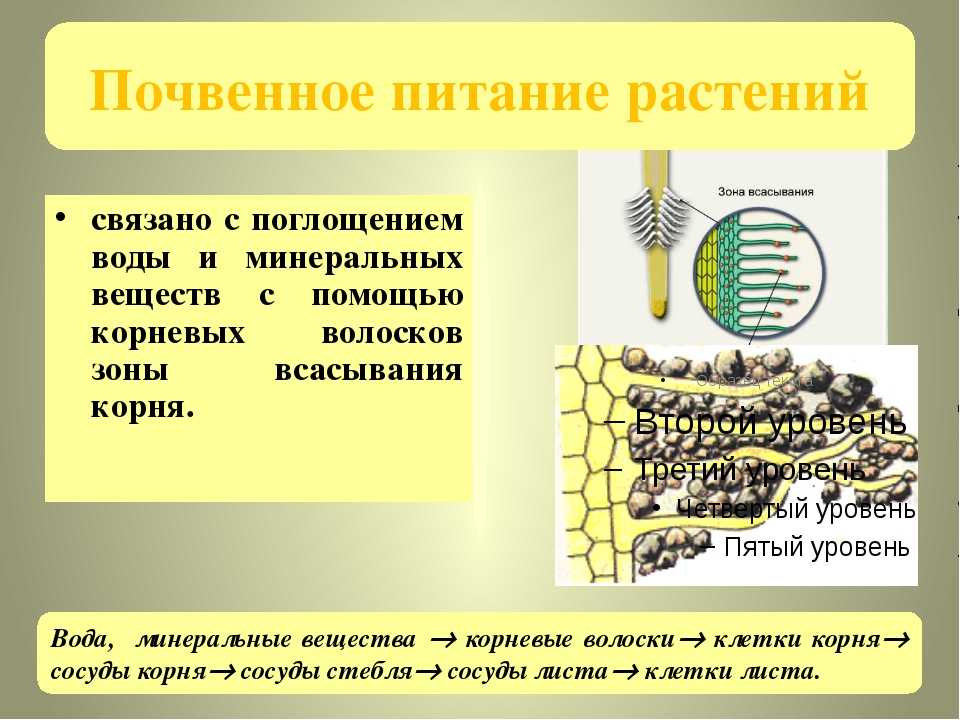
Содержание
Открытие амилоидов у растений может помочь создать сорта с более питательными и гипоаллергенными семенами
18 ноября, 2020 08:44
Источник:
Пресс-служба СПбГУ
Группа исследователей, куда вошли ученые СПбГУ, впервые показала, что особые фибриллярные белки амилоиды содержатся в растениях — они отвечают за «консервацию» питательных веществ внутри семян. Благодаря этой работе, как недавно отметил авторитетный научный журнал Trends in Plant Science, теперь известно, что функциональные амилоиды используют представители почти всех крупных групп живых организмов: бактерии, археи, животные, грибы и растения.
Поделиться
Это фундаментальное открытие в будущем может помочь повысить пищевую ценность семян растений и даже создать менее аллергенные сорта бобовых.
На упаковках доброй половины закусок и сладостей, в составе которых может и вовсе не быть орехов, часто встречается предупреждающая фраза: «Возможно содержание следов арахиса». У некоторых людей аллергия на этот продукт настолько сильна, что мельчайшие частицы ореха или даже его пыль вызывает неприятную и иногда опасную реакцию — от простой сыпи до серьезных отеков. В семенах арахиса содержится множество белков, часть из которых способна вызывать аллергию. Одним из самых сильных аллергенов является вицилин, который есть у различных бобовых, включая арахис и горох.
В своем исследовании, опубликованном в журнале PLOS Biology, группа ученых из СПбГУ, ВНИИ сельскохозяйственной микробиологии (ВНИИСХМ), Института цитологии РАН, Института теоретической и экспериментальной биофизики РАН, Казанского федерального университета и Университета Бургундии во Франции смогла впервые экспериментально показать (ранее им удалось предсказать это с помощью методов биоинформатики), что в семенах гороха посевного содержатся особые фибриллярные белки — амилоиды. Их раньше встречали у бактерий, архей, животных и грибов, но впервые нашли у растений. Что интересно, большую часть амилоидов в семенах гороха образует именно упомянутый выше белок вицилин.
Их раньше встречали у бактерий, архей, животных и грибов, но впервые нашли у растений. Что интересно, большую часть амилоидов в семенах гороха образует именно упомянутый выше белок вицилин.
Вицилин — это один из важнейших пищевых аллергенов, который содержится в бобовых. Механизм его аллергенности потенциально может быть связан с открытыми нами амилоидными свойствами этого белка. Мы показали, что запасные белки, которые являются основным резервуаром питательных веществ для зародыша, накапливаются в семенах в виде амилоидов. В перспективе изучение этих механизмов может помочь создать менее аллергенные сорта гороха, арахиса и других бобовых.
Руководитель научного коллектива, доцент кафедры генетики и биотехнологии СПбГУ, заведующий лабораторией в ВНИИСХМ Антон Нижников
«Интересно, что запасные белки семян не только гороха, но целого ряда растений, не относящихся к бобовым, по полученным нами биоинформатическим данным, оказались обогащены участками, склонными к формированию амилоидов, то есть компактных и стабильных фибриллярных агрегатов.Это объясняет способность семян переживать различные неблагоприятные условия и прорастать спустя долгие годы», — отметил первый автор исследования, доцент кафедры цитологии и гистологии СПбГУ, старший научный сотрудник ВНИИСХМ Кирилл Антонец.
 Это объясняет способность семян переживать различные неблагоприятные условия и прорастать спустя долгие годы», — отметил первый автор исследования, доцент кафедры цитологии и гистологии СПбГУ, старший научный сотрудник ВНИИСХМ Кирилл Антонец.
Это объясняет способность семян переживать различные неблагоприятные условия и прорастать спустя долгие годы», — отметил первый автор исследования, доцент кафедры цитологии и гистологии СПбГУ, старший научный сотрудник ВНИИСХМ Кирилл Антонец.
Еще одно возможное прикладное значение этой работы — создание в будущем культур растений со сверхпитательными семенами. Эксперименты ученых in vitro (в пробирке) показали, что млекопитающие не способны полностью переваривать растительные амилоиды — их не могут расщепить пищеварительные ферменты. Как объясняет Антон Нижников, амилоиды значительно ухудшают пищевую ценность семян, поэтому важно понять, каким образом можно снизить образование амилоидов в семенах растений, чтобы получить сорта с бóльшим количеством обычных белков — такие культуры могут стать для человека особенно полезными и питательными.
«Сегодня мы также занимаемся изучением амилоидов клубеньковых бактерий — эти микроорганизмы способны вступать в симбиоз с бобовыми и связывать атмосферный азот, чтобы растения могли получать больше питательных веществ, — рассказал Антон Нижников.
 — Есть предположение, что во взаимовыгодном симбиотическом процессе амилоиды тоже могут играть важную роль, по крайней мере, у клубеньковых бактерий, как мы показали, тоже есть амилоиды. Надеемся, что наши открытия будут полезны для развития биологии растений и микробиологии, а также для сельского хозяйства».
— Есть предположение, что во взаимовыгодном симбиотическом процессе амилоиды тоже могут играть важную роль, по крайней мере, у клубеньковых бактерий, как мы показали, тоже есть амилоиды. Надеемся, что наши открытия будут полезны для развития биологии растений и микробиологии, а также для сельского хозяйства».
Напомним, что особая фибриллярная форма белков — амилоиды — стала известна благодаря своей связи с рядом заболеваний, вызванных аномальной агрегацией белка, называемых амилоидозами. При этих тяжелых недугах мономерные растворимые белки превращаются в полимерные фибриллярные формы, образующие амилоидные «бляшки» в различных тканях и органах. Всего существует более 40 болезней человека, связанных с амилоидами, причем они с большим трудом поддаются лечению или являются вовсе неизлечимыми.
Все же, как подтверждает и данное исследование, в последние десятилетия ученые по всему миру находят все больше свидетельств тому, что амилоиды функционируют и в здоровых организмах: зачастую такая форма белка позволяет «законсервировать» и стабилизировать различные вещества, к тому же она выступает в качестве своеобразной структурной «матрицы».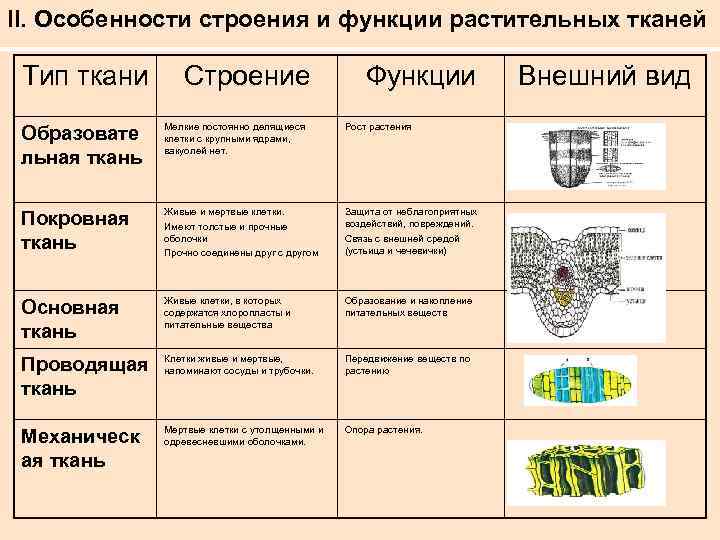 Происходит это не только в растениях. Например, у человека и животных часть гормонов запасается именно в форме амилоидов, а другие функциональные амилоиды участвуют в биосинтезе меланина и формировании долговременной памяти.
Происходит это не только в растениях. Например, у человека и животных часть гормонов запасается именно в форме амилоидов, а другие функциональные амилоиды участвуют в биосинтезе меланина и формировании долговременной памяти.
Исследование поддержано грантом Российского научного фонда. Экспериментальная часть работы была выполнена с использованием оборудования Научного парка СПбГУ.
Теги
Сельское хозяйство
ЛЕКЦИЯ 5
ЛЕКЦИЯ
5.
Производные
протопласта.Запасные
питательные вещества.Продукты
распада. Кристаллы.Физиологически
активные вещества.Химический
состав клеточного сока, его пигменты.Клеточная
стенка и ее видоизменения.
Характерной
особенностью живой
материи является способность к
постоянному обмену веществ, который
складывается из реакций синтеза
(ассимиляции) и реакций распада
(диссимиляции). Растительные клетки
отличаются интенсивной синтетической
деятельностью, причем синтез может
быть
первичным и вторичным. При первичном
синтезе происходит образование
органических веществ из минеральных.
Он идет при участии
энергии солнца и называется, как
известно, фотосинтезом. При
вторичном синтезе осуществляется
преобразование органических
соединений — из сахара образуется
крахмал, из аминокислот — белки
и т. п. Вторичный синтез протекает без
доступа света, за счет
внутриклеточной энергии, которая
выделяется при окислительных
процессах (дыхании) в клетке. Наряду с
реакциями синтеза в клетках совершается
процесс расщепления веществ на более
простые соединения, многие из которых
не участвуют в дальнейшем метаболизме.
В результате в клетке появляются
различные продукты
распада (катаболиты*). Все вещества,
вырабатываемые протопластом
в результате его жизнедеятельности,
составляют группу внутриклеточных
включений.
 Растительные клетки
Растительные клетки Вещества,
нерастворимые в воде, образуют в клетках
оформленные
включения
в
виде капель, зерен, кристаллов.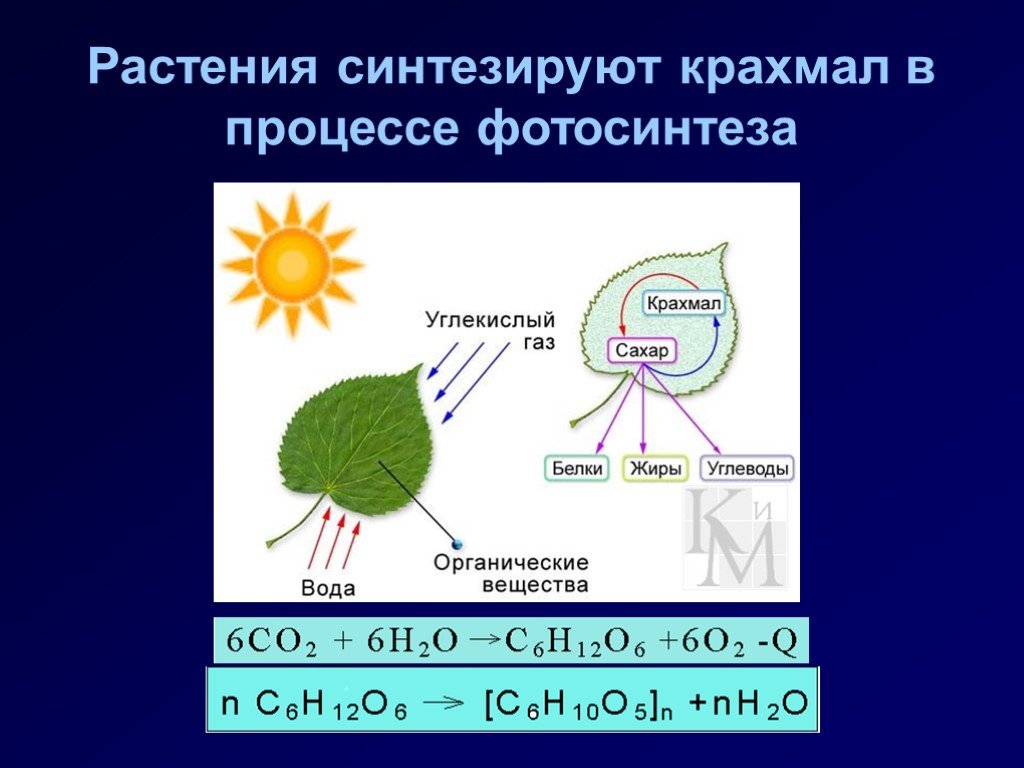 Растворимые
Растворимые
продукты обмена входят в состав клеточного
сока,
который накапливается в вакуолях и
относится к ж и д к и м(неоформленным)
включениям
клетки.
Включения
не являются постоянными компонентами,
они могут появляться и исчезать в
зависимости от физиологического
состояния клетки.
В
соответствии с ролью и значением в
жизнедеятельности клетки все
внутриклеточные включения можно
подразделить на 3 группы: запасные
питательные вещества, продукты распада
(катаболиты)’ и физиологически активные
вещества клетки.
Накопление
большого количества питательных веществ
является особенностью
растительных клеток. Эти вещества
частично используются
клеткой как
энергетический материал, окисляясь в
процессе дыхания,
в
результате чего освобождается энергия,
необходимая для всех
протекающих
в клетке жизненных процессов. Кроме
того, из запасных
питательных веществ образуются
конституционные вещества,
идущие
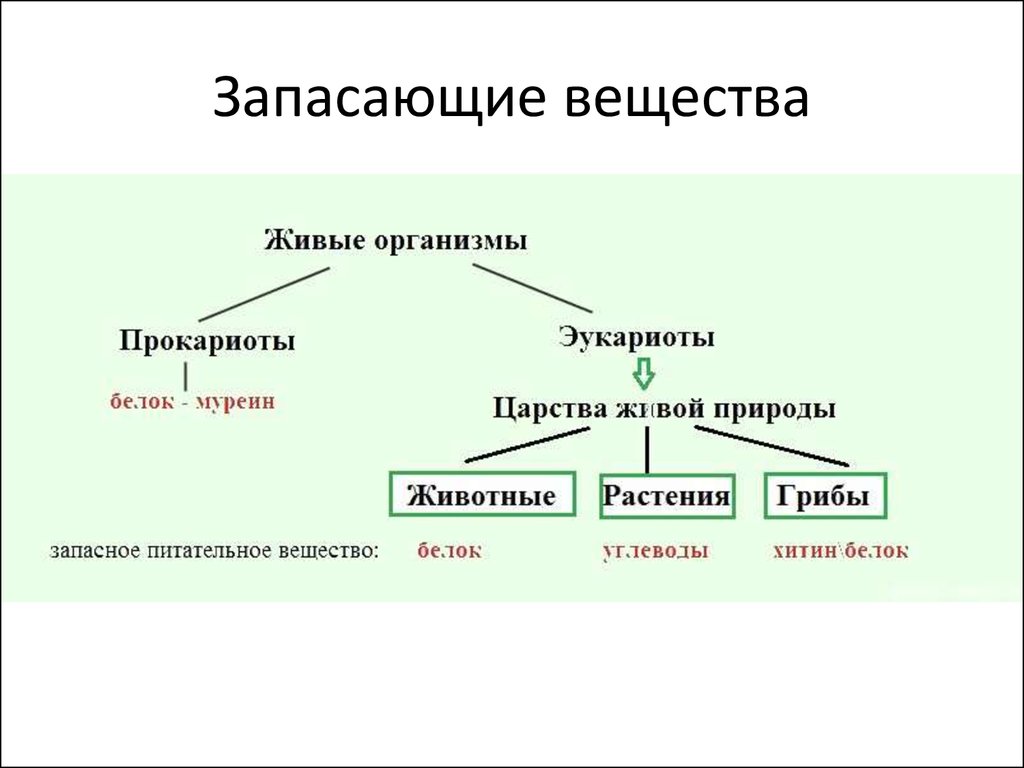
на построение тела растений. Запасные
питательные вещества
встречаются в растительных клетках в
виде углеводов, белков и
жиров. .
 Запасные
Запасные Углеводы
в
растительных клетках присутствуют в
виде полисахаридов,
дисахаридов и моносахаридов. Полисахари
д
ы представлены в основном крахмалом
(РАССМАТРИВАЕМ НА ПРАКТИКЕ!!!) однако
встречаются также гликоген,
инулин и гемицеллюлоза (полуклетчатка).
Крахмал является
одним из наиболее распространенных
углеводов, накапливающихся в клетках
растений в качестве запасного питательного
вещества.
В
его
образовании обязательно участвуют
пластиды. По
происхождению в растениях различают
крахмал ассимиляционный
(первичный), запасной (вторичный) и
транзиторный (передаточный).
Ассимиляционный
крахмал
синтезируется
в зеленых
частях растений и является одним из
первоначальных продуктов
фотосинтеза. Образование ассимиляционного
крахмала возможно
только в присутствии света и хлоропластов,
в которых он откладывается
в виде мельчайших зерен шаровидной или
палочковидной формы. Однако накопление
Однако накопление
крахмала в зеленых органах растений
в большом количестве, как правило, не
происходит. Образовавшийся в них
ассимиляционный крахмал под действием
фермента амилазы переводится в растворимую
форму, т. е. гидролизуется до сахара,
который и транспортируется в
запасающие органы растения, специально
приспособленные для накопления
питательных веществ. В этих органах
из притекающих к ним сахаров в присутствии
фермента а м и л о с и н т е а з ы снова
образуется крахмал— вторичный,
или запасной.
Запасной
крахмал накапливается в клубнях,
корневищах, корнях, семенах и других
органах растений.
Особенно много крахмала содержится
в клубнях картофеля (12…20%), семенах
риса (60…80%), кукурузы (65… 75%),
пшеницы (60…70%). Образование
вторичного крахмала осуществляется
при участии бесцветных пластид
лейкопластов и может
проходить без доступа света. Запасной
крахмал находится в клетках растений
в виде зерен различной величины — от
0,002 до
0,15 мм в диаметре.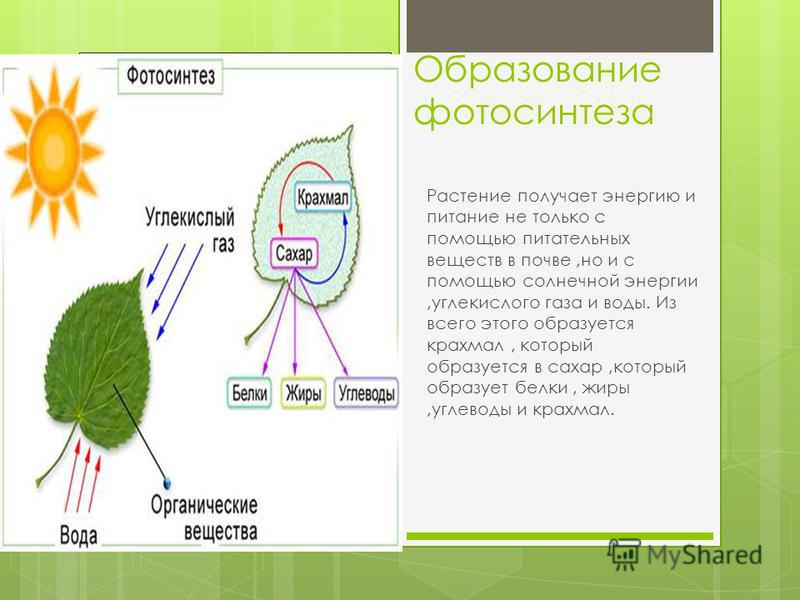 По форме они бывают
По форме они бывают
шаровидные, чечевицеобразные,
эллиптические, палочковидные и т. п.
Образование
крахмального зерна начинается с
возникновения в
лейкопласте образовательного центра,
вокруг которого стромой лейкопласта
слоями откладывается вещество крахмала.
Слои содержат
различное количество воды и имеют
различный коэффициент преломления
света, благодаря чему они хорошо видны
в микроскоп. Если
отдельные слои откладываются вокруг
образовательного центра
равномерно, формируются крахмальные
зерна с концентрической слоистостью
(злаки, бобовые). Если слой крахмала
откладываются вокруг
образовательного центра неравномерно,
возникают крахмальные
зерна с эксцентрической слоистостью
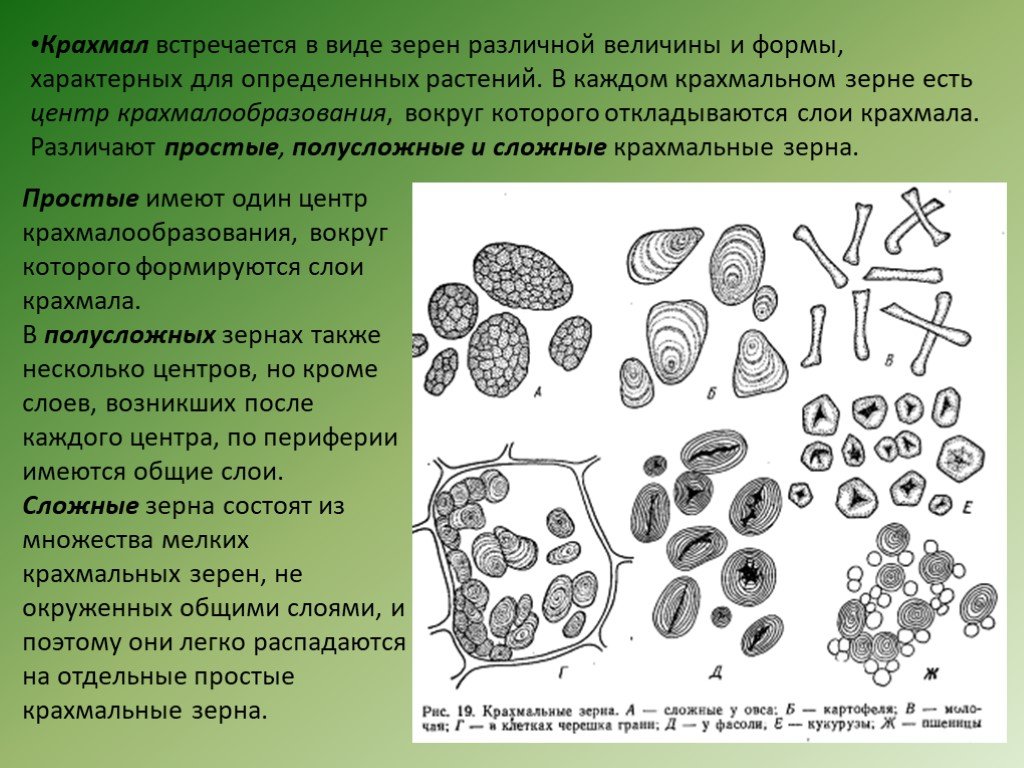
(картофель). Различают крахмальные
зерна простые, сложные и полусложные.
Простые
имеют один образовательный центр.
Сложные состоят из
множества очень мелких простых крахмальных
зерен, имеющих каждое свой образовательный
центр и слоистость.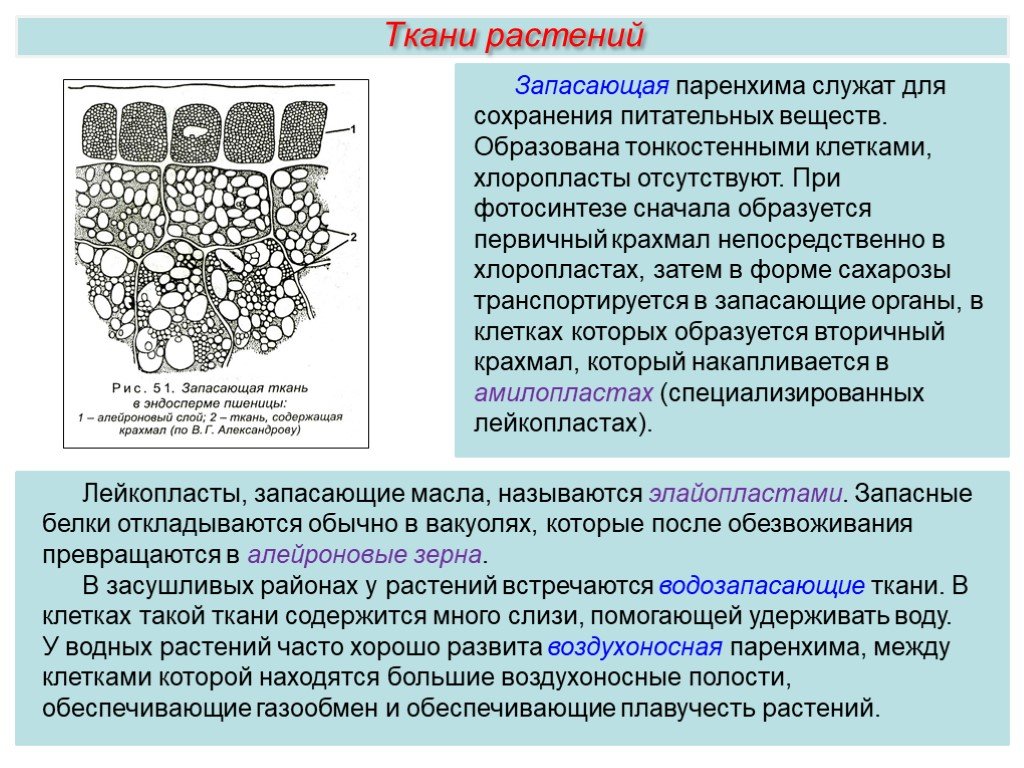 В состав сложного
В состав сложного
зерна может входить несколько тысяч
простых зерен (шпинат). В
полусложных крахмальных зернах — 2
образовательных центра, окруженных
общими слоями. Все крахмальные зерна
представляют собой сферокристаллы,
состоящие из тончайших радиально
расположенных
игл.
Форма
и величина крахмальных зерен специфичны
для отдельных
семейств, родов и даже видов растений.
Так, у картофеля они отличаются
неправильной .формой, эксцентрической
слоистостью и
Достигают размера .70… 100 мкм. Крахмальные
зерна бобовых значительно
мельче, овальные, с концентрической
слоистостью, и в центре у них обычно
образуется продольная трещина. У риса,
овса,
гречихи крахмальные зерна
сложные, легко распадающиеся
на множество простых
зернышек неправильной формы.
Транзиторный
крахмал
нередко
образуется
на путях следования сахаров
от фотосинтезирующих
органов к запасающим. Крахмал
Крахмал
окрашивается йодом
в синий цвет., медным
купоросом
и едким калием —в
фиолетовый цвет. Он нерастворим в
холодной воде, а в горячей
набухает, образуя клейстер.
Крахмал имеет как
питательное вещество, необходимое
растениям, животным и человеку, но и как
сырье для промышленного
производства глюкозы и спирта.
У
незеленых растений — бактерий, грибов,
а также некото
рых
водорослей — вместо крахмала накапливается
запасной поли
сахарид
гликоген,
более характерный для клеток
животных
организмов.
В отличие от крахмала гликоген является
воднорастворимым
веществом. . х
Другим
углеводом, заменяющим у некоторых
растений крахмал,
является и н у л и н. Он образуется в
клубнях земляной груши, корнях
цикория, одуванчика и вообще характерен
для представителей семейства
сложноцветные (астровые). Подобно
Подобно
гликогену, инулин
растворяется в воде, но под действием
спирта выпадает из раствора в виде
сферокристаллов. По химическому составу
гликоген и инулин близки к крахмалу и
имеют одинаковую с
ним эмпирическую формулу.
Г
е м и ц е л л ю л о з a
(CsH804)
n
встречается
в семенах кофейного
дерева, финиковой пальмы, многих видов
люпина, представителей
семейства лилейные и др., накапливаясь
в клеточных оболочках.
Под действием ферментов гемицеллюлоза,
подобно крахмалу и целлюлозе, может
превращаться в сахар.
Моносахариды
и дисахариды встречаются в клетках
растений в виде различных сахаров в
растворенном состоянии.
Моносахариды
(СвН12Ое)
представлены виноградным сахаром
— глюкозой и плодовым сахаром —
фруктозой. Эти сахара накапливаются
преимущественно в плодах (яблоня, груша,
виноград),
а также в стеблях (кукуруза, сорго),
листьях (лук) и других органах
растений.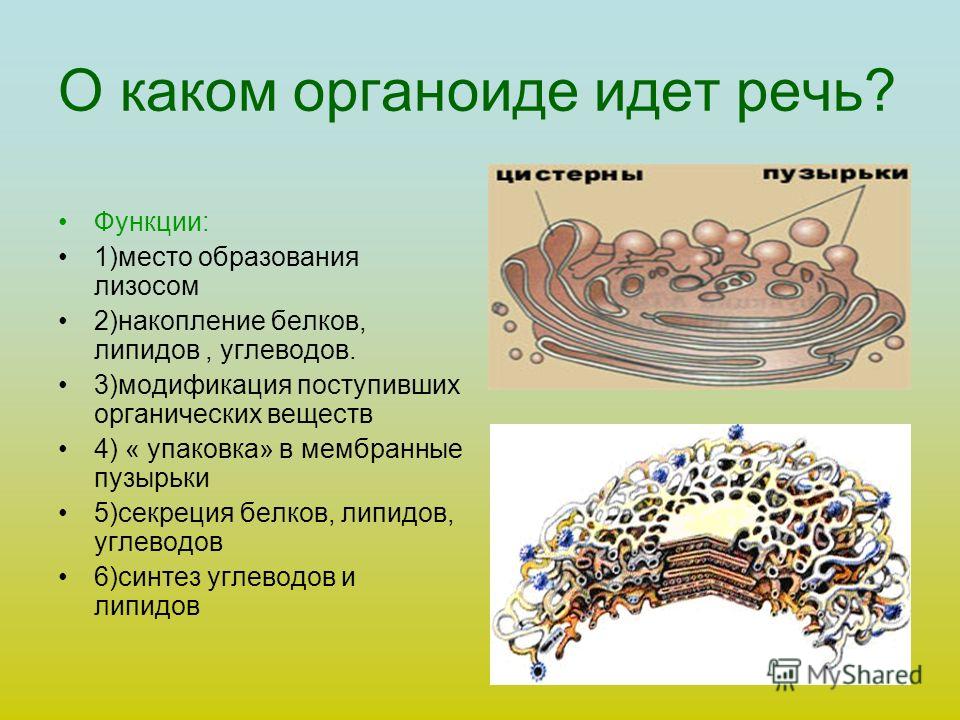 .
.
Дисахариды
(С12Н22Ои)
встречаются обычно в виде тростникового
или свекловичного сахара (сахарозы) и
накапливаются в
корнеплодах сахарной свеклы, стеблях
сахарного тростника, плодах
арбуза и других растений.
Белки,
накапливающиеся
в клетках в качестве запасного
питательного вещества, необходимо
отличать от конституционных живых
белков, составляющих основу протопласта.
Запасные белки — протеины — являются
простыми белками. В отличие от сложных
(конституционных) белков они состоят,
только из аминокислот. Для запасных
белков характерна инертность, в силу
которой они с большим трудом вступают
в различные реакции. Запасные белки
откладываются в форме алейроновых
(протеиновых) зерен (в семенах злаков,
бобовых)
или в виде кристаллоидов (в клубнях
картофеля),
которые отличаются от настоящих
кристаллов
способностью к набуханию и окрашиванию.
Алейроновые зерна образуются из
вакуолей
в результате их обезвоживания,
что наблюдается при созревании семян.
В прорастающих
семенах происходит обратный процесс —
набухание, и
алейроновые
зерна снова превращаются в вакуоли.
Размеры и
строение
алейроновых зерен очень изменчивы, но
характерны для
определенных групп
растений и могут служить
систематическим
признаком.
Алейроновые зерна бывают простые и
сложные. Простые
содержат аморфный белок, в сложных
имеются еще кристаллоид
белка и особое округлое тельце глобоид,
в
состав
которого входят кальций, магний и фосфор.
Содержание
белка в сельскохозяйственных растениях
также весьма
различно. Так, в семенах люпина белки
составляют 35% от массы
сухого вещества, фасоли — 25%, гороха
29%, пшеницы — 12%,
кукурузы—10%, картофеля — 8…10%.
От
йода, белковые зерна окрашиваются в
темно-желтый цвет. В
горячей воде, кислотах и щелочах запасные
белки растворяются почти
полностью.
Жиры
(жирные
масла) представляют собой сложные эфиры
— соединение
жирных кислот с глицерином. Они состоят
из тех же химических
элементов, что и углеводы, но отличаются
от них меньшим
содержанием кислорода (С/гН2д02).
Запасные жиры широко
распространены
в растительных клетках и обычно
сосредоточены
в
цитоплазме, пластидах и митохондриях.
По-видимому, жиры возникают
непосредственно в цитоплазме, а также
образуются в особом
типе лейкопластов — олеопластах.
Наиболее богаты ими семена
и
плоды растений. Особенно много жиров
содержится в семенах
масличных
культур:
в среднем у подсолнечника — 46…51% от
массы
сухого вещества, у льна — 37%, у хлопчатника
— 23%,
у
конопли — 34%. Жиры
не растворяются вводе, но хорошо
растворяются
в бензине, серном эфире, хлороформе и
т. д. По сравнению
с
другими питательными веществами жиры
являются наиболее
калорийными:
в среднем 1 г жира дает 38,9 кДж (9,3 ккал),
белка —
23,8
кДж (5,7 ккал), крахмала — 17,6 кДж (4,1 ккал).
У подавляющего
большинства растений жирные масла
жидкие и встречаются
в клетках в виде
капель различного размера. Твердые жиры
характерны
для семян шоколадного дерева и кокосовой
пальмы. Жиры
имеют большое значение
не только как высококалорийные
пита
тельные
вещества, но также применяются человеком
в лакокрасочной,
мыловаренной промышленности и в качестве
смазочных мате
риалов.
Продукты
распада
(к
а т а б о л и т ы). Наряду
с
запасными питательными веществами в
клетках растений образуются вещества,
которые обычно не участвуют в дальнейших
химических
процессах и называются катаболитами.
Они могут накапливаться
в специальных вместилищах или выделяются
в окружающую среду. К ним относятся
эфирные масла, алкалоиды, гликозиды,
дубильные вещества, соли щавелевой
кислоты, смолы, каучук и др.
Эфирные
масла
встречаются
значительно реже, чем жирные,
и характерны только для растений семейств
зонтичные (сельдерейные),
рутовые, губоцветные (яснотковые) и
некоторых других.
Обычно эфирные масла обладают летучестью
и сильным специфическим
запахом. Они встречаются в виде небольших
капелек и
скапливаются в различных частях растений
— корнях, корневищах,
листьях, стеблях, плодах и других органах.
Эфирные масла защищают растения от
поедания животными, многие из них
обладают
бактерицидными свойствами. Особенно
Особенно
богаты эфирными маслами
такие растения, как мята, эвкалипт, роза,
тмин, апельсин и некоторые другие. Многие
растения (кориандр, мята, герань)
возделываются
в широких масштабах в качестве
эфирномасличных культу]). Эфирные масла
широко используются в технике, медицине,
парфюмерии, кондитерской и других
отраслях промышленности.
Алкалоид
ы
представляют собой азотистые соли
органических
кислот — яблочной, лимонной, винной и
др. Они образуются во всех частях растений
— в корнях (белладонна), клубнях
(картофель), листьях (табак, чайное
дерево), плодах (мак, кофейное дерево),
семенах (дурман, люпин, какао) и т. д. В
настоящее время известно
свыше 1000 различных алкалоидов. Они имеют
для растений защитное
значение — предохраняют их от поедания
животными, иногда
играют роль запасных веществ, а также
фитогормонов и стимуляторов,
вызывающих усиление процессов обмена
веществ на тех или иных фазах роста.
Народохозяйственное
значение алкалоидов и алколоидоносных
растений очень велико. Многие алколоиды
Многие алколоиды
(никотин, атропин, кокаин, кофеин, хинин
и др.)широко применяются в медицине,
ветеринарии и сельском хозщяйстве.
Гликозиды
представляют
собой соединения глюкозы со спиртами
и другими безазотистыми веществами.
Они имеют горький вкус
и обладают ядовитыми свойствами,
благодаря чему предохраняют
растения от поедания животными. Гликозиды
многих растений
(ландыш, наперстянка и др.) применяются
в медицине. Для промышленности
большое значение имеют глнкозиды-красители.
Соли
щавелевой кислоты в растительных клетках
чаще всего встречаются
в виде щавелевокислого
кальция,
который
образует кристаллический песок,
сферокристаллы или кристаллы
иной формы в зависимости от вида растений.
Различают
одиночные
кристаллы,
встречающиеся в сухих наружных
чешуях луковиц репчатого лука и чеснока;
друз
ы, представляющие
собой сростки многочисленных кристаллов
звездчатой
формы (в плодах жимолости, в коре многих
древесных растений),
и рафиды
— игольчатые кристаллы, часто образующие
пучки
(в клетках мякоти плодов фуксии, листьев
лилии). Все формы, кристаллов
Все формы, кристаллов
локализуются в вакуолях. Благодаря
образованию кристаллов щавелевокислого
кальция происходит нейтрализация
щавелевой кислоты, обладающей ядовитыми
свойствами.
Кроме
щавелевокислого кальция, у некоторых
растений (фикус,
конопля) образуется у г л е к.и с л ы й
кальций,
который пропитывает
выросты клеточной оболочки, вдающиеся
в полость клетки.
В результате получаются своеобразные
гроздевидные образования
— цистолиты.
Кристаллы,
являясь конечным продуктом обмена
веществ в клетке,
как правило, тем или иным способом
удаляются из организма.
Обычно
они накапливаются в тех частях растения,
которые со временем от него отделяются,
— в листьях, плодах, наружных
слоях коры. Однако в некоторых случаях
кристаллы могут растворяться вновь и
участвовать в обмене веществ, как это
наблюдается
в плодах апельсина и некоторых
других растений.
Смолы
являются
комплексными соединениями,
образующимися из углеводов в процессе
нормальной жизнедеятельности
клеток или в результате их разрушения,
У одних растений смолы накапливаются
в виде капель в клетках, у других
выделяются в окружающую среду.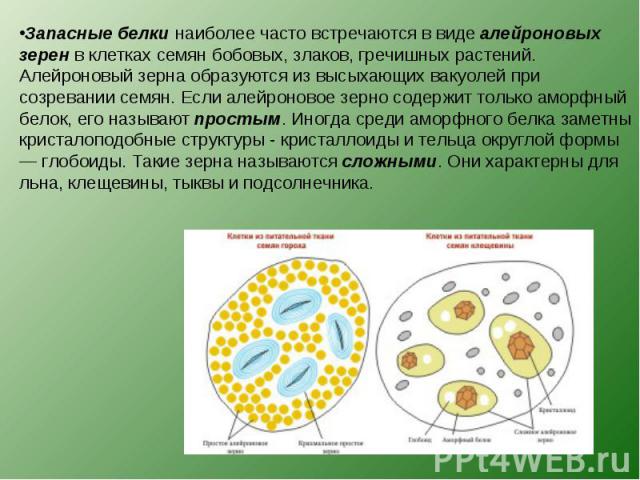
Будучи нерастворимыми в воде, смолы не
пропускают влагу, они непроницаемы для
микроорганизмов, обладают
антисептическими свойствами.
В
практической деятельности человека
смолы применяются при изготовлении
лаков, смазочных масел, в медицине.
Особое значение имеет смола вымерших
растений — янтарь.
Д
у б и л ь н ы е (дубящие)
вещества
представляют собой сложные органические
безазотистые вещества вяжущего вкуса.
Они широко распространены среди высших
растений, причем особенно
богаты ими клетки коры деревьев (дуб,
ель, ива), листья чая,
семена кофе. Обладая антисептическими
свойствами, дубильные вещества
защищают растения от поражения различными
микроорганизмами, иногда они могут
использоваться в качестве запасных
питательных
веществ..
Дубильные
вещества применяются в кожевенной
промышленности
для дубления кож, а также в медицине как
вяжущее средство.
Физиологически
активные вещества обусловливают
нормальную жизнедеятельность
клетки и всего организма в целом. Они
обладают специфическим
действием и неразрывно связаны с
метаболизмом клетки. К
этим веществам принадлежат ферменты,
витамины, фитогормоны, антибиотики,
фитонциды и ингибиторы. Все эти вещества
вырабатываются
протопластом клетки.
 Они
Они Ферменты
(энзимы) представляют собой сложные
вещества белковой
природы и являются биологическими
катализаторами, присутствие которых
необходимо для возбуждения и ускорения
биохимических реакций, протекающих в
клетке. Важнейшие, жизненные
процессы — дыхание, фотосинтез, синтез
и распад белков и
др. — могут совершаться только под
воздействием определенных ферментов.
Ферменты отличаются от неорганических
катализаторов
высокой специфичностью, т. е. действие
каждого фермента строго ограничено
одним веществом или группой близких
веществ. Специфичность
действия ферментов является их важнейшим
биологическим
свойством, без которого невозможен
нормальный метаболизм клетки. Активность
ферментов зависит от температуры,
кислотности среды
и от присутствия в окружающей среде
различных веществ, усиливающих или
подавляющих их каталитическое действие.
В настоящее
время известно свыше 800 различных
ферментов.
Начало
изучения ферментов относится к 1814 г.,
когда русский ученый
К. С. Кирхгоф показал, что в прорастающем
зерне имеется вещество, способное
превращать крахмал в сахар. В дальнейших
исследованиях
ферментов большая роль принадлежит
советским ученым
А. И. Опарину, А. Л. Курсанову, Н. М. Сисакяну,
Б. А. Рубину
и другим, впервые начавшим изучать
ферменты в живых растениях
и заложившим основу биологии ферментов.
Важным
свойством ферментов является их
способность сохранять активность
вне живой клетки. На этом свойстве
основано применение
ферментов в различных отраслях пищевой
промышленности — хлебопечении,
виноделии, производстве сахара, чая,
какао, табака и др.
Витамины(НА
ПРАКТИКЕ!!!)
представляют
собой органические вещества раз
личной
химической природы и почти исключительно
растительного
происхождения.
Однако, несмотря на большое разнообразие,
их
объединяют
в одну группу благодаря той исключительной
роли,
которую
они играют в обмене веществ. Витамины,
Витамины,
действующие
в
очень малых дозах, совершенно необходимы
для нормальной жизнедеятельности
как растительных, так и животных
организмов.
Хотя
витамины не являются непосредственными
источниками энергий,
они вместе с ферментами регулируют
энергетические изменения внутри
клетки, а многие из них даже входят в
состав ферментов.
В
настоящее время известно несколько
десятков различных витаминов,
каждый из которых обладает специфическим
действием. Так,
витамин В3
стимулирует рост корней, витамин С
(аскорбиновая кислота)
способствует прорастанию семян,
регулирует дыхание и
т. д. Однако значение витаминов для
растений изучено еще недостаточно.
Гораздо больше сведений имеется о роли
витаминов в жизнедеятельности
животных организмов. Отсутствие витаминов
в пище животных и человека вызывает
тяжелые заболевания.
Основоположником
учения о витаминах является русский
ученый
Н. И. Лунин, который еще в 1880 г. доказал
И. Лунин, который еще в 1880 г. доказал
необходимость витаминов для нормальной
жизнедеятельности животных организмов.
В результате дальнейшего изучения
витаминов была установлена
их химическая природа, что позволило
организовать промышленное производство
большинства витаминов как из растительного
сырья,
так и синтетическим путем.
Гормоны,
вырабатываемые протопластом растительной
клетки, получили название ф и т о горм
о нов.
Они представляют собой группу
веществ, способных усиливать различные
физиологические процессы
— рост, размножение, деление клеток и
др. Наиболее изучены
в настоящее время гормоны роста —
ауксины, впервые исследованные Н. Г.
Холодным. Ауксины усиливают доступ
кислорода
и приток питательных веществ к клеткам,
расположенным в
растущих частях растения, и таким образом
создают оптимальные условия
для ростовых процессов.
Наряду
с ауксином, который вырабатывается
клетками высших
растений, известны ростовые вещества,
вырабатываемые низшими
растениями — грибами. К таким веществам
К таким веществам
относится гиббереллин,
выделенный из почвенных грибов Gibberella
и Fusarium
и
обладающий совершенно исключительной
и многосторонней физиологической
активностью.
В
настоящее время ростовые вещества
получили широкое применение
в практике сельского хозяйства.
Синтетически получаемый гетероауксин
используется для укоренения черенков,
для борьбы с
опадением бутонов и плодов, для повышения
семенной продуктивности растений и
т. д. Гиббереллин применяется для
получения высокорослых
и сильно облиственных растений (соя,
табак, конопля), повышения
урожая овощных культур (томата, огурца,
баклажана) и
винограда. С
помощью гиббереллинов удается прерывать
период
покоя у семян, спящих почек, клубней,
ускорять цветение и
плодоношение, вызывать образование
бессемянных плодов. С помощью
гиббереллина можно также превращать
двулетние растения (морковь, свекла,
капуста) в однолетние, плодоносящие в
1-й год жизни.
Антибиотики
и
фитонциды
— это особые вещества, которые
вырабатываются в клетках растений и
имеют для них защитное
значение, предохраняя от поражения
болезнетворными микроорганизмами
и другими паразитами.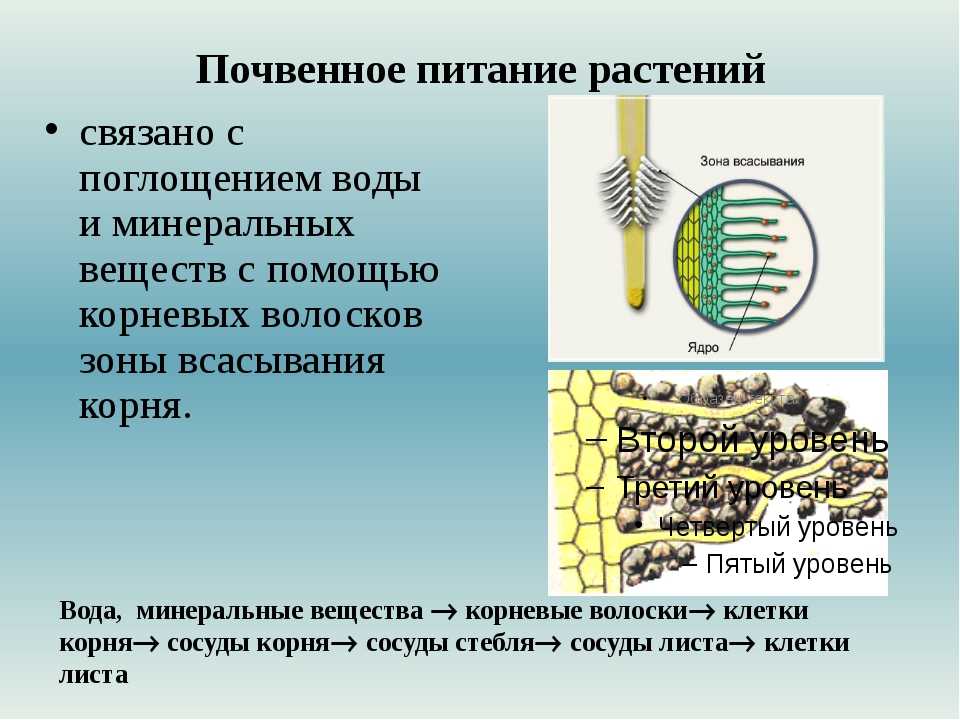 Принято называть
Принято называть
бактерицидные
вещества, образующиеся в клетках низших
растений (грибов и
некоторых бактерий),-антибиотиками, а
аналогичные вещества, выделяемые
клетками цветковых растений (лука,
чеснока, черемухи
и др.), — фитонцидами. Основоположником
учения о фитонцидах
является советский ученый Б. П. Токин.
Бактерицидные вещества
обладают способностью оказывать
губительное действие на различные
микроорганизмы, убивая или сильно
задерживая рост.
Как фитонциды, так и антибиотики действуют
избирательно, вследствие
чего для одних организмов они весьма
токсичны, тогда как
для других—совершенно безвредны.
Фитонциды некоторых растений обладают
настолько сильным действием, что убивают
насекомых
и даже мелких млекопитающих. В настоящее
время многие антибиотики получили
широкое применение в медицине в
качестве лечебных препаратов для борьбы
с тяжелыми инфекционными
болезнями. Общеизвестны такие препараты,
как пенициллин, стрептомицин,
синтомицин и др., получаемые в большом
количестве
заводским путем.
В
практике сельского хозяйства начинают
применяться фитонцидные препараты
для борьбы с различными заболеваниями
растений.
Так, например, протравливание зерен
проса, зараженных пыльной
головней, фитонцидами сарептской горчицы
повышает урожай
проса больше чем в 3 раза. Фитонциды
репчатого лука, чеснока,
цитрусовых губительно действуют на
гриб фитофтору, поражающий
картофель.
Ингибиторами
называют
вещества, подавляющие активность
ферментов и таким образом способствующие
торможению некоторых
физиологических процессов, протекающих
в растении. Тормозящее
действие ингибиторов имеет большое
биологическое значение.
Благодаря ингибиторам при преждевременном
потеплении
ранней весной задерживается распускание
почек. Ингибиторы обеспечивают период
покоя растений, во время которого не
происходит
прорастания клубней, семян и т. д.
Клеточный
сок. Как
уже отмечалось, растворимые продукты
обмена веществ образуют водный раствор,
называемый клеточным
соком. Он постепенно накапливается в
вакуолях, и для взрослой,
полностью дифференцированной клетки
характерна одна крупная
центральная вакуоль, объем которой
часто почти равен объему всей
клетки. Состав клеточного сока весьма
разнообразен и в первую очередь
зависит от вида растения. У большинства
растений клеточный
сок имеет кислую реакцию, исключение
составляют огурец, дыня
и некоторые другие растения, у которых
реакция клеточного сока
щелочная.
 Он постепенно накапливается в
Он постепенно накапливается в Помимо
веществ, рассмотренных выше (растворимые
углеводы, белки,
алкалоиды и др.), клеточный сок содержит
различные кислоты,
соли и пигменты. Из органических кислот
чаще встречаются
яблочная (в плодах яблони, малины, рябины,
листьях табака), щавелевая
(в листьях щавеля, кислицы, ревеня),
винная (в плодах винограда,
томата) и лимонная (в плодах лимона,
смородины, крыжовника,
земляники). К органическим кислотам
принадлежит также
бензойная кислота, содержащаяся в плодах
брусники и клюквы
и обладающая способностью предохранять
эти растения от различных
болезней. Органические кислоты выполняют
Органические кислоты выполняют
в клетках растений
разнообразные физиологические функции,
например участвуют в процессе дыхания.
Минеральные соли представлены в
клеточном
соке нитратами, фосфатами, хлоридами и
другими соединениями. Высоким содержанием
нитратов отличаются крапива, щирица,
картофель, подсолнечник, фасоль. В
молодых частях растений обычно
накапливаются фосфаты — у лука, чеснока
и др. Хлориды характерны для растений,
произрастающих на засоленных почвах.
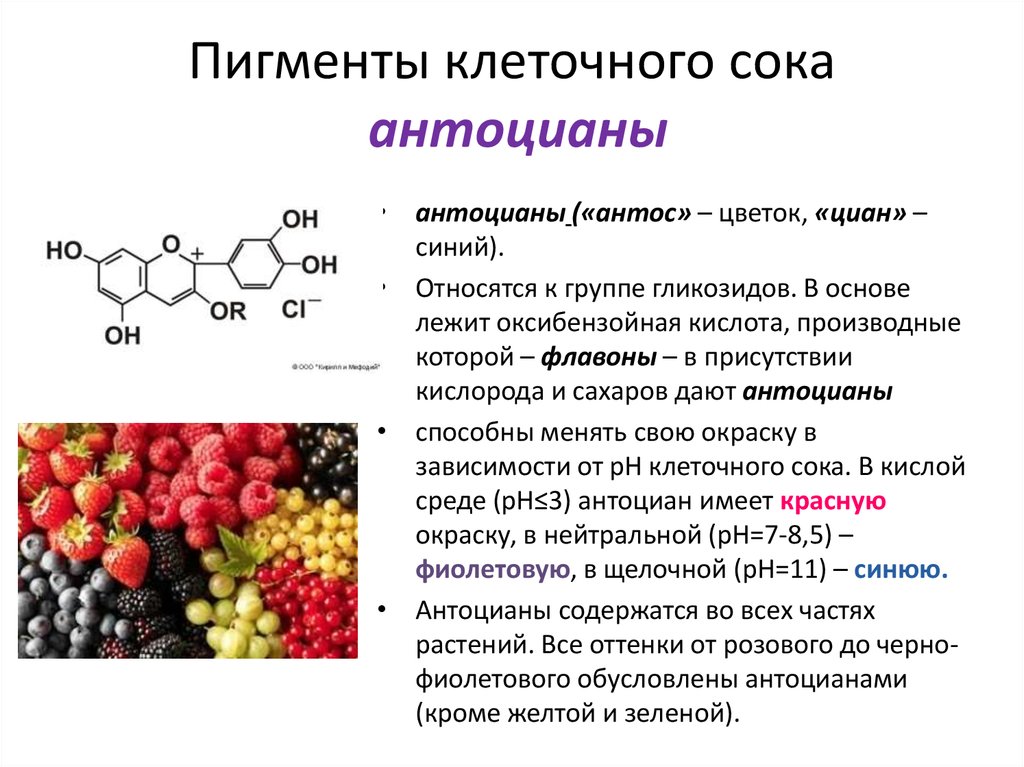
Наряду
с пигментами пластид у растений известны
пигменты клеточного
сока, из которых наиболее распространены
антоциан и
антохлор, относящиеся к гликозидам.
Особенностью антоциана является
изменение его окраски в зависимости от
кислотности среды: в нейтральной среде
он фиолетовый, в щелочной — синий и
в кислой — красный. Антоциан
встречается во всех органах растений
— корнях, листьях, цветках, плодах и в
зависимости от его
концентрации и особенностей организма
может давать самые разнообразные
окраски — от ярко-красных и синих до
почти черных.
Часто присутствие антоциана в клетках
связано с приспособлением
растений к неблагоприятным условиям
внешней среды и обеспечивает
повышение зимостойкости растений.
Антохлор
встречается
преимущественно в венчиках цветков,
которым придает желтую
окраску (у льнянки, георгина, коровяка
и др.), а также в плодах некоторых
цитрусовых.
Клеточный
сок некоторых растений имеет белую
(молочную) окраску,
вследствие чего получил название
млечного
сока.
Млечный
сок (латекс) вырабатывается многими
травянистыми и древесными
растениями. Он представляет собой
эмульсию или суспензию и содержит до
80 % воды, в которой находятся как запасные
питательные вещества (сахара, белки,
жиры), так и катаболиты
(алкалоиды, гликозиды, смолы, дубильные
вещества, а также каучук
и гуттаперча). Часто в нем встречаются
крахмальные зерна своеобразной
формы. У некоторых растений млечный сок
имеет, желтую
(мак) или оранжевую (чистотел) окраску,
что обусловлено присутствием
различных пигментов. Млечный сок
скапливается в
специальных элементах — млечниках.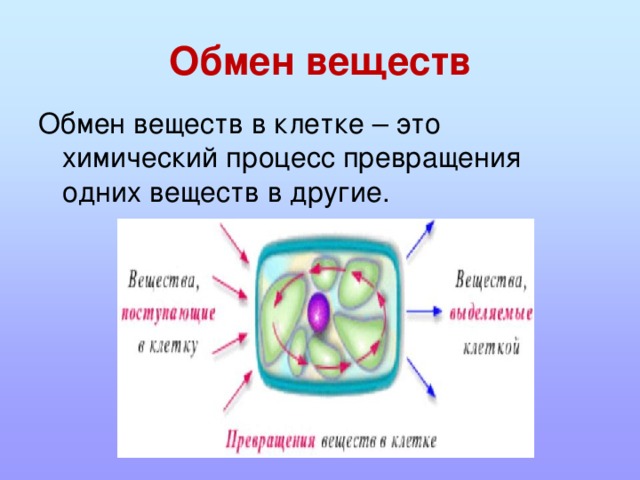
Роль млечного сока в жизни растений
отчасти связана с хранением питательных
веществ, с защитой
от поедания животными, однако значение
его еще недостаточно выяснено.
Синтез, процессинг и отложение белков семян: пути синтеза и отложения белков в клетке
Абэ С., Ю В. и Дэвис Э. (1991) вплетены в F-актин. Протоплазма , 165 , 139–149.
Перекрёстная ссылка
КАСGoogle ученый
Ахмед, С.У., Бар-Пелед, М. и Райхель, Н.В. (1997) Клонирование и субклеточное расположение Белок, подобный рецептору Arabidopsis , имеет общие черты с рецепторами сортировки белков эукариотических клеток. Физиология растений , 114 , 325–336.
Перекрёстная ссылка
пабмед
КАСGoogle ученый
Альбертс, Б., Брей, Д., Льюис, Дж., Рафф, М., Робертс, К. и Уотсон, Дж. Д. (1994) Молекулярная биология клетки , третье изд.
, Garland Publishing Inc. ., Нью-Йорк.Google ученый
Остин, Б.М. и Вествуд, О.М.Р. (1991) Нацеливание на белок и секреция, издательство Оксфордского университета, Нью-Йорк.
Google ученый
Bagga, S., Adams, H.P., Rodriguez, F.D., Kemp, J.D. and Sengupta-Goplan, C. (1997) Коэкспрессия генов δ-зеина и β-зеина кукурузы приводит к стабильному накоплению δ-зеина в белковых телах эндоплазматического ретикулума, образованных β-зеином. Растительная клетка, 9 , 1683–1696.
ПабМед
КАСGoogle ученый
Бар-Пелед, М. и Райхель, Н.В. (1997) Характеристика AtSEC12 и AtSAR 1, белков, вероятно, участвующих в эндоплазматическом ретикулуме и транспорте Гольджи. Физиология растений, 114 , 315–324.
Перекрёстная ссылка
пабмед
КАСGoogle ученый
Беднарек С.
Ю. и Райхель, Н.В. (1992) Внутриклеточный перенос секреторных белков. Молекулярная биология растений , 20, 133–150.Перекрёстная ссылка
пабмед
КАСGoogle ученый
Бьюли, Дж. Д. и Блэк, М. (1994) Семена. Физиология развития и прорастания , второе изд., Plenum Press, Нью-Йорк.
Google ученый
Блонд-Эльгинди, С., Квирла, С.Э., Дауэр, В.Дж., Липшуц, Р.Дж., Спранг, С.Р., Самбрук, Дж.Ф. и Гетинг, М.-Дж.Х. (1993). Аффинное панорамирование библиотеки пептидов, представленных на бактериофагах, показывает специфичность связывания BiP. Сотовый , 75 , 717–728.
Перекрёстная ссылка
пабмед
КАСGoogle ученый
Боллини, Р., Витале, А. и Криспилс, М. Дж. (1983) In vivo и in vitro процессинг резервного белка семян в эндоплазматическом ретикулуме: свидетельство двух стадий гликозилирования.
Журнал клеточной биологии , 96 , 999–1007.Перекрёстная ссылка
пабмед
КАСGoogle ученый
Boston, R.S., Fontes, E.B.P., Shank, B.B. и Wrobel, R.L. (1991) Повышенная экспрессия гомолога белка b-70, связывающего иммуноглобулин кукурузы, у трех регуляторных мутантов зеина. Растительная клетка , 3 , 497–505.
ПабМед
КАСGoogle ученый
Буллейд, Нью-Джерси и Фридман, Р. Б. (1988) Дефектное котрансляционное образование дисульфидных связей в микросомах с дефицитом протеиндисульфид-изомеразы. Природа , 335 , 649–652.
Перекрёстная ссылка
пабмед
КАСGoogle ученый
Бустос М.М., Калкан Ф.А., Ван ден Бош К.А. и Холл, Т.С. (1991) Дифференциальное накопление четырех гликоформ фазеолина в трансгенном табаке. Молекулярная биология растений , 16 , 381–395.
Перекрёстная ссылка
пабмед
КАСGoogle ученый
Casey, R.D. (1979) Иммуноаффинная хроматография как средство очистки бобовых от семян Pisum (гороха). Биохимический журнал , 177 , 509–520.
ПабМед
КАСGoogle ученый
Chrispeels, MJ (1991) Сортировка белков в секреторной системе. Ежегодный обзор физиологии растений и молекулярной биологии растений , 42 , 21–53.
Перекрёстная ссылка
КАСGoogle ученый
Chrispeels, M.J. and Raikhel, N.V. (1992) Короткие пептидные домены нацеливают белки на растительные вакуоли. Сотовый , 68 , 613–616.
Перекрёстная ссылка
пабмед
КАСGoogle ученый
Криспилс М.Дж. и Таг Б.В. (1990) Транспорт и нацеливание белков в вакуоли для хранения белков (белковые тела) в развивающихся семенах, в Recent Advances in Development and Germination of Seeds , (под редакцией R.
B. Taylorson), Plenum Publ. Корп., Нью-Йорк.Google ученый
Conceicao, A.D.S., Marty-Mazars, Bassham, D.C., Sanderfoot, A.A., Marty, F. and Raikhel, N.V. (1997) Гомолог синтаксин AtPEP12p находится в позднем отделе пост-Гольджи у растений. Растительная клетка , 9 , 571–582.
Google ученый
Крейг С. и Гудчайлд Д.Дж. (1984) Опосредованное Гольджи накопление вициллина в клетках семядолей гороха перенаправляется монензином и нигерицином. Протоплазма , 122 , 91–97.
Перекрёстная ссылка
КАСGoogle ученый
Крауч М.Л., Тенбардж К.М., Саймон А.Е. и Ферл Р. (1983) Клоны кДНК белков семян Brassica napus : данные анализа нуклеотидной последовательности свидетельствуют о том, что обе субъединицы напина отщеплены от полипептида-предшественника. Журнал молекулярной и прикладной генетики , 2 , 273–283.
ПабМед
КАСGoogle ученый
Денеке, Дж., Голдман, М.Х.С., Демолдер, Дж., Сеуринк, Дж. и Боттерман, Дж. (1991) Белок, связывающий просвет табака, кодируется мультигенным семейством. Растительная клетка , 3 , 1025–1035.
ПабМед
КАСGoogle ученый
D’Hondt, K., Bosch, D., Van Damme, J., Goethals, M., Vandekerckhove, J. и Krebbers, E. (1993) Аспарагиновая протеиназа, присутствующая в семенах, расщепляет Arabidopsis 2S предшественники альбумина in vitro . Журнал биологической химии , 268 , 20884–20891.
ПабМед
Google ученый
Д’Ондт, К., Ван Дамм, Дж., Ван Ден Босше, К., Лиджираджумнеун, С., Де Райке, К., Дерксен, Дж., Вандекеркхове, Дж. и Кребберс, Э. ( 1993) Изучение роли пропептидов альбумина Arabidopsis thaliana 2S. Физиология растений , 102 , 425–433.
Перекрёстная ссылка
пабмедGoogle ученый
Дикинсон, К.Д., Флоенер, Л.А., Лилли, Г.Г. и Нильсен, Северная Каролина (1987) Самосборка проглицинина и гибридного проглицинина , синтезированного in vitro из кДНК. Труды Национальной академии наук США , 84 , 5525–5529.
Перекрёстная ссылка
КАСGoogle ученый
Дикинсон, К.Д., Хуссейн, Э.Х.А. и Nielsen, NC (1989) Роль посттрансляционного расщепления в сборке глицинина. Растительная клетка , 1 , 459–469.
ПабМед
КАСGoogle ученый
Дикинсон, К.Д., Скотт, П.М., Хусейн, Э.Х.А., Аргос, П. и Нильсен, Н.К. (1990) Влияние структурных модификаций на сборку субъединицы глицина. Растительная клетка , 2 , 403–413.
ПабМед
КАСGoogle ученый
Домбровски Дж.
Э., Шредер М.Р., Беднарек С.Ю. и Райхель, Н.В. (1993) Определение функциональных элементов в пределах вакуолярного нацеливающего сигнала ячменного лектина. Растительная клетка , 5 , 587–696.ПабМед
КАСGoogle ученый
Дрейк-младший, Р.Р., Каушал, Г.П., Пастушак, И. и Эльбейн, А.Д. (1991) Частичная очистка, фотоаффинная маркировка и свойства УДФ-глюкозы бобов мунг: долихолфосфатглюкозилтрансфераза. Физиология растений , 97 , 396–401.
Перекрёстная ссылка
пабмед
КАСGoogle ученый
Faye, L. and Chrispeels, M.J. (1989) Явное ингибирование секреции β-фруктозидазы туникамицином можно объяснить расщеплением негликозилированного белка во время секреции. Физиология растений , 89 , 845–851.
Перекрёстная ссылка
пабмед
КАСGoogle ученый
Фэй, Л.
, и др. (1992), в Посттрансляционная модификация и ее значение в развитии растений (редакторы Н.М. Бэтти, Х.Г. Дикинсон и А.М. Хизерингтон), Cambridge University Press, стр. 213-242.Google ученый
Фонтес, Э.Б.П., Шэнк, Б.Б., Врубель, Р.Л., Мус, С.П., О’Брайан, Г.Р., Вуртцель, Э.Т. и Бостон, Р.С. (1991) Характеристика гомолога иммуноглобулин-связывающего белка в мутантном эндосперме мучнистого-2 кукурузы. Растительная клетка , 3 , 483–496.
ПабМед
КАСGoogle ученый
Галь С. и Райхель Н.В. (1993) Сортировка белков в эндомембранной системе растительных клеток. Текущие мнения в области клеточной биологии , 5 , 636–640.
Перекрёстная ссылка
КАСGoogle ученый
Галили Г., Альтшулер Ю. и Леванони А. (1993) Сборка и транспорт запасных белков семян.
Тенденции в клеточной биологии , 3 , 437–442.Перекрёстная ссылка
пабмед
КАСGoogle ученый
Гели, М.И., Торрент, М. и Людевид, Д. (1994) Два структурных домена опосредуют два последовательных события в нацеливании γ-зеина: удержание белка в эндоплазматическом ретикулуме и образование белкового тела. Растительная клетка , 6 , 1911–1922 гг.
ПабМед
КАСGoogle ученый
Гетинг, М.-Дж. и Sambrook, J. (1992) Сворачивание белков в клетке. Природа , 355 , 33–45.
Перекрёстная ссылка
пабмед
КАСGoogle ученый
Гилликин Дж.В., Чжан Ф., Коулман С.Е., Басс Х.В., Ларкинс Б.А. и Бостон, Р.С. (1997) Дефектный сигнальный пептид привязывает зеин fluory-2 к мембране эндоплазматического ретикулума. Физиология растений , 114 , 345–352.
Перекрёстная ссылка
пабмед
КАСGoogle ученый
Gomez, L. and Chrispeels, M.J. (1993) Тонопласт и растворимые вакуолярные белки действуют по разным механизмам. Растительная клетка , 5 , 1113–1124.
ПабМед
КАСGoogle ученый
Hara-Nishimura, I., Takeuchi, Y., Inoue, K. and Nishimura, M. (1993) Транспорт везикул и процессинг предшественника 2S-альбумина в тыкве. Заводской журнал , 4 , 793–800.
перекрестная ссылка
Google ученый
Hattori, T., Ichihara, S. и Nakamura, K. (1987) Обработка растительного предшественника вакуолярного белка in vitro . Европейский журнал биохимии , 166 , 533–538.
Перекрёстная ссылка
пабмед
КАСGoogle ученый
Герман Э.
М., Пуэйо Дж.Дж. и Chrispeels, M.J. (1994) Контроль качества запасного белка семян с дестабилизирующим эпитопом происходит в пост-ER секреторной системе. Физиология растений , 105 (Прил.), 94.Google ученый
Hinz, G., Menze, A., Hohl, I. and Vaux, D. (1997) Выделение пролегумина из развивающихся семян гороха: его связывание с эндомембранами и сборка в гексамеры пролегумина в вакуоли для хранения белка. Журнал экспериментальной ботаники , 48 , 139–149.
Перекрёстная ссылка
Google ученый
Хоффман Л.М., Дональдсон Д.Д. и Herman, E.M. (1988) Модифицированный запасной белок синтезируется, обрабатывается и расщепляется в семенах трансгенных растений. Молекулярная биология растений , 11 , 717–729.
Перекрёстная ссылка
КАСGoogle ученый
Höfte, H.
и Chrispeels, MJ (1992) Сортировка белков на вакуолярной мембране. Растительная клетка , 4 , 995–1004.ПабМед
Google ученый
Хох, Б., Хинц, Г., Чон, Б.-К. и Робинсон, Д.Г. (1995) Вакуоли для хранения белков образуются de novo во время развития семядолей гороха. Journal of Cell Science , 108 , 299–310.
ПабМед
КАСGoogle ученый
Hohl, I., Robinson, D.G., Chrispeels, M.J. and Hinz, G. (1996) Транспорт запасных белков в вакуоль опосредован везикулами без клатриновой оболочки. Journal of Cell Science , 109 , 2539–2550.
ПабМед
КАСGoogle ученый
Гольштейн, С.Э.Х., Друкер, М. и Робинсон, Д.Г. (1994) Идентификация адаптина β-типа в везикулах растений, покрытых клатрином. Journal of Cell Science , 107 , 945–953.
ПабМед
КАСGoogle ученый
Holwerda, B.
C., Galvin, N.J., Baranski, T.J. и Роджерс, Дж. К. (1990) In vitro обработка алейраина, вакуолярной тиоловой протеазы ячменя. Растительная клетка , 2 , 1091–1106.ПабМед
КАСGoogle ученый
Хертли, С.М. и Хелениус, А. (1989) Олигомеризация белков в эндоплазматическом ретикулуме. Ежегодный обзор клеточной биологии , 5 , 277–307.
Перекрёстная ссылка
пабмед
КАСGoogle ученый
Kelleher, D.J., Kreibich, G. and Gilmore, R. (1992) Активность олигосахарилтрансферазы связана с белковым комплексом, состоящим из рибофоринов I и II и белка массой 48 кДа. Сотовый , 69 , 55–65.
Перекрёстная ссылка
пабмед
КАСGoogle ученый
Кермоуд, А.Р. (1996) Механизмы внутриклеточного транспорта и нацеливания белков в растительных клетках.
Критические обзоры в области наук о растениях , 15 , 285–423.Перекрёстная ссылка
КАСGoogle ученый
Ким В.Т. и Окита Т.В. (1988)Структура, экспрессия и гетерогенность проламинов семян риса. Физиология растений , 88 , 649–655.
Перекрёстная ссылка
пабмед
КАСGoogle ученый
Ким, В.Ю., Чеонг, Н.Е., Дже, Д.Ю., Ким, М.Г., Лим, К.О., Бахк, Дж.Д., Чо, М.Дж. и Ли, С.Ю. (1997) Наличие семейства генов Sari в Brassica campestris , которое подавляет мутацию дрожжевого везикулярного транспорта Sec 12-1 . Молекулярная биология растений, 33 , 1025–1035.
Перекрёстная ссылка
пабмед
КАСGoogle ученый
Кирш, Т. и Биверс, Л. (1993) Снятие оболочки с клатрин-покрытых везикул путем снятия оболочки с АТФазы из развивающихся горошин.
Физиология растений , 103 , 205–212.Перекрёстная ссылка
пабмед
КАСGoogle ученый
Кирш, Т., Пэрис, Н., Батлер, Дж. М., Биверс, Л. и Роджерс, Дж. К. (1994) Очистка и начальная характеристика потенциального растительного вакуолярного нацеливающего рецептора. Труды Национальной академии наук США , 91 , 3403–3407.
Перекрёстная ссылка
КАСGoogle ученый
Kirsch, T., Saalbach, G., Raikhel, N.V. and Beevers, L. (1996) Взаимодействие потенциального вакуолярного нацеливающего рецептора с амино- и карбоксильными концевыми нацеливающими детерминантами. Физиология растений , 111 , 469–474.
Перекрёстная ссылка
пабмед
КАСGoogle ученый
Кришнан Х.Б., Франчески В.Р. и Окита, Т.В. (1986) Иммунохимические исследования роли комплекса Гольджи в формировании белковых тел в семенах риса.
Планта , 169 , 471–480.Перекрёстная ссылка
КАСGoogle ученый
Крочко, Дж. Э. и Бьюли, Дж. Д. 1988. Использование электрофоретических методов для определения состава запасных белков семян люцерны. Электрофорез
9 , 751–763.Перекрёстная ссылка
пабмед
КАСGoogle ученый
Ларкинс, Б.А. и Хуркман, У. Дж. (1978) Синтез и отложение зеина в белковых телах эндосперма кукурузы. Физиология растений , 62 , 256–263
CrossRef
пабмед
КАСGoogle ученый
Ларкинс Б.А., Педерсен К., Ханда А.К., Хуркман В.Дж. и Смит Л.Д. (1979)Синтез и процессинг запасных белков кукурузы в ооцитах Xenopus laevis . Труды Национальной академии наук США , 76 , 6448–6452.
Перекрёстная ссылка
КАСGoogle ученый
Lending, C.
R. and Larkins, B.A. (1989) Изменения зеинового состава белковых тел во время развития эндосперма кукурузы. Растительная клетка , 1 , 1011–1023.ПабМед
КАСGoogle ученый
Ли, X., Франчески, В.Р. и Окита, Т.В. (1993a) Сегрегация мРНК запасных белков на мембранах шероховатого эндоплазматического ретикулума клеток эндосперма риса. Сотовый , 72 , 869–879.
Перекрёстная ссылка
пабмед
КАСGoogle ученый
Ли, X., Ву, Ю., Чжан, Д.-З., Гилликин, Дж.В., Бостон, Р.С., Франчески, В.Р. и Окита, Т.В. (1993b) Биогенез проламинового белкового тела риса: BiP-опосредованный процесс. Наука , 262 , 1054–1056.
Перекрёстная ссылка
пабмед
КАСGoogle ученый
Lupattelli, F., Pedrazzini, E., Bollini, R., Vitale, A. and Ceriotti, A. (1997) Скорость сборки фазеолина контролируется состоянием глюкозилирования его N-связанных олигосахаридных цепей.
Растительная клетка , 9 , 597–609.ПабМед
КАСGoogle ученый
Matsuoka, K., Bassham, D.C., Raikhel, N.V. and Nakamura, K. (1995) Различная чувствительность к вортманнину двух вакуолярных сигналов сортировки указывает на наличие различных механизмов сортировки в клетках табака. Журнал клеточной биологии , 130 , 1307–1318.
Перекрёстная ссылка
пабмед
КАСGoogle ученый
Мацуока К., Хигучи Т., Маэсима М. и Накамура К. (1997) Н+-АТФаза вакуолярного типа в невакуолярной органелле необходима для сортировки растворимых вакуолярных белков-предшественников в клетках табака. . Растительная клетка , 9 , 533–546.
ПабМед
КАСGoogle ученый
Мацуока, К. и Накамура, К. (1991) Пропептид предшественника растительного вакуолярного белка, необходимого для нацеливания на вакуоли.
Труды Национальной академии наук США , 88 , 834–838.Перекрёстная ссылка
КАСGoogle ученый
Морел, К. (1997) Аквапорины и водопроницаемость растительных мембран. Ежегодный обзор физиологии растений и молекулярной биологии растений , 48 , 399–429.
Перекрёстная ссылка
пабмед
КАСGoogle ученый
Мюнх Д.Г., Ву Ю., Чжан Ю., Ли Х., Бостон, Р.С. и Окита, Т.В. (1997)Молекулярное клонирование, экспрессия и субклеточная локализация гомолога BiP из ткани эндосперма риса. Физиология растений и клеток , 38 , 404–412.
Перекрёстная ссылка
пабмед
КАСGoogle ученый
Манро, С. и Пелхэм, Х.Р.Б. (1986) Hsp70-подобный белок в ER: идентичность с белком, регулируемым глюкозой 78 кДа, и белком, связывающим тяжелую цепь иммуноглобулина. Сотовый , 46 , 291–300.
Перекрёстная ссылка
пабмед
КАСGoogle ученый
Мунц, К. (1996) Протеазы и протеолитическое расщепление запасных белков в развивающихся и прорастающих двудольных семенах. Журнал экспериментальной ботаники , 47 , 605–622.
Перекрёстная ссылка
Google ученый
Nagai, K., Shibata, K. and Yamaguchi, H. (1993) Роль внутримолекулярных цепей с высоким содержанием маннозы в фолдинге и сборке соевых ( Glycine max) лектиновых полипептидов: исследования комбинированного использования спектроскопия и размерный анализ методом гель-фильтрации. Журнал биохимии , 114 , 830–834.
ПабМед
КАСGoogle ученый
Накамура, К. и Мацуока, К. (1993) Нацеливание белков на вакуоли в растительных клетках. Физиология растений , 101 , 1–5.
Перекрёстная ссылка
пабмед
КАСGoogle ученый
Нельсон, Д.
Е., Глаунсингер, Б. и Бонерт, Х. (1997) Обильное накопление связывающего кальций молекулярного шаперона calreticulum в определенных цветочных тканях Arabidopsis thaliana . Физиология растений, 114 , 29–37.Перекрёстная ссылка
пабмед
КАСGoogle ученый
Neuhaus, J.-M., Pietrzak, M. and Boiler, T. (1994) Анализ мутаций С-концевого вакуолярного пептида-мишени хитиназы табака: низкая специфичность системы сортировки и постепенный переход между внутриклеточными задержка и секреция во внеклеточное пространство. Заводской журнал , 5 , 45–54.
Перекрёстная ссылка
пабмед
КАСGoogle ученый
Окита Т.В. и Роджерс, Дж. К. (1996) Компартментация белков в эндомембранной системе растительных клеток. Ежегодный обзор физиологии растений и молекулярной биологии растений , 47 , 327–350.
Перекрёстная ссылка
пабмед
КАСGoogle ученый
Paris, N.
, Stanley, C.M., Jones, R.L. and Rogers, J.C. (1996) Растительные клетки содержат два функционально различных вакуолярных компартмента. Ячейка , 85 , 563–572.Перекрёстная ссылка
пабмед
КАСGoogle ученый
Пэрис, Н., Роджерс, С.В., Цзян, Л., Кирш, Т., Биверс, Л., Филлипс, Т.Е. и Роджерс, Дж. К. (1997) Молекулярное клонирование и дальнейшая характеристика вероятного вакуолярного сортирующего рецептора растений. Физиология растений , 115 , 29–39.
Перекрёстная ссылка
пабмед
КАСGoogle ученый
Пелхэм, Х.Р.Б. (1989) Контроль выхода белка из эндоплазматического ретикулума. Ежегодный обзор клеточной биологии , 5 , 1–23.
Перекрёстная ссылка
пабмед
КАСGoogle ученый
Прайер, Н. К., Вюстехубе, Л. Дж. и Шекман, Р. (1992) Сортировка белков, опосредованная везикулами.
Ежегодный обзор биохимии , 61 , 471–516.Перекрёстная ссылка
пабмед
КАСGoogle ученый
Рапопорт Т.А. (1992) Транспорт белков через мембрану эндоплазматического ретикулума. Наука , 258 , 931–936.
Перекрёстная ссылка
пабмед
КАСGoogle ученый
Робинсон Д.Г. и Hinz, G. (1997) Биогенез вакуолей и транспорт в вакуоли растений: сравнение с вакуолями дрожжей и лизосомами млекопитающих. Протоплазма , 197 , 1–25.
перекрестная ссылка
КАСGoogle ученый
Робинсон Д.Г., Баумер М., Хинц Г. и Холь И. (1998) Везикулярный перенос запасных белков в вакуоль: роль аппарата Гольджи и мультивезикулярных телец. Журнал физиологии растений , 152 , 659–667.
Перекрёстная ссылка
КАСGoogle ученый
Роджерс, Дж.
К. (1998) Компартментация белков растительных клеток в отдельных литических вакуолях и вакуолях для хранения белков. Журнал физиологии растений , 152 , 653–658.Перекрёстная ссылка
КАСGoogle ученый
Rothman, J.E. (1989) GTP и метиониновые щетинки. Природа , 340 , 433–434.
Перекрёстная ссылка
пабмед
КАСGoogle ученый
Saalbach, G., Jung, R., Kunze, G., Saalbach, L, Adler, K. and Muntz, K. (1991) Различные домены белка легумина действуют как вакуолярные нацеливающие сигналы. Растительная клетка , 3 , 695–708.
ПабМед
КАСGoogle ученый
Shewry, P.R. и Tatham, A.S. (1990) Запасные белки проламинов семян злаков: структура и эволюция. Биохимический журнал , 267 , 1–12.
ПабМед
КАСGoogle ученый
Шимада Т.
, Хираива Н., Нисимура М. и Хара-Нисимура И. (1994) Фермент вакуолярного процессинга, превращающий пропротеины в соответствующие зрелые формы. Физиология растений и клеток , 35 , 713–718.ПабМед
КАСGoogle ученый
Саймон, П. и Джонс, Р.Л. (1988) Синтез и секреция каталитически активных изоформ α-амилазы ячменя ооцитами Xenopus , инъецированными мРНК ячменя. Европейский журнал клеточной биологии , 47 , 213–221.
КАС
Google ученый
Сингер, Р. Х., Ланжевин, Г. Л. и Лоуренс, Дж. Б. (1989) Ультраструктурная визуализация мРНК цитоскелета и связанных с ними белков с использованием двойной метки гибридизации in situ. Журнал клеточной биологии , 108 , 2343–2353.
Перекрёстная ссылка
пабмед
КАСGoogle ученый
Штурм А., Джонсон К.
Д., Шумило Т., Эльбейн А.Д. и Криспилс М.Дж. (1987) Субклеточная локализация гликозидаз и гликозилтрансфераз, участвующих в процессинге N-связанных олигосахаридов. Физиология растений , 85 , 741–745.Перекрёстная ссылка
пабмед
КАСGoogle ученый
Сан, С.С.М., Альтенбах, С.Б. и Leung, F.W. (1987) Свойства, биосинтез и переработка богатого серой белка в бразильском орехе (Bertholletia excelsa HBK). Европейский журнал биохимии , 162 , 477–483.
Перекрёстная ссылка
пабмед
КАСGoogle ученый
Taylor, C. (1997) Контроль качества секреторных белков в растениях. Растительная клетка , 9 , 1697–1699.
КАС
Google ученый
Taylor, J.R.N., Schussler, L. and Liebonberg, N.V.D.W. (1985) Формирование белковых тел в крахмалистом эндосперме развивающихся Sorghum bicolor (L) Семена Moench.
Южноафриканский журнал ботаники , 51 , 35–40.Google ученый
Вернер К. и Шац Г. (1988) Транслокация белков через мембраны. Наука
241 , 1307–1313.Перекрёстная ссылка
пабмед
КАСGoogle ученый
Витале, А., Чериоти, А. и Денеке, Дж. (1993) Роль эндоплазматического ретикулума в синтезе, модификации и внутриклеточном транспорте белков. Журнал экспериментальной ботаники , 44 , 1417–1444.
Перекрёстная ссылка
КАСGoogle ученый
Von Schaewen, A. and Chrispeels, MJ (1993) Идентификация информации о сортировке вакуолей в фитогемагглютинине, необработанном вакуолярном белке. Журнал экспериментальной ботаники , 44 (Прил.), 339–342.
Google ученый
Ся, Ж.
-Х. и Кермоуд, А.Р. (1998) Анализы для определения роли незрелости зародыша в поддержании покоя желтого кедра ( Chamaecyparis nootkatensis) семян: Синтез и накопление запасных белков и белков, участвующих в устойчивости к высыханию. Журнал экспериментальной ботаники , в печати.Google ученый
 , Garland Publishing Inc. ., Нью-Йорк.
, Garland Publishing Inc. ., Нью-Йорк.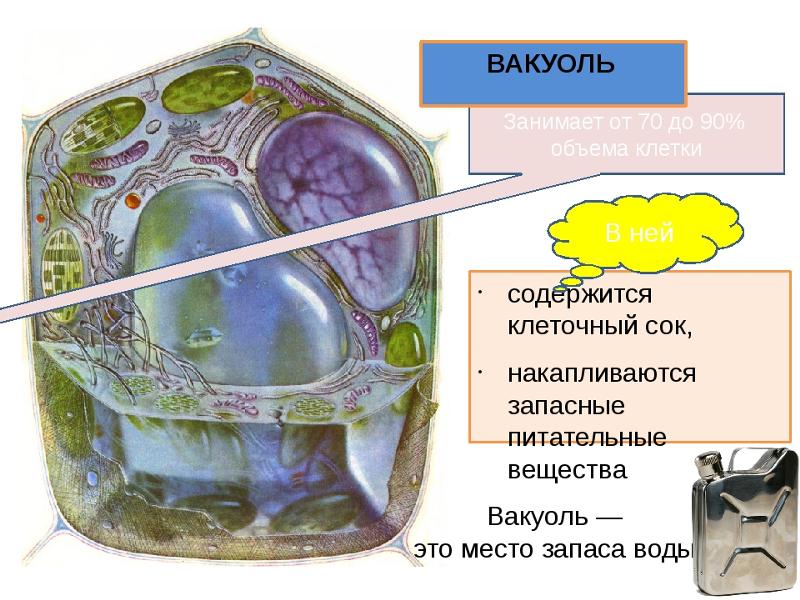 Ю. и Райхель, Н.В. (1992) Внутриклеточный перенос секреторных белков. Молекулярная биология растений , 20, 133–150.
Ю. и Райхель, Н.В. (1992) Внутриклеточный перенос секреторных белков. Молекулярная биология растений , 20, 133–150. Журнал клеточной биологии , 96 , 999–1007.
Журнал клеточной биологии , 96 , 999–1007.
 B. Taylorson), Plenum Publ. Корп., Нью-Йорк.
B. Taylorson), Plenum Publ. Корп., Нью-Йорк.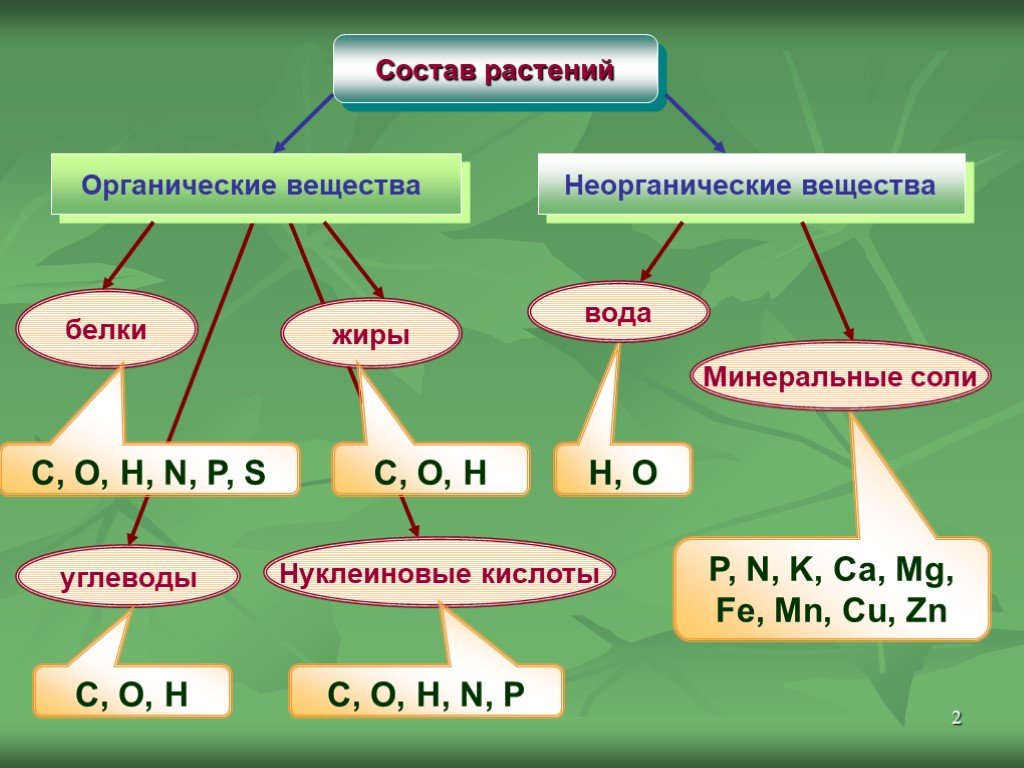
 Э., Шредер М.Р., Беднарек С.Ю. и Райхель, Н.В. (1993) Определение функциональных элементов в пределах вакуолярного нацеливающего сигнала ячменного лектина. Растительная клетка , 5 , 587–696.
Э., Шредер М.Р., Беднарек С.Ю. и Райхель, Н.В. (1993) Определение функциональных элементов в пределах вакуолярного нацеливающего сигнала ячменного лектина. Растительная клетка , 5 , 587–696.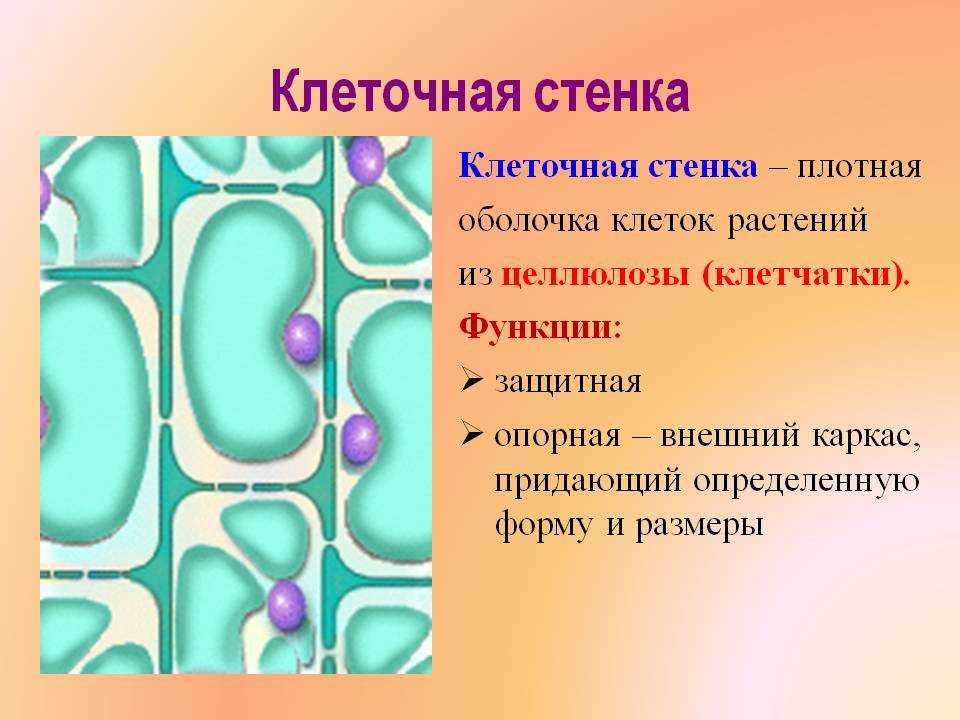 , и др. (1992), в Посттрансляционная модификация и ее значение в развитии растений (редакторы Н.М. Бэтти, Х.Г. Дикинсон и А.М. Хизерингтон), Cambridge University Press, стр. 213-242.
, и др. (1992), в Посттрансляционная модификация и ее значение в развитии растений (редакторы Н.М. Бэтти, Х.Г. Дикинсон и А.М. Хизерингтон), Cambridge University Press, стр. 213-242. Тенденции в клеточной биологии , 3 , 437–442.
Тенденции в клеточной биологии , 3 , 437–442.
 М., Пуэйо Дж.Дж. и Chrispeels, M.J. (1994) Контроль качества запасного белка семян с дестабилизирующим эпитопом происходит в пост-ER секреторной системе. Физиология растений , 105 (Прил.), 94.
М., Пуэйо Дж.Дж. и Chrispeels, M.J. (1994) Контроль качества запасного белка семян с дестабилизирующим эпитопом происходит в пост-ER секреторной системе. Физиология растений , 105 (Прил.), 94.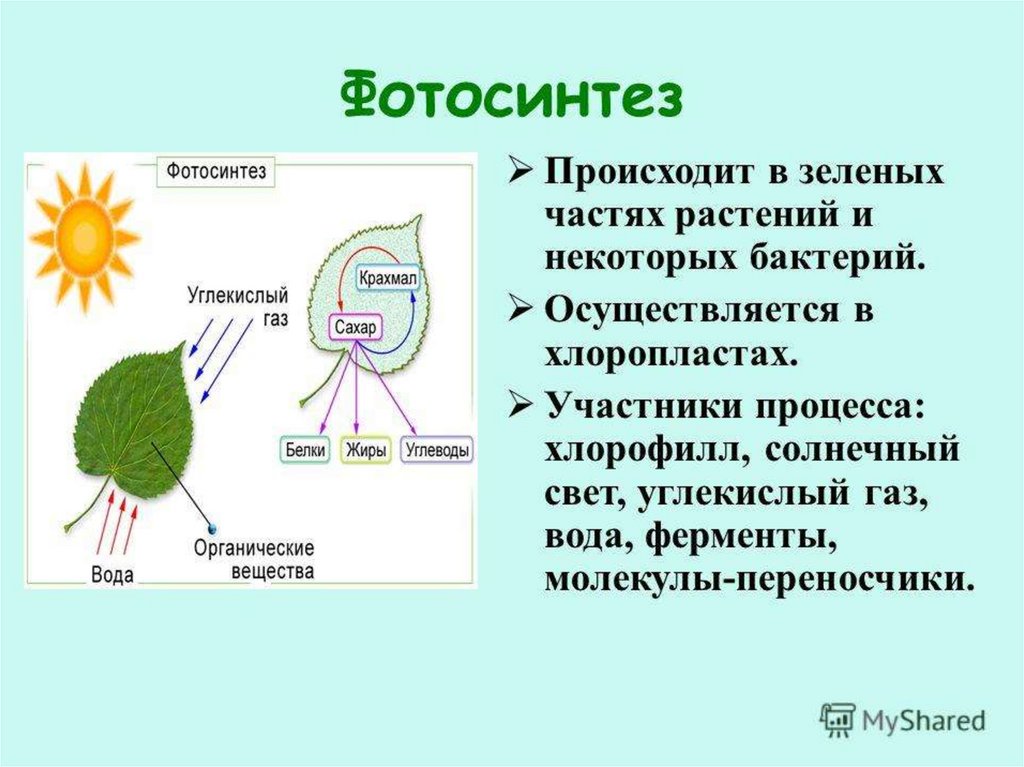 и Chrispeels, MJ (1992) Сортировка белков на вакуолярной мембране. Растительная клетка , 4 , 995–1004.
и Chrispeels, MJ (1992) Сортировка белков на вакуолярной мембране. Растительная клетка , 4 , 995–1004. C., Galvin, N.J., Baranski, T.J. и Роджерс, Дж. К. (1990) In vitro обработка алейраина, вакуолярной тиоловой протеазы ячменя. Растительная клетка , 2 , 1091–1106.
C., Galvin, N.J., Baranski, T.J. и Роджерс, Дж. К. (1990) In vitro обработка алейраина, вакуолярной тиоловой протеазы ячменя. Растительная клетка , 2 , 1091–1106. Критические обзоры в области наук о растениях , 15 , 285–423.
Критические обзоры в области наук о растениях , 15 , 285–423. Физиология растений , 103 , 205–212.
Физиология растений , 103 , 205–212. Планта , 169 , 471–480.
Планта , 169 , 471–480. R. and Larkins, B.A. (1989) Изменения зеинового состава белковых тел во время развития эндосперма кукурузы. Растительная клетка , 1 , 1011–1023.
R. and Larkins, B.A. (1989) Изменения зеинового состава белковых тел во время развития эндосперма кукурузы. Растительная клетка , 1 , 1011–1023. Растительная клетка , 9 , 597–609.
Растительная клетка , 9 , 597–609. Труды Национальной академии наук США , 88 , 834–838.
Труды Национальной академии наук США , 88 , 834–838.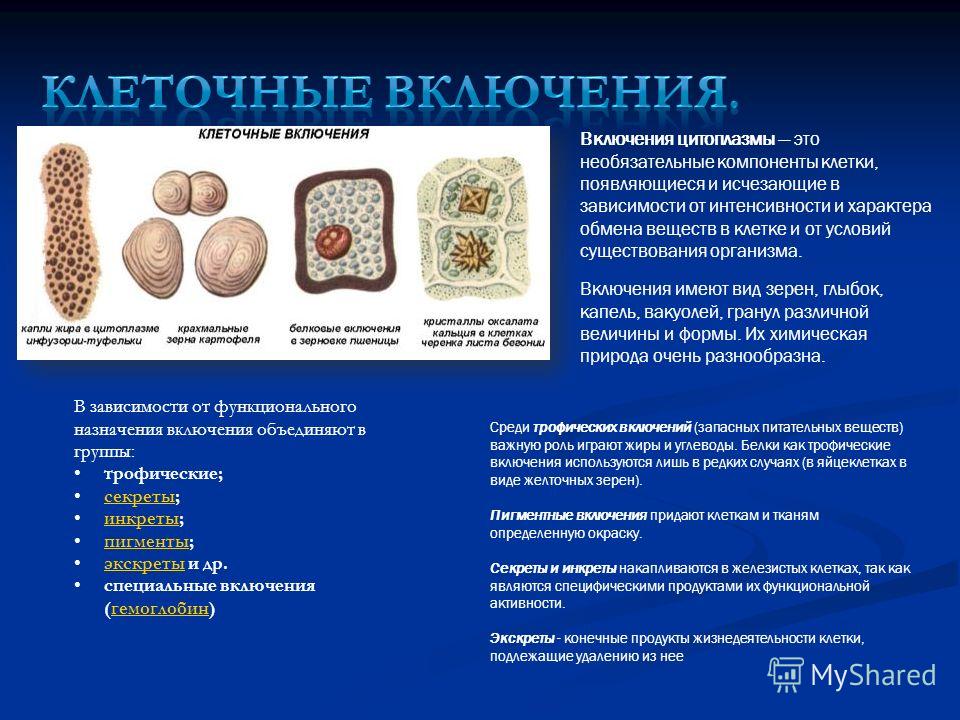
 Е., Глаунсингер, Б. и Бонерт, Х. (1997) Обильное накопление связывающего кальций молекулярного шаперона calreticulum в определенных цветочных тканях Arabidopsis thaliana . Физиология растений, 114 , 29–37.
Е., Глаунсингер, Б. и Бонерт, Х. (1997) Обильное накопление связывающего кальций молекулярного шаперона calreticulum в определенных цветочных тканях Arabidopsis thaliana . Физиология растений, 114 , 29–37.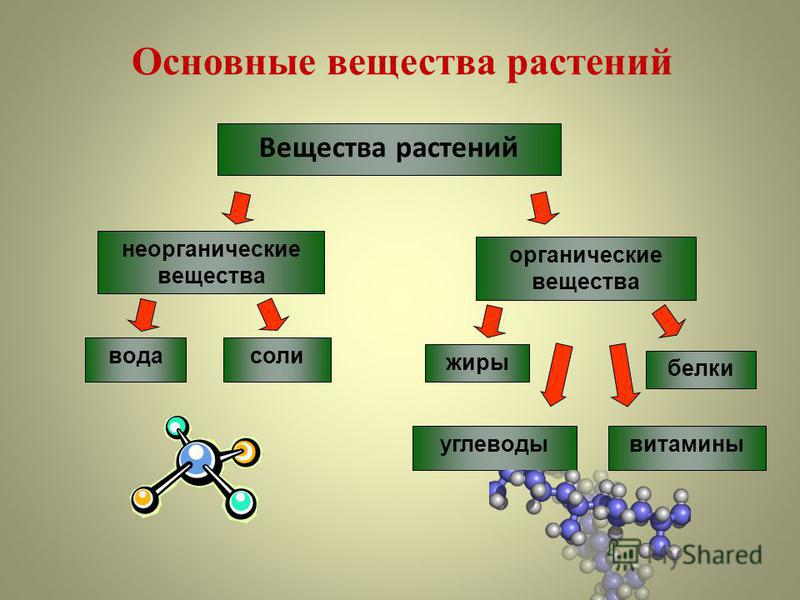 , Stanley, C.M., Jones, R.L. and Rogers, J.C. (1996) Растительные клетки содержат два функционально различных вакуолярных компартмента. Ячейка , 85 , 563–572.
, Stanley, C.M., Jones, R.L. and Rogers, J.C. (1996) Растительные клетки содержат два функционально различных вакуолярных компартмента. Ячейка , 85 , 563–572. Ежегодный обзор биохимии , 61 , 471–516.
Ежегодный обзор биохимии , 61 , 471–516. К. (1998) Компартментация белков растительных клеток в отдельных литических вакуолях и вакуолях для хранения белков. Журнал физиологии растений , 152 , 653–658.
К. (1998) Компартментация белков растительных клеток в отдельных литических вакуолях и вакуолях для хранения белков. Журнал физиологии растений , 152 , 653–658.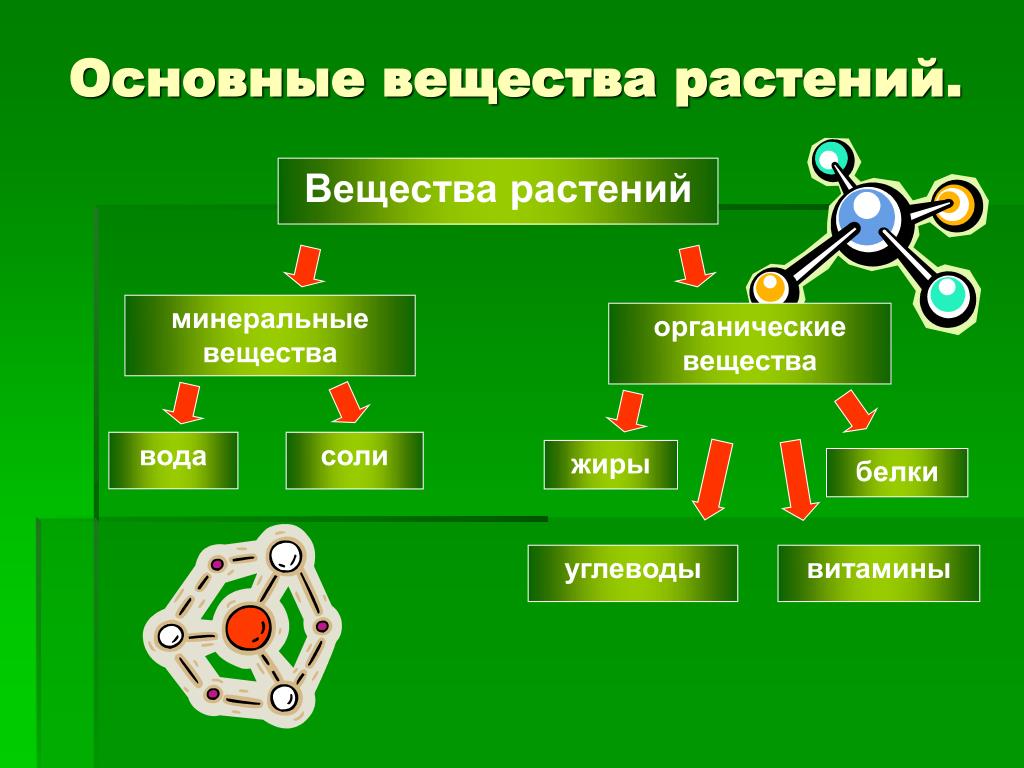 , Хираива Н., Нисимура М. и Хара-Нисимура И. (1994) Фермент вакуолярного процессинга, превращающий пропротеины в соответствующие зрелые формы. Физиология растений и клеток , 35 , 713–718.
, Хираива Н., Нисимура М. и Хара-Нисимура И. (1994) Фермент вакуолярного процессинга, превращающий пропротеины в соответствующие зрелые формы. Физиология растений и клеток , 35 , 713–718. Д., Шумило Т., Эльбейн А.Д. и Криспилс М.Дж. (1987) Субклеточная локализация гликозидаз и гликозилтрансфераз, участвующих в процессинге N-связанных олигосахаридов. Физиология растений , 85 , 741–745.
Д., Шумило Т., Эльбейн А.Д. и Криспилс М.Дж. (1987) Субклеточная локализация гликозидаз и гликозилтрансфераз, участвующих в процессинге N-связанных олигосахаридов. Физиология растений , 85 , 741–745.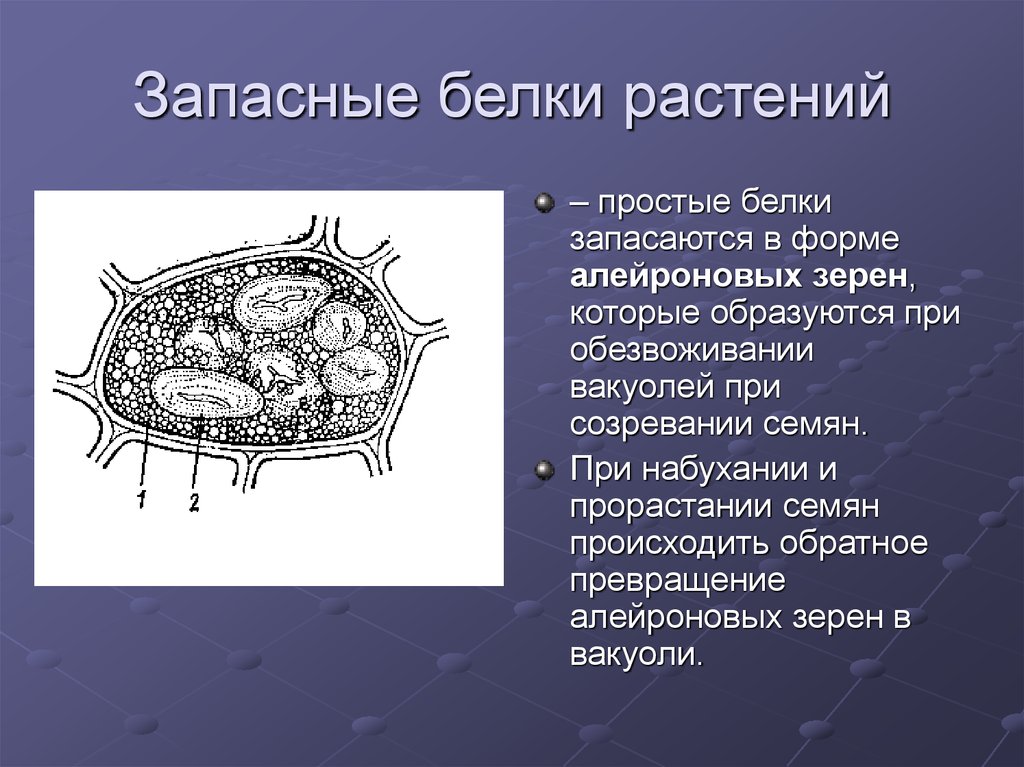 Южноафриканский журнал ботаники , 51 , 35–40.
Южноафриканский журнал ботаники , 51 , 35–40.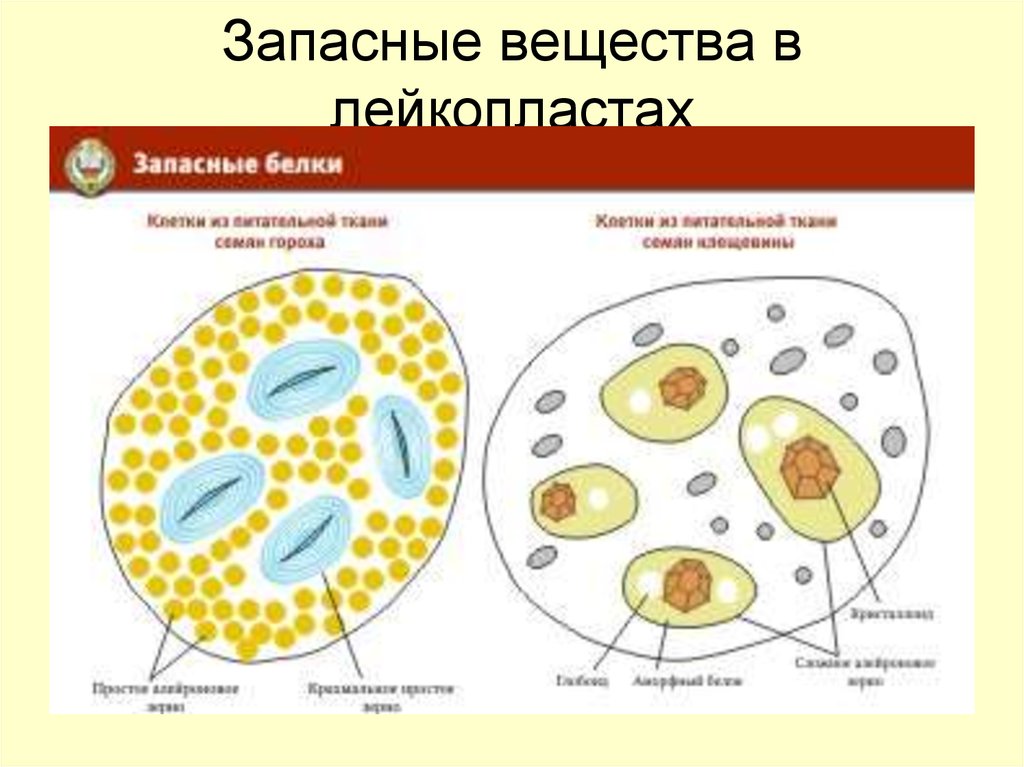 -Х. и Кермоуд, А.Р. (1998) Анализы для определения роли незрелости зародыша в поддержании покоя желтого кедра ( Chamaecyparis nootkatensis) семян: Синтез и накопление запасных белков и белков, участвующих в устойчивости к высыханию. Журнал экспериментальной ботаники , в печати.
-Х. и Кермоуд, А.Р. (1998) Анализы для определения роли незрелости зародыша в поддержании покоя желтого кедра ( Chamaecyparis nootkatensis) семян: Синтез и накопление запасных белков и белков, участвующих в устойчивости к высыханию. Журнал экспериментальной ботаники , в печати.Ссылки для скачивания
Plantae | Разрушение шаблона: снижение запасов белка в семенах Brassica napus вызывает неожиданные структурные изменения
сильно различаются между типами семян. Из-за очевидных сельскохозяйственных последствий исследователи долго искали способы манипулировать разделением ресурсов, импортируемых из материнского растения, чтобы увеличить урожай или качество желаемых продуктов хранения. Метаболическая инженерия и усилия по селекции обычно обнаруживают, что когда дело доходит до продуктов для хранения, вы не можете иметь все это. В масличных семенах запасные белки и липиды накапливаются в одних и тех же тканях (Borisjuk et al.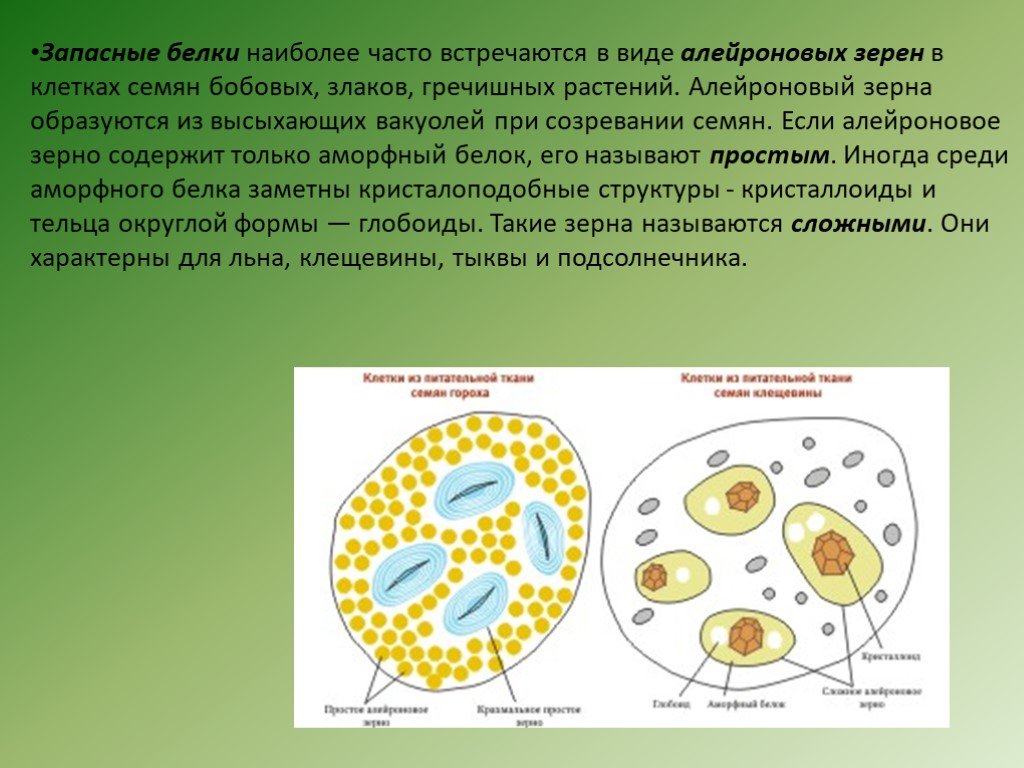 , 2013) и, по-видимому, конкурируют за одни и те же углеродные ресурсы, что приводит к взаимной зависимости между содержанием масла и белка (Weselake et al., 2009).).
, 2013) и, по-видимому, конкурируют за одни и те же углеродные ресурсы, что приводит к взаимной зависимости между содержанием масла и белка (Weselake et al., 2009).).
В новом исследовании Rolletschek et al. (2020) создали и охарактеризовали линий Brassica napus РНКи со значительно сниженной экспрессией и накоплением круциферина и напина, двух основных запасных белков масличных культур. Этот подход был направлен на снижение производства запасных белков семян и, таким образом, перенаправление большего количества углерода в триацилглицериды, основной сельскохозяйственный продукт B. napus (рапс/рапс). Однако полученные трансгенные семена не соответствовали ожидаемой плесени, потому что уменьшение содержания запасного белка сопровождалось аналогичным уменьшением содержания запасных липидов, что приводило к семенам с меньшими зародышами. Кроме того, наблюдались поразительные различия в субклеточной организации трансгенных эмбрионов.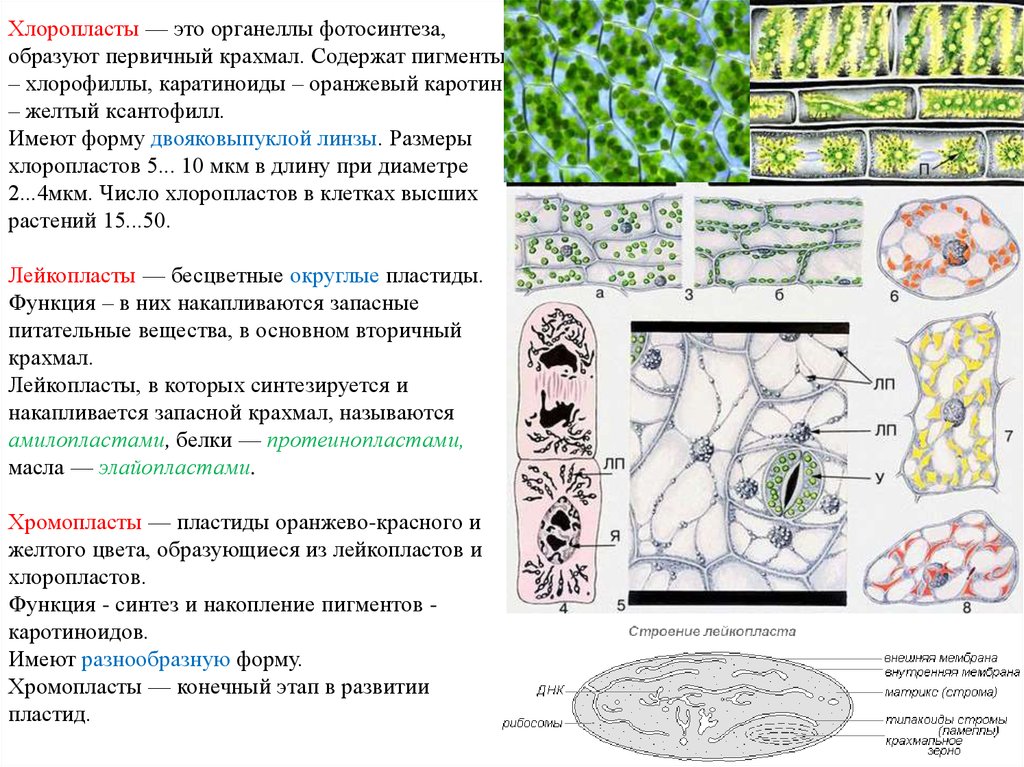 Как правило, запасной белок накапливается в виде белковых тел в вакуолях, тогда как липиды накапливаются в основном в виде триацилглицеридов, упакованных в цитозольные липидные капли. Как и ожидалось, трансгенные эмбрионы содержали гораздо меньшие белковые тела на протяжении всего развития, что согласуется со сниженной экспрессией запасных белков (рис.). Однако трансгенные линии также демонстрировали более мелкие липидные капли по сравнению с WT и заметное накопление необычных уплощенных мембранных стопок, напоминающих ER, которых не было в семенах WT (рисунок). Как ключ к их происхождению, эти стопки мембран содержали большое количество олеозина, белка, обычно связанного с образованием липидных капель.
Как правило, запасной белок накапливается в виде белковых тел в вакуолях, тогда как липиды накапливаются в основном в виде триацилглицеридов, упакованных в цитозольные липидные капли. Как и ожидалось, трансгенные эмбрионы содержали гораздо меньшие белковые тела на протяжении всего развития, что согласуется со сниженной экспрессией запасных белков (рис.). Однако трансгенные линии также демонстрировали более мелкие липидные капли по сравнению с WT и заметное накопление необычных уплощенных мембранных стопок, напоминающих ER, которых не было в семенах WT (рисунок). Как ключ к их происхождению, эти стопки мембран содержали большое количество олеозина, белка, обычно связанного с образованием липидных капель.
Поскольку природа метаболических и структурных изменений в трансгенных семенах не была очевидной, авторы провели всестороннее метаболомное, липидомное, транскриптомное и протеомное сравнение дикого типа и трансгенных семян. Данные об уровне метаболитов и транскриптов выявили накопление свободных аминокислот и подавление ассимиляции серы в трансгенных семенах, что согласуется со снижением синтеза запасного белка. Протеомный анализ выявил более высокие уровни олеозина и нескольких ферментов в пути синтеза пластидных жирных кислот, что свидетельствует о повышенной продукции липидов. Однако липидомный анализ показал, что общее содержание липидов было на 20% ниже в трансгенных семенах, наряду со значительными изменениями в составе липидов, включая более высокие уровни фосфатидилхолина и более высокую степень ненасыщенности липидов в целом. Эти наборы данных, по-видимому, частично согласуются с первоначальными ожиданиями снижения синтеза запасных белков и повышения синтеза липидов; но в конечном итоге трансгенные семена продемонстрировали загадочный дефицит накопления триглицеридов.
Протеомный анализ выявил более высокие уровни олеозина и нескольких ферментов в пути синтеза пластидных жирных кислот, что свидетельствует о повышенной продукции липидов. Однако липидомный анализ показал, что общее содержание липидов было на 20% ниже в трансгенных семенах, наряду со значительными изменениями в составе липидов, включая более высокие уровни фосфатидилхолина и более высокую степень ненасыщенности липидов в целом. Эти наборы данных, по-видимому, частично согласуются с первоначальными ожиданиями снижения синтеза запасных белков и повышения синтеза липидов; но в конечном итоге трансгенные семена продемонстрировали загадочный дефицит накопления триглицеридов.
Так почему же не накапливались триацилглицериды? Авторы отмечают несколько возможностей, включая обратную зависимость между уровнями олеозина и размером липидных капель, наблюдаемую в некоторых тканях семян (Siloto et al., 2006). Более высокое содержание олеозина могло действовать на увеличение отношения площади поверхности к объему липидных тел, что делало их меньше, а также могло вызывать образование необычных уплощенных мембранных стопок. Авторы предполагают, что такие структуры, хотя и многочисленны, сохраняют аналогичный общий объем по сравнению с липидными каплями у эмбрионов WT и потенциально способствуют неэффективному заполнению триглицеридами. Будущие исследования должны определить, может ли оптимизированная экспрессия олеозина способствовать, наряду с другими механизмами, метаболической инженерии повышенного накопления триглицеридов в семенах масличных культур.
Авторы предполагают, что такие структуры, хотя и многочисленны, сохраняют аналогичный общий объем по сравнению с липидными каплями у эмбрионов WT и потенциально способствуют неэффективному заполнению триглицеридами. Будущие исследования должны определить, может ли оптимизированная экспрессия олеозина способствовать, наряду с другими механизмами, метаболической инженерии повышенного накопления триглицеридов в семенах масличных культур.
Brendan M O’Leary
Центр передового опыта в энергетической биологии растений
Университет Западной Австралии
9915 . или orcid: 000010015151515151515151515. -8770-155X
ССЫЛКИ
Борисюк Л., Нойбергер Т., Швендер Дж., Хайнцель Н., Сандерхаус Дж. , Чирш Х., Браун Х. П., Денольф П., Ламберт Б., Якоб П. М. и Роллетчек Х. (2013). Структура семян формирует метаболизм зародыша масличного рапса. Растительная клетка 25: 1625-1640.
Структура семян формирует метаболизм зародыша масличного рапса. Растительная клетка 25: 1625-1640.
Роллетчек, Х., Швендер, Дж., Кениг, К., Чепмен, К.Д., Ромсдал, Т., Лоренц, К., Браун, Х.-П., Денольф, П., Ауденхове, К.В., Мунц Э., Хайнцель Н., Ортлеб С., Руттен Т., МакКоркл С., Борисюк Т., Гендель А., Ши Х., Аувермеулен М.В., Буро С., Борисюк , Л. (2020). Клеточная пластичность в ответ на подавление запасных белков у Brassica napus эмбрионов. Plant Cell https://doi.org/10.1105/tpc.19.00879
Силото, Р.М., Финдли, К., Лопес-Вильялобос, А., Юнг, Э.К., Никифорук, К.Л., и Молони, М.М. (2006 г.). Накопление олеозинов определяет размер семенных масличных телец арабидопсиса. Растительная ячейка 18: 1961-1974.
Веселаке, Р.Дж., Тейлор, Д.К., Рахман, М.Х., Шах, С., Ларош, А., МакВетти, П.Б.Е., и Харвуд, Дж.Л. (2009). Увеличение поступления углерода в масло семян. Биотехнолог.