
Содержание
Что такое вирусы и как они влияют на животных и показатели продуктивности?
В создании статьи принимал участие д-р Марко Реболло, двм
Часть 1 из 4 серий статей, посвященных роли вирусов в продуктивности животных.
Вирусы оказывают значительное влияние на продуктивность животных по всему миру. Вирусы могут поражать практически любые ткани в организме, включая кишечник, легкие, печень, почки, матку, яичники, позвоночный столб, мозг и даже кожу.
Когда вирус проникает в организм животного, он может угнетать рост, продуктивность, репродуктивные качества и даже приводить к летальному исходу. Контроль опасных вирусных болезней, таких как грипп, инфекционный бронхит, респираторно-синцитиальная инфекция КРС и многих других – ключ к получению здоровых высокопродуктивных животных. Для начала, необходимо понимать, что такое вирусы и как они взаимодействуют с организмом животного.
Вирус – это микроскопический инфекционный агент, способный размножаться только внутри живой клетки; вне организма хозяина вирус не может долго существовать. Существуют вирусы растений, животных, насекомых, они находятся в окружающей среде по всему миру, приводя к нарушению функции и структур в организмах животных.
Существуют вирусы растений, животных, насекомых, они находятся в окружающей среде по всему миру, приводя к нарушению функции и структур в организмах животных.
Контроль вирусов, достигаемый применением вакцин, усиливает биологическую безопасность, а противовирусные препараты, наравне с другими методами борьбы, а также обеспечение необходимого количества цинка в кормовых источниках, помогает животным продлить сроки жизни, повысить продуктивность и репродуктивные качества.
Как действуют вирусы животных?
Вирусы состоят из одно-или двух цепочечной нуклеиновой кислоты, РНК или ДНК, покрытой капсулой или оболочкой. Вирусы могут проникать в организм алиментарным (с кормом), аэрогенным (с вдыхаемым воздухом), контактным (через кожные покровы) или половым путем при половом акте.
Как только животное заражается вирусом, последний может внедрить свой генетический материал в здоровую клетку. После чего он заставляет клетку работать на себя, производя вирусные частицы, которые затем выходят из клетки и распространяются по организму, выходят во внешнюю среду и заражают других животных, а в редких случаях даже и людей.
На протяжении уже многих лет вирусы считаются этиологическим агентом болезни, однако не все вирусы сразу вызывают заболевания. Они могут находиться в спящем состоянии внутри клеток животных, пока организм не подвергнется воздействию сильного стресс-фактора. Это может быть транспортировка, слишком высокая или низкая температура, высокая плотность посадки. Стресс может приводить к тому, что вирус выходит из состояния спячки и начинает вызывать клиническое проявление болезни.
Хотя некоторые вирусы могут приводить к летальному исходу, большинство вирусных инфекций не вызывают гибели животных. Вирусы угнетают нервную систему,что значительно облегчает бактериальным патогенам проникновение внутрь животного, приводя с смерти от бактериальных инфекций. Такое часто встречается у откормочного скота.
А что насчет коронавирусов?
Вообще, у всех видов животных существуют коронавирусы – респираторные или кишечные – которые могут поражать организм. Коронавирусы достаточно широко распространены среди животных. Например, у скота это коронавирус КРС, вызывающий респираторные заболевания. У птиц вирус инфекционного бронхита (ИБЦ) относится к семейству коронавирусов. Эпизоотическая диарея свиней (ЭЗС) – коронавирус, способный поражать молодых свиней. . Наши домашние любимцы – кошки и собаки – могут заразиться кошачьим инфекционным перитонитом и другими респираторными заболеваниями. Эти коронавирусы, поражающие сельскохозяйственных животных, отличаются от человеческих (SARS-CoV2, SARS-CoV и MERS-CoV). Вероятность того, что они могут заразить и вызвать заболевания у человека при контакте с животными крайне мала.
Например, у скота это коронавирус КРС, вызывающий респираторные заболевания. У птиц вирус инфекционного бронхита (ИБЦ) относится к семейству коронавирусов. Эпизоотическая диарея свиней (ЭЗС) – коронавирус, способный поражать молодых свиней. . Наши домашние любимцы – кошки и собаки – могут заразиться кошачьим инфекционным перитонитом и другими респираторными заболеваниями. Эти коронавирусы, поражающие сельскохозяйственных животных, отличаются от человеческих (SARS-CoV2, SARS-CoV и MERS-CoV). Вероятность того, что они могут заразить и вызвать заболевания у человека при контакте с животными крайне мала.
Трудности с вирусами животных
Основная проблема состоит в том, что вирусы микроскопически малы – их размер составляет менее 300 нанометров. Это значит, что вирусы могут распределяться в воздухе, и распространяться с воздушными потоками, особенно внутри закрытых помещений. Поэтому чем выше плотность посадки, тем легче вирусу заражать животных.
Например считается, что вспышка ящура – заболевания, поражающего многих парнокопытных, произошедшая в Великобритании и Франции, изначально была принесена вирусом с воздушными массами через Ла-Манш.
Итак, вирус может распространяться по воздуху, однако это не единственный его механизм распространения. Например, вирус энцефалита лошадей может передаваться комарами и птицами. Кроме того, летучие мыши – частые переносчики таких вирусов как бешенство, грипп и коронавирус, причем сами животные не проявляют клинических симптомов.
Другая проблема, с которой сталкиваются производители – способность вирусов к мутации. Так же как животные, развивающие свой иммунитет, чтобы побороть болезни, вирусы могут мутировать и изменяться. Это значит, что вирус может становиться более или менее патогенным, более или менее вирулентым, а может начать заражать и другие виды животных и даже человека. Именно этого больше всего опасаются в случаях, например, с птичьим и свиным гриппом. Когда вирус начинает адаптироваться к новому виду, он может стать чрезвычайно патогенным. Итак, вирус проникает в организм нового вида и начинает приспосабливаться к нему. Требуется время пока вирус адаптируется и наладит контакт с организмом хозяина. Именно поэтому сейчас мы имеем дело с массовыми вспышками и переживаем нынешнюю пандемию – это вирус, который адаптируется.
Именно поэтому сейчас мы имеем дело с массовыми вспышками и переживаем нынешнюю пандемию – это вирус, который адаптируется.
Контроль вирусных заболеваний у животных
Во всем мире созданы системы животноводства, которые способствуют увеличению распространения вирусных инфекций, в основном из-за высокой плотности посадки.
В результате производители животных могут уменьшить распространение вирусов, уменьшив плотность посадки. Многие птицеводческие системы уже делают это, поскольку индустрия переходит на производство без антибиотиков, что, естественно, привело к тому, что в каждом птичнике размещается меньше птиц.
Другой способ снизить распространение вирусов – применение противовирусных препаратов и вакцин. Против многих вирусов, в том числе гриппа, болезни Ньюкасла и болезни Марека разработаны эффективные вакцины, доступные для применения. Однако противовирусные препараты зачастую стоят слишком дорого, в то время как для производства вакцин требуется время, исследования и технологии. Поэтому лучший способ профилактики вирусных инфекций – усилить биологическую безопасность, чтобы удерживать вирус как можно дальше от животных.
Поэтому лучший способ профилактики вирусных инфекций – усилить биологическую безопасность, чтобы удерживать вирус как можно дальше от животных.
Усиление биологической безопасности включает в себя:
- Раздельное размещение помещений для содержания и кормления
- Дезинфекция транспортных средств, прибывающих или покидающих хозяйство и/или ограничение перемещения оборудования с одного хозяйства в другое, особенно если на комплексе содержатся птицы и свиньи разных возрастов
- Ввести правила для сотрудников: принимать душ и сменять одежду перед входом и при выходе с каждого комплекса
- Носить защитные маски, чепчики и перчатки при работе в помещениях, мыть руки каждый раз при переходе из одного птичника в другой
- Приезжающие грузовые машины должны быть продизенфицированы, а водители должны оставаться внутри транспортных средств или также подвергаться дезинфекции
- Обеспечьте достаточное время перерывов между заполнениями групп – более 10 дней – чтобы вирусы успели погибнуть, а все оборудование и помещения подверглись дезинфекции
Помещение прибывающих животных на карантин – отличный способ снизить вероятность заноса заболевания в стада в хозяйстве. В случае с рогатым скотом, при поступлении новых животных на комплекс, необходимо помещение их на 21-дневный карантин. Вирус диареи крупного рогатого скота, в частности, может сохраняться внутри организма животных в течение 18 дней до проявления симптомов.
В случае с рогатым скотом, при поступлении новых животных на комплекс, необходимо помещение их на 21-дневный карантин. Вирус диареи крупного рогатого скота, в частности, может сохраняться внутри организма животных в течение 18 дней до проявления симптомов.
В дополнение к биологической безопасности, важно убедиться в том, что у животных достаточно развит иммунитет. Необходимо обеспечить животных достаточным количеством белка и энергии, а также поддерживать оптимальное физическое состояние. Также важно, чтобы у животных имелся достаточный запас минералов и витаминов в организме, чтобы улучшить работу белых кровяных телец. И наконец, у животных всегда должен быть доступ к чистой свежей воде, чтобы профилактировать попадание бактерий и вирусов в пищеварительную систему.
Ознакомьтесь с Частью 2 из 4 серий статей о том, как вирусы влияют на продуктивность животных. В следующей части подробный рассказ о том, каким образом иммунная система защищает организм от вирусных инфекций. После чего в 3 части вы узнаете о роли незаменимых микроэлементов в защите животных от вирусных инфекций.
Биобезопасность вирусов растений для человека и животных — статья
В связи с техническими работами в центре обработки данных, возможность загрузки и скачивания файлов временно недоступна.
скрыть
Статья опубликована в журнале из списка RSCI Web of Science
Статья опубликована в журнале из перечня ВАК
- Авторы:
Никитин Н.А.,
Трифонова Е.А.,
Карпова О.В.,
Атабеков И.Г.
- Журнал:
Вестник Московского университета. Серия 16: Биология - Том:
3 - Год издания:
2016 - Издательство:
Изд-во Моск. ун-та
ун-та - Местоположение издательства:
М. - Первая страница:
20 - Последняя страница:
26 - Аннотация:
В последнее время вирионы и вирусоподобные частицы (ВПЧ) вирусов растений рассматриваются в качестве основы для создания новых биотехнологий в медицине и ветеринарии, в том числе для получения современных безопасных вакцин, систем адресной доставки и новых диагностических препаратов, а также для продукции терапевтических белков в растениях. Несмотря на то, что вирусы растений не могут размножаться в организме позвоночных, существуют данные, что они способны воспроизводить тот или иной этап инфекционного цикла в клетках млекопитающих. Более того, показано, что фитовирусы могут постоянно присутствовать в организме животных и человека, и могут использовать его в качестве переносчика. В обзоре представлены результаты по биосовместимости, токсичности, тератогенности и биораспределению вирусов растений. Основываясь на последних данных, можно утверждать, что фитовирусы безопасны для животных и человека. Показано, что вирионы биодеградируемы и легко выводятся из организма лабораторных животных. При этом вирионы и ВПЧ вирусов растений высокоиммуногенны и, при презентации антигенных детерминант патогенов человека и животных на их поверхности, позволяют смоделировать безопасную вирусную частицу, способную заменить живые аттенуированные вакцины. - Добавил в систему:
Никитин Николай Александрович
 ун-та
ун-та В обзоре представлены результаты по биосовместимости, токсичности, тератогенности и биораспределению вирусов растений. Основываясь на последних данных, можно утверждать, что фитовирусы безопасны для животных и человека. Показано, что вирионы биодеградируемы и легко выводятся из организма лабораторных животных. При этом вирионы и ВПЧ вирусов растений высокоиммуногенны и, при презентации антигенных детерминант патогенов человека и животных на их поверхности, позволяют смоделировать безопасную вирусную частицу, способную заменить живые аттенуированные вакцины.
В обзоре представлены результаты по биосовместимости, токсичности, тератогенности и биораспределению вирусов растений. Основываясь на последних данных, можно утверждать, что фитовирусы безопасны для животных и человека. Показано, что вирионы биодеградируемы и легко выводятся из организма лабораторных животных. При этом вирионы и ВПЧ вирусов растений высокоиммуногенны и, при презентации антигенных детерминант патогенов человека и животных на их поверхности, позволяют смоделировать безопасную вирусную частицу, способную заменить живые аттенуированные вакцины.Прикрепленные файлы
| № | Имя | Описание | Имя файла | Размер | Добавлен |
|---|
Биобезопасность вирусов растений для человека и животных
Balique, F., Colson, P. и Raoult, D., Вирус табачной мозаики в сигаретах и слюне курильщиков, J. Clin. Вирол. , 2012, том. 55, нет. 4, стр. 374–376 http://dx.doi.org/10.1016/j.jcv.2012.08.012.
Артикул
пабмедGoogle ученый
Куделка, К.Дж., Дестито, Г., Пламмер, Э.М., Траугер, С.А., Сьюздак, Г. и Манчестер, М., Нацеливание на эндотелий вируса мозаики вигны (CPMV) через поверхностный виментин, PLoS Патог. , 2009, том.
5, нет. 5, e1000417 http://dx.doi.org/10.1371/journal.ppat.1000417.Артикул
пабмед
ПабМед ЦентральныйGoogle ученый
Ли, Л., Ван, Л., Сяо, Р., Чжу, Г., Ли, Ю., Лю, К., Ян, Р., Тан, З., Ли, Дж., Хуанг , W., Chen, L., Zheng, X., He, Y. и Tan, J., Инвазия РНК вируса табачной мозаики индуцирует аутофагию, связанную со стрессом эндоплазматического ретикулума, в клетках HeLa, Biosci. Отчет , 2012, том. 32, нет. 2, стр. 171–186 http://dx.doi.org/10.1042/BSR20110069.
Артикул
КАС
пабмедGoogle ученый
Медейрос, Р.Б., Резенде Рде, О., и де Авила, А.С., Вирус растений Тосповирус пятнистого увядания томатов активирует иммунную систему своего основного переносчика насекомых, Frankliniella occidentalis, J. Virol. , 2004, том. 78, нет. 10, стр. 4976–4982 http://dx.doi.org/10.1128/JVI.78.10.4976-4982.
2004.КАС
пабмедGoogle ученый
Стаффорд, К.А., Уокер, Г.П., и Ульман, Д.Е., Заражение вирусом растений изменяет поведение переносчика при кормлении, Proc. Натл. акад. науч. США , 2011, том. 108, нет. 23, стр. 9350–9355 http://dx.doi.org/10.1073/pnas.1100773108.
Артикул
КАС
пабмед
ПабМед ЦентральныйGoogle ученый
Чжан, Т., Брейтбарт, М., Ли, У.Х., Ран, Дж.К., Вэй, К.Л., Сох, С.В., Хибберд, М.Л., Лю, И.Т., Ровер, Ф., и Руан, Ю., Р.Н.А. вирусное сообщество в фекалиях человека: преобладание вирусов фитопатогенов, PLoS Биол. , 2006, том. 4, нет. 1, e3 http://dx.doi.org/10.1371/journal.pbio.0040003.
Артикул
Google ученый
Бландино А., Лико К., Баскьери С., Барберини Л., Чиротто К., Блази П. и Санти Л.
, Оценка токсичности вируса растений in vitro и in vivo наноносители, Colloids Surf., B , 2015, vol. 129, стр. 130–136 http://dx.doi.org/10.1016/j.colsurfb.2015.03.039.Артикул
КАСGoogle ученый
Lee, K.L., Twyman, R.M., Fiering, S. и Steinmetz, N.F., Наночастицы на основе вирусов как технологии платформы для современных вакцин, Wiley Interdiscip. Рев.: Наномед. Нанобиотехнология. , 2016, том. 8, нет. 4, стр. 554–578. doi http://dx.doi.org/10.1002/wnan.1383
CAS
Google ученый
Брюнель, Ф.М., Льюис, Дж.Д., Дестито, Г., Стейнмец, Н.Ф., Манчестер, М., Стулманн, Х. и Доусон, П.Е., Стратегия лигирования гидразона для сборки многофункциональных вирусных наночастиц для визуализации клеток и нацеливания на опухоли , Нано Летт. , 2010, том. 10, нет. 3, стр. 1093–1097 http://dx.doi.org/10.1021/nl1002526.
Артикул
КАС
пабмед
ПабМед ЦентральныйGoogle ученый
Steinmetz, N.F., Ablack, A.L., Hickey, J.L., Ablack, J., Manocha, B., Mymryk, J.S., Luyt, L.G., and Lewis, J.D., Прижизненная визуализация рака предстательной железы человека с использованием вирусных наночастиц, нацеленных на пептидные рецепторы, высвобождающие гастрин, Small , 2011, vol. 7, нет. 12, стр. 1664–1672 http://dx.doi.org/10.1002/smll.201000435.
Артикул
КАС
пабмед
ПабМед ЦентральныйGoogle ученый
Rioux, G., Mathieu, C., Russell, A., Bolduc, M., Laliberte-Gagné, M.E., Savard, P., and Leclerc, D., Наночастицы PapMV улучшают иммунный ответ слизистой оболочки на трехвалентный вакцина против гриппа инактивированная J. Nanobiotechnol. , 2014, том. 12, нет. 1, 19 http://dx.doi.org/10.1186/1477-3155-12-19.
Артикул
Google ученый
Folwarczna, J., Moravec, T., Plchova, H., Hoffmeisterova, H., and Cerovska, N., Эффективная экспрессия онкобелка E7 вируса папилломы человека 16, слитого с C-концом белка оболочки вируса табачной мозаики (TMV) с использованием молекулярных шаперонов в Escherichia coli, Белковая экспрессия Purif. , 2012, том. 85, нет. 1, стр. 152–157 http://dx.doi.org/10.1016/j.pep.2012.07.008.
Артикул
КАСGoogle ученый
Карпова О., Никитин Н., Чирков С., Трифонова Е., Шевелева А., Лазарева Е., Атабеков Ю. Иммуногенные композиции, собранные из шаровидных форм, генерируемых вирусом табачной мозаики платформы частиц и чужеродные антигены, J.
Gen. Virol. , 2012, том. 93, нет. 2, стр. 400–407 http://dx.doi.org/10.1099/вир.0.036293-0.Артикул
КАС
пабмедGoogle ученый
Паскуаль, Д.В., Вакцины к обеду, Proc. Натл. акад. науч. США , 2007, том. 104, нет. 26, стр. 10757–10758 http://dx.doi.org/10.1073/pnas.0704516104.
Артикул
КАС
пабмед
ПабМед ЦентральныйGoogle ученый
Ночи Т., Такаги Х., Юки Ю., Ян Л., Масумура Т., Медзима М., Наканиси У., Мацумура А., Уодзуми А., Хирои Т., Морита С., Танака К., Такайва Ф. и Киёно Х., Мукозальная вакцина на основе риса как глобальная стратегия холодовой цепи и безыгольной вакцинации, Проц. Натл. акад. науч. США , 2007, том. 104, нет. 26, стр. 10986–10991 http://dx.doi.org/10.1073/pnas.0703766104.
Артикул
КАС
пабмед
ПабМед ЦентральныйGoogle ученый
McCormick, A.A., Corbo, T.A., Wykoff-Clary, S., Palmer, K.E., и Pogue, G.P. Бивалентные гибридные вакцины с химическим конъюгатом TMV-пептид улучшают клеточный иммунитет и защиту от опухолей, Bioconjugate Chem. , 2006, том. 17, нет. 5, стр. 1330–1338 http://dx.doi.org/10.1021/bc060124m.
Артикул
КАСGoogle ученый
Staczek, J., Bendahmane, M., Gilleland, L.B., Beachy, R.N., and Gilleland, H.E. Иммунизация химерным вирусом табачной мозаики, содержащим эпитоп белка наружной мембраны F Pseudomonas aeruginosa, обеспечивает защиту от заражения P.
aeruginosa, Вакцина , 2000, том. 18, нет. 21, стр. 2266–2274 http://dx.doi.org/10.1016/S0264-410X(99)00571-X.Артикул
КАС
пабмедGoogle ученый
Фудзияма К., Саеджунг В., Янагихара И., Накадо Дж., Мисаки Р., Хонда Т., Ватанабэ Ю. и Секи Т. Производство иммуногенных полиовирусный пептид с использованием векторной системы на основе вируса табачной мозаики, J. Biosci. биоинж. , 2006, том. 101, нет. 5, стр. 398–402 http://dx.doi.org/10.1263/jbb.101.398.
Артикул
КАС
пабмедGoogle ученый
Yin, Z., Nguyen, H.G., Chowdhury, S., Bentley, P., Bruckman, M.A., Miermont, A., Gildersleeve, J.C., Wang, Q. и Huang, X., Вирус табачной мозаики как новый носитель углеводных антигенов, ассоциированных с опухолью, Биоконъюгат Хим. , 2012, том. 23, нет. 8, стр. 1694–1703 http://dx.doi.org/10.1021/bc300244a.
Артикул
КАСGoogle ученый
Bruckman, M., Randolph, L., VanMeter, A., Hern, S., Shoffstall, A., Taurog, R., Steinmetz, N.F., Биораспределение, фармакокинетика и совместимость крови нативных и пегилированных нано-палочки и -сферы вируса табачной мозаики у мышей, Virology , 2014, vol. 449, стр. 163–173 http://dx.doi.org/10.1016/j.virol.2013.10.035.
КАС
пабмедGoogle ученый
Ниль, А., Аппаикс, Ф., Боска, С., ван дер Санден, Б., Никуд, Дж. Ф., Больце, Ф. и Хайнлайн, М., Флуоресцентные бионаночастицы, полученные из вируса табачной мозаики для прижизненной двухфотонной визуализации, Фронт. Растениевод. , 2016, том. 6, с. 1244 http://dx.doi.org/10.3389/fpls.2015.01244.
Артикул
пабмед
ПабМед ЦентральныйGoogle ученый
Wetter, C., Вирус табачной мозаики и вирус паратабачной мозаики в сигаретах, Naturwissenschaften , 1975, vol. 62, нет. 11, с. 533 http://dx.doi.org/10.1007/BF00609072.
Артикул
КАС
пабмедGoogle ученый
Bothwell, P.W., Рак легких и вирус табачной мозаики, Lancet , 1960, vol. 275, нет. 7125, стр. 657–658 http://dx.doi.org/10.1016/S0140-6736(60)
-1.
Артикул
Google ученый
Ифтихар Ю., Джексон Р. и Нойман Б.В., Обнаружение тобамовируса табачной мозаики в сигаретах с помощью ОТ-ПЦР, пак. Дж. Агрик.
науч. , 2015, том. 52, нет. 3, стр. 667–670.Google ученый
Накамура С., Ян К.С., Сакон Н., Уэда М., Туган Т., Ямасита А., Гото Н., Такахаши К., Ясунага Т., Икута, К., Мизутани Т., Окамото Ю., Тагами М., Морита Р., Маэда Н., Каваи Дж., Хаясидзаки Ю., Нагаи Ю., Хории Т., Иида, Т. и Накая Т., Прямое метагеномное обнаружение вирусных патогенов в носовых и фекальных образцах с использованием беспристрастного высокопроизводительного секвенирования, PLoS One , 2009 г., том. 4, нет. 1, e4219 http://dx.doi.org/10.1371/journal. поне.0004219.
Артикул
пабмед
ПабМед ЦентральныйGoogle ученый
Hymowitz, N., Курение и рак: обзор общественного здравоохранения и клинических последствий, J. Natl. Мед. доц. , 2011, том. 103, нет. 8, стр. 695–700 http://dx.doi.org/10.1016/S0027-9684(15)30408-9.
Артикул
пабмедGoogle ученый
- 033.
Артикул
КАСGoogle ученый
Пауэрс К.М., Кей Д.М., Фактор С.А., Забетян С.П., Хиггинс Д.С., Самии А., Натт Дж.Г., Гриффит А., Лейс Б., Робертс Дж.В., Мартинес Э.Д. , Монтимурро Дж.С., Чековей Х. и Паями Х. Комбинированные эффекты курения, стр. 9.0005 кофе и НПВП на риск болезни Паркинсона, Mov. Беспорядок. , 2008, том. 23, нет. 1, стр. 88–95 http://dx.doi.org/10.1002/mds.21782.
Google ученый
Лю, Р., Вайшнав, Р. А., Робертс, А. М., и Фридланд, Р. П., У людей есть антитела против вируса растений: доказательства вируса табачной мозаики, PLoS One , 2013, vol.
8, нет. 4, e60621 http://dx.doi.org/10.1371/journal.pone.0060621.Артикул
Google ученый
Эриксон Дж.О., Армен Д.М. и Либби Р.Л. Стойкость антигена у мышей, J. Immunol. , 1953, том. 71, стр. 30–37.
КАС
пабмедGoogle ученый
Эриксон Д.О., Хенсли Т.Дж., Филдс М., Либби Р.Л. Внутриклеточная локализация вируса табачной мозаики в Вестнике биологических наук Московского университета. 71 № 3 2016 БИОБЕЗОПАСНОСТЬ РАСТИТЕЛЬНЫХ ВИРУСОВ ДЛЯ ЧЕЛОВЕКА И ЖИВОТНЫХ 133 печень мыши, J. Иммунол. , 1957, том. 78, нет. 2, стр. 95–103.
КАС
пабмедGoogle ученый
Бусбиа, С., Папазян, Л., Ла Скола, Б., и Рауль, Д., Обнаружение растительной ДНК в бронхоальвеолярном лаваже пациентов с вентилятор-ассоциированной пневмонией, PLoS One , 2010, vol.
. 5, нет. 6, e11298 http://dx.doi.org/10.1371/journal.pone.0011298.Артикул
Google ученый
Le Clair, R.A., Извлечение культивируемого вируса табачной мозаики из мокроты и жидкостей после торакоцентеза, полученных от курильщиков сигарет с легочными заболеваниями в анамнезе, Am. Преподобный Респир. Дис. , 1967, том. 95, нет. 3, стр. 510–511.
Google ученый
Katsilambros, L., Вирус табачной мозаики и рак легких, Lancet , 1960, vol. 2, с. 934 http://dx.doi.org/10.1016/S0140-6736(60)92014-6.
Артикул
Google ученый
Knowland, J., Синтез белка, управляемый РНК из растительного вируса в нормальной клетке животного, Genetics , 1974, vol. 78, нет. 1, стр. 383–394.
КАС
пабмед
ПабМед ЦентральныйGoogle ученый
Карпова О., Иванов К., Родионова Н., Дорохов Ю., Атабеков Ю., Нетранслируемость и различное поведение в растениях и протопластах комплексов вирусной РНК и транспортных белков, образованных in vitro, Вирусология , 1997, вып. 230, нет. 1, стр. 11–21 http://dx.doi.org/10.1006/viro.1997,8472.
Артикул
КАС
пабмедGoogle ученый
Димитриадис, Г.Дж. и Georgatsos, J.G., Синтез белка оболочки вируса табачной мозаики после миграции вирусной РНК в изолированные митохондрии печени мыши, Nucleic Acids Res. , 1975, том. 2, нет. 10, стр.
1719–1726 http://dx.doi.org/10.1093/nar/2.10.1719.Артикул
КАС
пабмед
ПабМед ЦентральныйGoogle ученый
Балик Ф., Колсон П., Барри А.О., Наппез К., Ферретти А., Муссави К.А., Нгунга Т., Лепиди Х., Гиго Э., Меже Дж. ., Lecoq, H. и Raoult, D., Вирус табачной мозаики в легких мышей после интратрахеальной инокуляции, PLoS One , 2013, vol. 8, нет. 1, e54993 http://dx.doi.org/10.1371/journal. пон.0054993.
Артикул
Google ученый
Атабеков Ю., Никитин Н., Архипенко М., Чирков С., Карпова О. Термический переход нативного вируса табачной мозаики и безРНК вирусных белков в сферические наночастицы, Дж. Ген. Вирол. , 2011, том. 92, нет. 2, стр. 453–456 http://dx.doi.org/10.1099/vir.0.024356-0.
Артикул
КАС
пабмедGoogle ученый
Атабеков Дж., Добров Е., Карпова О. и Родионова Н., Вирус картофеля X: структура, разборка и восстановление , Mol. Завод Патол. , 2007, том. 8, нет. 5, стр. 667–675 http://dx.doi.org/10.1111/j.1364-3703.2007.00420.x.
Артикул
КАС
пабмедGoogle ученый
Марусич, К., Рицца, П., Латтанци, Л., Манчини, К., Спада, М., Беларделли, Ф., Бенвенуто, Э. и Капоне, И., Частицы химерного вируса растений в виде иммуногены для индукции иммунного ответа мыши и человека против вируса иммунодефицита человека 1, 9 типов0005 Дж.
Вирол. , 2001, том. 75, нет. 18, стр. 8434–8439 http://dx.doi.org/10.1128/JVI.75.18.8434-8439.2001.Артикул
КАС
пабмед
ПабМед ЦентральныйGoogle ученый
Shukla, S., Wen, A.M., Commandeur, U., и Steinmetz, N.F., Презентация эпитопов HER2 с использованием платформы для вакцинации на основе нитчатых вирусов растений, J. Mater. хим. В , 2014, т. 2, с. 2, стр. 6249–6258 http://dx.doi.org/10.1039/C4TB00749Б.
Артикул
КАСGoogle ученый
Lico, C., Mancini, C., Italiani, P., Betti, C., Boraschi, D., Benvenuto, E., and Baschieri, S., Растительно-продуцируемые химерные частицы вируса картофеля X, демонстрирующие пептид вируса гриппа активирует специфические CD8+ Т-клетки у мышей, Vaccine , 2009, vol. 27, нет. 37, стр. 5069–5076 http://dx.doi.org/10.1016/j.vaccine.2009.06.045.
Артикул
КАС
пабмедGoogle ученый
Джобсри Дж., Аллен А., Раджагопал Д., Шиптон М., Канюка К., Ломоносов Г.П., Оттенсмейер К., Диболд С.С., Стивенсон Ф.К., Савельева Н. ., Частицы растительного вируса, несущие опухолевый антиген, активируют TLR7 и индуцируют высокие уровни защитных антител, PLoS One , 2015 г., том. 10, нет. 2, e0118096 http://dx.doi.org/10.1371/journal.pone.0118096.
Артикул
Google ученый
Steinmetz, N.F., Mertens, M.E., Taurog, R.E., Johnson, J.E., Commandeur, U., Fischer, R. и Manchester, M., Картофельный вирус X как новая платформа для биомедицинских приложений, Nano Lett .
, 2010, том. 10, нет. 1, стр. 305–312 http://dx.doi.org/10.1021/nl53.Артикул
КАС
пабмед
ПабМед ЦентральныйGoogle ученый
Эсфандиари Н., Арзанани М.К., Сулеймани М., Кохи-Хабиби М. и Свендсен В.Е. Новое применение наночастиц растительного вируса для доставки лекарств при раке молочной железы, Tumor Biol. , 2016, том. 37, нет. 1. С. 1229–1236. http://dx.doi.org/10.1007/s13277-015-3867-3
Статья
КАС
пабмедGoogle ученый
Шукла, С., Аблэк, А.Л., Вен, А.М., Ли, К.Л., Льюис, Дж.Д., и Стейнмец, Н.Ф., Повышенное самонаведение опухоли и проникновение в ткани вирусной наночастицы нитевидного растения Вирус картофеля X, мол. Фармацевтика. , 2013, том. 10, нет. 1, стр. 33–42 http://dx.doi.org/10.1021/mp300240m.
Артикул
КАСGoogle ученый
Толлин, П., Бэнкрофт, Дж. Б., Ричардсон, Дж. Ф., Пейн, Н. С., и Беверидж, Т. Дж., Дифракционные исследования вируса мозаики папайи, Virology , 1979, vol. 98, нет. 1, стр. 108–115 http://dx.doi.org/10.1016/0042-6822(79)
-4.
Артикул
КАС
пабмедGoogle ученый
Сит, Т.Л., Абухайдар, М.Г., и Холи, С., Нуклеотидная последовательность РНК вируса мозаики папайи, J. Gen. Virol. , 1989, том. 70, нет. 9, стр. 2325–2331 http://dx.doi.org/10.1099/0022-1317-70-9-2325.
Артикул
КАС
пабмедGoogle ученый
Денис Дж., Акоста-Рамирес Э., Чжао Ю., Хамелен М.Э., Кукавица И., Баз М., Абед Ю., Савар К., Паре К., Лопес Масиас С., Бойвен Г. и Леклерк Д., Разработка универсальной вакцины против гриппа А на основе пептида М2е, слитого с печенью мышей вируса мозаики папайи (PapMV), J. Immunol. , 1957, том. 78, нет. 2, стр. 95–103.
Google ученый
Савар, К., Герен, А., Друэн, К., Болдук, М., Лалиберте-Гань, М.Е., Дюма, М.С., Мажо, Н., и Леклерк, Д., Улучшение трехвалентной инактивированной вакцина против гриппа с использованием наночастиц PapMV, PLoS One , 2011 г.
, том. 6, нет. 6, e21522 http://dx.doi.org/10.1371/journal.pone.0021522.Артикул
Google ученый
Дуглас, Т. и Янг, М., Инкапсуляция материалов хозяин-гость с помощью собранных вирусных белковых клеток, Nature , 1998, vol. 393, стр. 152–158 http://dx.doi.org/10.1038/30211.
Артикул
КАСGoogle ученый
Gillitzer, E., Willits, D., Young, M. и Douglas, T., Химическая модификация вирусной клетки для поливалентной презентации, Хим. коммун. , 2002, том. 21, стр. 2390–2391 http://dx.doi.org/10.1039/b207853h.
Артикул
Google ученый
Брамфилд С., Уиллитс Д., Танг Л., Джонсон Дж. Э., Дуглас Т. и Янг М. Гетерологическая экспрессия модифицированного белка оболочки хлоротической пятнистости бромовируса вигны приводит к сборке белка клетки с измененной архитектурой и функциями, J.
Gen. Virol. , 2004, том. 85, нет. 4, стр. 1049–1053 http://dx.doi.org/10.1099/vir.0.19688-0.Артикул
КАС
пабмедGoogle ученый
Hassani-Mehraban, A., Creutzburg, S., van Heereveld, L., and Kormelink, R., Возможность использования вирусоподобных частиц вигны хлоротической крапчатости в качестве каркаса для презентации эпитопов, BMC Biotechnol. , 2015, том. 15, с. 80 http://dx.doi.org/10.1186/s12896-015-0180-6.
Артикул
пабмед
ПабМед ЦентральныйGoogle ученый
Дестито, Г., Йе, Р., Рэй, К.С., Финн, М.Г., и Манчестер, М., Опосредованное фолиевой кислотой нацеливание частиц вируса мозаики вигны на опухолевые клетки, Chem. биол. , 2007, том. 14, нет. 10, стр. 1152–1162 http://dx.doi.org/10.1016/j.chembiol.2007.08.015.
Артикул
КАС
пабмед
ПабМед ЦентральныйGoogle ученый
McLain, L., Durrani, Z., Wisniewski, L.A., Porta, C., Lomonossoff, G.P., and Dimmock, N.J., Стимуляция нейтрализующих антител к вирусу иммунодефицита человека типа 1 у трех линий мышей, иммунизированных 22-аминокислотный пептид gp41, экспрессированный на поверхности растительного вируса, Вакцина , 1996, том. 14, нет. 8, стр. 799–810 http://dx.doi.org/10.1016/0264-410X(95)00229-T.
Артикул
КАС
пабмедGoogle ученый
Сингх, П., Прасун, Д., Йех, Р.М., Дестито, Г.
, Рэй, К.С., Осборн, К., Финн, М.Г., и Манчестер, М., Биораспределение, токсичность и патология коровьего гороха наночастицы вируса мозаики in vivo, J. Controlled Release , 2007, vol. 120, вып. 1–2, стр. 41–50 http://dx.doi.org/10.1016/j.jconrel.2007.04.003.КАС
Google ученый
Лин, Т., Чен, З., Уша, Р., Штауффахер, К., Дай, Дж., Шмидт, Т. и Джонсон, Дж., Уточненная кристаллическая структура вируса мозаики вигны при 2,8 Резолюция, Virology , 1999, vol. 265, нет. 1, стр. 20–34 http://dx.doi.org/10.1006/viro.1999.0038.
Артикул
КАС
пабмедGoogle ученый
Ломоносов Г. и Джонсон Дж. Синтез и структура капсидов комовирусов, стр. Прогр. Биофиз. Мол. биол. , 1991, том. 55, нет. 2, стр. 107–137 http://dx.doi.org/10.1016/0079-6107(91)
- -B.
Артикул
КАС
пабмедGoogle ученый
Рэй, К.С., Хор, И.В., Ван, К., Дестито, Г., Гонсалес, М.Дж., Сингх, П., Томас, Д.М., Эстрада, М.Н., Пауэлл, Э., Финн, М.Г., и Манчестер М., Системный перенос наночастиц растительного вируса у мышей пероральным путем, Вирусология , 2005, том. 343, нет. 2, стр. 224–235 http://dx.doi.org/10.1016/j.virol.2005.08.017.
Артикул
КАС
пабмедGoogle ученый
Кайзер, Ч.Р., Фленникен, М.Л., Гиллитцер, Э., Хармсен, А.Л., Хармсен, А.Г., Ютила, М.А., Дуглас, Т. и Янг, М.Дж., Исследования биораспределения наночастиц белковых клеток демонстрируют широкое распределение в тканях и быстрый клиренс in vivo, Междунар.
Дж. Наномед. , 2007, том. 2, нет. 4, стр. 715–733.КАС
Google ученый
 «>
«>Colson, P., Richet, H., Desnues, C., Balique, F., Moal, V., Grob, J.J., Berbis, P., Lecoq, H. ., Harle, J.R., Berland, Y. и Raoult, D., Вирус легкой крапчатости перца, , растительный вирус, связанный со специфическими иммунными реакциями, лихорадкой, болями в животе и зудом у людей, PLoS One , 2010, vol. 5, нет. 4, e10041 http://dx.doi.org/10.1371/journal.pone.0010041.
Google ученый
 5, нет. 5, e1000417 http://dx.doi.org/10.1371/journal.ppat.1000417.
5, нет. 5, e1000417 http://dx.doi.org/10.1371/journal.ppat.1000417. 2004.
2004. , Оценка токсичности вируса растений in vitro и in vivo наноносители, Colloids Surf., B , 2015, vol. 129, стр. 130–136 http://dx.doi.org/10.1016/j.colsurfb.2015.03.039.
, Оценка токсичности вируса растений in vitro и in vivo наноносители, Colloids Surf., B , 2015, vol. 129, стр. 130–136 http://dx.doi.org/10.1016/j.colsurfb.2015.03.039.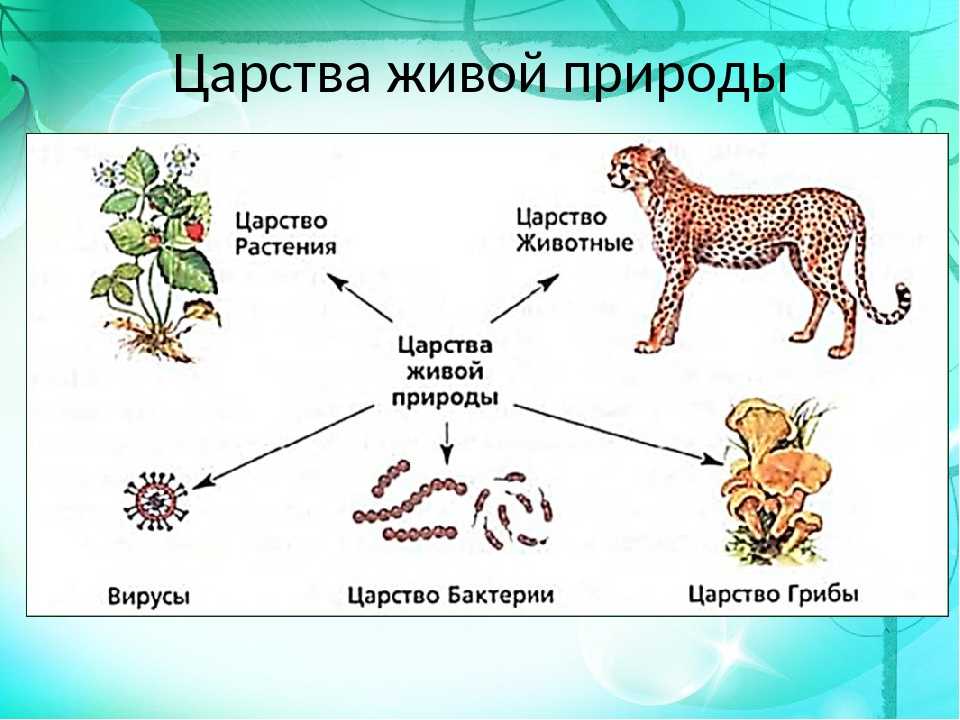
 «>
«>Каспар, Д.Л., Сборка и стабильность частиц вируса табачной мозаики, Adv. Белок хим. , 1963, том. 18, стр. 37–121 http://dx.doi.org/10.1016/S0065-3233(08)60268-5.
Артикул
КАС
пабмед
Google ученый
 Gen. Virol. , 2012, том. 93, нет. 2, стр. 400–407 http://dx.doi.org/10.1099/вир.0.036293-0.
Gen. Virol. , 2012, том. 93, нет. 2, стр. 400–407 http://dx.doi.org/10.1099/вир.0.036293-0. «>
«>Карасев А.В., Фоулке С., Велленс К., Рич А., Шон К.Дж., Звежиньски И., Хон Д., Копровски Х. и Рейц М., На основе растений Кандидат на вакцину против ВИЧ-1: белок Tat, полученный из шпината, Vaccine , 2005, vol. 23, нет. 15, стр. 1875–1880 http://dx.doi.org/10.1016/j.vaccine.2004.11.021.
Артикул
КАС
пабмед
Google ученый
 aeruginosa, Вакцина , 2000, том. 18, нет. 21, стр. 2266–2274 http://dx.doi.org/10.1016/S0264-410X(99)00571-X.
aeruginosa, Вакцина , 2000, том. 18, нет. 21, стр. 2266–2274 http://dx.doi.org/10.1016/S0264-410X(99)00571-X.
 «>
«>Wahyuni, W.S., Hanapi, M., и Hartana, I., Присутствие вируса табачной мозаики в экстракте компоста остатков сигарного табака, HAYATI J. Biosci. , 2008, том. 15, нет. 3, стр. 118–122.
Google ученый
 науч. , 2015, том. 52, нет. 3, стр. 667–670.
науч. , 2015, том. 52, нет. 3, стр. 667–670. «>
«>Антонисен, Н.Р., Коннетт, Дж.Е., Кайли, Дж.П., Альтос, М.Д., Бейли, В.К., и др., Влияние курения и использования ингаляционных антихолинергических бронходилататоров на скорость снижения ОФВ1. Исследование здоровья легких, J. Am. Мед. доц. , 1994, том. 272, нет. 19, стр. 1497–1505 http://dx.doi.org/10.1001/jama.1994.035201
 8, нет. 4, e60621 http://dx.doi.org/10.1371/journal.pone.0060621.
8, нет. 4, e60621 http://dx.doi.org/10.1371/journal.pone.0060621. . 5, нет. 6, e11298 http://dx.doi.org/10.1371/journal.pone.0011298.
. 5, нет. 6, e11298 http://dx.doi.org/10.1371/journal.pone.0011298. «>
«>Саломон, Р. и Бар-Джозеф, М., Трансляционная конкуренция между родственными видами вирусной РНК в бесклеточных системах, Дж. Ген. Вирол. , 1982, том. 62, нет. 2, стр. 343–347 http://dx.doi.org/10.1099/0022-1317-62-2-343.
Артикул
КАС
пабмед
Google ученый
 1719–1726 http://dx.doi.org/10.1093/nar/2.10.1719.
1719–1726 http://dx.doi.org/10.1093/nar/2.10.1719. «>
«>Никитин Н., Малинин А., Рахнянская А., Трифонова Е., Карпова О., Ярославов А., Атабеков Ю. Использование поликатионного спейсера для нековалентной иммобилизации альбумина на термомодифицированных вирусных частицах, Полим. наук, сер. А , 2011, том. 53, нет. 11, стр. 1026–1031 http://dx.doi.org/10.1134/S0965545С11110083.
Артикул
КАС
Google ученый
 Вирол. , 2001, том. 75, нет. 18, стр. 8434–8439 http://dx.doi.org/10.1128/JVI.75.18.8434-8439.2001.
Вирол. , 2001, том. 75, нет. 18, стр. 8434–8439 http://dx.doi.org/10.1128/JVI.75.18.8434-8439.2001. «>
«>Лико С., Бенвенуто Э. и Баскьери С. Двуликий картофельный вирус X: от патогена растений до умных наночастиц, Front. Растениевод. , 2015, том. 6, с. 1009 http://dx.doi.org/10.3389/fpls.2015.01009.
Артикул
пабмед
ПабМед Центральный
Google ученый
 , 2010, том. 10, нет. 1, стр. 305–312 http://dx.doi.org/10.1021/nl53.
, 2010, том. 10, нет. 1, стр. 305–312 http://dx.doi.org/10.1021/nl53. «>
«>Лебель, М.Э., Шартран, К., Леклерк, Д. и Ламарр, А., Вирусы растений как вакцины и адъюванты на основе наночастиц, Вакцины , 2015, том. 3, нет. 3, стр. 620–637 http://dx.doi.org/10.3390/vaccines3030620.
Артикул
пабмед
ПабМед Центральный
Google ученый
 «>
«>Zhang, H., Todderud, E., и Stubbs, G., Кристаллизация и предварительный рентгеновский анализ белка оболочки вируса мозаики папайи, J. Mol. биол. , 1993, том. 234, нет. 3, стр. 885–887 http://dx.doi.org/10.1006/jmbi.1993.1634.
Артикул
КАС
пабмед
Google ученый
 , том. 6, нет. 6, e21522 http://dx.doi.org/10.1371/journal.pone.0021522.
, том. 6, нет. 6, e21522 http://dx.doi.org/10.1371/journal.pone.0021522.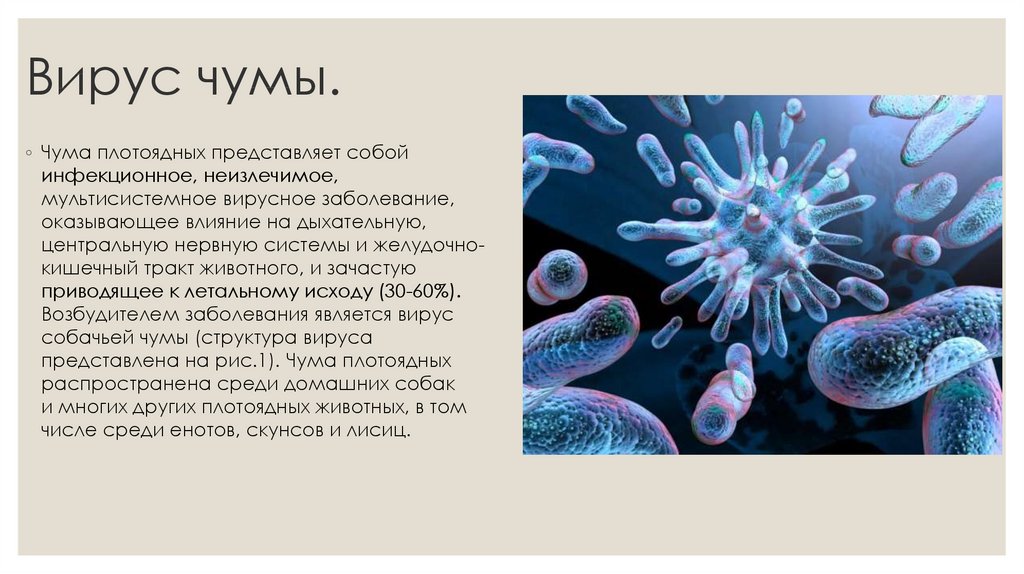 Gen. Virol. , 2004, том. 85, нет. 4, стр. 1049–1053 http://dx.doi.org/10.1099/vir.0.19688-0.
Gen. Virol. , 2004, том. 85, нет. 4, стр. 1049–1053 http://dx.doi.org/10.1099/vir.0.19688-0. «>
«>Льюис, Дж. Д., Дестито, Г., Зейлстра, А., Гонсалес, М. Дж., Куигли, Дж. П., Манчестер, М., и Стулманн, Х., Вирусные наночастицы как инструменты для прижизненной визуализации сосудов, Нац. Мед. , 2006, том. 12, нет. 3, стр. 354–360 http://dx.doi.org/10.1038/nm1368.
Артикул
КАС
пабмед
ПабМед Центральный
Google ученый
 , Рэй, К.С., Осборн, К., Финн, М.Г., и Манчестер, М., Биораспределение, токсичность и патология коровьего гороха наночастицы вируса мозаики in vivo, J. Controlled Release , 2007, vol. 120, вып. 1–2, стр. 41–50 http://dx.doi.org/10.1016/j.jconrel.2007.04.003.
, Рэй, К.С., Осборн, К., Финн, М.Г., и Манчестер, М., Биораспределение, токсичность и патология коровьего гороха наночастицы вируса мозаики in vivo, J. Controlled Release , 2007, vol. 120, вып. 1–2, стр. 41–50 http://dx.doi.org/10.1016/j.jconrel.2007.04.003. «>
«>Wang, Q., Kaltgrad, E., Lin, T., Johnson, J. и Finn, M., Природные надмолекулярные строительные блоки: вирус мозаики вигны дикого типа, Chem. биол. , 2002, том. 9, нет. 7, стр. 805–811 http://dx.doi.org/10.1016/S1074-5521(02)00165-5.
Артикул
КАС
пабмед
Google ученый
 Дж. Наномед. , 2007, том. 2, нет. 4, стр. 715–733.
Дж. Наномед. , 2007, том. 2, нет. 4, стр. 715–733.Скачать ссылки
Различные носители и их вирусы
Результаты обучения
- Объяснить передачу и болезни вирусов, поражающих животных
- Объясните передачу и болезни вирусов, поражающих растения
Как вы уже знаете, вирусы часто заражают очень конкретных носителей, а также определенные клетки внутри хозяина. Эта особенность вируса делает его специфичным для одного или нескольких видов жизни на Земле. С другой стороны, на Земле существует так много различных типов вирусов, что почти у каждого живого организма есть свой набор из 9 вирусов.0601 вирусы пытаются заразить его клетки. Даже прокариоты, самые маленькие и простые клетки, могут быть атакованы определенными типами вирусов. В следующем разделе мы рассмотрим некоторые особенности вирусной инфекции прокариотических клеток. Как мы узнали, вирусы, поражающие бактерии, называются бактериофагами (рис. 1). У архей есть свои похожие вирусы.
1). У архей есть свои похожие вирусы.
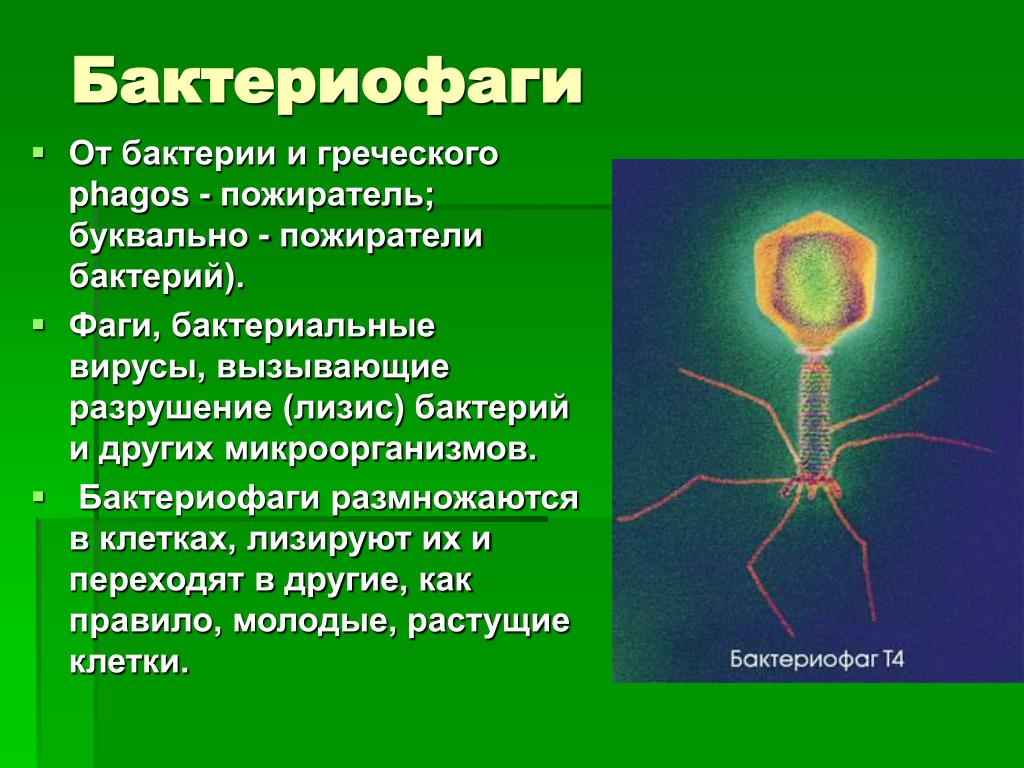
Бактериофаги
Рис. 1. Бактериофаги, прикрепленные к клетке-хозяину (электронная микрофотография). У бактериофага с хвостами, подобного показанному здесь, хвосты служат проходом для передачи генома фага. (кредит: модификация работы доктора Грэма Бердса; данные масштабной линейки от Мэтта Рассела)
Большинство бактериофагов являются вирусами двухцепочечной ДНК, которые используют ферменты хозяина для репликации ДНК и транскрипции РНК. Фаговые частицы должны связываться со специфическими поверхностными рецепторами и активно внедрять геном в клетку-хозяина. (Сложные хвостовые структуры, наблюдаемые у многих бактериофагов, активно участвуют в переносе вирусного генома через стенку прокариотической клетки.) Когда инфицирование клетки бактериофагом приводит к образованию новых вирионов, говорят, что инфекция является продуктивной . Если вирионы высвобождаются путем разрыва клетки, вирус реплицируется с помощью литического цикла (рис. 2). Примером литического бактериофага является T4, который инфицирует Escherichia coli , обнаруженную в желудочно-кишечном тракте человека. Однако иногда вирус может оставаться внутри клетки, не высвобождаясь. Например, когда умеренный бактериофаг заражает бактериальную клетку, он реплицируется с помощью лизогенного цикла (рис. 2), и геном вируса включается в геном клетки-хозяина. Когда ДНК фага встраивается в геном клетки-хозяина, это называется профаг . Примером лизогенного бактериофага является вирус λ (лямбда), который также поражает E . coli бактерия. Вирусы, поражающие клетки растений или животных, могут иногда подвергаться инфекциям, когда они не продуцируют вирионы в течение длительного времени. Примером могут служить вирусы герпеса животных , включая вирусы простого герпеса, вызывающие оральный и генитальный герпес у людей.
2). Примером литического бактериофага является T4, который инфицирует Escherichia coli , обнаруженную в желудочно-кишечном тракте человека. Однако иногда вирус может оставаться внутри клетки, не высвобождаясь. Например, когда умеренный бактериофаг заражает бактериальную клетку, он реплицируется с помощью лизогенного цикла (рис. 2), и геном вируса включается в геном клетки-хозяина. Когда ДНК фага встраивается в геном клетки-хозяина, это называется профаг . Примером лизогенного бактериофага является вирус λ (лямбда), который также поражает E . coli бактерия. Вирусы, поражающие клетки растений или животных, могут иногда подвергаться инфекциям, когда они не продуцируют вирионы в течение длительного времени. Примером могут служить вирусы герпеса животных , включая вирусы простого герпеса, вызывающие оральный и генитальный герпес у людей.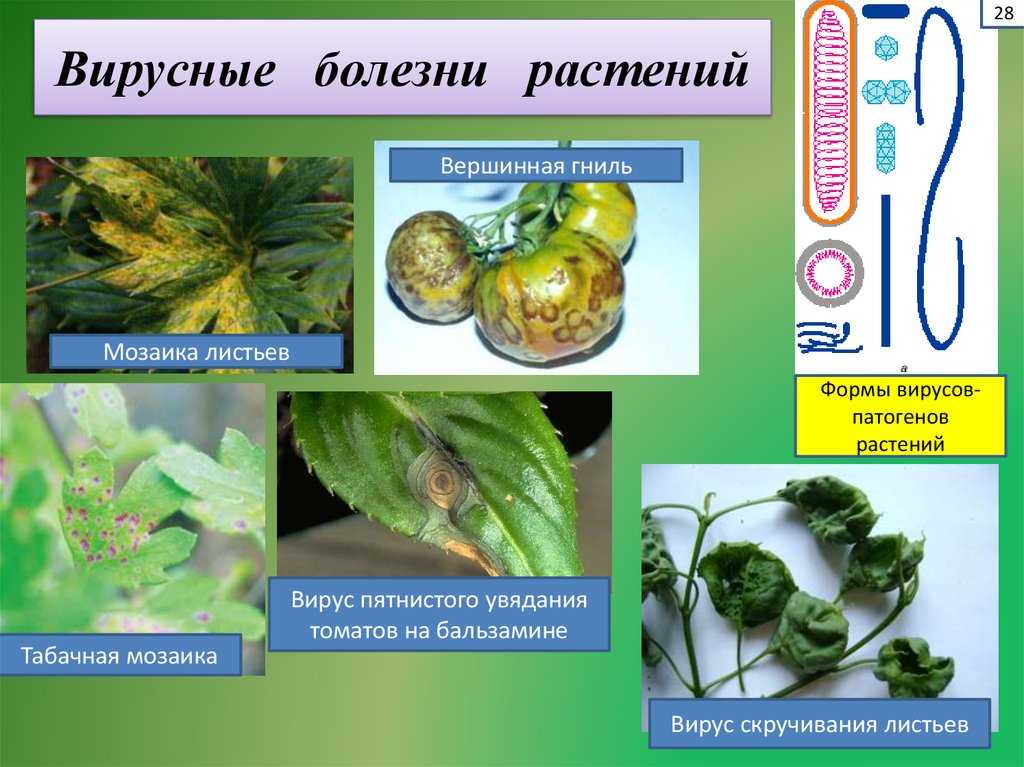 В процессе, называемом латентность , эти вирусы могут существовать в нервной ткани в течение длительных периодов времени, не производя новых вирионов, только для того, чтобы периодически выходить из латентного состояния и вызывать поражения кожи, где вирус реплицируется. Несмотря на то, что между лизогенией и латентностью есть сходство, термин лизогенный цикл обычно используется для описания бактериофагов. Задержка будет описана более подробно в следующем разделе.
В процессе, называемом латентность , эти вирусы могут существовать в нервной ткани в течение длительных периодов времени, не производя новых вирионов, только для того, чтобы периодически выходить из латентного состояния и вызывать поражения кожи, где вирус реплицируется. Несмотря на то, что между лизогенией и латентностью есть сходство, термин лизогенный цикл обычно используется для описания бактериофагов. Задержка будет описана более подробно в следующем разделе.
Рисунок 2. Бактериофаг умеренного пояса имеет как литический, так и лизогенный циклы. В литическом цикле фаг реплицируется и лизирует клетку-хозяина. В лизогенном цикле ДНК фага включается в геном хозяина, где передается последующим поколениям. Стрессовые факторы окружающей среды, такие как голодание или воздействие токсичных химических веществ, могут привести к вырезанию профага и его включению в литический цикл.
Практический вопрос
Какое из следующих утверждений неверно?
- В литическом цикле образуются новые фаги, которые высвобождаются в окружающую среду.
- В лизогенном цикле ДНК фага включается в геном хозяина.
- Фактор окружающей среды может привести к тому, что фаг инициирует лизогенный цикл.
- Лизис клеток происходит только в литическом цикле.

Показать ответ
Вирусы животных
Вирусы животных, в отличие от вирусов растений и бактерий, не должны проникать через клеточную стенку, чтобы получить доступ к клетке-хозяину. Вирус может даже побудить клетку-хозяина сотрудничать в процессе заражения. Безоболочечные или «голые» вирусы животных могут проникать в клетки двумя разными путями. Поскольку белок в вирусном капсиде связывается со своим рецептором на клетке-хозяине, вирус может попасть внутрь клетки через везикулу во время нормального клеточного процесса рецептор-опосредованный эндоцитоз . Альтернативный метод проникновения в клетку, используемый безоболочечными вирусами, заключается в том, что капсидные белки претерпевают изменения формы после связывания с рецептором, создавая каналы в мембране клетки-хозяина. Затем вирусный геном «инъецируется» в клетку-хозяина через эти каналы способом, аналогичным тому, который используется многими бактериофагами.
Затем вирусный геном «инъецируется» в клетку-хозяина через эти каналы способом, аналогичным тому, который используется многими бактериофагами.
Вирусы с оболочкой также имеют два пути проникновения в клетки после связывания с их рецепторами: опосредованный рецептором эндоцитоз или слияние . Многие вирусы с оболочкой проникают в клетку посредством эндоцитоза, опосредованного рецептором, подобно тому, как это наблюдается у некоторых безоболочечных вирусов. С другой стороны, слияние происходит только с оболочечными вирионами. Эти вирусы, в том числе ВИЧ, используют в своей оболочке специальные белки слияния, чтобы заставить оболочку слиться с плазматической мембраной клетки, тем самым высвобождая геном и капсид вируса в цитоплазму клетки.
Создав свои белки и скопировав свои геномы, вирусы животных завершают сборку новых вирионов и покидают клетку. Как мы уже обсуждали на примере вируса гриппа, оболочечные вирусы животных могут отпочковываться от клеточной мембраны по мере того, как они собираются, захватывая при этом часть клеточной плазматической мембраны. С другой стороны, безоболочечное вирусное потомство, такое как риновирусы, накапливается в инфицированных клетках до тех пор, пока не появится сигнал для лизиса или апоптоза, и все вирионы высвобождаются вместе.
С другой стороны, безоболочечное вирусное потомство, такое как риновирусы, накапливается в инфицированных клетках до тех пор, пока не появится сигнал для лизиса или апоптоза, и все вирионы высвобождаются вместе.
Как вы узнаете из следующего модуля, вирусы животных связаны с различными заболеваниями человека. Некоторые из них следуют классической модели острого заболевания , когда симптомы ухудшаются в течение короткого периода времени, после чего вирус удаляется из организма иммунной системой и в конечном итоге излечивается от инфекции. Примерами острых вирусных заболеваний являются простуда и грипп. Другие вирусы вызывают длительные хронические инфекции , например вирус, вызывающий гепатит С, тогда как другие, такие как вирус простого герпеса, вызывают только интермиттирующие симптомы. Тем не менее, другие вирусы, такие как вирусы герпеса человека 6 и 7, которые в некоторых случаях могут вызывать легкую детскую болезнь розеолу, часто успешно вызывают продуктивные инфекции, не вызывая вообще никаких симптомов у хозяина, и поэтому мы говорим, что у этих пациентов бессимптомное заболевание. инфекция .
инфекция .
При инфекциях гепатита С вирус растет и размножается в клетках печени, вызывая незначительные повреждения печени. Ущерб настолько низок, что инфицированные люди часто не подозревают о том, что они инфицированы, и многие инфекции обнаруживаются только при рутинном анализе крови у пациентов с такими факторами риска, как внутривенное употребление наркотиков. С другой стороны, поскольку многие симптомы вирусных заболеваний вызваны иммунными ответами, отсутствие симптомов является признаком слабого иммунного ответа на вирус. Это позволяет вирусу избежать элиминации иммунной системой и сохраняться у людей в течение многих лет, при этом производя низкие уровни потомства вирионов при так называемом хроническом вирусном заболевании. Хроническая инфекция печени этим вирусом приводит к гораздо большей вероятности развития рака печени, иногда даже через 30 лет после первоначальной инфекции.
Как уже говорилось, вирус простого герпеса может оставаться в латентном состоянии в нервной ткани месяцами и даже годами. Поскольку вирус «прячется» в ткани и производит мало вирусных белков, если вообще производит их, иммунному ответу не на что воздействовать, и иммунитет к вирусу постепенно снижается. При определенных условиях, включая различные виды физического и психологического стресса, латентный вирус простого герпеса может реактивироваться и подвергаться литическому циклу репликации в коже, вызывая поражения, связанные с заболеванием. Как только в коже образуются вирионы и синтезируются вирусные белки, иммунный ответ снова стимулируется и устраняет кожные поражения за несколько дней или недель путем уничтожения вирусов в коже. В результате этого типа репликативного цикла появление герпеса и вспышек генитального герпеса происходят только периодически, даже если вирусы остаются в нервной ткани на всю жизнь. Скрытые инфекции распространены и при других герпесвирусах, включая вирус ветряной оспы, вызывающий ветряную оспу. После перенесенной ветряной оспы в детстве вирус ветряной оспы может оставаться латентным в течение многих лет и реактивироваться у взрослых, вызывая болезненное состояние, известное как «опоясывающий лишай» (рис.
Поскольку вирус «прячется» в ткани и производит мало вирусных белков, если вообще производит их, иммунному ответу не на что воздействовать, и иммунитет к вирусу постепенно снижается. При определенных условиях, включая различные виды физического и психологического стресса, латентный вирус простого герпеса может реактивироваться и подвергаться литическому циклу репликации в коже, вызывая поражения, связанные с заболеванием. Как только в коже образуются вирионы и синтезируются вирусные белки, иммунный ответ снова стимулируется и устраняет кожные поражения за несколько дней или недель путем уничтожения вирусов в коже. В результате этого типа репликативного цикла появление герпеса и вспышек генитального герпеса происходят только периодически, даже если вирусы остаются в нервной ткани на всю жизнь. Скрытые инфекции распространены и при других герпесвирусах, включая вирус ветряной оспы, вызывающий ветряную оспу. После перенесенной ветряной оспы в детстве вирус ветряной оспы может оставаться латентным в течение многих лет и реактивироваться у взрослых, вызывая болезненное состояние, известное как «опоясывающий лишай» (рис. 3).
3).
Рисунок 3. (a) Varicella-zoster, вирус, вызывающий ветряную оспу, имеет икосаэдрический капсид в оболочке, видимый на этой трансмиссионной электронной микрофотографии. Его двухцепочечный геном ДНК включается в ДНК хозяина и может повторно активироваться после латентного периода в форме (b) опоясывающего лишая, часто проявляющегося сыпью. (кредит a: модификация работы доктора Эрскина Палмера, B.G. Martin, CDC; кредит b: модификация работы «rosmary»/Flickr; данные масштабной линейки от Matt Russell)
Рисунок 4. ВПЧ или вирус папилломы человека ( кредит: модификация работы NCI, NIH; данные масштабной линейки от Matt Russell)
Некоторые вирусы, заражающие животных, включая вирус гепатита С, описанный выше, известны как онкогенные вирусы: они способны вызывать рак. Эти вирусы нарушают нормальную регуляцию клеточного цикла хозяина либо путем введения генов, стимулирующих нерегулируемый рост клеток (онкогены), либо путем вмешательства в экспрессию генов, подавляющих рост клеток. Онкогенные вирусы могут быть ДНК- или РНК-вирусами.
Онкогенные вирусы могут быть ДНК- или РНК-вирусами.
Известные виды рака, связанные с вирусными инфекциями, включают рак шейки матки, вызванный вирусом папилломы человека (ВПЧ), рак печени, вызванный вирусом гепатита В, Т-клеточный лейкоз и несколько типов лимфомы.
ВПЧ, или вирус папилломы человека (как показано на рис. 4), имеет икосаэдрический капсид, видимый на этой трансмиссионной электронной микрофотографии, и двухцепочечный ДНК-геном, встроенный в ДНК хозяина. Вирус, передающийся половым путем, является онкогенным и может привести к раку шейки матки.
Посетите интерактивные анимации, показывающие различные этапы репликативных циклов вирусов животных, и щелкните ссылки на флэш-анимации.
Вирусы растений
Большинство вирусов растений, таких как вирус табачной мозаики, имеют геном одноцепочечной (+) РНК. Однако в большинстве других категорий вирусов также есть растительные вирусы. В отличие от бактериофагов вирусы растений не имеют активных механизмов доставки вирусного генома через защитную клеточную стенку. Чтобы растительный вирус проник в новое растение-хозяин, должно произойти какое-то механическое повреждение. Этот ущерб часто вызван погодой, насекомыми, животными, пожаром или деятельностью человека, такой как сельское хозяйство или ландшафтный дизайн. Перемещению от клетки к клетке внутри растения может способствовать вирусная модификация плазмодесм (цитоплазматических нитей, переходящих от одной растительной клетки к другой). Кроме того, потомки растений могут наследовать вирусные заболевания от родительских растений. Вирусы растений могут передаваться различными переносчиками, через контакт с соком инфицированного растения, через живые организмы, такие как насекомые и нематоды, а также через пыльцу. Перенос вируса с одного растения на другое известен как горизонтальная передача , тогда как наследование вируса от родителя называется вертикальной передачей .
Чтобы растительный вирус проник в новое растение-хозяин, должно произойти какое-то механическое повреждение. Этот ущерб часто вызван погодой, насекомыми, животными, пожаром или деятельностью человека, такой как сельское хозяйство или ландшафтный дизайн. Перемещению от клетки к клетке внутри растения может способствовать вирусная модификация плазмодесм (цитоплазматических нитей, переходящих от одной растительной клетки к другой). Кроме того, потомки растений могут наследовать вирусные заболевания от родительских растений. Вирусы растений могут передаваться различными переносчиками, через контакт с соком инфицированного растения, через живые организмы, такие как насекомые и нематоды, а также через пыльцу. Перенос вируса с одного растения на другое известен как горизонтальная передача , тогда как наследование вируса от родителя называется вертикальной передачей .
Симптомы вирусных заболеваний различаются в зависимости от вируса и его хозяина (см. таблицу ниже). Одним из распространенных симптомов является гиперплазия , аномальная пролиферация клеток, вызывающая появление опухолей растений, известных как галлы . Другие вирусы вызывают гипоплазию или уменьшение роста клеток в листьях растений, вызывая появление тонких желтых участков. Другие вирусы поражают растение, непосредственно убивая растительные клетки, процесс, известный как 9.0701 некроз клеток . Другие симптомы вирусов растений включают уродливые листья, черные полосы на стеблях растений, изменение роста стеблей, листьев или плодов и кольцевые пятна, которые представляют собой круглые или линейные области обесцвечивания на листе.
таблицу ниже). Одним из распространенных симптомов является гиперплазия , аномальная пролиферация клеток, вызывающая появление опухолей растений, известных как галлы . Другие вирусы вызывают гипоплазию или уменьшение роста клеток в листьях растений, вызывая появление тонких желтых участков. Другие вирусы поражают растение, непосредственно убивая растительные клетки, процесс, известный как 9.0701 некроз клеток . Другие симптомы вирусов растений включают уродливые листья, черные полосы на стеблях растений, изменение роста стеблей, листьев или плодов и кольцевые пятна, которые представляют собой круглые или линейные области обесцвечивания на листе.
| Таблица 1. Некоторые общие симптомы вирусных заболеваний растений | |
|---|---|
| Симптом | Выглядит как |
| Гиперплазия | Галлы (опухоли) |
| Гипоплазия | Истонченные желтые пятна на листьях |
| Некроз клеток | Мертвые, почерневшие стебли, листья или плоды |
| Аномальные модели роста | Деформированные стебли, листья или плоды |
| Обесцвечивание | Желтые, красные или черные полосы или кольца на стеблях, листьях или плодах |
Вирусы растений могут серьезно нарушать рост и развитие сельскохозяйственных культур, существенно влияя на наши запасы продовольствия.