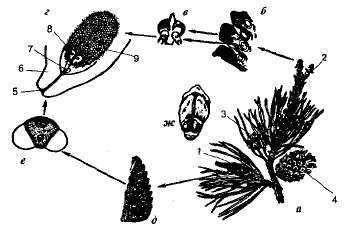
Строение растительной клетки Ткани растений. Клетки растений могут ли иметь жгутики
Жгутик — Википедия
Материал из Википедии — свободной энциклопедии
Жгу́тик — поверхностная структура, присутствующая у многих прокариотических и эукариотических клеток и служащая для их движения в жидкой среде или по поверхности твёрдых сред. Жгутики прокариот и эукариот принципиально различаются: бактериальный жгутик имеет толщину 10—20 нм и длину 3—15 мкм, он пассивно вращается расположенным в мембране мотором; жгутики же эукариот толщиной до 200 нм и длиной до 200 мкм, они могут самостоятельно изгибаться по всей длине. У эукариот часто также присутствуют реснички, идентичные по своему строению жгутику, но они двигаются по-другому: жгутик "вкручивается" в жидкость, а ресничка совершает волнообразные колебания (вперёд-назад).
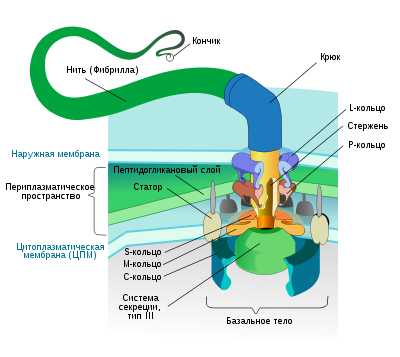
Жгутикование бактерий: A — монотрихиальное, B — лофотрихиальное, C — амфитрихиальное, D — перитрихиальное.Жгутики бактерий состоят из трёх субструктур:
- Филамент (фибрилла, пропеллер) — полая белковая нить толщиной 10—20 нм и длиной 3—15 мкм, состоящая из флагеллина, субъединицы которого уложены по спирали. Полость внутри используется при синтезе жгутика — он происходит в направлении от плазматической мембраны. По полости к собираемому в настоящий момент участку переносятся субъединиц
ru.wikipedia.org
Микротельца, жгутики и реснички относятся к какой клетке?
otvet.mail.ru
Жгутики эукариот - стр. 17
7. Жгутики эукариот
Почти у всех эукариот хотя бы некоторые клетки имеют реснички или жгутики. Они представляют собой органы плавания клетки в жидкой среде (у одноклеточных форм и стадий и у не очень крупных многоклеточных) или, наоборот, перемещения жидкой среды вдоль клетки (у многоклеточных форм). Что, по сути, есть одно и то же. Отличие ресничек от жгутиков состоит в длине и характере движения. Ресничка короткая, ее движение состоит из резкого изгиба у основания при сохранении прямизны по остальной длине (при этом она действует как весло), сменяющейся медленным изгибом основания в обратном направлении и расслабленного состояния основной длины (так что она принимает форму, максимально ослабляющую сопротивление жидкости). Как правило, ресничек много и их движения скоординированы и волнообразны. Жгутики длинны и совершают постоянные волнообразные движения. Диаметр жгутиков или ресничек – около 0,25 мкм, а длина варьирует от 10 до 200 мкм.
Структура и у жгутиков, и у ресничек одинаковая и весьма стандартная. В ее основе лежит красивая структура из микротрубочек (которые мы рассмотрели выше) – по периметру расположено девять пар сдвоенных микротрубочек (дублетов) и еще две отдельные микротрубочки проходят по центру образованного ими цилиндра (рис. 8.6). При этом обе центральные микротрубочки полноценны, в то время как в боковых парах только одна микротрубочка полная, другая же пристроена к ней и в месте контакта не имеет собственной стенки.
С интервалом в 24 нм от боковых дублетов по направлению к соседнему дублету отходят «ручки», состоящие из одной молекулы белка динеина, другой белок образует эластичные связки между дублетами. От них же по направлению к центральной паре микротрубочек отходят «спицы», состоящие из другого белка. Движение жгутиков обеспечивается скольжением боковых дублетов друг относительно друга, за которое ответствен белок динеин, который, гидролизуя АТФ, смещается относительно той микротрубочки, к которой он направлен.
В основании любого жгутика или реснички лежит базальное тельце, или базонема, совершенно идентичная центриоли, от которой у животных отходят цитоплазматические микротрубочки.
Неслучайно структура центриоли в чем-то аналогична структуре жгутика – и там и тут мы имеем девять продольных структур, образованных микротрубочками. Подобно тому как центриоль служит затравкой для построения цитоплазматических микротрубочек, базальное тельце служит затравкой для построения жгутика. Однако жгутик отходит не от некоего недифференцированного вещества возле него, а непосредственно от базального тельца. Микротрубочки боковых дублетов отходят от плюс-концов двух из трех микротрубочек каждого бокового триплета базального тельца. Центральная пара микротрубочек образуется непонятным пока образом без видимой затравки. Сам жгутик можно оторвать, и он восстановится из базального тельца. Но без базального тельца жгутик не возникает. Базальные тельца формируются около центриолей клеточного центра. В случае ресничек у человека вокруг каждой материнской центриоли возникают ювенильные базальные тельца. Каким-то образом центриоль служит матрицей для образующихся около нее базальных телец (равно как и для дочерней центриоли при делении клеточного центра). Как и в случае ювенильных центриолей, у них сначала вместо девяти триплетов имеется девять одиночных микротрубочек по периферии. В дальнейшем они дозревают до нормальных базальных телец, по структуре идентичных центриолям.
Базальные тельца и центриоли могут быть идентичны не только по структуре, но и по функции. Зеленый жгутиконосец хламидомонада имеет два жгутика, но перед делением клетки он их теряет, а их базальные тельца функционируют как центриоли при делении клетки (как происходит деление – рассмотрим в следующей лекции).
Жгутики (или реснички) изначально присущи эукариотической клетке. Они есть у многих одноклеточных – у жгутиконосцев (жгутики) и инфузорий (реснички). Их нет у высших грибов и высших (цветковых) растений, но они есть у низших грибов и растений. Почти у всех животных они так или иначе присутствуют в каких-то клетках, как минимум у сперматозоидов. Единственная группа животных, где они полностью утрачены, это круглые черви нематоды. У человека жгутики есть не только в сперматозоидах, но и у клеток так называемого ресничного эпителия, который выстилает полость легких и пазух носа и создает ток приповерхностной слизи (грубо говоря, соплей и мокроты), благодаря чему внутренняя поверхность легких очищается от пыли.
О жгутиках можно рассказать интересный факт. В раннем эмбриональном развитии человека есть так называемая стадия бластулы, когда человек еще представляет собой лишь однослойную сферу из клеток, на которые поделилась оплодотворенная яйцеклетка. Но клетки в ней уже неодинаковы и готовы к дальнейшим превращениям. На этой стадии одна совершенно определенная клетка несет один жгутик, который характерен тем, что в нем нет двух внутренних микротрубочек. Биение этого жгутика создает некоторый ток жидкости вокруг бластулы. Вместе с этим течением в определенную сторону перемещаются и определенные морфогены – вещества, выделяемые одними клетками, чтобы влиять на развитие других. Этим потоком морфогенов задается первоначальная право-левая ассимметрия человеческого тела. У человека есть мутация, нарушающая подвижность жгутиков. Такие люди вполне живут, но у них есть определенные проблемы со здоровьем и наблюдается мужское бесплодие, так как у них неподвижны реснички эпителия нижних дыхательных путей и жгутики сперматозоидов. И, что удивительно, сердце с равной вероятностью находится слева или справа. Как выяснилось, ничего удивительного в этом нет – отсутствие упомянутого жгутика у бластулы проиводит к тому, что тока морфогенов нет и право-левая асимметрия закладывается случайным образом.
8. Жгутики и движение бактерий
Жгутики бактерий не имеют ничего общего со жгутиками эукариот. Они имеют другую химическую основу, другой принцип движения и сами по себе к активному движению не способны – вращается лишь их основание. Жгутики бактерий представляют собой тонкие нити, берущие начало от цитоплазматической мембраны, имеют большую длину, чем сама клетка. Толщина жгутиков 12–20 нм, длина 3–15 мкм. Они состоят из трех частей: спиралевидной нити, крюка и базального тельца, содержащего стержень со специальными дисками (одна пара дисков – у грамположительных и 2 пары – у грамотрицательных бактерий) (рис. 8.7). Дисками жгутики прикреплены к мембране и клеточной стенке. При этом создается эффект электромотора со стержнем-ротором, вращающим жгутик. Жгутики состоят из белка – флагеллина (от лат. flagellum – жгутик). Бактериальный жгутик закручен спиралевидным образом.
Число жгутиков у бактерий различных видов варьирует от одного (например, у холерного вибриона) до десятка и сотен жгутиков, отходящих по периметру бактерии, у кишечной палочки, протея и др., либо же имеется пучок жгутиков на одном из концов клетки.
Рассмотрим движение бактерии со многими жгутиками. Все жгутики одной бактерии непрерывно вращаются, причем в одну и ту же сторону. Однако во времени направление вращения периодически меняется на обратное. При вращении спирально закрученных жгутиков против часовой стрелки они собираются в один общий пучок и создают тягу, толкающую клетку в противоположную сторону, в результате чего она движется поступательно. При вращении в обратном направлении, по часовой стрелке, пучок распадается на отдельные жгутики, которые толкают клетку в разных направлениях, так что она кувыркается на месте хаотическим образом.
Именно за счет чередования этих двух режимов – направленного плавания и кувыркания – бактерия в итоге может двигаться по направлению возрастания концентрации благоприятных ей веществ (например, сахара) либо убывания концентрации неблагоприятных веществ. Механизм такого движения необыкновенно прост (в соответствии с простотой организма), но в то же время весьма эффективен. Бактерия слишком мала, чтобы отслеживать изменения концентрации вдоль длины своей клетки. Но она может отслеживать изменения концентрации во времени. Если концентрация благоприятного вещества со временем нарастает, она проводит больше времени в движении и меньше времени в кувырканиях, тем самым продвигаясь в нужном направлении. Если концентрация нужного вещества во времени убывает, бактерия вскоре останавливает движение и начинает кувыркаться, «в надежде» случайным образом принять более благоприятное направление движения.
Здесь важнее всего то, что бактерия должна реагировать не на определенную концентрацию вещества, а на изменение этой концентрации во времени. Это достигается посредством молекулярной адаптации химических рецепторов (особых трансмембранных белков, связывающихся с интересующими бактерию веществами), расположенных на мембране. Адаптация рецепторов происходит путем обратимого ковалентного присоединения к ним нескольких метильных групп. А регуляция направления вращения жгутиков – за счет фосфорилирования и дефосфорилирования нескольких белков цитоплазмы, которые передают сигнал на белки, приводящие во вращение ротор жгутика.
Лекция 9. ДЕЛЕНИЕ КЛЕТОК И КЛЕТОЧНЫЙ ЦИКЛ
На двух предыдущих лекциях мы с вами рассмотрели строение клетки. Большая часть нашего времени ушла на знакомство с эукариотической клеткой, потому что она действительно гораздо сложнее устроена, чем прокариотическая. Сейчас мы переходим к клеточному делению, и здесь эта разница будет проявляться еще рельефнее.
Деления бактериальной клетки мы уже коснулись. Кольцевая молекула ДНК прикреплена в определенном месте ко внешней клеточной мембране. В ходе репликации молекула ДНК, называемая бактериальной хромосомой, удваивается, причем каждая из двух молекул по прежнему прикреплена к мембране. После этого мембрана между местами прикрепления двух молекул ДНК начинает расти. Одновременно растет и клеточная стенка. Когда обе хромосомы расходятся на достаточное расстояние, мембрана и клеточная стенка образуют перетяжку, которая делит клетку на две.
Деление эукариотической клетки протекает сложнее. Это связано с необходимостью точно поделить между двумя дочерними клетками гораздо большее количество ДНК, да еще подразделенное на многие хромосомы. Деление каждой эукариотической клетки сопровождается впечатляющими событиями, которые видно даже под средненьким световым микроскопом.
Деление эукариотической клетки называется митоз. Однако этот термин относится именно к процессам, наблюдаемым в микроскоп. В промежутках между делениями – митозами – клетка проходит стадии, которые под микроскопом не увидишь, но которые очень важны. Поэтому правильнее будет говорить о митотическом цикле – промежутке времени, включающем один промежуток между делениями клетки и одно клеточное деление (именно в такой последовательности).
«Нормальное» состояние клетки между делениями называется интерфазой. (Смысл термина в том, что это промежуток между делениями.) На самом деле интерфаза делится на три стадии – G1, S и G2 (здесь буква G всего лишь от английского gap – промежуток).
Фаза G1 – это основное рабочее состояние клетки. В этом состоянии идет транскрипция и трансляция, восстановление объема и внутреннего содержания клетки, которые были ополовинены в ходе предыдущего деления, идет размножение пластид и митохондрий (очень похожее на деление бактерий, только без необходимости наращивать клеточную стенку, которую они утеряли). Именно на этой стадии клетки многоклеточного организма выполняют все свои специфические функции, организму необходимые.
S-фаза – это период, когда ДНК в ядре удваивается. Репликация ДНК начинается во многих, однако строго определенных, местах, причем где-то раньше, где-то позже; тем не менее, к концу S-фазы каждая молекула ДНК удваивается полностью. Как вы понимаете, одновременно с ДНК должно удвоиться количество гистонов и негистоновых белков хроматина – появление огромной отрицательно заряженной молекулы ДНК в ядре «в голом виде» было бы физико-химической катастрофой. Поэтому в S-фазе в клетке активно синтезируются гистоны и прочие белки хроматина.
Здесь есть важный момент. Среди белков хроматина имеется очень малая по количеству, но очень разнообразная и важная часть – специфические генные регуляторы - это те белковые репрессоры и активаторы, которые включают и выключают гены. Они связываются с регуляторной областью практически любого эукариотического гена и определяют его выключенное или включенное состояние (т. е. транскрибируется он или нет). Не нужно напоминать, что коль скоро регуляторы – это специальные белки, то каждый из них тоже кодируется своим геном, экспрессия которого также регулируется какими-то регуляторами – так реализованы генные сети управления. Генов – десятки тысяч. Регуляторов меньше, так как каждый включает или выключает многие гены – иначе мы имели бы свой отдельный регулятор на каждый ген и впали бы в дурную бесконечность. Важно подчеркнуть, что каждая клетка многоклеточного организма несет в себе все гены, присущие этому организму, но в каждой конкретной клетке работает только малая часть генов, тогда как остальные нужны в других типах клеток или в другие периоды жизни. Гены включаются и выключаются по мере необходимости, но при делении клеток определенного типа важно, чтобы включенные и выключенные состояния генов, характерные для данного типа, в целом были унаследованы. При репликации ДНК удваивается, и надо, чтобы регуляторные белки не только были дополнительно синтезированы в таком же количестве, как и исходно наличествовавшее, но и сели на свои места. Обычно это достигается посредством обратной связи – такой белок регулирует и свой же собственный ген, поэтому когда этот ген активен, регуляторный белок производится, присутствует в клетке в нужном количестве и связывается с теми регуляторными сайтами, к которым он специфичен. Если активность гена данного регуляторного белка потеряна, то она как правило уже не восстанавливается ввиду отсутствия его продукта. Воспроизведение включенного и выключенного состояния генов при удвоении ДНК может достигаться и за счет кооперативного эффекта, который проявляют регуляторные белки, – наличие молекулы белка, связанной с ДНК, провоцирует в своей непосредственной близости связывание со вновь синтезированной ДНК такого же белка. О подобном явлении – наследовании определенного состояния генов в ряду клеточных поколений без изменения структуры самого гена на уровне ДНК принято говорить как об эпигенетическом наследовании.
И в то же время репликация – это именно тот критический момент, когда многие гены выключаются или включаются в ходе индивидуального развития. В течение G1 среди других белков могут быть синтезированы новые регуляторы, и во время S они могут успешно конкурировать со старыми за вновь синтезированные регуляторные области ДНК. Или, наоборот, бывают недосинтезированы старые регуляторы, в результате созданные заново регуляторные области ДНК оказываются не занятыми или занятыми регуляторами, сродство которых к ним меньше. Регуляторные сигналы могут поступить извне, в виде гормонов и цитокинов (веществ, при помощи которых клетки воздействуют друг на друга на ближних дистанциях) – они связываются с белками-рецепторами, которые за счет этого приобретают свойства генных регуляторов. Кроме того, каждый белок-регулятор в моменты репликации ДНК вынужден конкурировать за те участки вновь синтезируемой ДНК, к которым он специфичен, с гистонами. Мы упоминали «гистоновый код» - набор посттрансляционных модификаций коровых гистонов, определяющий свойства хроматина, в том числе активность находящихся в нем генов, то есть доступность ДНК для белков-регуляторов и элементов транскрипционной машины. Кроме того, некоторые регуляторные белки имеют определенное сходство с линкерным гистоном Н1 и конкурируют за свои сайты связывания на ДНК именно с ним. Так в S-фазе, за счет некоторых изменений в присутствии-отсутсвии регуляторов на регуляторных последовательностях ДНК тех или иных генов, в ходе индивидуального развития многоклеточного организма клетки и приобретают новые свойства.
Наконец, в клетке есть еще одна структура, удваивающаяся именно в S-фазе. Это центросома. В фазе G1 центросома выглядит так, как было описано выше: аморфное образование, внутри него находятся две расположенные перпендикулярно друг другу центриоли (но у растений центриолей нет). Как мы помним, центросома является местом, откуда формируется такой элемент цитоскелета, как микротрубочки. В интерфазе миркотрубочки растут от центросомы по направлению ко всей клеточной периферии. Некоторые из них становятся нестабильными и быстро разбираются на отдельные молекулы тубулина. В конце фазы G1 центриоли расходятся на несколько микрон. А в S-фазе рядом с каждой центриолью строится вторая центриоль, и центросома удваивается.
Следующая – G2 – подготовка к делению. На данной стадии нарабатываются определенные белки. В это время завершается формирование двух центросом, а система интерфазных микротрубочек начинает разрушаться, высвобождая тубулин, из которого микротрубочки состоят. Хромосомы в это время уже начинают дополнительно конденсироваться, но в микроскоп этого пока не видно.
Собственно митоз (М-фаза) также делится на несколько стадий (рис. 9.1). Это кульминация клеточного цикла, протекающая в виде жесткой последовательности событий, которые достаточно драматичны даже при простом взгляде в микроскоп. В солидном учебнике клеточной биологии Альбертса с соавт. сказано, что хромосомы в митозе напоминают покойника на похоронах – они стимулируют события, но сами не принимают никакого активного участия в действиях.
Митоз наступает при появлении в клетке специального митоз-стимулирующего фактора, который не может возникнуть, пока в клетке не закончилась репликация ДНК и другие подготовительные процессы. Под действием этого фактора запускается каскад фосфорилирования множества белков. В фосфорилированном состоянии они начинают активно функционировать. Один из наиболее интенсивно фосфорилируемых белков (до 6 фосфатных групп на молекулу) – это упомянутый выше гистон Н1. При этом он теряет в сродстве с ДНК (так как его положительный заряд частично компенсируется отрицательно заряженными фосфатными группами), и с ней связываются другие белки, специфичные именно для митоза, что приводит к гораздо более плотной упаковке хромосом, чем в интерфазе. Еще один белок, который фосфорилируется в том же каскаде, запускающем митоз – когезин. В нефосфорилированном состоянии он соединяет вместе две сестринские хроматиды, образовавшиеся в результате репликации ДНК в S-фазе, образуя своего рода кольца вокруг пары хроматид, подобно ободам в бочке. Фосфорилирование когезина в начале мейоза приводит к раскрытию колец и рассоедниению сестринских хроматид, за исключением одного их района - центромеры, о коротом речь пойдет ниже. Имеется механизм, который в этом районе снова фосфорилирует когезин, так что именно здесь сестринские хроматиды остаются соединенными друг с другом.
Первая стадия митоза – профаза. Главное, что происходит в профазе, – дополнительная упаковка (конденсация) хромосом. В такой степени, что они становятся похожими сначала на спутанные нити, видимые в световой микроскоп.
В профазе происходят важные события и в цитоплазме. Имевшиеся в клетке микротрубочки деполимеризуются. При этом клетка как правило теряет свою специфическую форму и округляется. Вокруг центросом образуется так называемая звезда – система из радиально расходящихся микротрубочек, которые постепенно удлиняются. Микротрубочки в процессе митоза начинают обновляться в 20 раз быстрее, чем в интерфазе, и небольшому числу длинных стабильных микротрубочек приходит на смену множество коротких нестабильных. Интенсивная сборка и разборка микротрубочек необходима для правильного течения митоза.
Когда микротрубочки, исходящие от двух полюсов (клеточных центров) навстречу друг другу, вступают в контакт, они соединяются друг с другом определенными белками, которые стабилизируют их и предотвращают от деполимеризации. Такие микротрубочки формируют веретено деления. Микротрубочки звезды, растущие в других направлениях, либо постояно разрушаются, либо фиксируются на клеточном кортексе вблизи полюсов.
Затем наступает прометафаза, которая знаменуется важнейшим событием – ядерная мембрана дефрагментируется на пузырьки и ядро исчезает как структура. При этом происходит деполимеризация ламины ядерного скелета, состоящего из филаментов определенных белков, подстилающих ядерную мембрану (этот процесс также связан с их фосфорилированием). Содержимое ядра объединяется с цитоплазмой. Тем самым восстанавливается состояние, похожее на прокариотическое, при котором ДНК находится в том же компартменте, что и рибосомы. Во время деления ядро исчезает. Это, по-видимому, указывает на то, что ядро – это временная рабочая структура, призванная разобщить тарнскрипцию и трансляцию, хотя бы ценой существенных энергетических затрат на ядерный транспорт и на то, чтобы от него, ядра, избавляться при всяком делении клетки и восстанавливать после него.
Смысл этого эволюционного решения не вполне ясен. Возможно, таким путем достигается защита ДНК от каких-то ферментов или других веществ в цитозоле. Допустим, в ходе клеточного дыхания в митохондриях образуется какое-то количество реакционноспособных свободных радикалов, которые опасны для ДНК. Дополнительная двойная мембрана может ставить для них дополнительный барьер. Цитозоль содержит РНКазы, разрушающие мРНК после того как они становятся ненужными, наверное, нежелательно, чтобы они контактировали с мРНК в процессе их синтеза (но у прокариот это не составляет проблемы).
В прометафазе хромосомы конденсируются окончательно и принимают вид парных образований, напоминающих двойные палочки или червяков, причем каждая пара соединяется центромерой, где образуется своего рода перетяжка – это называется метафазные хромосомы. Такое изображение хромосомы вам всем конечно же хорошо знакомо. На рис. 9.2 она показана с некоторыми подробностями. (Теломера – это конец хромосомы, имеющий специфическую последовательность нуклеотидов. Вторичная перетяжка соответствует ядрышку – это место, где находятся гены рРНК – оно не конденсируется в той же степени, что и остальная хромосома. Спутник – это участок «нормальной» хромосомы за вторичной перетяжкой. Вторичная перетяжка и соответственно спутник есть далеко не на всех хромосомах, поэтому они помогают их идентифицировать.)
Необходимо помнить, что метафазная хромосома – это хромосома в нерабочем состоянии, упакованная для деления. В рабочем состоянии, т. е. в интерфазе, хромосома представляет собой кисель, заваренный вокруг линейной молекулы ДНК, и ее не увидишь под микроскопом.
Метафазная хромосома – двойная. Две ее протяженные составляющие соответствуют двум линейным молекулам ДНК, образовавшимся при репликации. Они называются сестринские хроматиды.
Как мы уже сказали, место соединения хроматид называется центромера. Она удваивается позже остальной ДНК, но в метафазной хромосоме центромера, так же как и вся хромосома, состоит из двух хроматид, только в этом месте соединенных определенными белками. Местоположение центромеры на молекуле ДНК (хромосоме) не вполне определяется специфической первичной структурой. Центромера всегда расположена там, где локализованы определенные последовательности, как многократно повторенные голова к хвосту. Это тандемные повторы. Таких участков на хромосоме много, структуру они имеют разную, и некоторые из них обладают способностью служить центром организации центромеры. Если центромерный участок ДНК удалить, центромера как правило организуется на другом участке подходящей структуры. И в то же время в норме центромера сохраняет свою позицию на ДНК при удвоении последней. Это означает, что вся центромера как сложная структура также наследуется эпигенетически, как некая передаваемая по наследству надстройка к подходящему сайту, причем на хромосоме она всегда одна. После всего вышесказанного не выглядет удивительным, что структура центромерных повторов может быть разной у разных видов и даже у разных хромосом одного вида.
В прометафазе происходит следующее. На центромере каждой из хроматид формируется определенная структура, называемая кинетохор (рис. 9.3). Он состоит, как вы, наверное, догадались, из определенных белков, в его состав также может входить некодирующая РНК. Подчеркнем, что каждая хромосома несет два кинетохора, по одному – на каждую из своих хроматид. Кинетохор связывается с растущими концами микротрубочек, отходящих от полюсов клетки. К каждому кинетохору прикрепляется несколько десятков микротрубочек (но вот у дрожжей – только одна). При этом кинетохоры разных хроматид одной хромосомы связываются с микротрубочками, отходящими от разных полюсов. В прометафазе хромосомы, как правило, активно блуждают по цитоплазме. Поначалу оба кинетохора могут связываться с микротрубочками одного полюса, однако вскоре происходит определенная перестройка контактов кинетохора с микротрубочками, так что центромера одной хроматиды оказывается связанной с микротрубочками, идущими только от одного из полюсов веретена деления.
Любопытно, что связь кинетохора с микротрубочками, по-видимому, стабилизируется натяжением последних. В опытах, когда хромосому, оба кинетохора которой были связаны с микротрубочками одного и того же полюса, механически тянули от него, такая связь сохранялась. Если же усилия не прилагать, то рано или поздно каждый кинетохор окажется связанным с микротрубочками своего полюса, что и создаст стабилизирующее эту связь напряжение.
В прометафазе микротрубочки активно растут, и именно с того конца, который прикреплен к кинетохору. В метафазе этот рост компенсируется деполимеризацией концов микротрубочек у центросомы, так что молекулы тубулина постепенно перемещаются от концов к полюсам, а микротрубочка остается натянутой и сохраняет постоянную длину.
refdb.ru
Опорно-двигательная система клетки » СтудИзба
Лекция № 7.
Количество часов: 2
ОПОРНО-ДВИГАТЕЛЬНАЯ СИСТЕМА КЛЕТКИ
1. Центриоли, структура, репликация, участие в делении клетки
2. Строение ресничек и жгутиков эукариотических клеток
3. Фибриллярные структуры цитоплазмы
Одним из важнейших свойств любой клетки является подвижность. Подвижность клетки проявляется различными формами движения (внутриклеточное, амебовидное, мерцательное, сократительное). Несмотря на разнообразие форм, в основе любых двигательных реакций лежат общие молекулярные механизмы. Кроме того, двигательные аппараты (реснички, жгутики) структурно связаны с существованием опорных внутриклеточных структур. Это позволяет говорить об опорно-двигательной системе клеток.
1. Центриоли, структура, репликация, участие в делении клетки
Клеточный центр, или центросома, представляет собой немембранный органоид, локализованный около ядра и состоящий из двух центриолей и центросферы. Причем постоянным и наиболее важным компонентом клеточного центра являются центриоли. Этот органоид обнаружен в клетках животных, низших растений и грибов.
Центриоли (от лат. centrum – срединная точка, центр) представляют два перпендикулярно расположенных друг к другу цилиндра, стенки которых образованы микротрубочками и соединены системой связок. Конец одного цилиндра (дочерняя центриоль) направлен к поверхности другого (материнская центриоль). Совокупность сближенных между собой материнской и дочерней центриолей называетя диплосомой. Впервые центриоли были обнаружены и описаны в 1875 В. Флемингом. В интерфазных клетках центриоли часто располагаются возле комплекса Гольджи и ядра.
Ультрамикроскопическое строение центриолей было изучено только с помощью электронного микроскопа. Стенку центриолей составляют расположенные по окружности 9 триплетов микротрубочек, образующих полый цилиндр. Системы микротрубочек центриоли можно описать формулой (9X3) + 0, подчеркивая отсутствие микротрубочек в центральной части. Ширина центриоли составляет около 0,2 мкм, длина - 0,3-0,5 мкм (однако, есть центриоли, достигающие в длину нескольких микрометров). Кроме микротрубочек в состав центриоли входят дополнительные структуры - "ручки", соединяющие триплеты.
Центросфера - плотный слой цитоплазмы вокруг центриолей, в котором часто содержатся микротрубочки, расположенные лучами.
Центриолярный цикл. Строение и активность центриолей меняются в зависимости от периода клеточного цикла. Это позволяет говорить о центриолярном цикле. В начале периода G1 от поверхности материнской центриоли начинается рост микротрубочек, которые растут и заполняют цитоплазму. По мере роста микротрубочки теряют связь с областью центриолей и могут находиться в цитоплазме длительное время. В периоде S или G2 происходит удвоение числа центриолей. Этот процесс заключается в том, что центриоли в диплосоме расходятся и около каждой из них происходит закладка процентриолей. В начале вблизи и перпендикулярно исходной центриоли закладываются девять одиночных микротрубочек. Затем они преобразуются в девять дуплетов, а потом в девять триплетов микротрубочек новых центриолей. Этот способ увеличения числа центриолей был назван дупликацией. Следует отметить, что удвоение числа центриолей не связано с их делением, почкованием или фрагментацией, а происходит путем образования процентриолей. Таким образом, в результате дупликации в клетке содержатся четыре попарно связанные центриоли. В этом периоде материнская центриоль продолжает играть роль центра образования цитоплазматических микротрубочек. В периоде G2 обе материнские центриоли покрываются фибриллярным гало (зона тонких фибрилл), от которого в профазе начнут отрастать митотические микротрубочки. В этом периоде в цитоплазме происходит исчезновение микротрубочек и клетка стремиться приобрести шаровидную форму. В профазе митоза диплосомы расходятся к противоположным полюсам клетки. От фибриллярного гало материнской центриоли отходят микротрубочки, из которых формируется веретено деления митотического аппарата. Таким образом, центриоли являются центрами организации роста микротрубочек. В телофазе происходит разрушение веретена деления. Следует отметить, что в клетках высших растений, некоторых водорослей, грибов, ряда простейших центры организации роста микротрубочек центриолей не имеют. У некоторых простейших центрами индукции образования микротрубочек выступают плотные пластинки, связанные с мембраной.
2. Строение ресничек и жгутиков эукариотических клеток
Реснички и жгутики – органоиды специального назначения, выполняющие двигательную функцию и выступающие из клетки. Различий в ультрамикроскопическом строении ресничек и жгутиков нет. Жгутики отличаются от ресничек лишь длиной. Длина ресничек составляет 5-10 мкм, а длина жгутиков может достигать 150 мкм. Диаметр их составляет около 0,2 мкм. Причем клетки, имеющие реснички и жгутики, в свободном состоянии обладают способностью двигаться. Неподвижные клетки, благодаря движению ресничек, способны перемещать жидкости и частички веществ.
Жгутик – это органоид движения у бактерий, ряда простейших, зооспор и сперматозоидов. В клетке обычно бывает от 1 до 4 жгутиков.
Ресничка – это органоид движения или рецепции у клеток животных и некоторых растений. Траектория движения ресничек очень разнообразна. В различных клетках это движение может быть маятникообразным, крючкообразным, воронкообразным или волнообразным.
Ресничка представляет собой тонкий цилиндрический вырост цитоплазмы, покрытый цитоплазматической мембраной. Внутри выроста расположена аксонема ("осевая нить"), состоящая в основном из микротрубочек. В основании реснички находится базальное тело, погруженное в цитоплазму. Диаметры аксонемы и базального тельца одинаковы (около 150 нм). Базальное тельце, как и центриоли, состоит из 9 триплетов микротрубочек и имеет "ручки". Часто в основании реснички лежит не одна, а пара базальных телец, располагающихся под прямым углом друг к другу подобно диплосоме - центриоли. Аксонема в отличие от базального тельца или центриоли имеет 9 дублетов микротрубочек с "ручками", образующих стенку цилиндра аксонемы. Кроме периферических дублетов микротрубочек, в центре аксонемы располагается пара центральных микротрубочек. В целом систему микротрубочек реснички описывают как (9 х 2) + 2 в отличие от (9 х 3) + 0 системы центриолей и базальных телец. Базальное тельце и аксонема структурно связаны друг с другом и составляют единое целое: две микротрубочки триплетов базального тельца являются микротрубочками дублетов аксонемы.
Для объяснения способа движения ресничек и жгутиков используется гипотеза "скользящих нитей". Считается, что незначительные смещения дублетов микротрубочек друг относительно друга могут вызвать изгиб всей реснички. Если такое локальное смещение будет происходить вдоль жгутика, то возникает волнообразное движение.
3. Фибриллярные структуры цитоплазмы
Цитоплазма клетки представляет собой вязкую жидкость, поэтому из-за поверхностного натяжения клетка должна иметь шаровидную форму. Однако помимо шаровидной встречается множество других форм клеток (кубические, призматические, звездчатые, дисковидные, с разнообразными отростками и другие). Форма определяется с помощью жестких, параллельно расположенных волокон. Эти волокна называются фибриллярными структурами цитоплазмы. К ним относятся микротрубочки, микрофиламенты и промежуточные филаменты. Эти структуры образуют цитоскелет клетки (опорно-двигательная система). Цитоскелет определяет форму клетки, участвует в передвижении клетки, во внутриклеточном транспорте органоидов и отдельных соединений.
Микротрубочки - немембранные органоиды, представляющие собой полые цилиндры длиной около 200 нм и толщиной около 25 нм. Микротрубочки можно обнаружить в цитоплазме практически всех эукариотных клеток. В больших количествах они находятся в цитоплазматических отростках нервных клеток, фибробластов и других изменяющих свою форму клеток. Местом организации роста микротрубочек цитоскелета в интерфазной клетке является центриоль. Микротрубочки различного происхождения (реснички простейших, клетки нервной ткани, веретено деления) имеют сходный состав и содержат белки - тубулины. Очищенные тубулины при определенных условиях способны собираться в микротрубочки. Добавление алкалоида колхицина предотвращает самосборку микротрубочек или приводит к разборке уже существующих. В клетке тубулины существуют в двух формах - свободной и связанной. Сдвиг равновесия между этими формами приводит или к диссоциации микротрубочек, или к их росту. Ни тубулины в чистом виде, ни построенные из них микротрубочки не способны к сокращению. В клетках микротрубочки принимают участие в создании ряда временных (цитоскелет интерфазных клеток, веретено деления) или постоянных структур (центриоли, базальные тельца, реснички, жгутики). Кроме того, микротрубочки способствуют ориентированному движению внутриклеточных компонентов.
Микрофиламенты представляют собой тонкие (около 7 нм) белковые нити, встречающиеся практически во всех типах эукариотических клеток. Они расположены пучками или слоями в кортикальном слое цитоплазмы, непосредственно под плазмолеммой. Сеть микрофиламентов выявлена в большинстве клеток. В состав микрофиламентов входят сократительные белки: актин, миозин, тропомиозин, альфа-актинин. Функции микрофиламентов заключаются в образовании цитоскелета и обеспечении большинства внутриклеточных форм движения (токи цитоплазмы, движение вакуолей, митохондрий).
Промежуточные филаменты имеют толщину около 10 нм и также являются белковыми структурами. Это тонкие неветвящиеся, часто располагающиеся пучками нити. Характерно, что их белковый состав различен в разных тканях. В эпителии в состав промежуточных филаментов входит кератин. Пучки кератиновых промежуточных филаментов в эпителиальных клетках образуют тонофибриллы. В состав промежуточных филаментов клеток мезенхимальных тканей (например, фибробластов) входит другой белок - виментин, в мышечные клетки - десмин, в нервных клетках в состав их нейрофиламентов также входит особый белок. Промежуточные филаменты выполняют опорно-каркасную функцию.
studizba.com
| Мембраны – динамические, подвижные структуры, которые постоянно изменяют свою форму и площадь. На подвижности мембран основана концепция эндоплазматической системы. Согласно этой концепции, внутренние мембраны цитоплазмы, кроме мембран митохондрий и пластид, представляют собой единое целое и берут начало от эндоплазматического ретикулума. Новые цистерны диктиосом образуются из эндоплазматического ретикулума через стадию промежуточных пузырьков, а секреторные пузырьки, отделяющиеся от диктиосом, в конечном итоге способствуют формированию плазматической мембраны. Таким образом, эндоплазматический ретикулум и диктиосомы образуют функциональное целое, в котором диктиосомы играют роль промежуточных структур в процессе преобразования мембран, подобных эндоплазматическому ретикулуму, в мембраны, подобные плазматической. В тканях, клетки которых слабо растут и делятся, постоянно происходит обновление мембранных компонентов. Микротрубочки обнаружены практически во всех эукариотических клетках. Представляют собой цилиндрические структуры диаметром около 24 нм. Длина их варьирует. Каждая трубочка состоит из субъединиц белка, называемого тубулином. Субъединицы образуют 13 продольных нитей, окружающих центральную полость. Микротрубочки – это динамические структуры, они регулярно разрушаются и образуются на определенных стадиях клеточного цикла. Их сборка происходит в особых местах, которые называются центрами организации микротрубочек. В растительных клетках они имеют слабовыраженную аморфную структуру. Функции микротрубочек: участвуют в образовании клеточной оболочки; направляют пузырьки диктиосом к формирующейся оболочке, подобно нитям веретена, которые образуются в делящейся клетке; играют определенную роль в формировании клеточной пластинки (первоначальной границы между дочерними клетками). Кроме того, микротрубочки – важный компонент жгутиков и ресничек, в движении которых, играют немаловажную роль. Микрофиламенты, подобно микротрубочкам, найдены практически во всех эукариотических клетках. Представляют собой длинные нити толщиной 5 – 7 нм, состоящие из сократительного белка актина. Пучки микрофиламентов встречаются во многих клетках высших растений. По-видимому, играют важную роль в токах цитоплазмы. Микрофиламенты вместе с микротрубочками образуют гибкую сеть, называемую цитоскелетом. Основное вещество довольно долго считали гомогенным (однородный) богатым белком раствором с малым количеством структур или вообще бесструктурным. Однако в настоящее время, используя высоковольтный электронный микроскоп, было установлено, что основное вещество представляет трехмерную решетку, построенную из тонких (диаметром 3 – 6 нм) тяжей, заполняющих всю клетку. Другие компоненты цитоплазмы, включая микротрубочки и микрофиламенты, подвешены к этой микротрабекулярной решетке. Микротрабекулярная структура представляет собой решетку из белковых тяжей, пространство между которыми заполнено водой. Вместе с водой решетка имеет консистенцию геля, гель имеет вид студенистых тел. К микротрабекулярной решетке прикреплены органеллы. Решетка осуществляет связь между отдельными частями клетки и направляет внутриклеточный транспорт. Липидные капли – структуры сферической формы, придающие гранулярность цитоплазме растительной клетки под световым микроскопом. На электронных микрофотографиях они выглядят аморфными. Очень похожие, но более мелкие капли встречаются в пластидах. Липидные капли, принимая за органеллы, называли их сферосомами и считали, что они окружены одно- или двуслойной мембраной. Однако последние данные показывают, что у липидных капель мембран нет, но они могут быть покрыты белком. Эргастические вещества – это «пассивные продукты» протопласта: запасные вещества или отходы. Они могут появляться и исчезать в разные периоды клеточного цикла. Кроме зерен крахмала, кристаллов, антоциановых пигментов и липидных капель. К ним относятся смолы, камеди, танины и белковые вещества. Эргастические вещества входят в состав клеточной оболочки, основного вещества цитоплазмы и органелл, в том числе вакуолей. Жгутики и реснички – это тонкие, похожие на волоски структуры, которые отходят от поверхности многих эукариотических клеток. Имеют постоянный диаметр, но длина колеблется от 2 до 150 мкм. Условно более длинные и немногочисленные из них называют жгутиками, а более короткие и многочисленные - ресничками. Четких различий между этими двумя типами структур не существует, поэтому для обозначения обоих используют термин жгутик. У некоторых водорослей и грибов жгутики являются локомоторными органами, с помощью которых они передвигаются в воде. У растений (например, мхов, печеночников, папоротников, некоторых голосеменных) только половые клетки (гаметы) имеют жгутики. Каждый жгутик имеет определенную организацию. Наружное кольцо из 9 пар микротрубочек окружает две дополнительные микротрубочки, расположенные в центре жгутика. Содержащие ферменты «ручки» отходят от одной микротрубочки каждой из наружных пар. Это основная схема организации 9 + 2 обнаружена во всех жгутиках эукариотических организмов. Считают, что движение жгутиков основано на скольжении микротрубочек, при этом наружные пары микротрубочек движутся одна вдоль другой без сокращения. Скольжение пар микротрубочек относительно друг друга вызывает локальное изгибание жгутика. Жгутики «вырастают» из цитоплазматических цилиндрических структур, называемых базальными тельцами, образующимися и базальную часть жгутика. Базальные тельца имеют внутреннее строение, напоминающее строение жгутика, за исключением того, что наружные трубочки собраны в тройки, а не в пары, а центральные трубочки отсутствуют. Клеточная стенка. Клеточная стенка отграничивает размер протопласта и предохраняет его разрыв за счет поглощения воды вакуолью. Клеточная стенка имеет специфические функции, которые важны не только для клетки и ткани, в которой клетка находится, но и для всего растения. Клеточные стенки играют существенную роль в поглощении, транспорте и выделении веществ, а, кроме того, в них может быть сосредоточена лизосомальная, или переваривающая активность. Компоненты клеточной стенки. Наиболее типичным компонентом клеточной стенки является целлюлоза, которая в значительной степени определяет её архитектуру. молекулы целлюлозы состоят из повторяющихся молекул глюкозы, соединенных конец к концу. Длинные тонкие молекулы целлюлозы объединены в микрофибриллы толщиной 10 – 25 нм. Микрофибриллы перевиваются и образуют тонкие нити, которые в свою очередь могут обматываться одна вокруг другой, как пряди в канате. Каждый такой «канат», или макрофибрилла, имеет толщину около 0,5 мкм, достигая в длину 4 мкм. Макрофибриллы прочны, как равная по величине стальная проволока. Целлюлозный каркас клеточной стенки заполнен переплетающимися с ним целлюлозными молекулами матрикса. В его состав входят полисахариды, называемые гемицеллюлозами, и пектиновые вещества, или пектины, химически очень близкие к гемицеллюлозам. Другой компонент клеточной стенки – лигнин – является самым распространенным после целлюлозы полимером растительных клеток. Лигнин увеличивает жесткость стенки и обычно содержится в клетках, выполняющих опорную или механическую, функцию. Кутин, суберин, воска – обычно откладываются в оболочках защитных тканей растений. Кутин, например, содержится в клеточных оболочках эпидермы, а суберин - вторичной защитной ткани, пробки. Оба вещества встречаются в комбинации с восками и предотвращают чрезмерную потерю воды растением. Слои клеточной стенки. Толщина стенки растительных клеток варьирует в широких пределах в зависимости от роли клеток в структуре растений и возраста самой клетки. Под электронным микроскопом просматривается в растительной клеточной стенке два слоя: срединная пластинка (называемая также межклеточным веществом), и первичной клеточной стенки. Многие клетки откладывают ещё один слой – вторичную клеточную стенку. Срединная пластинка располагается между первичными стенками соседних клеток. Вторичная стенка, если она есть, откладывается протопластом клетки на внутреннюю поверхность первичной клеточной стенки. Срединная пластинка. Срединная пластинка состоит в основном из пектиновых веществ. Там, где должна возникнуть клеточная стенка, между двумя вновь образующимися клетками, вначале отмечается густое сплетение из канальцев эндоплазматической сети и цистерны аппарата Гольджи (диктиосом). Затем в этом месте появляются пузырьки, заполнены пектиновым веществом (из полисахаридов). Пузырьки эти отделяются от цистерн аппарата Гольджи. Ранняя клеточная стенка содержит различные полисахариды, основные из которых пектины и гемицеллюлоза. Позже в её состав входят более плотные вещества – целлюлоза и лигнин. Первичная клеточная оболочка. Это слой целлюлозной оболочки, который откладывается до начала или во время роста клетки. Помимо целлюлозы, гемицеллюлоз и пектина первичные оболочки содержат гликопротеин. Первичные оболочки могут лигнифицироваться. Пектиновый компонент придаёт пластичность, которая позволяет первичной оболочке, растягивается по мере удлинения корня, стебля или листа. Активно делящиеся клетки (большинство зрелых клеток, вовлеченных в процессы фотосинтеза, дыхания и секреции) имеют первичные оболочки. Такие клетки с первичной оболочкой и живым протопластом способны утрачивать характерную форму, делиться и дифференцироваться в новый тип клеток. Именно они участвуют в заживлении ран и регенерации тканей у растений. Первичные клеточные оболочки не одинаковы по толщине на всем своем протяжении, а имеют тонкие участки, которые называются первичными поровыми полями. Тяжи цитоплазмы, или плазмодесмы, соединяющие протопласты соседних клеток, обычно проходят через первичные поровые поля. Вторичная клеточная оболочка. Несмотря на то, что многие растительные клетки имеют только первичную оболочку, у некоторых к центру клетки протопласт откладывает вторичную оболочку. Обычно это происходит после прекращения роста клетки и площадь первичной оболочки более не увеличивается. По этой причине вторичная оболочка отличается от первичной. Вторичные оболочки особенно нужны специализированным клеткам, укрепляющим растение и проводящим воду. После отложения вторичной оболочки протопласт этих клеток, как правило, отмирает. Во вторичных оболочках больше целлюлозы, чем в первичных, а пектиновые вещества и гликопротеины в них отсутствуют. Вторичная оболочка растягивается с трудом, ее матрикс состоит из гемицеллюлозы. Во вторичной оболочке можно выделить три слоя – наружный, средний и внутренний (S1, S2, S3). Слоистая структура вторичных оболочек значительно увеличивает их прочность. Микрофибриллы целлюлозы во вторичной оболочке откладывается плотнее, чем в первичной. Лигнин – обычный компонент вторичных оболочек древесины. Поры в оболочках контактирующих клеток расположены напротив друг друга. Две лежащие друг против друга поры и поровая мембрана образуют пару пор. В клетках, имеющих вторичные оболочки, существуют два основных типа пор: простые и окаймленные. В окаймленных порах вторичная оболочка нависает над полостью поры. В простых порах этого нет. Рост клеточной оболочки. По мере роста клетки увеличивается толщина и площадь клеточной оболочки. Растяжение оболочки – процесс сложный. Он контролируется протопластом и регулируется гормоном ауксином. В клетках, растущих во всех направлениях равномерно, отложение миофибрилл носит случайный характер. Эти миофибриллы образуют неправильную сеть. Такие клетки обнаружены в сердцевине стебля, запасающих тканях и при культивировании клеток in vitro. В удлиняющихся клетках миофибриллы боковых оболочек откладывается под прямым углом к оси удлинения. Вещества матрикса – пектины, гемицеллюлозы и гликопротеины переносятся к оболочке в пузырьках диктиосом. При этом пектины более характерны для растущих клеток, а гемицеллюлозы преобладают в не растущих клетках. Целлюлозные микрофибриллы синтезируются на поверхности клетки с помощью ферментного комплекса, связанного с плазматической мембраной. Ориентация микрофибрилл контролируется микротрубочками, расположенными у внутренней поверхности плазматической мембраны. Плазмодесмы. Это тонкие нити цитоплазмы, которые связывают между собой протопласты соседних клеток. Плазмодесмы либо проходят сквозь клеточную оболочку в любом месте, либо сосредоточены на первичных поровых полях или в мембранах между парами пор. Под электронным микроскопом плазмодесмы выглядят как узкие каналы, выстланные плазматической мембранной. По оси канала из одной клетки в другую тянется цилиндрическая трубочка меньшего размера – десмотрубочка, которая сообщается с эндоплазматическим ретикулумом обеих смежных клеток. Многие плазмодесмы формируются во время клеточного деления, когда трубчатый эндоплазматический ретикулум захватывается развивающейся клеточной пластинкой. Плазмодесмы могут образовываться и в оболочках неделящихся клеток. Эти структуры обеспечивают эффективный перенос некоторых веществ от клетки к клетке. Деление клеток. У многоклеточных организмов деление клеток наряду с увеличением их размеров является способом роста всего организма. Новые клетки, образовавшиеся во время деления, сходны по структуре и функциям, как с родительской клеткой, так и между собой. Процесс деления у эукариот можно подразделить на две частично перекрывающиеся стадии: митоз и цитокинез. Митоз – это образование из одного ядра двух дочерних ядер, морфологически и генетически эквивалентных друг другу. Цитокинез – это деление цитоплазматической части клетки с образованием дочерних клеток. Клеточный цикл. Живая клетка проходи ряд последовательных событий, составляющих клеточный цикл. Продолжительность самого цикла варьирует в зависимости от типа клетки и внешних факторов, например от температуры или обеспеченности питательными веществами. Обычно цикл делится на интерфазу и четыре фазы митоза. Интерфаза. Период между последовательными митотическими делениями. Интерфазу делят на три периода, обозначаемые как G1, S, G2. В период G1, который начинается после митоза. В этот период увеличивается количество цитоплазмы, включая различные органеллы. Кроме того, согласно современной гипотезе, в период G1 синтезируются вещества, которые либо стимулируют, либо ингибируют период S и остальную часть цикла, определяя, таким образом, процесс деления. В период S следует за периодом G1, в это время происходит удвоение генетического материала (ДНК). В период G2, который следует за S, формируются структуры, непосредственно участвующие в митозе, например, компоненты веретена. Некоторые клетки проходит неограниченный ряд клеточных циклов. Это одноклеточные организмы и некоторые клетки зон активного роста (меристем). Некоторые специализированные клетки после созревания теряет способность к размножению. Третья группа клеток, например образующих раневую ткань (каллус), сохраняет способность делиться только в специальных условиях. Митоз, или деление ядра. Это непрерывный процесс, подразделяемый на четыре фазы: профазу, метафазу, анафазу, телофазу. В результате митоза генетический материал, удвоившийся в интерфазе, делится поровну между двумя дочерними ядрами. Одним из самых ранних признаков перехода клетки к делению служит появление узкого, кольцеобразного пояска из микротрубочек непосредственно под плазматической мембраной. Это относительно плотный поясок окружает ядро в экваториальной плоскости будущего митотического веретена. Так как он проявляется перед профазой, его называют препрофазным пояском. Он исчезает после митотического веретена, задолго до появления в поздней телофазе клеточной пластинки, которая растет от центра к периферии и сливается с оболочкой материнской клетки в области, ранее занятой препрофазным пояском. Профаза. В начале профазы хромосомы напоминают длинные нити, разбросанные внутри ядра. Затем, по мере того как нити укорачиваются и утолщаются, можно увидеть, что каждая хромосома состоит не из одной, а из двух переплетенных нитей, называемых хроматидами. В поздней профазе две укороченные спаренные хроматиды каждой хромосомы лежат рядом параллельно, соединённые узким участком, называемым центромерой. Она имеет определённое положение на каждой хромосоме и делит хромосому на два плеча различной длины. Микротрубочки располагаются параллельно поверхности ядра вдоль оси веретена. Это само раннее проявление сборки митотического веретена. К концу профазы ядрышко постепенно теряет чёткие очертания и наконец исчезает. Вскоре после этого распадается и ядерная оболочка. продолжение |
coolreferat.com
Особенности клетки водорослей
Клетки многих примитивных, со жгутиками водорослей, а также зооспоры и гаметы большинства водорослей покрыты только цитоплазматической мембраной, или плазмалеммой. Протопласты большинства водорослей одеты клеточной оболочкой. Они состоят из аморфного матрикса, образованного гемицеллюлозами и пектиновыми веществами, и погруженных в него обычно целлюлозных микрофибрилл. В оболочке или на ее поверхности откладываются добавочные компоненты (карбонат кальция, кремний, железо, альгиновая кислота, хитин и др.). Клеточная оболочка может состоять из двух и более частей, может иметь различные выросты. Существуют и другие типы клеточных покровов (например, перипласт, тека), которые рассматриваются в соответствующих отделах водорослей.
Цитоплазма у большинства взрослых клеток образует тонкий постенный слой. Вакуоль с клеточным соком занимает обычно центр клетки. Такая вакуоль отсутствует в клетках синезеленых водорослей и в монадных клетках. Пресноводные монадные формы образуют пульсирующие, или сократительные, вакуоли, синезеленые водоросли – газовые вакуоли.
Ядерный аппарат у большинства водорослей представлен в виде типичных ядер (эукариотический тип клеточной организации). У динофитовых ядро примитивное (мезокариотический тип клеточной организации). Синезеленые водоросли и прохлорофиты не имеют оформленного ядра. Ту часть клетки, в которой содержится ДНК, называют нуклеоидом или нуклеоплазматической областью (прокариотический тип клеточной организации).
Фотосинтетический аппарат у водорослей представлен разными уровнями организации. У синезеленых и прохлорофитовых он морфологически не оформлен. Другие имеют хроматофоры, которые чрезвычайно разнообразны по форме, строению и расположению в протопласте клетки. В большинстве случаев хроматофоры занимают постенное положение (периетальные хроматофоры) и могут быть пластинчатыми, кольцевидными, спиральными, чашевидными, цилиндрическими, в виде зерен или дисков. Реже хроматофор занимает центральное положение в клетке. В этом случае он имеет массивную центральную часть, от которой отходят лопасти, или гребни. Непрерывная оболочка хроматофора окружает тонкозернистый материал матрикса или стромы, в которой расположены уплощенные, одетые мембраной тилакоиды. В тилакоидах сосредоточены фотосинтезирующие пигменты: хлорофиллы, а также каротины и ксантофиллы. Кроме того, в матриксе хроматофора находятся рибосомы, липидные гранулы, фибриллы ДНК и особые образования – пиреноиды. Пиреноид представляет собой структуру белковой природы, в основном состоит из фермента рибулозодифосфаткарбоксилазы. У зеленых водорослей вокруг пиреноида в матриксе хроматофора откладываются зерна крахмала.
Кроме того, крахмал располагается и в матриксе. У других водорослей запасные полисахариды откладываются вне связи с пиреноидами.
Хроматофоры водорослей в деталях тонкого строения, касающихся оболочки, расположения тилакоидов и фибрилл ДНК, формы пиреноидов, места образования и отложения зерен запасных полисахаридов, обнаруживают весьма четкие различия. Это позволяет использовать их наряду с набором пигментов, продуктами запаса и строением жгутикового аппарата в качестве таксономических признаков больших групп – отделов водорослей. Так, у зеленых, харовых и красных водорослей оболочка хроматофора состоит только из двух параллельных мембран, у динофитовых и эвгленовых – из трех, у золотистых, желтозеленых, диатомовых, криптофитовых, рафидофитовых и бурых – из четырех. Последние имеют хлоропластную эндоплазматическую сеть, которая формируется в результате связи хлоропластов с мембранами эндоплазматического ретикулюма.
Расположение тилакоидов в матриксе хроматофоров также неодинаковое в разных отделах водорослей. Наиболее простое расположение у красных водорослей, у которых тилакоиды рассредоточены в матриксе по одному. У остальных эукариотических водорослей тилакоиды сгруппированы в ламеллы. Причем, количество тилакоидов в одной ламелле в пределах больших групп, объединяющих родственные водоросли, постоянно. У криптофитовых тилакоиды соединяются по два; у золотистых, желтозеленых, диатомовых бурых, динофитовых и эвгленофитовых – преимущественно по три. У зеленых, харовых, эвгленофитовых число тилакоидов может достигать 20. В таких случаях тилакоиды в стопках настолько тесно прижаты друг к другу, что пространство между соседними тилакоидами исчезает, тогда эти стопки называют гранами. В хроматофорах с трехтилакоидными ламеллами находятся периферические, или опоясывающие, ламеллы, которые расположены параллельно оболочке хроматофора и окружают остальные ламеллы, пересекающие строму хроматофора. Среди красных водорослей у одних опоясывающие тилакоиды есть, у других их нет. У зеленых, харовых, эвгленовых опоясывающие ламеллы не выявлены.
В матриксе хроматофора между ламеллами и вокруг пиреноида у зеленых водорослей (у харовых – только в матриксе) откладывается крахмал, а у всех остальных водорослей запасные вещества: хризоламинарин, ламинарин, парамилон, багрянковый крахмал и другие – в цитоплазме.
Подвижные формы водорослей имеют фоторецепторную органеллу – стигму, или глазок. Структурную основу стигмы составляют плотно прилегающие друг к другу пигментнонесущие глобулы, ограниченные мембранами. Стигма обычно расположена в хроматофоре. Особенно сложно устроенная стигма имеется у динофитовых.
Монадные клетки снабжены жгутиками. Жгутики имеют единый план строения и состоят из свободной части (ундулиподии), переходной зоны, базальных тел (кинетосом) и корней. Наружный отрезок (ундулиподия) покрыт мембраной, являющейся продолжением цитоплазматической мембраны. Под мембраной жгутика находится белковый матрикс, в котором расположены микротрубочки, или фибриллы. Вся система фибрилл в жгутике называется аксонемой. В главном стержне на поперечном сечении расположение микротрубочек следующее: две одинарные центральные микротрубочки окружены кольцом из 9 пар (дуплетов) микротрубочек. В апикальной части жгутика центральная пара микротрубочек может быть длиннее периферических, тогда на кончике жгутика имеется тонкий вырост – акронема.
В жгутиках эвгленовых, динофитовых, некоторых золотистых, репродуктивных клеток некоторых зеленых водорослей между поверхностной мембраной жгутика и его аксонемой расположен параксиальный тяж. Он представляет собой электронно-плотное образование, состоящее обычно из спирально закрученных относительно друг друга пучков микрофиламентов, прикрепленных на всем протяжении к одному или двум периферическим дуплетам аксонемы. Параксиальный тяж значительно увеличивает толщину жгутика.
Переходная зона представляет собой часть жгутика от верхнего конца базального тела до места основания центральных микротрубочек аксонемы, а также структуры, расположенные вокруг этого основания. Структурами переходной зоны жгутиков являются поперечная пластинка, звездчатая структура, переходная спираль, переходный цилиндр и др. Элементы переходной зоны играют роль в укреплении жгутика в месте его выхода из клетки.
Базальное тело представляет собой короткий цилиндр, по периферии которого располагается девять триплетов (у диатомовых – дуплетов) микротрубочек. Базальные тела жгутиков идентичны по своей структуре центриолям и во многих случаях могут превращаться друг в друга. Одной (и единственной) функцией центриолей является образование жгутиков. В начале митоза жгутики исчезают, а базальные тела перемещаются вглубь клетки, ближе к ядру, где служат центрами организации митотического веретена деления. По окончании митоза центриоли снова становятся базальными телами, от которых отрастают жгутики.
Корни жгутиков (корешковая система) прикреплены к базальным телам и закрепляют эти тела в протопласте. Корни жгутиков представлены микротрубочковыми и фибриллярными структурами. Микротрубочковые корни образованы одиночными микротрубочками или пучками и лентами микротрубочек. Фибриллярные корни могут быть простыми и поперечно-исчерченными. Поперечно-исчерченные корни могут состоять из структурного белка ассамблина или из белка центрина. В первом случае они ассоциированы с микротрубочками и не сократимы, во втором – не связаны с микротрубочками и могут медленно сокращаться.
Микротрубочковые корни жгутиков у большинства видов водорослей располагаются симметрично, образуя характерные крестообразные фигуры. У харовых и некоторых зеленых водорослей корешковая система жгутиков имеет асимметричное строение, обусловленное наличием многослойной структуры. Многослойная структура расположена в цитоплазме клетки вблизи базальных тел, ассоциирована со жгутиковым аппаратом и имеет сложное строение. Ее верхний слой состоит из 22-25 тесно расположенных микротрубочек; под ним находятся два слоя филаментов; в основании лежит электронно-плотная пластинка. Многослойная структура водорослей напоминает таковую в сперматозоидах некоторых высших растений.
У некоторых водорослей (желтозеленые, золотистые, рафидофитовые) имеется ризопласт – сложный ветвящийся корешок, соединяющий кинетосомы жгутиков с ядром. Для криптофитовых водорослей отмечается ризостиль – многослойная структура в виде ленты из 6-8 микротрубочек, идущая к заднему концу клетки.
Число жгутиков может варьировать от одного до многих (табл. 1) Большинство же монадных клеток имеет по два жгутика. Различают жгутики терминальные – отходящие от конца клетки, и латеральные – расположенные сбоку клетки. Двужгутиковые клетки могут нести изоконтные – одинаковой длины жгутики и гетероконтные – разной длины; изоморфные – одинакового строения и гетероморфные – разной формы. Гетероморфные формы имеют обычно один из жгутиков с волосовидными образованиями – мастигонемами. На поверхности жгутиков могут быть различные чешуйки, шипы.
Для многих водорослей с пальмеллоидным талломом отмечены псевдоцилии, представляющие собой неподвижные жгутиковидные выросты. Тонкое строение псевдоцилий напоминает жгутики, но свободная часть их не имеет центральных микротрубочек. Нередко псевдоцилии достигают значительной длины и могут обильно ветвиться.
У некоторых золотистых водорослей, помимо жгутиков, имеется гаптонема – направленный вперед вырост, служащий для временного прикрепления клетки к субстрату и парения в воде. По длине гаптонема может быть короче, равна или длиннее жгутиков. Гаптонема может находиться в скрученном или выпрямленном состоянии. Внутри нее находится лента из 6-8 микротрубочек, которая окружена каналом гладкой эндоплазматической сети. Центральные микротрубочки у гаптонемы отсутствуют. У основания гаптонемы число микротрубочек может увеличиваться. Микротрубочки гаптонемы отходят от одного из базальных тел жгутикового аппарата. У гаптонемы найдены фибриллярные корни.
Таблица 1. Особенности строения жгутиков у водорослей из разных отделов
|
Отделы водорослей |
Число жгутиков |
Длина |
Форма |
Поверхность жгутика |
Особенности тонкого строения |
||||
| изоко-нтные | гетеро-контные | изо-морфные | гетеро-морфные | с мастиго-немами, волосками |
с чешуй-ками | гладкая | |||
| Эвгленовые |
1 – 7 |
+ |
+ |
+ |
Параксиальный тяжо | ||||
| Динофитовые |
2 |
+ |
+ |
+ |
+ |
Одна или две поперечные пластинки (п. з.), параксиальный тяж |
|||
| Криптофитовые |
2 |
+ |
+ |
+ |
+ |
Система двойных септ (пластинчатых структур) (п. з.), ризостиль |
|||
| Рафидофитовые |
2 (1) |
+ |
+ |
+ |
+ |
Одна поперечная пластинка (п. з.), ризопласт |
|||
| Золотистые |
1-2 (4) (у некоторых гаптонема) |
+ |
+ |
+ |
+ |
+ |
+ |
+ |
Поперечная пластинка, переходная спираль, переходный цилиндр (п. з.), ризопласт, у некоторых – параксиальный тяж |
| Желтозеленые |
2 (1) |
+ |
+ |
+ |
+ |
Переходная спираль (п. з.), ризопласт | |||
| Диатомовые |
1 |
+ |
(9–9) + 0 (с. ж.), базальные тела из дуплетов микротрубочек, корешковая система не выражена |
||||||
| Бурые |
2 (1) |
+ |
+ |
+ |
+ |
+ |
Поперечная пластинка (п. з.) | ||
| Зеленые |
2 – много |
+ |
+ |
+ |
+ |
+ |
Звездчатая структура, редко совместно с переходной спиралью (п. з.), редко ризопласт | ||
| Харовые |
2 |
+ |
+ |
+ |
Звездчатая структура (п. з.) | ||||
Примечание: с. ж. – стержень жгутика; п. з. – переходная зона.
Деление клетки водорослей имеет свои особенности. У прокариот отсутствуют митоз и мейоз, а имеет место бинарное деление с удвоением ДНК. При этом новая поперечная перегородка возникает в виде кольцевидного валика, который постепенно растет от периферии к центру клетки, где смыкается. Формирование новых поперечных перегородок может начинаться еще до окончания предыдущего деления клетки (ускоренное деление). У эукариотических водорослей имеют место митоз (непрямое деление) и мейоз. Центриоли встречаются далеко не у всех водорослей (хлорококковые, рафидофитовые, желтозеленые, бурые). У некоторых красных водорослей есть полярные кольца – короткие полые цилиндры, около которых иногда могут собираться микротрубочки.
Закрытый митоз – митоз, при котором ядерная оболочка остается непрерывной (желтозеленые, динофитовые, эвгленовые, эдогониальные и др.). Полузакрытый митоз – в ядерной оболочке образуются полярные отверстия либо оболочка фрагментируется, но не исчезает (бурые, красные, вольвоксовые, хлорококковые, ульвовые). Открытый митоз – ядерная оболочка дезинтегрируется, как у высших растений (конъюгаты, харовые, диатомовые, золотистые, криптофитовые и др.).
В большинстве случаев за митозом следует цитокинез – деление цитоплазмы путем образования борозды деления (центростремительно) или клеточной пластинки в экваториальной плоскости клетки между дочерними клетками (центробежно). И тот, и другой тип цитокинеза может сопровождаться образованием фикопласта или фрагмопласта.
Фикопласт – микротрубочки, расположенные перпендикулярно оси веретена, но параллельно плоскости деления клетки и образующейся перегородке. Считается, что микротрубочки фикопласта помогают разделить дочерние ядра, которые расположены очень близко в области поперечной перегородки. Фрагмопласт представляет собой микротрубочки, ориентированные параллельно оси веретена деления и перпендикулярно к плоскости деления клетки. Фрагмопласт считается более эволюционно продвинутым в сравнении с фикопластом. Компоненты новой клеточной стенки находятся в везикулах, которые образуются в диктиосомах.
biofile.ru
Строение и регенерация жгутиков
Строение жгутиков
Монадные клетки снабжены жгутиками. Жгутики всех водорослей имеют единый план строения. Жгутик можно подразделить на наружный и внутренний – эстра- и интрацеллюлярный отрезки. Наружный отрезок, или свободная часть жгута, в свою очередь, состоит из трех частей: кончика, главного стержня и переходной зоны. Внутренний - интрацеллюлярный – отрезок охватывает основание жгута и корни жгута.
Свободная часть жгута всегда одета жгутиковой мембраной, которая не что иное, как приподнятая цитоплазматическая мембрана. Так, у бурой водоросли диктиоты единственный жгутик несколько раз свернут по спирали вокруг молодого сперматозоида. На срезе через такой сперматозоид под мембраной, одевающей клетку, можно видеть микротрубочки жгута, характерным образом расположенные, без какой-либо собственной мембраны. После освобождения сперматозоида первоначально закрученный по спирали жгутик разматывается, приподнимая при этом цитоплазматическую мембрану клетки, которая на этой стадии должна быть в очень лабильном состоянии. Наоборот, при втягивании жгутиков, например при остановке синзооспор вошерии, их мембрана становится частью цитоплазматической мембраны. Кнаружи от жгутиковой мембраны находится чехол – слой материала, покрывающего жгут на большем протяжении его длины.
Под мембраной жгута находится матрикс, образованный белками. Весь жгутиковый аппарат пронизан пучком белковых фибрилл или микротрубочек – аксонемой, поперечный разрез которой выглядит неодинаково на разных уровнях жгута: в его кончике, главном стержне, переходной зоне и базальном теле.
На поперечном срезе главного стержня видно типичное для всех водорослей расположение микротрубочек или фибрилл: дольцо из девяти пар микротрубочек окружает две одинарные центральные микротрубочки – структура +2. Каждый дублет образован «А» и «В» микротрубочками, «А» трубочка каждого дублета снабжена двумя короткими выступами–боковыми «руками», обращенными к следующему дублету, и радиальным тяжем, соединяющим периферический дублет с центральными микротрубочками, которые могут быть заключены в общий чехол. Кроме того, имеются интердублетные соединения – тяжи, соединяющие дублеты, и ряд дополнительных образований.
А последовательность растворения микротрубочек жгутика: удаление мембраны жгутика оставляет нетронутой аксонему. содержащую девять наружных дублетов и две центральные трубочки. При обработке аксонемы возрастающими концентрациями саркозила удаляется одна из центральных трубочек, затем растворяется оставшаяся центральная трубочка, часто вместе с наружной частью трубочки В. Б- интактные наружные дублеты растворяются в такой последовательности: наружная часть трубочки В, оставшаяся часть трубочки В, наружная часть трубочки А, А –остаток трубочки А, за исключением грех первичных нитей, составляющих перегородку между трубочками А и В. В схема, иллюстрирующая гипотетическое расположение тубулина 1 и тубулина 2 в наружном дублете.
Рис. 1. Схема строения дублетов в жгутике Chlamydopionas reinhurdtir
Тонкая структура микротрубочек, составляющих наружные дублеты жгутиков и ресничек, стала объектом многочисленных исследований. Подробно были изучены и дублеты жгутиков простейшей монадной зеленой водоросли хламидомонаса. Дублеты образованы субъединицами первичными нитями, в свою очередь состоящими из одного ряда сферических субъединин. Каждый дублет содержит 23 первичные нити: микротрубочка «А» образована 13 первичными нитями, из которых 3 расположены медианно. создавая перегородку между просветами «А» и «В» микротрубочек. «В» микротрубочка включает 10 первичных нитей.
Г. Витман с соавт. разработали методы изоляции жгутиков хламидомонаса и их фракционирования на мембрану, мастигонемы, матрикс и компоненты аксонемы. Наружные дублеты затем были разделены на «А» и «В» микротрубочки и выделена перегородка между ними, образованная тремя первичными нитями. Каждый и) компонентов был изучен электронно-микроскокически и биохимически. Изолированные мастигонемы образованы эллипсоидными субъединицами, соединенными конец к концу, и содержат один гликопротеин. Фракция матрикса включает ряд белков. Изолированные аксонемы удерживают сначала интактную структуру, а дальнейшее их фракционирование осуществляется путем нагревания или под действием детергента. Возрастающие концентрации детергента саркозила растворяют микротрубочки аксонемы в следующем порядке: одна из двух центральных микротрубочек, вторая центральная микротрубочка и наружная стенка «В» микротрубочки дублета, остальные части «В» микротрубочки, наружная стенка «А» трубочки, остаток «А» трубочки, за исключением трех первичных нитей, составляющих «перегородку» между просветами «А» и «В» микротрубочек.
Анализ наружных дублетов показал, что они образованы двумя белками-тубулином 1 и тубулином 2 соответственно с молекулярной массой 56000 и 53000 и разным аминокислотным составом. Как в микротрубочках «А», так и в микротрубочках «В» присутствуют оба тубулина, в трех же первичных нитях, образующих стенку между просветами «А» и «В» микротрубочек, обнаружен только тубулин 1. В остальных частях микротрубочек каждая первичная нить содержит только один из тубулинов и они чередуются парами: пары первичных нитей, образованных тубулином 1, перемежаются с парами нитей, содержащими тубулин 2.
Кончик – это участок жгута, тупой или слегка заостренный, у которого нарушается описанное для главного стержня жгута нормальное +2 расположение микротрубочек. По мере приближения к кончику утрачивается материал матрикса и периферические дублеты один за другим постепенно теряют одну из микротрубочек, становясь одинарными, но сохраняя при этом изначальное круговое расположение. Затем постепенно сокращается число периферических микротрубочек, пока не останутся только центральные микротрубочки, которые сохраняются почти до самого кончика жгута.
У гамет бурой водоросли эктокарпуса оба жгутика несут длинный, часто закрученный спирально терминальный придаток – акронему, в который из стержня жгута переходят две центральные микротрубочки.
Переходная зона – между свободной частью жгута и базальным телом – внешне легко отличается от стержня, так как здесь жгутик перетянут: жгутиковая мембрана плотно подходит к дублетам аксонемы. У проксимального – морфологически нижнего конца наружной части жгута находится базальный диск, у которого оканчиваются две центральные микротрубочки, Периферические дублеты продолжаются ниже базального диска, где к ним вскоре добавляются дополнительные микротрубочки, превращающие дублеты в триплеты базального тела.
Базальное тело, находящееся в основании каждого жгутика, расположено в цитоплазме вблизи поверхности клетки и представляет собой короткий цилиндр, по периферии которого располагается девять триплетов микротрубочек. Триплеты соединяются друг с другом тонкими нитями между «А» и «С» – микротрубочками соседних триплетов. По-видимому, жгутики всех водорослей имеют «корни», прикрепленные обычно к их базальным телам, закрепляющие эти тела в протопласте. Обычно корни бывают двух типов: группы исчерченных полосатых волокон и группы микротрубочек. Ориентация корней по отношению к базальным телам и в клетке различна. Например, у хламидомонады базальные тела двух жгутиков связаны друг с другом широким полосатым корнем, от которого отходят четыре четырехчленных микротрубчатых корня, идущих назад в клетку непосредственно под цитоплазматической мембраной.
Такая комбинация исчерченных и трубчатых корней встречается часто. Совсем иной тип «корневой системы» у представителей празинофициевых, хризофитовых, рафидофитовых водорослей. У этих организмов широкая исчерченная лента – ризопласт – тянется непосредственно от базальных тел к ядру, где она, по-видимому, прикрепляется к ядерной оболочке. В некоторых группах водорослей, например зеленых, особенностям жгутиковых корней в последнее время придается важное значение в связи с выявлением родственных отношений между видами и родами.
При едином общем плане строения жгутики разных водорослей могут отличаться в деталях, причем эти различия весьма константны и характерны для больших групп – отделов. Эти постоянные различия касаются как особенностей тонкого строения жгута, так и таких признаков, как число жгутиков, место прикрепления, длина, характер поверхности и пр. Так, у диатомовых водорослей в жгутике сперматозоидов отсутствуют центральные микротрубочки и вместо обычного строения 4–2 трубочки расположены по формуле +0, а у зеленой водоросли голенкинии в стержне жгутика сперматозоида имеется только одна центральная микротрубочка +1.
Особенно вариабильной сказалась переходная зона, которая у зеленых водорослей характеризуется так называемым звездчатым телом. У золотистых водорослей между периферическими дублетами и центральными трубочками в этой зоне имеется спиральное тело, а у динофитовых переходная зона благодаря диафрагмам разделена на камеры. Число жгутиков может варьировать от одного до многих, хотя преобладают двужгутиковые формы; прикрепление их к клетке – к концу или сбоку.
Если у двужгутиковых клеток оба жгутика одинаковой длины, то они изоконтные, если разной длины, – то гетерокоптные. Одинакового строения жгутики называют изоморфными, различающиеся по форме – гетероморфными, В последнем случае один из жгутиков может быть гладким, а другой несет прикрепленные к его мембране волосовидные образования – мастигонемы. Мастигонемы имеют сложное и разнообразное строение и расположение. Поверхность жгутиков бывает покрыта различной формы чешуйками или шипами.
Регенерация жгутиков
Перед клеточным делением у одних монадных форм старые жгутики сбрасываются; при этом связь с базальным телом прерывается у проксимального конца переходной зоны в месте превращения триплетов в дублеты. В других случаях половина старого кинетического аппарата может, переходить к дочерним клеткам, а отсутствующие половинки образуются заново. Жгутики не способны делиться и всегда берут начало от базальных тел, репликация которых предшествует клеточному делению: от дистального конца базального тела вырастает стержень жгута, приподнимающий плазмалемму, которая по мере вытягивания жгутика также вытягивается и превращается в его мембрану. При репликации базальных тел формирование нового базального тела индуцируется уже имеющимся, по соседству с которым образуются девять новых триплетов микротрубочек. Базальные тела гомологичны, а в некоторых случаях идентичны с центриолями – центрами, от которых во время митоза у многих водорослей расходятся нити веретена.
Центриоли располагаются парами, образуя дюносому с осями, ориентированными под прямым углом друг к другу, в особом участке гомогенной основной цитоплазмы – центросфере, где нет ни рибосом, ни эндоплазматической сети, ни каких-либо иных клеточных органелл. Центриоли, как и базальные тела, образованы девятью триплетами микрогрубочек, расположенными по кругу. При возникновении из единичной центриоли диплосомы в гомогенной центросфере перпендикулярно к оси имеющейся центриоли появляются девять очень коротких триплетов микротрубочек, которые растут, пока не достигнут длины трубочек материнской центриоли. Никаких контактов между двумя системами микротрубочек обнаружить нельзя – между ними всегда сохраняется расстояние. По-видимому, как и в случае репликации базального тела, каждая существующая центриоль является центром дифференцировки микротрубочек.
Во время митоза центриоли расходятся в противоположные стороны, образуя полюса ядра, к которым сходятся нити веретена. Центриоли могут располагаться вне ядра или же при образовании в оболочке ядра полярных отверстий они вместе с микротрубочками проникают внутрь ядра. Центриоли встречаются не только у монадных форм и водорослей, жизненный цикл которых включает репродуктивные подвижные клетки, они были обнаружены и у водорослей, лишенных подвижных стадий. В одних случаях центриоли мигрируют от ядра к поверхности клетки и начинают функционировать в качестве базальных тел, например у хорды, у хары в вегетативных клетках центриоли отсутствуют, однако имеются в клетках сперматогенных нитей, где они связаны с веретеном, а позднее принимают участие в образовании жгутиков. Во многих других случаях базальные тела функционируют во время митоза в качестве цептриолей.
Псевдоцилии
У зеленых водорослей с пальмеллоидной структурой клетки многих представителей несут неподвижные отростки – псевдоцилии. Электронно-микроскопическое исследование показало, что эти структуры – производные жгутиков, базальные тельца и корневые системы которых почти идентичны таковым хламидомонады; переходная зона также имеет звездчатый узор, характерный для жгутиков зеленых водорослей. Однако свободная часть органеллы не имеет центральных микротрубочек, и «В» трубочки периферических дублетов большей частью очень коротки. Таким образом, под мембраной псевдоцилий микротрубочки имеют расположение 9 –0 вместо характерного для типичных жгутиков +2.
Гаптонема
Ряд золотистых водорослей, помимо двух жгутиков, обладает особой органеллой – гаптонемой. Это расположенный между двумя жгутиками обычно более короткий отросток; он может находиться в скрученном или выпрямленном состоянии. У одного из видов хризохромулины с помощью цейтраферной киносъемки удалось определить скорость скручивания гапотнемы и ее вытягивания. Существует мнение, что гаптонема может служить для временного прикрепления клетки к субстрату. С другой стороны, у видов с очень длинной гаптонемой она может служить приспособлением, облегчающим парение в толще воды, как и всякий вырост клетки. На эту мысль наводят наблюдения за поведением гаптонемы: если клетка движется с помощью жгутиков, гаптонема свернута, если же жгутики перестают работать, гаптонема оказывается далеко вытянутой.
Как и в жгутиках, внутри гаптонемы проходят микротрубочки, расположение которых иное. Гаптонема, как правило, содержит лишь один круг из 5–8 простых микротрубочек. Центральные микротрубочки полностью отсутствуют. У вхождения гаптонемы в клетку добавляются еще 2–3 микротрубочки, так что число их достигает 9. Кроме того, в отличие от жгутиков кольцо трубочек гаптонемы окружено тремя концентрическими обвертками, из которых самая наружная служит продолжением плазмалеммы клетки. Очевидно, микротрубочки это те структурные элементы, которые обусловливают движение как жгутиков, так и гаптонемы; корней у гаптонемы нет.
biofile.ru