Содержание
Высшие споровые и семенные растения
Похожие презентации:
Эндокринная система
Анатомо — физиологические особенности сердечно — сосудистой системы детей
Хронический панкреатит
Топографическая анатомия верхних конечностей
Анатомия и физиология сердца
Мышцы головы и шеи
Эхинококкоз человека
Черепно-мозговые нервы
Анатомия и физиология печени
Топографическая анатомия и оперативная хирургия таза и промежности
1. Появление высших споровых и семенных растений
Выполнила:
Белявская Евгения
Ученица 5 – Б класса
ПСШ 6 им.Е.Ищенко
Учитель биологии
Чигрина Надежда
Александровна
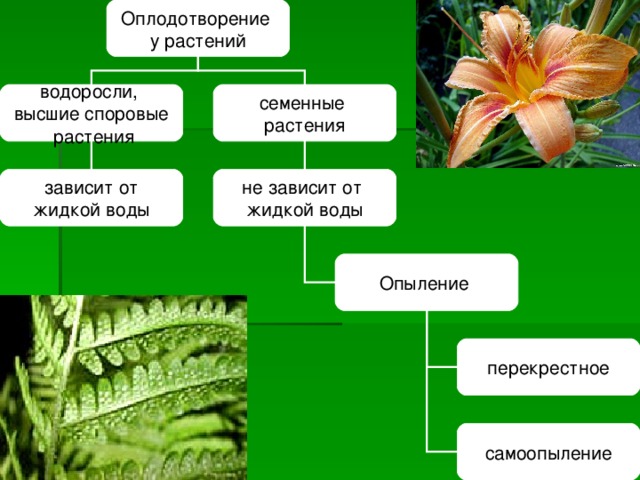
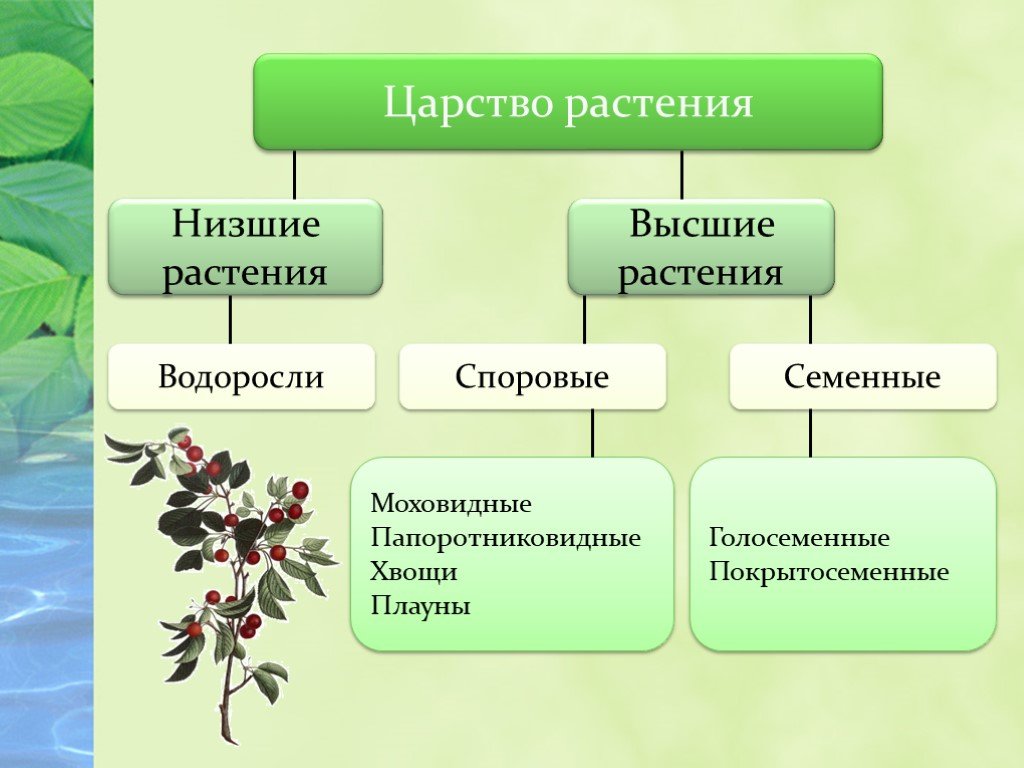
2. Высшие растения
Имеют хорошо выраженные ткани:
Образовательную
Покровную
Проводящие
Механические
Выделительные
Основные
Запасающие
3. Ткани высших растений образуют органы
органы
вегетативные
корень
стебель
генеративные
лист
спорангии
цветки
плод
4.
 Индивидуальное развитие
Индивидуальное развитие
период
эмбриональный
постэмбриональный
5. Высшие растения
Споровые
Семенные
Споровые растения
7. Моховидные
8. Плауновидные
9. Хвощевидные
10. Папоротниковидные
11. Отличительная черта споровых:
Менее четкая по сравнению с семенными
специализация тканей
Бесполое размножение при помощи спор
Половое размножение связано с водной средой
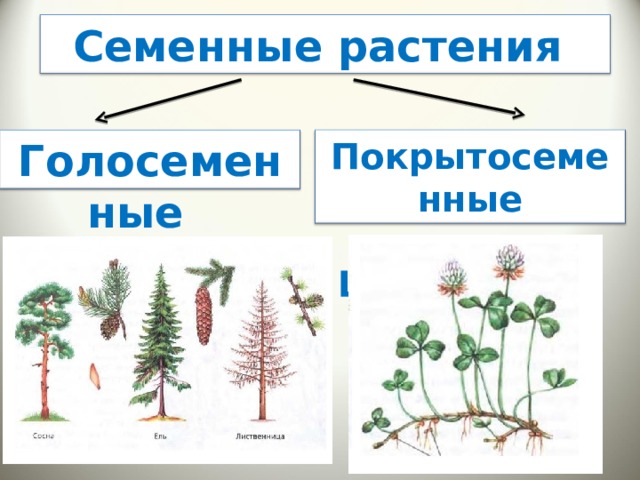
Семенные растения
13. Голосеменные растения
14. Покрытосеменные растения
15. Особенности семенных растений:
Размножаются при помощи семян и
вегетативно(частями тела)
Отчетливо выраженные системы органовпобеговая и корневая
16. Отдел Моховидные
17. Моховидные произошли 350 млн. лет назад от псилофитов.
18. Моховидные
Многолетние растения
Размеры: от
миллиметра до
нескольких
сантиметров
19. Строение мха. Кукушкин лен.
20. Автотрофы
21. Сфагновые мхи
Распространены от гор
тропиков и
субантарктической
зон, но особенно
широко представлены
в умеренной зоне
Северного полушария
22.
 Строение сфагнума
Строение сфагнума
Спорангии
Верхушечные листья
Торчащие листья
Свисающие листья
Стебель
23. Сфагнум
Растут во влажных
местах, обычно
находятся в воде,
поэтому у них нет
ризоидов.
Сфагнум может
выпить воды в 20-25
раз больше
собственной массы.
24. Внутреннее строение листа сфагнума (стр. 55)
Хлорофиллоносные
клетки
Водоносные клетки
Функции:
Хлорофиллоносные
клетки- фотосинтез;
Водоносные- запас
воды
25. Значение сфагновых мхов:
Вызывают
заболачивание
Отмирающие части
растения формируют
торф
26. Болота и леса, где произрастают мхи
Служат
накопителями влаги
и влияют на водный
режим соседних
территорий.
English
Русский
Правила
«Семенные и споровые растения» | План-конспект урока по биологии (7 класс):
Тема урока. Семенные и споровые растения
Задачи:
- Показать сходство и различия между споровыми и семенными растениями;
- Развивать практические умения работать с ручной лупой, зарисовывать изучаемый объект и обозначать органы;
- Формировать ответственное отношение к различным растениям.


Средства обучения.
«Органы цветкового растения», «Папоротники», живые комнатные растения, гербарии папоротников и цветковых растений в цветущем состоянии, спороносные листья папоротника, лупа и препаровальная игла (на каждого ученика или одна на парту).
Планируемые результаты обучения:
Предметные:
- Формирование знаний о споровых и семенных растениях о функциях споры и семени и о их различиях;
- Соблюдение правил поведения в кабинете биологии.
Метапредметные:
- Развитие умения сравнивать и анализировать растительные обьекты, находить черты их сходства и различия, делать выводы.
Личностные:
- Формирование ответственного отношения к учению, развитию устойчивых, познавательных интересов в области ботаники;
- Формирование коммуникативной компетентности в общении и сотрудничестве со сверстниками в процессе образовательной деятельности.
Общие рекомендации.
Перед началом работы необходимо напомнить учащимся о технике безопасности, так как работа связана с потенциально опасными предметами
Ход урока I. Актуализация знаний – Ответьте на вопросы. 1. В чем основное отличие высших растений от низших? 2. Какие растения относят к высшим, а какие – к низшим? 3. Что такое вегетативные и генеративные органы растения? 4. Каковы их основные функции?
Актуализация знаний – Ответьте на вопросы. 1. В чем основное отличие высших растений от низших? 2. Какие растения относят к высшим, а какие – к низшим? 3. Что такое вегетативные и генеративные органы растения? 4. Каковы их основные функции?
II. Практическая работа 1. СТРОЕНИЕ ЦВЕТКОВОГО РАСТЕНИЯ
Цель: изучить внешнее строение цветкового растения.
Оборудование: гербарий цветковых растений в цветущем состоянии, по возможности с плодами (на каждого ученика или один на парту), лупа (на каждого ученика или одна на парту), препаровальная игла (на каждого ученика или одна на парту), линейка (на каждого ученика)
Общее знакомство с растениями один на парту), лупа (на каждого ученика или одна на парту), препаровальная игла (на каждого ученика или одна на парту), линейка (на каждого ученика).
Общие рекомендации. Гербарии лучше приготовить заранее в большем, чем это необходимо, количестве. Лучше всего для этой работы подходят пастушья сумка, сурепка, так как у этих растений на одном стебле одновременно можно увидеть и плоды, и семена. Собрать и засушить необходимое количество этих растений не составляет труда.
Собрать и засушить необходимое количество этих растений не составляет труда.
Ход работы
1. Рассмотрите экземпляр растения на вашем столе. Найдите у него вегетативные органы. Какие из вегетативных органов вы видите? (Корень, стебель, листья, на некоторых видны почки.)
2. Определите цвет и размер корня, цвет и длину стебля, цвет, размер и примерное количество листьев.
3. Найдите генеративные органы растения. Какие из генеративных органов вы видите? (Цветы, плоды).
4. Определите размер и окраску (по возможности) цветов и плодов. Аккуратно при помощи препаровальной иглы вскройте плод и найдите там семена. Определите размер семян данного растения.
5. Зарисуйте растение в тетрадь для практических работ, укажите все органы, которые вам удалось увидеть. Не забудьте указать название исследуемого растения.
6. Заполните таблицу.
7. Сделайте вывод о том, что это растение относится к высшим цветковым растениям, объясните почему
III. Закрепление знаний об особенностях споровых и семенных растений,их строение и размножении. Обобщающая беседа
Обобщающая беседа
Домашнне задание: изучить рисунки и текст п.4 учебника, выполнить задания в конце параграфа
7.1: Введение в семенные растения
- Последнее обновление
- Сохранить как PDF
- Идентификатор страницы
- 47643
Цели обучения
- Объяснить, как гетероспория привела к эволюции семян.
- Соедините r- и k-стратегии с пропагулами, производимыми растениями (например, со спорами, гаметами, пыльцой и семенами).
- Перечислите несколько различий в жизненном цикле голосеменных и покрытосеменных растений.
Окаменелости девонского периода показывают папоротниковидные растения, которые были гетероспористыми ; то есть произвел два вида спор: микроспоры и мегаспоры . Мегаспоры не отделялись от родительского спорофита. Оплодотворение происходило внутри ткани родительского спорофита, что освобождало его от зависимости от поверхностных вод. Однако необходимость переноса микроспор с одного растения на другое для достижения мегагаметофита лишала их ценности как агентов распространения. Эту функцию взял на себя семян — покоящихся, защищенных, зачаточных спорофитов.
Мегаспоры не отделялись от родительского спорофита. Оплодотворение происходило внутри ткани родительского спорофита, что освобождало его от зависимости от поверхностных вод. Однако необходимость переноса микроспор с одного растения на другое для достижения мегагаметофита лишала их ценности как агентов распространения. Эту функцию взял на себя семян — покоящихся, защищенных, зачаточных спорофитов.
Гетероспория: следующий большой шаг к жизни на суше
Позвоночные животные стали полностью наземными только тогда, когда их оплодотворение стало полностью независимым от воды. Растения начали выполнять подобные «эволюционные усилия» еще раньше, но пока рептилии активно приближаются к половому партнеру, у растений выработались другие решения. Вместо активного секса растения используют «ковровую бомбардировку» спорами; это увеличило вероятность того, что две споры приземлятся рядом, а расстояние между сперматозоидом и яйцеклеткой будет минимальным.
Однако увеличение количества спор также приводит к пустой трате ресурсов, поэтому растения минимизировали размер спор; это также позволит увеличить расстояние распространения и уменьшить количество ресурсов, вложенных в каждую спору.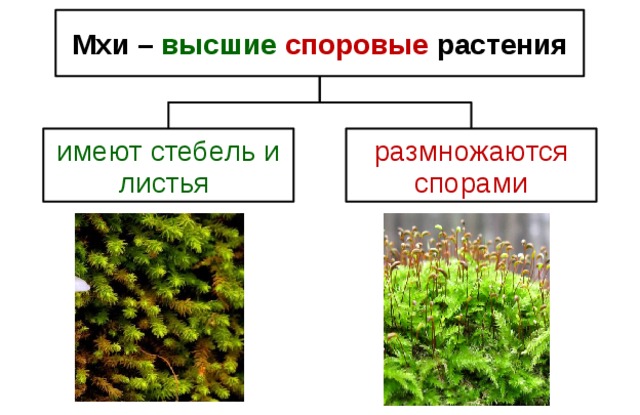 С другой стороны, некоторые споры должны оставаться большими, потому что зародыш (если произойдет оплодотворение) будет нуждаться в поддержке питающего гаметофита. Следовательно, растения оказались с разделением труда: многочисленные крохотные микроспоры вырастают в микрогаметофиты только с антеридиями, а несколько крупных мегаспор превращаются в мегагаметофиты, производящие только архегонии (рис. \(\PageIndex{1}\)). Мегаспоры крупнее, потому что они богаты питательными веществами, готовы к оплодотворению и питанию развивающегося спорофита.
С другой стороны, некоторые споры должны оставаться большими, потому что зародыш (если произойдет оплодотворение) будет нуждаться в поддержке питающего гаметофита. Следовательно, растения оказались с разделением труда: многочисленные крохотные микроспоры вырастают в микрогаметофиты только с антеридиями, а несколько крупных мегаспор превращаются в мегагаметофиты, производящие только архегонии (рис. \(\PageIndex{1}\)). Мегаспоры крупнее, потому что они богаты питательными веществами, готовы к оплодотворению и питанию развивающегося спорофита.
Рисунок \(\PageIndex{1}\): Селагинелла, одно из бессемянных сосудистых растений, является гетероспоровым. Несколько мегаспор (А) образуются в мегаспорангии (С), в то время как многие микроспоры (В, слишком маленькие, чтобы различить их по отдельности) образуются в микроспорангии (D). Мегаспоры вырастут в гаметофиты, производящие архегонии (мегагаметофиты), а микроспоры разовьются в гаметофиты, производящие антерид (микрогаметофиты). Масштабная линейка соответствует 0,7 мм.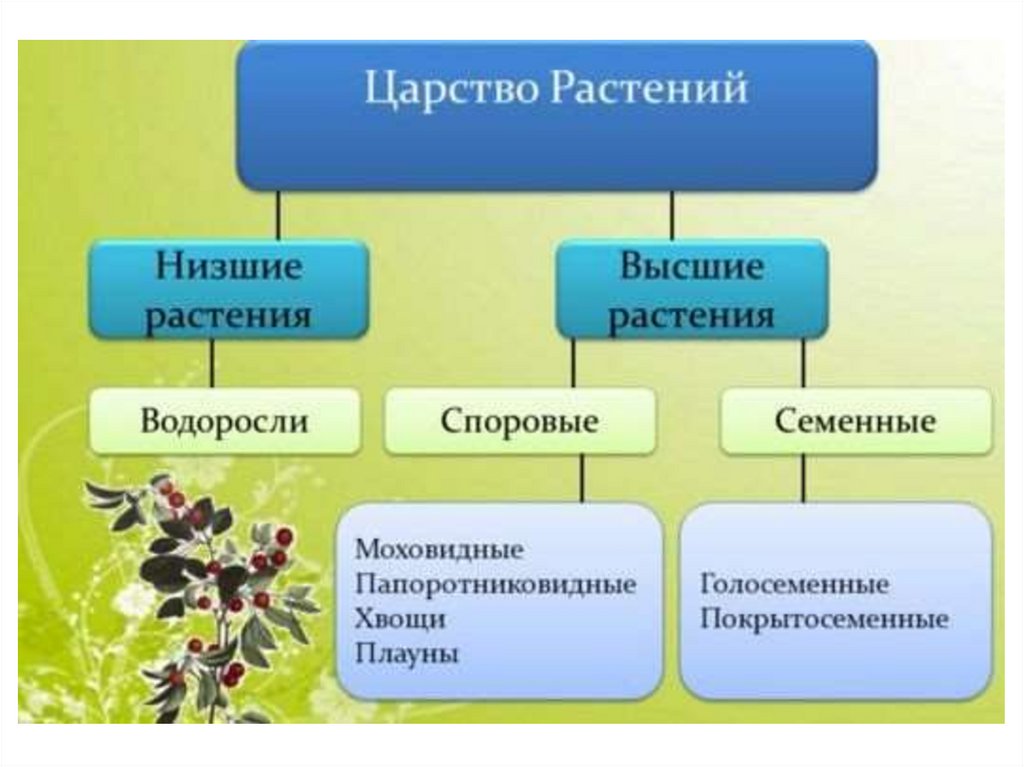 Фото Джона Хаусмана, CC BY-NC, с Викисклада.
Фото Джона Хаусмана, CC BY-NC, с Викисклада.
Этот цикл гетероспор делает оплодотворение менее зависимым от воды и более зависимым от распределения спор и особенностей гаметофита (Рисунок \(\PageIndex{2}\)). Это также позволяет увеличить вариации для действия отбора. Разделение труда позволяет использовать ресурсы более эффективно, а также ограничивает самоопыление. В эволюции растений потребность в гетероспории была высока, поскольку она независимо возникла в нескольких группах бессемянных сосудистых растений (например, Selaginella 9).0055 и Azolla ) и даже среди мхов.
По мере продвижения по эволюционной траектории гетероспории некоторые мегаспоры не покидают материнское растение, а вместо этого прорастают там, ожидая оплодотворения от соседнего микрогаметофита: шаг к первым семенам. Мужские гаметофиты становятся настолько мелкими, что их можно легко транспортировать целиком. Подвижной стадией становятся целые мужские гаметофиты, а не только гаметы — так происходит опыление .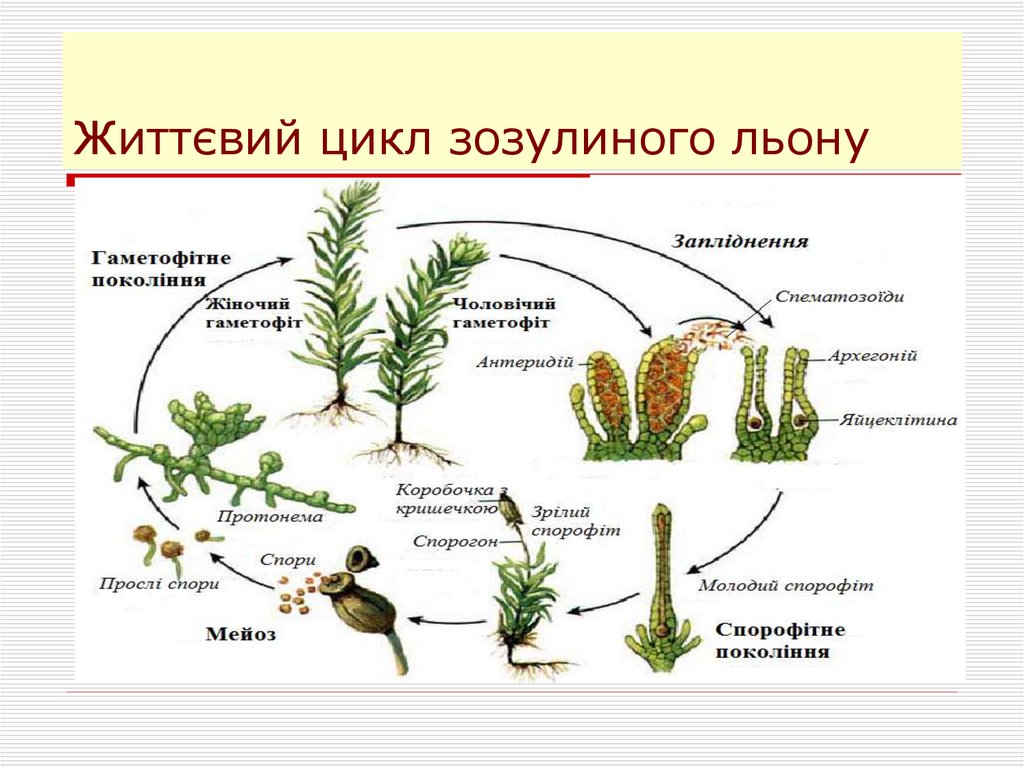
Рисунок \(\PageIndex{2}\): Гетероспоровый жизненный цикл с раздельнополыми спорофитами. Жизненные циклы растений могут быть довольно сложными! У двудомных растений микроспорангии образуются на микроспорофите, а мегаспорангии — на мегаспорофите. У однодомных растений микроспорангии и мегаспорангии образуются на одном и том же спорофите. Если образуются спорангии двух разных типов (мега- и микро-), жизненный цикл гетероспорический. Описательный текст: на этой диаграмме микроспорофит производит микроспоры, которые превращаются в микрогаметофиты, производящие сперму. Мегаспорофит производит мегаспоры, которые превращаются в мегагаметофиты, производящие яйца. Происходит оплодотворение, в результате которого образуется либо мега-, либо микроспорофит. Схема Питера Коксхеда, общественное достояние, через Wikimedia Commons.
Рост: вторичный рост и переход от семян к спорам
Когда растения развили вторичный рост , тип роста, который допускал боковое расширение и накопление древесной ткани, они могли расти намного больше.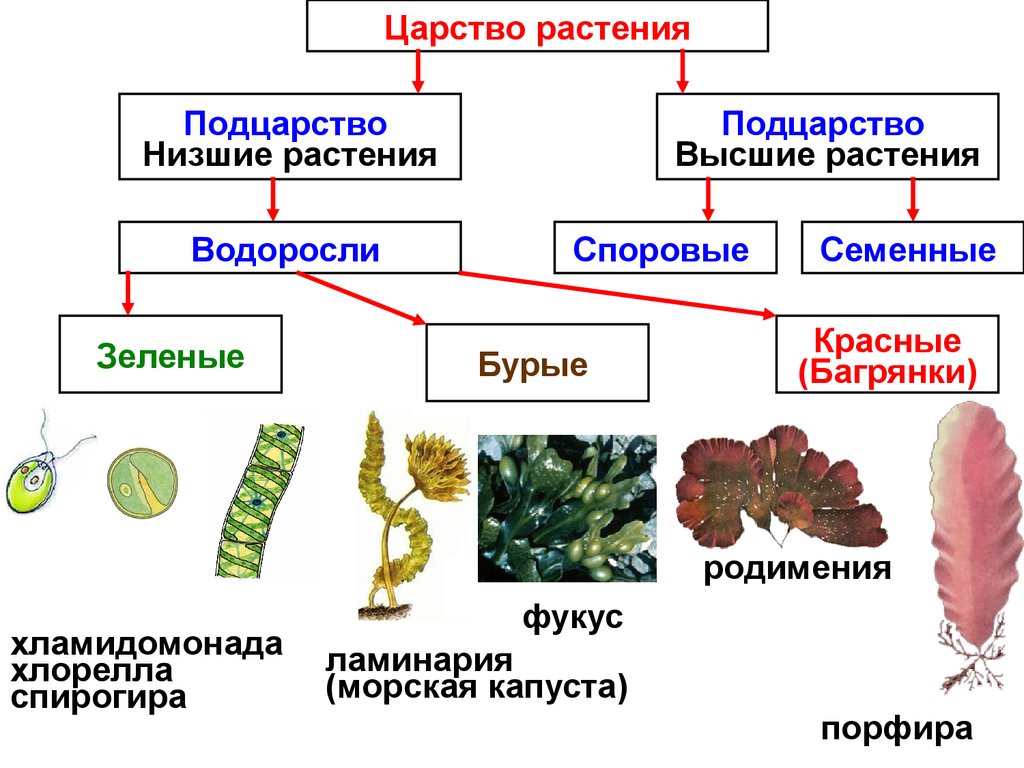 Однако эти гиганты столкнулись с новой проблемой. Крупные животные, такие как слоны, львы и киты, как правило, производят небольшое количество потомства, но увеличивают заботу о детях, чтобы обеспечить выживание. Другая стратегия, обычно используемая более мелкими организмами, состоит в том, чтобы производить большое количество потомков с небольшими затратами на каждое из них (например, споры большинства грибов), хотя большинство из них не выживает (рис. \(\PageIndex{3}\)).
Однако эти гиганты столкнулись с новой проблемой. Крупные животные, такие как слоны, львы и киты, как правило, производят небольшое количество потомства, но увеличивают заботу о детях, чтобы обеспечить выживание. Другая стратегия, обычно используемая более мелкими организмами, состоит в том, чтобы производить большое количество потомков с небольшими затратами на каждое из них (например, споры большинства грибов), хотя большинство из них не выживает (рис. \(\PageIndex{3}\)).
Рисунок \(\PageIndex{3}\): На этом изображении слон размножается с использованием двух разных стратегий. Хотя мы можем быть знакомы с первым слоном, второй тип слона не существует. Почему это может быть?
Представьте себе эти большие растения со вторичными утолщениями спор: они производят миллиарды спор с небольшим количеством питательных веществ или защиты для развивающихся потомков. Естественно, лишь немногие из этих миллиардов доживут до оплодотворения. Размножение спорами дешево и эффективно, но результаты непредсказуемы. Хуже того, эти леса споровых деревьев были вовсе не стабильны: в случайно хороших условиях многие споры выживали и образовывали спорофиты, которые все начинали расти одновременно, а затем подавляли друг друга. Но если условия окружающей среды плохие, то ни один из гаметофитов не выживет, поэтому не будет новых саженцев взамен старых деревьев.
Хуже того, эти леса споровых деревьев были вовсе не стабильны: в случайно хороших условиях многие споры выживали и образовывали спорофиты, которые все начинали расти одновременно, а затем подавляли друг друга. Но если условия окружающей среды плохие, то ни один из гаметофитов не выживет, поэтому не будет новых саженцев взамен старых деревьев.
Растения нашли решение этой загадки: семя . Идея семени состоит в том, чтобы скрыть большую часть жизненного цикла гетероспор внутри родительского растения. У семенных растений все происходит непосредственно на родительском спорофите: развитие гаметофитов, сингамия и рост потомства спорофита. Следовательно, мегаспора никогда не покидает спорангий. Он прорастает внутри, ждет оплодотворения, а затем зигота превращается в зародыш, все еще внутри того же спорангия. Что в конце концов покинет материнское растение, так это весь мегаспорангий с мегагаметофитом и зародышем внутри. Это химерная конструкция семени: спорофит, мегагаметофит, зародыш и (у покрытосеменных) генотипы эндосперма заключены в единую структуру.
Осталась одна проблема. Как сперматозоиды попадут в сохранившийся мегагаметофит и яйцеклетку? Теперь цель, возможно, находится высоко над землей, на ветке гигантского дерева. Единственное возможное решение – опыление. Опыление — это распространение всего мужского гаметофита, также называемого пыльцой . Растения физически не могут найти себе пару, поэтому им всегда нужна третья сторона для переноса пыльцы, обычно ветер или насекомые.
Как только пыльца попадет в пункт назначения, как сперматозоиды попадут в яйцеклетку? Некоторые семенные растения выделяют каплю жидкости из верхушки семязачатка (а пыльцевая капля ) для создания текучей связи между пыльцевым зерном и семязачатком. Другой инструмент доставки спермы, пыльцевая трубка (рис. \(\PageIndex{4}\)), сделана из одной из клеток пыльцевого зерна некоторых растений. Оплодотворение пыльцевой трубкой часто называют сифоногамией .
Рисунок \(\PageIndex{4}\): Открытое оплодотворение семенных растений (слева) с помощью жидкости, выделяемой материнским растением; и оплодотворение пыльцевой трубкой (справа), или сифоногамией, с помощью пыльцевой трубки, растущей из мужского гаметофита. Среди современных семенных растений только саговники и гинкго имеют открытое оплодотворение.
Следовательно, семенные растения с пыльцевой трубкой не имеют жгутиков на мужских гаметах; эти клетки сперматозоиды : неподвижные мужские гаметы. Пыльцевая трубка допускает только две мужские гаметы на гаметофит. Мужские гаметы обычно конкурируют за оплодотворение — это отбирает лучшие генотипы; тогда как у высших семенных растений конкуренция происходит между пыльцевыми трубками. Гаплоидная пыльцевая трубка растет внутри чужеродной ткани диплоидного спорофита, поэтому у многих семенных растений этот рост крайне медленный. Однако пыльцевые трубки покрытосеменных быстро росли.
Благодаря всем этим революционным приспособлениям семенные растения первыми заселили действительно засушливые места и, в свою очередь, позволили всем остальным живым существам выжить в засушливом климате. Узнайте больше об эволюции семян в этой статье от сентября 2020 года.
Узнайте больше об эволюции семян в этой статье от сентября 2020 года.
Голосеменные и покрытосеменные растения
Семенные растения состоят из двух основных линий: голосеменных (что означает голые семена) и покрытосеменных (что означает покрытые семена). Эти две линии представляют собой основные различия в чертах истории жизни, связанных с эволюционными условиями, в которых каждая из них развивалась.
Размножение голосеменных растений отличается от размножения покрытосеменных растений несколькими способами. У голосеменных мегагаметофит содержится в семязачатке, присутствует на открытых прицветниках мегастробила, где семена развиваются и часто высвобождаются пассивно (хотя некоторые из них рассеиваются животными). У покрытосеменных семязачаток находится внутри закрытой завязи. Затем завязь превращается в плод , специализирующийся на защите и распространении семян. Двойное оплодотворение является ключевым событием в жизненном цикле покрытосеменных, образуя второй запас питательных веществ для развивающегося зародыша ( эндосперм ), но отсутствует у большинства голосеменных растений. Структуры гаметофитов производятся отдельными стробилами у голосеменных растений, тогда как у покрытосеменных они часто являются частью одного и того же цветка . Ветер играет важную роль в опылении голосеменных, потому что пыльца переносится ветром и попадает на женские шишки. Хотя многие покрытосеменные также опыляются ветром, опыление животными более распространено, что приводит к появлению у цветов специализированных синдромов опыления.
Структуры гаметофитов производятся отдельными стробилами у голосеменных растений, тогда как у покрытосеменных они часто являются частью одного и того же цветка . Ветер играет важную роль в опылении голосеменных, потому что пыльца переносится ветром и попадает на женские шишки. Хотя многие покрытосеменные также опыляются ветром, опыление животными более распространено, что приводит к появлению у цветов специализированных синдромов опыления.
7.1: Introduction to Seed Plants распространяется по незаявленной лицензии, автором, ремиксом и/или куратором является LibreTexts.
- Наверх
- Была ли эта статья полезной?
- Тип изделия
- Раздел или Страница
- Версия лицензии
- 4,0
- Программа OER или Publisher
- Программа ASCCC OERI
- Теги

Развитие семян – неодушевленная жизнь
- Рис. 1 Знакомые семена гороха, кукурузы и фасоли на протяжении тысячелетий отбирались из-за большого размера.
- Рис. 2. Многие семена намного меньше, чем семена гороха, кукурузы или бобов. На этой фигуре семя гороха сравнивается с типичным семенем горчицы.
На первый взгляд производство семян (рис. 1-2) напоминает производство потомства у знакомых животных: внутри диплоидного родителя развивается член «следующего поколения», который взращивается внутри своего родителя на критических ранних стадиях развития, а затем депонируется вне своего родителя, чтобы закончить свою жизнь. Но помните, что все растения демонстрируют чередование поколений, поэтому, если диплоидное (спорофитное) растение дает новое диплоидное (спорофитное) растение в семени, необходимо учитывать поколение гаплоидного гаметофита, которое должно было появиться между двумя поколениями спорофитов. И нужно также понимать, что семена НЕ заменяют споры, на самом деле споры имеют решающее значение для производства семян. Появление семян (как в смысле эволюции, так и в смысле развития) — сложная история, включающая в себя схему «чередования поколений», проявляющуюся у всех растений. В свете этой закономерности семена представляют собой «бабушку» (русскую куклу) с несколькими поколениями, находящимися друг в друге. Понимание этого «поколения в поколении» необходимо для понимания того, «как появились семена» как в эволюции, так и в процессе развития.
И нужно также понимать, что семена НЕ заменяют споры, на самом деле споры имеют решающее значение для производства семян. Появление семян (как в смысле эволюции, так и в смысле развития) — сложная история, включающая в себя схему «чередования поколений», проявляющуюся у всех растений. В свете этой закономерности семена представляют собой «бабушку» (русскую куклу) с несколькими поколениями, находящимися друг в друге. Понимание этого «поколения в поколении» необходимо для понимания того, «как появились семена» как в эволюции, так и в процессе развития.
Хотя долгое время предполагалось, что такая сложная структура, как семена, возникла однажды, многие теперь считают, что семена эволюционировали несколько раз. Следовательно, семена могут представлять собой пример конвергентной эволюции, когда несколько линий сошлись на общем признаке. Независимо от того, так это на самом деле или нет, мы можем указать несколько особенностей, которые позволили семенам развиваться, и некоторые из этих особенностей проявляются в группах, которые не производят семян. Центральное место в появлении семян, как в смысле развития, так и в эволюционном смысле, занимает появление семязачатков, динамических объектов, состав которых меняется, в конечном итоге превращаясь в семя. В этой главе мы рассмотрим преобразования в жизненном цикле растений, которые позволили развиться семенам. В следующей главе мы рассмотрим специфические структуры и закономерности, наблюдаемые у хвойных и цветковых растений. Хотя мы сосредоточимся на семени, мы также рассмотрим сопутствующую сущность, которая необходима для развития семян: пыльцевое зерно, которое, как мы увидим, представляет собой миниатюрный подвижный мужской гаметофит.
Центральное место в появлении семян, как в смысле развития, так и в эволюционном смысле, занимает появление семязачатков, динамических объектов, состав которых меняется, в конечном итоге превращаясь в семя. В этой главе мы рассмотрим преобразования в жизненном цикле растений, которые позволили развиться семенам. В следующей главе мы рассмотрим специфические структуры и закономерности, наблюдаемые у хвойных и цветковых растений. Хотя мы сосредоточимся на семени, мы также рассмотрим сопутствующую сущность, которая необходима для развития семян: пыльцевое зерно, которое, как мы увидим, представляет собой миниатюрный подвижный мужской гаметофит.
- Структура семян
- Сокращение
- Удержание
- Замедленное развитие
- Обеспечение
- Упаковка
Структура семени
Рис. 3 Структура семени двудольного растения с двумя семядолями и осью корень/побег
Семя состоит из трех компонентов: зародыша растения-спорофита, ткани, которая обеспечивает питание этого зародыша, и «семенная оболочка», ткань-контейнер, в которой развиваются зародыш и питательная ткань.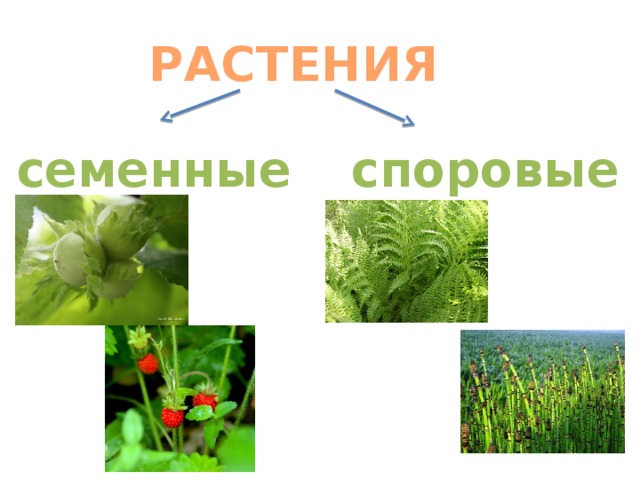 Эмбриональное растение диплоидно и развивается из зиготы, образованной слиянием яйцеклетки и спермия. Семенная кожура также диплоидна и также происходит от растения-спорофита, но это более раннее поколение спорофита, чем зародыш. Как во временном, так и в физическом смысле семя — это поколение «бабушка, матрешка», с «вложенными» поколениями. Есть два поколения спорофита, старое (семенная кожура) снаружи и новое (эмбрион) внутри, с поколением гаметофитов или остатками одного, зажатым между ними. Семена являются следствием того, что мегаспоры не рассеиваются, а вместо этого сохраняются в спорофите, который их производит. Споры прорастают, и, следовательно, женские гаметофиты, производящие яйца, присутствуют на/в спорофите. Позже на/в спорофите также присутствуют зародыши, возникающие в результате оплодотворения яиц, продуцируемых гаметофитами. Структура, в которой находится оставшаяся спора и где в конечном итоге развивается семя, называется яйцеклетка . В конечном итоге семязачатки развиваются в семена, содержащие новый спорофит, «упакованный» в семенную оболочку, ткань, полученную из исходного спорофита.
Эмбриональное растение диплоидно и развивается из зиготы, образованной слиянием яйцеклетки и спермия. Семенная кожура также диплоидна и также происходит от растения-спорофита, но это более раннее поколение спорофита, чем зародыш. Как во временном, так и в физическом смысле семя — это поколение «бабушка, матрешка», с «вложенными» поколениями. Есть два поколения спорофита, старое (семенная кожура) снаружи и новое (эмбрион) внутри, с поколением гаметофитов или остатками одного, зажатым между ними. Семена являются следствием того, что мегаспоры не рассеиваются, а вместо этого сохраняются в спорофите, который их производит. Споры прорастают, и, следовательно, женские гаметофиты, производящие яйца, присутствуют на/в спорофите. Позже на/в спорофите также присутствуют зародыши, возникающие в результате оплодотворения яиц, продуцируемых гаметофитами. Структура, в которой находится оставшаяся спора и где в конечном итоге развивается семя, называется яйцеклетка . В конечном итоге семязачатки развиваются в семена, содержащие новый спорофит, «упакованный» в семенную оболочку, ткань, полученную из исходного спорофита. До этого семяпочка содержит женский гаметофит; до этого семязачатки содержат спору, из которой образуется женский гаметофит; еще раньше они содержат материнскую клетку мегаспоры, которая производит эту спору. Обнаружение гаметофитов, как мужских, так и женских, и понимание их развития является ключом к пониманию как эволюции, так и развития семян.
До этого семяпочка содержит женский гаметофит; до этого семязачатки содержат спору, из которой образуется женский гаметофит; еще раньше они содержат материнскую клетку мегаспоры, которая производит эту спору. Обнаружение гаметофитов, как мужских, так и женских, и понимание их развития является ключом к пониманию как эволюции, так и развития семян.
Семенные растения и их предки являются гетероспористыми, производящими два типа спор, которые развиваются в два типа гаметофитов, один мужской и один женский. Как эволюция семян, так и развитие любого отдельного семени включают модификации как мужского, так и женского гаметофита, модификации структур, которые их производят, а также изменения времени и места важных процессов развития.
Рис. 4 Сравнение жизненных циклов семенных и несеменных растений.
Мы можем описать преобразования, которые позволили развиться семенам, а также сделать возможным развитие семян с помощью аббревиатуры RRAPP: Reduction, Retention, Arrested development, Provisioning, Packaging.
Редукция
И мужской, и женский гаметофиты семенных растений значительно уменьшены в размерах по сравнению с гаметофитами других растений. Гаметофиты большинства групп растений менее выражены, чем спорофиты, но у семенных растений они настолько редуцированы, что закономерность чередования поколений трудно увидеть, и ошибочное представление о том, что растения размножаются подобно знакомым животным, т. е. что чередования нет поколений часто предполагают, что единственными гаплоидными клетками являются яйцеклетка и сперматозоиды. Мужской гаметофит семенных растений представляет собой пыльцу, организм из 3-6 клеток, который начинает свое развитие из споры на одном спорофитном растении и завершает свое развитие на другом спорофитном растении, в месте, близком к женскому гаметофиту. У некоторых групп пыльца высвобождает подвижный жгутиковый сперматозоид, но у знакомых групп, цветковых растений и хвойных, мужской гаметофит сравним с грибами в том смысле, что гаметофит прирастает к яйцеклетке (а не приплывает к ней). женский гаметофит сливается с ним и дает ядро спермия, которое соединяется с ядром яйцеклетки, образуя диплоидную зиготу. Хотя размеры мужского гаметофита сильно уменьшены, он может жить до года в хвойных, начиная свою жизнь на одном спорофите и заканчивая на другом спорофите.
женский гаметофит сливается с ним и дает ядро спермия, которое соединяется с ядром яйцеклетки, образуя диплоидную зиготу. Хотя размеры мужского гаметофита сильно уменьшены, он может жить до года в хвойных, начиная свою жизнь на одном спорофите и заканчивая на другом спорофите.
Женский гаметофит семенных растений также сильно редуцирован: у цветковых растений он составляет всего семь клеток, а у хвойных может превышать тысячу клеток. У всех семенных растений женский гаметофит существует исключительно внутри тканей породившего его спорофита, не имея никакого самостоятельного существования. Помимо уменьшения размера женского гаметофита, происходит также уменьшение количества женских гаметофитов, которые продуцируются каким-либо конкретным мегаспорангием. Помните, что женские гаметофиты вырастают из мегаспор, которые образуются после мейотического «клеточного деления» (на самом деле это процесс, включающий два клеточных деления) материнской клетки мегаспоры.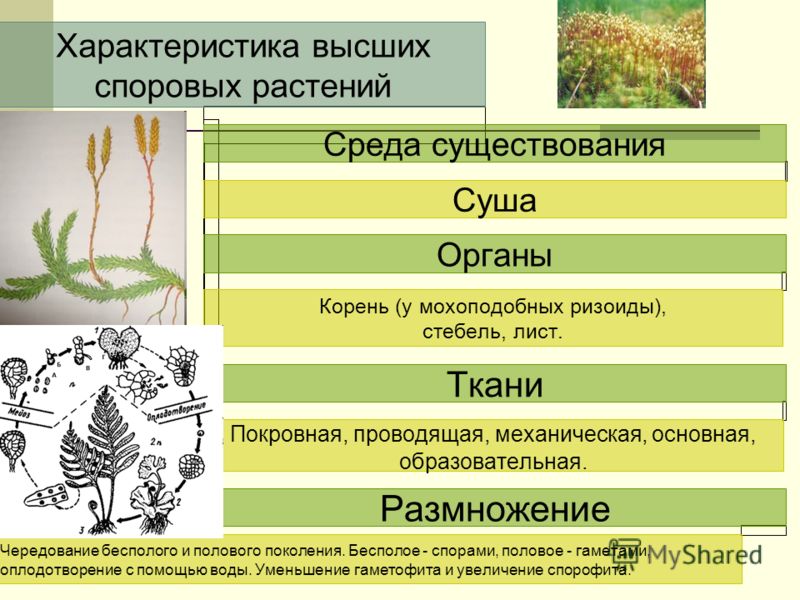 У всех семенных растений внутри мегаспорангия образуется только одна материнская клетка мегаспоры. И хотя, как правило, материнская клетка мегаспоры производит четыре споры после мейоза, у семенных растений три гаплоидных ядра дегенерируют после мейоза, оставляя одну мегаспору внутри мегаспорангия. Структура его совсем не спороподобна, специальной споровой стенки нет вообще; он не нуждается в защите, потому что всегда находится внутри ткани спорофита. Точно так же мегаспорангий, контейнер, в котором развиваются мегаспоры, сильно отличается от спорангиев несеменных штанов, потому что это не контейнер, подвергающийся воздействию окружающей среды, а вместо этого контейнер, встроенный в ткань спорофита. Развитие женского гаметофита семенных растений происходит в семязачатке, внутри мегаспорангия, развивающегося в семязачатке. У семенных растений мегаспорангий называется нуцеллусом, а у некоторых групп нуцеллус остается признаком полностью развитого семени.
У всех семенных растений внутри мегаспорангия образуется только одна материнская клетка мегаспоры. И хотя, как правило, материнская клетка мегаспоры производит четыре споры после мейоза, у семенных растений три гаплоидных ядра дегенерируют после мейоза, оставляя одну мегаспору внутри мегаспорангия. Структура его совсем не спороподобна, специальной споровой стенки нет вообще; он не нуждается в защите, потому что всегда находится внутри ткани спорофита. Точно так же мегаспорангий, контейнер, в котором развиваются мегаспоры, сильно отличается от спорангиев несеменных штанов, потому что это не контейнер, подвергающийся воздействию окружающей среды, а вместо этого контейнер, встроенный в ткань спорофита. Развитие женского гаметофита семенных растений происходит в семязачатке, внутри мегаспорангия, развивающегося в семязачатке. У семенных растений мегаспорангий называется нуцеллусом, а у некоторых групп нуцеллус остается признаком полностью развитого семени.
Удержание
Существует несколько этапов удержания, связанных с производством семян, и только последние этапы относятся только к семенным растениям.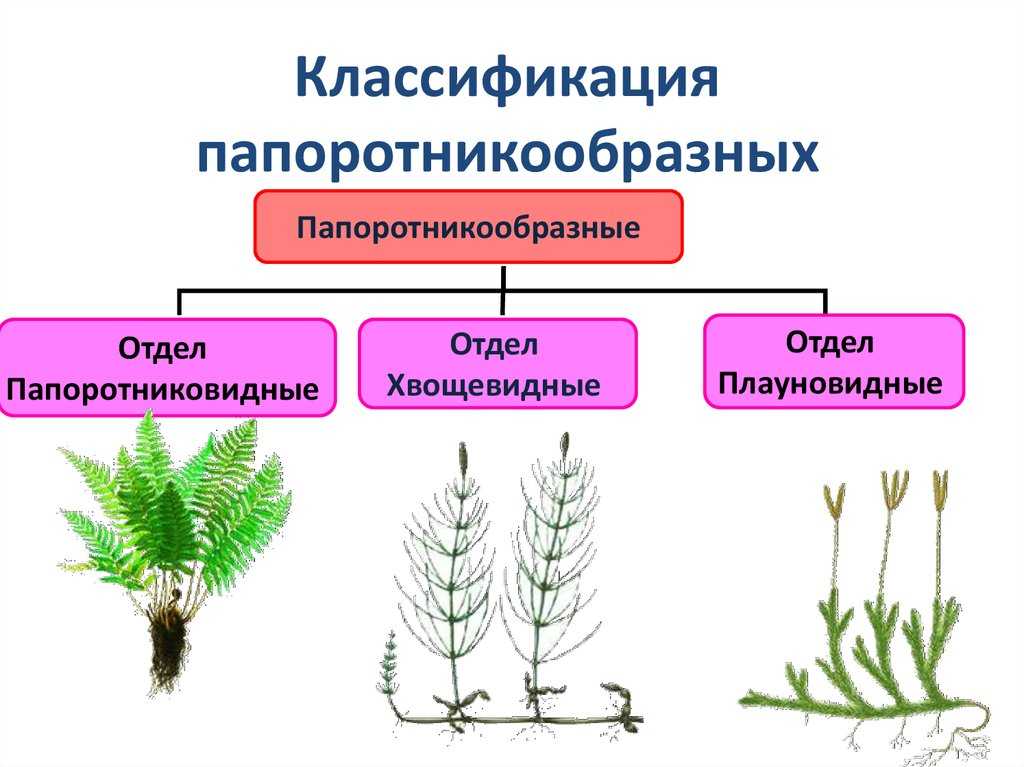 Во-первых, должна быть задержка яйцеклетки в организме, который ее производит. У организмов, у которых гаметы дифференцированы на «сперматозоид» (подвижную, обычно жгутиковую клетку, высвобождаемую из организма, который ее производит) и «яйцеклетку» (определяемую как более крупная и неподвижная гамета), удержание яйцеклетки является часто (например, Oedogonium , водяная плесень), но не всегда (например, Fucus) , случай. Удержание яйцеклетки требует, чтобы оплодотворение происходило внутри/на яйцеклетирующем организме. Это относится к млекопитающим. Для организмов, демонстрирующих чередование поколений, это означает, что яйцеклетка оплодотворяется в гаметофите. Следующее обнаруженное удержание характерно для всех растений, поэтому эту группу иногда называют «эмбриофитами». Это сохранение зиготы и вырастающего из нее зародыша в женском гаметофитном растении. Сохранение яйца, зиготы и эмбриона в гаметофите позволяет раннему развитию диплоидного поколения происходить в более контролируемой среде с ресурсами, предоставляемыми гаметофитом.
Во-первых, должна быть задержка яйцеклетки в организме, который ее производит. У организмов, у которых гаметы дифференцированы на «сперматозоид» (подвижную, обычно жгутиковую клетку, высвобождаемую из организма, который ее производит) и «яйцеклетку» (определяемую как более крупная и неподвижная гамета), удержание яйцеклетки является часто (например, Oedogonium , водяная плесень), но не всегда (например, Fucus) , случай. Удержание яйцеклетки требует, чтобы оплодотворение происходило внутри/на яйцеклетирующем организме. Это относится к млекопитающим. Для организмов, демонстрирующих чередование поколений, это означает, что яйцеклетка оплодотворяется в гаметофите. Следующее обнаруженное удержание характерно для всех растений, поэтому эту группу иногда называют «эмбриофитами». Это сохранение зиготы и вырастающего из нее зародыша в женском гаметофитном растении. Сохранение яйца, зиготы и эмбриона в гаметофите позволяет раннему развитию диплоидного поколения происходить в более контролируемой среде с ресурсами, предоставляемыми гаметофитом.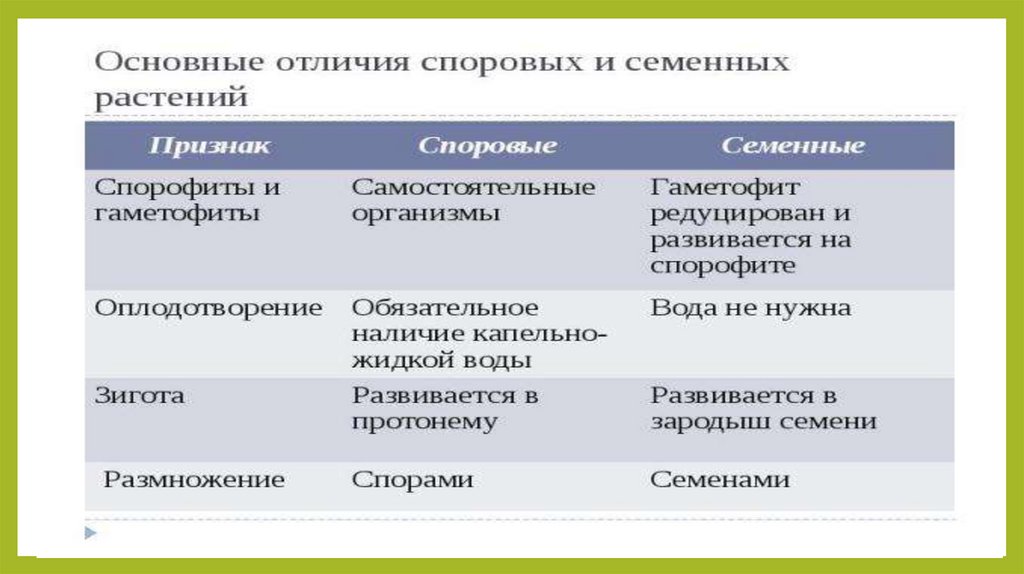 Обратите внимание, что это удержание приводит к тому, что спорофит вырастает из гаметофита, что опять же встречается у всех растений.
Обратите внимание, что это удержание приводит к тому, что спорофит вырастает из гаметофита, что опять же встречается у всех растений.
Следующим этапом удержания является сохранение гаметофита на спорообразующем растении. Гаметофиты развиваются из спор, образующихся в спорангии (спорангии), присутствующей внутри/на спорофитном растении. У семенных растений (и очень немногих растений, не дающих семена, например, у некоторых колосовидных мхов) гаметофиты сохраняются на спорофитном растении, которое их производит, потому что спора остается в спорангии спорообразующего растения, а не рассеивается из него. . Для женских гаметофитов семенных растений это сохранение является постоянным, женский гаметофит обитает только на спорофитном растении / в мегаспорангии, структуре, в которой образовались мегаспоры (крупные споры, которые развиваются в женские гаметофиты). Мужской гаметофит также проявляет задержку, но только временно; микроспоры (мелкие споры, которые развиваются в мужские гаметофиты) сохраняются в микроспорангии родительского (спорофитного) растения, и там происходит начальное развитие мужского гаметофита с образованием пыльцевого зерна. Затем он рассеивается и завершает свое развитие на другом спорофитном растении, в структуре которого находится женский гаметофит (женская шишка хвойных, цветок покрытосеменных).
Затем он рассеивается и завершает свое развитие на другом спорофитном растении, в структуре которого находится женский гаметофит (женская шишка хвойных, цветок покрытосеменных).
Задержка развития
Важным аспектом размножения является расселение. Особенно для растений с их неопределенным образом жизни размножение не имеет большого значения, если только нет возможности рассеяться на новое место. Рассеивание также является важным аспектом секса. Хотя процессы сингамии и мейоза могут порождать изменчивость даже при самооплодотворении, они гораздо эффективнее производят изменчивость, исходный материал для эволюции, если в них участвуют генетически различные организмы. Для малоподвижных растений это требует перемещения одной особи к другой, т. е. расселения. Для несеменных растений на распространение влияют два агента распространения: спора и сперма. Споры обладают огромной способностью к распространению, потому что они малы и могут (как правило) разноситься ветром на большие расстояния. Это возможно только потому, что живое внутри споры, отдельная клетка, крайне «безжизненно»; то есть, если бы кто-то наблюдал его, он бы увидел очень небольшую биологическую активность. Метаболизм минимален, происходит очень мало химических реакций; это очень устойчивая структура, находящаяся в состоянии «приостановки жизнедеятельности», нормальные функции жизни временно приостановлены. Эти жизненные функции возобновляются, если спора достигает среды обитания, которая может вызвать прорастание споры, которое возвращает спору в оживленное состояние. Напротив, другой подвижный агент несеменных растений, сперматозоиды, очень активны, у них очень высокая скорость метаболизма не только из-за их подвижности (обмен веществ необходим для движения жгутиков), но и из-за их общей структуры. по организации мембран и цитозоля гораздо более типичен для живых существ, чем для покоящихся спор. Сперматозоиды потребляют запасы энергии («еда») для поддержания своих жизненных функций. Поскольку сперматозоиды имеют очень ограниченную способность добывать пищу, продолжительность их жизни определяется объемом хранимых резервов, которыми они снабжаются при рассеивании.
Это возможно только потому, что живое внутри споры, отдельная клетка, крайне «безжизненно»; то есть, если бы кто-то наблюдал его, он бы увидел очень небольшую биологическую активность. Метаболизм минимален, происходит очень мало химических реакций; это очень устойчивая структура, находящаяся в состоянии «приостановки жизнедеятельности», нормальные функции жизни временно приостановлены. Эти жизненные функции возобновляются, если спора достигает среды обитания, которая может вызвать прорастание споры, которое возвращает спору в оживленное состояние. Напротив, другой подвижный агент несеменных растений, сперматозоиды, очень активны, у них очень высокая скорость метаболизма не только из-за их подвижности (обмен веществ необходим для движения жгутиков), но и из-за их общей структуры. по организации мембран и цитозоля гораздо более типичен для живых существ, чем для покоящихся спор. Сперматозоиды потребляют запасы энергии («еда») для поддержания своих жизненных функций. Поскольку сперматозоиды имеют очень ограниченную способность добывать пищу, продолжительность их жизни определяется объемом хранимых резервов, которыми они снабжаются при рассеивании. Следствием этого является то, что сперматозоиды не живут очень долго и не перемещаются очень далеко.
Следствием этого является то, что сперматозоиды не живут очень долго и не перемещаются очень далеко.
У семенных растений также есть два агента распространения: семена и пыльца. Оба они находятся «в анабиозе» точно так же, как и споры. Это позволяет им обоим преодолевать значительные расстояния. Важно отметить, что и семя, и пыльца не являются отдельными клетками, это частично развитые организмы, которые начали свое развитие на «родительском» растении, но остановили свое развитие и вошли в «стадию покоя», где они могут пережить неблагоприятные условия. и жить без каких-либо дополнительных ресурсов. В то время как производство спящих клеток (например, спор) встречается у различных групп организмов, остановка развития многоклеточного организма и перевод его в спящую стадию встречается гораздо реже, но это необходимо для развития семян. Мужские гаметофиты начинают свое развитие внутри растения-спорофита, когда образуются микроспоры, которые превращаются в пыльцу. Затем они рассеиваются в спящем состоянии только для того, чтобы возродиться, если они достигают места женского гаметофита, расположенного на спорофитном растении или внутри него.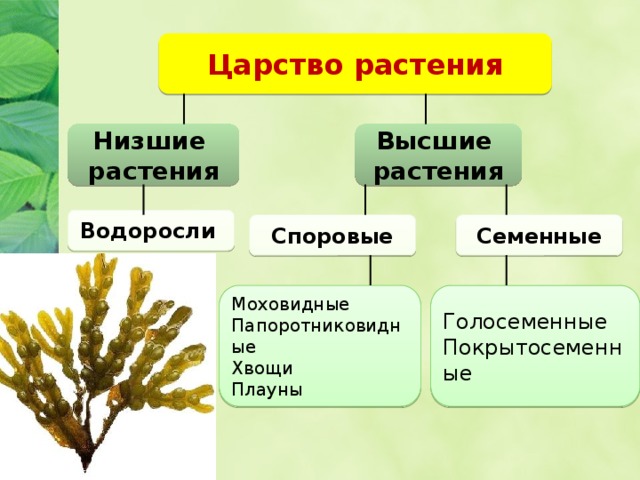 Здесь их развитие возобновляется, и они способны произвести клетку, способную оплодотворить яйцеклетку.
Здесь их развитие возобновляется, и они способны произвести клетку, способную оплодотворить яйцеклетку.
Точно так же новое поколение спорофита, обнаруженное в семени, демонстрирует задержку развития. После оплодотворения образующаяся зигота обычно быстро делится и развивается в зародыш с осью корень/побег, апикальными меристемами корня и побега и одним или двумя листьями, которые являются «семенными листьями» = семядолями. Затем развитие прекращается, не происходит ни деления, ни дифференцировки клеток, несмотря на то, что питательные вещества доступны и условия окружающей среды благоприятны, по крайней мере временно. Скорость обмена веществ падает до очень низкого уровня, а толерантность эмбриона к экстремальным температурам и т. д. существенно возрастает. В этом состоянии семя рассеивается и возобновляет рост и развитие только при соблюдении условий, благоприятных для прорастания.
Обеспечение
Семена содержат питательные вещества, как питательные вещества, которые могут «поставлять энергию», т.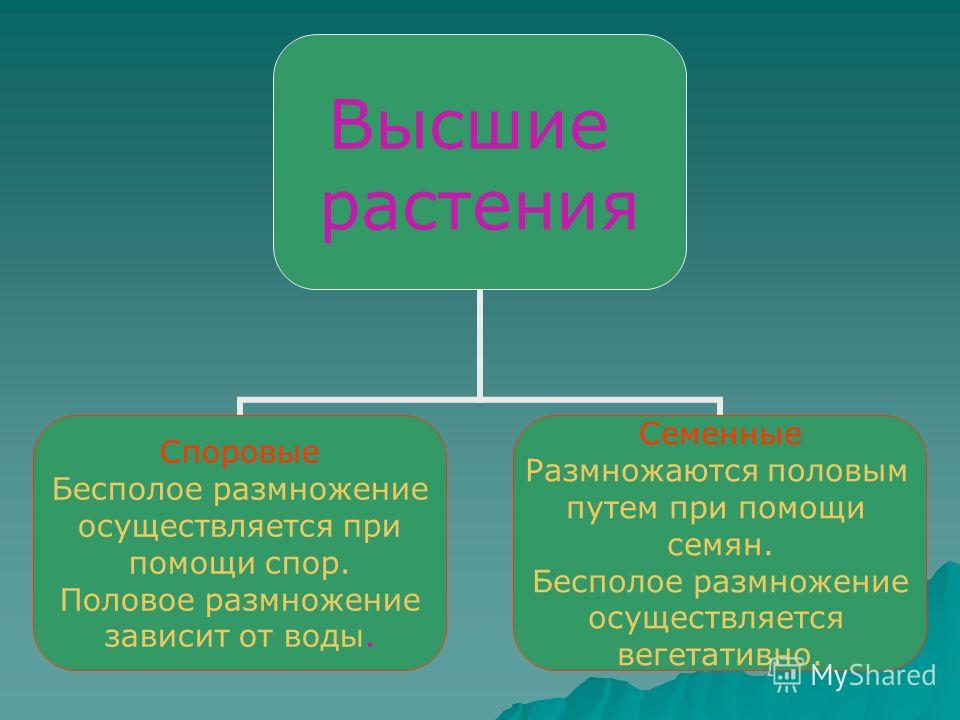 е. материалы, которые можно использовать в клеточном дыхании, так и питательные вещества, которые являются строительными материалами для строительства большего количества растительных клеток. Эти энергетические и материальные запасы очень важны для молодого автотрофа, потому что «чтобы делать деньги, нужны деньги»: чтобы прокормить себя, растению нужна структура, но структура невозможна без запасов энергии и материалов. Обеспечение позволяет организму создавать структуру, которая может самостоятельно получать материю и энергию. Обеспечение осуществляется спорами, но в меньшей степени; у них действительно есть некоторые материалы, но их количество ограничено, потому что спора, как правило, представляет собой только одну клетку и, как правило, небольшого размера, что способствует распространению.
е. материалы, которые можно использовать в клеточном дыхании, так и питательные вещества, которые являются строительными материалами для строительства большего количества растительных клеток. Эти энергетические и материальные запасы очень важны для молодого автотрофа, потому что «чтобы делать деньги, нужны деньги»: чтобы прокормить себя, растению нужна структура, но структура невозможна без запасов энергии и материалов. Обеспечение позволяет организму создавать структуру, которая может самостоятельно получать материю и энергию. Обеспечение осуществляется спорами, но в меньшей степени; у них действительно есть некоторые материалы, но их количество ограничено, потому что спора, как правило, представляет собой только одну клетку и, как правило, небольшого размера, что способствует распространению.
Семена снабжены материалами, которые хранятся в нескольких различных тканях:
- Материалы могут храниться в женском гаметофите, который иногда является компонентом семени: в семенах хвойных женский гаметофит представляет собой структуру примерно из 1000 клеток, все из которых могут хранить материалы,
- Материал может храниться в самом зародыше, в частности, в семядолях, «семенном листе/листьях», производимых зародышевым растением при росте внутри семени. У многих покрытосеменных семядоли увеличиваются во время развития и хранят значительное количество материалов. Многие из наших видов сельскохозяйственных культур обладают большими семядолями, которые занимают большую часть объема семян и содержат большую часть сохраняемого материала.
- Материал может храниться в ткани эндосперма, уникальной ткани, обнаруженной в семенах покрытосеменных, которые образуются в результате второго события оплодотворения, помимо того, которое создало зиготу. В следующей главе мы обсудим, как эта ткань формируется и развивается. Хотя все покрытосеменные имеют ткань эндосперма в какой-то момент своего развития, некоторые покрытосеменные теряют свой эндосперм по мере расширения семядолей, и семя остается с увеличенными семядолями, но с небольшим количеством эндосперма.
 У многих покрытосеменных семядоли увеличиваются во время развития и хранят значительное количество материалов. Многие из наших видов сельскохозяйственных культур обладают большими семядолями, которые занимают большую часть объема семян и содержат большую часть сохраняемого материала.
У многих покрытосеменных семядоли увеличиваются во время развития и хранят значительное количество материалов. Многие из наших видов сельскохозяйственных культур обладают большими семядолями, которые занимают большую часть объема семян и содержат большую часть сохраняемого материала. Рис. 5 Обратите внимание на сосудистые следы («нити»), которые соединяются с каждым семенем. Они содержат сосудистую ткань, которая позволяет снабжать семена материалом родительского растения. Каждая горошина – это созревшая семяпочка.
Каждая горошина – это созревшая семяпочка.
Где бы и как бы они ни хранились, питательные вещества поступают от родительского растения-спорофита через ткань флоэмы. Тщательное изучение развивающихся семян покажет, что каждое из них прикреплено к родительскому растению посредством сосудистой нити.
Упаковка
Семена в конечном итоге рассыпаются в виде упаковки, внутри которой находится зародыш и хранящаяся пища. Наружная оболочка (семенная кожура) развивается из тканей спорофита родительского растения, называемых интегументами, одним или несколькими слоями ткани спорофита, образующими наружные слои семязачатка. Частью превращения семязачатка в семя является превращение покровов (или части покровов) в семенную кожуру. Семенная оболочка развивается в процессе деления и дифференциации клеток, образуя то, что обычно представляет собой жесткую внешнюю оболочку семени. Эти клетки обычно имеют толстые вторичные клеточные стенки.
Таким образом, семя представляет собой зародыш и хранящуюся в семенной оболочке пищу.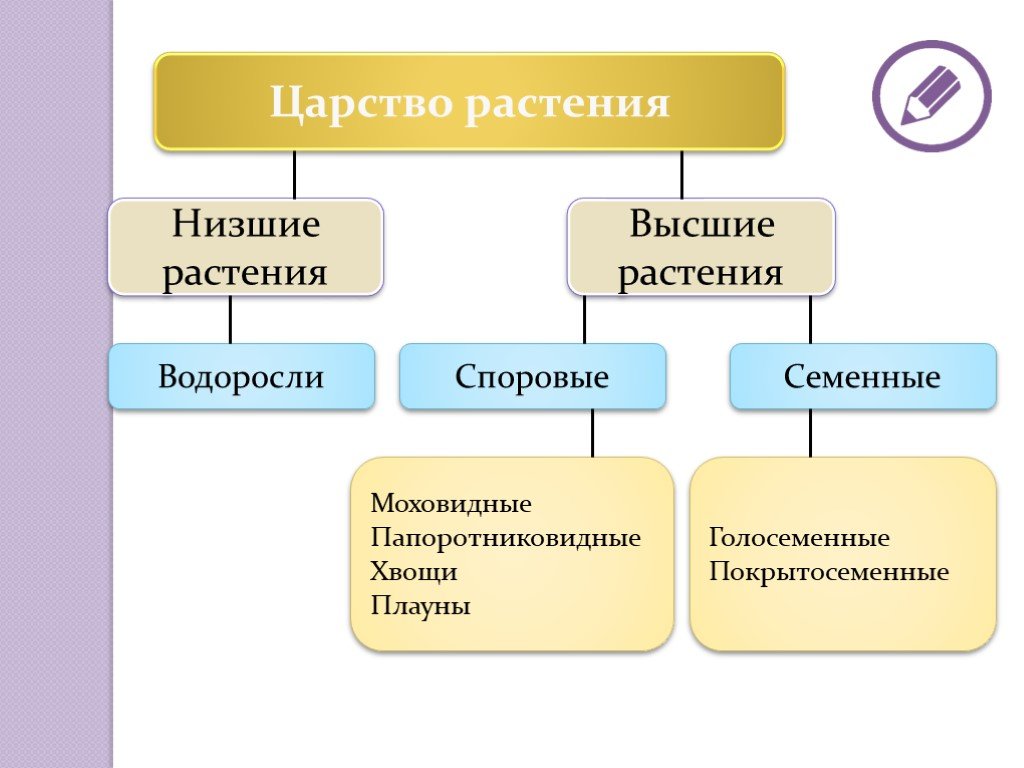 Развитие семени, также известное как созревание семязачатка, включает три различных процесса развития:
Развитие семени, также известное как созревание семязачатка, включает три различных процесса развития:
- Развитие зиготы в эмбрион. Зигота – новое поколение спорофита. Он делится, чтобы произвести новые клетки, которые растут и развиваются, чтобы произвести зародыш как с корневой, так и с апикальной меристемой побега, а также с одним или несколькими листьями, называемыми семядолями, или семенными листьями.
- Превращение тканей спорофита семязачатка в семенную кожуру. Обычно это включает производство новых клеток, что позволяет яйцеклетке увеличиваться в размерах, и дифференцировку этих клеток для создания защитного контейнера.
- Развитие питательной ткани для питания семян после их рассеивания. Как указывалось выше, питательная ткань у разных семян развивается из разных источников, но она всегда связана с пролиферацией клеток и расширением этих клеток по мере поступления в них материала от родительского спорофита.
Растения с семенами были успешными по нескольким причинам, включая: (1) раннее развитие спорофита происходит в очень защищенном месте внутри предыдущего поколения спорофита, (2) рассеиваются два элемента: пыльца, распространение которой сосредоточено на соединение гамет и семена, распространение которых направлено на доставку следующего поколения спорофитов в новые места; утвердиться.