлюлозы могут соединяться в пучки или волокна. В клеточной оболочке целлюлоза образует фибриллы, которые состоят из субмикроскопических микрофибрилл толщиной до 25 нм, которые в свою очередь состоят из множества параллельно лежащих цепей молекул целлюлозы.
Количественные соотношения целлюлозы к веществам матрикса (гемицеллюлозы) могут быть весьма различными у разных объектов. Свыше 60% сухого веса первичных оболочек составляет их матрикс и около 30% приходится на скелетное вещество - целлюлозу. В сырых клеточных оболочках почти вся вода связана с гемицеллюлозами, поэтому вес основного вещества в набухшем состоянии достигает 80% сырого веса всей оболочки, тогда как содержание волокнистых веществ сводится всего к 12%. В случае же другого примера, волоски хлопчатника, целлюлозный компонент составляет 90%; в древесине целлюлоза составляет 50% от компонентов клеточной стенки.
Кроме целлюлозы, гемицеллюлозы и пектинов в состав клеточных оболочек входят дополнительные компоненты, придающие им особые свойства. Так, инкрустация (включение внутрь) оболочек лигнином (полимер кониферилового спирта) приводит к одревеснению клеточных стенок, повышению их прочности (рис. 159). Лигнин замещает в таких оболочках пластические вещества матрикса и играет роль основного вещества, обладающего высокой прочностью. Часто матрикс бывает укреплен минеральными веществами (SiO2, CaCO3 и др.).
На поверхностях клеточной оболочки могут скапливаться различные адкрустирующие вещества, например кутин и суберин, приводящие к опробковению клеток. В клетках эпидермиса на поверхности клеточных оболочек откладывается воск, который образует водонепроницаемый слой, препятствующий потере клеткой воды.
Из-за своего пористого, рыхлого строения клеточная стенка растений проницаема в значительной степени для низкомолекулярных соединений, таких как вода, сахара и ионы. Но макромолекулы проникают через целлюлозные оболочки плохо: величина пор в оболочках, позволяющая свободную диффузию веществ составляет всего лишь 3-5 нм.
Опыты с мечеными соединениями показали, что при росте клеточной оболочки выделение веществ, из которых она строится, происходит по всей поверхности клетки. Аморфные вещества матрикса, гемицеллюлозы и пектины синтезируются в вакуолях аппарата Гольджи и выделяются через плазмолемму путем экзоцитоза. Фибриллы целлюлозы синтезируются специальными ферментами, встроенными в плазмолемму.
Оболочки дифференцированных, зрелых, клеток обычно многослойные, в слоях фибриллы целлюлозы ориентированы по-разному, и количество их также может значительно колебаться. Обычно описывают первичные, вторичные и третичные клеточные оболочки (рис. 158). Для того чтобы разобраться в строении и появлении этих оболочек, необходимо познакомиться с тем, как же возникают, образуются клеточные оболочки после деления клеток.
При делении клеток растений после расхождения хромосом в экваториальной плоскости клеток появляется скопление мелких мембранных пузырьков, которые в центральной части клеток начинают сливаться друг с другом (рис. 160). Этот процесс слияния мелких вакуолей происходит от центра клетки к периферии и продолжается до тех пор, пока мембранные пузырьки не сольются между собой и с плазматической мембраной боковой поверхности клетки. Так образуется клеточная пластинка или фрагмопласт. В центральной части ее располагается аморфное вещество матрикса, которое наполняло сливающиеся пузырьки. Доказано, что эти первичные вакуоли происходят от мембран аппарата Гольджи. В состав первичной клеточной стенки входит также небольшое количество белка (около 10%), богатого гидроксипролином. имеющего множество коротких олигосахаридных цепей, что определяет этот белок как гликопротеид. По периферии клеточной пластинки при наблюдении ее в поляризованном свете обнаруживается заметное двойное лучепреломление, вызванное тем, что в этом месте располагаются ориентированные фибриллы целлюлозы. Таким образом, растущая первичная клеточная стенка состоит уже из трех слоев: центральный - срединная пластинка, состоящая только из аморфного матрикса, и два периферических - первичная оболочка, содержащая гемицеллюлозу и целлюлозные фибриллы. Если срединная пластинка - это продукт активности еще исходной клетки, то первичная оболочка образуется за счет выделения гемицеллюлозы и фибрилл целлюлозы уже двумя новыми клеточными телами. И все дальнейшее увеличение толщины клеточной (вернее, межклеточной) стенки будет происходить за счет активности двух дочерних клеток, которые с противоположных сторон будут выделять вещества клеточной оболочки, утолщающейся путем подслаивания все новых и новых пластов. Так же как и с самого начала, выделение веществ матрикса происходит за счет подхода к плазматической мембране пузырьков аппарата Гольджи, слияния их с мембраной и высвобождения их содержимого за пределы цитоплазмы. Здесь же вне клетки на ее плазматической мембране идет синтез и полимеризация целлюлозных фибрилл. Так постепенно образуется вторичная клеточная оболочка. С достаточной точностью определить и суметь отличить первичную оболочку от вторичной трудно, так как они соединены между собой несколькими промежуточными слоями.
Основную массу закончившей свое формирование клеточной стенки составляет вторичная оболочка. Она придает клетке ее окончательную форму. После разделения клетки на две дочерних происходит рост новых клеток, увеличение их объема и изменение формы; клетки часто вытягиваются в длину. Одновременно с этим идет наращивание толщины клеточной оболочки и перестройка ее внутренней структуры.
При образовании первичной клеточной оболочки в ее составе еще мало целлюлозных фибрилл, и они располагаются более или менее перпендикулярно будущей продольной оси клетки, позже в период растяжения (удлинения клетки за счет роста вакуолей в цитоплазме) ориентация этих поперечно-направленных фибрилл подвергается пассивным изменениям: фибриллы начинают размещаться под прямым углом друг к другу и в конечном счете оказываются вытянутыми более или менее параллельно продольной оси клетки. Постоянно идет процесс: в старых слоях (ближе к центру оболочки) фибриллы подвергаются пассивным сдвигам, а отложение новых фибрилл во внутренних слоях (ближайших к мембране клетки) продолжается в соответствии с исходным планом конструкции оболочки. Этот процесс создает возможность скольжения фибрилл относительно друг друга, а перестройка арматуры клеточной оболочки возможна из-за студенистого состояния компонентов ее матрикса. В дальнейшем при замещении в матриксе гемицеллюлозы на лигнин подвижность фибрилл резко снижается, оболочка становится плотной, происходит одревеснение.
Часто под вторичной оболочкой обнаруживают третичную оболочку, которую можно рассматривать как засохший остаток дегенерировавшего слоя собственно цитоплазмы.
Следует отметить, что при делении клеток растений формированию первичной оболочки не во всех случаях предшествует образование клеточной пластинки. Так, у зеленой водоросли спирогиры новые поперечные перегородки возникают путем образования на боковых стенках исходной клетки выступов, которые, постепенно разрастаясь к центру клетки, смыкаются и делят клетку надвое.
Как уже говорилось, если в водной гипотонической среде лишить клетку ее оболочки, то произойдет лизис, разрыв клетки. Оказалось, что, подбирая соответствующие концентрации солей и сахаров, можно уравнять осмотические давления снаружи и внутри клеток, лишенных своих оболочек. При этом такие протопласты приобретают шаровидную форму (сферопласты). Если в среде, где находятся протопласты, будет достаточное количество питательных веществ и солей (среди них необходим Ca2+), то клетки снова восстанавливают , регенерируют свою клеточную оболочку. Более того, они способны в присутствии гормонов - ауксинов - делиться и создавать клеточные колонии, которые могут дать начало для роста целого растения, от которого была взята клетка.
Главный волокнистый компонент клеточной стенки больших групп грибов (базибиомицеты, аскомицеты, зигомицеты) - хитин, полисахарид, где основным сахаридом является N-ацетилглюкозамин. В состав клеточной стенки грибов кроме хитина могут входить вещества матрикса, гликопротеиды и различные белки, синтезированные в цитоплазме и выделенные клеткой наружу.
Клеточные оболочки бактерий
Опорным каркасом клеточной стенки бактерий и синезеленых водорослей также служит в значительной степени однородный полимер - пептидогликан или муреин. Жесткий каркас, окружающий бактериальную клетку, представляет собой одну гигантскую мешковидную молекулу сложного полисахарида-пептида. Каркас этот называют муреиновым мешком. Основа структуры муреинового мешка - сеть параллельных полисахаридных цепей, построенных из чередующихся дисахаридов (ацетилглюкозамин, соединенный с ацетилмурамовой кислотой), связанных многочисленными пептидными поперечными связями (рис. 161). Длина цепочек гликана может быть огромной - до нескольких сот дисахаридных блоков. Основу пептидной части муреина составляют тетрапептиды, образованные различными аминокислотами.
Бактериальная стенка может составлять до 20-30% от сухого веса бактерии. Это связано с тем, что в ее состав кроме многослойного муреинового каркаса входит большое количество дополнительных компонентов, как и в матриксе стенки растений. У грамположительных бактерий (при окраске по Граму (окраска кристаллическим фиолетовым, обработка иодом, отмывка спиртом) бактерии по-разному воспринимают краситель: грамположительные остаются окрашенными после обработки спиртом; грамотрицательные обесцвечиваются). сопутствующими компонентами служат полимерные вещества, сложным образом вплетенные в муреиновую сеть. К ним относятся тейхоевые кислоты, полисахариды, полипептиды и белки. Клеточная стенка грамположительных бактерий обладает большой жесткостью, ее муреиновая сеть многослойна.
Стенки грамотрицательных бактерий содержат однослойную муреиновую сеть, составляющую 12% сухой массы стенки. Сопутствующие компоненты составляют до 80% сухой массы. Это липопротеиды, сложные липополисахариды. Они образуют сложную наружную липопротеиновую мембрану. Следовательно, периферия грамотрицательных бактерий содержит наружную мембрану, затем однослойную муреиновую сеть, ниже нее расположена плазматическая мембрана (рис. 162). Наружная мембрана обеспечивает структурную целостность клетки, служит барьером, ограничивающим свободный доступ разных веществ к плазматической мембране. На ней также могут располагаться рецепторы для бактериофагов. Она содержит белки-порины, которые участвуют в переносе многих низкомолекулярных веществ. Молекулы порина образуют тримеры, проходящие сквозь толщу мембраны. Одна из функций этих белков - формирование в мембране гидрофильных пор, через которые происходит диффузия молекул, не более 900 дальтон. Через поры проходят свободно сахара, аминокислоты, небольшие олигосахариды и пептиды. Поры образованы разными поринами, обладают разной проницаемостью.
Между внешней липопротеидной мембраной бактериальной стенки и плазматической мембраной лежит периплазматическое пространство или периплазма. Ее толщина обычно составляет около 10 нм, она содержит тонкий (1-3 нм) муреиновый слой и раствор, содержащий специфические белки двух типов - гидролитические ферменты и транспортные белки. Из-за наличия гидролаз иногда периплазму рассматривают как аналог лизосомного компартмента эукариот. Периплазматические транспортные белки связывают и переносят сахара, аминокислоты и др. от внешней мембраны к плазмолемме.
Предшественники стенок бактерий синтезируются внутри клетки, сборка стенок происходит снаружи от плазматической мембраны.
Под действием фермента лизоцима можно разорвать муреиновый каркас и растворить бактериальную стенку. В гипотонических условиях клетки при этом разрушаются, как разрушаются голые клетки животных и растений; в изотонических условиях образуются шаровидные протопласты, которые способны снова вырабатывать свою клеточную стенку.
Интересно, что протопласты бактерий нечувствительны к действию бактериофагов- вирусов, паразитирующих на бактериях. Следовательно, компоненты бактериальной стенки обладают антигенной специфичностью по отношению к этим вирусам.
studfiles.net
КЛЕТОЧНАЯ ОБОЛОЧКА
Оболочки клеток в основном представляют собою гели веществ, находящихся в коллоидном состоянии. Клеточные оболочки состоят из веществ, не растворимых в воде, но способных впитывать воду и растворенные в ней вещества и набухать при этом до известного предела.
СОСТАВ И СТРОЕНИЕ КЛЕТОЧНОЙ ОБОЛОЧКИ
Целлюлоза. В большинстве случаев клеточные оболочки у вычеши х растений состоят преимущественно из целлюлозы1, или клетчатки — углевода типа полисахарида с эмпирической формулой С6h20O5.
Целлюлоза анизотропна, т. е. показатели её физических свойств не по всем направлениям одинаковы. Анизотропность может быть показана на конкретном примере волокон льна: после набухания в воде поперечные размеры их увеличиваются на 20%, а длина лишь на 0,1%. Анизотропны - клеточные оболочки по упругости и прочности, по оптическим свойствам и т. д.
Целлюлоза в клеточной оболочке — сложно организованное высокомолекулярное вещество тонкокристаллической структуры, находящейся за пределами видимости в микроскоп. Картину строения, целлюлозы удалось выяснить с помощью комплекса исследований биохимических и оптических, с использованием микроскопа, с применением поляризационного микроскопа и методов рентгеноскопии и рентгенографии.
Сущность выводов из исследований сводится к следующему.
Основными структурными единицами целлюлозы являются кристаллиты, или мицеллы. Схематическая формула мицеллы имеет вид:
[(С6Н10О5)·х]·у,
где у (коэффициент ассоциации) — число цепочек в мицелле, а х (коэффициент полимеризации) — число звеньев в цепочке.
Цепочка (макромолекула) целлюлозы состоит из глюкозных остатков (С6Н10О5), соединенных по типу глюкозидов главными валентностями в ряд. Каждый из остатков глюкозы повернут на 180° относительно соседних.
1 От латинского слова cellula--клетка.
Раньше полагали, что коэффициент полимеризации (х) целлюлозы равен приблизительно 660, затем принимали его равным 5,4*80° относительно соседних.10. Исследования О. П. Головой, В. М. Иванова и И. Н. Николаевой (1948) и затем О. П. Головой и В. И. Иванова (1949)1 дали для молекулярного веса целлюлозы в эксперименте величину 1,62*106, т. е. в З раза более высокую 2. Авторы вычислили путем экстраполирования, что при идеально безупречной постановке эксперимента молекулярный вес оказался бы в 1,5 раза выше (2,43*106).
Величину коэффициента ассоциации (у) считают равной 40 — 60, т. е. полагают, что 40—60 цепей .(макромолекул) целлюлозы, расположенных параллельно и ассоциированных силами остаточного (побочного) сродства и пучок, составляют одну целлюлозную мицеллу.
Схема элементарной кристаллической ячейки целлюлозы преде топлена на рис. 14.
Мицеллы, числом до 100, группируются в мицеллярные ряды с поперечными размерами около 60mµ. Несколько сот мицеллярных рядов составляют фибриллу. Поперечные размеры фибрилл таковы, что фибриллы доступны микроскопическому видению при известной обработке препарата и при достаточно сильной оптической части микроскопа.
Например, диаметр фибрилл набухших волокон льна, применяющихся в текстильном деле, составляет 0,3—0,6µ.
Целлюлоза относится к индифферентным веществам: она не обладает ни кислотными, ни щелочными свойствами. Она стойка по отношению к тепловым воздействиям и может быть нагрета без разложения до + 200°С и насколько выше. Целлюлоза не растворима в воде, в спирте, в эфире и в других нейтральных растворителях. Не растворяется она при нормальном давлении и при обычных температурах в разведенных кислотах и щелочах.
1 О. П. Голова и В. И. Иванов, О молекулярном весе целлюлозы. Академия наук СССР. Институт органической химии. Изд. АН СССР, М. — Л. 1949.
2 Определение молекулярного веса целлюлозы производилось по вязкости ее медно-аммиачных растворов, с многократным откачиванием системы в высоком вакууме и промывкой ее инертным иолом наивысшей очистки.
Если целлюлоза и растворяется при известных условиях под действием кислот и щелочей, то это растворенке сспровождается более или менее сильным разрушением ее организации либо даже гидролизом ее вещества.
Без разрушения мицелл целлюлоза может быть растворена в швейцеровом реактиве — аммиачной растворе окиси меди. Целлюлоза, осажденная еодсй из раствора в реактиве Швейцера (гидрат-целлюлоза), представляет собою набухший гель с беспорядочным расположением мицелл. При изесстной физико-механической обработке расположение мицелл может быть упорядочено.
Известны две кристаллические формы целлюлозы: одна, — названная природной, и другая — «мерсеризованная».
Мерсеризованная неллюлеза отличается, по данным рентгеноскопического анализа, тем, что глюкозкые остатки в ее цепях повернуты, по сравнению с «природной» клетчаткой, на 30°, и кристаллические ячейки имеют иную форму. Природная целлюлоза сравнительно неустойчива (мстастабильна) в физико-химическом смысле слова, мерсеризованная же более устойчива (стабильна).
Сравнительно недавно обнаружено, что и в природе можно встретить «мерсеризованную» целлюлозу. Она была найдена в оболочках зеленой морской водоросли галицистис (Halicystis cvalis).
Полуклетчатки (гемицеллюлозы). Нередко в состав клеточных стенок растений входят вещества, относящиеся к так называемым полуклетчаткам (гемицеллюлозы). Это— углеводы, а именно—твердые полисихариды гексозаны (С6Н10О5)n и пентозаны (С5Н8О4)n.
Строение гемицеллюлоз аналогично строению целлюлозы: они имеют цепные молекулы из остатков гексоз либо пентоз, связанных кислородными мостиками. Гемицеллюлозы гораздо менее стойки по отношению к химическим воздействиям, нежели целлюлозы.
Гемицеллюлозы в организме растения играют 1) механическую роль, входя в состав клеточных стенок, и 2) роль запасных веществ, откладывающихся и затем расходующихся.
В качестве запасных питательных веществ гемицеллюлозы отлагаются в клеточных стенках семян многих растений — пальм (финиковой пальмы, пальмы Phylelephas macrocarpa)1, лилейных (птицемлечника, рябчика), бобовых (цареградских рожков — Сегаtonia siliqua, люпинов) и т. д.
Полуклетчатки, не всегда используемые в качестве запасных веществ, имеются в древесине и хвойных, и лиственных пород в в количестве, составляющем 20 — 25% сухого веса клеточных стенок.
Пектиновые вещества. Целлюлозные клеточные стенки строятся, как отмечено выше, из мицелл и пучков их, примыкающих друг к другу не вплотную, а с промежутками, поперечные размеры которых имеют геличину порядка 10 mµ. Промежутки нередко заполняются пектиноными веществами. Химики определяют эти вещества как «кальциевые-магниевые соли полимерной а-галактуроновой кислоты, у которых часть водородов карбоксильных групп замещена метиловыми радикалами»1.
1 Семена этой пальмы, очень крупные и твердые («растительная слоновая кость») используются для вытачивания из них пуговиц.
В клеточных стенках содержатся пектиновые вещества с весьма сложной молекулой; в воде они не растворяются, а только набухают. По отношению к химическим агентам они менее стойки, нежели целлюлоза: легко разрушаются при действии щелочей, а после обработки кислотами легко растворяются в кислотах.
Пектиновые вещества клеточных стенок широко распространены в растительном царстве, в частности — у высших растений. Клеточные стенки в ранней стадии развития состоят в преобладающей мере из пектинозых веществ. Межклеточное вещество, как бы цементирующее оболочки соседних клеток, оэычно состоит главным образом из пектиновых веществ, преимущественно из пектата кальция.
Пектиновые вещества, хотя бы в очень небольших количествах, имеются и в толще взрослых клеточных оболочек.
Пектиновые вещества межклетных пластинок нередко претерпевают химические изменения и под действием энзимов разрушаются, после чего клетки разъединяются: такое явление происходит, например, при дозревании сочных плодов (арбузов, груш и иных). На разрушении межклеточного вещества под действием энзима пектиназы, вырабатываемой некоторыми бактериями и грибками, основана мочка льна. Такое разрушение межклеточного вещества и разъединение тканей на отдельные клетки носит название мацерации.
Фитомеланы. Весьма интересны как чрезвычайно богатые углеродом вещества клеточных стенок, названные фитомеланами. Фитомеланы — безазотистые органические вещества, в которых содержится водород и кислород в том же, приблизительно, соотношении, как в углеводах, и углерод в количестве 69 — 76%. Фитомеланы чрезвычайно стойки по отношению к реактивам.
Фитомеланы найдены у некоторых растений из семейства сложноцветных: в скорлупе семянок (подсолнечника, георгины, дурнишника, череды и других), в листочках обвертки одчого из бессмертников (Аmmоbium), в корнях рудбекии. В скорлупе семянок оболочки несколько слогв клеток иод кожицей превращаются в «угольный слой», темнобурую или черную фитомелановую массу.
Борисовым иРаздорским (1931) фитомеланы были обнаружены в корнях василька иволистного (Centaureu salocifolla).
www.botanik-learn.ru