
Содержание
биофак СПбГУ
биофак СПбГУ
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Главная / учебные материалы
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
 gif»>
gif»>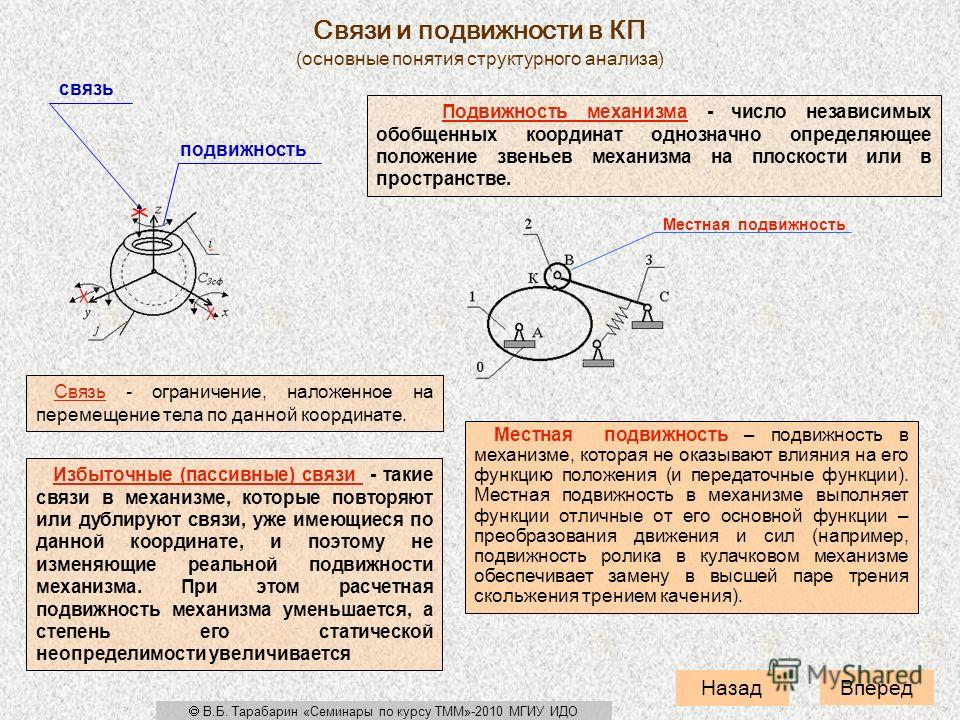 gif»>
gif»>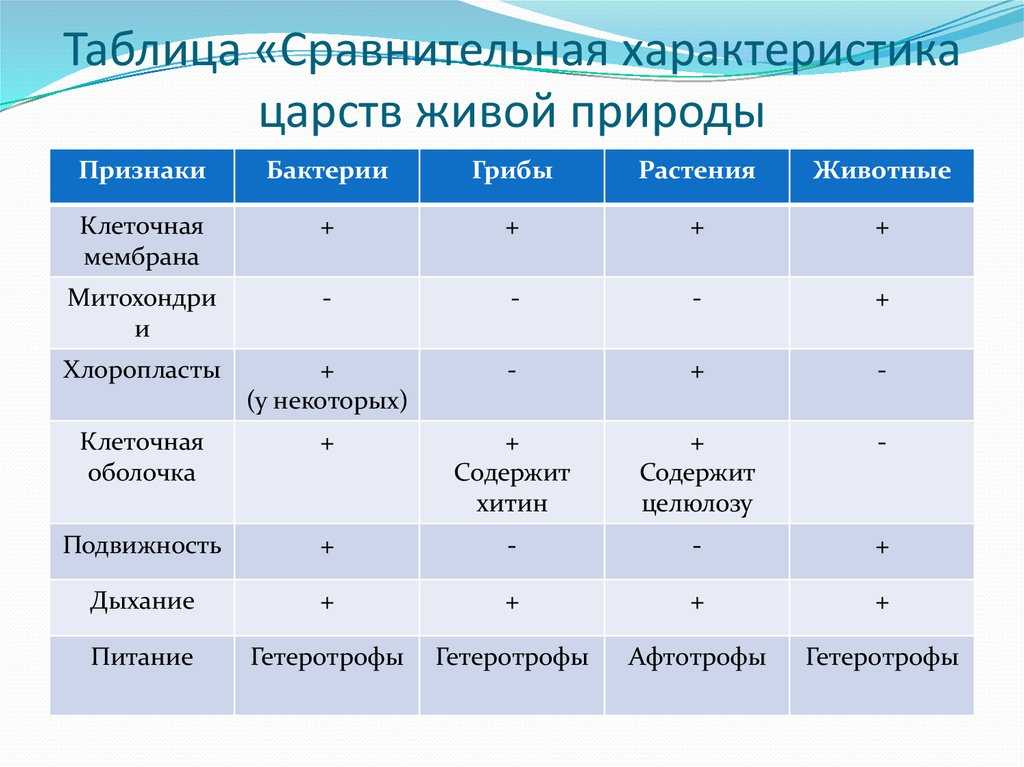 gif»>
gif»> gif»>
gif»> gif»>
gif»> gif»>
gif»> gif»>
gif»> gif»>
gif»> gif»>
gif»> gif»>
gif»> gif»>
gif»> gif»>
gif»> gif»>
gif»>контакты
карта сайта почтовый сервер
управление поддержка
199034, Санкт-Петербург, Университетская наб., 7-9
© Санкт-Петербургский государственный университет, 2006-2017
Микроэлементы и их роль в жизни растений
23.07.2021
Интенсивное земледелие способствует значительному изменению химического состава почв. За счет вывоза большей части сельскохозяйственной продукции за пределы агроценоза из биологического круговорота выключается заметная масса химических элементов. Важную роль в получении высоких и стабильных урожаев сельскохозяйственных культур играют микроэлементы. Изучение их содержания, распределения и подвижности необходимо для агроэкологической оценки состояния плодородия почв и оптимизации минерального питания растений.
Микроэлементами называются такие элементы минерального питания растений и животных, которые необходимы живому организму лишь в очень небольших количествах. К ним относятся бор, молибден, медь, марганец, кобальт, цинк, йод и другие. Микроэлементы, жизненно важные для растений, это такие, которые не могут быть заменены другими элементами в их специфической биохимической роли и которые имеют прямое влияние на организм, то есть без них он не может ни расти, ни завершить некоторые метаболические циклы.
К ним относятся бор, молибден, медь, марганец, кобальт, цинк, йод и другие. Микроэлементы, жизненно важные для растений, это такие, которые не могут быть заменены другими элементами в их специфической биохимической роли и которые имеют прямое влияние на организм, то есть без них он не может ни расти, ни завершить некоторые метаболические циклы.
Учение о микроэлементах было создано трудами многих отечественных и зарубежных ученых (В.И. Вернадский, А.П. Виноградов, К.К. Гедройц, Д.Н. Прянишников, В.В. Ковальский, В.А.Ковда, Р. Митчел, А. Пейлж, Дж. Ходсон, Н.Г. Зырин).
Микроэлементы поступают в почву различными способами, в том числе: осаждаясь из атмосферы, при выщелачивании, при разложении надземных частей растений, применении средств химической обработки, внесении удобрений. Формы нахождения микроэлементов в почвенном профиле зависят от их химических форм, унаследованных от материнской породы.
Обеспеченность почвы микроэлементами меняется в течение вегетационного периода, а также зависит от интенсивности осадков, испарения влаги из почвы и т. д. В зависимости от этих факторов, концентрации микроэлементов в почвенных растворах могут изменяться более чем в 10 раз. Это необходимо учитывать при проведении анализов почвы. При этом концентрации макроэлементов, хотя так же зависят от упомянутых факторов, изменяются в меньшей степени.
д. В зависимости от этих факторов, концентрации микроэлементов в почвенных растворах могут изменяться более чем в 10 раз. Это необходимо учитывать при проведении анализов почвы. При этом концентрации макроэлементов, хотя так же зависят от упомянутых факторов, изменяются в меньшей степени.
В почвах Ростовской области редко проявляется дефицит таких микроэлементов, как марганца и меди, причем степень обеспеченности марганцем, как правило, высокая > 20 мг/кг, медью – средняя – 0,21-0,50 мг/кг. Степень обеспеченности почв кобальтом и цинком низкая, соответственно < 0,15 и 2,0 мг/кг.
В течение вегетационного периода растения потребность и степень усваиваемости одного и того же элемента может сильно отличаться. Поскольку все растительные организмы обладают избирательной способностью, то поглощение веществ, необходимых на данном этапе их развития, происходит более активно, чем всех остальных.
Для некоторых типов почв изначально характерен недостаток какого-либо из элементов. Так, плодородные, богатые органикой грунты отличаются низким содержанием в них меди, на дерновых почвах растения будут ощущать острую нехватку молибдена, в щелочных грунтах снижается доступность бора, меди, марганца, а в кислых отмечается повышенная усваиваемость марганца, который в больших количествах токсичен для растений. Принимая во внимание такие особенности, в каждом конкретном случае необходимо учитывать потребности каждого вида растений в конкретном микроэлементе, поскольку для разных культур они могут сильно отличаться. Основной недостаток микроэлементов, находящихся в почве – их труднодоступность для потребления растениями. Поэтому, чтобы восполнить недостающие в питании растений микроэлементы, применяют корневые либо внекорневые подкормки культур микроудобрениями.
Так, плодородные, богатые органикой грунты отличаются низким содержанием в них меди, на дерновых почвах растения будут ощущать острую нехватку молибдена, в щелочных грунтах снижается доступность бора, меди, марганца, а в кислых отмечается повышенная усваиваемость марганца, который в больших количествах токсичен для растений. Принимая во внимание такие особенности, в каждом конкретном случае необходимо учитывать потребности каждого вида растений в конкретном микроэлементе, поскольку для разных культур они могут сильно отличаться. Основной недостаток микроэлементов, находящихся в почве – их труднодоступность для потребления растениями. Поэтому, чтобы восполнить недостающие в питании растений микроэлементы, применяют корневые либо внекорневые подкормки культур микроудобрениями.
Материал подготовлен специалистами отдела защиты растений и агрохимии ФГБУ «Ростовский референтный центр Россельхознадзора».
- Предыдущая новость
- Возврат к списку
- Следующая новость
животных | Определение, типы и факты
животное
Посмотреть все СМИ
- Ключевые люди:
- Чарльз Элтон
Карл Э. Экли
Экли
Спенсер Фуллертон Бэрд
Карл П. Шмидт
Росс Грэнвилл Харрисон
- Похожие темы:
- социальное поведение животных
рептилия
рыбы
поведение животных
насекомое
Просмотреть весь связанный контент →
Популярные вопросы
Что такое животное?
Животные — это многоклеточные эукариоты, клетки которых связаны между собой коллагеном. Животные доминируют в человеческих представлениях о жизни на Земле из-за их размера, разнообразия, изобилия и мобильности. Наличие мышц и подвижность — одна из основных характеристик животного мира.
Каковы две основные группы животных?
Двумя основными группами животных являются позвоночные и беспозвоночные. У позвоночных есть позвоночник, а у беспозвоночных его нет.
Когда впервые появились животные?
Животные впервые появились в эдиакарский период, примерно от 635 до 541 миллионов лет назад, в виде мягкотелых форм, оставивших следы своего тела в мелководных отложениях.
Каковы основные функциональные системы животных?
Основные функциональные системы животных включают костно-мышечную систему, обеспечивающую поддержку и движение тела; нервная система для приема и обработки сенсорной информации и передачи сигналов для управления мышечной и гормональной активностью; эндокринная система для секреции гормонов для химического контроля функций организма; пищеварительная система для приема и переработки пищи; кровеносная система для доставки питательных веществ и кислорода к клеткам и выведения их отходов; и репродуктивная система для производства потомства, обеспечивающего выживание вида.
Чем животные отличаются от растений и грибов?
Животные отличаются от растений и грибов фундаментальными различиями в морфологии и физиологии, особенно в отношении развития мышц и подвижности животных.
Сводка
Прочтите краткий обзор этой темы
Посмотрите бой сурикатов и посмотрите, как доминирующая альфа-самка суриката изгоняет подчиненного из стаи
Посмотреть все видео к этой статье
животное , (царство Animalia), любой из группы многоклеточных эукариотических организмов (т. е. в отличие от бактерий их дезоксирибонуклеиновая кислота, или ДНК, содержится в мембраносвязанном ядре). Считается, что они произошли независимо от одноклеточных эукариот. Животные отличаются от представителей двух других царств многоклеточных эукариот, растений (Plantae) и грибов (Mycota), фундаментальными различиями в морфологии и физиологии. Во многом это связано с тем, что у животных развиты мышцы и, следовательно, подвижность, характеристика, которая стимулировала дальнейшее развитие тканей и систем органов.
Животные доминируют в представлениях людей о жизни на Земле не только из-за их размера, численности и явного разнообразия, но и из-за их подвижности, общей для людей черты. Движение к представлениям о животных настолько неотъемлемо, что губки, лишенные мышечной ткани, долгое время считались растениями. Только после того, как в 1765 году были замечены их небольшие движения, животная природа губок стала постепенно признаваться.
По своим размерам животные на суше уступают растениям, среди листвы которых они часто прячутся. Напротив, фотосинтезирующие водоросли, которыми питаются открытые океаны, обычно слишком малы, чтобы их можно было увидеть, но морские животные размером с китов. Разнообразие форм, в отличие от размера, лишь периферически воздействует на человеческое осознание жизни и, следовательно, менее заметно. Тем не менее, животные составляют три четверти или более видов на Земле, и это разнообразие отражает гибкость в питании, защите и размножении, которую им дает мобильность. Животные следуют практически всем известным образам жизни, описанным для земных существ.
Животные перемещаются в поисках пищи, партнеров или укрытия от хищников, и это движение привлекает внимание и интерес, особенно когда становится очевидным, что поведение некоторых существ не так уж сильно отличается от поведения человека. Помимо простого любопытства, люди изучают животных, чтобы узнать о себе, которые являются совсем недавним продуктом эволюции животных.
Викторина «Британника»
Брачное поведение животных
Что вы знаете о брачном поведении животных? Пройдите этот тест и узнайте.
Животные произошли от одноклеточных эукариот. Наличие ядерной мембраны у эукариот позволяет разделить две фазы синтеза белка: транскрипцию (копирование) дезоксирибонуклеиновой кислоты (ДНК) в ядре и трансляцию (декодирование) сообщения в белок в цитоплазме. По сравнению со структурой бактериальной клетки это дает больший контроль над тем, какие белки вырабатываются. Такой контроль позволяет специализировать клетки, каждая из которых имеет идентичную ДНК, но с возможностью точно контролировать, какие гены успешно отправляют копии в цитоплазму. Таким образом, ткани и органы могут развиваться. Полужесткие клеточные стенки растений и грибов, ограничивающие форму и, следовательно, разнообразие возможных типов клеток, отсутствуют у животных. Если бы они присутствовали, нервные и мышечные клетки, центр подвижности животных, были бы невозможны.
Функциональное разнообразие растений и животных способствует сборке мутуалистической сети на градиенте высот
1. Thompson JN. Совместно развивающаяся паутина жизни. Являюсь. Нац. 2009; 173:125–140. дои: 10.1086/595752. [PubMed] [CrossRef] [Google Scholar]
2. Bascompte J, Jordano P. Мутуалистические сети растений и животных: архитектура биоразнообразия. Анну. Преподобный Экол. Эвол. Сист. 2007; 38: 567–593. doi: 10.1146/annurev.ecolsys.38.091206.095818. [CrossRef] [Google Scholar]
3. Кремень С. Управление экосистемными услугами: что нам нужно знать об их экологии? Экол. лат. 2005; 8: 468–479.. doi: 10.1111/j.1461-0248.2005.00751.x. [PubMed] [CrossRef] [Google Scholar]
4. Васкес Д.П., Блютген Н., Каньоло Л., Чакофф Н.П. Объединяющий паттерн и процесс в мутуалистических сетях растений и животных: обзор. Анна. Бот. 2009; 103:1445–1457. doi: 10.1093/aob/mcp057. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
5. Hillebrand H, Mattiessen B. Биоразнообразие в сложном мире: консолидация и прогресс в исследованиях функционального биоразнообразия: консолидация и прогресс в исследованиях BDEF. Экол. лат. 2009 г.;12:1405–1419. doi: 10.1111/j.1461-0248.2009.01388.x. [PubMed] [CrossRef] [Google Scholar]
6. Блютген Н., Кляйн А.М. Функциональная взаимодополняемость и специализация: роль биоразнообразия во взаимодействии растений и опылителей. Базовое приложение Экол. 2011;12:282–291. doi: 10.1016/j.baae.2010.11.001. [CrossRef] [Google Scholar]
7. Schleuning M, Fründ J, García D. Прогнозирование функций экосистем на основе биоразнообразия и мутуалистических сетей: расширение концепций, основанных на признаках, на взаимодействие растений и животных. Экография. 2015; 38:1–13. doi: 10.1111/ecog.00983. [CrossRef] [Google Scholar]
8. Gravel D, Albouy C, Thuiller W. Значение функционального состава пищевых сетей для функционирования экосистемы. Филос. Транс. Р. Соц. Лонд. Б. 2016; 371:20150268. doi: 10.1098/rstb.2015.0268. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
9. Vollstädt MGR, et al. Прямое и косвенное влияние климата, вмешательства человека и особенностей растений на функциональное разнообразие птиц. Глоб. Экол. Биогеогр. 2017;26:963–972. doi: 10.1111/geb.12606. [Перекрестная ссылка] [Академия Google]
10. Хантер, доктор медицинских наук, цена PW. Игра в желоба и лестницы: неоднородность и относительная роль восходящих и нисходящих сил в естественных сообществах. Экология. 1992; 73: 724–732. [Google Scholar]
11. Cardinale BJ, et al. Влияние биоразнообразия на функционирование экосистем: сводка 164 экспериментальных манипуляций с видовым богатством: Экологический архив E090-060. Экология. 2009; 90: 854–854. дои: 10.1890/08-1584.1. [CrossRef] [Google Scholar]
12. Scherber C, et al. Влияние разнообразия растений снизу вверх на мультитрофические взаимодействия в эксперименте по биоразнообразию. Природа. 2010; 468: 553–556. дои: 10.1038/nature09492. [PubMed] [CrossRef] [Google Scholar]
13. Díaz S, et al. Функциональные черты, филогения функций и уязвимость экосистемных услуг. Экол. Эвол. 2013;3:2958–2975. doi: 10.1002/ece3.601. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
14. Gagic V, et al. Функциональная идентичность и разнообразие животных предсказывают функционирование экосистем лучше, чем индексы, основанные на видах. проц. биол. науч. 2015;282:20142620. doi: 10.1098/rspb.2014.2620. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
15. Duffy JE, et al. Функциональная роль биоразнообразия в экосистемах: включая трофическую сложность. Экол. лат. 2007; 10: 522–538. doi: 10.1111/j.1461-0248.2007.01037.x. [PubMed] [CrossRef] [Google Scholar]
16. Schleuning M, et al. Специализация и сила взаимодействия в сети тропических растений и плодоядных различаются в разных лесных ярусах. Экология. 2011;92:26–36. дои: 10.1890/09-1842.1. [PubMed] [CrossRef] [Google Scholar]
17. Станг М., Клинкхамер П.Г.Л., ван дер Мейден Э. Ограничения по размеру и изобилие цветов определяют количество взаимодействий в сети посетителей растений и цветов. Ойкос. 2006; 112:111–121. дои: 10.1111/j.0030-1299.2006.14199.х. [CrossRef] [Google Scholar]
18. Юнкер Р.Р., Блютген Н., Келлер А. Функциональное и филогенетическое разнообразие растительных сообществ по-разному влияет на структуру взаимодействия цветок-посетитель и выявляет конвергенцию цветочных признаков. Эволют. Экол. 2015; 29: 437–450. doi: 10.1007/s10682-014-9747-2. [CrossRef] [Google Scholar]
19. Junker RR, et al. Специализация по признакам как основа для широты ниши посетителей цветов и как механизм структурирования экологических сетей. Функц. Экол. 2013;27:329–341. doi: 10.1111/1365-2435.12005. [CrossRef] [Google Scholar]
20. Schlinkert H, et al. Размер растения влияет на мутуалистические и антагонистические взаимодействия и репродуктивный успех у 21 вида Brassicaceae. Экосфера. 2016;7:e01529. doi: 10.1002/ecs2.1529. [CrossRef] [Google Scholar]
21. Дафни А., Поттс С.Г. Роль наклона, глубины и высоты цветка в предпочтениях жука-опылителя (Coleoptera: Glaphyridae) J. Insect Behav. 2004; 17: 823–834. doi: 10.1023/B:JOIR.0000048991.45453.73. [CrossRef] [Google Scholar]
22. Хён П., Чарнтке Т., Тилианакис Дж. М., Штеффан-Девентер И. Разнообразие функциональных групп пчел-опылителей повышает урожайность. проц. биол. науч. 2008; 275:2283–2291. doi: 10.1098/rspb.2008.0405. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
23. Parsche S, Fründ J, Tscharntke T. Экспериментальное изменение окружающей среды и мутуалистические и антагонистические взаимодействия растения-цветка-посетителя. Перспектива. Завод Экол. Эвол. Сист. 2011;13:27–35. doi: 10.1016/j.ppees.2010.12.001. [Перекрестная ссылка] [Академия Google]
24. Dehling DM, et al. Функциональные отношения, выходящие за рамки моделей видового богатства: соответствие признаков взаимного обмена растениями и птицами в разных масштабах. Глоб. Экол. Биогеогр. 2014;23:1085–1093. doi: 10.1111/geb.12193. [CrossRef] [Google Scholar]
25. Dray S, et al. Сочетание методов четвертого угла и RLQ для оценки реакции признаков на изменение окружающей среды. Экология. 2014;95:14–21. дои: 10. 1890/13-0196.1. [PubMed] [CrossRef] [Google Scholar]
26. Dolédec S, Chessel D, ter Braak CJF, Champely S. Сопоставление видовых признаков с переменными окружающей среды: новый метод ординации с тремя таблицами. Окружающая среда. Экол. Стат. 1996;3:143–166. doi: 10.1007/BF02427859. [CrossRef] [Google Scholar]
27. ter Braak CJF, Cormont A, Dray S. Улучшенное тестирование взаимосвязей видовых признаков и окружающей среды в задаче четвертого угла. Экология. 2012;93:1525–1526. дои: 10.1890/12-0126.1. [PubMed] [CrossRef] [Google Scholar]
28. Moran MD. Аргументы в пользу отказа от последовательного Бонферрони в экологических исследованиях. Ойкос. 2003; 100:403–405. doi: 10.1034/j.1600-0706.2003.12010.x. [CrossRef] [Академия Google]
29. Блютген Н., Фрунд Дж., Васкес Д.П., Мензель Ф. Что метрики сети взаимодействия говорят нам о специализации и биологических чертах? Экология. 2008; 89: 3387–3399. дои: 10.1890/07-2121.1. [PubMed] [CrossRef] [Google Scholar]
30. Wheelwright NT. Размер плодов, ширина зева и рацион птиц, питающихся фруктами. Экология. 1985; 66: 808–818. дои: 10.2307/1940542. [CrossRef] [Google Scholar]
31. Пайк Г.Х., Пуллиам Х.Р., Чарнов Э.Л. Оптимальный поиск пищи: выборочный обзор теории и тестов. Q. Преподобный Биол. 1977;52:137–154. дои: 10.1086/409852. [CrossRef] [Google Scholar]
32. Haskell JP, Ritchie ME, Olff H. Фрактальная геометрия предсказывает различные отношения масштабирования размера тела для участков обитания млекопитающих и птиц. Природа. 2002; 418: 527–530. doi: 10.1038/nature00840. [PubMed] [CrossRef] [Google Scholar]
33. Fontaine C, Collin CL, Dajoz I. Универсальный поиск пищи для опылителей: расширение рациона при высокой плотности. Дж. Экол. 2008;96:1002–1010. doi: 10.1111/j.1365-2745.2008.01405.x. [Перекрестная ссылка] [Академия Google]
34. Albrecht J, et al. Вырубка леса и лесные опушки уменьшают избыточность в сетях растений и плодоядных в старовозрастных европейских лесах. Дж. Экол. 2013; 101:990–999. doi: 10.1111/1365-2745.12105. [CrossRef] [Google Scholar]
35. Нойшульц Э.Л., Браун М., Фарвиг Н. Частые перемещения птиц по сильно фрагментированному ландшафту: роль видовых признаков и лесной матрицы. Аним. Консерв. 2013;16:170–179. doi: 10.1111/j.1469-1795.2012.00582.x. [CrossRef] [Google Scholar]
36. Eklöf A, et al. Размерность экологических сетей. Экол. лат. 2013; 16: 577–583. doi: 10.1111/ele.12081. [PubMed] [CrossRef] [Академия Google]
37. Спасоевич М.Ю., Судинг К.Н. Вывод механизмов сборки сообщества из моделей функционального разнообразия: важность множественных процессов сборки. Дж. Экол. 2012; 100: 652–661. doi: 10.1111/j.1365-2745.2011.01945.x. [CrossRef] [Google Scholar]
38. Hawkins BA, et al. Энергия, вода и широкомасштабные географические модели видового богатства. Экология. 2003;84:3105–3117. дои: 10.1890/03-8006. [CrossRef] [Google Scholar]
39. Currie DJ, et al. Прогнозы и проверка основанных на климате гипотез широкомасштабных изменений таксономического богатства. Экол. лат. 2004;7:1121–1134. doi: 10.1111/j.1461-0248.2004.00671.x. [Перекрестная ссылка] [Академия Google]
40. Эванс К.Л., Уоррен П.Х., Гастон К.Дж. Видо-энергетические отношения в макроэкологическом масштабе: обзор механизмов. биол. 2005 г.; 80:1–25. doi: 10.1017/S1464793104006517. [PubMed] [CrossRef] [Google Scholar]
41. Kreft H, Jetz W. Глобальные закономерности и детерминанты разнообразия сосудистых растений. проц. . Натл акад. науч. США. 2007; 104: 5925–5930. doi: 10.1073/pnas.0608361104. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
42. Kissling WD, Field R, Böhning-Gaese K. Пространственные модели разнообразия древесных растений и птиц: функциональные отношения или влияние окружающей среды? Глоб. Экол. Биогеогр. 2008; 17: 327–339.. doi: 10.1111/j.1466-8238.2007.00379.x. [CrossRef] [Google Scholar]
43. Маккейн CM. Глобальный анализ высотного разнообразия птиц. Глоб. Экол. Биогеогр. 2009; 18: 346–360. doi: 10.1111/j.1466-8238.2008. 00443.x. [CrossRef] [Google Scholar]
44. Peters MK, et al. Предикторы высотных градиентов биоразнообразия меняются от отдельных таксонов до уровня сообщества с несколькими таксонами. Нац. коммун. 2016;7:13736. doi: 10.1038/ncomms13736. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
45. Schellenberger Costa D, et al. Средневзвешенные значения сообщества и функциональная дисперсия функциональных признаков растений вдоль градиентов окружающей среды на горе Килиманджаро. Дж. Вег. науч. 2017; 28: 684–695. doi: 10.1111/jvs.12542. [CrossRef] [Google Scholar]
46. Classen A, Steffan-Dewenter I, Kindeketa WJ, Peters MK. Интеграция внутривидовой изменчивости в экологию сообщества объединяет теории об изменении размера тела в зависимости от климатических градиентов. Функц. Экол. 2017; 31: 768–777. doi: 10.1111/1365-2435.12786. [Перекрестная ссылка] [Академия Google]
47. Спасоевич М.Дж., Грейс Дж.Б., Харрисон С., Дамшен Э.И. Функциональное разнообразие поддерживает гипотезу физиологической толерантности к богатству видов растений в зависимости от климатических градиентов. Дж. Экол. 2014; 102: 447–455. doi: 10.1111/1365-2745.12204. [CrossRef] [Google Scholar]
48. Tylianakis JM, Didham RK, Bascompte J, Wardle DA. Глобальные изменения и взаимодействие видов в наземных экосистемах. Экол. лат. 2008; 11:1351–1363. doi: 10.1111/j.1461-0248.2008.01250.x. [PubMed] [CrossRef] [Академия Google]
49. Альбрехт М., Ризен М., Шмид Б. Сборка сети растений-опылителей вдоль хронологической последовательности форланда ледника. Ойкос. 2010; 119:1610–1624. doi: 10.1111/j.1600-0706.2010.18376.x. [CrossRef] [Google Scholar]
50. Classen A, et al. Температура по сравнению с ограниченностью ресурсов: какие факторы определяют разнообразие пчел на горе Килиманджаро, Танзания? Глоб. Экол. Биогеогр. 2015; 24:642–652. doi: 10.1111/geb.12286. [CrossRef] [Google Scholar]
51. Андерсон К.Дж., Джетц В. Широкомасштабная экология расхода энергии эндотермами: ограничения на энергетику эндотермов. Экол. лат. 2005; 8: 310–318. doi: 10.1111/j.1461-0248.2005.00723. x. [Перекрестная ссылка] [Академия Google]
52. Конопля А. Континуум или зональность? Высотные градиенты в лесной растительности горы Килиманджаро. Завод Экол. 2006; 184:27–42. doi: 10.1007/s11258-005-9049-4. [CrossRef] [Google Scholar]
53. Конопля А. Растительность Килиманджаро: скрытые эндемики и недостающий бамбук. фр. Дж. Экол. 2006; 44: 305–328. doi: 10.1111/j.1365-2028.2006.00679.x. [CrossRef] [Google Scholar]
54. Guimarães PR, Pires MM, Jordano P, Bascompte J, Thompson JN. Косвенные эффекты стимулируют коэволюцию в мутуалистических сетях. Природа. 2017; 550:511–514. doi: 10.1038/nature24273. [PubMed] [CrossRef] [Академия Google]
55. Vollstädt MGR, et al. Сети распространения семян по-разному реагируют на воздействие ресурсов в открытых и лесных местообитаниях. Ойкос. 2017; 127:847–854. doi: 10.1111/oik.04703. [CrossRef] [Google Scholar]
56. Циммерман Д. А., Тернер Д. А., Пирсон Д. Дж. и Уиллис И. Птицы Кении и Северной Танзании (A & C Black, Лондон, Великобритания, 1996).
57. Eck, S. et al. Измерение птиц — Vögel Vermessen (Deutsche Ornithologen-Gesellschaft, Вильгельмсхафен, Германия, 2011 г.).
58. Террилл, В. Б. и Милн-Рэдхед, Э. Флора тропической Восточной Африки (Агенты короны для колоний, Лондон, Великобритания, 1952).
59. Wilman H, et al. EltonTraits 1.0: Особенности кормодобывания птиц и млекопитающих мира на уровне вида: Экологический архив E095-178. Экология. 2014;95:2027–2027. дои: 10.1890/13-1917.1. [CrossRef] [Google Scholar]
60. Dunning, JB CRC Handbook of Avian Body Masses (CRC Press, Бока-Ратон, Флорида, США, 2008 г.).
61. Krenn HW, Plant JD, Szucsich NU. Ротовые органы насекомых, посещающих цветы. Структура членистоногих. Дев. 2005; 34:1–40. doi: 10.1016/j.asd.2004.10.002. [CrossRef] [Google Scholar]
62. Bascompte J, Jordano P, Olesen JMM. Асимметричные коэволюционные сети облегчают поддержание биоразнообразия. Наука. 2006; 312: 431–433. doi: 10. 1126/science.1123412. [PubMed] [CrossRef] [Google Scholar]
63. Дрей С., Лежандр П. Проверка взаимосвязей видовых признаков и окружающей среды: новый взгляд на проблему четвертого угла. Экология. 2008;89: 3400–3412. дои: 10.1890/08-0349.1. [PubMed] [CrossRef] [Google Scholar]
64. Дрей С., Петторелли Н., Чессел Д. Сопоставление наборов данных из двух разных пространственных выборок. Дж. Вег. науч. 2002; 13: 867–874. doi: 10.1111/j.1654-1103.2002.tb02116.x. [CrossRef] [Google Scholar]
65. Лалиберте Э., Лежандр П. Основанная на расстоянии структура для измерения функционального разнообразия по множеству признаков. Экология. 2010;91:299–305. дои: 10.1890/08-2244.1. [PubMed] [CrossRef] [Google Scholar]
66. Квадратичная энтропия Ботта-Дуката З. Рао как мера функционального разнообразия на основе множества признаков. Дж. Вег. науч. 2005; 16: 533–540. doi: 10.1111/j.1654-1103.2005.tb02393.х. [CrossRef] [Google Scholar]
67. Nakagawa S, Santos ESA. Методологические проблемы и достижения в области биологического метаанализа.